Embed Size (px)
Citation preview

lable at ScienceDirect
Soil Biology & Biochemistry 42 (2010) 543e550
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lb io
Carbon decomposition in broiler litter-amended soils
Alassane Sissoko, Kokoasse Kpomblekou-A*
Department of Agricultural and Environmental Sciences, Tuskegee University, Tuskegee, AL 36088, USA
a r t i c l e i n f o
Article history:Received 5 March 2009Received in revised form6 October 2009Accepted 8 October 2009Available online 30 October 2009
Keywords:Organic carbonAnimal manuresMineralization of organic CC sequestration in soilsHalf-life of organic COrganic matter
* Corresponding author. Tel.: þ1 334 724 4521; faxE-mail address: [email protected] (K. Kpombleko
0038-0717/$ e see front matter � 2009 Elsevier Ltd.doi:10.1016/j.soilbio.2009.10.007
a b s t r a c t
Organic carbon (OC) is generally low in Alabama (U.S.A.) soils and varies considerably with croppingsystems. Information on decomposition rates of the added C is a prerequisite to designing strategies thatimprove C sequestration in farming systems. Different models including exponential models have beenused to describe OC mineralization in soils as well as to describe its potential as CO2 to be released intothe environment. We investigated the decomposition of broiler litter added to ten non-calcareous soils(Appling, Troup, Cecil, Decatur, Sucarnoochee, Linker, Hartsells, Dothan, Maytag, and Colbert soils). Anon-linear regression approach for N mineralization was used to estimate the potentially mineralizableOC pools (Co) and the first-order rate constant (k) in the soil samples. Results showed that the non-amended soils have distinct differences in their ability to release their native OC as CO2 and can bedivided into four groups depending on their potentially mineralizable C (Co) and their ability to protectstable organic matter. Sucarnoochee soil represents the first group and contains a moderate amount ofOC (11.4 g C kg�1) but had the highest Co (7.30 g C kg�1 soil). The second distinct group of soils has Co
varying between 5.50 and 5.00 g C kg�1 soil (Decatur, Hartsells, Dothan, and Maytag). The third group hasCo between 5.00 and 4.00 (Appling, Cecil, and Linker). The fourth group has Co less than 4.00 g C kg�1 soil(Troup and Colbert). Half-life of C remaining in non-amended soils varied from 26 days in Maytag soil to139 days in Cecil soil. The OC in these non-amended soils represents a very stable form of organic C andthus, not easily decomposed by soil microorganisms. In the broiler litter-amended soils, the Co variedfrom 3.82 g C kg�1 in Appling soil amended with broiler litter 1e7.04 g C kg�1 soil in Maytag amendedwith broiler litter 2. Decomposition of the added OC proceeded in two phases with less than 31%decomposed in 43 days. Potentially mineralizable organic C (Co) was related to soil organic C (r ¼ 0.661**)and soil C/N ratio (r ¼ 0.819*).
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Soil organic matter (SOM) is composed of a living part repre-sented by soil organisms and their remains and a non-living partrepresented by dissolved organic matter, particulate organicmatter, humus, and inert organic matter. The amount of SOMpresent at any given time is determined by the balance between theamounts of C added (input) and decomposed (output). This balanceitself depends on soil forming factors such as parent material,climate, topography, organisms, and time that determine soil types.Thus, generally, organic carbon (OC) levels tend to vary among soiltypes and correlate well with a zonal distribution of soils in relationto climatic factors. Traditional agricultural practices such as landclearing and continuous cultivation of soil have led to land degra-dation characterized by a rapid decomposition of SOC witha subsequent loss of C as CO2 emitted into the atmosphere. During
: þ1 334 724 4529.u-A).
All rights reserved.
the last decade, the amount of broiler litter generated on poultryfarms has increased steadily and faced environmental andeconomic challenges related to its disposal (Albiach et al., 2001).High organic matter content of chicken manure makes it attractivefor use as a soil organic amendment, especially in sandy soils inwhich organic matter tends to be very low. In these sandy soils,SOM tends to decrease rapidly with inappropriate managementpractices (Garcia-Gil et al., 2000). Thus, application of broiler littermay restore biological activity in soil and improve soil physico-chemical properties.
