Embed Size (px)
Citation preview

INTRODUCTION
The interaction of cells with the extracellular matrix isimportant in regulating cell growth, differentiation, migrationand survival (Hynes, 1990). Integrins are a major class oftransmembrane receptors that mediate cell adhesion tofibronectin (Hynes, 1992; Pytela et al., 1985) as well as to otherextracellular matrix proteins (Hynes, 1992; Ruoslahti, 1988;Yamada, 1989). Binding of extracellular matrix proteins tointegrins leads to the generation of intracellular signals, manyof which are similar to intracellular signals generated bygrowth factor stimulation (Assoian, 1997; Juliano, 1996;Schwartz, 1997). Integrin-extracellular matrix interactions arealso important for cell survival, as disruption of integrin-mediated attachment to the extracellular matrix inducesapoptosis in epithelial and endothelial cells (Frisch and
Francis, 1994; Meredith et al., 1993; Ruoslahti and Reed,1994). Integrins and integrin-dependent signalling events alsoregulate the deposition of fibronectin into the extracellularmatrix (Giancotti and Ruoslahti, 1990; Wu et al., 1993; Wu etal., 1998).
Fibronectin also binds to cell surface proteoglycans,including syndecans (Carey, 1997; Saunders and Bernfield,1988) and CD44 (Naor et al., 1997). Binding of fibronectin tosyndecan involves the interaction of fibronectin’s carboxyl-terminal heparin-binding domain with heparan sulfateglycosaminoglycan chains on syndecan (Saunders andBernfield, 1988). Much data have suggested that the heparin-binding domain of fibronectin contributes to cell spreading andstress fiber formation in a variety of cell types (Bloom et al.,1999; Izzard et al., 1986; Woods et al., 1986). The addition ofheparin-binding fragments of fibronectin in solution or
4287Journal of Cell Science 113, 4287-4299 (2000)Printed in Great Britain © The Company of Biologists Limited 2000JCS1610
Many aspects of cell behavior are regulated by cell-extracellular matrix interactions, including cell migrationand cell growth. We previously showed that the addition ofsoluble fibronectin to collagen-adherent fibronectin-nullcells enhances cell growth. This growth-promoting effect offibronectin depended upon the deposition of fibronectininto the extracellular matrix; occupancy and clustering offibronectin-binding integrins was not sufficient to triggerenhanced cell growth. To determine whether the bindingof integrins to fibronectin’s RGD site is required forfibronectin-enhanced cell growth, the ability of fibronectinlacking the integrin-binding RGD site (FN∆RGD) topromote cell growth was tested. FN∆RGD promoted cellgrowth when used as an adhesive substrate or when addedin solution to collagen-adherent fibronectin-null cells.Addition of FN∆RGD to collagen-adherent fibronectin-nullcells resulted in a 1.6-1.8× increase in cell growth incomparison with cells grown in the absence of fibronectin.The growth-promoting effects of FN∆RGD and wild-typefibronectin were blocked by inhibitors of fibronectinpolymerization, including the anti-fibronectin antibody,L8. In addition, FN∆RGD-induced cell growth was
completely inhibited by the addition of heparin, and waspartially blocked by either heparitinase-treatment or byaddition of recombinant fibronectin heparin-bindingdomain. Heparin and heparitinase-treatment also partiallyblocked the growth-promoting effects of wild-typefibronectin, as well as the deposition of wild-typefibronectin into the extracellular matrix. These datasuggest that cell surface heparan-sulfate proteoglycanscontribute to the growth-promoting effects of FN∆RGDand wild-type fibronectin. Addition of heparin, treatmentwith heparitinase, or incubation with monoclonal antibodyL8 all inhibited the formation of short linear FN∆RGDfibrils on the cell surface. Inhibitory β1 integrin antibodieshad no effect on FN∆RGD fibril formation, FN ∆RGD-induced cell growth, or cell adhesion on FN∆RGD-coatedsubstrates. These data suggest that fibronectin fibrilformation can promote cell growth by a novel mechanismthat is independent of RGD-integrin binding, and thatinvolves cell surface proteoglycans.
Key words: Fibronectin, Extracellular matrix, Cell growth
SUMMARY
Fibronectin polymerization stimulates cell growth by RGD-dependent and
-independent mechanisms
Jane Sottile 1,*, Denise C. Hocking 2 and Kurt J. Langenbach 3,‡
1Department of Medicine, Center for Cardiovascular Research and 2Department of Pharmacology and Physiology, University ofRochester Medical Center, 601 Elmwood Ave, Box 679, Rochester, NY 14642, USA 3Department of Physiology and Cell Biology, Albany Medical College, Albany, NY 12208, USA*Author for correspondence (e-mail: [email protected])‡Present address: Department of Neurology and Neurological Sciences, Stanford University School of Medicine, Stanford, CA 94305, USA
Accepted 26 September; published on WWW 7 November 2000

4288
adsorbed to the substrate can restore stress fiber formation incells adherent to the cell-binding fragment of fibronectin(Bloom et al., 1999; Woods et al., 1986). In some systems, therequirement for the heparin-binding domain of fibronectincan be bypassed by treatment of cells with activators ofprotein kinase C (PKC) (Woods and Couchman, 1992) or Rho(Saoncella et al., 1999). These data suggest that cell surfaceproteoglycans cooperate with integrins in mediating maximalcell adhesion and spreading through PKC- and/or Rho-dependent mechanisms.
Our recent data demonstrate that deposition of fibronectininto the extracellular matrix stimulates adhesion-dependentcell growth, and that ligation and clustering of fibronectin-binding integrins are not sufficient to promote enhanced cellgrowth (Sottile et al., 1998). These data are consistent withother studies showing that inhibition of fibronectin deposition,or disruption of a preformed fibronectin matrix, decreases cellproliferation (Bourdoulous et al., 1998; Clark et al., 1997;Mercurius and Morla, 1998). Others have shown that cellgrowth in response to fibronectin can also be regulated by thethree-dimensional organization of fibronectin fibrils within thematrix (Sechler and Schwarzbauer, 1998). Taken together,these data indicate that extracellular matrix fibronectin playsan important role in regulating adhesion-dependent cellgrowth.
We previously demonstrated that integrin ligation is notsufficient to promote fibronectin-dependent cell growth (Sottileet al., 1998). To determine whether RGD-integrin binding isrequired for fibronectin-induced cell growth, we examined theeffects of fibronectin lacking the RGD sequence (FN∆RGD)on the growth of fibronectin-null cells. Our data demonstratethat cells adhere, spread and grow on substrates coated withFN∆RGD, and that addition of FN∆RGD to substrate-adherentfibronectin-null cells results in a 1.6-1.8× increase in cellgrowth in comparison with cells grown in the absence offibronectin. FN∆RGD stimulated cell growth to approximately50-60% of the levels induced by wild-type fibronectin,indicating that both RGD-dependent and -independentmechanisms are involved in the cell growth response tofibronectin. The ability of FN∆RGD to enhance growth ofcollagen-adherent cells was completely inhibited by theaddition of heparin, and was partially blocked by heparitinasetreatment, or by addition of recombinant fibronectin heparin-binding domain. These treatments also blocked the formationof short linear FN∆RGD fibrils on the cell surface.Interestingly, our data also demonstrate that αv and β3 integrinantibodies inhibit cell adhesion to FN∆RGD. However, αv andβ3 integrin antibodies did not inhibit either FN∆RGD fibrilformation or the ability of FN∆RGD to induce growth ofcollagen-adherent cells. These data indicate that fibronectinfibril formation and fibronectin-induced cell growth can occurin the absence of RGD, and suggest a role for proteoglycansin the cell growth response to fibronectin polymerization.
MATERIALS AND METHODS
Immunological reagentsMonoclonal antibody L8, which recognizes an epitope in fibronectin’sI-9 and III-1 modules, was a generous gift of Dr Michael Chernousov(Penn State College of Medicine, Danville, PA, USA) (Chernousov et
al., 1991). Rabbit polyclonal antibody to α5 integrin cytoplasmicdomain (a gift of Dr Susan LaFlamme, Albany Medical College,Albany, NY, USA) and polyclonal anti-fibronectin antibody werepreviously described (LaFlamme et al., 1992; Sottile and Mosher,1993). Polyclonal anti-vinculin antibody was from Sigma (St Louis,MO, USA). FITC-phalloidin was purchased from Molecular Probes(Eugene, OR, USA). Monoclonal anti-integrin antibodies to α5, αv,β1 and β3 subunits, and control hamster IgG and IgM were purchasedfrom Pharmingen (San Diego, CA, USA).
