Upload
others
View
0
Download
0
Embed Size (px)
Citation preview
Perception & Psychophysics2000, 62 (2), 386-401
Categorical perception occurs in newly learnedfaces, other-race faces, and inverted faces
DANIEL T. LEVINKent State University, Kent, Ohio
and
JAMES M. BEALEGrand Valley State University, Allendale, Michigan
On the basis of findings that categorical perception (CP) is possible in complex visual stimuli suchas faces, the present study tested for CP on continua between unfamiliar face pairs. Results indicatethat CP can be observed for unfamiliar faces, in both familiar (same-race) and unfamiliar (other-race)groups. In addition, significant CP effects were observed in inverted faces. Finally, half-continua weretested where midpoint stimuli became endpoints. This was done to ensure that stimulus artifacts didnot account for the observed CP effects. Consistent with the perceptual rescaling associated with Cp,half-continua showed a rescaled CP effect. Weargue that these CP effects are based on the rapid ac-quisition of perceptual equivalence classes.
Categorical perception (CP) occurs when discrimina-tions between stimuli within one category are more dif-ficult than discriminations between stimuli that span theboundary between categories. The question is, under whatconditions does this occur? Initial research suggested thatCP would be observed only for naturally occurring con-tinua and was perhaps a sign of innate or highly practicedpredispositions to break these continua into equivalenceclasses (e.g., Liberman, Harris, Kinney, & Lane, 1961).For example, Etcoff and Magee (1992) observed CP oncontinua between different facial expressions and sug-gested this supports the hypothesis that the perception offacial expression is innate. However, other findings sug-gest that such predispositions are not necessary to ob-serve CP. In another study using faces, Beale and Keil(1995) found that participants perceive continua betweenindividual familiar faces categorically. This is the casedespite the likelihood that the continua between thesefaces have never been experienced by participants.
IfCP is possible along artificial continua, then hypothe-ses assuming that CP is a sign of innate or even highly fa-miliar perceptual categories are incorrect. Here, we ex-plore two basic questions about the degree to which CPcan be observed in the absence ofprevious learning. First,will it be possible to observe CP on novel continua be-tween endpoints that do not have long-standing represen-
The authors thank Robert Goldstone and Bill Merriman for readingdrafts of this report and Stevan Hamad and Andy Youngfor many help-ful discussions. Correspondence should be addressed to D. T. Levin,Kent State University, Department of Psychology, Kent Hall, Kent, OH44121-0001 (e-mail: [email protected]). J. M. Beale can be reached atthe Department of Psychology, Grand Valley State University, Allen-dale, MI 49401 (e-mail: [email protected]).
-Accepted by previous editor, Myron L. Braunstein
tations associated with them as do familiar faces? Anumber of recent findings suggest that this kind of rep-resentation is not a necessary precondition for CPO Thisbrings up the possibility that CP might be observed oncontinua between newly learned faces. Second, ifCP oc-curs on novel continua between novel endpoints, thenunder what conditions will such effects be found? In thispaper, we explored CP in newly learned faces and manip-ulated the degree to which face-specific coding processeswere available by using faces from unfamiliar races andinverted faces.
Categorical Perception: The Basic Phenomenonand Reasons to Expect It in Unfamiliar Faces
The phenomenon of CP is interesting for a number ofreasons, but perhaps the most basic is that CP involvescognitive and perceptual equivalence classes that affectour ability to perceive continuous variation among stim-uli. In the basic CP paradigm, a set of stimuli are createdthat vary continuously along a single physical dimen-sion. The simplest example is the color spectrum, whichvaries continuously in terms ofwavelength. Despite con-tinuity along the physical dimension of wavelength, weperceive a series of discrete color categories. Empiricalverification of this impression usually comes from linkedclassification and discrimination tasks. The classifica-tion task, in which participants classify a set of stimuliusing whatever labels are appropriate, determines the lo-cation of the boundaries between categories. For exam-ple, participants might be presented with a series of IIhues ranging from 500 nm (red) to 600 nm (yellow) inl O-nm steps and might be asked simply to label each"red" or "yellow." Label assignments can then be used todetermine the boundary between red and yellow. Oncethis boundary is known, ifthese stimuli are perceived cate-
Copyright 2000 Psychonomic Society, Inc. 386
gorically, one would predict more accurate discrimina-tion between members of stimulus pairs that straddle theboundary between the colors, whereas discriminationbetween stimuli that lie within one of the regions will beless accurate (e.g., see Grahm, 1965). Facilitated discrim-ination accuracy between stimuli that straddle categoryboundaries implies that cognitive and perceptual pro-cesses act to divide physical stimulus continua into dis-crete equivalence classes. Differences among variants fall-ing within a class are psychologically attenuated, whereasdifferences among variants that fall in a region betweenclasses are amplified.
Initially, research on CP was confined to spoken lan-guage perception, where it appeared as though the con-tinua between different phonemes were broken up by aninnate language-specific process. For example, there ex-ists a clear continuum between the phonemes Ibl and Iplthat can be defined in terms of the difference between theonset of voicing and the plosive burst. This difference isreferred to as voice onset time (VOT). Ifvoicing and the"burst" are simultaneous (VOT = 0), the phoneme Ibl isperceived. If the VOT is greater than approximately30 msec, the percept is Ip/. It is therefore possible to cre-ate a continuum between these two phonemes by simplyvarying the VOT from 0 to 80 msec in 10-msec incre-ments using a voice synthesizer. Each stimulus can thenbe paired with other stimuli on the continuum in a dis-crimination paradigm to test for differences in discrim-inability across the continuum. In the case of Ibl and Ip/,pairs of stimuli that straddle the category boundary (forexample, a 20-msec VOT vs. a 40-msec VOT) are easierto discriminate than pairs both falling to one side of the30-msec boundary (Liberman, Cooper, Shankweiler, &Studdert-Kennedy, 1967).
At first, CP was observed only with these stimuli, andnot other language-like stimuli such as "inverted" syn-thetic phonemes (e.g., the high-frequency and low-frequency formants were switched and their envelopesinverted; Liberman et al., 1961). This research was takento support the hypothesis that CP was produced either byan innate perceptual screening mechanism particular tolanguage that enhances the contrast between similar pho-nemes (Stevens, 1981) or by a perceptual inference basedon the motor patterns required to make speech (Libermanet al., 1967). More recently, Etcoffand Magee (1992) ar-gued that CP on continua between different facial expres-sions was similarly indicative ofinnate expression-specificperceptual mechanisms. For example, a continuum wascreated between a happy expression and a disgusted ex-pression worn by the same individual. The presence ofCP along this continuum was interpreted by the authorsto imply that the particular facial expressions at each endof the continuum are specified innately. If one's theoret-ical preference is to avoid nativistic extrapolation, thenthe hypothesis that facial expressions are simply over-learned facial configurations also fits these findings. Thekey hypothesis in the above research is that CP results
CATEGORICAL PERCEPTION 387
from specific long-standing representations that specifythe continuum endpoints and, according to Etcoffand Ma-gee, the actual continua themselves.
A number ofexperiments show, however, that CP doesnot necessarily imply innate programming, or even over-learning of the relevant stimuli. First, Kuhl (1981) foundthat chinchillas produced CP effects for the same phonemecontinua that had been tested in humans, reproducing theexact VOT boundaries between different phoneme pairsas found in humans. This finding, in concert with otherson infrahuman species (Kuhl & Padden, 1982), castdoubt on the presumption that CP must necessarily reflectspecific inborn phoneme boundaries. Kuhl (1987) insteadsuggests that low-level perceptual properties of the con-tinua themselves contain naturally information-rich re-gions that the evolution of language has selected as di-viding lines between phonemes.
In addition, the original findings that auditory CP waslimited to speech sounds has been reversed. For example,Sawusch and Gagnon (1995) observed CP in nonspeechphoneme analogs. In this case, the critical manipulationthat allowed CP was a task in which the entire stimuluscontinuum was played before alternate discrimination tri-als. Also, Burns and Ward (1978) found that music ex-perts perceive musical intervals categorically. Smith,Kemler Nelson, and Appleton (1994) found that even mu-sical novices could perceive continua between two-toneintervals categorically if each interval was associated witha well-known folk song (the subjects were informed thatone tone interval was the first two notes of one song andthe other represented the first two notes ofanother song).Thus, expertise allows CP to occur in novel unlabeledstimuli that mimic the practiced domain. Even where per-ceivers are less expert, if stimulus continua can be an-chored by familiar endpoints, then CP is again observed.
The question is, how do Beale and Keil's (1995) find-ings fit with findings in music experts? They found CPon continua between familiar faces, but not unfamiliarfaces. On the basis of this finding, they argue that CP isdriven not by innate perceptual continua but rather bylearned representations ofthe invariant structure ofknownfaces. However, they reported no CP effects for unfamil-iar faces, which appears to conflict with the above data onmusical experts' perception of musical intervals. In thedomain of music, expertise was associated with an abil-ity to immediately represent new stimuli such that con-tinua between them were perceived categorically. Noviceswere different in that they had to be provided with a di-rect mapping from the test stimulus to familiar music ex-emplars for CP effects to be found. In the domain offaces,it is safe to say that most adults are experts, but Beale andKeil's results make face experts similar to music novicesin that neither showed CP effects on continua betweennovel exemplars.
