Embed Size (px)
Citation preview

1388 Blood, Vol 67, No 5 (May), 1986: pp 1388-1394
Activation of Human Neutrophils by Monoclonal Antibody PMN7C3:Cell Movement and Adhesion Can Be Triggered Independently
From the Respiratory Burst
By David A. Melnick, Tova Meshulam, Aurelio Manto, and Harry L. Malech
Anti-neutrophil monoclonal antibody PMN7C3 (IgG3) rec-
ognizes glycoproteins bearing the oligosaccharide lacto-
N-fucopentaose III, including the C3bi receptor, LFA-1 , and
p1 50.95 on the plasma membrane and a group of granule-
associated glycoproteins. We have previously shown that
binding of this antibody to polymorphonuclear leukocytes
(PMNs) stimulates a transient rise in cytosolic free calcium
concentration but does not trigger the neutrophil respira-
tory burst. We now demonstrate that binding of PMN7C3
(and five other monoclonal antibodies recognizing the same
antigen) to human neutrophils activates several other
cellular responses. Addition of PMN7C3 to monolayers of
neutrophils induces a rapid change in cell shape followed
by pseudopod formation and increased migration. With
P OLYMORPHONUCLEAR leukocytes (PMNs) can
be activated to migrate to sites of inflammation and to
generate toxic mediators of the inflammatory response. The
events that link receptor-ligand binding to cell activation
remain uncertain.
PMN7C3 is an anti-human neutrophil murine monocbonal
antibody that inhibits opsonized zymosan-induced respira-
tory burst but enhances phagocytosis.’2 It recognizes glyco-
proteins bearing the obigosaccharide bacto-N-fucopentaose
III, including the C3bi receptor (CR3), LFA-l, and p150,95
on the plasma membrane and a group of granule-associated
glycoproteins.2 Binding of PMN7C3 results in capping of
antigen-antibody complexes on the cell surface and induces
a transient rise in free cytosolic calcium concentration,
(Ca2� ) but does not trigger the respiratory burst.’3 Thus, a
transient rise in (Ca2� ), cannot be a sufficient signal for
activation of this response.
In the present study, we show that binding of PMN7C3 to
human neutrophils stimulates shape changes, leukoaggluti-
nation, chemokinesis, and chemotaxis. These data suggest
that activation pathways beading to cell movement and
adhesion can be triggered independently from those leading
to the respiratory burst.
MATERIALS AND METHODS
Cell preparation. Venous blood from human volunteers was
mixed with heparin (10 U/mL). PMNs (>95% purity) were
From the Evans Memorial Department of Clinical Research and
the Department of Medicine, Boston University Medical Center;
and the Department of Medicine, Yale University. New Haven,
Conn.
Supported in part by grants No. A106937 and AI2I 972from the
National institute of Allergy and Infectious Diseases. National
Institutes ofHealth. DAM. was a Daland Fellow ofthe American
Philosophical Society during the course ofthis work.
Submitted July 29. 1985; accepted Jan 3. 1986.
Address reprint requests to Dr David A. Melnick, Section of
Infectious Diseases, Room E-529, University Hospital. 75 E New-
ton St. Boston, MA 02118.
(c) I 986 by Grune & Stratton, Inc.
0006-4971/86/6705-0036$03.00/0
incubation at 37 ‘C. the neutrophils aggregate in clusters
(leukoagglutination). Quantitation of cell movement in a
multiwell chemotaxis assembly or by migration of PMNs
under agarose revealed that PMN7C3 is both chemotactic
and chemokinetic. Pretreatment with the antibody inhibits
subsequent chemotactic response to other stimuli. Mono-
clonal antibodies binding to other neutrophil antigens do
not mimic these effects. These data suggest that cell
movement and adhesion can be triggered independently
from the respiratory burst. PMN7C3 may be a useful probe
with which to study the events that link receptor-ligand
binding to cellular response.
a 1986 by Grune & Stratton. Inc.
prepared by sedimentation of erythrocytes with 3% dextran followedby centrifugation of leukocytes over Ficoll-Hypaque gradients (His-
topaque 1083, Sigma Chemical Co., St. Louis).4 Residual erythro-cytes were removed by hypotonic lysis. PMNs were suspended inice-cold Hank’s balanced salt solution (HBSS) until use.
Preparation ofmonoclonal antibodies. Murine hybridoma linessecreting anti-human neutrophil monoclonab antibodies wereselected as previously described.’2 Five 1gM antibodies (95G8,43B9, 74D8, 66B7, and 80C9) bind to the same antigen as PMN7C3(IgG3) as determined by Western blotting pattern and competitivebinding studies. Control antineutrophil monocbonal antibodies
(31D8, l3F6, 41C5, 8C7, and 55F3) bind to other antigens. Mono-cbonal immunogbobulin was partially purified from malignant ascites
by two successive precipitations with 45% ammonium sulfate. Anti-bodies were stored in phosphate-buffered saline (PBS) at - 70 #{176}Cuntil thawed for use. Additional monoclonab antibodies were kindlyprovided by Dr Samuel Wright (IB4; anti-CR3) and Dr John
O’Shea (3D9; anti-CRI). Hybridoma line Ml/70, secreting anti-MAC- 1, was obtained from the American Type Culture Collection.
