Embed Size (px)
Citation preview

Communication
cDNA cloning, gene organization and expression analysis ofhuman peptidylarginine deiminase type VI��
Jiayi Zhang, Jianliang Dai, Enpeng Zhao, Yun Lin, Li Zeng, Jinzhong Chen, Huari
Zheng, Yu Wang, Xin Li, Kang Ying�, Yi Xie and YuMin Mao�
State Key Laboratory of Genetic Engineering, School of Life Sciences, Fudan University,Shanghai, P. R. China
Received: 13 January, 2004; revised: 26 October, 2004; accepted: 26 November, 2004
Key words: peptidylarginine deiminase, ePAD, ovary, hPADVI
Peptidylarginine deiminase (PAD) catalyzes the post-translational modification ofprotein through the conversion of arginine to citrulline in the presence of calciumions. Human, similar to rodents, has four isoforms of PAD (type I, II, III and IV/V),each of which is distinct in substrate specificity and tissue specific expression. In ourlarge-scale sequencing project, we identified a new human PAD cDNA from a humanfetal brain cDNA library. The putative protein encoded by this cDNA is designatedhPADVI. Expression analysis of hPADVI showed that it is mainly expressed in adulthuman ovary and peripheral blood leukocytes. We conclude that hPADVI may beorthologous to mouse ePAD, basing on sequence comparison, chromosome localiza-tion and exon-intron structure analysis. PAD-mediated deimination of epithelial cellkeratin resulting in cytoskeletal remodeling suggests a possible role for hPADVI incytoskeletal reorganization in the egg and in early embryo development. This studydescribes a new important member of the human PAD family.
Vol. 51 No. 4/2004
1051–1058
QUARTERLY
�This work was supported by the National Project 863 of P. R. China (grant number 2003AA221020) andthe National Nature Science Foundation of P.R. China (grant number 30270519).�The nucleotide sequence reported in this paper has been submitted to GenBank under accession num-ber AY443100.
�Correspondence to: YuMin Mao: State Key Laboratory of Genetic Engineering, School of Life Sciences,Fudan University, Shanghai 200433, P. R. China; tel.: (86 21) 6564 2502; e-mail: [email protected];Kang Ying, tel.: (86 21) 6598 8366 6096; e-mail: [email protected]
Abbreviation: PAD, peptidylarginine deiminase.

Peptidylarginine deiminases (protein-L-argi-nine iminohydrolase, EC 3.5.5.15, PAD) are agroup of enzymes that convert peptide boundarginyl residues to citrullinyl residues in pro-teins (Rothnagel & Rogers, 1984). Enzymaticdeimination abolishes positive charges of na-tive protein molecules, inevitably causing sig-nificant alteration in their structure and func-tion (Lamensa & Moscarello, 1993; Imparl etal., 1995; Tarcsa et al., 1996). All the enzymesknown to date show absolute requirement forcalcium ion (Nakashima et al., 1999). Dei-mination of arginine residues of vimentin,desmin and glial fibrillary acidic protein(GFAP) by PAD interferes with the ability ofthese proteins to polymerize (Inagaki et al.,1989). Deimination of trichohyalin results inloss of secondary structure and such modifiedprotein is then more easily cross-linked by atransglutaminase (Tarcsa et al., 1996; 1997).Early research described four isoforms ofPADs in rodents (Ishigami et al., 1998). Theseisoforms displayed nearly identical amino-acid sequences, but different tissue-specificexpression (Ishigami et al., 2001). Recently,oocyte and early embryo abundant peptidyl-arginine deiminase-like protein, ePAD, hasbeen reported in mouse (Wright et al., 2003).Concerning human tissues, four types of PADhave been cloned, i.e., PAD type I (Guerrin etal., 2003), PADII (Ishigami et al., 2002),PADIII (Kanno et al., 2000), and PADIV/V(Guerrin et al., 2003). Human PADI mRNAswere detected by reverse transcriptase-PCRin various organs, including epidermis, testis,placenta, spleen and thymus (Guerrin et al.,2003). Human PADII mRNA was detected inthe epidermis, the type II enzyme was ex-pressed in all the living epidermal layers, sug-gesting that PADII is functionally importantduring terminal differentiation of epidermalkeratinocytes. Human PADIII is the predomi-nant isoform in hair follicles and may func-tion as a modulator of hair structural pro-teins, including trichohyalin during hair andhair follicle formation (Kanno et al., 2000).Human PADIV/V is present in human
myeloid leukemia HL-60 cells induced to dif-ferentiate into granulocytes by retinoic acidand later in peripheral blood granulocytes(Nakashima et al., 1999).Here we report a new gene, which encodes
PAD, whose transcript is detected mainly inthe ovary and peripheral blood leukocytes. Abioinformatic analysis suggests that it is anorhtologous gene to mouse ePAD.
