Embed Size (px)
Citation preview

Copyright 0 1989 by the Genetics Society of America
cdc2 and the Regulation of Mitosis: Six Interacting mcs Genes
Lisa Molz, Robert Booher, Paul Young’ and David Beach Cold Spring Harbor Laboratory, Cold Spring Harbor, New York I I724
Manuscript received January 19, 1989 Accepted for publication May 4, 1989
ABSTRACT A cdc2-3w weel-50 double mutant of fission yeast displays a temperature-sensitive lethal phenotype
that is associated with gross abnormalities of chromosome segregation and has been termed mitotic catastrophe. In order to identify new genetic elements that might interact with the cdc2 protein kinase in the regulation of mitosis, we have isolated revertants of the lethal double mutant. The suppressor mutations define six mcs genes (rncs: mitotic catastrophe suppressor) that are not allelic to any of the following mitotic control genes: cdc2, weel, cdcl3, cdc25, sucl or niml. Each mcs mutation is recessive with respect to wild-type in its ability to suppress mitotic catastrophe. None confer a lethal phenotype as a single mutant but few of the mutants are expected to be nulls. A diverse range of genetic interactions between the mcs mutants and other mitotic regulators were uncovered, including the following examples. First, mcs2 cdc2w or mcs6 cdc2w double mutants display a cell cycle defect dependent on the specific wee allele of cdc2. Second, both mcsl cdc25-22 or mcs4 cdc25-22 double mutants are nonconditionally lethal, even at a temperature normally permissive for cdc25-22. Finally, the characteristic suppression of the cdc25 phenotype by a loss-of-function weel mutation is reversed in a mcs3 mutant background. The mcs genes define new mitotic elements that might be activators or substrates of the cdc2 protein kinase.
I N eukaryotes, entry of the interphase cell into mitosis is induced by a nonspecies specific activity
known as M-phase promoting factor (MPF) (MASUI and MARKERT 197 1 ; K I ~ H I M ~ T ~ and KANATANI 1976; SUNKARA, WRIGHT and RAO 1979; NELKIN, NICHOLS and VOCELSTEIN 1980; WEINTRAUB et al. 1982; SO- RENSEN, CYERT and PEDERSEN 1985; TACHIBANA, YANAGISHIMA and KISHIMOTO 1987). MPF is a pleio- tropic meiotic activator that induces the full range of mitotic events including breakdown of the nuclear membrane, chromosome condensation and formation of the mitotic spindle (MIAKE-LYE, NEWPORT and KIRSCHNER 1983). MPF is usually detected by its ability to induce G2 arrested Xenopus oocytes to enter meiotic “phase, but entirely cell-free mitotic assays have also been developed (MIAKE-LYE and KIRSCHNER 1985; LOHKA and MALLER 1985).
Recently, it has been shown that the frog homolog of the fission yeast c d c 2 protein kinase is one compo- nent of MPF (DUNPHY et al. 1988; GAUTIER et al. 1988). Further components of MPF appear to be the mitotic cyclins (EVANS et al. 1983; SWENSON, FARRELL and RUDERMAN 1986), which are related to the c d c l 3 gene product of fission yeast (BOOHER and BEACH 1988; SOLOMON et al. 1988; COEBL and BYERS 1988). Cyclins are proteolytically degraded at each cell divi- sion, probably causing MPF to be inactivated and allowing the cell to return to interphase.
ton, Ontario K7L 3N6, Canada. ‘Permanent address: Department of Biology, Queens University, Kings-
of page charges. This article must therefore be hereby marked “advertisement” T h e publication costs o f this article were partly defrayed by the payment
in accordance with U.S.C. $1734 solely to indicate this fact.
Generics 122: 773-782 (August, 1989)
c d c 2 of fission yeast, which is the homolog of the budding yeast CDC28 gene (BEACH, DURKACZ and NURSE 1982), is required both for entry into mitosis and also independently for the initiation of DNA synthesis (NURSE, THURIAUX and NASMYTH 1976; NURSE and BISSETT 1981). The finding that the cells of higher eukaryotes, including man (DRAETTA et al. 1987; LEE and NURSE 1987), have a c d c 2 protein kinase that plays a role in regulation of mitosis focuses attention on elements that interact with yeast cdc2 .
Genetic screens in fission yeast have identified two genes whose products have been shown to physically interact with the c d c 2 protein kinase. First, s u c l was isolated as a plasmid-borne sequence that could rescue some but not all temperature-sensitive c d c 2 mutants (HAYLES et al. 1986; HINDLEY et al. 1987). The 13- kD s u c l product appears to be a subunit of the c d c 2 protein kinase in fission yeast and also in human cells (BRIZUELA, DRAETTA and BEACH 1987; DRAETTA et al. 1987; DRAETTA and BEACH 1988).
Second, an allele of the c d c l 3 gene that conferred a temperature-sensitive cell cycle defect was isolated as an extragenic suppressor of a cold-sensitive allele of c d c 2 (BOOHER and BEACH 1987). c d c l 3 is required only for entry into mitosis and not for DNA synthesis (BOOHER and BEACH 1987, 1988). The allele of c d c 2 that is suppressible by a compensating c d c l 3 mutation is presumed to be defective only in its G2 function (BOOHER and BEACH 1987). The product of c d c l 3 is homologous to mitotic cyclins (SOLOMON et al. 1988; GOEBL and BYERS 1988) and has recently been shown

774 L. Molz et al.
to be a further subunit of the cdc2 protein kinase. T w o genes that interact with cdc2 in the regulation
of mitosis, but whose products do not necessarily physically complex with the protein kinase include cdc25 and weel. Genetic evidence suggests that these two genes act antagonistically (RUSSELL and NURSE 1986, 1987), cdc25 as an activator (RUSSELL and NURSE 1986) and weel as an inhibitor of cdc2 function in GS (NURSE 1975; NURSE and THURIAUX 1980). Either loss-of-function alleles of weel or dominant so- called wee alleles of cdc2 (cdc2-w), bypass the require- ment for the cdc25 gene product (FANTES 1979, 198 1).
Combining a particular activated allele of cdc2 (cdc2-3w) with a recessive temperature-sensitive w e e l mutation (wee 1-50) yields a temperature-sensitive lethal strain. The phenotype of this strain, described in greater detail below, has been termed mitotic ca- tastrophe (RUSSELL and NURSE 1987). These cells do not display cell cycle arrest, but instead enter mitosis prematurely and do not complete it effectively. We have taken advantage of the lethal cdc2-3w wee 1-50 strain to isolate extragenic revertants that might be expected to target further genes whose products in- teract closely with the cdc2 protein kinase in mitotic regulation.
MATERIALS AND METHODS
Media and inhibitors: Strains were grown in standard Schizosaccharomyces pombe media (BEACH, RODGERS and GOULD 1985). Thiabendazole (Sigma) was prepared in di- methylsulfoxide as a 5 mg/ml stock solution and stored in the dark at 40". Thiabendazole was added to autoclaved YEA medium at concentrations of 10, 15, and 20 rg/ml alter cooling to 55". Phloxin B (Sigma) was added to solid medium at 20 Pg/ml in some cases as an indicator of cell viability (GUTZ et al. 1974).
Flow cytometry: Flow cytometry was performed as pre- viously described (BEACH, RODCERS and GOULD 1985). Cells were fixed in ethanol, digested with RNase, stained with propidium iodide and analyzed in a Coulter Electronics Epic C system.
Cell viabillty: Cell viability was determined by particle and colony counts. A sample of the cell suspension was briefly sonified and plated on a YEA plate. At 24 hr the plate was examined microscopically and the percentage of cells forming colonies was scored.
Cytology: S. pombe strains were stained with diamidino- phenylindole (DAPI) according to the method of MARKS and HYAMS (1985).