Leukoagglutination assay. Leukoagglutination of adherentneutrophils was assayed by modification of the technique of Lalezari
and colleagues.56 PMNs (5 x 106/mL HBSS) were added to each of24 wells on plastic tissue culture plates (Costar, Cambridge, Mass).After a ten-minute incubation at 37 #{176}C,nonadherent cells wereaspirated, leaving behind a homogeneous monolayer of neutrophils.The medium was then replaced with HBSS alone or buffer contain-ing 3.3 x l0� mob/L of monocbonal antibody. In some experiments,
cytochalasin B (5 �zg/mL) was added five minutes prior to theaddition of monoclonal antibody. The monolayers were then incu-bated for 60 minutes at 37 #{176}C.After incubation, the monolayerswere examined under phase optics and scored for beukoagglutination
(0, homogeneous monolayer; 4 + , all cells aggregated in clusters) bytwo observers blinded to the daily protocol (concordance > 95%). Insome experiments (as indicated below), incubation was carried out
at room temperature rather than at 37 #{176}C.Time-lapse videomicroscopy. The events that followed binding
of monoclonal antibody to the surface of adherent neutrophils wererecorded on time-lapse videotapes. Monolayers of PMN were pre-
pared as above and observed on the temperature-controlled stage ofan inverted phase microscope. Images were recorded at lOOxmagnification using a Panasonic video camera and time-lapse videocassette recorder with internal clock. These tapes were used to assessthe time sequence of shape and motility changes which occurredafter addition of monoclonal antibody or other stimuli to theincubation buffer.
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom

NEUTROPHIL ACTIVATION BY ANTIBODY PMN7C3 1389
Chemotaxis and chemokinesis. Neutrophib migration was
quantitated using a multiwell chemotaxis assembly (Neuro Probe,Bethesda, Md) modeled after that designed by Falk et al.7 Potential
chemoattractants (in gelatin veronal buffer) were loaded in the
lower wells and separated from PMNs [in Gey’s buffer with 2%bovine serum albumin (BSA)] by filters with 3-zm pores (Neuro
Probe, Bethesda, Md). After incubation (30 minutes at 37 #{176}C),thefilters were fixed, stained, and examined at 40x magnification. Thedistance from the top of the filter to the last focal plane in which two
neutrophils were clearly visualized was measured as an index of
migration as originally described by Zigmond and Hirsch.8 Random
migration was defined as the distance migrated toward buffer
without added chemoattractant. The distance migrated toward a
potential chemoattractant was defined as directed migration. To
differentiate between chemotactic and chemokinetic effects, a
“checkerboard analysis” was carried out in which the compounds tobe tested were added both above and below the filter to vary thegradient systematically.89
Chemotaxis was also assayed by migration of neutrophils onplastic plates under 1% agarose.’0” Plastic tissue culture dishes (60
mm) were coated with 0.5% gelatin which was then overlayed with 6mL of 1% agarose in minimal essential medium (MEM) containing20 mmol/L of HEPES, 10 mmol/L of NaHCO3, and 0.5% gelatin.
A metal dye was then used to punch a radial array of triplicate wellsin the agarose layer. A potential chemoattractant was placed in theouter well, and buffer was placed in the inner well of each triplicate
set; 2.5 x l0� PMNs were then added to the center well, and theplates were incubated at 37 #{176}Cfor two or three hours. Afterincubation, plates were fixed by sequential additional of methanol
and formaldehyde, and the agarose layer was removed. PMNs
adherent to the plastic dish were then stained with Giemsa. Thedistance from the edge of the center well to the beading front of
PMNs advancing toward chemoattractant (directed migration) andtoward buffer (random migration) was measured at 40x magnifica-
tion using a micrometer. In experiments in which monoclonal
antibodies were used as potential chemoattractants, plates wereincubated overnight prior to addition of PMNs to allow for gradient
formation.
Inhibition of chemotaxis by monoclonal antibodies. Inhibition
of chemotactic response by monocbonal antibodies was examined bypretreating neutrophils with varying concentrations of monocbonal
immunoglobubin for I 5 minutes at 4 #{176}C,after which cells werewashed with buffer to remove unbound antibody. Migration toward
N-formyl norleucyl leucyl phenylalanine (fNLLP) or zymosan-
activated serum (ZAS) was then measured by the techniques above.
Migration by antibody-treated cells was compared to migration by
control PMNs pretreated with buffer lacking monocbonal antibody.
Reagents. ZAS was prepared on the day of use by incubatingpooled human serum with prehydrated zymosan (10 mg/mL) for 20
minutes at 37 #{176}C.All reagents were purchased from Sigma except as
indicated.Human experimentation. All blood specimens were collected
following guidelines approved by the Institutional Review Board forHuman Experimentation of University Hospital.
RESULTS
PMN7C3 induces adherent neutrophils to leukoaggluti-
nate. Lalezari and co-workers observed that naturally
occurring anti-neutrophil antisera (derived from mothers of
infants with isoimmune neonatal neutropenia) could induce
adherent PMNs to migrate into large clusters in an active,
energy-dependent process. He termed this response leukoag-
gbutination.5’6 We screened a panel of anti-neutrophil mono-
clonal antibodies for the ability to induce this response.
Preformed PMN monolayers in multiwell plastic tissue
culture plates were incubated with monocbonab antibody
(3.3 x l0� mob/L) or buffer for 60 minutes at 37 #{176}C.As
shown in Fig 1 , incubation with PMN7C3 (but not a control
anti-neutrophil monocbonal IgG) induced the adherent neu-
trophils to migrate into clusters. This reaction did not occur
if cells were incubated at room temperature rather than at
37 #{176}C,suggesting that it is an active process. Furthermore,
fixation of neutrophils with I % paraformaldehyde prior to
antibody treatment blocked beukoagglutination, although it
does not inhibit binding of PMN7C3 as detected by
enzyme-linked immunosorbent assay (ELISA).”2 In addi-
tion, leukoagglutination was blocked by pretreatment of
neutrophils with cytochalasin B (S zg/mL), suggesting that
this response requires an intact cytoskeleton.
The results of leukoagglutination assays with a panel of
anti-neutrophil monocbonal antibodies are summarized in
Table 1. In addition to PMN7C3 (IgG3), we have isolated
five other monocbonab antibodies (all 1gM) which bind to the
Fig 1 . Phase photomicrographs of monolayers of neutrophilsafter incubation (60 minutes, 37 ‘C) with (A) 3.3 x i0� mol/L ofPMN7C3 or (B) control anti-neutrophil lgG 31 D8. In the presenceof PMN7C3. virtually all the neutrophils have aggregated in clus-ters. These photomicrographs are from a representative experi-ment of ten, each using granulocytes from a different donor.