ProteinsHuman fibronectin was purified from Cohn’s fractions I and 2 (agenerous gift from Dr Ken Ingham, American Red Cross, Bethesda,MD, USA) as previously described (Miekka et al., 1982). Rat plasmafibronectin was purified on columns of gelatin-Sepharose. Full-lengthrecombinant rat fibronectin, and fibronectin lacking the RGD site(FN∆RGD) were expressed in insect cells and purified from insectcell conditioned medium as described (Hocking et al., 2000).Production and purification of recombinant rat 70 kDa and 40 kDafibronectin fragments have been previously described (Sottile andMosher, 1997). Recombinant III12-13 was produced in bacteria andpurified as described (Hocking et al., 1999).
Cell cultureMouse embryo cells were derived from fibronectin-null embryos andadapted to grow under serum-free conditions in defined medium (a1:1 mixture of Cellgro (Mediatech, Herndon, VA, USA) and Aim V(Life Technologies, Gaithersburg, MD, USA) as described (Sottile etal., 1998). These media do not require serum supplementation. Thus,the cells are cultured under conditions where no exogenous source offibronectin or other extracellular matrix proteins is present. The insectcell line IPLB-SF-21, adapted to grow in the serum-free mediumSF900-II, was obtained from Life Technologies (Gaithersburg, MD,USA). SF21 cells do not produce any detectable endogenousfibronectin. Thus, recombinant proteins produced by these cells do notcontain any contaminating fibronectin.
ImmunofluorescenceFibronectin-null cells were plated onto 18 mm glass coverslipsprecoated with vitronectin (5 µg/ml), recombinant fibronectin (10µg/ml) or FN∆RGD (10 µg/ml). Cells were seeded in definedmedium, and incubated at 37°C for various lengths of time. Definedmedium contains 1.5 mg/ml albumin, which would effectively blockany remaining protein binding sites on the dishes. After allowing cellsto attach and spread for 6 hours, some wells were supplemented withfibronectin, FN∆RGD or heparin. Cells were then fixed withparaformaldehyde, permeabilized with 0.5% Triton X-100, andincubated with the indicated primary antibodies for 60 minutes. Afterwashing, cells were incubated with fluorescein isothiocyanate (FITC)-or Texas Red-conjugated antibodies for 30 minutes. Followingwashing, cells were mounted in glycerol gel (Sigma) and examinedusing an Olympus BX60 microscope equipped with epifluorescence.
Cycloheximide treatmentFibronectin-null cells were preincubated for 2 hours with 20 µg/mlcycloheximide (Sigma). Cells were then trypsinized, washed twicewith medium containing soybean trypsin inhibitor (Sigma) andcycloheximide, then replated onto vitronectin-coated 18mm glasscoverslips in defined medium in the presence of cycloheximide (20µg/ml). Control cells were cultured in the absence of cycloheximide.Cells were allowed to attach and spread for 1-3 hours before beingprocessed for immunofluorescence.
Cell adhesion assaysTissue culture dishes (96-well) were coated with wild-type fibronectinor FN∆RGD at 10 µg/ml in PBS at 4°C overnight. Collagen wascoated onto dishes in 0.02 N acetic acid at 50 µg/ml at 4°C overnight.
J. Sottile, D. C. Hocking and K. J. Langenbach

4289Cell growth regulation by fibronectin
Wells were washed with phosphate-buffered saline (PBS), thenblocked with 1% bovine serum albumin (BSA) in PBS for 60 minutesat 37°C. Cells were preincubated with anti-integrin or controlantibodies at 25-50 µg/ml, or with various fibronectin fragments for30 minutes at room temperature prior to plating at 5×105 cell/ml in0.1 ml of defined medium. The cells were allowed to attach for 30-60 minutes at 37°C. Cells were washed with PBS, then fixed with 1%paraformaldehyde for 30 minutes at room temperature. Cells werestained with 0.5% Crystal Violet, then air dried. The dye wassolubilized with 1% sodium dodecyl sulfate, and the absorbance at592 nm was determined using a Wallac Victor2 (Gaithersberg, MD,USA) plate reader. Background absorbance of protein-coated wells inthe absence of cells was subtracted from each data point.
Cell growth assays on fibronectin- and FN ∆RGD-coateddishesTissue culture dishes (24 well; Corning, Cambridge, MA, USA) werecoated with recombinant fibronectin or recombinant FN∆RGD in PBSat 2.5-20 µg/ml at 37°C overnight. Wells were washed with PBSbefore seeding cells at 0.5×104 cell/cm2 in 1 ml of defined medium.The cells were allowed to grow for various lengths of time at 37°C.Cells were washed with PBS, then fixed with 1% paraformaldehydefor 30 minutes at room temperature. Cells were stained with 0.5%Crystal Violet, and the absorbance determined on a spectrophotometeras described (Hocking et al., 1998; Sottile et al., 1998).
Cell growth assays on collagen-coated dishesCollagen was coated onto dishes in 0.02 N acetic acid at 50 µg/ml at4°C overnight. Fibronectin-null cells cultured on collagen-coateddishes have a doubling time of approximately 20 hours (Sottile et al.,1998). Collagen-coated wells were washed with PBS before seedingcells at 0.5×104 cell/cm2 in 1 ml of defined medium. 4-6 hours afterseeding, plasma fibronectin, recombinant wild-type fibronectin orrecombinant FN∆RGD (20 nM) were added to some of the wells. Forinhibition experiments, heparin, recombinant fibronectin fragments,purified L8 IgG or integrin antibodies were added at the time ofaddition of soluble fibronectin. Control wells received PBS,
nonimmune mouse IgG (Cappel), hamster IgG or IgM (Pharmingen)or recombinant 70 kDa amino-terminal or 40 kDa gelatin-bindingfragments of fibronectin. Cells were allowed to grow for the indicatedtimes, then assayed for cell number using Crystal Violet, as describedabove.
Heparitinase and chondroitinase treatmentFibronectin-null cells were seeded in defined medium onto collagen-coated dishes. Following a 4 hour incubation, wells were eithersupplemented with 0.005 U/ml heparitinase or 0.6 U/mlchondroitinase ABC (Seikagaku America, Falmouth, MA, USA)(Chen et al., 1996a; Minden et al., 1995; Shishido et al., 1995).Following a 2 hour incubation, some wells were supplemented with20 nM wild-type (WT) fibronectin or 20 nM FN∆RGD. Freshheparitinase or chondroitinase were added daily. Cells were allowedto grow for 3 days, then processed for immunofluorescencemicroscopy or assayed for cell number using Crystal Violet, asdescribed above.
RESULTS
Cell spreading and focal contact formation onFN∆RGDIdentifying the role of fibronectin and fibronectinpolymerization in regulating adhesion-dependent cell growthhas been complicated by the presence of fibronectin in theserum used to culture cells, and by the ability of most adherentcells to constitutively synthesize and deposit fibronectin intothe extracellular matrix. To circumvent these problems, weestablished fibronectin-null cell lines that do not makefibronectin, but are capable of polymerizing exogenouslyadded fibronectin (Sottile et al., 1998).
In many cell types, the α5β1 integrin plays a major role inmediating cell attachment to fibronectin. We previously
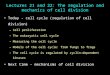
Fig. 1.Cell spreading on wild-type and ∆RGD fibronectin. Tissue culture dishes were precoated with 10 µg/ml of wild-type (WT) or ∆RGDfibronectin. Fibronectin-null cells were seeded onto coated wells in defined medium. 1 hour (A) or 3 hours (B) after seeding, the cells werefixed with 2.5% paraformaldehyde, permeabilized with 0.5% Triton X-100, then incubated with FITC-phalloidin (to detect actin) and a mousemonoclonal antibody to vinculin. After washing, cells were incubated with Texas Red-anti mouse antibody to detect vinculin. Actin (Panels1,3,5,7) and vinculin (Panels 2,4,6,8) staining was detected using an Olympus microscope equipped with epifluorescence. Photographs weretaken with a Spot digital camera. Bar, 10 µm.