Strain constructions: All strains were derived from wild- type S. pombe strains 972 (h-S) 975 or 968 (h9') intro- duced by U. LEUPOLD. Conventions for the genetic nomen- clature of S . pombe have been previously described (KOHLI 1987). The isolation and characterization of the cdcl3-117, cdc25-22 (NURSE, THURIAUX and NASMYTH 1976), cdc2- Iw, wee 1-50 (NURSE and THURIAUX 1980), cdc2-?w (FANTES 1981), cdcl3-cl and cdc2-59 (BOOHER and BEACH 1987) mutations have been previously described. Heteroth- allic strains listed in the text as h+ and h- refer to the mating types h+"'and h?, respectively. All diploids used in this study were constructed by the method of complementing ade6-
210 and ade6-216 alleles as previously described (GUTZ et al. 1974). Standard genetical procedures used for S. pombe were performed as previously described (GUTZ et al. 1974), except that matings were done on complete minimal media (PMA) plates rather than on malt extract plates.
Mutagenesis and isolation of the mitotic catastrophe rev- ertant strains have been previously described (BOOHER and BEACH 1987). One representative triple mutant strain from each linkage group strain was sequentially crossed to wild- type strains to obtain the mcs mutations in weel-50, cdc2- ?w, and wild-type backgrounds.
mcsl-77 cdc2-3w weel-50 (SP772) was crossed to a wild- type strain (SP202) and a tetrad was identified with two mitotic catastrophe segregants. The other two segregants in the tetrad were presumed to be the mcsl-77 mutation alone. These segregants displayed a cold-sensitive leaky cell cycle defect described in detail in the text. One of these segregants (SP820) was crossed to strains carrying the cdc2-3w (SP661) and weel-50 (SP616) mutations and tetrads were dissected. In both cases, a segregation of parental ditypes (PD), non- parental ditypes (NPD) and tetratypes (T) was observed, and mcsl-77 cdc2-3w and mcsl-77 weel-50 progeny could be identified. mcsl-77 weel-50 (SP829) and mcsl-77 cdc2-3w (SP831) double-mutant strains displayed a phenotype of heterogeneous cell size described in detail in the text. The genotype of these segregants was confirmed by crossing them with a mitotic catastrophe strain to demonstrate that they suppressed mitotic catastrophe.
mcs2-75 cdc2-3w weel-50 (SP881) was outcrossed to a wild-type strain (972) and a segregant with an elongated phenotype was chosen for further analysis. The genotype of this segregant was determined to be mcs2-75 cdc2-3w (SP834) by outcrossing it to cdc2-3w (SP661), weel-50 (SP616), and wild-type strains (972). In a cross of mcs2-75 cdc2-3w (SP834) with a wild-type strain (972), a NPD tetrad could be identified by the segregation of two wee progeny. The other two progeny in these tetrads displayed a wild- type phenotype and were presumed to carry the mcs2-75 mutation alone. The presence of the mcs2-75 mutation (SP887) was confirmed by sequentially crossing the cdc2-3w (SP662) and weel-50 (SP546) into the strain and demon- strating that the mutation suppressed mitotic catastrophe.
mcs3-12 cdc2-3w weel-50 (SP759) was crossed to a weel- 50 cdc25-22 (SP552) strain and conditional lethal progeny were observed. One of these progeny (SP803) was crossed with a strain carrying the weel-50 cdc25-22 (SP552) muta- tions and a 2:2 segregation of cdc+:cdc- progeny was ob- served. Thus, this conditional lethal strain was confirmed to have the genotype mcs3-12 weel-50 cdc25-22, and its phe- notype is described in detail in the text. This strain was then crossed to a strain containing only the cdc25-22 (SP587) mutation. Nonparental ditypes were detected as tetrads segregating two wee progeny, which are presumed to be of the genotype weel-50 cdc25-22. The other two progeny segregating in the tetrad are presumed to have the genotype mcs3-12 cdc25-22. Next, this strain was crossed to a wild- type strain and one tetrad presumed to be a tetratype was observed. The tetrad segregated a 2:2 ratio of cdc+:cdc- progeny. Both of the cdc+ progeny displayed wild-type length cells, but one stained slightly darker with phloxin B on a YEA plate. This segregant was then sequentially crossed back into the mitotic catastrophe background to confirm the presence of mcs3-12 (SP862).
mcs6-13 cdc2-3w weel-50 (SP760) was outcrossed to a wild-type strain (972) and a conditional lethal cdc- pheno- type segregant was chosen for further analysis. When this strain (SP833) was outcrossed to a wild-type strain, the cross could not be interpreted simply as a two-point cross, but as a cross involving more markers. When this segregant was

Six Interacting mcs Genes 775
crossed to a strain carrying the cdc2-3w mutation (SP661), the cross could be interpreted as a two-point cross segregat- ing parental ditypes, nonparental ditypes and tetratypes. Thus, the conditional lethal segregant was determined to carry two mutations in addition to the cdc2-3w mutation. Both of the mutations were crossed into a mitotic catastro- phe background and both suppressed mitotic catastrophe. They were designated mcs4-13 and mcs6-13. An mcs4-13 cdc2-3w double-mutant strain was outcrossed to a wild-type strain and the cross could be interpreted as a two-point cross, segregating parental ditypes, nonparental ditypes and tetratypes. Progeny presumed to be the mcs6-13 mutation (SP849) alone displayed the nutritionally dependent elon- gated phenotype described in detail in the text. The mcs6- 13 cdc2-3w double mutant (SP879) displayed an elongated phenotype described in detail in the text. The mcs6-13 cdc2-3w double-mutant strain (SP879) was outcrossed to a wild-type strain (SP202) and the cross could be interpreted as a two-point cross. NPD could be identified as tetrads which segregated two wee progeny. In all cases the other two progeny, presumed to be the mcs4-13 mutation (SP886), alone displayed a phenotype indistinguishable from wild-type. The presence of the mcs6-13 mutation was con- firmed by crossing it back into a mitotic catastrophe strain and demonstrating suppression of mitotic catastrophe.
mcs5-74 wee 1-50 cdc2-3w (SP761) was outcrossed to a wild-type strain (SP202) and one tetrad was obtained which segregated two mitotic catastrophe progeny. The other two progeny, presumed to carry only the mcs5-74 mutation, displayed a phenotype indistinguishable from wild type. The presence of the mcs5-74 mutation (SP863) was confirmed by sequentially crossing the mutation back into a mitotic catastrophe background and then demonstrating suppres- sion of mitotic catastrophe.
mcs cdc2-3w cdc25-22 and mcs weel-50 cdc25-22 triple mutant strains were constructed by crossing the mcs weel- 50 and mcs cdc2-3w mutant strains against wee 1-50 cdc25- 22 (SP552) and cdc2-3w cdc25-22 (SP627) strains, respec- tively. Genetic interactions were detected for the mcs2-75 cdc2-3w cdc25-22 (SP755), mcs6-13 cdc2-3w cdc25-22 (SP902), mcs3-12 wee 1-50 cdc25-22 (SP803), mcs4-13 weel-50 cdc25-22 (SP890), and mcs6-13 weel-50 cdc25-22 (SP897) strains as the segregation of a new phenotypic class. The genotype of triple mutant strains was confirmed by crossing the strains back to the weel-50 cdc25-22 and cdc2- 3w cdc25-22 parental strains.
Strains containing the mitotic catastrophe suppressors in a wild-type background (SP819, SP821, SP847, SP887, SP862, SP849, SP863 and SP878) were crossed against strains containing cdc2-lw (SP623 and SP903), cdc2-59 (SP652 and SP653), cdc2-33 (SP354 and SP355), cdc25-22 (SP586 and SP587), cdcl3-cl (SP692 and SP693), and cdc13-117 (SP731 and SP732), and the phenotypes were observed at 25 O and 35 O . Genetic interactions were detected as the segregation of a new phenotypic class and are de- scribed in detail in the text. The genotype of the double mutants was confirmed by crossing the presumed double mutant against the parental strains, and by outcrossing the double mutants with a wild-type strain.