The importance of SOMas a sink in C sequestrationprocesses hasbeen recognized (Lal and Bruce, 1999; Follett, 2001) and strategiesfor increasing its sequestration in soils have been investigatedincluding conservation tillage, soil amendments with biosolids,organic wastes, and improved crop rotations (Lal, 2004; Smith,2004). Martyniuk and Wagner (1978) reported positive effects oforganic amendments on soil biology in diverse agricultural systems.Swift (2001) linked potential soil C sequestration and extend of Caccumulation in soil to a soil's ability to protect stable organicstructures such as humic substances and mineral-associated

A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550544
organic matter. Because C decomposition in soil is a function ofdifferent factors including soil properties, climatic factors, andorganic amendments, it is important to have better understandingon C mineralization in native soils in order to select managementpractices that help maintain adequate C cycling levels in those soils.
Different soils vary in their response to external carbon addi-tions. Previous research had shown that soil texture for examplehas a profound influence on SOC decomposition. Decomposition ofOC proceeds faster in soils with high sand content than in thosewith high clay content (Khalil et al., 2005; Six et al., 2001). In sandysoils, microorganisms have more access to OC than in heavy-textured soils where microbial access is limited because of sorptionof the OC onto soil minerals. For the past several decades, broilerlitter has been added in large quantities to low organic mattercontent soils in the southeastern United States. However, thedecomposition rates of the added organic amendments are not wellunderstood. An understanding of the mineralization of this OC isa prerequisite for predicting contributions of this OC pools to globalCO2 balance and helping to develop strategies to sequester moreOC into cultivated soils. The objectives of our study were to: 1)compare native OC decomposition in ten soils, 2) assess thedecomposition of two well characterized broiler litter samples inthe selected soils, and 3) establish relationships between poten-tially mineralizable organic C (Co) and soil properties.
2. Materials and methods
2.1. Soil sampling and analysis
Ten surface soil (0e15 cm) samples were collected from variousparts of Alabama and included representative soil series witha wide range of organic C contents such as: Appling, Troup, Cecil,Decatur, Sucarnoochee, Linker, Hartsells, Dothan, Maytag, andColbert soils (Table 1). The soils collected are non-calcareousagricultural soils that to our knowledge have never received broilerlitter. Counties where the soils were collected were shown inSissoko (2009, M. Sc. Thesis, unpublished observations). Thesamples were air-dried and ground to pass through a 2-mm sieve. Asubsample was finely ground to pass through a 100-mesh (<149-mm) sieve for analysis of organic C and total N by Vario EL IIIAutomated Analyzer (CHNS Analyzer, Hanau, Germany). Soil pHwas determined by a combination glass electrode (soil:water ratio1:2.5); inorganic N by steam distillation (Keeny and Nelson, 1982);
Table 1Selected properties of the soils used.
Soil seriesa pH Organiccarbon
Totalnitrogen
Inorganic N Texture
NH4þ (NO2
� þ NO3�) Clay Silt Sand
g kg�1 mg kg�1 g kg�1
Appling 5.95 26.4 3.90 14.3 9.57 75 125 800Troup 6.64 20.1 3.30 2.16 5.07 125 75 800Cecil 5.87 19.0 3.38 14.4 20.8 75 225 700Decatur 5.90 14.3 3.91 8.91 8.73 e 100 900Sucarnoochee 5.91 11.4 2.82 6.87 3.39 75 475 450Linker 6.31 9.71 2.73 3.10 7.02 25 350 625Hartsells 6.06 9.58 2.67 3.91 5.39 125 300 575Dothan 6.48 9.37 2.57 12.0 21.2 75 650 275Maytag 5.95 7.37 2.55 2.55 3.05 125 325 550Colbert 6.05 6.88 2.53 13.1 23.0 e 125 875
a Appling: Fine, kaolinitic thermic kanapludult; Troup: Loamy, siliceous, ThermicGrossarenic Paleudults; Cecil: Fine, kaolinitic thermic, Typic kanapludults; Decatur:Clayey, kaolinitic thermicRhodicPaleudults;Sucarnoochee:montmorillonitic, thermicchromic Epiaqurand; Linker: Fine-loamy, siliceous semiactive thermic Typic Haplu-dults; Hartsells: Fine-loamy, siliceous subactive thermic Typic Hapludults; Dothan:Fine-loamy, siliceous, thermic Plinthic Paleudults; Maytag: Fine montmorillonitic,thermic, oxyaquic Hapluderts; Colbert: Fine, smetitic, thermic Vertic Hapludults.
and particle-size distribution by the pipette method (Kilmer andAlexander, 1949).