Perhaps the conflict between the music expert dataand Beale and Keil's (1995) face data can be resolved byspecific aspects of their task. The experiment began with
388 LEVIN AND BEALE
an instruction screen containing the names and imagesof the eight endpoint faces. Although four of the faceswere well known and therefore probably required littlelearning, the other four were unfamiliar to most partici-pants and therefore would have required at least somelearning. Where they manipulated familiarity, Beale andKeil randomly mixed discrimination trials from continuabetween all eight faces within a single block. This inter-mixing might account for the lack of CP effects for theunfamiliar faces. If sufficient familiarity with novel facesmust be built up during the discrimination trials, thenBeale and Keil's task may not have allowed sufficient ex-perience with each of the face continua to adequatelyrepresent all of the endpoints, especially for the novelstimuli. This possibility is supported by recent findingsfrom Stevenage (1998), who had subjects complete ashort-training task in which they learned to differentiatea pair of identical twins by classifying pictures of eachwith feedback. After the task, participants gave highersimilarity ratings to different pictures of the same twinand lower ratings to pictures ofdifferent twins. The sim-ilarity ratings therefore suggested a CP effect based onshort-term learning ofpreviously unfamiliar faces. How-ever, none of the above findings imply that long-term fa-miliarity is unrelated to observing CPO Rather, the impli-cation is that prior familiarity facilitates CP by eliminatingthe necessity that continuum endpoints be learned atsome point during the experiment or on line during theactual task.
The present research therefore used a methodologysimilar to that of Beale and Keil (1995), with the excep-tion that unfamiliar face pairs were presented from onlyone continuum in any given block of trials. If CP is pos-sible with novel visual stimuli, then it may appear for un-familiar faces if the learning task is made easier.
Testing the Limits of CP in FacesIf CP can be observed on novel continua and between
novel endpoints, then it is important to ask about the ex-tent of the phenomenon. It is possible that it will be ob-served on just about any continuum with sufficiently dis-criminable endpoints. Therefore, extensive expertisewith a given stimulus domain may not be necessary toobserve CPo For example, Goldstone (1994) found that itwas possible to observe CP-like effects using continuaof artificial visual stimuli given a small amount of clas-sification training. The present experiments assessedthe limits of CP in two ways. First, they included con-tinua between faces from a less familiar race (Blackfaces for the White and Asian participants tested here),in addition to a more familiar race (White faces). Alsoincluded were continua that traverse the races. Given thewell-known other-race recognition deficit (e.g., seeShapiro & Penrod, 1986) whereby participants recognizefaces from an unfamiliar race less well than those from afamiliar race, it is straightforward to predict that no CPwill be observed on continua between Black faces if thefull compliment of face coding skills is a necessary pre-condition.
An additional test for the limits of CP was included inExperiment 2 in which all faces were presented upsidedown. This common manipulation is thought to differen-tially disrupt coding subtle variations in the configurationof facial features (Diamond & Carey, 1986; Farah, Ta-naka, & Drain, 1995; Rhodes, Brake, & Atkinson, 1993).This configural coding process is considered the hall-mark of face coding and is probably the source of ourability to accurately differentiate members of this ho-mogeneous class (Diamond & Carey, 1986; Rhodes, Tan,Brake, & Taylor, 1989). Therefore, if CP depends onexpertise-based configural coding, any CP effects thatappeared in Experiment I were expected to disappear inthe inverted faces in Experiment 2.
Summary and PredictionsThe goal of the present experiments was to improve
our understanding of the CP phenomenon in complex vi-sual stimuli such as faces by testing for CP on continuabetween unfamiliar faces. In Experiment 1, continua be-tween pairs of faces were tested for the presence of CPOIn this experiment, discrimination trials were presentedin blocks including stimuli from a single continuum. Inaddition, continua within familiar and less familiar racialcategories were included along with continua that spannedthe two categories. In Experiment 2, the same stimulusset and procedures were used but the faces were inverted.This allowed a test for CP in the absence of expertise-induced configural coding. Finally, Experiment 3 was de-signed to eliminate the possibility that stimulus artifactswere responsible for the positive findings in Experi-ments 1 and 2.
EXPERIMENT 1
Experiment 1 was a standard CP paradigm based onthe methodologies used in Etcoff and Magee (1992) andBeale and Keil (1995). Twolinked tasks were used in thisexperiment: a discrimination task and a categorizationtask. The discrimination task was similar to one used byBeale and Keil in which participants viewed pairs offaces and were asked to respond which was more simi-lar to one of the endpoint images. This task was used inplace ofthe more traditional ABX paradigm primarily tofocus participants' attention on the association betweena given name and a face representation. In the categoriza-tion task, participants categorized each face used in thediscrimination task according to their similarity to one orthe other endpoint. This task therefore provided an esti-mate of the category boundary that was used to predictwhich pairs in the discrimination task should show im-proved performance. Pairs that straddle this boundaryshould show improved discrimination relative to the otherface pairs.
In this experiment, and in all of the following experi-ments reported here, each participant completed dis-crimination tasks along three different continua: one withtwo Black faces as endpoints, one with two White end-points,and one with one Blackendpointand one White end-
point. Each continuum was presented in a different blockof trials, and four different sets of three face pairs wereused across different participants.
MethodParticipants. A total of 17 participants completed Experiment I.
One participant who did not consistently classify the faces with oneof the endpoints was dropped from the analysis. Of the 16 remain-ing participants, I was Asian and IS were White. The participantscompleted the experiment in exchange for extra credit in a psy-chology course.
Stimuli. Sixteen unfamiliar male faces (8 Black and 8 White)were used as stimuli in this experiment. These were the same unfa-miliar faces used in Levin (1996) and were equated for average dis-tinctiveness (and rated familiarity) across the races. The 8 faceswithin each race were paired to make 4 continua within each race.In addition, 4 of the faces in each race were paired across race cat-egories for a total of 12 face pairs. To equalize similarity amongfaces in each ofthe three groups (Black-Black, White-White, andBlack-White continua), 10 judges rated the similarity of all possi-ble pairings among the 16 faces. Judges were instructed to rate sim-ilarity based on the overall similarity offace shape using a 7-pointscale (I = very similar, 7 = very dissimilar) and were further in-structed to ignore differences in race when viewing other-race pairs.On the basis of these ratings, sets offace pairs were chosen to equal-ize average similarity ratings between the three stimulus groups(mean similarity ratings: Black-Black pairs, 4.15; White-White pairs,4.08; Black-White pairs, 4.05; SD for all chosen pairs = 0.5823).
CATEGORICAL PERCEPTION 389
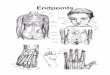
Once the faces were paired, a set of 9 intermediate stimuli wascreated for each pair (using the morph program from Gryphon Soft-ware) by linearly interpolating the shape and luminance values on10% intervals between the endpoint averages (see Beale & Keil,1995, for details). Together with the continuum endpoints, the par-ticipants viewed II different stimuli representing controlled blendsof the two faces (see Figure I). As a matter of convention, 0% willbe assigned to one end ofthe continuum and 100% to the other withintermediates described in terms of percentage locations on thisscale. Therefore, the 50% stimulus is the intermediate that sits in themiddle ofthe continuum. In the case ofthe Black-White (B-W) con-tinua, the scale always begins with the Black face (0%) and endswith the White face (100%). On the White-White (W-W) and Black-Black (B-B) continua, the 0% and 100% faces were arbitrarily cho-sen. However, as will become important later, the instruction screenscontaining the faces and names associated with them always hadthe 0% face on the left and the 100% face on the right.
Once the intermediates were created, all images were blurredslightly (0.5-pixel-radius Gaussian blur using Photoshop 2.0) to re-duce the visibility of small image defects and interference patternscaused by scanning. Each image was approximately 240 X 336 pixels(horizontal X vertical) and was presented in 256-level gray scale.
Apparatus. The experiment was presented on Macintosh LCcomputers attached to 12-in. color monitors set to 8-bit gray scalemode. Stimuli were viewed from approximately 60 em, and mea-sured approximately 8.5 X 12.5 em on the monitor.
Procedure. The participants first completed three blocks ofdis-crimination trials, one each with B-B, W-w, and B-W continua.Before each block, the participants viewed an instruction screen
B-B
B·W
W·W
0% End1000/0 Start
200/0 End800/0 Start
40% End60% Start
60% End400/0 Start
8004End20% Start
100% End0% Start
Figure 1. Sample set of stimuli. 8--B continua have two Black faces as endpoints, 8--W continua start at a Black face and end ata White one, and W-W continua have two White faces as endpoints. Each type (8--B, W-W, and 8--W) was represented by fourdifferent continua with eight different faces as endpoints.