MATERIAL AND METHODS
cDNA library construction. A cDNA li-brary was constructed in a modifiedpBluescript II SK (+) vector (Stratagene). Themodified vector was constructed by introduc-ing two SfiI recognition sites, i.e. SfiIA (5�gg-ccattatggcc 3�) and SfiIB (5�ggccgcctcggcc 3�)between the EcoRI and NotI sites ofpBluescript II SK (+). Fetal brain mRNA waspurchased from Clontech. Double-strandedcDNA was synthesized and inserted into pBSvector between the above sites usingSMARTTM cDNA Library Construction Kit(Clontech) following manufacturer’s instruc-tions. The cDNA inserts were sequenced onan ABIPRISMTM 377 DNA sequencer (Per-kin-Elmer) using the BigDye Terminator Cy-cle Sequencing Kit (Perkin-Elmer) with–21M13 primer, M13Rev primer and syn-thetic internal-walking primers designed ac-cording to the obtained cDNA sequence frag-ments. Each part of the insert was sequencedat least three times bidirectionally. Subse-quent editing and assembly of all the se-quences from one clone were performed us-ing Acembly (Sanger Center).Bioinformatic analysis. To verify the new
full length cDNAs, a database search was per-formed with the basic local alignment searchtools (BLAST) network service at NCBI(http://www.ncbi.nlm.nih.gov/BLAST). Pro-file scan and alignment were done athttp://www.expasy.org/pfscan. Other se-quence analysis was performed online.
1052 J. Zhang and others 2004

RT-PCR. To investigate the expression pat-tern of hPADVI in different tissues, a multi-ple tissue cDNA (MTC, Clontech) basedRT-PCR was employed. Panel I/II and Advan-tage 2 Kit (Clontech) were used in the reac-tion. The hPADVI specific primer pairs(hPADVIF: 5�cagcagcttttaccccagtgcagaggg3�
and hPADVIR: 5�tcttgcccatcacaatcatccgca-acag3�) were designed to amplify a 500 bpfragment. A glyceraldehyde-3-phosphate de-hydrogenase (G3PDH) control primer pair in-cluded in the panels was used to verify thenormalization of the MTC panel. The se-quences of the primers for amplifying G3PDHwere 5�tgaaggtcggagtcaacggatttggt3� (G3P-DHF) and 5�catgtgggccatgaggtccaccac3�
(G3PDHR). A total of 35 cycles of amplifica-tion was performed in a total volume of 50 �l.The cycling conditions were as follows: 5 minat 94�C, followed by 35 cycles of 95�C for 30 s,68�C for 60 s, 72�C for 5 min. HPADVI andG3PDH cDNAs were amplified in a parallelRT-PCR reaction. Five microlitters of eachproduct was later resolved on 1.5% agarosegels.
RESULTS
Sequence characterization
The nucleotide sequence and deduced amino-acid sequence of this gene are shown inFig. 1. An open reading frame encodes a pro-tein of 694 amino acids. The molecular massand isoelectric point of the predicted transla-tion product are calculated to be 77.4 kDa and5.02, respectively. Comparison of the se-
Vol. 51 Human peptidylarginine deiminase type VI 1053
Figure 1. Nucleotide and deduced amino-acid se-quence of human hPADVI (GenBank AccessionNo. AY443100).
The nucleotide sequence is shown in the top lines, andthe deduced amino-acid sequence below in thesinger-letter code. The ORF extends from nucleotide 52to 2136 and encodes a protein of 694 amino acids. Anasterisk represents the stop codon; at the 3� end thepossible polyadenylation signal (AATAAA) is boxed.