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom

1390 MELNICK ET AL
Table 1 . Monoclonal Antibod y-lnduced Leukoagglutination
Antibody Leukoagglutination
Anti-PMN7A9t
7C3 3-4+
95G8 4+
43B9 2-3+
74D8 4+
66B7 4+
80C9 4+
Control*
31D8 0
13F6 0-2+
M1/70 0
41C5 0
8C7 0-1+
55F3 0
‘Plates were scored for formation of cell clusters by two observers
blinded to the daily protocol (0, homogeneous monolayer; 4 + , all cells
aggregated in clusters). Data represent the range of values recorded in at
least five experiments with each antibody, using granulocytes isolated
from different donors. The two groups of antibodies differ significantly in
their ability to induce leukoagglutination (P < .001 . Wilcoxon rank-sum
analysis).
tAntibodies that bind to the same antigen as PMN7C3 as detected byWestern blotting pattern and competitive binding studies.
�MonocIonaI antibodies binding to other neutrophil antigens.
same antigen as detected by Western blotting pattern and
competitive binding studies.2 These five antibodies also
induce PMNs to beukoaggbutinate. In contrast, six control
monocbonal antibodies binding to other neutrophil antigens
did not induce leukoagglutination (P < .001, Wilcoxon
rank-sum analysis). Control antibodies included Ml/70
(anti-Mac I) which recognizes an antigen associated with
the C3bi receptor,’2”3 but does not compete with PMN7C3
for binding in a crossed radioimmunoassay. The ability of
different antibodies to induce beukoagglutination did not
correlate with the amount of each which binds to the cell
surface as detected by ELISA (data not shown). These
results suggest that leukoagglutination is not due to passive
crosslinking of cells coated with multivalent ligands.
Binding of PMN7C3 to neutrophils induces shape
changes and increased migration. Immunoflourescence
microscopy has demonstrated that after binding to neutro-
phils, PMN7C3 undergoes capping to the uropod of the cell.2
The time course of other cellular events which follow binding
of PMN7C3 was examined using time-lapse videotapes
recorded through a phase microscope. A sequence of still
photographs taken from a representative experiment is pre-
sented in Fig 2. Monolayers of adherent PMNs were formed
by preincubation ofcells in HBSS at 37 #{176}C(Fig 2a). Cellular
changes became evident 30 to 40 seconds after the addition
of PMN7C3 (3.3 x l0� mob/L) to the buffer. Initially cells
became less rounded, and many assumed a triangular shape
(Fig 2b). This was quickly followed by formation of pseudo-
pods, with a rapid increase in the rate of migration (Fig 2c).
Within two to three minutes of the addition of PMN7C3,
small clusters ofcebls started to form (Fig 2d). These changes
did not occur after addition of control antineutrophil IgG
(PMN3ID8) tothe incubation medium.
PMN7C3 is both chemoauractant and chemokinet-
Ic. The effect of PMN7C3 on neutrophil migration was
quantitated by modification of the leading front assay
described by Zigmond and Hirsch.8 When used in the
chemoattractant webb, PMN7C3 induced a significant incre-
ment in migration in concentrations �3.3 x l0� mol/L (Fig
3). This effect increased in a dose-dependent fashion with
antibody concentrations up to 3.3 x 10_s mol/L. Higher
concentrations of PMN7C3 in the chemoattractant well did
not produce any further increment in neutrophib migration.
The directed migration induced by l0� mol/L of PMN7C3
(74.7 ± 3.5 �m) was similar to that produced by an equimo-
bar concentration of fNLLP (69.5 ± 2.5 zm). In contrast,
monocbonal IgG binding to other neutrophil antigens
(PMN31D8, Ml/70) did not induce increased migration
(Fig 4). Similar results were obtained when chemotaxis was
assayed by migration of PMNs under 1% agarose (data not
shown).
To differentiate between chemokinetic (increased random
migration) and chemotactic (increased directed migration)
effects of PMN7C3, we performed a checkerboard analysis
as originally described by Zigmond and Hirsch.8’9 In these
experiments, varied concentrations of PMN7C3 were added
to both upper (PMN) and lower (chemoattractant) wells to
alter the gradient across the filter. These results are summa-
rized in Table 2. As the concentration of PMN7C3 in the
lower wells was increased (vertical columns), neutrophil
migration increased in a dose-dependent fashion. Reversal of
the gradient by addition of increasing concentrations of
PMN7C3 to the upper wells (horizontal rows) inhibited
migration of neutrophils into the filter. These data suggest
that PMN7C3 is a chemoattractant. Exposure of neutrophils
to PMN7C3 in the absence of a chemotactic gradient
(diagonal row) also produced a dose-dependent increase in
neutrophil migration, although of a smaller magnitude than
that seen in the presence of a gradient across the filter. Thus,
PMN7C3 also induces chemokinesis.