4290
showed that fibronectin-null cells haveα5- and αv-containingintegrins on their cell surface, but no detectable cell surface α4integrins (Sottile et al., 1998). The major binding site infibronectin for α5β1 is the Arg-Gly-Asp (RGD) sequence thatis contained within the III-10 module (Pierschbacher andRuoslahti, 1984a; Pierschbacher and Ruoslahti, 1984b). Celladhesion to fibronectin can be disrupted by addition of RGDpeptides, demonstrating the importance of the RGD sequencein mediating cell adhesive events (D’Souza et al., 1991;Pierschbacher and Ruoslahti, 1984a). To determine whetherregions of fibronectin other than the RGD sequence are able topromote cell attachment and growth, recombinant fibronectinthat lacks the RGD sequence (FN∆RGD) was coated ontotissue culture dishes, and the ability of fibronectin-null cells toattach, spread and grow was monitored. As shown in Fig. 1,cells attached, spread and formed vinculin-containing focalcontacts (top panels) on FN∆RGD-coated substrates. Cellspreading and stress fiber formation on FN∆RGD was delayedin comparison with spreading on wild-type fibronectin(compare Fig. 1A,B). However, by 3 hours, cells seeded onFN∆RGD were well spread and contained prominent stressfibers (Fig. 1, Panel 5).
Others have shown that the presence of α5β1 in focalcontacts depends upon ligand occupancy (LaFlamme et al.,1992). The α5β1 integrin is not present in focal contacts offibroblasts (LaFlamme et al., 1992) or fibronectin-null cells(Sottile et al., 1998) adherent to surfaces coated withextracellular matrix proteins other than fibronectin, butredistributes to focal contacts following addition of RGD-containing fibronectin fragments or peptides (LaFlamme et al.,
1992; Sottile et al., 1998). Although RGD is the major bindingsite in fibronectin for α5β1 (Pierschbacher and Ruoslahti,1984a; Pierschbacher and Ruoslahti, 1984b), α5β1 has alsobeen shown to interact with the amino-terminal 70 kDa portionof fibronectin (Hocking et al., 1998). Therefore, we askedwhether adhesion to FN∆RGD resulted in the clustering ofα5β1 in focal contacts in an RGD-independent manner. Asshown in Fig. 2, α5-integrin was not detected in focal contactsof cells seeded on FN∆RGD (Fig. 2F,H). As expected, cellsadherent to wild-type fibronectin contained α5 integrin in theirfocal contacts (Fig. 2B,D).
Since cell spreading was delayed on FN∆RGD incomparison with spreading on wild-type fibronectin (Fig. 1),we next tested whether cell spreading on FN∆RGD wasmediated by production of cell-derived adhesive moleculesother than fibronectin. Fibronectin-null cells cultured in thepresence of the protein synthesis inhibitor, cycloheximide,spread (Fig. 2C,D,G,H) and formed vinculin-containing focalcontacts (Fig. 2C,G) when seeded onto FN∆RGD (Fig. 2G,H)or wild-type (Fig. 2C,D) fibronectin. Cells did not spread onuncoated tissue culture wells in the absence (Fig. 2I,J) orpresence (not shown) of cycloheximide. Taken together, thesedata indicate that cell spreading on FN∆RGD does not requirethe production of cell-derived adhesive factors, and thatfibronectin sequences other than the integrin-binding RGD siteare able to mediate cell spreading and focal contact formation.
Cell adhesion to FN ∆RGD is αv-integrin dependentThe α5β1 integrin has been shown to bind to the 70 kDa aminoterminal region of fibronectin (Hocking et al., 1998).
J. Sottile, D. C. Hocking and K. J. Langenbach
Fig. 2. Ability of FN∆RGD to promote cell spreading and focal contact formation. Fibronectin-null cells were seeded in defined medium ontouncoated coverslips (I,J) or onto coverslips coated with wild-type fibronectin (WT) or FN∆RGD in the absence (−CHX: A,B,E,F,I,J) orpresence (+CHX: C,D,G,H) of cycloheximide. 3 hours after seeding, cells were fixed then incubated with a polyclonal antibody toα5 integrinand a monoclonal antibody to vinculin, followed by a Texas Red-conjugated anti-rabbit antibody to visualizeα5 (B,D,F,H, J) and a FITC-anti-mouse antibody to visualize vinculin (A,C,E,G,I). Few cells attached to uncoated coverslips; to visualize cells that did attach, the cells shown inI and J were not washed prior to fixation. Cells were examined using an Olympus microscope equipped with epifluorescence, and photographedwith a Spot digital camera. Bar, 10 µm.

4291Cell growth regulation by fibronectin
Therefore, to determine whether adhesion of fibronectin-nullcells to FN∆RGD is mediated by 70 kDa-integrin interactions,we tested whether addition of the 70 kDa fragment inhibitedcell adhesion to FN∆RGD. As shown in Fig. 3, the 70 kDaamino-terminal fragment and the control 40 kDa gelatin-binding fragment of fibronectin did not inhibit adhesion toFN∆RGD or wild-type fibronectin. To determine whether celladhesion to FN∆RGD is integrin mediated, we tested theability of integrin antibodies to block adhesion to FN∆RGD.As shown in Fig. 3A, inhibitory α5 and β1 integrin antibodiesdid not block adhesion to FN∆RGD. In contrast, adhesion tocollagen I-coated dishes was blocked by β1 antibodies (notshown). Surprisingly, β3 integrin antibodies partially inhibited(40%) cell adhesion to FN∆RGD (Fig. 3A), but had little effecton adhesion to wild-type fibronectin (Fig. 3B). Whenantibodies to αv and β3 integrins were added together, >70%of adhesion to FN∆RGD was inhibited (Fig. 3A). Controlantibodies had no effect on cell adhesion to wild-typefibronectin or FN∆RGD. These data indicate that in theabsence of RGD-integrin binding, αv- and β3-containingintegrins mediate cell adhesion to fibronectin.
Cell growth in response to FN ∆RGDCell spreading is critical for cell cycle progression (Chen et al.,1997; Hansen et al., 1994; Huang et al., 1998). However,spreading by itself is not sufficient to promote cell growth(Davey et al., 1999). Therefore, to determine whetherFN∆RGD was able to promote cell growth in addition to cellspreading, fibronectin-null cells were seeded onto tissueculture plates coated with increasing concentrations ofFN∆RGD or wild-type fibronectin. As shown in Fig. 4, cellsgrown on dishes coated with FN∆RGD achieved cell densitiesapproximately 60% of those observed with cells seeded onwild-type fibronectin. This cell density is similar to thoseachieved by fibronectin-null cells adherent to type I collagen(Sottile et al., 1998). Maximal cell growth on both fibronectin
and FN∆RGD occurred at a coating concentration ofapproximately 10 µg/ml.
Adhesion and growth of cells on FN∆RGD-coated substrateswas not blocked by function-blocking anti-β1 integrinantibodies (Fig. 5), indicating that cell growth in response toFN∆RGD is β1 integrin-independent. In contrast, cell growthon collagen I-coated substrates was drastically reduced by β1antibodies. The effect of β3 antibodies on cell growth couldnot be assessed, since β3 antibodies inhibit cell adhesion toFN∆RGD (Fig. 3A).
We previously showed that addition of soluble fibronectin tocollagen, fibronectin, or laminin-adherent fibronectin-null cells
0 20 40 60 80 100 120
40K
70K
IgG+IgM
α5
β3+ αv
β1+β3
β3
β1
αv
∆RGD
A
0 20 40 60 80 100 120
40K
70K
IgG+IgM
β1+β3+ αv
β3
β1
αv
α5
WT FN
B
% Control Adhesion
Fig. 3. Effect of integrin antibodies on cell adhesion to fibronectin and ∆RGD fibronectin. Fibronectin-null cells were incubated with antibodies(25 µg/ml) or with the 70 kDa or 40 kDa fragments (1.42 µM) of fibronectin for 30 minutes at room temperature prior to seeding onto tissueculture dishes precoated with 10 µg/ml wild-type fibronectin (WT FN) or FN∆RGD (∆RGD). Cells were allowed to attach for 30 (WT) or 60minutes (∆RGD) and were then washed, fixed with 1% paraformaldehyde, then stained with 0.5% Crystal Violet. The absorbance wasdetermined using a plate reader as described in Materials and Methods. Similar results were seen when the adhesion assays on FN∆RGD weredone for 30 minutes. Data are presented as percentage adhesion of cells plated in the absence of competitor proteins. Values are means ± s.e.m.of triplicate determinations.
