Embed Size (px)
Citation preview

Aquatic Botany, 43 (1992) 365-377 365 Elsevier Science Publishers B.V., Amsterdam
Carex litter decomposition and nutrient release in mires with different water chemistry
J.T.A. Verhoeven and H.H.M. Arts Department of Plant Ecology and Evolutionary Biology, University of Utrecht, P.O. Box 800.84,
3508 TB Utrecht, Netherlands
(Accepted 23 April ! 992)
ABSTRACT
Verhoeven, J.T.A. and Arts, H.H.M, 1992° Carex litter decomposition and nutrient release in rnire~ with ,:l_~ft'erent water chemistry. Aquat. Bot., 43: 365-377.
Losses of dry weight, nitrogen, phosphorus and potassium from shoot litter ofCarex diandra Schrank and Carex acutiformis Ehrh. were studied in a 2-year litter hag experiment in two fens, each domi- nated by one of these species. Weight losses over time showed negative exponen:ial curves dr~.ng Year 1, with k values of 0.0018 and 0.0009 day -~ for Carex diandra and Carex acutiformis, respec- tively. Total weight loss during Year I equalled that during Year 2, and amounted to 45% and 27% of the initial weight for Carex diandra and Carex acutiformis, respectively.
Total nitrogen in the litter bags remained approximately constan~ throughout the 2-year-period for Carex acutiformis, whereas it remained constant during Year I and then dropped to 23% of the initial value during Year 2 for Carex diandra. Total phosphorus showed a similar pattern: no cha, ges for Carex acutiformis, and a substantial decrease to 15% of the initial value during Year 2 for Carex diandra.
A study of cotton strip tensile strength loss showed that conditions for cellulose decay are much more suitaole in the groundwater fed (base-rich) fen dominated by Carex diandra than in the rain- water fed (base-poor) fen with Carex acutiformis, which is in accordance with the results of the litter bag study.
The slower nutrient release from Carex acutiformis litter does not lead to a lower mineralization rate of soil organic matter in the fen dominated by this species. The high release rates of inorganic nitrogen and phosphorus in this fen are probably because of the presence of Sphagnum-derived or- ganic matter.
INTRODUCTION
The main question addressed in this paper is whether differences in the decomposition rate of the dominant vascular plants explain differences in nutrient availability among fens with different hydrological regimes. Decom-
Correspondence to: J.T.A. Verhoeven, Department o f Plant Ecology and Evolutionary Biology, University of Utrecht, P.O. Box 800.84, 3508 TB Utrecht, Netherlands.
© 1992 Elsevier Science Publishers B.V. All rights reserved 0304-3770/92/$05.00

366 #.T.A. VERHOEVEN AND H.H.M. ARTS
position of plant litter plays a major role in determining the availability of nutrients to plants in terrestrial and wetland ecosystems (Swift et at., 1979). Nitrogen and P tend to remain tied up in the litter and the associated micro- bial community for variable amounts of time before eventually being re- leased. The residence time of the nutrients in the litter depends on the rate of decomposition. This rate is determined by the chemical composition of the litter (e.g. N, P and lignin content; Parnas, 1975; Berg and McClaugherty, 1987; Berendse et at., 1989) and by environmental conditions (temperature, rcdox status, pH, nutrient richness; Godshalk and Wetzel, 1978; Swift et al., 1979). In a study of the nutrient dynamics of small fens, the rates of release of
inorganic N and P in the soil were found to be higher in fens in groundwater recharge areas than in fens in groundwater discharge areas (Verhoeven and Arts, 1987; Verhoeven et al., 1990). These fen types also differ in vegetation and water chemistry. The vegetation is dominated by short Carex species in base-rich groundwater discharge fens and by tall Carex species in base-poor recharge fens in which a well developed Sphagnum layer is also present (Ver- meer and Verhocven, 1987; Beltman and Verhoeven, 1988).
In this paper, studies of decomposition and nutrient release from Carex shoot litter, decomposition of cotton strips and mineralization of soil organic matter in fens of both types are presented. These studies investigate the de- gree to which inorganic N and P release in the soil are determined by decom- position of vascular plant litter, and the degree to which the rates of decom- position and mineralization processes are affected by water chemistry, arc presented.
