Embed Size (px)
Citation preview

409
the cardiac plexus. The cardiac plexus, which is divided into the superficial (ventral) and deep (dorsal) cardiac plexus, gives rise to sympathetic nerve branches that run along the coronary arteries on the epicardium and penetrate the myocardium with the vascu-lature. Thus, the nerves are located primarily around blood vessels and between myocytes oriented along their long axis. Further, a gradient in sympathetic innervation is thought to exist from base to apex, as well as from epicardium to endocardium.7,8
Data collected over the past four decades indicate that exten-sive neural processing occurs in the cardiac plexus, otherwise known as the intrinsic cardiac nervous system. This plexus contains afferent neurons, interconnecting interneurons, and sympathetic and parasympathetic efferent postganglionic neurons.9 Further-more, it is composed of seven ganglionic subplexi, containing more than 800 epicardial ganglia. Each subplexus or group of subplexi innervates different chambers of the heart. One sub-plexus innervates the right ventricle, three innervate the left ven-tricle, and the rest innervate the atria. The highest density of epicardial ganglia, approximately 50%, exists near the hilum of the heart, especially on the dorsal and dorsolateral surfaces of the left atrium. The number of neurons in these ganglia decreases with age, from approximately 94,000 neurons in neonates and children to 43,000 intrinsic neurons in the plexus of adults.10 A complex feedback regulatory system allows the cardiac nervous system to modulate sympathetic and parasympathetic input to the heart.
Cardiac nerves can be demonstrated by labeling nerve-specific markers such as S100 protein, neurofilament, and synaptophysin using immunohistochemistry techniques,5,6 and myelinated fibers can be identified by myelin markers, such as myelin basic protein. Sympathetic nerves can be identified by immunolabeling tyrosine hydroxylase. Tyrosine hydroxylase and myelin basic protein stain-ing have confirmed the presence of myelinated sympathetic nerves on all surfaces of the epicardium and endocardium.11
Sympathetic Modulation of Myocardial Excitability
The major neurotransmitter mediating sympathetic response in the heart is norepinephrine. Epinephrine release from intracar-diac neural endings is negligible.12 Along the length of terminal axons are a series of localized swellings known as varicosities. Most of the norepinephrine storage vesicles in a terminal axon are concentrated in these varicosities, which act as specialized sites of norepinephrine storage and release.13 The overall effect of norepinephrine release through multiple signaling pathways is shortening of the ventricular action potential duration (APD) and the refractory period.14 Most norepinephrine undergoes reuptake into nerve terminals by the presynaptic norepinephrine trans-porter. A small fraction diffuses into the vascular space, where it can be measured in coronary sinus blood. Norepinephrine spill-over (both interstitial and in the coronary sinus) can be used to infer that sympathetic outflow to the heart can also be assessed in humans.15
CHAPTER OUTLINE
Anatomy of Cardiac Sympathetic Innervation 409
Sympathetic Modulation of Myocardial Excitability 409
Neural Remodeling in the Presence of Myocardial Pathology 410
Proarrhythmic Effects of Sympathetic Stimulation 412
Effect of Neuraxial Modulation in Reducing Risk of Ventricular Arrhythmias 413
Surgical Sympathetic Denervation in Humans 414
Conclusion 415
The autonomic nervous system plays a significant role in the genesis and maintenance of ventricular arrhythmias, and power-fully modulates the underlying substrate in a dynamic manner.1-3 Blockade of the sympathetic nervous system, whether through medications or neuraxial modulation, has been associated with a reduction in the risk of sudden cardiac death and the burden of ventricular arrhythmias. In this chapter, the anatomy of the cardiac sympathetic nervous system and pathological changes associated with neural remodeling in the setting of scar and car-diomyopathy will be reviewed. Subsequently, the role of sympa-thetic activation in promoting arrhythmias, and the effects of neuraxial modulation in reducing the burden of these arrhyth-mias will be discussed. Finally, cardiac denervation in humans, including feasibility, surgical techniques, and complications, will be considered.
Anatomy of Cardiac Sympathetic Innervation
The preganglionic sympathetic neurons that innervate the heart reside in the intermediate zone of the thoracic spinal cord.4 These preganglionic sympathetic nerves pass through the white rami communicantes, enter the sympathetic trunk, and terminate in the cervicothoracic ganglion as well as in the T2-T4 ganglia. Of note, preganglionic neurons may synapse on neurons within the ganglia at the same thoracic level or may travel within the sympa-thetic chain and synapse on neurons of ganglia at other spinal levels. The preganglionic neurotransmitter within the ganglia is acetylcholine. The right and left cervicothoracic ganglia, which are often a product of the fusion of the C8 and T1 ganglia, are called the left stellate (LSG) and right stellate (RSG) ganglia. LSG and RSG, along with the ganglia of T2-T4, give rise to postgan-glionic axons that target organs including the heart, the esopha-gus, the trachea, and the head and neck (Figure 41-1). Postganglionic nerve fibers from these ganglia then join the branches of the vagus nerve to form the left and right cardiac (cardiopulmonary) nerves, destined for the heart.5,6 These nerves subsequently form a network of neurons on the epicardium, called
41 Sympathetic Innervation, Denervation, and Cardiac Arrhythmias
Marmar Vaseghi, Olujimi A. Ajijola, Aman Mahajan, and Kalyanam Shivkumar

410 NEURAL CONTROL OF CARDIAC ELECTRICAL ACTIVITY
innervation; certain viable sites shorten their repolarization, while others show no response. All denervated areas show dener-vation super-sensitivity, defined as an exaggerated response to norepinephrine infusion.21 The cellular mechanisms for this response do not involve differences in the β-adrenergic receptor or the α-subunit of stimulatory G-protein density.22,23
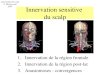
In patients with recurrent ventricular arrhythmias undergoing ventricular tachycardia (VT) ablation procedures, activation recovery interval (ARI) measurements show a reduced response to indirect sympathetic stimulation via nitroprusside in dense scar and in the viable peri-infarct myocardium, suggesting denerva-tion.24 These sites demonstrate an exaggerated response to iso-proterenol infusion, suggesting the presence of denervation super-sensitivity in humans (Figure 41-3). Further, the response to sympathetic stimulation is extremely heterogeneous, with a greater than 2-fold increase in dispersion in ARI with nitroprus-side infusion.24
These studies were important because they demonstrated that transmural MI and heart failure not only can alter the myocardial substrate for arrhythmias, they can also disrupt innervation to histologically viable myocardium, leading to denervation super-sensitivity and a non-uniform electrophysiological response to sympathetic stimulation. This contributes to the genesis of ven-tricular arrhythmias in both acute and chronic MI.