Fig. 4.Comparison of cell growth on wild-type and ∆RGDfibronectin-coated dishes. Tissue culture dishes were precoated withvarious amounts of wild-type (WT, s) or ∆RGD (d) fibronectin.Fibronectin-null cells were seeded onto coated wells in definedmedium. Cells were incubated for 4 days, then washed, fixed with1% paraformaldehyde, stained with 0.5% Crystal Violet and theabsorbance at 540 nm determined. Values are means of duplicatedeterminations, and error bars the range.
0
0.5
1
1.5
2
2.5
3
Abs
orba
nce
(540
nm)
0 5 10 15 20 25
WT
∆RGD
µg/ml

4292
resulted in a 2-5× increase in cell growth (Sottile et al., 1998).This fibronectin-enhanced cell growth depended upondeposition of fibronectin into the extracellular matrix (Sottileet al., 1998). Fibronectin deposition is a cell-mediated process(McDonald, 1988; McKeown-Longo and Mosher, 1983) in
which binding of fibronectin to cell surface receptors triggershomophilic binding interactions between fibronectin molecules(Aguirre et al., 1994; Chernousov et al., 1991; Hocking et al.,1994; Morla and Ruoslahti, 1992). To determine whetherfibronectin can enhance cell growth of collagen-adherent cellsin the absence of RGD-integrin interactions, collagen-adherent
J. Sottile, D. C. Hocking and K. J. Langenbach
0
20
40
60
80
100
120
% C
ontr
ol G
row
th
Control IgM
Collagen
∆RGD
β1
Fig. 5.Effect of β1 integrin antibodies on cell growth. Fibronectin-null cells in defined medium were seeded onto dishes precoated withcollagen type I (white bars) or FN∆RGD (black bars). After allowingcells to attach and spread for 2 (FN∆RGD) or 6 (collagen) hours, thecell culture medium was supplemented with β1 integrin or controlIgM antibodies at 25 µg/ml. Cells were allowed to grow for 4 daysand were then processed as described in Fig. 4. Data are presented aspercentage growth of cells incubated in the absence of antibodies.Values are means of duplicate determinations, and error bars therange.
00.5
11.5
22.5
33.5
44.5
Abs
orba
nce
(540
nm)
20 40 60 80 100 120
Time, hours
pFN
WTFN
FN∆RGD
-FN
Fig. 6. Effect of FN∆RGD on cell growth. Fibronectin-null cellswere seeded on collagen-coated wells in defined medium. 6 hoursafter seeding, cells were supplemented with 20 nM human plasmafibronectin (pFN, d), wild-type recombinant fibronectin (WTFN,s), recombinant FN∆RGD (h) or were given an equivalent volumeof PBS (−FN, j). Various times after seeding, cells were processedas described in Fig. 4. Values are means of duplicate determinations,and error bars the range.
Fig. 7. Deposition of wild-type and FN∆RGD into the extracellular matrix. Fibronectin-null cells were seeded in defined medium ontovitronectin-coated dishes. Following a 6 hour incubation, wells were either supplemented with 20 nM fibronectin (FN; A-D) or FN∆RGD (E-H). At the time of fibronectin addition, some wells were supplemented with the 350 nM 70 kDa amino-terminal fragment of fibronectin (70K)or the 40 kDa gelatin-binding fragment of fibronectin (40K). Following a 3 day incubation, cells were fixed, then permeabilized with 0.5%Triton-X-100. Cells were incubated with a polyclonal antibody to fibronectin followed by incubation with fluorescein-conjugated goat anti-rabbit IgG (A,C-E,G,H). Cells were examined using an Olympus microscope equipped with epifluorescence. (B,F) The corresponding phasepictures to A and E, respectively. Bar, 10 µm.

4293Cell growth regulation by fibronectin
fibronectin-null cells were incubated in the presence ofFN∆RGD, and cell growth was monitored over the course of4 days. Fig. 6 demonstrates that addition of FN∆RGD tofibronectin-null cells resulted in a 1.8× increase in cell growthover that observed in cells adherent to collagen alone. Wild-type recombinant fibronectin and plasma fibronectin increasedcell growth 3.5 and 4×, respectively, over that observed in cellsadherent to collagen alone (Fig. 6). These data indicate thatfibronectin can stimulate the growth of collagen-adherent cellsin the absence of RGD-integrin interactions.
Monoclonal antibody L8 inhibits cell surface stainingof FN∆RGD and inhibits FN ∆RGD-induced growthPrevious studies have shown that fibronectin lacking the RGDsite is impaired in its ability to form fibronectin fibrils; in CHOcells, short linear fibrils formed at the periphery of cells, mostlyin areas of cell-cell contact (Sechler et al., 1996). Addition ofFN∆RGD to fibronectin-null cells results in the formation ofshort stitch-like fibrils on the cell surface (Fig. 7E). In contrast,cells incubated with wild-type fibronectin (Fig. 7A) elaboratean extensive fibronectin matrix. Formation of FN∆RGD fibrilswas blocked by agents known to inhibit formation of wild-typefibronectin matrix, including the 70 kDa amino-terminalfragment of fibronectin (Fig. 7G) and the anti-fibronectinantibody, L8 (see below).
The III-1 module of fibronectin is thought to be involved infibronectin-fibronectin interactions that are important duringfibronectin fibrillogenesis (Chernousov et al., 1991; Hockinget al., 1994; Morla and Ruoslahti, 1992; Sechler et al., 1996).
Previous studies have shown that antibodies that bind to III-1,such as 9D2 and L8, inhibit fibronectin polymerization(Chernousov et al., 1987; Chernousov et al., 1991). Theinhibition of wild-type fibronectin polymerization by 9D2blocks the ability of fibronectin to promote cell growth (Sottileet al., 1998). To determine whether III-1 plays a role in theformation of FN∆RGD fibrils and in the ability of FN∆RGDto enhance cell growth, we asked whether these events couldbe blocked by monoclonal antibody L8. As shown in Fig. 8,addition of L8 to fibronectin-null cells blocked the depositionof FN∆RGD (Fig. 8E) and wild-type fibronectin (Fig. 8B) onthe cell surface, as detected by indirect immunofluorescencemicroscopy; control IgG had no effect (Fig. 8C,F). Addition ofL8 did not result in any change in cell morphology (not shown).To determine whether L8 could block the ability of FN∆RGDto promote cell growth, collagen-adherent fibronectin-null cellswere incubated with FN∆RGD in the presence or absence ofL8. As shown in Fig. 9, addition of L8 blocked >80% ofFN∆RGD-induced cell growth. Addition of L8 resulted in asimilar inhibition of wild-type fibronectin-induced cell growth(Fig. 9). Taken together, these data indicate that theorganization of FN∆RGD into short linear fibrils depends uponthe 70 kDa amino-terminal domain, as well as III-1-fibronectininteractions, and thus occurs by a mechanism similar to thatused to polymerize wild-type fibronectin. These data alsoindicate that III-1-fibronectin interactions are critical for thecell growth response to FN∆RGD.
Integrins are known to participate in fibronectin fibrilformation (Akiyama et al., 1989; Fogerty et al., 1990; Giancotti
Fig. 8. Effect of L8 on fibronectin and FN∆RGD deposition. Fibronectin-null cells were seeded in defined medium onto vitronectin-coateddishes. Following a 6 hour incubation, wells were either supplemented with 20 nM wild-type fibronectin (WT; A-C) or FN∆RGD (D-F). At thetime of fibronectin addition, some wells were also supplemented with 50 µg/ml monoclonal antibody L8 (B,E) or mouse IgG (C,F). Controlwells received an equal volume of PBS (A,D). Following a 3 day incubation, cells were fixed, then permeabilized with 0.5% Triton-X-100.Cells were incubated with a polyclonal antibody to fibronectin followed by incubation with fluorescein-conjugated goat anti-rabbit IgG. Cellswere examined using an Olympus microscope equipped with epifluorescence to visualize fibronectin and FN∆RGD. Bar, 10 µm.