quence against the NCBI nonredundant data-base using the BLAST algorithm found thatthe sequence was 89% identical to a recentlysubmitted putative peptidylarginine deimi-nase protein sequence (XP_372767), whichwas predicted by the NCBI’s automated anno-tation tool GNOMON. The protein has 42%,43%, 41% and 42% homology to humanhPADI, hPADII, hPADIII, and hPADIV/V, re-spectively (Fig. 2A). It shares a higher
homology (65% identity) with mouse ePAD(Fig. 2B). We term this gene hPADVI follow-ing the HUGO Nomenclature Committee(http://www.gene.vcl.ac.vk/nomenclature).
Chromosomal localization
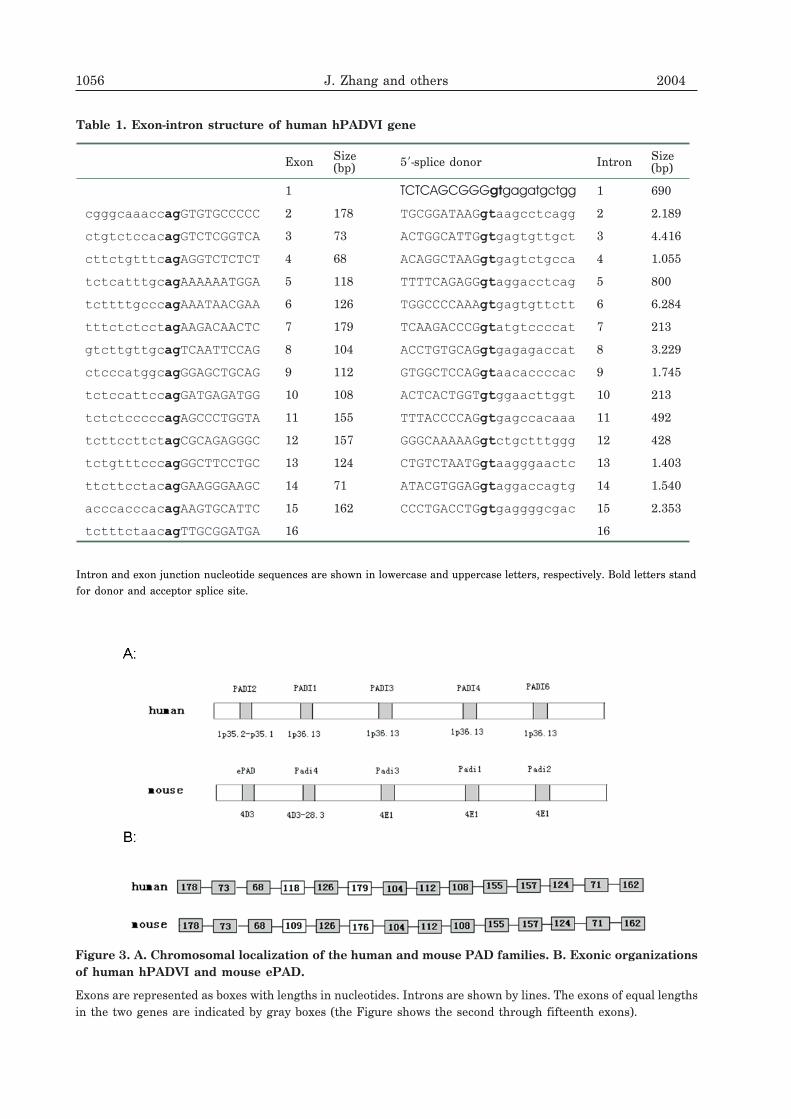
Using the international human genome data-base on NCBI, we found that hPADVI localizeson 1p36.13. The gene spans 28.8 kbp and con-
1054 J. Zhang and others 2004
Figure 2. A. Alignment of human PAD types I, II, III, IV/V, and VI.