MATERIALS AND METHODS
Study area
The study was carried out in two small fens in the Vechtplassen area, the Netherlands. Both fens have floating mats with emergent vegetation and water tables a few centimetres below the mat surface. They are located in different polders (i.e. areas with a regulated water table) and differ in vegetation, water chemistry and nutrient dynamics (Verhoeven et al., 1983, 1988a; Verhoeven and Arts, 1987).
The fen in the Westbroek polder is characterized by a species-rich vegeta- tion of small sedges dominated by Carex diandra Schrank; the moss layer is of small quantitative importance and contains species such as Calliergon cor- difolium (Hedw.) Kindb. and Bryum pseudotriquetrum (Hedw.) G.M.S. This fen is located in a groundwater discharge area and has a 'base-rich' water chemistry characterized by relatively high pH values and calcium, magne- sium and bicarbonate concentrations (Table 1; Koerselman et al., 1990a).
The fen in the Molenpoldei has a tall but sparse phanerogam vegetation

DECOMPOSITION OF CAREXLITTER 367
TABLE 1
Some characteristics (mean + s.e.) of the interstitial soil water (n--~ 14) at the surface of the fens studied (after Koerselman, 1990)
Parameter Fen dominated by
Carex diandra Carex acutiformis
pH 6.2+ 0.1 5.3+ 0.1 EC(ms/m) 1980 +109 !100 +100 Na(ppm) 20.2+ 5.5 11.1+ 2.1 K(ppm) 1.7+ 0.3 6.5+ 0.9 Ca (ppm) 32.7+ 2.0 6.4+ 0.7 Mg (ppm) 2.9+ 0.2 1.0+ 0.2 HCO3 (ppm) 92.3 + 9.8 25.7+ 6.0 SO,, (ppm) 11.9+ 2.4 15.3+ 1.7 CI (ppm) 13.1+ 1.6 24.4+ 3.4
dominated by Carex acutiformis Ehrh. and Phragmites australis ( Cav. ) Trin. ex Steud. and a well-developed moss layer dominated by Sphagnum species (Sphagnum squarrosum Crome in Hoppe, Sphagnum fimbriatum Wils. ). It is located in a groundwater recharge area and has a 'base-poor' water chem- istry (Table 1 ).
The two fens differ in nutrient dynamics. Levels ofphosphate are higher in the water of the Molenpolder fen (Koerselman et al., 1990b) and the miner- alization of organic nitrogen and phosphorus are also faster in this fen (Ver- hoeven et al., 1990).
Methods
Standing dead shoot litter of Carex diandra and Carex acutiformis was col- lected at the beginning of March 1985 in quaking fens in Westbroek and Mo- lenpolder, respectively. Data collected in connection with a concurrent de- mographic study of both species (Verhoeven et al., 1988b) have shown that the litter had originated from shoots formed in 1984 between the mowing of the plants in July and the death of the shoots at the end of November.
After air-drying of the litter (8 h at 30 °C), 1 g subsamples were enclosed in 10 cm × 10 cm nylon litter bags with 0.2 mm mesh. One hundred and twenty litter bags containing Carex diandra litter, and 120 bags containing Carex acutiformis litter were placed at the surface of the floating mat (in contact with the interstitial soil water) at the sites where the litter had been collected (Westbroek fen and Molenpolder fen, respectively).
After 0, 1, 2, 4, 8, 12, 16, 24, 32, 42, 52 and 110 weeks, ten litter bags were retrieved from each fen and taken to the laboratory. Samples were dried (48 h at 70°C) and weighed. Carbon and N contents of ground samples were

368 J.T.A. VERHOEVEN AND H.H.M. ARTS
determined gaschromatographically in a Carlo Erba Elemental Analyzer. Phosphorus contents were determined colorimetrically after acid digestion with sulfuric and salicylic acid.