2.2. Laboratory incubation studies
A 20-g (oven-dry) soil sample (<2-mm) was mixed with orwithout broiler litter sample 1 or 2 to give 9 g organic C kg�1 soil.The samples were placed in 250-mL French square bottles andincubated for 43 days. Selected properties of the broiler littersamples used are shown in Table 2. The incubation apparatus usedwas similar to that described previously (Cheng and Coleman,1989;Ajwa and Tabatabai, 1994). In brief, air from a laboratory outlet wasconnected to a scrubber consisting of 4 M NaOH (2 L) used toremove CO2 from the flowing air through a tygon tube. Concen-trated H2SO4 was used to remove hydrocarbon contaminants andammonia from the air while a manometer was used to regulate airflow. The French square bottles were connected to test tubes (Tube,culture Kimax-51, Kimble Glass Inc.) containing 25 mL of knownmolarity KOH (0.2 M) used to trap the CO2 evolved. Blanks wereincluded to account for any trace amount of CO2 that might havecome from the scrubber. The air flow to each incubation bottle wasset at about 10 mL min�1 using a pich clamp. The test tubescontaining the trapping KOH solutions were replaced periodicallyand the CO2 trapped was determined potentiometrically bytitrating a 20-mL aliquot against a standard HCl solution (0.2 M),after precipitating carbonate with 6 mL of 0.375 M BaCl2. Resultsare averages of duplicate samples and are expressed on moisturefree-basis. Moisture was determined after 72 h of drying.
2.3. Model description and statistical analysis
To estimate the potentially mineralizable organic pools (Co) andthe first-order rate constant (k), the non-linear regression approachfor N mineralization of Smith et al. (1980) was used.
Cm ¼ Co�1� exp�kt�; where
Cm is the organic C mineralized (mg kg�1) at a specific time (t). TheStatistical Analysis System (SAS) computer language was used tocalculate Co and k (Barr et al., 1976). The decomposition rates (ki) ofthe OC pool was calculated from slopes of the linear segments ofcurves obtained from plotting the natural log of OC remainingagainst time as described by (Gilmour et al., 1977). The ki valueof the resistant fraction was used in calculating the half-life (t1/2)of the most resistant C fraction in the broiler litter samples(t½ ¼ 0.693/ki). The fitting of the mathematical models was doneusing SAS-ProcNLIN (SAS Institute,1990), an iterative method usingMARQUADT (1963) algorithm.
3. Results
3.1. Carbon decomposition in soils
To determine the amount of OC present in the soil samples afteraddition of the broiler litter samples, the soil-broiler litter (<100
Table 2Selected properties of the broiler litter used.a
Broilerlitter ID
pH Organic Total Inorganic N C/N Beddingmaterial
Litter age
C N NH4þ (NO2
� þ NO3�) Month
g kg�1
1 8.4 229 27.5 1.61 1.03 8.32 Pine sawdust 92 8.6 351 46.1 4.39 1.95 7.61 Peanut hulls 9
a From Kokoasse Kpomblekou-A (2006).