PMN7C3 inhibits chemotactic response to ZAS and
JNLLP. Exposure of neutrophils to a chemotactic stimulus
such as N-formyl-met-beu-phe (fMLP) may inhibit subsequent
response to the formyl peptide and to other chemotactic
agents, a process which has been termed deactivation.’�’6 To
examine the effect of PMN7C3 on response to other che-
moattractants, neutrophibs were pretreated with the antibody
(6.7 x 10� mol/L) for 15 minutes at 4 #{176}Cand then washed
to remove unbound IgG. Migration of pretreated neutrophils
toward ZAS or fNLLP was then measured. Chemotaxis
toward both agents over a range of concentrations was
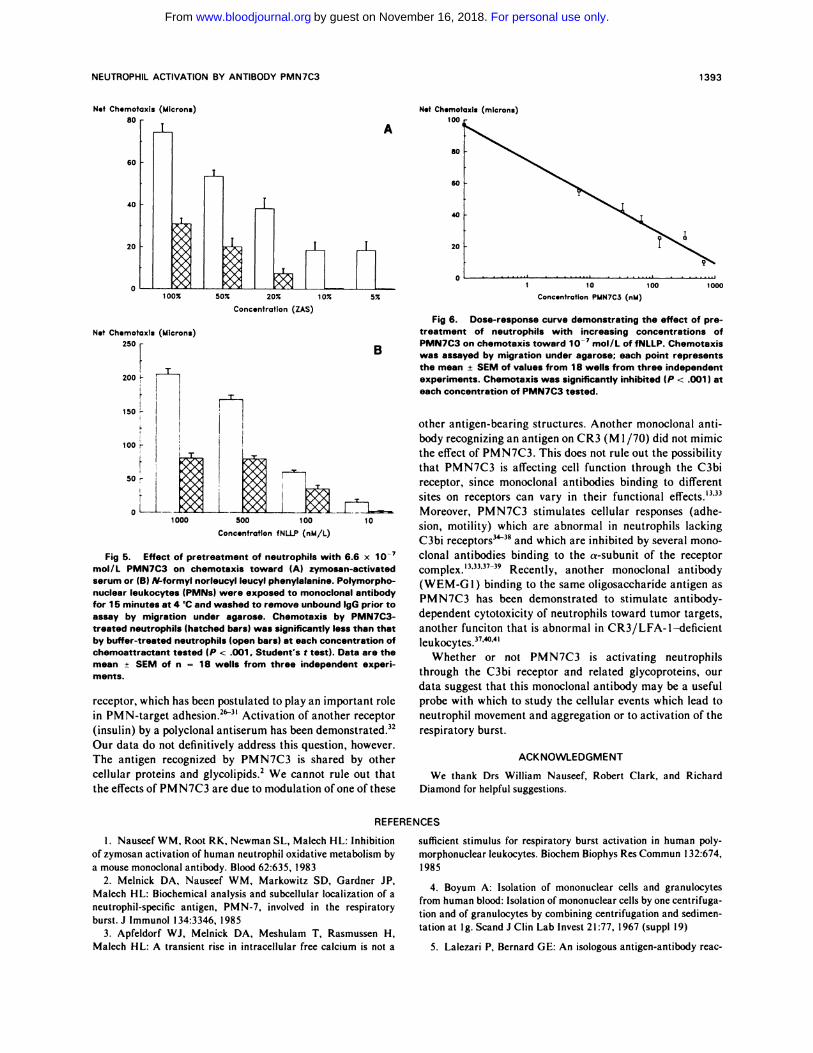
significantly inhibited by pretreatment with PMN7C3 (Fig
5). Five other monocbonal antibodies that bind to the same
antigen as PMN7C3 had a similar effect on chemotaxis
toward ZAS, inhibiting migration to 22.8% ± 3% of control
(mean ± SEM, P < .001 ). In contrast, six monoclonal
antibodies binding to other neutrophil antigens did not
inhibit chemotaxis (mean migration 94.6% ± I 7% of con-
trol). The degree of inhibition of chemotaxis by PMN7C3
was dependent on the concentration of monocbonal antibody
used to pretreat the neutrophils. Inhibition was seen with
antibody concentrations as low as 6.7 x l0� mol/L (Fig 6).
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom
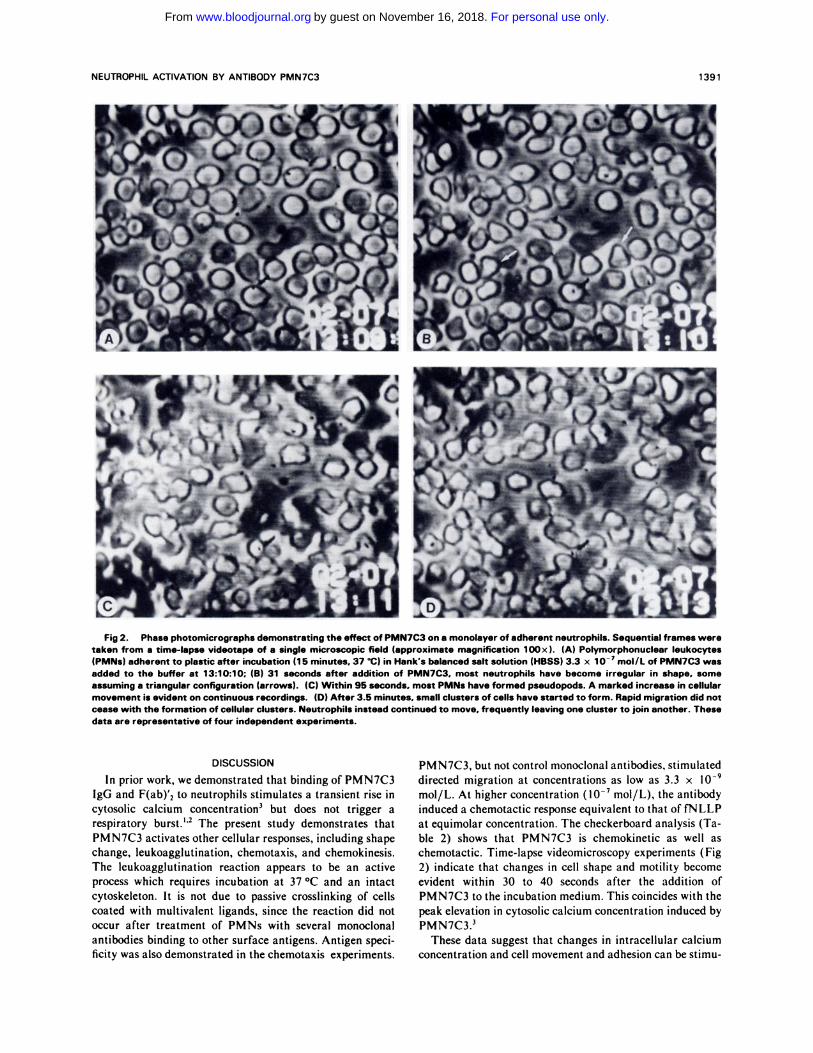
NEUTROPHIL ACTIVATION BY ANTIBODY PMN7C3 1391
Fig 2. Phase photomicrographs demonstrating the effect of PMN7C3 on a monolayer of adherent neutrophils. Sequential frames weretaken from a time-lapse videotape of a single microscopic field (approximate magnification lOOx ). (A) Polymorphonuclear leukocytes(PMNs) adherent to plastic after incubation (1 5 minutes. 37 ‘C) in Hank’s balanced salt solution (HBSS) 3.3 x iO� mol/L of PMN7C3 was
added to the buffer at 13:10:10; (B) 31 seconds after addition of PMN7C3. most neutrophils have become irregular in shape, someassuming a triangular configuration (arrows). (C) Within 95 seconds, most PMNs have formed pseudopods. A marked increase in cellularmovement is evident on continuous recordings. (D) After 3.5 minutes. small clusters of cells have started to form. Rapid migration did notcease with the formation of cellular clusters. Neutrophils instead continued to move. frequently leaving one cluster to join another. These
data are representative of four independent experiments.