In situ soil incubation experiments for measuring N and P mineralization and cotton strip experiments for measuring decomposition were carried out at five stations at each of the two sites. The incubation period lasted for 6 weeks and started on 10 September 1987. At each station, soil material was collected from depths of 10 and 25 cm. Living roots and coarse plant parts were removed and the material was divided into two polyethylene bottles ( 100 ml). One of the bottles was taken to the laboratory, the other bottle was placed back into the soil at the depth where the material had been sampled and left there for 6 weeks. The methodology has been described in more detail by Verhoeven et al. (1990).
Substrate samples (fresh as well as incubated) were extracted the day after they had been collected. A wet equivalent of I g dry soil was extracted with 100 ml ofmedium ( 1 h at 100 r.p.m.). Distilled water was used for extraction of nitrate and phosphate, and 0.2 M KCI for extraction of ammonium. The extracts were filtered and stored, and refrigerated overnight. Analyses were made colorimetrically with a continuous-flow analyzer (see Verboeven et al., 1988b).
Mineralization rates were calculated as extractable N or P from incubated soil minus that from fresh soil. For N, the amounts of nitrate and ammonium found in the water and the KCI extracts, respectively, were added as elemen- tary N. Nitrate concentrations in the extracts were very low (below the detec- tion level in most cases).
At each station, close to the bottles, a cotton strip measuring 30 cm × 12 cm was inserted vertically in the soil to a depth of 25 cm in a sawn slit 15 cm wide. The strips were left in place for 6 weeks. A second strip was inserted and removed immediately to serve as a control. Test ah~i control cotton strips were washed gently in deionized water and air-dried before dispatch to the laboratory. Strips were cut into horizontal substrips 3 cm wide and reduced by fraying to 2 cm, to give test substrips corresponding to 0-2 cm, 3-5 cm, 6- 8 cm, etc. Tensile strength was measured using a tensometer (Monsanto Type W, University of Exeter, Department of Geography, Exeter, UK). The gen- eral procedure is described in full and illustrated in Harrison et al. (1988). Tensile-strength loss was calculated from the difference between test and con- trol strips and duration of exposure.
Data were analyzed statistically with the General Linear Models procedure of the Statistical Analysis Systems Institute (1985). Statistical significance of differences between means was tested with the Tukey-Cramer test (Sokal and Rohlf, 1981 ).

DECOMPOSITION OF CAREX LITTER 369
RESULTS
Dry weight loss of Carex litter occurred throughout the 2-year incubation period in both fens studied (Fig, 1 ). Carex diandra litter in the Westbroek fen had lost 45% of its initial dry weight after one year and 88% after 2 years, whereas these figures were 27% and 52% for Carex acutiforrnis in the Molen- polder fen. The amounts ol dry weight lost in Year 1 and Year 2 were almost equal for both species. Statistical testing of the actual weight loss by both spe- cies (calculated as weight at time t minus weight at t-I divided by t minus (t- 1) ) revealed a significantly faster decomposition rate for Carex diandra (P<0.001).
The total amounts of N in the bags (Fig. 2) were initially higher for Carex diandra because the litter of this species was higher in N. The data for both species showed some irregular changes during the first 12 weeks. After 16 weeks, the amount of N in the bags remained constant until the end of the study period in Carex acutiforrais, whereas it showed a non-significant in- crease until 32 weeks and a highly significant decrease (P<0.01) thereafter in Carex diandra. After 1 year, the amount of N in Carex diandra litter was still similar to the initial amount but after 2 years only 23% remained.
The total amounts of P in the bags (Fig. 3) were also initially higher for Carex diandra. There were some initial irregular fluctuations during the first 12 weeks; the high values for Carex diandra at 4 weeks and for Carex acuti-
formis at 2 weeks are significantly different (P< 0.05) fror~ some ofthe other
1.0
0.8
-- c 0.6 m
~ Carex d landra
.~ = C. acutiformis 0.4
G) 5=
o 0.2.
0.0' o 2'0 ,'0 e'0 8'0 ,oo ,2o
T ime ( w e e k s )
Fig. 1. Dry weight of litter in litter bags in the course of the experiment. Vertical bars indicate standard errors.

3 7 0 J.T.A. VERHOEVEN AND H.H.M. ARTS
20
£
m
'= 10 0'-,-- Carex diandra
• C. aculiformis
_e
z 5
o | ~ i w i 0 20 4 60 80 100 120
T i m e (weeks )
Fig. 2. The amount of nitrogen in the litter bags in the course of the experiment. Vertical bars indicate standard errors.