4294
and Ruoslahti, 1990; Zhang et al., 1993), and antibodies to α5and β1 integrins partially inhibit fibronectin polymerization
(Akiyama et al., 1989; Fogerty et al., 1990). To compare theeffects of anti-integrin antibodies on fibronectin and FN∆RGDfibril formation, and fibronectin- and FN∆RGD-inducedgrowth, function-blocking β1 and β3 antibodies were added tofibronectin-null cells that were incubated with either FN∆RGDor with non-mutant fibronectin. β1 and β3 antibodies did notprevent the deposition of FN∆RGD on the cell surface (Fig.10B). Similarly, addition of αv and β3 antibodies did notinhibit FN∆RGD fibril formation (not shown). In contrast,fibril formation of non-mutant fibronectin was partiallyblocked by the presence of β1 and β3 antibodies, resulting inthe formation of shorter, less extensive fibrils (Fig. 10E). Todetermine whether β3 integrin antibodies inhibit cell growth,we asked whether FN∆RGD-induced growth on collagen-coated dishes was blocked by addition of β3 antibodies. Asshown in Fig. 11, β3 integrin antibodies partially blocked cellgrowth both in the absence (29%) and presence (22%) ofFN∆RGD. However, the relative increase in cell growthinduced by FN∆RGD was unchanged by β3 integrin antibodytreatment (without anti-β3=1.7×; with anti-β3=1.8×). Nofurther decrease in cell growth was observed when αv and β3integrin antibodies were added together (not shown).
Heparin and heparitinase-treatment inhibit FN ∆RGDinduced cell growthAntibodies to β3 integrins did not prevent enhanced cellgrowth induced by FN∆RGD (Fig. 11). These data suggest thatbinding of FN∆RGD to a non-integrin receptor may contributeto the cell growth response. Since fibronectin is known to bindto proteoglycans, we asked whether heparin could block the
J. Sottile, D. C. Hocking and K. J. Langenbach
1
1.25
1.5
1.75
2
2.25
2.5
Fol
d In
crea
se
- +L8 +IgG - +L8
∆RGDWT + + +
+ +- -
- - -
Fig. 9. Effect of L8 on fibronectin- and FN∆RGD-induced growth.Fibronectin-null cells were seeded in defined medium onto collagen-coated dishes. Following a 6 hour incubation, wells weresupplemented with either 20 nM wild-type fibronectin (WT) orFN∆RGD. At the time of fibronectin addition, some wells were alsosupplemented with 50 µg/ml monoclonal antibody L8 or mouse IgG.Following a 5 day incubation, cells were fixed with 1%paraformaldehyde, then stained with 0.5% Crystal Violet and theabsorbance at 540 nm determined. Data is expressed as fold increasein growth; growth of control wells (cells incubated in the absence offibronectin) was set equal to 1. Values are means of duplicatedeterminations, and error bars the range.
Fig. 10. Effect of integrin antibodies on fibronectin and FN∆RGD deposition. Fibronectin-null cells were seeded in defined medium ontovitronectin-coated dishes. The day after seeding, wells were supplemented with 20 nM nonmutant fibronectin (D-F) or FN∆RGD (A-C). At thetime of fibronectin addition, some wells were also supplemented with β1 and β3 (25 µg/ml each) antibodies (B,E) or with control IgG and IgMantibodies (25 µg/ml each; C,F). Control wells were supplemented with an equal volume of PBS (A,D). Following a 4 day incubation, cellswere fixed, then permeabilized with 0.5% Triton-X-100. Cells were incubated with a polyclonal antibody to fibronectin followed by incubationwith fluorescein-conjugated goat anti-rabbit IgG. Cells were examined using an Olympus microscope equipped with epifluorescence tovisualize fibronectin or FN∆RGD. Bar, 10 µm.

4295Cell growth regulation by fibronectin
ability of FN∆RGD to promote growth. As shown in Fig. 12A,the growth-promoting effects of FN∆RGD were completelyabolished by the addition of 1 mg/ml heparin (H1), and werepartially (45%) inhibited by addition of the recombinantfibronectin heparin-binding domain containing modulesIII 12,13 (HBD). The addition of heparin (1 mg/ml) tofibronectin-null cells also decreased growth induced by wild-
type fibronectin by 45% (Fig. 12A). Lower doses of heparin(10 µg/ml) partially inhibited growth induced by FN∆RGD(70%) and wild-type fibronectin (30%). Heparin (1 mg/ml) hadlittle effect (<15% decrease) on cell growth in the absence ofadded fibronectin (data not shown). The ability of heparin andheparin-binding fragments of fibronectin to inhibit cell growthsuggests that fibronectin-proteoglycan binding may be acritical component of fibronectin- and FN∆RGD-inducedgrowth.
To further explore the possibility that cell surfaceproteoglycans may be involved in fibronectin-induced cellgrowth, we examined the effect of heparitinase andchondroitinase treatment on fibronectin-induced cell growth.As shown in Fig. 12B, heparitinase treatment (+H) offibronectin-null cells attenuated the growth-promoting effectsof both wild-type fibronectin and FN∆RGD, causing a 43%reduction in the growth-promoting effects of FN∆RGD and a34% decrease in the growth-promoting effects of wild-typefibronectin. Chondroitinase treatment (+C) had no effect onFN∆RGD or fibronectin induced cell growth (Fig. 12B). Takentogether, these data support a role for cell surface heparansulfate proteoglycans in mediating the growth-promotingeffects of fibronectin.
Cell surface staining of FN ∆RGD is attenuated byheparin and heparitinase treatmentMany studies have shown that clustering of cell surfacereceptors, including growth factor receptors and integrins, isnecessary for transducing downstream signalling events(Akiyama et al., 1994; Clark and Brugge, 1995; Heldin, 1995;Miyamoto et al., 1995; Weiss and Schlessinger, 1998). Todetermine whether the heparin-binding activity of fibronectinis important for the clustering of FN∆RGD into fibrils on thecell surface, we asked whether FN∆RGD fibril formation could
0
50
100
150
200
250
300%
Con
trol
Gro
wth
None IgG
+FN
+∆RGD
None
β3
Fig. 11. Effect of β3 integrin antibodies on fibronectin- andFN∆RGD-induced cell growth. Fibronectin-null cells were seeded oncollagen-coated wells in defined medium. 6 hours after seeding, cellswere supplemented with 20 nM rat plasma fibronectin (+FN, hatchedboxes), FN∆RGD (white boxes) or were given an equivalent volumeof PBS (none; black boxes). At the time of fibronectin addition, somewells were also supplemented with β3 integrin antibodies (25 µg/ml)or with control IgG (25 µg/ml). 3 days after seeding, cells wereprocessed as described in Fig. 4. Growth of cells incubated in theabsence of antibodies and in the absence of added fibronectin was setequal to 100%. Values are means of duplicate determinations, anderror bars the range.
A
∆RGD
WT
1.0
1.2
1.4
1.6
1.8
2.0
Fol
d In
crea
se
- H.01 H1 HBD - H.01 H1 HBD
+ + + ++-
-- +- - -
- - + +
1.0
1.5
2.0
2.5
3.0
- +H +C - +H +C
+ + ++ +- - - +- - -
B
Fig. 12. Effect of heparin and heparitinase on fibronectin- and FN∆RGD-induced cell growth. (A) Fibronectin-null cells were seeded in definedmedium onto collagen-coated dishes. Following a 6 hour incubation, wells were either supplemented with 10 nM wild-type fibronectin (WT) orFN∆RGD. At the time of fibronectin addition, some wells were also supplemented with 0.01 (H.01) or 1 mg/ml heparin (H1), 0.5 µM III 12,13(HBD), or with an equivalent volume of PBS (−). Cells were allowed to grow for 5 days, then processed as described in Fig. 4. (B) Fibronectin-null cells were seeded onto collagen-coated dishes and allowed to attach for 4 hours. Wells were then either supplemented with 0.005 U/mlheparitinase (H) or 0.6 U/ml chondroitinase ABC (C). Following a 2 hour incubation, some wells were supplemented with 20 nM wild-type(WT) fibronectin or 20 nM FN∆RGD. Fresh heparitinase or chondroitinase were added daily. Cells were allowed to grow for 3 days, thenprocessed as described in Fig. 4. Data are expressed as fold increase in growth; growth of control wells (cells incubated in the absence offibronectin) was set equal to 1. Values are means of duplicate determinations, and error bars the range.

4296
be blocked by either the addition of heparin or by heparitinasetreatment (Fig. 13). FN∆RGD staining was eliminated whencells were incubated in the presence of heparin (Fig. 13G), orwhen cells were treated with heparitinase (Fig. 13I). Bothtreatments also attenuated fibril formation in cells incubatedwith wild-type fibronectin (Fig. 13B,D). Addition of heparinor treatment with heparitinase did not cause any noticeabledecrease in cell attachment or spreading (not shown). Inaddition, chondroitinase treatment did not affect the depositionof either wild-type fibronectin (Fig. 13E) or FN∆RGD (Fig.13J). These data suggest that heparan sulfate proteoglycansmay participate in FN∆RGD fibril formation on the cellsurface.