DISCUSSION
In prior work, we demonstrated that binding of PMN7C3
IgG and F(ab)’2 to neutrophibs stimulates a transient rise in
cytosolic calcium concentration3 but does not trigger a
respiratory burst.’2 The present study demonstrates that
PMN7C3 activates other cellular responses, including shape
change, beukoagglutination, chemotaxis, and chemokinesis.
The leukoagglutination reaction appears to be an active
process which requires incubation at 37 #{176}Cand an intact
cytoskeleton. It is not due to passive crosslinking of cells
coated with multivalent ligands, since the reaction did not
occur after treatment of PMNs with several monocbonal
antibodies binding to other surface antigens. Antigen speci-
ficity was also demonstrated in the chemotaxis experiments.
PMN7C3, but not control monocbonal antibodies, stimulated
directed migration at concentrations as low as 3.3 x l0�
mob/L. At higher concentration (l0� mob/L), the antibody
induced a chemotactic response equivalent to that of fNLLP
at equimolar concentration. The checkerboard analysis (Ta-
ble 2) shows that PMN7C3 is chemokinetic as well as
chemotactic. Time-lapse videomicroscopy experiments (Fig
2) indicate that changes in cell shape and motility become
evident within 30 to 40 seconds after the addition of
PMN7C3 to the incubation medium. This coincides with the
peak elevation in cytosolic calcium concentration induced by
PMN7C3.3
These data suggest that changes in intracellular calcium
concentration and cell movement and adhesion can be stimu-
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom
80
70
50
10
Concentration PMN7C3 (nil) HI Serum FN�P ZAS 181/70 3108 PWN7C3
1392 MELNICK ET AL
represent the mean ± SEM of values from two experiments.
Migration (microns)
90
so
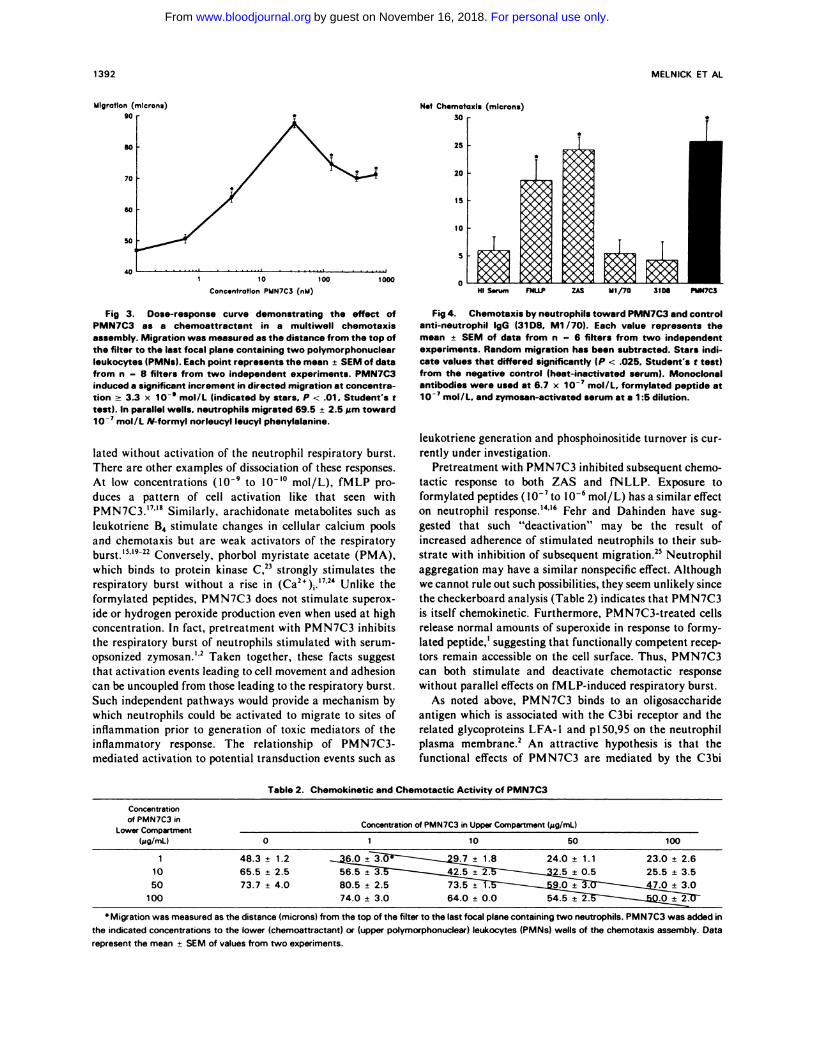
Fig 3. Dose-response curve demonstrating the effect of
PMN7C3 as a chemoattractant in a multiwell chemotaxisassembly. Migration was measured as the distance from the top ofthe filter to the last focal plane containing two polymorphonuclearleukocytes (PMNs). Each point represents the mean ± SEM of datafrom n - 8 filters from two independent experiments. PMN7C3induced a significant increment in directed migration at concentra-tion � 3.3 x 1O�’ mol/L (indicated by stars. P < .01. Student’s t
test). In parallel wells. neutrophils migrated 69.5 ± 2.5 �zm towardio� mol/L N-formyl norleucyl leucyl phenylalanine.
bated without activation of the neutrophib respiratory burst.