Plasmids and yeast transformations: The S.pombe repli- cating vectors pDB248x (BEACH and NURSE 1981) and pIRT3 were used. The construction of pIRT3 has been previously described (BOOHER and BEACH 1988). The LEU2 genes of S. cereuisiae can rescue leu1 mutants of fission yeast (BEACH and NURSE 1981). pcdc2-89 used in this study is pIRT3 with a 1.3-kb Hind111 insert bearing the cdc2+ gene. psucl-1 has been previously described (HAYLES et al. 1986). pcdc25-1 carries an 8-kb genomic insert in pWH5 (WRIGHT et al. 1986) encompassing the cdc25+ gene (P. YOUNG, un-
published results). pwee2-1 carries a 5.5-kb genomic insert in pWH5 encompassing the weel+ gene (G. DRAETTA, un- published results). The latter two plasmids were derived from a sized Sau3A partial gene bank constructed in the BclI site of pWH5 (YOUNG and BEACH, unpublished results). pniml-1 was a generous gift from PETER FANTES. The construction of pcdcl3-X 1 b has been previously described (BOOHER and BEACH 1988). Transformation of S. pombe has been previously described (BEACH and NURSE 1981).
RESULTS
Mitotic catastrophe phenotype: The lethal mitotic catastrophe phenotype was first described in strains that overexpress the c d c 2 5 inducer but lack w e e l gene function (RUSSELL and NURSE 1986). Cell viability of this strain was drastically reduced within 8 hr of shifting an exponential culture to a restrictive tem- perature. We first investigated the phenotype of the c d c 2 - 3 w w e e l - 5 0 mitotic catastrophe strain, which has not previously been described in any detail.
A c d c 2 - 3 w w e e l - 5 0 double mutant (SP638) was transferred to minimal medium lacking a nitrogen source at a permissive temperature of 25 '. Flow cy- tometric analysis of the DNA content of the cells after 12 hr revealed that 70% had accumulated in GI, a figure typical for this organism in nitrogen limited medium (COSTELLO, RODGERS and BEACH 1986) (Fig- ure 1B). The culture was subsequently transferred to complete medium at a restrictive temperature (35", w e e l - 5 0 is temperature-sensitive) and samples taken at intervals for estimation of cellular DNA content and viability. In a wild-type strain, DNA replication and cell division occur within 6 hr of nutritional and temperature shift-up and the cell viability remains at 100% in a plating test (Figure 1, B and C). A cdc2-3w w e e l - 5 0 strain likewise underwent DNA replication but gradually began to lose cell viability at 6 hr. Within 28 hr less than 1 % of cells could form colonies at 25 O
(Figure IC). Cytological examination revealed that many cells divided abnormally, often leaving one daughter without a nucleus, or with abnormally small nuclei (Figure 1A). This is reflected in the very dis- persed cellular DNA content that develops within 10 hr of shift-up of the double mutant (Figure 1B).
These observations indicate that the lethal pheno- type of the cdc2-3w wee 1-50 strain is not due to grossly premature entry into mitosis without prior DNA replication, but more likely due to aberrant execution of mitosis, particularly with respect to chro- mosome segregation and septum formation.
Six mcs genes: The isolation of seventeen extra- genic revertants in a c d c 2 - 3 w w e e l - 5 0 double-mutant strain has been described previously (BOOHER and BEACH 1987). Briefly, this temperature-sensitive strain was subjected to chemical mutagenesis and mu- tants capable of forming colonies at 37' were selected. The revertants were each back-crossed to a wild-type strain and in 17 cases segregants displaying the mitotic

776
A
L. Molz et al.
TABLE 1
Identification and initial characterization of six rncs loci
m:i ~h~~~~~ rncs phenotype in a
No. of background backgount' Domi- nance/
B Wlld-type cdc2-3w woel-50
loo\ 80
Tome (hrs) after shlft-up
FIGURE I.-Phenotype of mitotic catastrophe strain. (A) wild- type (972) and cdr2-3w weel-50 (SP638) double-mutant cultures were shifted to 35" for 5 hr. prior to staining with DAPI. (B) Flow- cytometric analysis of a wild-type and cdc2-3w weel-50 double mutant strains. Cultures were starved of a nitrogen source at the permissive temperature (25") and then shifted to complete media at the restrictive temperature (35'). Samples were taken at 0 , 7 and IO hr after shift up. Open triangles represent IC DNA content and closed triangles 2C. A G I peak is not detected at 10 hr because in exponentially growing cells DNA synthesis occurs before cell sepa- ration and thus the newly born daughter cells already have a 2N DNA content. (C) Fmction of wild-type (circles) and double-mutant (squares) cells able to form colonies on a g r plates at 25". at time intervals following shift-up to 3.5". The closed triangle represents the time at which an increase in the cell number is first observed i n a wild-type culture.
catastrophe phenotype were found. These 17 there- fore carry suppressors that are extragenic to either cdc2 or weel.
Each of the original revertant strains was inter- crossed pair-wise to determine the number of linkage groups (Table 1). Initially the revertants defined five unlinked groups called mcs (mitotic catastrophe sup- pressor). However, upon further genetic analysis, it transpired that one triple mutant strain had developed a secondary spontaneous mutation which was un- linked to the first but also independently suppressed mitotic catastrophe (see MATERIALS AND METHODS). This mutation was designated mrs4-13 and the origi- nal suppressor mcs6-13 (Table I ) .
One allele was chosen from each linkage group for further physiological and genetic analysis. First we tested whether the mcs mutations are dominant or recessive with respect in their ability to suppress mi- totic catastrophe. Diploids heterozygous for the rncs mutation but homozygous for wee 1-50 and cdc2-3w
Linkage alleles group isolated 25" 55" 25" 35'
recessive- nessD
mcsl 1 1 wee' wee' cdc-.' cdc+ Rec mcs2 3 wee' c ~ c - . ~ cdc+ cdc+ Rec mcs3 1 wee wee cdc' cdc+ Rec mcs4 I wee' wee' cdc-.' cdc-.' Rec mcs5 1 wee' wee' cdc+ cdc+ Rec mcs6 1 wee' cdc-.d cdc+ cdc+ Rec
a The mcs mutations were determined to be dominant or reces-
' These strains formed colonies of cells that ranged from wee to
' Poorly growing colonies of elongated cells.
sive w i t h respect to their ability to suppress mitotic catastrophe.
wild-type in length at cell division.
Poorly growing colonies of elongated cells which ranged from wee to highly elongated in length at cell division, and accumulated multiple nuclei and division septa. ' Poorly growing colonies of cells which divided at a length
approximately 1.5 times larger than wild-type on minimal media, but at a wild-type cell length on yeast extract media.
were constructed at 25" (see MATERIALS AND METH- ODS) and assayed at 35 ". Each of the diploids was not viable at the raised temperature indicating that each rncs mutation is recessive to wild-type in its ability to suppress mitotic catastrophe.
We further tested whether any mcs mutant might be allelic to any known mitotic control gene. Classical linkage analysis excluded cdc2+, weel+ (since the mu- tants are extragenic to cdc2-3w weel-50) and also cdc25+ (see below). We also introduced plasmids car- rying sucl+, cdc25+, cdcl3+or nimP into each triple mutant strain and observed no reversion of mitotic catastrophe suppression. By this criterion, none of the rncs mutants appear to be an allele of any of these genes.
The possibility of allelism between the mcs mutants and either winl+ (OGDEN and FANTES 1986) or either of the two cdr+ genes (YOUNG and FANTES 1987) was not tested.