390 LEVIN AND BEALE
that had the two endpoint faces, each associated with an arbitrarilychosen name (e.g., "Fred"). The participants were instructed to payclose attention to this screen and to be sure that they could identifyeach face and associate the correct name with it. On each trial ofthediscrimination task, the participants saw a response cue telling themto indicate which of the upcoming faces looked more like one oftheendpoints. For example, if one of the endpoint faces was named"Fred," the participants might be asked, "Which of these faceslooks more like Fred?" After reading the cue, the participants hit thespace bar on the computer keyboard indicating their readiness tocontinue. They then saw two faces side by side for 1,000 msec. In allcases, the two faces in the stimulus display were separated by twosteps, or 20% of the total continuum. After the faces disappeared,the participants responded by pressing the" I" key if the left facelooked more like the target face or the "2" key ifthe right face did.In each block, there were 8 trials (two trials for each screen side andcorrect response) for each ofthe 9 face pairs along the continua fora total of 72 trials. The order of discrimination blocks was counter-balanced across participants such that half saw the other-race pairfirst and halfsaw one of the within-race pairs first and the other sec-ond. The order of the two within-race pairs was also counterbalanced.
After completing the discrimination trials, the participants com-pleted three blocks of classification trials. The order of classificationblocks followed the same order as the discrimination blocks foreach participant. In each block, the participants classified individ-ual faces along one of the continua from the discrimination trials.At the beginning of each block, the two endpoint faces were pre-sented, again paired with their names. The participants were in-structed to hit the "I" key if they thought an individual face wasmore similar to the first face or to hit the "2" key if it was more sim-ilar to the second face. Each of the II faces was presented threetimes for a total of33 trials in each of the three blocks.
ResultsFor all analyses reported in this paper, data were col-
lapsed across the four continua in each condition.Classification task. The classification task allowed for
selection of cross-boundary discrimination pairs usingthe same criterion as in Beale and Keil (1995) and Etcoffand Magee (1992). Pairs including one individual faceclassified with one of the endpoints on more than 66%oftrials and another classified with the other endpoint onmore than 66% oftrials were selected as cross-boundarypairs to be compared with the other pairs that were con-sidered within-category pairs.
By the above criteria, the 40%-60% pair was selectedas the cross-boundary pair for the global analysis for allcontinuum types (B-B, W-W, and B-W; see Figure 2).This pair was also selected for individual analyses of theB-Band B-W continua. For the W- W continua, boththe 40%-60% and 50%-70% pairs straddled the 66%-33% boundary, so both were selected to represent cross-boundary discrimination accuracy (de Gelder, Teunisse,& Benson, 1997).
Discrimination task. Data from the discriminationtask were entered into one single-factor (AB Pair; 1%-20% to 80%-100%) analysis of variance (ANaYA) totest for CP across all continua in addition to three sepa-rate ANOVAs,one for each condition (B-B, B-W, W-W).To test for significant categorical perception effects,planned contrasts were computed between cross-boundarypairs and within category pairs to test (using one-tailed
t tests) for increased discrimination accuracy on the cross-boundary pair.
The overall CP effect was highly significant [F( 1,15) =25.005, MSe = 0.004,p = .0001]. In addition, significantcategorical effects were observed for all three continuumtypes (see Figure 2). For the B-W and W-W continua,cross-boundary pairs were more accurately discriminatedthan within-boundary pairs [B-W, F(1,15) = 14.775,MSe = 0.008,p = .0008;W-W,F(1,15) = 28.782,MSe =0.0 II, p < .000 I]. The categorical effect was smaller butalso significant for the B-B condition [F(1, 15) = 3.609,MSe = 0.015, p = .0385].
Average discrimination accuracy was 81.9% in theB-B condition, 84.3% in the B-W condition, and 82.3%in the W-W condition. An ANOVA revealed no signifi-cant effect ofcontinuum type on discrimination accuracy(F < 1).
Finally, as Figure 2 makes clear, discrimination accu-racy was higher at the "beginning" of the continua thanat the "end." Duncan tests indicate that discrimination ac-curacy was significantly lower for the 0%-20% pair thanfor the 80%-100% pair (p < .05) and also lower for the10%-30% pair than for the 70%-90% pair (p < .0 I). Alook at the individual continuum types shows that theB-B and W-W continua drive this effect. For the B-Bcontinua, accuracy was significantly lower for the 10%-30% pair than for the 70%-90% pair (p < .05); for theW-W continua, accuracy was lower for both the 0%-20% pair and the 10%-30% pair than for the 70%-90%and 80%-100% pairs, respectively (p < .0 I). None ofthese comparisons was significant for the B-W continua.
DiscussionIn Experiment I, face pairs on all continua showed a
significant categorical effect, although the effect wassmaller for the B-B pairs. These findings suggest thatprevious familiarity with continuum endpoints is unnec-essary to observe CPO Instead, so long as participants geta chance to familiarize themselves with the stimuli, asufficient representation is available that allows parsingof a visual continuum into equivalence classes.
One issue that arises when looking at Figure 2 is thatno clear sharp peak in discrimination accuracy was evi-dent for the 40%--60% stimulus pair. Instead, there appearsto have been a gradual increase in accuracy as stimuluspairs approached the category boundary. Furthermore,discrimination accuracy was far above chance through-out the continuum; even the endpoint pairs were dis-criminated with 68%-75% accuracy. Both of these find-ings might be seen to conflict with "pure" CP wherebydiscrimination is only possible to the extent that stimuluspairs cross the category boundary. Therefore, discrimi-nation accuracy within the perceptual categories shouldbe at chance and should only increase above that levelwhen one member of the pair falls into one category andthe other member falls into the other category. This kindof finding is, however, not the norm, even in the case ofphoneme discrimination (for review, see Pastore, 1987).
CATEGORICAL PERCEPTION 391
Classification Task
All Continua Individual Continuum Types
100 100
'E 'E·0 "0 80a. 80 a.'a 'ae t::W W
III 60 '" 60as '"'a 'aGI GI~ ;:: 40"ii 40 ·iiiIII IIIas asU U
B-B'E 20 'E 20• GI B-Wu u WoW.. ..GI •a. a. 00
I!-I!-I!-I!-I!-I!-I!-I!-I!-I!-I!- I!-I!-I!-I!-I!-I!-I!-I!-I!-I!-I!-
=========== ===========... Nf'l ..... IIl~I--QllCl\= ... Nf'l ..... IIl~I--QllCl\=... ...Stimulus Stimulus
Discrimination Task
Individual Continuum Types100.,.------------,
All Continua100 ...------,.-------,
90 90.. ..() ()ell ell... ...... ... 800 80 00 0.. ..c Cell 70 ell 70() f:!...ell ella. a. --- B-B60 60 ----.-- B-W
---0-- WoW
5050l!- I!- l!- I!- l!- I!- l!- I!- l!- I!- l!- I!- l!- I!- l!- I!- l!- I!-= = = = = = = = = = = = = = = = = =N f'l ..... III ~ I-- Qll CI\ =N f'l ..... III ~ I-- Qll CI\ = ==Q Q Q Q Q
. ...Q Q Q Q Q Q Q Q ... = I... N f'l ..... III ~ I-- Q ... N f'l ..... III ~ I-- =ee Qll
Pair Pair
Figure 2. Classification and discrimination data from Experiment 1. The x-axis of the classi-fication graph represents the continua, including both endpoint stimuli (0% and 100%) and in-termediates (10%-90%), and the x-axis of the discrimination graph represents stimulus pairsspanning the continua. The 0% and 100% endpoint designations are arbitrary for the W-W andB-B continua. For the B-W continua, the 0% endpoint is the Black face of the pair, and the100% endpoint is the White face. The classification graph contains the percentage oftrials onwhich each stimulus was classified in the same category as the 100% (endpoint) stimulus. Thediscrimination graph contains discrimination accuracy for stimulus pairs along each contin-uum. The vertical line in the discrimination graph for all continua represents the pair crossingthe category boundary.
Instead, discrimination accuracy is usually affected bysome combination offactors. For example, Pisoni (1975)describes a model ofCP that makes explicit the combinedinputs of both categorical effects and perceptual effects(such as comparisons of iconic or echoic sensory traces)to determine discrimination accuracy throughout a con-
tinuum. Therefore, above-chance discrimination withincategories and unsharp peaks in accuracy are probablycaused by noncategorical inputs to discrimination thatare overlaid on the CP effect.
In this context, it is interesting to discuss the asym-metry evident in Figure 2 whereby discrimination accu-
392 LEVIN AND BEALE
racy was higher at the "ends" of the continua than at their"beginnings." Although the between-race continua alwaysbegan with the Black face and ended with the White face,the within-race continua were not intended to have anysystematic difference between starting points and end-ing points. However, as noted in the Method section, theinstruction screen in which each face pair was learnedhad the starting-point stimulus on the left side and theendpoint stimulus on the right. It is possible that this po-sitioning in some way specified a relationship betweenthe faces that caused increased discrimination accuracyfor the right-side faces. One possibility is that the face onthe left served as the "base" for comparison with the faceon the left (Medin, Goldstone, & Gentner, 1993). In thecontext oflanguage, a sentence can specify a comparisonbetween two concepts such that one, the "base," speci-fies the features relevant to the comparison, whereas the"target" is evaluated with reference to these features. Ifsuch a situation has occurred in the present case, the ar-rangement of the faces may have caused features of theface on the right to be coded relative to those on the left.This kind of selection could account for the increase indiscrimination accuracy for variants of this face over thatfor the face on the left. While these possibilities are poten-tially interesting, a resolution of these issues will be re-served for future exploration and not further discussed here.