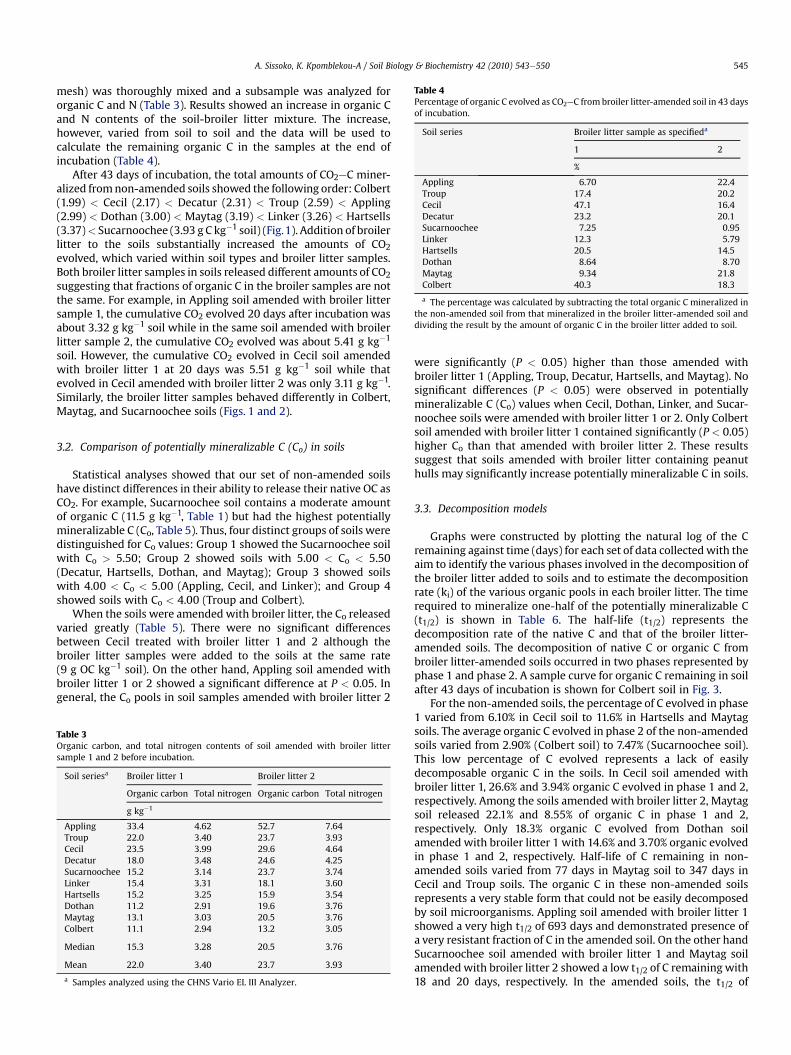
Table 4Percentage of organic C evolved as CO2eC from broiler litter-amended soil in 43 daysof incubation.
Soil series Broiler litter sample as specifieda
1 2
%
Appling 6.70 22.4Troup 17.4 20.2Cecil 47.1 16.4Decatur 23.2 20.1Sucarnoochee 7.25 0.95Linker 12.3 5.79Hartsells 20.5 14.5Dothan 8.64 8.70Maytag 9.34 21.8Colbert 40.3 18.3
a The percentage was calculated by subtracting the total organic C mineralized inthe non-amended soil from that mineralized in the broiler litter-amended soil anddividing the result by the amount of organic C in the broiler litter added to soil.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550 545
mesh) was thoroughly mixed and a subsample was analyzed fororganic C and N (Table 3). Results showed an increase in organic Cand N contents of the soil-broiler litter mixture. The increase,however, varied from soil to soil and the data will be used tocalculate the remaining organic C in the samples at the end ofincubation (Table 4).
After 43 days of incubation, the total amounts of CO2eC miner-alized fromnon-amended soils showed the following order: Colbert(1.99) < Cecil (2.17) < Decatur (2.31) < Troup (2.59) < Appling(2.99) < Dothan (3.00) < Maytag (3.19) < Linker (3.26) < Hartsells(3.37)< Sucarnoochee (3.93 g C kg�1 soil) (Fig.1). Addition of broilerlitter to the soils substantially increased the amounts of CO2evolved, which varied within soil types and broiler litter samples.Both broiler litter samples in soils released different amounts of CO2suggesting that fractions of organic C in the broiler samples are notthe same. For example, in Appling soil amended with broiler littersample 1, the cumulative CO2 evolved 20 days after incubation wasabout 3.32 g kg�1 soil while in the same soil amended with broilerlitter sample 2, the cumulative CO2 evolved was about 5.41 g kg�1
soil. However, the cumulative CO2 evolved in Cecil soil amendedwith broiler litter 1 at 20 days was 5.51 g kg�1 soil while thatevolved in Cecil amended with broiler litter 2 was only 3.11 g kg�1.Similarly, the broiler litter samples behaved differently in Colbert,Maytag, and Sucarnoochee soils (Figs. 1 and 2).
3.2. Comparison of potentially mineralizable C (Co) in soils
Statistical analyses showed that our set of non-amended soilshave distinct differences in their ability to release their native OC asCO2. For example, Sucarnoochee soil contains a moderate amountof organic C (11.5 g kg�1, Table 1) but had the highest potentiallymineralizable C (Co, Table 5). Thus, four distinct groups of soils weredistinguished for Co values: Group 1 showed the Sucarnoochee soilwith Co > 5.50; Group 2 showed soils with 5.00 < Co < 5.50(Decatur, Hartsells, Dothan, and Maytag); Group 3 showed soilswith 4.00 < Co < 5.00 (Appling, Cecil, and Linker); and Group 4showed soils with Co < 4.00 (Troup and Colbert).