Phenotype of rncs mutants: rncs cdc2-3w wee 1-50 triple-mutant strains were examined at 25" and 35" and three distinct phenotypic classes were observed (Table 1 , Figure 2). Members of the first class (mcsl- 77, mcs4-13, and mcs5-74; SP763, SP898, and SP761, respectively) displayed similar behavior at either 25" or 35". The cells were highly heterogeneous in length at division, ranging between wee (approximately half the wild-type length) to wild type (Figure 2 and Table 1). Members of the second class (mcs2-75 and mcs6- 13, SP881 and SP882, respectively) were tempera- ture-sensitive. At 35", cell length at division varied from small to highly elongated, whereas at 25" no elongated cells were observed (Figure 2 and Table 1). The third class was represented by the mcs3-I2 cdc2- 3w weel-50 triple mutant (SP759) which displayed a
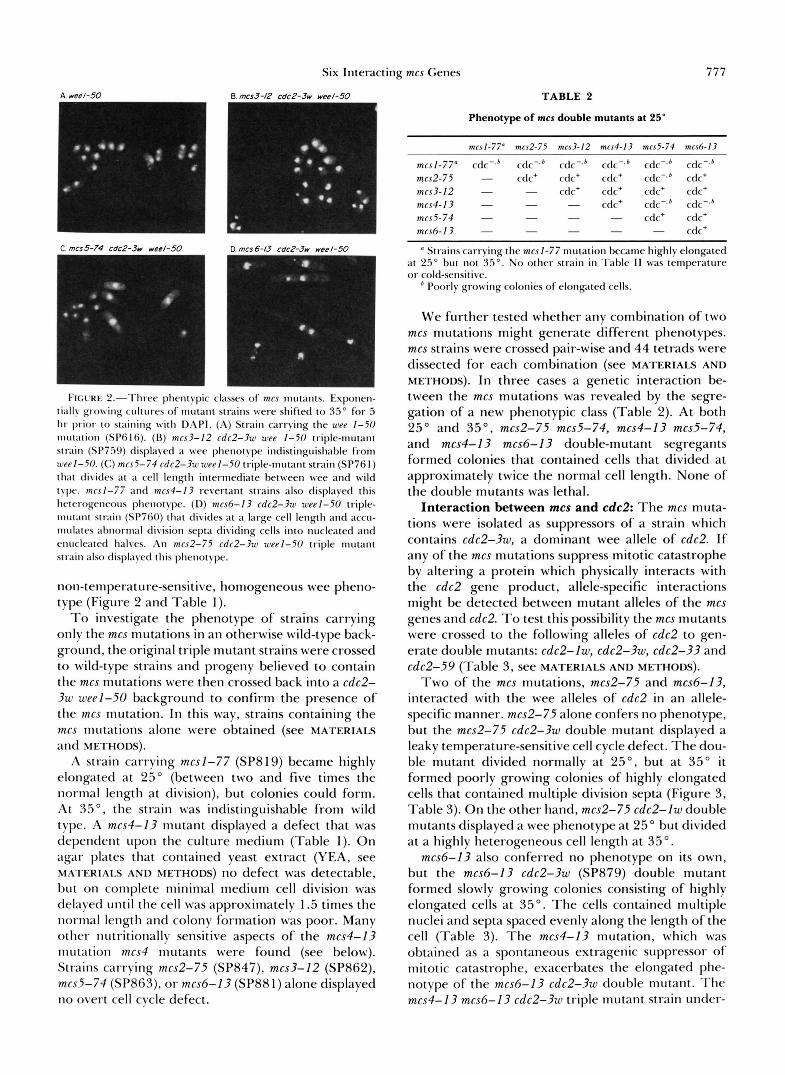
Six Interacting mcs Genes 777
TABLE 2
Phenotype of mcs double mutants at 25"
I I G L . R K ~ . - . I ' I I I C C plxm[yic classes 01. mcs ~nutants. I . S ~ O I I C I I -
t i a l l \ growing cultures of 1nut;tnt strains were shifted t o :+.So for 5 hr prior to staining with DAPI. (A) Strain carrying the utet 1-50 mut;ltion (SP(iI6). (B) mcs3-12 cdc2-3w wee 1-50 triple-mutant strain (SP759) displayed a wee phenotype indistinguishable from a w l - 5 0 . (C) mcs5-74 cdc2-3wweel-50 triple-mutant strain (SP761) that divides at a cell length intermediate between wee and wild type. mrsl-77 and mcs4-I3 revertant strains also displayed this heterogeneous phenotype. (D) mcs6-13 cdc2-3up weel-50 triple- mw;tnt strain (SP760) th;tt divides at a large cell length and accu- mulates abnornlal division septa dividing cells into nucleated and enucleated halves. An mcs2-75 cdc2-3w weel-50 triple mutant strain also displayed this phenotype.
non-temperature-sensitive, homogeneous wee pheno- type (Figure 2 and Table l).
T o investigate the phenotype of strains carrying only the mcs mutations in an otherwise wild-type back- ground, the original triple mutant strains were crossed to wild-type strains and progeny believed to contain the mcs mutations were then crossed back into a cdc2- 3w weel-50 background to confirm the presence of the mcs mutation. In this way, strains containing the rncs mutations alone were obtained (see MATERIALS and METHODS).
A strain carrying mcsl-77 (SP819) became highly elongated at 25" (between two and five times the normal length at division), but colonies could form. At 35", the strain was indistinguishable from wild type. A mcs4-13 mutant displayed a defect that was dependent upon the culture medium (Table 1). On agar plates that contained yeast extract (YEA, see MATERIALS AND METHODS) no defect was detectable, but on complete minimal medium cell division was delayed until the cell was approximately 1.5 times the normal length and colony formation was poor. Many other nutritionally sensitive aspects of the mcs4-13 mutation mcsl mutants were found (see below). Strains carrying mcs2-75 (SP847), mcs3-12 (SP862), mcs5-74 (SP863), or mcs6-13 (SP881) alone displayed no overt cell cycle defect.
~~
mcsl-77' mcs2-75 mcs3-12 mcs4-13 mcs5-74 mcs6-13
mcsl-77" cdc-.b c ~ c - . ~ c ~ c - . ~ cdc-.b cdc-.b cdc-.b mcs2-75 - cdc+ cdc+ cdc+ c ~ c - . ~ cdc+ mcs3-12 - cdc+ cdc' cdc' cdc+ mcs4-13 - - - cdc' cdc-.' CdC-.b mcs5-74 - - - - CdC+ cdc+ mcs6-13 - - - - - cdc'
-
a Strains carrying the mcsl-77 mutation became highly elongated ;kt 2.5" but not 35". N o other strain in Table I I was temperature or cold-sensitive.
Poorly growing colonies of elongated cells.
We further tested whether any combination of two mcs mutations might generate different phenotypes. mcs strains were crossed pair-wise and 44 tetrads were dissected for each combination (see MATERIALS AND METHODS). In three cases a genetic interaction be- tween the mcs mutations was revealed by the segre- gation of a new phenotypic class (Table 2). At both 25" and 35", mcs2-75 mcs5-74, mcs4-13 mcs5-74, and mcs4-13 mcs6-13 double-mutant segregants formed colonies that contained cells that divided at approximately twice the normal cell length. None of the double mutants was lethal.
Interaction between mcs and cdc2: The mcs muta- tions were isolated as suppressors of a strain which contains cdc2-3w, a dominant wee allele of cdc2. If any of the mcs mutations suppress mitotic catastrophe by altering a protein which physically interacts with the cdc2 gene product, allele-specific interactions might be detected between mutant alleles of the mcs genes and cdc2. To test this possibility the mcs mutants were crossed to the following alleles of cdc2 to gen- erate double mutants: cdc2-lw, cdc2-3w, cdc2-33 and cdc2-59 (Table 3, see MATERIALS AND METHODS).