12
1.0'
0.8
~ 0.6 ~ Camx d~andra
8 C. acuhlmmis
,.E 0.4 o.
0.2,
0,0 0 20 4 6 80 100 120
T i m e (weeks )
Fig. 3. The amount of phosphorus in the litter bags in the course of the experiment. Vertical bars indicate standard errors.

DECOMPOSITION OF CAREX LITTER 371
TABLE 2
C: N and C: P ratios of litter of Carex diandra ( Cd ) and C. acutiform~ (Ca) dunng the experiment
Time C: N ratio C: P ratio (weeks)
Cd Ca Cd Ca
0 34 67 4~5 898 1 33 68 399 933 2 38 49 461 616 4 28 51 327 740 8 29 49 438 880
12 25 54 357 1126 16 22 44 293 652 24 19 43 249 797 32 17 36 257 708 42 17 34 290 771 52 !8 38 253 569
110 18 27 314 556
Carex acutlformls fen
~" -~o
~ -13 e -16 .o
-19
-22
-25 ~ I
0.9 ,'.9 ;.o tensile strenoth 19SS (%/day)
Cerex dlandra fen
-, P'///////-/////////7////~
., ~/// / / / / / / / /1~/// / / / / / / ,~//1~
., ~///// / / / / / / / / / / / / / / / / / / / / / / / / /~ .,o ~//////////////////////////1~
,: .,3 ~/////// / / / / / / / / / / / / / / / / /~ e .,s ~// / / / / / / / / / / / / / / / / / /~
"o
.,9 ~//// / / / / / / / / / / / / /1~ -22
.2s ~ / / / / / / / / / / / / / / 1 , - . ~
o.o ~:o 219 tensile strength loss (%/gay)
Fig. 4. Tensile strength loss of cotton strips inserted 25 cm into the soil of the fens and left there for 6 weeks (September-November 1987). Horizontal lines indicate standard errors.
372 J.T.A. VEPHOEVEN AND H.H.M. ARTS
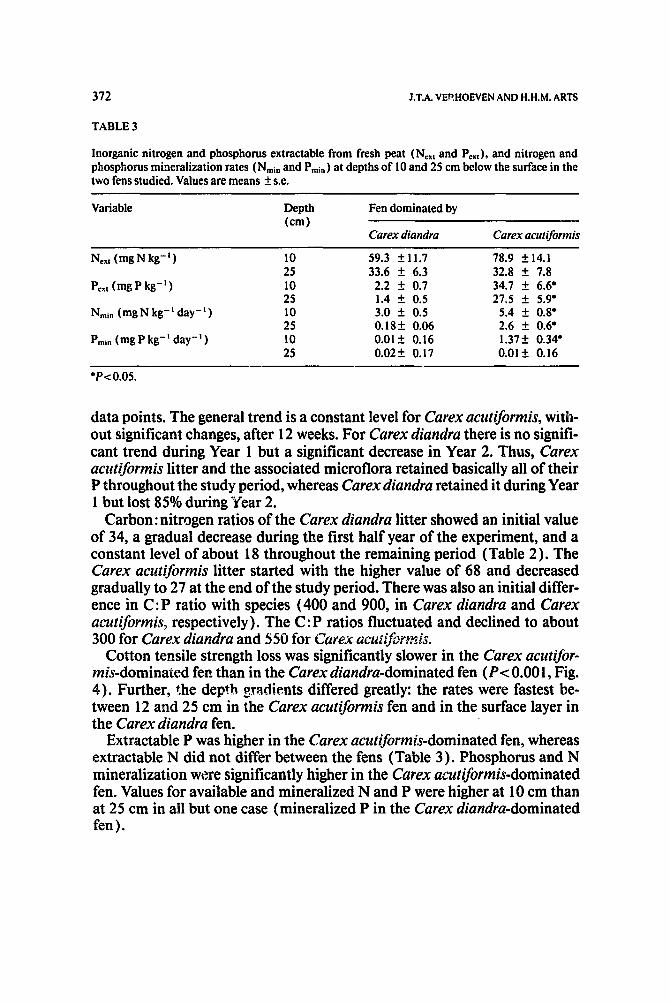
TABLE 3
Inorganic nitrogen and phosphorus extractable from fresh peat (Next and Pext), and nitrogen and phosphorus mineralization rates (Nmln and Pmin) at depths of 10 and 25 cm below the surface in the two fens studied. Values are means + s.e.