DISCUSSION
We previously demonstrated that fibronectin polymerizationinto the extracellular matrix promotes adhesion-dependentgrowth, and that integrin-ligation and clustering are notsufficient to promote enhanced cell growth in response tofibronectin (Sottile et al., 1998). In this study, we haveextended these findings by demonstrating that fibronectin canstimulate cell growth in the absence of RGD-integrin ligationby a mechanism that remains dependent on fibronectinpolymerization. FN∆RGD promoted cell growth toapproximately 50-60% of the levels induced by wild-typefibronectin (Figs 4, 6). Thus, it is likely that both RGD-dependent and -independent mechanisms are involved in thecell growth response to wild-type fibronectin.
We previously showed that fibronectin-null cells do notexpress α4 integrins on their cell surface (Sottile et al., 1998).
In addition, all of the remaining fibronectin-binding integrins,including those that contain αv (αvβ1, αvβ3, αvβ5, αvβ6)have been shown to interact with fibronectin via the RGD sitein III-10 (Chen et al., 1996b; Plow et al., 2000). Therefore, itwas surprising to find that cell adhesion to FN∆RGD could beblocked with antibodies to αv and β3 integrins (Fig. 3). To ourknowledge, this is the first demonstration that binding offibronectin to αv-containing integrins can occur independentlyof the RGD sequence. αvβ3 binds to a surprisingly largenumber of molecules, by both RGD-dependent andindependent mechanisms (Plow et al., 2000). In addition,binding of fibronectin to the platelet receptor, αIIbβ3, canoccur by RGD-dependent and −independent mechanisms(Bowditch et al., 1991). The site in fibronectin that mediatescell attachment to αv-containing integrins in the absence ofRGD is not currently known.
Our previous data indicate that the ability of fibronectin toenhance cell growth depends upon its ability to becomedeposited into the extracellular matrix (Sottile et al., 1998).FN∆RGD does not polymerize into robust fibrils in theextracellular matrix of fibronectin-null cells, but forms shortstitch-like fibrils on the cell surface (Fig. 7). Others have shownthat FN∆RGD can be induced to form fibrils by an α4β1-dependent mechanism following integrin activation (Sechler etal., 2000). The FN∆RGD fibrils reported here are generated bya distinct mechanism, since fibronectin-null cells do not havecell surface α4 integrins (Sottile et al., 1998). The short linearFN∆RGD fibrils are abolished by the exogenous addition ofheparin, and by heparitinase-treatment, which also inhibitedthe growth-promoting effects of both FN∆RGD and wild-typefibronectin (Figs 12, 13). These data suggest that cell surfaceheparan sulfate proteoglycans may play an important role in
J. Sottile, D. C. Hocking and K. J. Langenbach
Fig. 13. Effect of heparin and heparitinase on fibronectin and FN∆RGD deposition. Fibronectin-null cells were seeded in defined medium ontovitronectin-coated dishes. Following a 6 hour incubation, wells were either supplemented with 20 nM wild-type fibronectin (A,B) or FN∆RGD(F,G) in the presence (+Hep; B,G) or absence (−Hep; A,F) of 1 mg/ml heparin. For cells treated with heparitinase and condroitinase, wells wereeither untreated (C,H), or supplemented with 0.005 U/ml heparitinase (D,I) or 0.6 U/ml chondroitinase ABC (E,J) 4 hours after seeding.Following a 2 hour incubation, some wells were supplemented with 20 nM wild-type (WT) fibronectin (C-E) or 20 nM FN∆RGD (H-J).Control wells received an equivalent volume of PBS. Fresh heparitinase or chondroitinase were added daily. Following a 3 day incubation, cellswere fixed, then permeabilized with 0.5% Triton-X-100. Cells were incubated with a polyclonal antibody to fibronectin followed by afluorescein-conjugated goat anti-rabbit IgG. Cells were examined using an Olympus microscope equipped with epifluorescence. Bars, 10 µm.

4297Cell growth regulation by fibronectin
fibronectin fibril formation, as well as in mediatingfibronectin’s growth-promoting effects.
Interestingly, both the formation of FN∆RGD fibrils on thecell surface, and the ability of FN∆RGD to induce cell growthwere blocked by the presence of monoclonal antibody L8. L8recognizes an epitope in fibronectin’s I9 and III1 modules(Chernousov et al., 1991), and inhibits fibronectin depositioninto the extracellular matrix, presumably by blockingfibronectin-fibronectin interactions (Chernousov et al., 1987;Chernousov et al., 1991). It is possible that binding ofFN∆RGD to the cell surface exposes fibronectin-interactivesites within III-1, as has previously been proposed for wild-type fibronectin (Hocking et al., 1994; Morla and Ruoslahti,1992; Sechler et al., 1996). The ability of L8 to block bothFN∆RGD fibril formation and the growth-promoting effects ofFN∆RGD suggests that exposure of self-interactive sites thatlead to clustering of fibronectin into fibrils is a critical eventduring fibronectin-stimulated cell growth and can occur in theabsence of RGD-integrin interactions. The loss of FN∆RGDfibrils with heparitinase treatment suggests the possibility thatheparan sulfate proteoglycans may play a role in exposingfibronectin self-interactive sites on the cell surface.
Our data are consistent with a model in which fibronectincan trigger cell growth by two mechanisms: (1) by serving asan adhesive substate for cells and thus allowing for cellspreading and adhesion-dependent cell cycle progression;and (2) as a consequence of its polymerization into fibrils inthe extracellular matrix. αv-containing integrins maypromote cell growth via effects on cell adhesion andspreading, since cell adhesion to FN∆RGD is dependent uponαv and β3 integrins (Fig. 3). β3 integrin antibodies partiallyinhibited the growth of collagen-adherent cells in thepresence and absence of FN∆RGD. However, the relativeincrease in growth induced by FN∆RGD was not affected byantibody treatment (Fig. 11). The effect of β3 integrinantibodies on cell growth could be due to the ability of theseantibodies to partially block adhesion (not shown) and growth(Fig. 11) on collagen I in the absence of FN∆RGD. Incontrast, our data suggest that heparan sulfate proteoglycansparticipate in the cell growth response to fibronectin by amechanism that depends upon fibronectin deposition into theextracellular matrix. The ability of heparitinase to inhibitFN∆RGD cell surface staining and to attenuate FN∆RGD-induced growth suggests that binding of fibronectin toheparan sulfate proteoglycans contributes to the growth-promoting effects of FN∆RGD. It is likely that proteoglycansalso contribute to the growth-promoting effects of wild-typefibronectin, as treatment of cells with heparitinase, oraddition of soluble heparin decreased deposition of wild-typefibronectin into the extracellular matrix (Fig. 13) and partiallyinhibited fibronectin-induced cell growth (Fig. 12).
Several heparin-binding domains have been identified withinthe fibronectin molecule. Heparin-binding activity has beenlocalized to modules I1-5 (the 27 kDa amino-terminal region),III 1, and the carboxyl-terminal heparin-binding domaincontaining III12-14 (Hynes, 1990; Litvinovich et al., 1992).Previous studies have demonstrated a role for fibronectinheparin-binding domains in fibronectin deposition into theextracellular matrix. The amino-terminal 27-kDa fragment offibronectin is important for binding of soluble fibronectin tothe surface of substrate-attached cells (McKeown-Longo and
Mosher, 1985; Quade and McDonald, 1988), while thecarboxyl-terminal heparin-binding domain has been shown topartially inhibit fibronectin binding and deposition into theextracellular matrix (Bultmann et al., 1998). Our datademonstrate that recombinant III12,13 partially inhibitedFN∆RGD-induced cell growth (Fig. 12); the 70 kDa amino-terminal fragment (which contains modules I1-5) also partiallyblocks FN∆RGD-induced growth (data not shown). The abilityof III 12,13 and the 70 kDa fragment to inhibit FN∆RGD-induced growth could be due to the direct involvement of thesedomains in fibronectin-proteoglycan interactions.