Two of the mcs mutations, mcs2-75 and mcs6-13, interacted with the wee alleles of cdc2 in an allele- specific manner. mcs2-75 alone confers no phenotype, but the mcs2-75 cdc2-3w double mutant displayed a leaky temperature-sensitive cell cycle defect. The dou- ble mutant divided normally at 25", but at 35" it formed poorly growing colonies of highly elongated cells that contained multiple division septa (Figure 3, Table 3). On the other hand, mcs2-75 cdc2-lw double mutants displayed a wee phenotype at 25" but divided at a highly heterogeneous cell length at 35".
mcs6-13 also conferred no phenotype on its own, but the mcs6-13 cdc2-3w (SP879) double mutant formed slowly growing colonies consisting of highly elongated cells at 35". The cells contained multiple nuclei and septa spaced evenly along the length of the cell (Table 3). The mcs4-13 mutation, which was obtained as a spontaneous extragenic suppressor of mitotic catastrophe, exacerbates the elongated phe- notype of the mcsb-13 cdc2-3w double mutant. The mcs4-13 mcs6-13 cdc2-3w triple mutant strain under-

778 L. Molz et al.
TABLE 3
Interactions between mcs and cdc2 mutants
cdc2+ c d c 2 - 3 ~ cdc2- I H' cdc2-59 cdc2-33
25" 35" 25" 35" 25" 35" 320 25' 350
mcs' cdc' cdc' wee wee wee wee cdc' cdc' cdc-." mcsl-77 CdC-.b cdc' wee' wee' wee' wee' CdC-.d CdC-.b cdc-." mcs2-75 cdc' cdc' cdc+ cdc-.' wee wee/ cdc-.d cdc' cdc-." mcs3-12 cdc' cdc' wee wee wee wee CdC-J cdc+ cdc-." mcs4- I 3 cdc' cdc' wee' wee' wee' weer cdc-.d CdC+ cdc-." mcs5-74 cdc+ cdc' wee wee wee wee cdc-.d cdc' cdc-." mcs6-13 cdc' cdc' cdc' cdc-.' wee wee cdc-.d cdc+ cdc-."
The phenotypes presented in Table 111 are the phenotypes the strains observed on yeast extract containing media. a These strains arrested in cell division as highly elongated cells.
These strains formed slowly growing colonies of elongated cells. These strains formed colonies of cells that ranged from wee to wild-type in length at cell division. Spores of this genotype germinated and divided several times before arresting as a microcolony consisting of approximately 5-20 highly
eloncrated cells. " ' These strains formed slowly growing colonies of elongated cells which accumulated multiple nuclei and division septa evenly spaced along
/These strains formed colonies of cells that ranged from wee to highly elonKated in cell length but did not accumulate multiple nuclei or the cell.
division septa.
C. mcs2-75 cdc2-3,
E. mcsZ-75 CdCZ-Iw
D mcs4-/3 mcs6-I3 cdc2-3w
FIGURE 3.-Interactions between mcs and cdr2 mutations. Ex- ponentially growing cultures of mutant strains were shifted to 35" for .5 hr prior to staining with DAPI. (A) mcs2-75 (SP847) single- mutant divides at a wild-type cell length. (B) An mcs2-75 cdc2-lw double-mutant strain (SP880) displays a highly heterogeneous phe- notype in which cells ranged in length from wee to highly elongated at division. (C) Cells of an mcs2-75 cdc2-3w double-mutant strain (SP834) divide at a highly elongted cell length and accumulate abnormal division septa. (I)) Cells of an mcs4-13 mcs6-13 cdc2-3w triple-mutant strain (SP833) undergo cell cycle arrest at 35" and accumulate abnornlal division septa.
goes immediate cell cycle arrest at 35" and accumu- lates multiple abnormal division septa and nuclei (Fig- ure 3). However, the mcs6-13 cdc2-lw double mutant displayed a wee phenotype indistinguishable from the phenotype of the cdc2-lw mutant alone (Table 3).
Therefore, the interactions between mcs2-75 and mcs6-13 with the wee alleles of cdc2 was highly allele specific. Among the other mcs mutants, slight effects were observed in mcsl-77 cdc2-3w and mcs4-13 cdc2- 3w double mutants. These mcs mutants partially sup- pressed the cdc2-3w wee phenotype, causing cell di- vision to occur at more normal cell lengths.
Since the cdc2-3w mutation is dominant or semi- dominant to wild type (FANTES 1981) we investigated whether cdc2-3w is dominant or recessive with respect to cdc2+ in its interactions with mcs2-75 and mcs6-13. mcs2-75 cdc2- 3wlmcs2-75 cdc2+ and mcs6-I3 cdc2- 3wlmcs6-I3 cdc2+ diploids were constructed (see MA- TERIALS AND METHODS) and their phenotypes were observed at 25" and 35". The cell length of each at division was approximately twice that of mcs2-751 mcs2-75 and mcs6-l3/mcs6-13 diploids, respectively. However, these diploids did not generate fully ar- rested cells that accumulate with multiple division septa, as do the mcs2-75 cdc2-3w and mcs6-I3 cdc2- 3w haploids. Thus cdc2-3w appears to be a semidom- inant mutation with respect to wild type in its inter- action with mcs2-75 and mcs6-13.
Given that the interaction of cdc2-3w with mcs2-75 or mcs6-13 was not fully dominant with respect to cdc2+, we tested whether overexpression of cdc2+ might suppress the effect of cdc2-3w. The cdc2+ gene was introduced on a multicopy vector into mcs2-75 cdc2-3w (SP88 1) and mcs6-I3 cdc2-3w (SP882) strains (see MATERIALS AND METHODS). The cell cycle defect of both strains at 35" was fully rescued by the plasmid. These strains were not rescued by the introduction of plasmids carrying cdc25+, weel+, sucI+, nim1+ or cdcl3+.
In addition to the wee alleles of cdc2, we have investigated whether loss-of-function cdc2 alleles in-

Six Interacting mcs Genes 779
teract with the mcs mutations. cdc2-59 was isolated as an intragenic suppressor of mitotic catastrophe (BOOHER and BEACH 1987). This allele presumably carries two lesions, the original cdc2-3w mutation and a secondary mutation which prevents mitotic catastro- phe in a weel-50 background and confers a cold- sensitive cdc phenotype at 25". cdc2-59 is the only known allele of cdc2 that appears to be defective for GZ, but not G1 function (BOOHER and BEACH 1987).
Strains carrying cdc2-59 (SP652 and SP653) were crossed with those containing each mcs mutation (SP821, SP847, SP862, SP849, SP863 and SP886). Tetrads were dissected and allowed to germinate at 33", a temperature normally permissive for cdc2-59 (see MATERIALS AND METHODS). In crosses containing the mcsl-77, mcs2-75, mcs4-13, mcs5-74 and mcs6- 13 frequent cdc- segregants were observed. These segregants, which were presumed to be the cdc2-59 mcs double mutants, germinated to form a microco- lony of less than 20 cells before arresting as highly elongated cells. These nonconditionally lethal segre- gants could be inferred to be the double mutants by scoring the phenotype of the surviving spores, with the exception of the mcs5-74 and mcs6-13 mutations which cannot be easily be scored in a wild-type back- ground. By contrast, the mcs3-12 cdc2-59 double mutant was viable at 33". Thus, with the exception of mcs3-12, each of the mcs mutations exacerbated the defect of cdc2-59.