Variable Depth Fen dominated by (cm)
Carex diandra Carex acutiformis
Next (rag N kg- I) 10 59.3 + 11.7 78.9 + 14.1 25 33.6 _+ 6.3 32.8 + 7.8
Pe~t(mgPkg - I ) 10 2.2 + 0.7 34.7 + 6.6* 25 1.4 + 0.5 27.5 + 5.9*
Nmin(mgNkg-lday - I ) 10 3.0 + 0.5 5.4 + 0.8* 25 0.18+ 0.06 2.6 + 0.6*
Pmin (mgP kg -I day - I ) 10 0.01 + 0.16 1.37+ 0.34* 25 0.02+ 0.17 0.01+ 0.16
*P<0.05.
data points. The general trend is a constant level for Carex acutiformis, with- out significant changes, after 12 weeks. For Carex diandra there is no signifi- cant trend during Year 1 but a significant decrease in Year 2. Thus, Carex acutiformis litter and the associated microflora retained basically all of their P throughout the study period, whereas Carex diandra retained it during Year 1 but lost 85% during Year 2.
Carbon: nitrogen ratios of the Carex diandra litter showed an initial value of 34, a gradual decrease during the first half year of the experiment, and a constant level of about 18 throughout the remaining period (Table 2). The Carex acutiformis litter started with the higher value of 68 and decreased gradually to 27 at the end of the study period. There was also an initial differ- ence in C:P ratio with species (400 and 900, in Carex diandra and Carex acutiformis, respectively). The C:P ratios fluctuated and declined to about 300 for Carex diandra and 550 for Carex acut~r~,~is.
Cotton tensile strength loss was significantly slower in the Carex acutifor- mis-dominated fen than in the Carex diandra-dominated fen (P< 0.001, Fig. 4). Further, ~:he dep*h gradients differed greatly: the rates were fastest be- tween 12 and 25 cm in the Carex acutiformis fen and in the surface layer in the Carex diandra fen.
Extractable P was higher in the Carex acutiformis-dominated fen, whereas extractable N did not differ between the fens (Table 3). Phosphorus and N mineralization were significantly higher in the Carex acutiformis-dominated fen. Values for available and mineralized N and P were higher at 10 cm than at 25 cm in all but one case (mineralized P in the Carex diandra-dominated fen).

DECOMPOSITION OF CAREXLITTER 373
DISCUSSION
The litter used for tile experiments had been collected about 3 months after the death of the shoots of both Carex species. A concurrent demographic study ofthe two species (Verhoeven et al., 1988a) revealed that the litter originated from the senescence (October) and death (November) of shoots formed after the harvesting of the stand (July 1984 in both sites). The material would certainly have been subject to leaching prior to collection but microbial activ- ity would have been very low in view of the low temperatures prevailing from December 1984 to February 1985; the monthly average air temperatures at De Bilt weather station, close to the fens, were 4.1 °C, -4 .9°C and -0 .4°C for December, January and February, respectively.
Leaching of dissolved organic matter from fresh litter is known to be a fast process, occurring primarily during the first 2 months after the death of the plants (Swift et al., 1979). Conditions for leaching were quite similar in the two fens: senescence and death of shoots were simultaneous for the two Carex species, and the fens are located 4 km apart in an open landscape of pastures and have carefully regulated water levels close to the soil surface. Hence, in our experiment~ the two litter types were at a similar 'after-leaching' stage when harvested and the observed losses in organic matter, N and P from the bags were therefore primarily associated with microbial breakdown.