Before going on to Experiment 2, one potentially prob-lematic aspect of the specific task used here needs to beaddressed. On each trial, one of two stimuli must bematched with a representation of one endpoint face orthe other. Therefore, on a subset of trials, the participantswere asked which of two versions ofone face looked likethe other. For example, the participants might be askedwhich of the upcoming faces looks most like "John" andthen be shown two faces that plainly look like "Fred."These trials might be particularly difficult because thespecified target is a face at one end of the continuum,whereas the presented stimuli are from the other end. Ifso, then the observed CP effect might be based on reducedaccuracy for these trials, especially because the problemwould be particularly acute for stimulus pairs close toeach endpoint. To eliminate this concern, the data weredivided according to the relationship between the targetspecification and the discrimination stimuli. One analy-sis included only trials on which the target (X) and dis-crimination (A and B) stimuli were from the same side("near-X" trials) of the continuum, and the other in-cluded only trials for which the target was the endpointopposite the discrimination stimuli ("far-X" trials). If thetask artifact caused the CP effect, then it should only besignificant for the far-X trials. However, this is plainly notthe case. The CP effect is significant in both cases and is,in fact, stronger for the near-X trials [far-X, F(I,15) =12.016,MSe = 0.005,p = .003;near-X,F(I,l5) = 25.137,MSe = 0.005, P = .0002]. The implications of this par-ticular pattern of results will be examined further in theGeneral Discussion section.
The significant CP effect observed in other-race facessuggests that CP in unfamiliar faces generalizes to facesfrom unfamiliar groups. Given findings by Fallshore andSchooler (1995) and Rhodes et al. (1989) implying thatconfigural coding is not present or reduced in other-racefaces, it is possible that CP does not depend on the avail-ability ofthis process. This runs counter to recent researchsuggesting that configural coding underlies the CP ef-fect (Campbell, Pascalis, Coleman, Wallace, & Benson,1997; de Gelder et aI., 1997). In both cases, CP was ob-served in upright faces and disappeared when the faceswere inverted. This finding suggests the importance ofcon-figural coding because inversion has been found to sub-stantially disrupt expertise-based coding ofthe relation-ships among features (Farah et aI., 1995; Rhodes et a!.,1993; Tanaka & Farah, 1993).
Therefore, Experiment 2 tested the limits ofCP by us-ing inverted faces. IfCP remained in inverted faces, thenit is likely that configural coding was not necessary toobserve this effect.
EXPERIMENT 2
MethodA total of 14 White participants completed Experiment 2 in ex-
change for course credit. Stimuli and procedures were identical tothose in Experiment I with the exception that all faces (includingthe ones in the sample screens) were inverted by rotating them 180°.
ResultsClassification task. Using the same criterion as in
Experiment I, the 40%-60% pair was again selected asthe cross-boundary pair for all conditions (see Figure 3).
Discrimination task. The analysis was identical to thatin Experiment I. The test for CP across all continua wasagain highly significant [F(I,13) = 12.376, MSe = 0.008,p = .0019]. Categorical effects were significant for theB-W continua [F(1,13) = 6.713, MSe = 0.016,p = .0112and the W-W continua [F(1,13) = 4.125, MSe = 0.026,p = .0316] but nonsignificant for the B-B continua(F< I) (see Figure 3).
Average discrimination accuracy was 74.2% in the B-Bcondition, 78.3% in the B-W condition, and 76.6% inthe W-W condition. None ofthe differences between con-ditions were significant, but discrimination accuracy was,overall, lower in Experiment 2 than in Experiment I [76.4%vs. 82.8%; F(I,28) = 5.497, MSe = 0.017,p = .0264].
DiscussionExperiment 2 demonstrated CP in inverted faces, sug-
gesting that configural coding processes are not a pre-requisite to observe CPO As is evident from comparing Fig-ures 2 and 3, inversion seems to have reduced the size ofthe CP effect in that discrimination accuracy for endpointswas minimally reduced, whereas that for stimulus pairsin the middle of the continua was lower. Therefore, itseems reasonable to conclude that configural coding might
~~~~~~~~~~~
394 LEVIN AND BEALE
Original Continuum
Figure 4. Predicted findings for half-continua assuming CP (A)or an artifact (B). IfExperiments 1 and 2 revealed CP-like resultsbecause midregion stimuli contain artifacts that make them easyto discriminate, then redoing the experiment with half-continua(in which the stimulus originally serving as a midpoint is now anendpoint) should produce no CP effect. Instead, AB pairs thatwere originally in midregions should still be most accuratelydiscriminated.
were from half ofa given continuum from Experiments Iand 2. Thus, one face was originally an endpoint, but theother was the midpoint of the original continua. If the cat-egorica� effect we have been observing is simply a stim-ulus artifact, accuracy should be high for stimulus pairsthat originally straddled midpoints (which we will here-after refer to as switch pairs) and low for all original end-points. This is illustrated in Figure 4B, where accuracyon the half-continua simply mirrors that of the originalcontinua. On the other hand, if the categorical effect re-sults from the rapid acquisition ofon-line representationsof the endpoints, then a newly scaled CP effect should beobserved where midregion stimuli that were originallythe most accurately discriminated are now less accuratelydiscriminated than the new midregion stimuli (see Fig-ure 4). This hypothesis is clearest for the B-B and W-Wcontinuum types, for which the half-continua bear norelationship to specific categories. For the B-W half-con-tinua, the switch pairs straddle the between-race bound-ary because they are always the middle of a continuumacross race categories. Therefore, although the B~Wmayshow rescaled CP effects, the switch pairs may well beaccurately discriminated for nonartifact reasons.
Two separate discrimination conditions were run inExperiment 3: a "2-step" condition in which face pairswere separated by 33% of the half-continua (or 20% ofthe original continua and therefore identical in terms ofphysical difference to the stimulus pairs in Experiment I),and a "l-step" condition in which face pairs were separatedby 17% ofthe half-continua (or 10% ofthe original con-tinua). In all cases, individual stimuli were presented up-right and were identical to those used in Experiment I.
ArtifactCategorical Perception
A
Although it is possible that CP can occur based onshort-term learning, its presence in within-race continuaand inverted faces does raise a number of concerns-most importantly, the possibility that the effect is a stim-ulus artifact. Whenever testing for CP between two com-plex visual stimuli, one must consider the possibility thatthe effect might be driven by accidental increases in theperceptual differences between midregion stimuli. Forexample, perhaps some aspect of the morphing processcauses midregion stimuli to have a large number ofsmalldiscontinuities or other artifacts that participants can useto enhance discrimination accuracy. In such a case, in-creased disciminability at category boundaries would beuninteresting. Although we attempted to reduce suchconcerns both by using Beale and Keil's (1995) "better-likeness" task and by slightly blurring the stimuli, neitherprecaution completely eliminated the possibility that ar-tifacts are driving our CP effects. Experiment 3 wastherefore designed to provide a critical test of the hy-pothesis that artifacts are driving our findings.
EXPERIMENT 3\
Experiment 3 was designed to test for artifacts by pre-senting participants with half-continua. In each of threeblocks of trials, participants were shown face pairs that
MethodParticipants. A total of 41 participants completed the discrimi-
nation and categorization tasks in exchange for course credit (22 inthe I-step condition, and 19 in the 2-step condition). Of these, 6were Asian, I was Indian, and 34 were White.
Stimuli. Stimuli were the same as those used in Experiments Iand 2 with the exception that only half of each continuum was pre-sented. Therefore, 2-step continua that previously consisted of nineface pairs, now consisted of only five pairs. The I-step continua in-cluded six discrimination pairs. These pairs included stimuli rang-ing from 0% to 60% (or 40% to 100%) of the original continua.This made it possible to use the discrimination pair that formerlystraddled the midpoint at the continuum end. These pairs were thesame as those used in Experiments I and 2, but, in the 2-step con-dition, they spanned 33% of the 7-step continua instead of20% ofthe II-step continua and, in the l-step continua, they covered 17%instead of 10% of the original continua. Two different versions ofeach continuum were created, each including one half or the otherof the original continua. Each participant therefore viewed continuathat represented one half or the other of the original continua, andno participant viewed two halves of the same original continuum.
Procedure. The procedure for the discrimination and catego-rization tasks was identical to that in Experiments I and 2 with theexception that fewer trials were included in each block because ofthe reduced number ofstimulus pairs. The instruction screens wherenames were paired with the faces were modified to include the for-mer midpoint stimuli where appropriate. For each participant, boththe discrimination and the categorization tasks used the same ab-breviated continua.