We also determined if any of the mcs mutations interacted with cdc2-33, a recessive temperature-sen- sitive allele of cdc2 which can undergo a cell cycle arrest at both the G1 or GZ restriction points. Strains containing the mcs mutations (SP821, SP847, SP862, SP849, SP863 and SP886) were crossed to strains carrying cdc2-33 (SP354 and SP355) and in all cases, the double mutants divided at a wild-type cell length at 25" and displayed a first cycle arrest at 35" (see MATERIALS AND METHODS). Therefore, the mcs muta- tions did not affect expression of the cdc2-33 pheno- type (Table 3).
mcs interaction with cdcl3: T w o conditional lethal alleles of the c d c l 3 gene have been isolated to date, cdc13-117 (NURSE, THURIAUX and NASMYTH 2976) and cdcl3-c l (BOOHER and BEACH 1987). Neither of these temperature-sensitive mutations is a full null- allele of the gene which causes total inhibition of mitotic initiation (BOOHER and BEACH 1988). cdcl3- I 1 7 allows some cells to enter mitosis and undergo aberrant septation. The cdcl3-c l allele is even more leaky and very poorly growing colonies can be ob- tained at 37". Both of these alleles of cdcl3+ interact in complex ways with cdc2 mutations (BOOHER and BEACH 1988).
mcs2-75, mcs5-74 and mcs6-13 each altered the phenotype of the cdc13-117 mutant. At 35", the mcs2-75 cdc13-117 (SP892), mcs5-I3 cdc13-117
(SP893), and mcs6-13 cdcl3-1 I 7 (SP894) double-mu- tant strains arrested as highly elongated cells which did not enter mitosis or accumulate multiple division septa (Table 4). Likewise, mcs2-75 cdcl3-cl (SP896) and mcs6-13 cdcl3-cl (SP895) double mutants dis- played first cycle arrest with division septa at the restrictive temperature (Table 4). No interactions were observed with any other mcs mutations and alleles of cdcl3 .
mcs interactions with weel and cdc25 weel-50 and cdc25-22 are temperature-sensitive loss-of-function mutations. weel+ has been proposed to be an inhibitor and cdc25+ an activator of cdc2+. Both mcsl-77 and mcs4-13 mutations interacted strongly with cdc25-22. cdc25-22 mutants are viable at 25" but slightly elon- gated. However, both cdc25-22 mcsl-77 and cdc25- 22 mcs4-I3 double mutants could not be propagated at this temperature. Spores of these genotypes ger- minated and divided to form a microcolony consisting of less than twenty cells before arresting as cdc- cells (Table 4). mcs2-75, mcs3-12, mcs5-74 and mcs6-13 did not dramatically alter the phenotype of cdc25-22 mutants.
Strains carrying the cdc25-22 mutation can be phe- notypically rescued by either dominant wee alleles of cdc2 (FANTES 1979, 1981) or loss-of-function alleles of the weel gene (FANTES 1979). We therefore inves- tigated whether either the weel-50 or cdc2-3w muta- tions might suppress the non-conditional lethal phe- notype of the mcsl-77 cdc25-22 and mcs4-13 cdc25- 22 double mutants. Both mcsl-77 weel-50 cdc25-22 (SP889) and mcs4-13 weel-50 cdc25-22 (SP890) triple mutants were constructed and found to be viable at 25 " and 35" on rich medium (see MATERIALS AND METHODS). The phenotype of the triple mutants was indistinguishable from that of mcsl-77 weel-50 (SP829) and mcs4-13 weel-50 (SP884) double mu- tants, namely cells divided at a length intermediate between wee and wild type. Interestingly, the mcs4- I3 weel-50 cdc25-22 (SP890) triple mutant displayed a nutritionally sensitive phenotype. As described above, no striking defect was noticed on rich medium, but on minimal agar at 35" the strain underwent immediate cell cycle arrest.
The cdc2+, weel+, S U C S , cdcl3+ and niml+ genes were introduced into a mcs4-13 weel-50 cdc25-22 triple-mutant strain in order to investigate if overex- pression of these genes might suppress the nutrition- ally sensitive cell cycle defect. Among these, only the cdc2+ gene rescued the lethality of the triple mutant allowing division at the same length as wild-type cells.
The proceeding experiments investigated the inter- actions between mcsl-77, mcs4-13, cdc25-22 and their suppressibility by weel-50. However, a more general question may be asked. Since weel or cdc2w mutants suppress cdc25-22, can any of the mcs mu- tants alleviate this suppression. Such an effect was

780 L. Molz et al.
TABLE 4
Interaction of mcs with cdc25, cdcl3 and wee1 mutants
+ weel-50 cdc25-22 cdc l3 -c l cdc13-117
25" 35" 25" 35" 25" 35" 25" 35" 25" 35"
mcs+ cdc+ cdc+ cdc' wee cdc+ cdc-." cdc+ cdc-,b cdc+ cdc-.' mcsl-77 cdc-.b cdc+ weed weed cdc-,' na cdc+ cdc-.b cdc+ cdc-.' mcs2-75 cdc+ cdc+ cdc+ wee cdc+ cdc-." cdc+ cdc-." cdc+ cdc-." mcs3-I2 cdc+ cdc+ cdc+ wee cdc+ cdc-." cdc+ cdc-,b cdc+ cdc-,' mcs4-13 cdc-J cdc-.f weed weed cdc-.' na cdc+ cdc-.b cdc+ cdc-.< mcs5-74 cdc+ cdc' cdc+ wee cdc+ cdc-." cdc+ cdc-.b cdc+ cdc-." mcs6-I3 cdc+ cdc+ cdc+ wee cdc+ cdc-," cdc+ cdc-." cdc+ cdc-."
NA, not applicable.
* These strains formed slowly growing colonies of highly elongated cells. ' These strains arrested in cell division as highly elongated cells which accumulated multiple division septa.
' Spores of this genotype germinated and divided several times before arresting as a microcolony consisting of approximately 5-20 highly
These strains arrested in cell division as highly elongated cells.
These strains formed colonies of cells that ranged from wee to wild-type in length at cell division.
elongated cells.
media, but at a wild-type cell length on yeast extract media. /These strains formed slowly growing colonies of cells which divided at a length approximately 1.5 times larger than wild-type on minimal
TABLE 5
Yeast strains
Strain Genotype
972 SP202 SP354 SP355 SP546 SP552 SP586 SP587 SP616 SP623 SP627 SP638 SP652 SP653 SP661 SP662 SP638 SP692 SP693 SP731 SP73 2 SP755 SP759 SP760 SP76 1 SP763 SP772 SP803 SP8 19 SP820 SP82 1
h- h+ leul-32 ade6-216 h+ cdc2-33 leul-32 ade6-210 h- cdc2-33 leul-32 ade6-210 h+ weel-50 h+ weel-50 cdc25-22 h- cdc25-22 leul-32 ura4 h+ cdc25-22 leul-32 ade6-210 ura4 h- weel-50 leul-32 ade6-210 h- cdc2-lw leul-32 h+ cdc2-3w cdc25-22 leul-32 h- cdc2-3w weel-50 h- cdc2-59 leul-32 ade6-216 h+ cdc2-59 leul-32 ade6-216 h- cdc2-3w leul-32 h+ cdc2-3w leul-32 ade6-216 h- cdc2-3w weel-50 h+ cdcl3-cl leul-32 ade6-210 h- cdcl3-cl leul-32 ade6-210 h- cdc13-117 leul-32 ade6-210 h+ cdc13-117 leul-32 ade6-210 h- mcs2-75 cdc2-3w cdc25-22 leul-32 h+ mcs3-12 cdc2-3w weel-50 ade6 h+ mcs6-13 cdc2-3w weell-50 ade6 h- mcs5-74 cdc2-3w weel-50 leul-32 ade6 h+ mcsl-77 cdc2-3w weel-50 leul-32 ade6 h- mcsl-77 cdc2-3w weel-50 Eeul-32 ade6 h- mcs3-12 weel-50 cdc25-22 leul-32 h+ mcsl-77 h+ mcsl-77 leul-32 ade6 h- mcsl-77
Strain Genotvve
SP829 SP83 1 SP833 SP834 SP847 SP849 SP862 SP863 SP878 SP879 SP880 SP881 SP882 SP883 SP884 SP885 SP886 SP887 SP888 SP889 SP890 SP89 1 SP892 SP893 SP894 SP895 SP896 SP897 SP898 SP902 SP903
h' mcsl-77 weel-50 leul-32 ade6 h+ mcsl-77cdc2-3w leul-32 ade6 h+ mcs4-13 mcs6-13 cdc2-3w leul-32 h- mcs2-75 cdc2-3w leul-32 ade6 h- mcs2-75 ade6-216 h- mcs4-13 h- mcs3-I2 ade6-216 ura4 h+ mcs5-74 leul-32 h+ mcs6-13 leul-32 h- mcs6-13 cdc2-3w leul-32 ade6 h+ mcs2-75 cdc2-lw ade6 h- mcs2-75 cdc2-3w weel-50 leul-32 ade6 h+ mcs6-13 cdc2-3w weel-50 leul-32 ade6 h- mcs l -77 cdc2- lw leu l -32 h+ mcs4-13 weel-50 leul-32 h+ cdc2-lw leul-32 h- mcs6-13 leul-32 h+ mcs2-75 ade6-216 h mcs4-13 weel-50 h' mcsl-77 weel-50 cdc25-22 h+ mcs4-13 weel-50 cdc25-22 h+ mcsl-77 cdc2-3w cdc25-22 h' mcs2-75 cdc l3-1 I7 l eu l -32 ade6-210 h+ mcs5-74 cdc13-117 leul-32 ade6-210 h- mcs6-13 cdcl3-117 h- mcs6-13 cdcl3-cl leul-32 h- mcs2-75 cdcl3-cl ade6-210 h- mcs6-13 weel-50 cdc25-22 leul-32 h+ mcs4-13 weel-50 cdc2-3w h+ mcs6-13 cdc2-3w cdc25-22 leul-32 h+ cdc2-lw leul-32 ade6-216
Note: The presence of a cdc2-3w or weel-50 mutation in strains carrying the ade6-210 or ade6-216 markers alters the red and pink colors of these strains, respectively, so that they cannot be easily scored.