Cardiac Neural Remodeling and Nerve Sprouting
After studies showed acute denervation, evidence of nerve sprout-ing and heterogeneous hyperinnervation was observed in chronic
Figure 41-1. The sympathetic chain and in particular the left stellate ganglion (LSG) and the ganglia of thoracic spinal 1-4 (T1-T4) are located in a paravertebral position behind the parietal pleura.
(Reproduced with permission from Abrahams PH, Spratt JD, Boon J: McMinn’s Clinical Atlas of Human Anatomy, 6th ed, St Louis, 2007, Mosby.)
LSG
T2 Ganglion
T3 Ganglion
T4 GanglionAorta
Leftventricle
Figure 41-2. Multiple organs mediate sympathetic outflow to the normal heart including the brain, the spinal cord, the sympathetic chain (left stellate ganglion [LSG], right stellate ganglion [RSG], and ganglia of T1-T4), the adrenal medulla by secreting catecholamines, and the renal nerves. E, Epinephrine; LSG, left stellate ganglion; NE, norepinephrine; RSG, right stellate ganglion; SAN, sinoatrial node; T1-T4, thoracic spinal level 1 through 4.
Brain andhigher centers
LSG
RSG
SAN
NENE
Adrenalmedulla
2/3 E
1/3 NE
Kidney
Thoracic
T1-T4
Spinalcord
Sympathetic stimulation, predominantly mediated by post-synaptic myocardial β-adrenergic receptors, has important effects on chronotropy, dromotropy, lusitropy, and inotropy. Discharge of the sinoatrial (SA) node and atrioventricular (AV) nodal con-duction are augmented, increasing chronotropy. In the atria and ventricles, contractility and relaxation are enhanced. Both β1 and β2 subtypes are present at a ratio of approximately 5 : 1 in the healthy human heart.16-18 Alpha adrenoreceptors are mainly present in the vascular wall, but are also found in ventricular myocardium, where they account for approximately 15% of cardiac adrenergic receptors.18
Cardiac sympathetic activation is complex and is mediated through multiple organs at multiple levels. The brain (higher centers), brain stem, spinal cord, sympathetic ganglia, adrenal medulla, and renal nerves can increase the net sympathetic output to the heart, dynamically modulating cardiac function (Figure 41-2).
Neural Remodeling in the Presence of Myocardial Pathology
Myocardial infarction (MI) and heart failure, in addition to leading to scar formation, cause remodeling of cardiac nerves. Initial denervation, followed by nerve-sprouting and trans-differentiation of sympathetic nerves, leads to heterogeneity in repolarization and can modulate reentry.
Cardiac Neural Remodeling and Denervation
Myocardial infarction causes death of sympathetic fibers within the scar and loss of efferent sympathetic innervation at nonin-farcted apical sites.19 Norepinephrine depletion in the scar is accompanied by increased production in noninfarcted basal areas.20 In response to LSG stimulation, viable myocardial sites apical to an infarct show evidence of heterogeneity in
SYMPATHETIC INNERVATION, DENERVATION, AND CARDIAC ARRHYTHMIAS 411
41
“swarm-like” pattern in the periphery of necrotic tissues and in perivascular regions have been observed, and these changes were more prominent in patients with a history of ventricular arrhythmias.29-31 These border zones of infarcts have also been shown to be frequent sites of origin of inducible ventricular tachycardia (VT)/ventricular fibrillation (VF).32,33 Furthermore, nerve sprouting can occur in a noninfarct setting such as stem
MI and heart failure models. Peripheral nerve injury resulting in Wallerian degeneration, which leads to nerve growth factor (NGF), triggered regeneration via nerve sprouting.25,26 Along with denervation, MIBG studies have shown localized re-innervation in injured myocardium in both ischemic and nonischemic cardiomyopathy patients.27,28 In human hearts with cardiomyopathy, local increases in sympathetic nerves in a
Figure 41-3. A recording multi-electrode catheter on fluoroscopy (left upper panels) and electroanatomical map (right upper panel) in this patient with ischemic cardiomy-opathy (ICM) and a large anteroapical scar are used to record unipolar electrograms from scar, border zone, and viable myocardium. On the electroanatomical map, the purple areas represent viable tissue (normal voltage) and gray represents dense scar. All other colors represent border zones (0.5 mV < voltage < 1.5 mV). The delta activa-tion recovery interval (ARI; change in ARI from baseline) within the cardiomyopathic and normal hearts is shown in the lower panel. Note that in response to isoproterenol, the delta ARI is greatest in the CM-normal site (viable myocardium) and scar regions of the cardiomyopathic heart. The border zones within each patient are the least responsive to isoproterenol. On the other hand, in response to nitroprusside, the scar and the CM-NL tissue appear to be the least responsive, even paradoxically increasing their ARI in comparison with border-zone regions. Therefore, it appears that the most denervated regions have the greatest response to catecholamines, consistent with denervation super-sensitivity.
Recordingcatheter
Recordingcatheter
AICD
10 mV
5 mV
0 mV
AICD
RAOCS
CS
RV
LAO
P = .016
20
Del
ta A
RI (
ms)
10
0
10
20
-30
-40
-50
-60
-70
-80
-90
P = .08
P = .01
P = .0002
Normal hearts CM-normal site CM-border zones CM-scars
Isoproterenol
Nitroprusside
1 2 34
56 7 8
9 10 12
11 13
1415

412 NEURAL CONTROL OF CARDIAC ELECTRICAL ACTIVITY
Figure 41-4. Human cardiomyopathy is associated with stellate ganglion neuronal hypertrophy. Shown in the figure are representative thionine-stained right stellate ganglion neurons from normal controls (NL) and from patients with ischemic (ICM) and nonischemic cardiomyopathy (NICM). Neurons from ICM and NICM are larger and show a more granular appearance compared with normal controls. (Magnification 20×; scale bar 50 µm.)