CATEGORICAL PERCEPTION 395
A Classification Task
All Continua Individual Continuum Types100~-------.--..",o..,100..---------:::=,.....,
c c'0 '0e,80 l1. 80
"C:lC "C:lell C
ellII)
II)III 60 III 60"C:l
".! .!!'iii =II) 40 III 40III III'0 III'0C ---.- B-BCII 20
i: 20 -...- B-WQ CII... 2 ---;:r-- wowCIIa. CIIa.
0 0~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~0 ,... l') 0 ,... l') 0 0 ,... l') 0 ,... l') 0.... l') II> q) lID 0 l') II> q) lID 0
Stimulus Stimulus
Discrimination Task
All Continua Individual Continuum Types
100 ,-------"T---r------, 100 ,--------------,
---.- B-B90 90 -...- B-W- - --0-- WoWU U
Gl f...... 80 ... 800 00 0- -c: e 70Gl 70 GlU U... ...Gl GlCl. Cl.
60 60
50 50~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~r- f'l ~ ..... f'l ~ e- f'l ~ e- f'l ~.... f'l III ~ QC ~ .... f'l III ~ QC ~Q r-:. .:, Q r-:. .... Q r-:. .:, Q r-:. ........ f'l III ~ .:, .... f'l III ~ .:,
QC QC
Pair Pair
Figure 5A. Classification and discrimination data from the I-step condition ofExperiment 3. Thex-axis of the classification graph represents the continua, including both endpoint stimuli (0% and100%) and intermediates (10%-90%), and the x-axis of the discrimination graph represents stim-ulus pairs spanning the continua. The 0% and 100% endpoint designations are arbitrary for theW-W and B-B continua. For the B-W continua, the 0% endpoint is the Black face of the pair, andthe 100% endpoint is the White face. The classification graph contains the percentage of trials onwhich each stimulus was classified in the same category as the 100% (endpoint) stimulus. The dis-crimination graph contains discrimination accuracy for stimulus pairs along each continuum. Thevertical lines in the discrimination graphs for all continua represent the pairs crossing the categoryboundary.
After completing the categorization and discrimination tasks, eachparticipant was presented with an additional rating task. In the rat-ing task, the participants saw each of the endpoint and midpointfaces used in the experiment (including faces from all four stimu-lus sets) and rated their distinctiveness on a 7-point scale, with an-chor points ranging from very undistinctive (rating of I) to very dis-tinctive (rating of 7). The participants were instructed to rate the
faces in terms of how well they would stand out in a crowd includ-ing male faces from many different races.
ResultsAs in Experiments I and 2, the pair or pairs straddling
the physical middle of all continua (the 33%-67% in the
396 LEVIN AND BEALE
B Classification TaskAll Continua Individual Continuum Types
100,---------"iii:::"--, 100.-------,loooo!e~
c C"0 '0Q, 80 Q, 80'tI 'tIc: c:II II
co60 II 60., III
'tI 'tI
.! ~Ui 40 Ui 40II II., .,u U B-BC
20 C 20 B-WCD IIU U WoW..
~II4- 4-
0 0f!. f!. f!. f!. f!. f!. f!. f!. f!. f!. f!. f!. f!. f!.0 ,.. .., 0 ,.. .., 0 0 ,.. .., 0 ,.. .., 0.., U'l '" co 0
.., U'l '" co 0Stimulus Stimulus
Discrimination Task
100 .,-------------,
~ B-B----.-- B-W--{)-- WOW
Individual Continuum Types
90
80
70
60
mm
All Continua
100
90-()Gl.... 8000-c 70Gl()..Gla.
60
50 50~ ~ ~ ~ ~ ~ ~ ~ ~ ~"'l Q e- "'l Q "'l Q r-- "'l Q"'l ~ ~ QC
Q "'l III '" QC Q¢ = ... = r.:. .;., = ...e- "'l r.:. r.:.... "'l III ... "'l III'" '"Pair Pair
Figure 5B. Classification and discrimination data from the 2-step condition of Experiment 3. Thex-axis ofthe classification graph represents the continua, including both endpoint stimuli (0% and100%) and intermediates (10%-90%), and the x-axis of the discrimination graph represents stim-ulus pairs spanning the continua. The 0% and 100% endpoint designations are arbitrary for theW-W and B-B continua. For the B-W continua, the 0% endpoint is the Black face ofthe pair, andthe 100% endpoint is the White face. The classification graph contains the percentage oftrials onwhich each stimulus was classified in the same category as the 100% (endpoint) stimulus. The dis-crimination graph contains discrimination accuracy for stimulus pairs along each continuum. Thevertical lines in the discrimination graphs for all continua represent the pairs crossing the categoryboundary.
2-step condition, and the 33%-50% and 50%-67% pairsin the l-step condition) also straddled the category bounds,as determined in the categorization tasks.
As can be seen in Figures 5A and 5B, small but signif-icant CP effects were observed for both the l-step and 2-step conditions [l-step, F(1,21) = 4.763, MSe = 0.009,p = .0203; 2-step, F(l,18) = 7.361, MSe = 0.002, p =
.0071]. Overall tests including only the two within-racecontinuum types (B-B + W-W) were significant in the2-step condition [F(l,18) = 14.93, MSe = 0.002, p =.0006] and marginally significant in the l-step condition[F(l,21) = 2.576, MSe = 0.013,p = .0617]. In the 1-step condition, a significant categorical effect was ob-served for the B-B continua [F(1,21) = 4.382, MSe =
0.013,p = .0243], whereas the effect was nonsignificantfor the W- W continua (F < 1), and marginally significantfor the B-W continua [F(l,21) = 2.391, MS e = 0.029,p = .0685]. In the 2-step condition, the CP contrasts forthe B-W and W-W continua were nonsignificant, al-though, for the W-W continua, they came close [B-W,F< I; W-w, F(l,18) = 2.365, MSe = 0.005,p = .0775].The CP contrast for the B-B continua was, however, sig-nificant [F(l,18) = 4.664, MSe = 0.013,p = .0223].
Planned contrasts were also run to test for increaseddiscrimination accuracy for stimuli that had been in themidregions of the original continua (as predicted by anartifact hypothesis). For the I-step condition, this in-volved a contrast between accuracies for the 0%-17% +17%-33% pairs and the 67%-83% + 83%-100% pairs(recall that the former pairs are switch pairs and the lat-ter are the continuum end). For the 2-step condition, thecontrast was computed between the 0%-33% + 17%-50% pairs and the 50%-83% + 67%-100% pairs. One-tailed tests were used in all cases. These contrasts werenonsignificant for the overall accuracies in both condi-tions[l-step,F< 1;2-step,F(I,18) = 1.979,MSe = 0.007,p = .0883]. Within the l-step condition, the contrast didnot approach significance for the B-Band W-W continua(Fs < I). The test was significant for the B-W continua[F(l,21) = 5.701, MS e = 0.032,p = .0132]. Within the2-step condition, the contrast was significant for the W- Wcontinua [F(l,18) = 3.375, MSe = 0.014, P = .0414]and nonsignificant for the B-Wand B-B conditions [B-W,F(I,18) = 2.520, MS e = 0.016,p = .0649; B-B, F< 1].Thus, in the overall tests and in three of the four criticalindividual tests (e.g., the B-B and W-W tests for I-stepand 2-step conditions; recall that existing race categoriescould cause switch pairs to be accurately discriminatedin the B-W continua), there was no evidence of an arti-fact contribution to discrimination accuracy.
In addition to testing for CP using contrasts betweencross-boundary pairs and within-category pairs, we testedfor significant linear and nonlinear trends using polyno-mial regression. This allowed a test of nonlinearities, aspredicted by CP, independent of possible linear trends,as predicted by stimulus artifacts. In all cases, the indepen-dent variable was pair number (I for the switch pair, and5 or 6 for the original endpoint pair), and the dependentvariable was percent accuracy for that pair across all con-tinuum types. For the I-step condition, a third-order re-gression was the most parsimonious model (r 2 = .963;fourth order, r2 = .966; second order, r2 = .605) and wasassociated with a near-significant overall Ftest [F(3,5) =17.5,p = .0545]. X, X2,and X3 terms were all associatedwith significant beta coefficients (Bs = .221, - .062, and.005; ps = .0281, .0363, and .0475, respectively). Thus,as predicted by a CP finding, there were significant non-linearities in the data. In addition, the significant linearcomponent was associated with a positive beta coefficient(increased accuracy for pairs near the original endpoint),goes in a direction opposite to that predicted by an artifacthypothesis (increased accuracy for the switch pair). None
CATEGORICAL PERCEPTION 397
of the regressions (linear, second-order, and third order)run on the data from the 2-step condition approached over-all signi ficance, so they were not analyzed further.'