Others have proposed that heparan sulfate proteoglycans actas coreceptors with integrins to promote maximal cellspreading on fibronectin (Saoncella et al., 1999; Woods et al.,1986). Our data demonstrate that fibronectin can promoteadhesion and focal contact formation in the absence offibronectin-RGD interactions, and that this adhesion dependsupon binding to αv- and β3-containing integrins. Our data alsosuggest an important role for fibronectin-proteoglycaninteractions in mediating FN∆RGD’s growth-promotingeffects, and in promoting the formation of FN∆RGD fibrils onthe cell surface. Although FN∆RGD can form short stitch-likefibrils on the cell surface, its inability to form extensive fibrilssuggests that exposure of additional fibronectin-fibronectininteractive sites critical for fibril growth may depend uponintegrin interactions with fibronectin’s RGD site. Our data alsosuggest that proteoglycans are important regulators of wild-type fibronectin function, since heparin and heparitinasetreatment attenuated both fibronectin deposition into theextracellular matrix and the growth-promoting effects of wild-type fibronectin. Fibronectin is known to interact with anumber of cell surface and cell-associated proteoglycans thathave been shown to regulate cell migration and cell growth,including syndecan, perlecan and CD44 (Carey, 1997; Mathiaket al., 1997; Naor et al., 1997). Our data suggest that onemechanism whereby proteoglycans affect cell growth may beby regulating fibronectin deposition into the extracellularmatrix. This hypothesis is supported by a recent study showingthat transfection of a mutant syndecan 2 into CHO cells caninhibit fibronectin deposition into the extracellular matrix(Klass et al., 2000). Together, these data suggest thatfibronectin fibril formation can promote cell growth by a novelmechanism that is independent of RGD-integrin binding, andthat involves cell surface proteoglycans.
This research was supported by grants HL50549 and HL03971 (toJ.S.), and HL60181 and HL64074 (to D.H.) from the NationalInstitutes of Health. K.L. was supported by National Institutes ofHealth Predoctoral Training Grant T32-HL07194. The authors thankDr Jean Schwartzbauer for providing fibronectin cDNAs, and for helpin expressing full length fibronectins; Drs Susan LaFlamme, DeaneMosher, and Michael Chernousov for providing antibodies; MsMichelle Arquiett for technical assistance; and Dr Susan LaFlammefor critically reading this manuscript.
REFERENCES
Aguirre, K. M., McCormick, R. J. and Schwarzbauer, J. E. (1994).Fibronectin self-association is mediated by complementary sites within theamino-terminal one-third of the molecule. J. Biol. Chem.269, 27863-27868.
Akiyama, S. K., Yamada, S. S., Chen, W. T. and Yamada, K. M.(1989).Analysis of fibronectin receptor function with monoclonal antibodies: Roles

4298
in cell adhesion, migration, matrix assembly, and cytoskeletal organization.J. Cell Biol.109, 863-875.
Akiyama, S. K., Yamada, S. S., Yamada, K. M. and LaFlamme, S. E.(1994). Transmembrane signal transduction by integrin cytoplasmicdomains expressed in single-subunit chimeras. J. Biol. Chem.269, 15961-15964.
Assoian, R. K. (1997). Anchorage-dependent cell cycle progression. J. CellBiol. 136, 1-4.
Bloom, L., Ingham, K. C. and Hynes, R. O.(1999). Fibronectin regulatesassembly of actin filaments and focal contacts in cultured cells via theheparin-binding site in repeat III13. Mol. Biol. Cell10, 1521-1536.
Bourdoulous, S., Orend, G., MacKenna, D. A., Pasqualini, R. andRuoslahti, E. (1998). Fibronectin matrix regulates activation of RHOand CDC42 GTPases and cell cycle progression. J. Cell Biol. 143, 267-276.
Bowditch, R. D., Halloran, C. E., Aota, S.-i., Obara, M., Plow, E. F.,Yamada, K. M. and Ginsberg, M. H. (1991). Integrin α11bβ3 (plateletGPIIb-IIIa) recognizes multiple sites in fibronectin. J. Biol. Chem. 266,23323-23328.
Bultmann, H., Santas, A. J. and Presciotta-Peters, D. M.(1998).Fibronectin fibrillogenesis involves the heparin II binding domain offibronectin. J. Biol. Chem.273, 2601-2609.
Carey, D. J. (1997). Syndecans: multifunctional cell-surface co-receptors.Biochem. J. 327, 1-16.
Chen, C. S., Mrksich, M., Huang, S., Whitesides, G. M. and Ingber, D. E.(1997). Geometric control of cell life and death. Science276, 1425-1428.
Chen, H., Strickland, D. K. and Mosher, D. F.(1996a). Metabolism ofthrombospondin 2. Binding and degradation by 3T3 cells andglycosaminoglycan-variant Chinese hamster ovary cells. J. Biol. Chem. 271,15993-15999.
Chen, J., Maeda, T., Sekiguchi, K. and Sheppard, D.(1996b). Distinctstructural requirements for interaction of the integrins α5β1, αvβ5, andαvβ6 with the central cell binding domain in fibronectin. Cell Adhes. Comm.4, 237-250.
Chernousov, M. A., Faerman, A. I., Frid, M. G., Pritseva, O. andKoteliansky, V. E. (1987). Monoclonal antibody to fibronectin whichinhibits extracellular matrix assembly. FEBS Lett. 217, 124-128.
Chernousov, M. A., Fogerty, F. J., Koteliansky, V. E. and Mosher, D. F.(1991). Role of the I-9 and III-1 modules of fibronectin in formation of anextracellular matrix. J. Biol. Chem. 266, 10851-10858.
Clark, E. A. and Brugge, J. S.(1995). Integrins and signal transductionpathways: the road taken. Science268, 233-239.
Clark, R. A. F., McCoy, G. A., Folkvord, J. M. and McPherson, J. M.(1997). TGF-β1 stimulates cultured human fibroblasts to proliferate andproduce tissue-like fibroplasia: A fibronectin matrix-dependent event. J.Cell. Physiol. 170, 69-80.
D’Souza, S. E., Ginsberg, M. H. and Plow, E. F.(1991). Arginyl-glycyl-aspartic acid (RGD): A cell adhesion motif. Trends Biochem. Sci. 16, 246-250.
Davey, G., Buzzai, M. and Assoian, R. K.(1999). Reduced expression ofα5β1 integrin prevents spreading-dependent cell proliferation. J. Cell Sci.112, 1663-1672.
Fogerty, F. J., Akiyama, S. K., Yamada, K. M. and Mosher, D. F.(1990).Inhibition of binding of fibronectin to matrix assembly sites by anti-integrin(α5β1) antibodies. J. Cell Biol. 111, 699-708.
Frisch, S. M. and Francis, H.(1994). Disruption of epithelial cell-matrixinteractions induces apoptosis. J. Cell Biol. 124, 619-626.
Giancotti, F. G. and Ruoslahti, E. (1990). Elevated levels of the α5β1fibronectin receptor suppress the transformed phenotype of Chinese hamsterovary cells. Cell 266, 8923-8931.
Hansen, L. K., Mooney, D. J., Vacanti, J. P. and Ingber, D. E.(1994). Integrinbinding and cell spreading on extracellular matrix act at different points incell cycle to promote hepatocyte growth. Mol. Biol. Cell5, 967-975.
Heldin, C.-H. (1995). Dimerization of cell surface receptors in signaltransduction. Cell 80, 213-223.
Hocking, D. C., Sottile, J. and Langenbach, K. J.(2000). Stimulation ofintegrin-mediated cell contractility by fibronectin polymerization. J. Biol.Chem. 275, 10673-10682.
Hocking, D. C., Sottile, J. and McKeown-Longo, P. J.(1994). Fibronectin’sIII-1 module contains a conformation-dependent binding site for the amino-terminal region of fibronectin. J. Biol. Chem. 269, 19183-19181.
Hocking, D. C., Sottile, J. and McKeown-Longo, P. J.(1998). Activation ofdistinct α5β1-mediated signaling pathways by fibronectin’s cell adhesionand matrix assembly domains. J. Cell Biol. 141, 241-253.
Hocking, D. C., Sottile, J., Reho, T., Fassler, R. and McKeown-Longo, P.J. (1999). Inhibition of fibronectin matrix assembly by the heparin-bindingdomain of vitronectin. J. Biol. Chem. 274, 27257-27264.
Huang, S., Chen, C. S. and Ingber, D. E.(1998). Control of cyclin D1,p27Kip1, and cell cycle progression in human capillary endothelial cells bycell shape and cytoskeletal tension. Mol. Biol. Cell9, 3179-3193.