NL ICM NICM
400
Mea
n ne
uron
al a
rea
(µm
2 )
350
300
250
NL ICM NICM
cell transplantation,34 in radiofrequency ablation,35 and in rapid pacing–induced heart failure in dogs36 and in hypercholesterol-emic rabbits, where it has been shown to cause QTc dispersion, increased heterogeneity of repolarization, and significantly increased episodes of VF, both spontaneous and induced.37 Fur-thermore, NGF infusion into the LSG promotes nerve sprouting in dogs with MI and complete AV block, and is associated with increased incidence of VF.38 Infusion of NGF into the RSG has not been shown to increase the risk of sudden cardiac death (SCD).39 Of note, NGF and GAP-43 levels are increased in the LSG of these dogs 3 days after MI, without a concomitant increase in mRNA levels, indicating possible retrograde trans-portation of these proteins to the LSG, which then triggers nerve sprouting at noninfarcted LV sites.40
Heart failure can cause trans-differentiation of cardiac sym-pathetic nerves. Cholinergic trans-differentiation of nerve sprouts by production of interleukin-6 cytokines through a gp-130 sig-naling pathway has been demonstrated in rodents,41,42 potentially further promoting heterogeneity in repolarization. However, the exact ramifications of this sympathetic rejuvenation and plasticity are yet unknown. Heart failure is also known to cause remodeling of cardiac ion channels, including increased L-type Ca (ICaL) density, decreased potassium current, decreased Ito density, and changes in Cl and Ca transporters and enzymes in the border zones surrounding the infarct.43-46 Thus, sympathetic stimulation could result in complex effects on the APD and restitution, which, along with increased ICaL density, can lead to intracellular Ca2+ overload–induced triggered activity, potentiating the risk of spontaneous ventricular arrhythmias.
Extracardiac Neural Remodeling
In addition to cardiac neural remodeling, electroanatomical remodeling of the LSG in the setting of MI and heart failure has been described. Increased nerve density along with increased mRNA levels of NGF and GAP-43 in the LSG of dogs with MI
and nerve sprouting has been reported.40 Increased stellate gan-glion nerve activity (SGNA) immediately after MI was associated with intramyocardial nerve sprouts, as well as increased neuronal size and synaptic density in the LSG and RSG.47 Although SCD was not observed in this study, Ogawa et al and Zhou et al had previously shown that sympathetic nerve discharges tend to precede ventricular arrhythmias in the same dog model of MI.48,49 Similarly, in human cadavers with evidence of cardiac scar, neu-ronal number in the LSG is increased as compared with cadavers without evidence of scar.50 Compared with cadavers with normal hearts, those with cardiomyopathy demonstrate increased neuro-nal size and synaptophysin density (Figure 41-4).51 These results provide a possible mechanistic link between neural remodeling (nerve sprouting within the myocardium) and remodeling of the stellate ganglion and ventricular arrhythmias, although further studies are required. Interaction between areas of denervation, regional nerve sprouting and trans-differentiation (neural remod-eling), electrical remodeling due to heart failure, and electroana-tomical remodeling of the stellate ganglia all combine to create a substrate that can be conducive for ventricular arrhythmias and sudden cardiac death (Figure 41-5).
Proarrhythmic Effects of Sympathetic Stimulation
Many studies have suggested that sympathetic nerve stimulation, particularly LSG stimulation, is proarrhythmic. Greater increases in the amplitude of early afterdepolarization have been observed with LSG stimulation.52 Priori et al showed that LSG stimulation caused delayed afterdepolarizations in vivo in cat hearts, suggest-ing triggered activity as the mechanism of ventricular arrhyth-mogenesis.53 Further, an increase in dispersion with sympathetic stimulation, both during ischemia and in normal canine and porcine hearts, has been observed.54-57 In the porcine heart, LSG stimulation increased ARI dispersion by 4-fold and caused VF in

SYMPATHETIC INNERVATION, DENERVATION, AND CARDIAC ARRHYTHMIAS 413
41
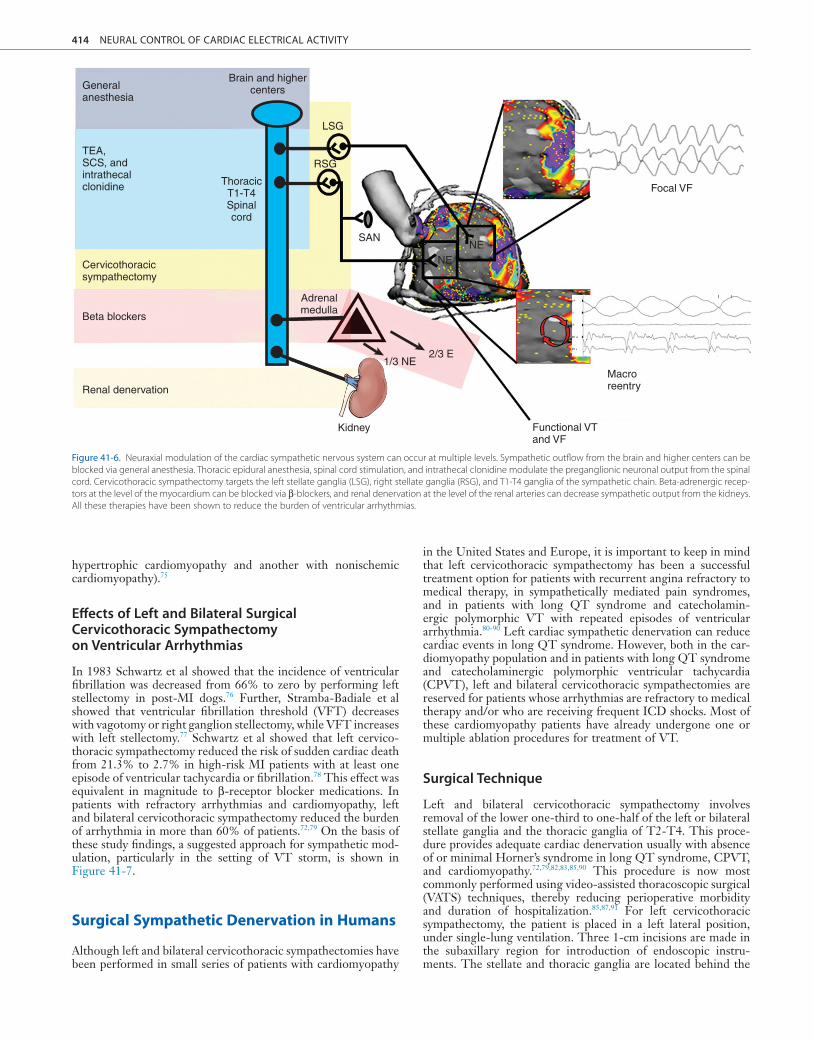
cord, the sympathetic chain, and the β-adrenergic receptors of the myocardium, and it can be attempted by renal denervation (Figure 41-6). General anesthesia, sedation, and intubation can suppress or significantly reduce the burden of ventricular arrhyth-mias and implantable cardioverter-defibrillator (ICD) shocks by reducing the sympathetic drive in electrical storm.62-65 At the spinal cord level, Issa et al demonstrated that in a canine model of ischemic cardiomyopathy, spinal cord stimulation at T1-T2 segments reduced the incidence of ventricular arrhythmia from 59% to 23% during ischemia.66 A simultaneous decrease in heart rate and systolic blood pressure, consistent with the antisympa-thetic effects of spinal cord stimulation, was also observed.67-70 Intrathecal clonidine, when delivered via a catheter at T2-T4 spinal segments, also significantly reduces the occurrence of ven-tricular tachycardia and fibrillation during transient myocardial ischemia.71 The benefit of thoracic epidural anesthesia (TEA) was reported in series of patients with cardiomyopathy and refractory VT. In 66% of the patients, TEA reduced the burden of arrhyth-mias by 80%.72