The decomposition of Carex diandra in the groundwater discharge fen was obviously much faster than that of Carex acutiformis in the recharge fen. For each species, total weight losses were very similar in Years I and 2. This con- stant annual weight loss has also been found in some other 2-year decompo- sition studies, e.g. by Mason and Bryant ( 1975 ) for Phragmites australis, An- dren and Paustian (1987) for Hordeum distichon L. (barley) and Seastedt ( 1988 ) for tallgrass prairie grasses.
The time courses of the dry weight losses show a steadily declining rate of decomposition during Year 1. The weight loss curves for this period can be described with a 'single source" first-order negative-exponential model (see Andren and Paustian, 1987). The relative decay constants (k) provide a gen- eral index for comparison of decay rates (Jenny et al., 1949; Webster and Benfield, 1986). The k values for Year 1 are 0.0018 for Carex diandra and 0.0009 for Carex acutiformis. Both k values are considerably lower than the mean value for Cyperaceae given by Webster and Benfield (1986): 0.0027. The value for Carex acutiformis is even in the range of the rather refractory pine litter.
Both environmental factors and litter quality may strongly influence litter decay rates (Godshalk and Wetzel, 1978; Swift et al., 1979). In our experi- ment, the most important environmental factor differentiating the two sites studied was water chemistry. Groundwater discharge in the Carex diandra- domim~ted fen led to a pH of about 6.5 and high concentrations of calcium

374 J.T.A. VERI-IOEVEN AND H.H.M. ARTS
and bicarbonate at the fen surface. The Carex acutiformis dominated fen had a more rainwater-like chemistry at the surface and also a thick Sphagnum carpet; pH was about 5.0 and calcium and bicarbonate levels were much lower.
The cotton strip assay, a relative measure of the suitability of environmen- tal conditions for decay (Latter and Howson, 1977; Hill et al., 1985; Harrison et al., 1988) showed that the acid, base-poor conditions in the Carex acutifor- mis.dominated fen were unfavourable for decomposition. Cellulose decay in the surface layer proceeded at a 75% reduced rate compared with the Carex diandra-dominated fen. This is a stronger reduction in decay rate even than that found in the litter bag study. Distinct differences in decay rate were also found by Ohlson (1987), who studied the decomposition of Carex rostrata Stokes leaves at two sites with different water chemistry within one fen. The differences in water chemistry described by him ('spring fen' vs. 'intermedi- ate fen') correspond well with those between our two sites, and he also found much lower decay rates in the 'intermediate-fen' site.
The differences in the chemical composition of the litters of the two species studied, as characterized by the C:N and C: P ratios, may also have contrib- uted to the difference in decay rate. Litter with high C:N and C: P ratios de- cays relatively slowly (Parnas, 1975; Swift et al., 1979). The strong differ- ences found in the cotton strip assay suggest, however, that, in our study, differences in water chemistry rather than litter quality caused the difference in decay rate found between Carex diandra and Carex acutiformis.
The results of the measurements of decay and nutrient release from Carex litter are surprising in that they showed a difference between the two fens opposite to that in inorganic N and P release rates measured in the soil mi- neralization experiments. Two factors may explain this different behaviour of fresh shoot litter and 'old' soil organic matter. The first factor is the actual depth at which most of the nutrient release from the litter occurs. The litter of Carex diandra loses most of its nutrients by Year 2 after senescence and death. If we assume that its annual weight loss remains constant after Year 2, the Care~" acutiformis litter would lose most of its nutrients in Years 3 and 4 after death; considering that C: N and C: P ratios were still relatively high at the end of Year 2, it is most probable that significant losses would not occur until Year 4. This would mean that N and P losses from Carex diandra shoot litter occur at or just beiow the soil surface, whereas they occur in somewhat deeper strata buried by litter additionally accumulated for 3 years for Carex acutiformis. Our soil incubation at a depth cf 10 cm may just miss a shallower nutrient release p~ak in the case of Carex diandra, whereas it may just be at the location of this pe~k in the case of Carex acutiformis. This explanation is, however, not supported by the fact that the difference in soil N mineraliza- tion also exists at a depth cf25 cm.