When the soils were amended with broiler litter, the Co releasedvaried greatly (Table 5). There were no significant differencesbetween Cecil treated with broiler litter 1 and 2 although thebroiler litter samples were added to the soils at the same rate(9 g OC kg�1 soil). On the other hand, Appling soil amended withbroiler litter 1 or 2 showed a significant difference at P < 0.05. Ingeneral, the Co pools in soil samples amended with broiler litter 2
Table 3Organic carbon, and total nitrogen contents of soil amended with broiler littersample 1 and 2 before incubation.
Soil seriesa Broiler litter 1 Broiler litter 2
Organic carbon Total nitrogen Organic carbon Total nitrogen
g kg�1
Appling 33.4 4.62 52.7 7.64Troup 22.0 3.40 23.7 3.93Cecil 23.5 3.99 29.6 4.64Decatur 18.0 3.48 24.6 4.25Sucarnoochee 15.2 3.14 23.7 3.74Linker 15.4 3.31 18.1 3.60Hartsells 15.2 3.25 15.9 3.54Dothan 11.2 2.91 19.6 3.76Maytag 13.1 3.03 20.5 3.76Colbert 11.1 2.94 13.2 3.05
Median 15.3 3.28 20.5 3.76
Mean 22.0 3.40 23.7 3.93
a Samples analyzed using the CHNS Vario EL III Analyzer.
were significantly (P < 0.05) higher than those amended withbroiler litter 1 (Appling, Troup, Decatur, Hartsells, and Maytag). Nosignificant differences (P < 0.05) were observed in potentiallymineralizable C (Co) values when Cecil, Dothan, Linker, and Sucar-noochee soils were amended with broiler litter 1 or 2. Only Colbertsoil amended with broiler litter 1 contained significantly (P < 0.05)higher Co than that amended with broiler litter 2. These resultssuggest that soils amended with broiler litter containing peanuthulls may significantly increase potentially mineralizable C in soils.
3.3. Decomposition models
Graphs were constructed by plotting the natural log of the Cremaining against time (days) for each set of data collectedwith theaim to identify the various phases involved in the decomposition ofthe broiler litter added to soils and to estimate the decompositionrate (ki) of the various organic pools in each broiler litter. The timerequired to mineralize one-half of the potentially mineralizable C(t1/2) is shown in Table 6. The half-life (t1/2) represents thedecomposition rate of the native C and that of the broiler litter-amended soils. The decomposition of native C or organic C frombroiler litter-amended soils occurred in two phases represented byphase 1 and phase 2. A sample curve for organic C remaining in soilafter 43 days of incubation is shown for Colbert soil in Fig. 3.