In humans, postganglionic sympathetic blockade, with medi-cations (β-adrenergic receptor blockers) or via percutaneous stel-late blockade (usually injection of bupivacaine 0.25%), has also been reported to reduce ventricular arrhythmias. Among patients early after MI with recurrent VF (electrical storm), those treated with sympathetic blockade had improved outcomes as compared with those treated with the standard Advanced Cardiac Life Support (ACLS) protocol.73 Sympathetic blockade was estab-lished with the use of LSG blockade in 6 patients and with infu-sions of either propranolol or esmolol in 21 patients without antiarrhythmic therapy, as recommended by ACLS. One-week and 1-year mortality were significantly higher in the group undergoing standard ACLS protocol compared with the sympathetic blockade group (82% vs. 22% at 1 week, 95% vs. 33% at 1 year, respectively).73 It is important to note that in humans with percutaneous RSG or LSG blockade, no significant changes in hemodynamics or ejection fraction during rest or with exercise have been reported.74 Finally, renal denervation has been reported to reduce the burden of ventricular arrhythmias in two patients, as well as electrical storm (one with nonobstructive
25% of the animals.57 In open-chest dogs, electrical stimulation of LSG, the left middle cervical or left caudal pole of the cardio-pulmonary nerve, or the ventrolateral nerve caused VT in 13 of 22 normal dog hearts, with isochronal mapping showing the earliest electrical excitation occurring on the posterior aspect of the ventricles.58 Electrical stimulation of the left ansa subclavia in open-chest dogs during left circumflex occlusion increased the incidence of VF from 35% to 73%.59 The incidence of induced ventricular arrhythmias in dogs with myocardial ischemia increased from 54% to 68% and 63% with LSG and bilateral stellate stimulation, respectively. RSG stimulation had no signifi-cant effect on the incidence of arrhythmias in this study.60 Further, by measuring local VF intervals, Opthof et al showed that LSG stimulation can increase dispersion in refractoriness by shorten-ing refractoriness across nonischemic sites while either not changing or increasing refractoriness at ischemic sites during coronary occlusion, thereby increasing dispersion across the ischemic border by 14% to 59%.55 Finally, subthreshold LSG stimulation has been shown to increase nerve sprouting in dogs with MI. These dogs have a greater number of episodes of VT compared with controls, suggesting that additional mechanisms underlie the increase in dispersion seen in infarcted hearts.61
Effect of Neuraxial Modulation in Reducing Risk of Ventricular Arrhythmias
As augmented sympathetic tone increases the risk of arrhythmias, interventions that aim to reduce sympathetic tone diminish the risk of sudden cardiac death and ventricular arrhythmias.
Strategies and Techniques of Cardiac Sympathetic Denervation and Effects on Ventricular Arrhythmias
Modulation and blockade of the sympathetic nervous system can be attempted at multiple levels, including the brain, the spinal
Figure 41-5. The schematic shows the various changes that occur as a result of myocardial infarction that cause neural remodeling and increased sympathetic activation. Myocardial infarction causes scar formation and denervation leading to release of nerve growth factor (NGF) and cytokines by the surrounding myocardium. This causes remodeling of the stellate ganglion and neural remodeling of the myocardium, including nerve sprout formation and cholinergic trans-differentiating, further causing heterogeneity in repolarization with sympathetic stimulation. Sympathetic activation then causes EADs, DADs, and increased dispersion in this substrate, leading to ven-tricular arrhythmias. DAD, delayed afterdepolarization; EAD, early afterdepolarization; NGF, neural growth factor.
Myocardialinfarction
Scar &denervation
NGFCytokines
Cytokin
es
NGF
Heterogenoussubstrate
Sympatheticactivation
Dispersion ofrepolarization
EADs
DADsVT/VF
+
+
Stellate ganglionremodeling
ScarNerve
sprouts
Normal
414 NEURAL CONTROL OF CARDIAC ELECTRICAL ACTIVITY
in the United States and Europe, it is important to keep in mind that left cervicothoracic sympathectomy has been a successful treatment option for patients with recurrent angina refractory to medical therapy, in sympathetically mediated pain syndromes, and in patients with long QT syndrome and catecholamin-ergic polymorphic VT with repeated episodes of ventricular arrhythmia.80-90 Left cardiac sympathetic denervation can reduce cardiac events in long QT syndrome. However, both in the car-diomyopathy population and in patients with long QT syndrome and catecholaminergic polymorphic ventricular tachycardia (CPVT), left and bilateral cervicothoracic sympathectomies are reserved for patients whose arrhythmias are refractory to medical therapy and/or who are receiving frequent ICD shocks. Most of these cardiomyopathy patients have already undergone one or multiple ablation procedures for treatment of VT.