There are other examples of dissociation of these responses.
At low concentrations (l0� to 10_b mol/L), fMLP pro-
duces a pattern of cell activation like that seen with
PMN7C3.’7’8 Similarly, arachidonate metabolites such as
beukotriene B4 stimulate changes in cellular calcium pools
and chemotaxis but are weak activators of the respiratory
burst . ‘ 5.1922 Conversely, phorbob myristate acetate (PMA),
which binds to protein kinase C,23 strongly stimulates the
respiratory burst without a rise in (Ca2�)1.’7’24 Unlike the
formybated peptides, PMN7C3 does not stimulate superox-
ide or hydrogen peroxide production even when used at high
concentration. In fact, pretreatment with PMN7C3 inhibits
the respiratory burst of neutrophils stimulated with serum-
opsonized zymosan.’2 Taken together, these facts suggest
that activation events leading to cell movement and adhesion
can be uncoupled from those leading to the respiratory burst.
Such independent pathways would provide a mechanism by
which neutrophibs could be activated to migrate to sites of
inflammation prior to generation of toxic mediators of the
inflammatory response. The relationship of PMN7C3-
mediated activation to potential transduction events such as
N.t Ch.motaxls (mIcrons)
30
25
20
15
10
0
Fig 4. Chemotaxis by neutrophils toward PMN7C3 and controlanti-neutrophil IgG (31 D8. Ml /70). Each value represents themean ± SEM of data from n = 6 filters from two independentexperiments. Random migration has been subtracted. Stars mdi-cate values that differed significantly (P < .025. Student’s t test)from the negative control (heat-inactivated serum). Monoclonalantibodies were used at 6.7 x iO� mol/L, formylated peptide atio� mol/L. and zymosan-activated serum at a 1 :5 dilution.
leukotriene generation and phosphoinositide turnover is cur-
rently under investigation.
Pretreatment with PMN7C3 inhibited subsequent chemo-
tactic response to both ZAS and fNLLP. Exposure to
formylated peptides (l0� to lO_6 mol/L) has a similar effect
on neutrophil response.’4”6 Fehr and Dahinden have sug-
gested that such “deactivation” may be the result of
increased adherence of stimulated neutrophils to their sub-
strate with inhibition of subsequent migration.25 Neutrophil
aggregation may have a similar nonspecific effect. Although
we cannot rule out such possibilities, they seem unlikely since
the checkerboard analysis (Table 2) indicates that PMN7C3
is itself chemokinetic. Furthermore, PMN7C3-treated cells
release normal amounts of superoxide in response to formy-
lated peptide,’ suggesting that functionally competent recep-
tors remain accessible on the cell surface. Thus, PMN7C3
can both stimulate and deactivate chemotactic response
without parallel effects on fMLP-induced respiratory burst.
As noted above, PMN7C3 binds to an oligosaccharide
antigen which is associated with the C3bi receptor and the
related glycoproteins LFA-I and p150,95 on the neutrophil
plasma membrane.2 An attractive hypothesis is that the
functional effects of PMN7C3 are mediated by the C3bi
Table 2. Chemokinetic and Chemotactic Activity of PMN7C3
Concentrationof PMN7C3 in
Lower Compartment(�ig/mL)
Concentration of PMN7C3 in Upper Compartment (�g/mL)
0 1 10 50 100
50110
100
73.7 ± 4.048.3 ± 1.265.5 ± 2.5
24.0 ± 1.1 23.0 ± 2.66.0 9.7 ± 1.8
56 5
74.0 64.0 ± 0.0 54.5±2. 0 2
Migration was measured as the distance (microns) from the top of the filter to the last focal plane containing two neutrophils. PMN7C3 was added in
the indicated concentrations to the lower (chemoattractant) or (upper polymorphonucleer) leukocytes (PMNs) wells of the chemotaxis assembly. Data
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom
Net Ch.motaxis (MIcrons) Hit Chimotaxls (microns)
80 100
I A
I-
60
40
20
0-
80
60
40
I I 10 100
Conc.ntration PUP47C3 (nil)
Nit Chimotoxis (Microns)
250
200 �
150 -
100 �
�:�
I 000
I000
1-I--I500 100 10
Conc.ntration fNLLP (nM/L)
other antigen-bearing structures. Another monocbonal anti-
body recognizing an antigen on CR3 (M1/70) did not mimic
the effect of PMN7C3. This does not rule out the possibility
that PMN7C3 is affecting cell function through the C3bi
receptor, since monocbonab antibodies binding to different
sites on receptors can vary in their functional effects.’3’33
Moreover, PMN7C3 stimulates cellular responses (adhe-
sion, motility) which are abnormal in neutrophils backing
C3bi receptors3�38 and which are inhibited by several mono-
clonal antibodies binding to the a-subunit of the receptor
3.33.3739 Recently, another monocbonal antibody
(WEM-Gl) binding to the same oligosaccharide antigen as
PMN7C3 has been demonstrated to stimulate antibody-
dependent cytotoxicity of neutrophils toward tumor targets,
another funciton that is abnormal in CR3/LFA-l-deficient
leukocytes.37’40’4’
Whether or not PMN7C3 is activating neutrophils
through the C3bi receptor and related glycoproteins, our
data suggest that this monocbonal antibody may be a useful
probe with which to study the cellular events which bead to
neutrophil movement and aggregation or to activation of the
respiratory burst.