A second explanation concerns the fact that Carex root litter is an addi- tional important litter type in both fens and that Sphagnum litter is of at least

DECOMPOSITION OF CAREX LFI'TER 375
equal quantitative importance as Carex litter in the Carex acutiformis domi- nated fen. Although root litter in peatlands is thought to decompose very slowly (Clymo, 1983 ), hardly any quantitative studies exist because of inher- ent methodological problems. Sphagnum litter is known to be particularly refractory (Clymo, 1965). In a l-year decomposition experiment Brock and Bregman (1989) showed that litter of Sphagnum recurvam P.B. var. mucro- nature lost only 17% of its initial dry weight but 45% of its N and 60e of its P. Apparently, the coupling between decomposition and net mineralization in Sphagnum litter is very different from that in most other plant litter (Swift et al., 1979). Studies in acid fens and bogs have revealed that N mineraliza- tion takes place at high C:N ratios in these systems, probably as a result of low microbial activity which does break down the easily degradable cyto- plasm but leaves Sphagnum cell wall material almost unchanged. Because of this lack of immobilizing microbial activity, inorganic N is released (Waugh- man, 1980; Damman, 1988; Verhoeven et al., 1990).
Hence, it can be concluded that water chemistry strongly influences the rate of decomposition and nutrient release from vascular plant litter in fens. The much slower rate of these processes in base-poor, add fens in comparison with base-rich fens does, however, not lead to slower nutrient release rates from soil organic matter. The presence of Sphagnum-derived organic matter in the soils of base-poor fens may even result in higher nutrient release rates than in base-rich fens.
ACKNOWLEDGEMENTS
We are much indebted to Dr. E. Maltby (Department of Geography, Uni- versity of Exeter) who provided and analyzed the cotton strips. This paper was completed while JTAV was at the Smithsonian Environmental Research Center in Edgewater, MD, USA, supported by a Smithsonian Senior Postdoc- toral Fellowship and a grant from the Netherlands Organization for Scientific Research (NWO). Dr. D.F. Whigham, Dr. W. Koerselman, Dr. B. Beltman, Dr. R. Aerts and Prof. Dr. P.J.M. van der Aart critically read the manuscript.
REFERENCES
Andren, O. and Paustian, K., 1987. Barley straw decomposition in the field: a comparisoa of models. Ecology, 68:1190--1200.
Beltman, B. and Verhoeven, J.T.A., 1988. Distribution of fen plant communities in relation to hydrochemical characteristics in the Vechtplassen area. In: J.T.A. Verhoeven, G.W. Heft and M.J.A. Werger (Editors), Structure of Vegetation in Relation to Carbon and Nutrient Ecow omy. SPB Academic, The Hague, pp. 121-136.
Berendse, F., Bobbink, R. and Rouwenhorst, G., 1989. A comparative study on nutrient cycling in wet heathland ecosystems. II. Litter decomposition and nutrient mineralization. Oecolo- gia, 78: 338-348.
Berg, B. and McClaugherty, C., 1987. Nitrogen release from litter in relation to the disappear- ance oflignin. Biogeochemistry, 4:219-224.
Brock, T.C.M. and Bregman, R., 1989. Periodicity in growth, productivity, nutrient content and

376 J.T.A. VERHOEVEN AND H.H.M. ARTS
decomposition of Sphagnum recurvum var. mucronatum in a fen woodland. Oecologia, 80: 44-52.