Another interesting pattern was evident in the 2-stepcondition. Note that, in this condition, the physical dif-ference between stimuli within each pair was identical tothat in Experiment 1. As compared with Experiment 1,however, overall discrimination accuracy significantly in-creased for stimulus pairs on the B-W continua [Exper-iment 1, 84.3%; Experiment 3,92.5%; F(l,33) = 8.265,MSe = 0.007,p = .007] and also increased for the W-Wcontinua, although the effect was only marginally sig-nificant [Experiment I, 82.3%; Experiment 3, 88.6%;F( 1,26) = 3.825, MS e = 0.009, p = .059]. Accuracy re-mained unchanged for the B-B continua (Experiment I,81.9%; Experiment 3, 79.7%; F< I).
DiscussionExperiment 3 provides evidence that the CP effects
observed in Experiments I and 2 were not driven bystimulus artifacts. In both the l-step condition and the2-step condition, the overall CP effect was again signif-icant. Switch pairs that formerly straddled the categoryboundary and therefore were most accurately discrimi-nated in Experiments I and 2, now included the new end-point and, in many cases, were least accurately discrim-inated. On the half-continua, the CP effect was rescaledrelative to that in Experiments 1 and 2, which demon-strates its dependence on the particular location of eachdiscrimination pair relative to the continuum endpoints.If the effects were artifacts, based on low-level percep-tual differences between the stimuli along the originalcontinua, then discrimination accuracy should reflecteach pair's place on these continua and be unaffected byits location on the new continua.
Although new CP effects were observed, a small asym-metry was observed for one of the individual continuumtypes in which switch pairs were more accurately dis-criminated than the corresponding end pairs for one ofthe four tests on critical continuum types (the W-W 2-step condition). Switch pairs were also somewhat moreaccurately discriminated in the I-step and 2-step B-Wcontinua, but, as mentioned above, this can be expectedbecause the switch pairs straddle the between-race cate-gory boundary even if they are defined as endpoints inthe context of the experiment. Thus, it appears that thelocation of stimulus pairs on the original continua mayhave a small effect on discrimination accuracy. Althoughthis effect might reflect the contribution ofa stimulus ar-tifact, there is another plausible explanation. This alter-native begins with the assumption that intermediates aregenerally less distinctive than endpoints because they areessentially averages of the endpoint face.
To confirm this, we asked II new participants to rate themidpoint and endpoint faces for distinctiveness and foundthat midpoint faces were less distinctive (rating of 3.46on a scale of 1-7) than endpoint faces [4.29; F( I, I0) =41.431, MS e = O.277,p = .0001], a finding confirmed
398 LEVIN AND BEALE
recently by Busey (1998). Given that distinctive faces aremore accurately recognized than typical ones (Going &Read, 1974; Light, Kayla-Stuart, & Hollander, 1979;Valentine & Bruce, 1986), it is likely that they are associ-ated with stronger or more robust representations. If thisis true, then discrimination accuracy might have beenreduced in continuum end regions because of a "percep-tual magnet" effect (Iverson & Kuhl, 1995; Kuhl, 1991)whereby strong representations of a stimulus cause nearvariants to be assimilated into the stimulus class and there-fore are inaccurately discriminated. It is possible that suchan effect contributed to the discrimination accuracies ob-served here. The key is that different locations on simplecontinua between faces vary in terms of distinctiveness,and this variation could contribute to categorical effects.Thus, the tendency for switch pairs to have a slight ad-vantage in discrimination could be an artifact, but it couldalso reflect a more interesting variant of Kuhl's percep-tual magnet effect. However, it is important to note thaton no half-continua were the former midpoints discrim-inated most accurately. This clearly suggests that locationon the newly defined continua was a strong determinantof discrimination accuracy.
In addition to the data from Experiment 3, other find-ings mitigate against an artifact explanation. First, Levin(1999) used the same stimulus set in a design where stim-uli from three different continua were intermixed in a sin-gle block oftrials. In this case, no CP effect was observed.In addition, the negative finding in unfamiliar faces re-ported by Beale and Keil (1995) was based on differentstimuli but the same distortion process. Therefore, bothof these findings converge to suggest the implausibilityofa simple artifact explanation. Ifthe CP effects we haveobserved here were based on a stimulus artifact inducedby the distortion process, then the effect should not dis-appear when mixing of different continua makes thelearning task more difficult. Finally,the CP effect size var-ied systematically with the difficulty of representing thecontinuum endpoints. In Experiment I, the effect wasweaker for other-race faces (same-race, F = 28.782; other-race, F = 3.609). Inversion in Experiment 2 also weakenedthe effect in same-race faces (F = 4.125), while com-pletely eliminating it in other-race faces (F < 1). Again,these results would not be expected if the effect were asimple perceptual artifact. Instead, the CP finding appearslinked to the relative difficulty that participants have inrepresenting the continuum endpoints.
One surprising finding from the 2-step condition isthat overalldiscrimination accuracy on the B-W and W-Wcontinua improved for the same-face pairs when they werepart of half-continua, as compared with full continua.Thus, when the continua included at least one face fromthe familiar group, discrimination accuracy seems to beheavily influenced by the percentage of the continuumspanned by each pair as opposed to the actual physicaldifference between pairs. The lack of between-experimentimprovement on B-B continua did not occur, becausethe participants spent little time looking at B-B end-
points, as compared with B-W and W-W endpoints. Themean inspection time for B-B instruction screens wasactually slightly greater than that for B-W and W-Wcontinua (B-B, 41.9 sec, SE = 5.2 sec; B-W: 38.9 sec,SE = 3.9 sec; W-W, 36.1 sec, SE = 5.6 sec).2 It thereforeappears likely that improved discrimination accuracy onB-Wand W- W continua was the result of rescaling theperceptual difference between face pairs based on the psy-chological difference between the endpoints. This pro-cess is similar to the range effects observed when mak-ing discriminations between different tone intensitiesand pitches (Gravetter & Lockhead, 1973; Lockhead &Hinson, 1986). For example, Lockhead and Hinson (1986)found that intensity discrimination between two tones ismore accurate ifthe task includes a third tone that is sim-ilar to the first two than if the third is very different. Thus,the perceptual difference between the two tones is effec-tively greater if they are part of a larger set that covers asmall intensity range.
GENERAL DISCUSSION
Twomain findings emerge from these data. First, CP ispossible on continua between novel faces. Second, CP isobserved even when the faces are inverted and when thefaces belong to an unfamiliar group. It therefore appearsthat CP can be observed in a wide variety of situations.Given this, it appears that a good understanding ofCP musttake into account the on-line learning processes that allowus to set up visual representations of individual objects.
Learning New Perceptual CategoriesIn the research reported here, there were two primary
opportunities for the participants to learn the faces. First,for each of three blocks of trials, they viewed an instruc-tion screen containing the two endpoint faces, each as-sociated with an arbitrarily chosen name. As reported inExperiment 3, it appears that the participants spent anaverage of approximately 40 sec viewing this screen.Given that these averages were based on only the secondand third blocks oftrials, it is probably accurate to assumethat this time was spent primarily on the faces and not onreading the instructions that were simply repeats ofthoseon the first screen. Most of this time was probably spentclosely comparing the faces with each other and attempt-ing to associate a name with each. Moreover, verbal in-structions given to the participants emphasized the im-portance ofgetting the face-name association correct andfurther emphasized that this task would take some thoughtand was not as easy as it appeared. These instructions weregiven to avoid the tendency to quickly look at the stim-uli and then continue the task, which could result in for-getting the correct face-name association, a necessaryprecondition for making responses.
The second major source oflearning occurs during thediscrimination trials themselves. During this task, partic-ipants are gradually exposed to the continuum and mustassess the degree to which the intermediates look more
or less like the endpoints. Again, the task context couldplaya central role in determining the extent ofthe CP ef-fect. In addition, it is important to ask about the timecourse of this learning: Will the CP effect be observed inthe first blocks ofdiscrimination trials, or does it only ap-pear late in the experiment? An exploratory analysis ofthe data from Experiment I shows that the CP effect ispresent even if only the first half of the discriminationtrials are analyzed [F(l,15) = 10.219, MSe = O.OII,p =.0060]. Therefore, it appears that CP reflects a quick parseof new perceptual categories into equivalence classes.
If the CP we have observed arises during the experi-ment itself, then what specific processes cause it? Onemeans of approaching the question is to ask whether theeffect is caused by reduced sensitivity to variation withinperceptual categories ("equalization"), thereby causingless accurate discrimination in continuum end regions, orby increased sensitivity to variation on the borders ofper-ceptual categories, thereby causing more accurate discrim-ination in continuum midregions ("sensitization"; Gold-stone, 1994; Pisoni, 1991). In the present case, there is nodirect evidence on the matter, but a comparison betweenExperiments 1 and 2 does give some hints. Inverting thefaces seems to have reduced the CP effect primarily by re-ducing discrimination accuracy in the continuum midre-gions. Discrimination accuracy for the 0%-20% and80%-100% stimulus pairs was relatively unaffected byinversion (upright, 72.8%; inverted, 70.5%), whereas ac-curacy for the other more central pairs was more clearlyreduced [upright, 85.7%; inverted, 78.0%; orientation Xregion (end/mid) interaction, F(l,28) = 4.127, MSe =0.003, p = .0518]. It therefore appears that inverting thefaces reduced the CP effect primarily by reducing dis-crimination accuracy in the midregion, implying that theeffect is based on sensitization more than equalization.