The alignment was performed by the Align X program of vector NTI suite 5.5, and amino acids are shaded accord-ing to the degree of conservation using GeneDoc (http://www.cris.com/�Ketchup/genedoc.shtml): black (100%similarity); gray (80–90% similarity); light gray (60–70% similarity). The accession numbers of the sequence datacited for comparison have the following designations: PADI, AB033768; PADII, AB030176; PADIII, AB026831;PADIV/V, AB017919; PADVI, AY443100. (continued on next page)

sists of 16 exons. All sequences of theexon-intron junctions are consistent with theAG-GT rule (Table 1). hPADII, hPADI, hPADIIIand hPADIV/V link with hPADVI in tandem,and mouse PADII, PADI, PADIII, PADIV andePAD were linked in the same order (Fig. 3A).The lengths of the corresponding exons ofhPADVI and ePAD were equal (Fig. 3B).
Expression pattern of hPADVI
The tissue distribution of hPADVI mRNAwas determined by RT-PCR. The resultshowed that hPADVI was expressed mainly inthe ovary and peripheral blood leukocytes,and slightly expressed in the liver, thymus,
testis, lung and spleen of the 16 tissues exam-ined (Fig. 4).
DISCUSSION
In a large-scale cDNA sequence study, weisolated a 2397 bp cDNA that encodes humanpeptidylarginine deiminase type VI gene. ThecDNA containing an ORF from 52 to 2136 bpencodes a protein of 694 residues. The puta-tive initiation ATG codon at 52 bp (CTG-AGGATGG) conformed to the Kozak consen-sus sequence (A/GXXATGG) apparently con-trolling the translational efficiency of mam-malian mRNAs (Kozak, 1987). The poly-
Vol. 51 Human peptidylarginine deiminase type VI 1055
Figure 2. B. Alignment of human PADVI and mouse ePAD.
1056 J. Zhang and others 2004
Exon Size(bp) 5�-splice donor Intron Size
(bp)
1 TCTCAGCGGGgtgagatgctgg 1 690
cgggcaaaccagGTGTGCCCCC 2 178 TGCGGATAAGgtaagcctcagg 2 2.189
ctgtctccacagGTCTCGGTCA 3 73 ACTGGCATTGgtgagtgttgct 3 4.416
cttctgtttcagAGGTCTCTCT 4 68 ACAGGCTAAGgtgagtctgcca 4 1.055
tctcatttgcagAAAAAATGGA 5 118 TTTTCAGAGGgtaggacctcag 5 800
tcttttgcccagAAATAACGAA 6 126 TGGCCCCAAAgtgagtgttctt 6 6.284
tttctctcctagAAGACAACTC 7 179 TCAAGACCCGgtatgtccccat 7 213
gtcttgttgcagTCAATTCCAG 8 104 ACCTGTGCAGgtgagagaccat 8 3.229
ctcccatggcagGGAGCTGCAG 9 112 GTGGCTCCAGgtaacaccccac 9 1.745
tctccattccagGATGAGATGG 10 108 ACTCACTGGTgtggaacttggt 10 213
tctctcccccagAGCCCTGGTA 11 155 TTTACCCCAGgtgagccacaaa 11 492
tcttccttctagCGCAGAGGGC 12 157 GGGCAAAAAGgtctgctttggg 12 428
tctgtttcccagGGCTTCCTGC 13 124 CTGTCTAATGgtaagggaactc 13 1.403
ttcttcctacagGAAGGGAAGC 14 71 ATACGTGGAGgtaggaccagtg 14 1.540
acccacccacagAAGTGCATTC 15 162 CCCTGACCTGgtgaggggcgac 15 2.353
tctttctaacagTTGCGGATGA 16 16
Intron and exon junction nucleotide sequences are shown in lowercase and uppercase letters, respectively. Bold letters standfor donor and acceptor splice site.
Table 1. Exon-intron structure of human hPADVI gene
Figure 3. A. Chromosomal localization of the human and mouse PAD families. B. Exonic organizationsof human hPADVI and mouse ePAD.
Exons are represented as boxes with lengths in nucleotides. Introns are shown by lines. The exons of equal lengthsin the two genes are indicated by gray boxes (the Figure shows the second through fifteenth exons).