Clymo, R.S., 1965. Experiments on the break-down of Sphagnum in bogs. J. Ecol., 53: 747-757. Clymo, R.S., 1983. Peat. In: A.J.P. Gore (Editor), Mires: Swamp, Bog, Fen and Moor. Ecosys-
tems of the World, Vol. 4A. Elsevier, Amsterdam, pp. 159-224. Damman, A.W.H., 1988. Regulation of nitrogen removal and retention in Sphagnum bogs and
other peatlands. Oikos, 51: 291-305. Godshalk, G.L. and Wetzel, R.G., 1978. Decomposition of aquatic angiosperms. III. Zostera
marina and a conceptual model of decomposition. Aquat. Bot., 5: 329-354. Harrison, A.F., Latter, P.M. and Walton, D.W.H. (Editors), 1988. Cotton strip assay: an index
of decomposition in soils. Institute of Terrestrial Ecology Symp. No. 24. Grange-over-Sands. Hill, M.O., Latter, P.M. and Bancroft, G., 1985. A standard curve for inter-site comparison of
cellulose degradation using the cotton strip method. Can. J. Soil Sci., 65:609-.619. Jenny, H., Gessel, S. and Bingham, F.T., 1949. Comparative study of decomposition of organic
matter in temperate and tropical regions. Soil Sci., 68: 419-432. Koerselman, W., Bakker, S.A. and Biota, M., 1990a. Nitrogen, phosphorus and potassium budgets
for two small fens surrounded by heavily fertilized pastures. J. Ecol., 7g: 498-424. Koerselman, W., Claessens, D., Ten Den, P. and van Winden, E., 1990b. Dynamic hydrochem-
ical and vegetation gradients in fens. Wetl. Ecol. Manage., 1: 73-84. Latter, P.M. and Howson, G., 1977. The use of cotton strips to indicate cellulose decomposition
in the field. Pedobiologia, 17: 145-155. Mason, C.F. and Bryant, R.J., 1975. Production, nutrient content and decomposition of Phrag-
mites communis Trin. and Typha angustifolia L.. J. Ecol., 63:71-95. Ohlson, M., 1987. Spatial variation in decomposition rate ofCarex rostrata leaves on a Swedish
mire. J. Ecol., 75:1191-1197. Parnas, H., 1975. Model for decomposition of organic material by microorganisms. Soil Biol.
Biochem., 7: 161-169. Seastedt, T.R., 1988. Mass, nitrogen and phosphorus dynamics in foliage and root detritus of
tallgrass prairie. Ecology, 69: 59-65. Sokal, R.R. and Rohh, F.J., 198L Biometry. Freeman, San Francisco, CA, 859 pp. Statistical Analysis Systems Institute, 1985. SAS/STAT Guide for Personal Computers. SAS
Institute, Cary, NC, 378 pp. Swift, M.J., Heal, O.W. and Anderson, J.M., 1979. Decomposition in Terrestrial Ecosystems.
Studies in Ecology 5. BlackweU, Oxford, 372 pp. Verhoeven, J.T.A. and Arts, H.H.M., 1987. Nutrient dynamics in small mesotrophic fens sur-
rounded by cultivated land. II. N and P accumulation in plant biomass in relation to the release of inorganic N and P in the peat soil. Oecologia (Berlin), 72:557-561.
Verhoeven, J.T.A., van Beek, S., Dekker, M. and Storm, W., 1983. Nutrient dynamics in small mesotrophic fens surrounded by cultivmed land. I. Productivity and nutrient uptake by the vegetation in relation to the flow ofeutrophicated ground water. Oecologia (Berlin), 60: 25- 33.
Verhoeven, J.T.A., Schmitz, M.B. and Pons, T.L., 1988a. Comparative demographic study of Carex rostrata Stokes, C. diandra Schrank and C. acutiformis Ehrh. in fens of different nu- trient slatus. Aquat. Bot., 30: 95-108.
Verhoeven, J.T.A., Koerselman, W. and Beltman, B., 1988b. The vegetation of fens in relation to their hydrology and nutrient dynamics: A case study. In: J.J. Symoens (Editor), The Veg- etation of Inland Waters. Handbook of Vegetation Science, Vol. 15. Kluwer, Dordrecht, pp. 249-282.
Vcrhoeven, J.T.A., Maltby, E. and Schmitz, M.B., 1990. Nitrogen and phosphorus mineraliza- tion in fens and bogs. J. Ecol., 78:713-726.
Vermeer, J.G. and Verhoeven, J.T.A., 1987. Species composition and biomass production of

DECOMPOSITION OF CAREX LITTER 377
mesotrophic fens in relation to the imtrient status of the organic soil. Acta Oecol./Oecol. Plant., 8(22): 321-330.
Waughman, G.J., 1980. Chemical aspects of the ecology of some South German peatlands. J. Ecol., 68: 1025-1046.
Webster, J.R. and Benfieid, E.F., 1986. Vascular plant breakdown in freshwater ecosystems. Ann. Rev. Ecol. Syst., 17: 567-594.





