For the non-amended soils, the percentage of C evolved in phase1 varied from 6.10% in Cecil soil to 11.6% in Hartsells and Maytagsoils. The average organic C evolved in phase 2 of the non-amendedsoils varied from 2.90% (Colbert soil) to 7.47% (Sucarnoochee soil).This low percentage of C evolved represents a lack of easilydecomposable organic C in the soils. In Cecil soil amended withbroiler litter 1, 26.6% and 3.94% organic C evolved in phase 1 and 2,respectively. Among the soils amended with broiler litter 2, Maytagsoil released 22.1% and 8.55% of organic C in phase 1 and 2,respectively. Only 18.3% organic C evolved from Dothan soilamended with broiler litter 1 with 14.6% and 3.70% organic evolvedin phase 1 and 2, respectively. Half-life of C remaining in non-amended soils varied from 77 days in Maytag soil to 347 days inCecil and Troup soils. The organic C in these non-amended soilsrepresents a very stable form that could not be easily decomposedby soil microorganisms. Appling soil amended with broiler litter 1showed a very high t1/2 of 693 days and demonstrated presence ofa very resistant fraction of C in the amended soil. On the other handSucarnoochee soil amended with broiler litter 1 and Maytag soilamendedwith broiler litter 2 showed a low t1/2 of C remaining with18 and 20 days, respectively. In the amended soils, the t1/2 of
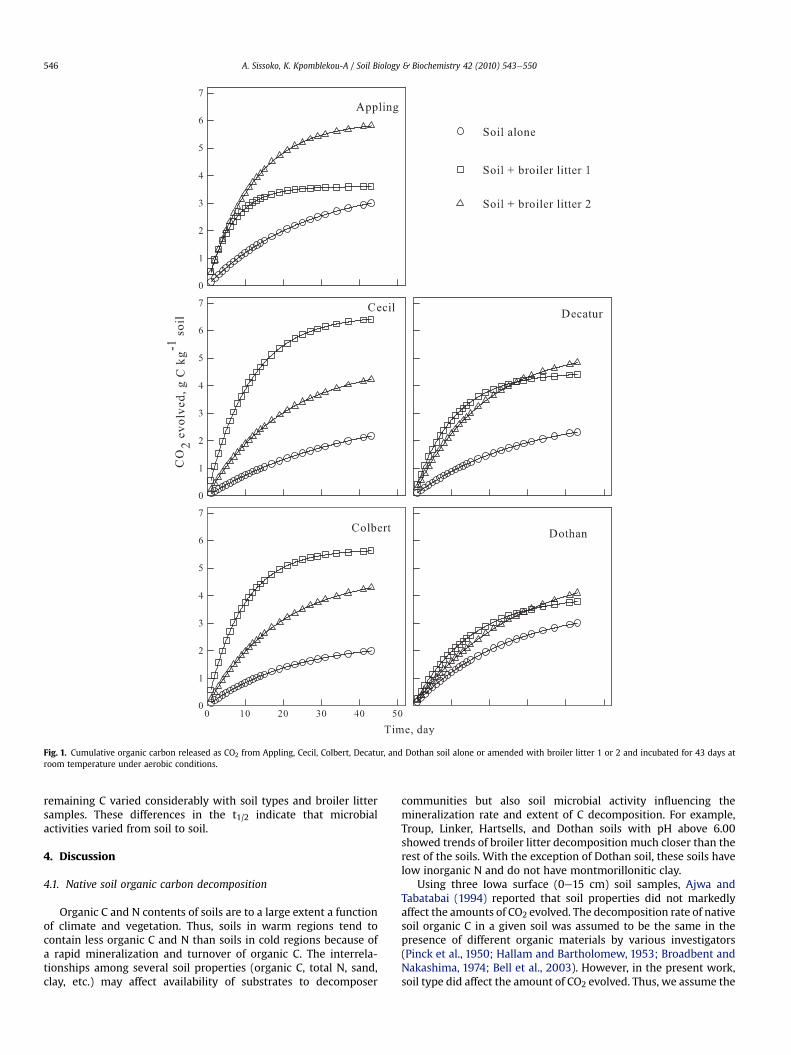
Fig. 1. Cumulative organic carbon released as CO2 from Appling, Cecil, Colbert, Decatur, and Dothan soil alone or amended with broiler litter 1 or 2 and incubated for 43 days atroom temperature under aerobic conditions.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550546
remaining C varied considerably with soil types and broiler littersamples. These differences in the t1/2 indicate that microbialactivities varied from soil to soil.
4. Discussion
4.1. Native soil organic carbon decomposition
Organic C and N contents of soils are to a large extent a functionof climate and vegetation. Thus, soils in warm regions tend tocontain less organic C and N than soils in cold regions because ofa rapid mineralization and turnover of organic C. The interrela-tionships among several soil properties (organic C, total N, sand,clay, etc.) may affect availability of substrates to decomposer
communities but also soil microbial activity influencing themineralization rate and extent of C decomposition. For example,Troup, Linker, Hartsells, and Dothan soils with pH above 6.00showed trends of broiler litter decomposition much closer than therest of the soils. With the exception of Dothan soil, these soils havelow inorganic N and do not have montmorillonitic clay.