Surgical Technique
Left and bilateral cervicothoracic sympathectomy involves removal of the lower one-third to one-half of the left or bilateral stellate ganglia and the thoracic ganglia of T2-T4. This proce-dure provides adequate cardiac denervation usually with absence of or minimal Horner’s syndrome in long QT syndrome, CPVT, and cardiomyopathy.72,79,82,83,85,90 This procedure is now most commonly performed using video-assisted thoracoscopic surgical (VATS) techniques, thereby reducing perioperative morbidity and duration of hospitalization.85,87,91 For left cervicothoracic sympathectomy, the patient is placed in a left lateral position, under single-lung ventilation. Three 1-cm incisions are made in the subaxillary region for introduction of endoscopic instru-ments. The stellate and thoracic ganglia are located behind the
hypertrophic cardiomyopathy and another with nonischemic cardiomyopathy).75
Effects of Left and Bilateral Surgical Cervicothoracic Sympathectomy on Ventricular Arrhythmias
In 1983 Schwartz et al showed that the incidence of ventricular fibrillation was decreased from 66% to zero by performing left stellectomy in post-MI dogs.76 Further, Stramba-Badiale et al showed that ventricular fibrillation threshold (VFT) decreases with vagotomy or right ganglion stellectomy, while VFT increases with left stellectomy.77 Schwartz et al showed that left cervico-thoracic sympathectomy reduced the risk of sudden cardiac death from 21.3% to 2.7% in high-risk MI patients with at least one episode of ventricular tachycardia or fibrillation.78 This effect was equivalent in magnitude to β-receptor blocker medications. In patients with refractory arrhythmias and cardiomyopathy, left and bilateral cervicothoracic sympathectomy reduced the burden of arrhythmia in more than 60% of patients.72,79 On the basis of these study findings, a suggested approach for sympathetic mod-ulation, particularly in the setting of VT storm, is shown in Figure 41-7.
Surgical Sympathetic Denervation in Humans
Although left and bilateral cervicothoracic sympathectomies have been performed in small series of patients with cardiomyopathy
Figure 41-6. Neuraxial modulation of the cardiac sympathetic nervous system can occur at multiple levels. Sympathetic outflow from the brain and higher centers can be blocked via general anesthesia. Thoracic epidural anesthesia, spinal cord stimulation, and intrathecal clonidine modulate the preganglionic neuronal output from the spinal cord. Cervicothoracic sympathectomy targets the left stellate ganglia (LSG), right stellate ganglia (RSG), and T1-T4 ganglia of the sympathetic chain. Beta-adrenergic recep-tors at the level of the myocardium can be blocked via β-blockers, and renal denervation at the level of the renal arteries can decrease sympathetic output from the kidneys. All these therapies have been shown to reduce the burden of ventricular arrhythmias.
Generalanesthesia
Brain and highercenters
LSG
RSG
SAN
Adrenalmedulla
Kidney Functional VTand VF
Macroreentry
NE
Focal VF
NE
ThoracicT1-T4Spinalcord
TEA,SCS, andintrathecalclonidine
Cervicothoracicsympathectomy
Beta blockers
Renal denervation
2/3 E1/3 NE

SYMPATHETIC INNERVATION, DENERVATION, AND CARDIAC ARRHYTHMIAS 415
41 parietal pleura, in the paravertebral position. Then, the lower one-third to one-half of the stellate ganglion, along with the chain from T2-T4, is completely removed. Histopathologic con-firmation of neuronal cell bodies is obtained during the proce-dure. The procedure can be typically performed on one side in less than 60 minutes.82
Risks and Complications of Cervicothoracic Sympathectomy
Left cervicothoracic sympathectomy can be accompanied imme-diately postoperatively by ipsilateral Horner’s syndrome. However, when the VATS approach is used, it occurs rarely and has been temporary because the sympathetic fibers directed toward the ocular regions usually cross the upper portion of the LSG, which is typically spared in this procedure.82,85 Mild lower-ing of the left eyelid appears transiently after surgery, but seldom persists.82
The effect of left cervicothoracic sympathectomy on LV func-tion has been studied using echocardiography. In patients with sympathetically mediated pain syndrome but normal ventricular function, hemodynamic parameters including heart rate and blood pressure remain unchanged, although afterload reduction—a desired effect in cardiomyopathy patients—does occur.89 Fur-thermore, global and regional systolic function also remains unchanged, although isovolumic relaxation time is slightly pro-longed.88,89 This small effect has not compromised ventricular function, and stroke volume has actually increased as a result of afterload reduction.89
Conclusion
The sympathetic nervous system plays a major role in the patho-genesis of ventricular arrhythmias. Therapies that inhibit the effects of this system have been shown to reduce the burden of arrhythmias. Left and/or bilateral sympathetic denervation tech-niques are feasible and safe options for patients with refractory arrhythmias who have not responded to medical therapy and/or ventricular tachycardia ablation. Further, they may be the only option for patients with refractory arrhythmias who are not can-didates for cardiac transplantation. Randomized studies with longer follow-up are required to assess the long-term risks and benefits of these procedures.
Acknowledgments
Some of the research reported in this chapter is supported by the National Heart, Lung, and Blood Institute grant R01HL084261 (Dr. Shivkumar) and American Heart Association AHA #11FTF6559994 (Dr. Vaseghi).Figure 41-7. A suggested approach for neuraxial modulation of a patient with
ventricular tachycardia (VT) storm is shown.
YES
NO
NO
NO
YES
NO
NO
YES
YES
YES
YES
NO
Adjust TEAlevel
Patient with severe ventricular
arrhythmias
Reversible causes suppressed
arrhythmias?Devise optimal medical therapy and consider substrate mapping if indicated.
Stabilize patient and plan elective catheter mapping and ablation.
Devise optimal medical therapy and ICD implantation if not already present.
Proceed with cardiac transplant work up and listing.
Perform left cardiac sympatheticdenervation (LCSD)
Consider bilateral cardiac sympathetic denervation (BCSD)
Pharmacologic therapy
successful?
Thoracic epidural anesthesia (TEA)
successful?
Emergent catheter mapping and ablation. VT
remains inducible?
Intubation and sedationsuccessful?
Cardiac transplant candidate?
Plan cardiac sympatheticdenervation
(CSD).
If arrhythmia is RVOT, A RVC,
PMVT, RV only scar, or no cardiac scar
If arrhythmia is LV origin, or LV-only scar.