found in two further cases. mcs3-12 wee 1-50 cdc25- cell cycle phenotype. In the former case this was a 22 (SP803) and mcs6-13 weel-50 cdc25-22 (SP897) tight first cycle arrest, while mcs6-13 weel-50 cdc25- triple-mutant strains displayed a temperature-sensitive 22 strain formed poorly growing colonies of very

Six Interacting mcs Genes 78 1
elongated cells. Neither mcs3-12 nor mcs6-13 muta- tions conferred any unusual phenotype in weel-50 or cdc25-22 single mutant backgrounds (Table 4).
Again the cdc2+, sucl+, cdcl3+ and n i m S genes were introduced into a mcs3-12 weel-50 cdc25-22 triple mutant strain and, as in the case of the equiva- lent mcs4+ strain, the cdc2+ gene was able to rescue the lethal temperature-sensitive cdc phenotype.
Thiabendazole effects: Antimitotic benzimidazole compounds such as benomyl and thiabendazole (TBZ) are believed to specifically inhibit microtubule func- tion in ascomycete fungi by binding to tubulin (DAV- IDSE 1973). In fission yeast, TBZ sensitive mutations have identified the tubulin genes (nda, UMESONO et al. 1983), but the cdcl3-117 mutation also confers mild TBZ hypersensitivity (BOOHER and BEACH 1988).
Wild-type S. pombe is sensitive to TBZ and at ap- proximately 20 pg/ml TBZ cells arrest in division and an aberrant septum frequently forms across the nu- cleus (UMESONO et al. 1983). We have investigated whether any mcs mutant might be hypersensitive to TBZ and found that mcs4 strains displayed such a phenotype. mcs4-13 was unable to form colonies at a concentration of 10 pg/ml TBZ, whereas equivalent inhibition of a wild-type strain occurred at 20 pg/ml.
One further effect was noticed. Wild-type strains arrest in mitosis in the presence of lethal doses of TBZ. However, at equivalent doses mcs3-12 cdc2-3w weel-50 and mcs4-13 cdc2-3w weel-50 triple mutant strains underwent mitotic catastrophe at 35", rather than mitotic arrest.
DISCUSSION
We have isolated extragenic suppressors that rescue the lethal mitotic catastrophe phenotype of cdc2-3w weel-50 double mutants. They fall into six linkage groups and appear to be newly identified genes. Al- though none of the mcs mutants alone conferred a tight cell cycle arrest phenotype, the striking interac- tions with other known mitotic control genes suggests that each of them is intimately involved in the regu- lation of mitosis.
The mcs mutants appear to fall into three general classes on the basis of their interactions with other genes. mcs2 and mcsb both displayed allele-specific interactions with cdc2 mutants, particularly the cdc2- w alleles. Both these mutants also exacerbated the phenotype of leaky cdcl3 mutants. In fact, the range of interactions between mcs2, mcsb and cdc2 were remarkably similar to those previously described be- tween cdc2 and cdc l3 (BOOHER and BEACH 1987). It is now known that cdcl3 encodes a mitotic cyclin (GOEBL and BYERS 1988; SOLOMON et al. 1988), that physically interacts with the cdc2 protein kinase (B. BOOHER and D. BEACH, personal communication). We therefore suspect that the mcs2 and mcsb gene prod-
ucts also interact directly with the cdc2lcdcl3 com- plex, possibly as substrates of the protein kinase.
Another class of genes is represented by mcsl and mcs4 mutants. Both of these mutants are noncondi- tionally lethal in a cdc25 mutant background, and as with the cdc25 mutant itself this lethality can be res- cued by loss of weel function. It has been proposed that the cdc25 gene product is an activator of cdc2 (FANTES 1979; RUSSELL and NURSE 1986). This hy- pothesis has not been elaborated in molecular detail, but is compatible with available genetic data. In this context, it is possible that mcsl and mcs4 act on the same pathway as cdc25, either as activators of its function or as intermediates lying between cdc25 and cdc2.
The final class of genes is represented by mcs3 alone. mcs3-12 is the most effective suppressor of mitotic catastrophe and appears to interact most closely with weel. A simple way in which the mitotic catastrophe phenotype could be rescued would be to negate the effect of loss of weel function. In the mcs3-12 cdc25- 22 wee 1-50 triple mutant this is exactly how the mcs3 mutant appears to behave since this strain displays a temperature-sensitive cdc phenotype. However, mcs3- 12 can not be completely suppressing loss of weel gene function since the mcs3-12 weel-50 double mutant still displayed a wee phenotype.
The genetic interactions described in this paper are extraordinarily complicated and begin to stretch the useful limits of genetic formalism as a tool to build models that illustrate possible biochemical interac- tions. However, the present study has introduced six new elements all of which interact with cdc2 in the regulation of mitosis and which can now be pursued at the molecular level.
We wish to thank DAVID GREENE, JIM DUFFY and CAROL MAR- CINCUK for their help in the preparation of the figures. This work was funded by National Institutes of Health GM34607 to D.B. and by NSERCC A9335 to P.Y.
LITERATURE CITED
BEACH, D., and P. NURSE, 1981 High frequency transformation of the fission yeast Schizosaccharomycespombe. Nature 290 140- 142.
BEACH, D., B. DURKACZ and P. NURSE, 1982 Functionally homol- ogous cell cycle genes in budding and fission yeast. Nature 300:
BEACH, D., L. RODCERS and J. COULD, 1985 RANI+ controls the transition from mitotic division to meiosis in fission yeast. Curr. Genet. 1 0 297-3 1 1 .
BOOHER, R., and D. BEACH, 1987 Interaction between cdcl3+and cdc2' in the control of mitosis in fission yeast; dissociation of the GI and Gn roles of the cdc2+ protein kinase. EMBO J. 6 3441-3447.
BOOHER, R . , and D. BEACH, 1988 Involvementofcdc13+in mitotic control in Schirosaccharomyces pombe: possible interaction of the gene product with microtubules. EMBO J. 7: 2321-2327.
706-709.

782 L. Molz et al.
BRIZUELA, L., G . DRAETTA and D. BEACH, 1987 p13s"""acts in the fission yeast cell division cycle as a component of the p3Vdc2 protein kinase. EMBO J. 6: 3507-3514.