This conclusion is similar to that arrived at by Gold-stone (1994), who tested the effects ofcategorization train-ing on the degree to which discrimination accuracy couldbe affected for only part ofa given dimension. Those ex-periments included a training task in which classificationswere based on dimensions, such as the size and luminanceofgray squares, such that two values fell into each of twocategories. Goldstone found that training increased dis-crimination accuracy selectively for values that straddledthe trained category boundaries and found no evidencethat it reduced accuracy for values within a category.
In contrast to Goldstone (1994), Livingston, Andrews,and Harnad (1998) recently observed learned CP effectscharacterized by equalization within categories. Similarto in Goldstone's study, subjects were exposed to a learn-ing task in which they classified stimuli on the contin-uum into two categories and then completed a discrimi-nation task. Compared with subjects who had not donethe learning task, subjects who learned the categoriesshowed a CP effect characterized by equalization withincategories. As Livingston et al. point out, a number offactors may have produced the contrast between their dataand Goldstone's. The most important is that they used a
CATEGORICAL PERCEPTION 399
similarity-judging task in which CP was indicated byjudg-ments of increased similarity within categories and de-creased similarity between categories. Goldstone used asame/different task in which subjects who learned thecategories were more accurate for between-category pairs.Livingston et al. discuss two task-relevant factors thatmay have produced this difference. First, their similarity-rating task allowed for continuous responses (a ratingranging from 1 to 9), whereas Goldstone's task involvedbinary responses. Furthermore, Goldstone's task involvedpresentation ofstimulus pairs that were the same, whereasLivingston et al.'s did not. Our data give tentative supportto the former hypothesis-we also used a task with a bi-nary response to reveal sensitization but did not presentpairs of identical stimuli. Another contrast between Liv-ingston et al.s, and Goldstone's and our data is that thelatter tasks were performance-based and the former weresubjective ratings. Perhaps the similarity ratings dependedmore on decision-stage processing, whereas the perfor-mance tasks depended on more immediate perceptualprocesses. In any case, the reasons behind this empiricalcontrast remain open to debate and could certainly provethe basis for fruitful exploration.
In addition to suggesting a putative sensitization ef-fect, our data go a step further and show that CP can beobserved for novel continua, even in the absence of pre-training. In particular, the sensitization we observed wasnot the result ofa training task where each member of thesensitized discrimination pairs were actually classifiedinto different categories. Instead, we observed sensitiza-tion based more directly on representations ofthe contin-uum endpoints. No experience classifying one memberof a pair in category A and the other in category B wasnecessary for the cross-boundary pair to be discriminatedmore accurately. This implies that the learned effects wehave observed are based less on sensitization producedby direct training and more on the way that representationsof the endpoints themselves cause increased sensitivity tovariation in intermediates. The comparison between thedata reported here and the data reported by Beale and Keil(1995) reinforces this conclusion. As mentioned above,Beale and Keil mixed different continua in a single blockofdiscrimination trials and found no CP on continua be-tween unfamiliar faces. Thus, increasing the number offaces that must be represented decreases the CP effect.
In suggesting the importance of endpoint representa-tions, these data run counter to adaptation level theory asa sole explanation for the CP effects we have observedhere. According to this explanation, CP is caused by alow-level adaptation process in which the mean value ofthe stimulus dimension becomes an adapted "neutralpoint" (Helson, 1948). During the discrimination task,stimuli are judged relative to the point. Pairs that straddlethe adapted region are therefore easy to discriminate, andpairs that fall within one side of the adaptation level aremore difficult to discriminate (see Wilson, 1987, for re-view). Thus, the range of physical stimulus values com-prising the continuum cause CP, not representations of
400 LEVIN AND BEALE
the endpoint per se. The close correspondence of CP ef-fect size with codability of the endpoints suggests thatsomething more than repetition-based adaptation causedthe CP effects we have observed here.
However, it might be possible to modify an adaptationlevel explanation ofCP to encompass our results. For ex-ample, it would be helpful to add some mechanism bywhich neutral points become less robust in difficult-to-code faces or when a number of different continua arepresented in a single block of trials. In the latter case,adaptation might overlap between continua to producediffuse neutral points that affect discrimination on morethan one continuum. This could reduce the CP,especiallyon continua between unfamiliar faces that vary betweenpreviously unknown values.
The near-pair and far-pair comparison in Experiment 1also suggests the incompleteness of a range-based ex-planation for these CP effects. It is possible that mid-region stimuli were more accurately discriminated be-cause they are effectively judged within a smaller rangethan end-region stimuli because no stimulus is highly dif-ferent from them. According to research discussed above,a given physical difference between stimuli might be-come psychologically smaller if they are coded as part ofa large range. These effects can occur both globally andlocally (Repp & Libermann, 1987). In the latter case, thetrials immediately preceding the stimuli to be discrimi-nated have a particularly strong effect on the effectiverange. Although range effects do appear to contribute tooverall discrimination accuracy (see discussion of Ex-periment 3), the near-X/far-X analysis in Experiment 1reveals a stronger CP effect when the stimulus pair wason the same side of the continuum as the target stimulus,a circumstance in which the effective stimulus range is re-duced. A range theory appears to predict a smaller CP ef-fect here if one assumes that the X stimulus is effective indefining a range locally. Therefore, the relationship be-tween the representation invoked on each trial and theperceived stimuli mediated the effect-a circumstanceoutside the scope ofmost range-based theories, which re-fer more directly to the effects of repeatedly viewing thestimuli themselves.
Summary and ConclusionsThese findings suggest that the scope of the CP phe-
nomenon should be expanded from the idea that it repre-sents innate perceptual constraints, or constraints broughton only by long-term familiarity with the particular stim-uli used in the experimental setting. Instead, it might behelpful to view CP as a dynamic on-line process that parsesperceptual input in response to both long-term needs andtask-specific constraints (Goldstone, 1994). This kind ofon-line classification is similar to that proposed by Barsa-lou (1993) for general-knowledge concepts and thereforehighlights the similarities in form between research onabstract concepts and perceptual classes discussed byMedin and Barsalou (1987). As a practical matter, we
should expect to observe CP in many situations whereparticipants have the opportunity to sufficiently differ-entiate continuum endpoints either during training trialsor when simply shown them. As Hamad (1987) pointsout, this process might exemplify the global similaritybetween psychophysical identification and naming inthat both processes are based on the need to quickly as-similate the extent ofequivalence classes. Although CP isfacilitated by expertise with the category to which thestimuli belong, this skill may not be strictly necessary. Assuch, the presence ofCP per se tells us little about whethercontinua or specific stimulus classes are innately speci-fied or are even highly learned. Instead, CP can serve as auseful index ofshort-term perceptual learning, which canhelp us understand the first steps in molding perceptualvariation into cognitively useful equivalence classes.
REFERENCES
BARSALOU. L. W. (1993). Flexibility, structure, and linguistic vagary inconcepts: Manifestations of a compositional system of perceptual sym-bols. In A. F. Collins, S. E. Gathercole, M. A. Conway, & P. E. Morris(Eds.), Theories ofmemory (pp. 29-10 I). Hove, England: Erlbaum.
BEALE, J. M., & KEIL, E C. (1995). Categorical effects in the percep-tionoffaces. Cognition, 57, 217-239.
BURNS, E. M., & WARD, W. D. (1978). Categorical perception-phenomenon or epiphenomenon: Evidence from experiments in theperception of melodic musical intervals. Journal of the AcousticalSociety ofAmerica, 63, 456-468.
BUSEY, T. A. (1998). Physical and psychological representations offaces: Evidence from morphing. Psychological Science, 9, 476-483.
CALDER, A. J., YOUNG, A. w., PERRETT, D. I., ET('OFF, N. L., & Row-LAND, D. (1996). Categorical perception ofmorphed facial expression.Visual Cognition, 3, 81-117.
CAMPBELL, R., PASCALlS, 0., COLEMAN, M., WALLACE, S. B., & BEN-SON, P. J. (1997). Are faces of different species perceived categori-cally by human observers? Proceedings ofthe Royal Society ofLon-don: Series B, 264,1429-1434.
DE GELDER, B., TEUNISSE, J., & BENSON, P.J. (1997). Categorical per-ception offacial expressions: Categories and their internal structure.Cognition & Emotion, 11, 1-23.
DIAMOND, R., & CAREY, S. (1986). Why faces are and are not special:An effect of expertise. Journal of Experimental Psychology: Gen-eral, 115, 107-117.
ETCOFF, N. L., & MAGEE, J. J. (1992). Categorical perception of facialexpression. Cognition, 44, 227-240.
FALLSHORE, M. E, & SCHOOLER, J. W. (1995). The verbal vulnerabilityofperceptual expertise. Journal ofExperimental Psychology: Learn-ing, Memory, & Cognition, 21,1608-1623.
FARAH, M. J., TANAKA, J. W., & DRAIN, H. M. (1995). What causes theface inversion effect? Journal ofExperimental Psychology: HumanPerception & Performance, 21, 628-634.