References1. Schwartz PJ, Vanoli E, Stramba-Badiale M, et al:
Autonomic mechanisms and sudden death: New insights from analysis of baroreceptor reflexes in conscious dogs with and without a myocardial infarction. Circulation 78:969–979, 1988.
2. Vaseghi M, Shivkumar K: The role of the auto-nomic nervous system in sudden cardiac death. Prog Cardiovasc Dis 50:404–419, 2008.
3. Zipes DP, Barber MJ, Takahashi N, et al: Influence of the autonomic nervous system on the genesis of
cardiac arrhythmias. Pacing Clin Electrophysiol 6:1210–1220, 1983.
4. Jänig W: Functional anatomy of the peripheral sympathetic and parasympathetic system. In Jänig W, editor: The Integrative Action of the Auto-nomic Nervous System: Neurobiology of Homeo-stasis, Cambridge, 2006, Cambridge University Press, pp 13–34.
5. Janes RD, Brandys JC, Hopkins DA, et al: Anatomy of human extrinsic cardiac nerves and ganglia. Am J Cardiol 57:299–309, 1986.
6. Kawashima T: Anatomy of the cardiac nervous system with clinical and comparative morpho-
logical implications. Anat Sci Int 86:30–49, 2011.
7. Angelakos E, King M, Millard R: Regional distri-bution of catecholamines in the heart of various species. Ann N Y Acad Sci 156:219–240, 1969.
8. Hutchins GD, Miller MA, Zips DP: Neurocardiac imaging. In Zipes DP, Jalife J, editors: Cardiac Electrophysiology: From Cell to Bedside, Philadel-phia, 2009, Saunders Elsevier, pp 391–403.
9. Armour JA: Myocardial ischaemia and the cardiac nervous system. Cardiovasc Res 41:41–54, 1999.
10. Pauza DH, Skripka V, Pauziene N, et al: Morphol-ogy, distribution, and variability of the epicardiac

416 NEURAL CONTROL OF CARDIAC ELECTRICAL ACTIVITY
neural ganglionated subplexuses in the human heart. Anat Rec 259:353–382, 2000.
11. Marron K, Wharton J, Sheppard MN, et al: Dis-tribution, morphology, and neurochemistry of endocardial and epicardial nerve terminal arboriza-tions in the human heart. Circulation 92:2343–2351, 1995.
12. Esler M, Jennings G, Lambert G, et al: Overflow of catecholamine neurotransmitters to the circula-tion: Source, fate, and functions. Physiol Rev 70:963–985, 1990.
13. Gabella G: Fine structure of post-ganglionic nerve fibers and autonomic neuroeffector junctions. In Burnstock G, Hoyle CHV, editors: Autonomic Neuroeffector Mechanisms, Chur, 1995, Harwood Academic Publishers, pp 1–31.
14. Rubart M, Zipes DP: Mechanisms of sudden cardiac death. J Clin Invest 115:2305–2315, 2005.
15. Meredith IT, Eisenhofer G, Lambert GW, et al: Cardiac sympathetic nervous activity in congestive heart failure: Evidence for increased neuronal nor-epinephrine release and preserved neuronal uptake. Circulation 88:136–145, 1993.
16. Bristow M: Changes in myocardial and vascular receptors in heart failure. J Am Coll Cardiol 22:61A–71A, 1993.
17. Bengel FM, Schwaiger M: Assessment of cardiac sympathetic neuronal function using PET imaging. J Nucl Cardiol 11:603–616, 2004.
18. Riemann B, Shafers M, Law M, et al: Radioligands for imaging myocardial alpha- and beta-adrenoreceptors. Nuklearmedizin 42:4–9, 2003.
19. Inoue H, Zipes DP: Time course of denervation of efferent sympathetic and vagal nerves after occlu-sion of the coronary artery in the canine heart. Circ Res 62:1111–1120, 1988.
20. Li W, Knowlton D, Van Winkle DM, et al: Infarc-tion alters both the distribution and noradrenergic properties of cardiac sympathetic neurons. Am J Physiol Heart Circ Physiol 286:H2229–H2236, 2004.
21. Barber MJ, Mueller TM, Henry DP, et al: Trans-mural myocardial infarction in the dog produces sympathectomy in noninfarcted myocardium. Cir-culation 67:787–796, 1983.
22. Kammerling JJ, Green FJ, Watanabe AM, et al: Denervation supersensitivity of refractoriness in noninfarcted areas apical to transmural myocardial infarction. Circulation 76:383–393, 1987.
23. Warner MR, Wisler PL, Hodges TD, et al: Mech-anisms of denervation supersensitivity in regionally denervated canine hearts. Am J Physiol 264:H815–H820, 1993.
24. Vaseghi M, Lux RL, Mahajan A, et al: Sympathetic stimulation increases dispersion of repolarization in humans with myocardial infarction. Am J Physiol Heart Circ Physiol 302:H1838–H1846, 2012.
25. Sjoberg J, Kanje M: The initial period of peripheral nerve regeneration and the importance of the local environment for the conditioning lesion effect. Brain Res 529:79–84, 1990.
26. Fu SY, Gordon T: The cellular and molecular basis of peripheral nerve regeneration. Mol Neurobiol 14:67–116, 1997.
27. Rabinovitch MA, Rose CP, Rouleau JL, et al: Metaiodobenzylguanidine [131I] scintigraphy detects impaired myocardial sympathetic neuronal transport function of canine mechanical-overload heart failure. Circ Res 61:797–804, 1987.
28. Parthenakis FI, Prassopoulos VK, Koukouraki SI, et al: Segmental pattern of myocardial sympathetic denervation in idiopathic dilated cardiomyopathy: Relationship to regional wall motion and myocar-dial perfusion abnormalities. J Nucl Cardiol 9:15–22, 2002.
29. Vracko R, Thorning D, Frederickson RG: Nerve fibers in human myocardial scars. Hum Pathol 22:138–146, 1991.
30. Vracko R, Thorning D, Frederickson RG: Fate of nerve fibers in necrotic, healing, and healed rat myocardium. Lab Invest 63:490–501, 1990.
31. Cao JM, Fishbein MC, Han JB, et al: Relationship between regional cardiac hyperinnervation and
ventricular arrhythmia. Circulation 101:1960–1969, 2000.
32. Dillon SM, Allessie MA, Ursell PC, et al: Influ-ences of anisotropic tissue structure on reentrant circuits in the epicardial border zone of subacute canine infarcts. Circ Res 63:182–206, 1988.