ACKNOWLEDGMENT
We thank Drs William Nauseef, Robert Clark, and Richard
Diamond for helpful suggestions.
REFERENCES
sufficient stimulus for respiratory burst activation in human poly-morphonuclear leukocytes. Biochem Biophys Res Commun 132:674,
1985
4. Boyum A: Isolation of mononuclear cells and granulocytesfrom human blood: Isolation of mononuclear cells by one centrifuga-
tion and of granulocytes by combining centrifugation and sedimen-tation at 1g. Scand J Clin Lab Invest 21:77, 1967 (suppl 19)
NEUTROPHIL ACTIVATION BY ANTIBODY PMN7C3 1393
5. Labezari P, Bernard GE: An isobogous antigen-antibody reac-
100% 50% 20% 10% 5%
Concentration (ZAS)
Fig 6. Dose-response curve demonstrating the effect of pre-
treatment of neutrophils with increasing concentrations of
D PMN7C3 on chemotaxis toward � mol/L of fNLLP. Chemotaxis� was assayed by migration under agarose; each point represents
the mean ± SEM of values from 1 8 wells from three independent
experiments. Chemotaxis was significantly inhibited (P < .001 ) at
each concentration of PMN7C3 tested.
Fig 5. Effect of pretreatment of neutrophils with 6.6 x i0�mol/L PMN7C3 on chemotaxis toward (A) zymosan-activatedserum or (B) N-formyl norleucyl leucyl phenylalanine. Polymorpho-nuclear leukocytes (PMNs) were exposed to monoclonal antibodyfor 1 5 minutes at 4 ‘C and washed to remove unbound lgG prior to
assay by migration under agarose. Chemotaxis by PMN7C3-treated neutrophils (hatched bars) was significantly less than thatby buffer-treated neutrophils (open bars) at each concentration ofchemoattractant tested (P < .001 . Student’s t test). Data are themean ± SEM of n = 1 8 wells from three independent experi-ments.
receptor, which has been postulated to play an important role
in PMN-target adhesion.2�3’ Activation of another receptor
(insulin) by a polycbonal antiserum has been demonstrated.32
Our data do not definitively address this question, however.
The antigen recognized by PMN7C3 is shared by other
cellular proteins and glycolipids.2 We cannot rule out that
the effects of PMN7C3 are due to modulation of one of these
I . Nauseef WM, Root RK, Newman SL, Malech HL: Inhibition
of zymosan activation of human neutrophil oxidative metabolism bya mouse monoclonal antibody. Blood 62:635, 1983
2. Melnick DA, Nauseef WM, Markowitz SD, Gardner JP,Malech HL: Biochemical analysis and subcellular localization of aneutrophil-specific antigen, PMN-7, involved in the respiratory
burst. J Immunol 134:3346, 19853. Apfeldorf WJ, Melnick DA, Meshulam T, Rasmussen H,
Malech HL: A transient rise in intracellular free calcium is not a
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom

1394 MELNICK ET AL
24. Sha’afi RI, White JR. Molski TFP, Shefcyk J, Volpi M,Naccache PH, Feinstein MB: Phorbol 12-myristate 1 3-acetate
tion with human neutrophils, rebated to neonatal neutropenia. J Clin
lnvest45:174l, 1966
6. Lalezari P. Radel E: Neutrophil-specific antigens: Immunol-
ogy and clinical significance. Semin Hematob 11:281, 19747. Falk W, Goodwin RH, Leonard EJ: A 48-well chemotaxis
assembly for rapid and accurate measurement of leukocyte migra-tion. J Immunol Methods 33:239, 1980
8. Zigmond SH, Hirsch JG: Leukocyte locomotion and chemo-taxis. J Exp Med 137:387, 1973
9. Sobel JD, Galbin JI: Polymorphonuclear leukocyte and mono-cyte chemoattractants produced by human fibroblasts. J Clin Invest
63:609, 197910. Nelson RD. Quie PG. Simmons RL: Chemotaxis under
agarose: A new and simple method for measuring chemotaxis andspontaneous migration of human polymorphonuclear leukocytes and
monocytes.J Immunol 115:1650, 1975
I I . Nelson RD. Fiegel VD, Simmons RL: Chemotaxis of human
polymorphonuclea r neutrophils under agarose: Morphologic
changes associated with the chemotactic response. J Immunol
117:1676, 1976
12. BelIer DI, Springer TA, Schreiber RD: Anti-Mac-I selec-tively inhibits the mouse and human type three complement recep-
tor. J Exp Med 156:1000, 1982I 3. Sanchez-Madrid F, Simon P, Thompson 5, Springer TA:
Mapping of antigenic and functional epitopes on the alpha and betasubunits of two related mouse glycoproteins involved in cell interac-tions, LFA-I and Mac-I. J Exp Med 158:586, 1983
14. Ward PA, Becker EL: The deactivation of rabbit neutrophils
by chemotactic factor and the nature of the activatabbe esterase. JExp Med 127:693, 1968
I 5. Sha’afi RI, Molski TFP, Borgeat P. Naccache PH: Deactiva-
tion of the effects of f-Met-Leu-Phe and leukotriene B4 on calcium
mobilization in rabbit neutrophils. Biochem Biophys Res Commun
103:766, 1981
16. Donabedian H, Gallin JI: Deactivation of human neutrophilchemotaxis by chemoattractants: Effect on receptors for the chemo-tactic factor f-met-leu-phe. J Immunol 127:839, 1981
17. Korchak HM, Vienne K, Rutherford LE, Wilkenfeld C,
Finkelstein MC, Weissmann G: Stimulus response coupling in thehuman neutrophil II. Temporal analysis of changes in cytosoliccalcium and calcium effiux. J Biol Chem 259:4076, 1984