Using three Iowa surface (0e15 cm) soil samples, Ajwa andTabatabai (1994) reported that soil properties did not markedlyaffect the amounts of CO2 evolved. The decomposition rate of nativesoil organic C in a given soil was assumed to be the same in thepresence of different organic materials by various investigators(Pinck et al., 1950; Hallam and Bartholomew, 1953; Broadbent andNakashima, 1974; Bell et al., 2003). However, in the present work,soil type did affect the amount of CO2 evolved. Thus, we assume the
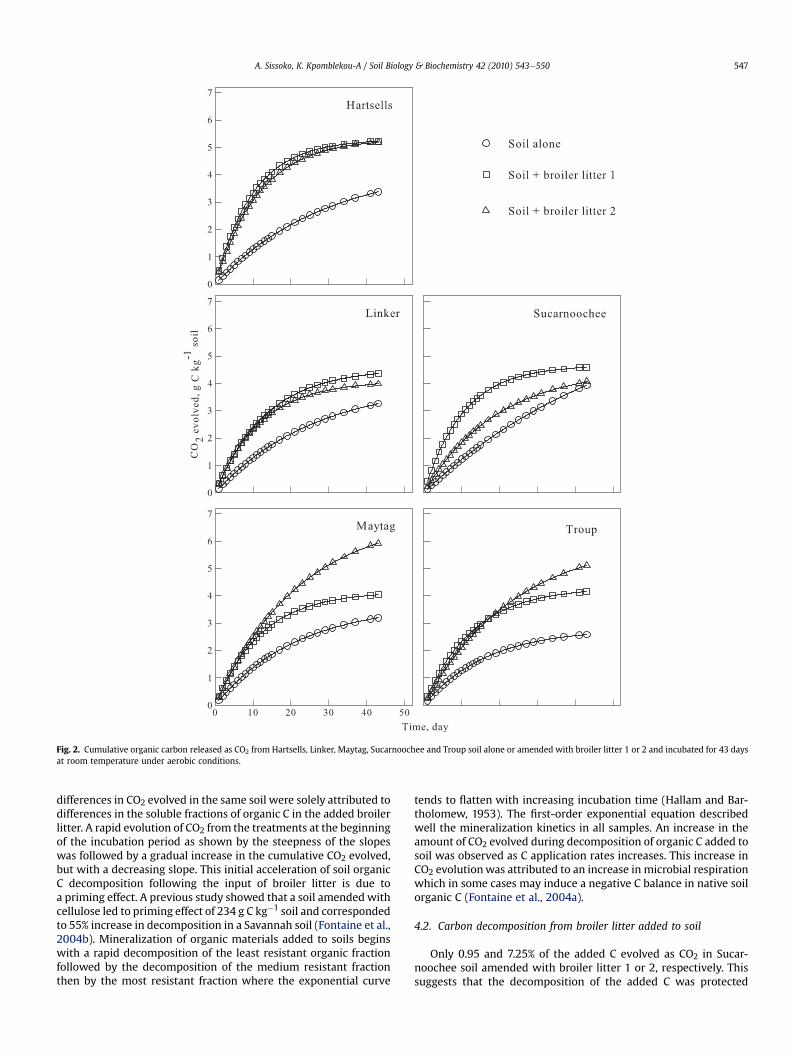
Fig. 2. Cumulative organic carbon released as CO2 from Hartsells, Linker, Maytag, Sucarnoochee and Troup soil alone or amended with broiler litter 1 or 2 and incubated for 43 daysat room temperature under aerobic conditions.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550 547
differences in CO2 evolved in the same soil were solely attributed todifferences in the soluble fractions of organic C in the added broilerlitter. A rapid evolution of CO2 from the treatments at the beginningof the incubation period as shown by the steepness of the slopeswas followed by a gradual increase in the cumulative CO2 evolved,but with a decreasing slope. This initial acceleration of soil organicC decomposition following the input of broiler litter is due toa priming effect. A previous study showed that a soil amended withcellulose led to priming effect of 234 g C kg�1 soil and correspondedto 55% increase in decomposition in a Savannah soil (Fontaine et al.,2004b). Mineralization of organic materials added to soils beginswith a rapid decomposition of the least resistant organic fractionfollowed by the decomposition of the medium resistant fractionthen by the most resistant fraction where the exponential curve
tends to flatten with increasing incubation time (Hallam and Bar-tholomew, 1953). The first-order exponential equation describedwell the mineralization kinetics in all samples. An increase in theamount of CO2 evolved during decomposition of organic C added tosoil was observed as C application rates increases. This increase inCO2 evolutionwas attributed to an increase in microbial respirationwhich in some cases may induce a negative C balance in native soilorganic C (Fontaine et al., 2004a).