Hynes, R. O.(1990). Fibronectins. New York: Springer-Verlag.Hynes, R. O.(1992). Integrins: Versatility, modulation, and signaling in cell
adhesion. Cell 69, 11-25.Izzard, C. S., Radinsky, R. and Culp, L. A.(1986). Substratum contacts and
cytoskeletal reorganization of BALB/c 3T3 cells on a cell-binding fragmentand heparin-binding fragments of plasma fibronectin. Exp. Cell Res. 165,320-336.
Juliano, R. (1996). Cooperation between soluble factors and integrin-mediated cell anchorage in the control of cell growth and differentiation.BioEssays18, 911-917.
Klass, C. M., Couchman, J. R. and Woods, A.(2000). Control ofextracellular matrix assembly by syndecan-2 proteoglycan. J. Cell Sci. 113,493-506.
LaFlamme, S. W., Akiyama, S. K. and Yamada, K. M.(1992). Regulationof fibronectin receptor distribution. J. Cell Biol. 117, 437-447.
Litvinovich, S. V., Novokhatny, V. V., Brew, S. A. and Ingham, K. C.(1992). Reversible unfolding of an isolated heparin and DNA bindingfragment, the first type III module from fibronectin. Biochim. Biophys.Acta 1119, 57-62.
Mathiak, M., Yenisey, C., Grant, D. S., Sharma, B. and Iozzo, R. V.(1997).A role for perlecan in the suppression of growth and invasion infibrosarcoma cells. Cancer Res. 57, 2130-2136.
McDonald, J. A. (1988). Extracellular matrix assembly. Ann. Rev. Cell Biol.4, 183-207.
McKeown-Longo, P. J. and Mosher, D. F.(1983). Binding of plasmafibronectin to cell layers of human skin fibroblasts. J. Cell Biol. 97, 466-472.
McKeown-Longo, P. J. and Mosher, D. F.(1985). Interaction of the 70,000-mol-wt amino-terminal fragment of fibronectin with the matrix-assemblyreceptor of fibroblasts. J. Cell Biol. 100, 364-374.
Mercurius, K. W. and Morla, A. O. (1998). Inhibition of vascular smoothmuscle cell growth by inhibition of fibronectin matrix assembly. Circ. Res.82, 548-556.
Meredith, J. E., Fazeli, B. and Schwartz, M. A.(1993). The extracellularmatrix as a cell survival factor. Mol. Biol. Cell4, 953-961.
Miekka, S. I., Ingham, K. C. and Menache, D.(1982). Rapid methods forisolation of human plasma fibronectin. Thromb. Res. 27, 1-14.
Minden, A., Lin, A., Claret, F.-X., Abo, A. and Karin, M. (1995). Selectiveactivation of the JNK signaling cascade and c-Jun transcriptional activity bythe small GTPases Rac and Cdc42Hs. Cell 81, 1147-1157.
Miyamoto, S., Akiyama, S. K. and Yamada, K. M.(1995). Synergistic rolesfor receptor occupancy and aggregation in integrin transmembrane function.Science267, 883-885.
Morla, A. and Ruoslahti, E. (1992). A fibronectin self-assembly site involvedin fibronectin matrix assembly: Reconstruction in a synthetic peptide. J. CellBiol. 118, 421.
Naor, D., Slonov, R. V. and Ish-Shalom, D.(1997). CD44: Structure,function, and association with the malignant process. Adv. Cancer Res. 71,241-319.
Pierschbacher, M. and Ruoslahti, E.(1984a). The cell attachment activityof fibronectin can be duplicated by small synthetic fragments of themolecule. Nature309, 30-33.
Pierschbacher, M. D. and Ruoslahti, E.(1984b). Variants of the cellrecognition site of fibronectin that retain attachment-promoting activity.Proc. Natl. Acad. Sci. USA81, 5985-5988.
Plow, E. F., Haas, T. A., Zhang, L., Loftus, J. and Smith, J. W.(2000).Ligand binding to integrins. J. Biol. Chem. 275, 21785-21788.
Pytela, R., Pierschbacher, M. D. and Ruoslahti, E.(1985). Identification andisolation of a 140 kD cell surface glycoprotein with properties of afibronectin receptor. Cell 40, 191-198.
Quade, B. J. and McDonald, J. A.(1988). Fibronectin’s amino-terminalmatrix assembly site is located within the 29 kDa amino terminal domaincontaining five type 1 repeats. J. Biol. Chem. 263, 19602-19609.
Ruoslahti, E. (1988). Fibronectin and its receptors. Ann. Rev. Biochem. 57,375-413.
Ruoslahti, E. and Reed, J. C.(1994). Anchorage dependence, integrins, andapotosis. Cell 77, 477-478.
Saoncella, S., Echtermeyer, F., Denhez, F., Nowlen, J. K., Mosher, D. F.,
J. Sottile, D. C. Hocking and K. J. Langenbach

4299Cell growth regulation by fibronectin
Robinson, S. D., Hynes, R. O. and Goetinck, P. F.(1999). Syndecan-4signals cooperatively with integrins in a Rho-dependent manner in theassembly of focal adhesions and actin stress fibers. Proc. Natl. Acad. Sci.USA96, 2805-2810.
Saunders, S. and Bernfield, M.(1988). Cell surface proteoglycan bindsmouse mammary epithelial cells to fibronectin and behaves as a receptor forinterstitial matrix. J. Cell Biol. 106, 423-430.
Schwartz, M. A. (1997). Integrins, oncogenes, and anchorage independence.J. Cell Biol. 139, 575-578.
Sechler, J. L., Cumiskey, A. M., Gazzola, D. M. and Schwarzbauer, J. E.(2000). A novel RGD-independent fibronectin assembly pathway initiatedby α4β1 integrin binding to the alternatively spliced V region. J. Cell Sci.113, 1491-1498.
Sechler, J. L. and Schwarzbauer, J. E.(1998). Control of cell cycleprogression by fibronectin matrix architecture. J. Biol. Chem273, 25533-25536.
Sechler, J. L., Takada, Y. and Schwarzbauer, J. E.(1996). Altered rate offibronectin matrix assembly by deletion of the first type III repeats. J. CellBiol. 134, 573-583.
Shishido, Y., Sharma, K. D., Higashiyama, S., Klagsbrun, M. and Mekada,E. (1995). Heparin-like molecules on the cell surface potentiate binding ofdiphtheria toxin to the diphtheria toxin receptor/membrane-anchoredheparin-binding epidermal growth factor-like growth factor. J. Biol. Chem.270, 29578-29585.
Sottile, J., Hocking, D. C. and Swiatek, P.(1998). Fibronectin matrixassembly enhances adhesion-dependent cell growth. J. Cell Sci. 111, 2933-2943.
Sottile, J. and Mosher, D. F.(1993). Assembly of fibronectin molecules withmutations or deletions of the carboxyl-terminal type I modules. Biochem.32, 1641-1647.
Sottile, J. and Mosher, D. F.(1997). N-terminal type I modules required forfibronectin binding to fibroblasts and to fibronectin’s III1 module. Biochem.J. 323, 61-60.
Weiss, A. and Schlessinger, J.(1998). Switching signals on or off by receptordimerization. Cell 94, 277-280.
Woods, A. and Couchman, J. R.(1992). Protein kinase C involvement infocal adhesion formation. J. Cell Sci. 101, 277-290.
Woods, A., Couchman, J. R., Johansson, S. and Hook, M.(1986). Adhesionand cytoskeletal organisation of fibroblasts in response to fibronectinfragments. EMBO J. 5, 665-670.
Wu, C., Bauer, J. S., Juliano, R. L. and McDonald, J. A.(1993). The α5β1integrin fibronectin receptor, but not the α5 cytoplasmic domain, functionsin an early and essential step in fibronectin matrix assembly. J. Biol. Chem.268, 21833-21888.
Wu, C., Keightley, S. Y., Leung-Hagesteijn, C., Radeva, G., Coppolino, M.,Goicoechea, S., McDonald, J. A. and Dedhar, S.(1998). Integrin-linkedprotein kinase regulates fibronectin matrix assembly, E-cadherin expressionand tumorigenicity. J. Biol. Chem. 273, 528-536.
Yamada, K. (1989). Fibronectin domains and receptors. In Fibronectin (ed.D. F. Mosher), pp. 47-121. New York: Academic Press.
Zhang, Z., Morla, A., Vuori, K., Bauer, J. S., Juliano, R. L. and Ruoslahti,E. (1993). The αvβ1 integrin functions as a fibronectin receptor but does notsupport fibronectin matrix assembly and cell migration on fibronectin. J.Cell Biol. 122, 235-242.