18. Showell JH, Freer RJ, Zigmond SH, Schiffman E, Aswani-kumar 5, Corcoran B, Becker EL: The structure-activity relations of
synthetic peptides as chemotactic factors and inducers of lysosomalenzyme secretion for neutrophils. J Exp Med 143:1154, 1976
19. Ford-Hutchinson AW, Bray MA, Doig MV, Shipley ME,
Smith MJH: Leukotriene B, a potent chemokinetic and aggregatingsubstance released from polymorphonuclear leukocytes. Nature286:264, 1980
20. Naccache PH, Molski TFP, Becker EL, Borgeat P. Vallerand
P, Sha’afi RI: Specificity of the effect of bipoxygenase metabolites of
arachidonic acid on calcium homeostasis in neutrophils. J Biol Chem
257:8608, 198221. Sha’afi RI, Naccache PH, Molski TFP, Borgeat P. Goetzl,
EJ: Cellular regulatory role of leukotriene B4: Its effect on cationhomeostasis in rabbit neutrophils. J Cell Physiob 108:401, 1981
22. Shak 5, Perez HD, Goldstein IM: A novel dioxygenationproduct of arachidonic acid possesses potent chemotactic activity for
human polymorphonuclear beukocytes. J Biol Chem 258:14948,
I983
23. Niedel JE, Kuhn U, Vandenbark GR: Phorbol diester recep-
tor copurifies with protein kinase C. Proc NatI Aced Sci USA 80:36,1983
activates rabbit neutrophils without an apparent rise in the level ofintracellular free calcium. Biochem Biophys Res Commun 1 14:638,1983
25. Fehr J, Dahinden C: Modulating influence of chemotacticfactor-induced cell adhesiveness on granulocyte function. J ClinInvest 64:8, 1979
26. Lay WH, Nussenzweig V: Receptors for complement onleukocytes. J Exp Med 128:991, 1968
27. Ehlenberger AG, Nussenzweig V: The role of membrane
receptors for C3b and C3d in phagocytosis. J Exp Med 145:357,I 977
28. Mantovani B: Different robes of IgG and complement recep-
tors in phagocytosis by polymorphonuclear leukocytes. J Immunol115:15, 1975
29. Newman SL, Johnston RB Jr: Role of binding through C3band IgG in polymorphonuclear neutrophil function: Studies withtrypsin-generated C3b. J Immunol I 23: 1 839, 1979
30. Ross GD, Lambris JD: Identification of a C3bi-specific
membrane complement receptor that is expressed on lymphocytes,monocytes, neutrophibs, and erythrocytes. J Exp Med 155:96, 1982
31 . Sanchez-Madrid F, Nagy JA, Robbins E, Simon P, SpringerTA: A human leukocyte differentiation antigen with distinct alpha
subunits and a common beta subunit: The lymphocyte function-
associated antigen (LFA- 1), the C3bi complement receptor(OKMI/Mac-l), and the p150/95 molecule. J Exp Med 158:1785,I983
32. Simpson IA and Hedo JA: Insulin receptor phosphorybation
may not be a prerequisite for acute insulin action. Science 223:130 1,I 984
33. Wright SD, Rao PE, Van Voorhis WC, Craigmyle LS, lidaK, Table MA, Westberg EF, Goldstein G, Silverstein SC: Identifica-tion of the C3bi receptor of human monocytes and macrophages by
using monocbonal antibodies. Proc NatI Acad Sci USA 80:5699,I983
34. Arnaout MA, Pitt J, Cohen HJ, Melamed J, Rosen FS,
Colten HR: Deficiency of a granulocyte-membrane glycoprotein(gpl 50) in a boy with recurrent bacterial infections. N Engb J Med306:693, 1982
35. Dana N, Todd RF, Pitt J, Springer TA, Arnaout MA:Deficiency of a surface membrane glycoprotein (Mol ) in man. JClin Invest 73:153, 1984
36. Arnaout MA, Spits H, Terhorst C, Pitt J, Todd RF: Defi-ciency of a leukocyte surface glycoprotein (LFA-l) in two patientswith Mol deficiency: Effects of cell activation on Mol/LFA-lsurface expression in normal and deficient leukocytes. J Clin Invest
74:1291, 198437. Kohl 5, Springer TA, Schmalstieg FC, Loo LS, Anderson
DC: Defective natural killer cytotoxicity and polymorphonuclearleukocyte antibody-dependent cellular cytotoxicity in patients withLFA-l/OKM-l deficiency. J Immunol 133:2972, 1984
38. Springer TA, Thompson WS, Miller U, Schmalsteig FC,Anderson DC: Inherited deficiency of the MAC-I, LFA-l, p150,95glycoprotein family and its molecular basis. J Exp Med 160:1901,I 984
39. Arnaout MA, Todd RF, Dana N, Melamed J, Schlossman
SF, Cobten HR: Inhibition of phagocytosis of complement C3- orimmunoglobubin G-coated particles and of C3bi binding by mono-
cbonal antibodies to a monocyte-granubocyte membrane glycopro-tein. J Clin Invest 72:171, 1983
40. Lopez AF, Vadas MA: Stimulation of human granulocytefunction by monocbonal antibody WEM-GI. Proc NatI Acad Sci
USA 81:1818, 198441 . Spitalnik PF, Spitalnik SL, Lopez AF, Vadas MA: Mono-
cbonal antibody WEM-GI stimulates granulocyte function andrecognizes a carbohydrate antigen. Clin Res 33:354A, 1985 (abstr)
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom

1986 67: 1388-1394
DA Melnick, T Meshulam, A Manto and HL Malech respiratory burstmovement and adhesion can be triggered independently from the Activation of human neutrophils by monoclonal antibody PMN7C3: cell
http://www.bloodjournal.org/content/67/5/1388.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of
For personal use only.on November 16, 2018. by guest www.bloodjournal.orgFrom





