COSTELLO, G., L. RODGERS and D. H. BEACH, 1986 Fission yeast enters the stationary phase Go state from either mitotic GI or GP. Curr. Genet. 11: 119-125.
DAVIDSE, L. C., 1973 Antimitotic activity of methyl benzimidazol- 2-yl carbamate in Aspergallus nidulans. Pest. Biochem. Physiol.
DRAETTA, G. , and D. BEACH, 1988 Activation of cdc2 protein kinase during mitosis in human cells: cell cycle-dependent phosphorylation and subunit rearrangement. Cell 5 4 17-26.
DRAETTA, G . , L. BRIZUELA, J. POTASHKIN and D. BEACH, 1987 Identification of p34 and p13, human homologs of the cell cycle regulators of fission yeast encoded by cdc2+ and sucI+. Cell 50: 3 19-325.
DUNPHY, W. G . , L. BRIZUELA, D. BEACH and J. NEWPORT, 1988 The Xenopus cdc2 protein is a component of MPF, a cytoplasmic regulator of mitosis. Cell 54: 423-431.
EVANS, T., E. T. ROSENTHAL, J. YOUNGBLOOM, K. DISTEL and T. HUNT, 1983 Cyclin: a protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell 33: 389-396.
FANTES, P., 1979 Epistatic gene interactions in the control of division in fission yeast. Nature 279: 428-430.
FANTES, P., 1981 Isolation of cell size mutants of a fission yeast by a new selective method: characterization of mutants and implications for division control mechanisms. J. Bacteriol. 146:
GAUTIER, J., C. NORBURY, M. LOHKA, P. NURSE and J. MALLER, 1988 Purified maturation-promoting factor contains the product of a Xenopus homolog of the fission yeast cell cycle control gene cdc2+. Cell 54: 433-439.
GOEBL, M., and B. BYERS, 1988 Cyclin in fission yeast. Cell 54: 739-740.
GUTZ, H., H. HESLOT, U. LEUPOLD and N. LOPRIENO, 1974 Chapter 25 in Handbook of Genetics, edited by R. C. KING. Plenum Press, New York.
HAYLES, J., D. BEACH, B. DURKACZ and P. NURSE, 1986 The fission yeast cell cycle control gene cdc2; isolation of a sequence sucl that suppresses cdc2 mutant function. Mol. Gen. Genet.
HINDLEY, J., G . PHEAR, M. STEIN and D. BEACH, 1987 sucl+ encodes a predicted 13-kilodalton protein that is essential for cell viability and is directly involved in the division cycle of Schizosaccharomycespombe. Mol. Cell. Biol. 7: 504-51 1 .
KISHIMOTO, T., and H. KANATANI, 1976 Cytoplasmic factors re- sponsible for germinal vesicle breakdown and meiotic matura- tion in starfish oocyte. Nature 2 6 0 321-322.
KOHLI, J.. 1987 Genetic nomenclature and gene list of the fission yeast Schizosaccharomyces pombe. Curr. Genet. 11: 575-589.
LEE, M., and P. NURSE, 1987 Complementation used to clone a human homolog of the fission yeast cell cycle control gene cdc2+. Nature 327: 31-35.
LOHKA, M. J., and J. L. MALLER, 1985 Induction of nuclear envelope breakdown, chromosome condensation, and spindle formation in cell-free extracts. J. Cell Biol. 101: 518-523.
MARKS, J., and J. S. HYAMS, 1985 Localization of F-actin through the cell division cycle of Schizosaccharomyces pombe. Eur. J. Cell Biol. 39: 27-32.
MASUI, Y., and C. L. MARKERT, 1971 Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J.
3: 3 17-325.
746-754.
202: 291-293.
EXP. ZOO^. 177: 129-146.
MIAHE-LYE, R., and M. W. HIRSCHNER, 1985 Induction of early mitotic events in a cell-free system. Cell 41: 165-175.
MIAKE-LYE, R., J. NEWWRT and M. KIRSCHNER, 1983 Maturation-promoting factor induces nuclear envelope break- down in cycloheximide-arrested embryos of Xenopus laeuis J. Cell Biol. 97: 81-91.
NELKIN, B., C. NICHOLS and B. VOGELSTEIN, 1980 Protein fac- tor($ from mitotic CHO cells induce meiotic maturation in Xenopus laevis oocytes. FEBS Lett. 1 0 9 233-238.
NURSE, P., 1975 Genetic control of cell size at cell division in fission yeast. Nature 2 5 6 547-55 1.
NURSE, P., and Y. BISSETT, 1981 Gene required in GI for com- mitment to cell cycle and in Gz for control of mitosis in fission yeast. Nature 292: 558-560.
NURSE, P., and P. THURIAUX, 1980 Regulatory genes controlling mitosis in the fission yeast Schizosaccharomyces pombe. Genetics
NURSE, P., P. THURIAUX and K. NASMYTH, 1976 Genetic control of the cell division cycle in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 146: 167-178.
OGDEN, J. E., and P. A. FANTES, 1986 Isolation of a novel type of mutation in the mitotic control of Schizosaccharomyces pombe whose phenotypic expression is dependent on the genetic back- ground and nutritional environment. Curr. Genet. 10: 509- 514.
RUSSELL, P., and P. NURSE, 1986 cdc.25' functions as an inducer of mitotic control of fission yeast. Cell. 45: 145-153.
RUSSELL, P., and P. NURSE, 1987 Negative regulation of mitosis by weel', a gene encoding a protein kinase homolog. Cell 4 9 559-567.
SOLOMON, M., R. BOOHER, M. KIRSCHNER and D. BEACH, 1988 Cyclin in fission yeast. Cell 54: 738.
SORENSON, R. A., M. S. CYERT and R. A. PEDERSEN, 1985 Active maturation-promoting factor is present in mature mouse oo- cytes. J. Cell Biol. 100: 1637-1640.
SUNKARA, P. S., D. A. WRIGHT and P. N . RAO, 1979 Mitotic factors from mammalian cells induce germinal vesicle break- down and chromosome condensation in amphibian oocytes. Proc. Natl. Acad. Sci. USA. 7 6 2799-2802.
SWENSON, K., K. M. FARRELL and J. V. RUDERMAN, 1986 The clam embryo protein cyclin A induces entry into M phase and the resumption of meiosis in Xenopus oocytes. Cell 47: 861- 870.
TACHIBANA, K., N. YANAGISHIMA and T . KISHIMOTO, 1987 Preliminary characterization of maturation promoting factor from yeast Saccharomyces cerevisiae. J. Cell Sci. 88: 273- 282.
UMENSONO, K., T . TODA, S. HAYASHI and M. YANAGIDA, 1983 Two cell division cycle genes NDAZ and NDA3 of the fission yeast Schizosaccharomyces pombe control microtubular organization and sensitivity to anti-mitotic benzimidazole com- pounds. J. Mol. Biol. 168: 271-284.
WEINTRAUB, H., M. BUSCAGLIA, M. FERREZ, S. WEILLER, A. Bou- LET, F. FABRE and E. E. BAULIEU, 1982 Mise en evidence d'une activite "MPF" chez Saccharomyces cereuisiae. C. R. Acad. Sci. (Paris) 295: 787-790.
WRIGHT, A , , K. MAUNDRELL, W. D. HEYER, D. BEACH and P. NURSE, 1986 Vectors for the construction of gene banks and the integration of cloned genes in Schizosaccharomyces pombe and Saccharomyces cerevisiae. Plasmid 15: 156- 158.
YOUNG, P. G., and P. A. FANTES, 1987 Schizosaccharomycespombe mutants affected in their division response to starvation. J. Cell Sci. 88: 295-304.
9 6 627-637.
Communicating editor: D. BOTSTEIN





