33. Gough WB, Mehra R, Restivo M, et al: Reentrant ventricular arrhythmias in the late myocardial infarction period in the dog: Correlation of activa-tion and refractory maps. Circ Res 57:432–442, 1985.
34. Pak HN, Qayyum M, Kim DT, et al: Mesenchymal stem cell injection induces cardiac nerve sprouting and increased tenascin expression in a Swine model of myocardial infarction. J Cardiovasc Electro-physiol 14:841–848, 2003.
35. Okuyama Y, Pak HN, Miyauchi Y, et al: Nerve sprouting induced by radiofrequency catheter abla-tion in dogs. Heart Rhythm 1:712–717, 2004.
36. Cha YM, Redfield MM, Shah S, et al: Effects of omapatrilat on cardiac nerve sprouting and struc-tural remodeling in experimental congestive heart failure. Heart Rhythm 2:984–990, 2005.
37. Liu YB, Wu CC, Lu LS, et al: Sympathetic nerve sprouting, electrical remodeling, and increased vul-nerability to ventricular fibrillation in hypercholes-terolemic rabbits. Circ Res 92:1145–1152, 2003.
38. Cao JM, Chen LS, KenKnight BH, et al: Nerve sprouting and sudden cardiac death. Circ Res 86:816–821, 2000.
39. Zhou S, Cao JM, Tebb ZD, et al: Modulation of QT interval by cardiac sympathetic nerve sprout-ing and the mechanisms of ventricular arrhythmia in a canine model of sudden cardiac death. J Car-diovasc Electrophysiol 12:1068–1073, 2001.
40. Zhou S, Chen LS, Miyauchi Y, et al: Mechanisms of cardiac nerve sprouting after myocardial infarc-tion in dogs. Circ Res 95:76–83, 2004.
41. Kanazawa H, Ieda M, Kimura K, et al: Heart failure causes cholinergic transdifferentiation of cardiac sympathetic nerves via gp130-signaling cytokines in rodents. J Clin Invest 120:408–421, 2010.
42. Kimura K, Ieda M, Fukuda K: Development, mat-uration, and transdifferentiation of cardiac sympa-thetic nerves. Circ Res 110:325–336, 2012.
43. Huang B, Qin D, El-Sherif N: Early down-regulation of K+ channel genes and currents in the postinfarction heart. J Cardiovasc Electrophysiol 11:1252–1261, 2000.
44. Gidh-Jain M, Huang B, Jain P, et al: Differential expression of voltage-gated K+ channel genes in left ventricular remodeled myocardium after experi-mental myocardial infarction. Circ Res 79:669–675, 1996.
45. Qin D, Zhang ZH, Caref EB, et al: Cellular and ionic basis of arrhythmias in postinfarction remod-eled ventricular myocardium. Circ Res 79:461–473, 1996.
46. Heath BM, Xia J, Dong E, et al: Overexpression of nerve growth factor in the heart alters ion channel activity and beta-adrenergic signalling in an adult transgenic mouse. J Physiol 512(Pt 3):779–791, 1998.
47. Han S, Kobayashi K, Joung B, et al: Electroana-tomic remodeling of the left stellate ganglion after myocardial infarction. J Am Coll Cardiol 59:954–961, 2012.
48. Zhou S, Jung BC, Tan AY, et al: Spontaneous stel-late ganglion nerve activity and ventricular arrhythmia in a canine model of sudden death. Heart Rhythm 5:131–139, 2008.
49. Ogawa M, Zhou S, Tan AY, et al: Left stellate ganglion and vagal nerve activity and cardiac arrhythmias in ambulatory dogs with pacing-induced congestive heart failure. J Am Coll Cardiol 50:335–343, 2007.
50. Wood A, Docimo S, Elkowitz DE: Cardiovascular disease and its association with histological changes of the left stellate ganglion. Clin Med Insights Pathol 3:19–24, 2010.
51. Ajijola OA, Wico J, Lambert HW, et al: Chronic myocardial infarction is associated with neural remodeling in human stellate ganglia. Heart Rhythm 9:261, 2012.
52. Ben-David J, Zipes DP: Differential response to right and left ansae subclaviae stimulation of early afterdepolarizations and ventricular tachycardia induced by cesium in dogs. Circulation 78:1241–1250, 1988.
53. Priori SG, Mantica M, Schwartz PJ: Delayed after-depolarizations elicited in vivo by left stellate gan-glion stimulation. Circulation 78:178–185, 1988.
54. Opthof T, Coronel R, Vermeulen JT, et al: Disper-sion of refractoriness in normal and ischaemic canine ventricle: effects of sympathetic stimulation. Cardiovasc Res 27:1954–1960, 1993.
55. Opthof T, Dekker LR, Coronel R, et al: Interac-tion of sympathetic and parasympathetic nervous system on ventricular refractoriness assessed by local fibrillation intervals in the canine heart. Car-diovasc Res 27:753–759, 1993.
56. Opthof T, Misier AR, Coronel R, et al: Dispersion of refractoriness in canine ventricular myocardium: Effects of sympathetic stimulation. Circ Res 68:1204–1215, 1991.
57. Vaseghi M, Zhou W, Shi J, et al: Sympathetic innervation of the anterior left ventricular wall by the right and left stellate ganglia. Heart Rhythm 9:1303–1309, 2012.
58. Cardinal R, Savard P, Armour JA, et al: Mapping of ventricular tachycardia induced by thoracic neural stimulation in dogs. Can J Physiol Pharma-col 64:411–418, 1986.
59. Euler DE, Nattel S, Spear JF, et al: Effect of sym-pathetic tone on ventricular arrhythmias during circumflex coronary occlusion. Am J Physiol 249:H1045–H1050, 1985.
60. Gantenberg NS, Hageman GR: Enhanced induc-tion of ventricular arrhythmias during sympathetic stimulation before and during coronary artery occlusion. Int J Cardiol 34:75–83, 1992.
61. Swissa M, Zhou S, Gonzalez-Gomez I, et al: Long-term subthreshold electrical stimulation of the left stellate ganglion and a canine model of sudden cardiac death. J Am Coll Cardiol 43:858–864, 2004.
62. Burjorjee JE, Milne B: Propofol for electrical storm; a case report of cardioversion and suppres-sion of ventricular tachycardia by propofol. Can J Anaesth 49:973–977, 2002.