adenylation signature (AATAAA) is located at2372 bp.The putative protein shows 42%, 43%, 41%
and 42% identify to human hPADI, hPADII,hPADIII, and hPADIV/V, respectively. More-over, it shows a higher identity (65%) tomouse ePAD. The domain that is conservedbetween the known PADs is also conserved inthis protein, suggesting that the putative pro-tein represents a new member of the PAD en-zyme family. We have named this proteinhPADVI, in agreement with HUGO Nomen-clature Committee (http://www.gene.vcl.ac.vk/nomenclature).Alignment of hPADVI cDNA against NCBI
database revealed that the cDNA sequencecovered 28.8 kb. The hPADVI gene consists of16 exons along the human chromosome1P36.13, where the human PAD gene familyis located as a cluster. Interestingly, mouseePAD gene also consists of 16 exons alongmouse chromosome 4D3, where the mousePAD gene family presents the same pattern(Fig. 3A). We also found that hPADVI andePAD have the same number of amino-acidresidues coded for corresponding exons ex-cept for the fifth and seventh exons, andthese two exons do not change the open read-ing frame (Fig. 3B). The similar gene organi-zation of hPADVI and ePAD suggests thathPADVI is the counterpart of ePAD.Peptidylarginine deiminases (PADs) are
posttranslational modification enzymes thatconvert protein arginine to citrulline residuesin a calcium-dependent ion manner. In ro-dents and human, different isoforms of PADare distinct in substrate specificity and tissuespecific expression. The relatively high se-quence conservation in the C-terminal regionsuggests that this enzyme is involved in suchcommon physiological functions as catalysisand calcium binding. The N-terminal regionmight be involved in selective recognition oftarget proteins in relevant tissues (Fig. 2).The RT-PCR showed that hPADVI is mainlyexpressed in the ovary and peripheral bloodleukocytes. Mouse ePAD localizes to egg cyto-
plasmic sheets, a unique keratin-containingintermediate filament structure found only inmammalian oocytes and in early embryos,and known to undergo reorganization at criti-cal stages of development. The specific local-ization of ePAD to oocytes in ovarian sections
and its homology to a well-characterized en-zyme family that has known in vitro and invivo substrates supports further developmentof small molecule inhibitors of new potentialcontraceptive targets (Wright et al., 2003).hPADVI, the counterpart gene of mouseePAD might have a similar role in cyto-skeletal reorganization in the egg and earlyembryo. Further study should be made toclarify the precise role of hPADVI.
R E F E R E N C E S
Guerrin M, Ishigami A, Mechin MC. (2003)Biochem J.; 370: 167–74.
Imparl JM, Senshu T, Graves DJ. (1995) ArchBiochem Biophys.; 318: 370–7.
Inagaki M, Takahara H, Nishi Y, Sugawara K,Sato C. (1989) J Biol Chem.; 264: 18119–27.
Ishigami A, Kuramoto M, Yamada M,Watanabe K, Senshu T. (1998) FEBS Lett.;433: 113–8.
Ishigami A, Asaga H, Ohsawa T, Akiyama K,Maruyama N. (2001) Biomed Res.; 22: 63–5.
Vol. 51 Human peptidylarginine deiminase type VI 1057
Figure 4. Multiple tissue cDNA based RT-PCR ex-pression pattern of hPADVI.
Twenty-six cycles (for G3PDH) and 35 cycles (forhPADVI) were performed with Advantage 2 Kit(Clontech).

Ishigami A, Ohsawa T, Asaga H. (2002) ArchBiochem Biophys.; 407: 25–1.
Kanno T, Kawada A, Yamanouchi J, et al.(2000) J Invest Dermatol.; 115: 813–23.
Kozak M. (1987) Nucleic Acids Res.; 15: 812–8.
Lamensa JW, Moscarello MA. (1993) JNeurochem.; 61: 987–96.
Nakashima K, Hagiwara T, Ishigami A. (1999)J Biol Chem.; 274: 27786–2.
Rothnagel JA, Rogers GE. (1984) MethodsEnzymol.; 107: 624–1.
Tarcsa E, Marekov LN, Mei G, Melino G, LeeSC, Steinert PM. (1996) J Biol Chem.; 271:30709–6.
Tarcsa E, Marekov LN, Andreoli J, Idler WW,Candi E, Chung S-I, Steinert PM. (1997) JBiol Chem.; 272: 27893–1.
Wright PW, Bolling LC, Calvert ME. (2003) DevBiol.; 256: 73–8.
1058 J. Zhang and others 2004





