GOING, M., & READ. J. D. (1974). Effects of uniqueness, sex of subject,and sex of photograph on facial recognition. Perceptual & MotorSkills,39,109-110.
GOLDSTONE, R. (1994). Influences of categorization on perceptual dis-crimination. Journal of Experimental Psychology: General, 123,178-200.
GRAHM, C. H. ( 1965). Discriminations that depend on wavelength. InC. H. Grahrn (Ed.), Vision and visual perception (pp. 452-503). NewYork: Wiley.
GRAVETTER, E, & LOCKHEED. G. R. (1973). Criterial range as a frame ofreference for stimulus judgement. Psychological Review, 80, 203-216.
HARNAD, S. (1987). Category induction and representation. In S. Har-nad (Ed.), Categorical perception: The groundwork ofcognition(pp. 535-565). New York: Cambridge University Press.
HELSON, H. (1948). Adaptation level as a basis for a quantitative theoryofframes of reference. Psychological Review, 55, 297-313.
IVERSON, P., & KUHL, P. K. (1995). Mapping the perceptual magnet ef-fect for speech using signal detection theory and multidimensionalscaling. Journal of the Acoustical Society ofAmerica, 97, 553-562.
KUHL, P. K. (1981). Discrimination of speech by nonhuman animals:Basic auditory sensitivities conducive to the perception of speech-sound categories. Journal ofthe Acoustical Society ofAmerica, 70,340-349.
KUHL, P. K. (1987). The special-mechanisms debate in speech research:Categorization tests on infants and animals. In S. K. Harnad (Ed.),Categorical perception: The groundwork ofcognition (pp. 355-386).New York: Cambridge University Press.
KUHL, P. K. (1991). Human adults and human infants show a "percep-tual magnet effect" for the prototypes of speech categories, monkeysdo not. Perception & Psychophysics, 50, 93-107.
KUHL, P. K., & PADDEN, D. M. (1982). Enhanced discriminability at thephonetic boundaries for the voicing feature in macaques. Perception& Psychophysics, 32, 542-550.
LEVIN, D. T. (1996). Classifying faces by race: The structure of facecategories. Journal ofExperimental Psychology: Learning. Memory.& Cognition, 22, 1364-1382.
LEVIN, D. T. (1999). Race as a visualfeature: Using visual search andperceptual discrimination tasks to understandface categories. Man-uscript submitted for publication.
LIBERMAN, A. M., COOPER, F. S., SHANKWEILER, D. P., & STUDDERT-KENNEDY, M. (1967). Perception of the speech code. PsychologicalReview, 74,431-461.
LIBERMAN, A. M., HARRIS, K. S., HOFFMAN, H. S., & GRIFFITH, B. C.(1957). The discrimination of speech sounds within and across pho-neme boundaries. Journal ofExperimental Psychology, 54, 358-368.
LIBERMAN, A. M., HARRIS, K. S., KINNEY, J., & LANE, H. (1961). Thediscrimination of relative onset-time of the components of certainspeech and nonspeech patterns. Journal ofExperimental Psychology,61,379-388.
LIGHT, L. L., KAYLA-STUART, F.. & HOLLANDER, S. (1979). Recognitionmemory for typical and unusual faces. Journal ofExperimental Psy-chology: Human Learning & Memory,S, 212-228.
LIVINGSTON, K. R., ANDREWS, J. K., & HARNAD, S. (1998). Categoricalperception effects induced by category learning. Journal ofExperi-mental Psychology: Learning. Memory & Cognition, 24, 732-753.
LOCKHEAD, G. R., & HINSON, J. (1986). Range and sequence effects injudgment. Perception & Psychophysics, 40, 53-61.
MEDIN, D. L., & BARSALOU, L. W. (1987). Categorization processes andcategorical perception. In S. Harnad (Ed.), Categorical perception:The groundwork ofcognition (pp. 455-490). New York: CambridgeUniversity Press.
MEDIN, D. L., GOLDSTONE, R. L., & GENTNER, D. (1993). Respects forsimilarity. Psychological Review, 100,254-278.
PASTORE, R. E. (1987). Categorical perception: Some psychophysicalmodels. In S. Harnad (Ed.), Categorical perception: The groundworkofcognition (pp. 535-565). New York: Cambridge University Press.
PISONI, D. B. (1975). Auditory short-term memory and vowel perception.Memory & Cognition, 3, 7-18.
PISONI, D. B. (1991). Modes of processing speech and nonspeech sig-nals. In I. G. Mattingly & M. Stewart-Kennedy (Eds.), Modularityand the motor theory ofspeech perception (pp. 225-238). Hillsdale,NJ: Erlbaum.
REPP, B. H., & LIBERMANN, A. M. (1987). Phonetic category bound-aries are flexible. In S. Harnad (Ed.), Categorical perception: Thegroundwork ofcognition (pp. 535-565). New York: Cambridge Uni-versity Press.
RHODES, G., BRAKE, S., & ATKINSON, A. P. (1993). What's lost in in-verted faces? Cognition, 47, 25-57.
RHODES, G., TAN, S., BRAKE, S., & TAYLOR, K. (1989). Expertise andconfigural encoding in face recognition. British Journal ofPsychol-ogy, 80, 273-283.
CATEGORICAL PERCEPTION 401
SAWUSCH, J. R., & GAGNON, D. A. (1995). Auditory coding, cues, andcoherence in phonetic perception. Journal ofExperimental Psychol-ogy: Human Perception & Performance, 21, 635-652.
SHAPIRO, P. N., & PENROD, S. (1986). Meta-analysis of facial identifi-cation studies. Psychological Bulletin, 100, 139-156.
SMITH, J. D., KEMLER NELSON, D. G., & ApPLETON, T. (1994). Whatchild is this? What interval was that? Familiar tunes and music per-ception in novice listeners. Cognition, 52, 23-54.
STEVENAGE, S. V. (1998). Which twin are you? A demonstration of in-duced categorical perception of identical twin faces. British JournalofPsychology, 89, 39-57.
STEVENS, K. N. (198 I). Constraints imposed by the auditory system onproperties used to classify speech sounds: Data from phonology,acoustics and psycho-acoustics. In T. F. Meyers, 1. Laver, & 1. An-derson (Eds.), The cognitive representation ofspeech (pp. 61-74).Amsterdam: North-Holland.
TANAKA, J. W., & FARAH, M. J. (1993). Parts and wholes in face recog-nition. Quarterly Journal ofExperimental Psychology, 46A, 225-245.
VALENTINE, T., & BRUCE, V. (1986). Recognizing familiar faces: Therole of distinctiveness and familiarity. Canadian Journal ofPsychol-ogy, 40, 300-305.
WILSON, M. (1987). Brain mechanisms in categorical perception. InS. Harnad (Ed.), Categorical perception: The groundwork ofcogni-tion (pp. 387-417). New York: Cambridge University Press.
NOTES
I. To allay the concern that the statistical test for CP used for Exper-iments 1-3 was too liberal, we reanalyzed the data using another tradi-tional test for CP (e.g., Liberman, Harris, Hoffman, & Griffith, 1957).In this kind of test, the categorization results are used to predict dis-crimination accuracy for each pair. Following Calder, Young, Perrett,Etcoff, and Rowland (1996), we started with a baseline estimate forwithin-category accuracy by averaging accuracy for the first and lastpairs in the continuum. Added to this was the absolute difference be-tween the A and B stimuli in probability of classification with a con-tinuum end multiplied by .25 to scale estimates in rough accord with theoverall accuracy data. Regressions tested the relationship between pre-dicted and observed discrimination accuracies in each condition. ForExperiment I, increased accuracy in continuum ends was considered anoncategorical input to discrimination accuracy and was accounted forby adding a "side" variable to a multiple regression. Accounting for con-tinuum side, the relationship between predicted and actual accuracy wassignificant (p < .05) overall and for all individual continuum types (par-tial rs = .74, .78, .77, and .82, for B-B, B-W, W-W, and overall aver-age continua, respectively). In Experiment 2, a simple regression (therewas no side effect in this experiment) predicted discrimination scoressignificantly for B-W, W-W, and overall average continua (rs = .80,.68, and .79, respectively), but not for B-B continua (r = .43). Datafrom the two conditions in Experiment 3 were combined to allow in-creased power (which was lacking because there were only 5 or 6 pairsin each condition), and a multiple regression including variables for pre-dicted accuracy and condition was run. Controlling for condition, pre-dicted accuracy was significant overall (partial r = .68) but not for in-dividual continuum types (partial rs = .50, .50, and .30, for B-B, B-W,and W-W continuum types, respectively).
2. These means include only the second two of three instructionscreen inspection times for each participant. The first instruction screenfor each participant was present on each computer before the participantsat down to begin the task; therefore, the "RT" from that stimulus wasnot a measure of the time spent actually looking at it. Thus, means acrossconditions reflected contributions from different sets of participants,and, therefore, no test of significance is reported.
(Manuscript received October 13, 1997;revision accepted for publication December 13,1998.)