63. Dijkman B, den Dulk K, Wellens HJ: Management of electrical instability after ICD implantation. Pacing Clin Electrophysiol 18:148–151, 1995.
64. Mulpuru SK, Patel DV, Wilbur SL, et al: Electrical storm and termination with propofol therapy: A case report. Int J Cardiol 128:e6–e8, 2008.
65. Tavernier R, Derycke E, Jordaens L: An arrhyth-mia storm dependent on adrenergic drive late after implantation of an internal cardioverter defi-brillator. Eur J Pacing Electrophysiol 7:33–36, 1997.
66. Issa ZF, Zhou X, Ujhelyi MR, et al: Thoracic spinal cord stimulation reduces the risk of ischemic ven-tricular arrhythmias in a postinfarction heart failure canine model. Circulation 111:3217–3220, 2005.
67. Foreman RD, Linderoth B, Ardell JL, et al: Modu-lation of intrinsic cardiac neurons by spinal cord stimulation: Implications for its therapeutic use in angina pectoris. Cardiovasc Res 47:367–375, 2000.
68. Mannheimer C, Eliasson T, Andersson B, et al: Effects of spinal cord stimulation in angina pectoris induced by pacing and possible mechanisms of action. Br Med J 307, 1993.
69. Sanderson JE, Ibrahim B, Waterhouse D, et al: Spinal electrical stimulation for intractable angina: Long-term clinical outcome and safety. Eur Heart J 15:810–814, 1994.
70. Eliasson T, Augustinsson LE, Mannheimer C: Spinal cord stimulation in severe angina pectoris-presentation of current studies, indications, and clinical experience. Pain 65:169–179, 1996.
71. Issa ZF, Ujhelyi MR, Hildebrand KR, et al: Intra-thecal clonidine reduces the incidence of ischemia-provoked ventricular arrhythmias in a canine postinfarction heart failure model. Heart Rhythm 2:1122–1127, 2005.
72. Bourke T, Vaseghi M, Michowitz Y, et al: Neur-axial modulation for refractory ventricular

SYMPATHETIC INNERVATION, DENERVATION, AND CARDIAC ARRHYTHMIAS 417
41 arrhythmias: Value of thoracic epidural anesthesia and surgical left cardiac sympathetic denervation. Circulation 121:2255–2262, 2010.
73. Nademanee K, Taylor R, Bailey WE, et al: Treat-ing electrical storm: Sympathetic blockade versus advanced cardiac life support-guided therapy. Cir-culation 102:742–747, 2000.
74. Gardner MJ, Kimber S, Johnstone DE, et al: The effects of unilateral stellate ganglion blockade on human cardiac function during rest and exercise. J Cardiovasc Electrophysiol 4:2–8, 1993.
75. Ukena C, Bauer A, Mahfoud F, et al: Renal sym-pathetic denervation for treatment of electrical storm: First-in-man experience. Clin Res Cardiol 101:63–67, 2012.
76. Schwartz PJ, Billman GE, Stone HL: Autonomic mechanisms in ventricular fibrillation induced by myocardial ischemia during exercise in dogs with healed myocardial infarction: An experimental preparation for sudden cardiac death. Circulation 69:790–800, 1984.
77. Stramba-Badiale M, Lazzarotti M, Schwartz PJ: Development of cardiac innervation, ventricular fibrillation, and sudden infant death syndrome. Am J Physiol 263:H1514–H1522, 1992.
78. Schwartz PJ, Motolese M, Pollavini G, et al: Pre-vention of sudden cardiac death after a first myo-cardial infarction by pharmacologic or surgical
antiadrenergic interventions. J Cardiovasc Electro-physiol 3:2–16, 1992.
79. Ajijola OA, Lellouche N, Bourke T, et al: Bilateral cardiac sympathetic denervation for the manage-ment of electrical storm. J Am Coll Cardiol 59:91–92, 2012.
80. Tygesen H, Wettervik C, Claes G, et al: Long-term effect of endoscopic transthoracic sympathicotomy on heart rate variability and QT dispersion in severe angina pectoris. Int J Cardiol 70:283–292, 1999.
81. Stephenson EA, Berul CI: Electrophysiological interventions for inherited arrhythmia syndromes. Circulation 116:1062–1080, 2007.
82. Schwartz PJ, Priori SG, Cerrone M, et al: Left cardiac sympathetic denervation in the manage-ment of high-risk patients affected by the long-QT syndrome. Circulation 109:1826–1833, 2004.
83. Li C, Hu D, Shang L, et al: Surgical left cardiac sympathetic denervation for long QT syndrome: Effects on QT interval and heart rate. Heart Vessels 20:137–141, 2005.
84. Bhandari AK, Scheinman MM, Morady F, et al: Efficacy of left cardiac sympathectomy in the treat-ment of patients with the long QT syndrome. Cir-culation 70:1018–1023, 1984.
85. Li J, Wang L, Wang J: Video-assisted thoraco-scopic sympathectomy for congenital long QT
syndromes. Pacing Clin Electrophysiol 26:870–873, 2003.
86. Epstein AE, Rosner MJ, Hageman GR, et al: Pos-terior left thoracic cardiac sympathectomy by surgi-cal division of the sympathetic chain: An alternative approach to treatment of the long QT syndrome. Pacing Clin Electrophysiol 19:1095–1104, 1996.
87. Ouriel K, Moss AJ: Long QT syndrome: an indica-tion for cervicothoracic sympathectomy. Cardio-vasc Surg 3:475–478, 1995.
88. Lobato EB, Kern KB, Paige GB, et al: Differential effects of right versus left stellate ganglion block on left ventricular function in humans: an echocar-diographic analysis. J Clin Anesth 12:315–318, 2000.
89. Schlack W, Dinter W: Haemodynamic effects of a left stellate ganglion block in ASA I patients: An echocardiographic study. Eur J Anaesthesiol 17:79–84, 2000.
90. Collura CA, Johnson JN, Moir C, et al: Left cardiac sympathetic denervation for the treatment of long QT syndrome and catecholaminergic poly-morphic ventricular tachycardia using video-assisted thoracic surgery. Heart Rhythm 6:752–759, 2009.
91. Josephs LG, Menzoian JO: Technical consider-ations in endoscopic cervicothoracic sympathec-tomy. Arch Surg 131:355–359, 1996.