4.2. Carbon decomposition from broiler litter added to soil
Only 0.95 and 7.25% of the added C evolved as CO2 in Sucar-noochee soil amended with broiler litter 1 or 2, respectively. Thissuggests that the decomposition of the added C was protected
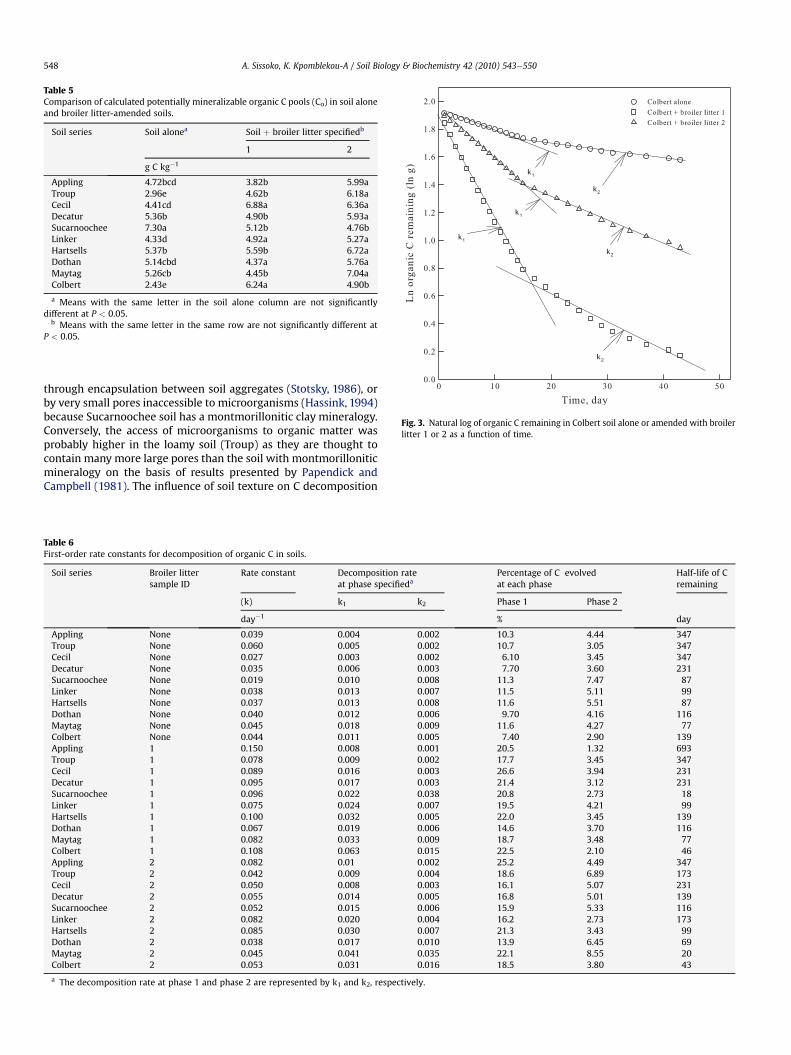
Table 5Comparison of calculated potentially mineralizable organic C pools (Co) in soil aloneand broiler litter-amended soils.
Soil series Soil alonea Soil þ broiler litter specifiedb
1 2
g C kg�1
Appling 4.72bcd 3.82b 5.99aTroup 2.96e 4.62b 6.18aCecil 4.41cd 6.88a 6.36aDecatur 5.36b 4.90b 5.93aSucarnoochee 7.30a 5.12b 4.76bLinker 4.33d 4.92a 5.27aHartsells 5.37b 5.59b 6.72aDothan 5.14cbd 4.37a 5.76aMaytag 5.26cb 4.45b 7.04aColbert 2.43e 6.24a 4.90b
a Means with the same letter in the soil alone column are not significantlydifferent at P < 0.05.
b Means with the same letter in the same row are not significantly different atP < 0.05.
Fig. 3. Natural log of organic C remaining in Colbert soil alone or amended with broilerlitter 1 or 2 as a function of time.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550548
through encapsulation between soil aggregates (Stotsky, 1986), orby very small pores inaccessible to microorganisms (Hassink, 1994)because Sucarnoochee soil has a montmorillonitic clay mineralogy.Conversely, the access of microorganisms to organic matter wasprobably higher in the loamy soil (Troup) as they are thought tocontain many more large pores than the soil with montmorilloniticmineralogy on the basis of results presented by Papendick andCampbell (1981). The influence of soil texture on C decomposition
Table 6First-order rate constants for decomposition of organic C in soils.
Soil series Broiler littersample ID
Rate constant Decomposition rateat phase specifieda
Percentage of C evolvedat each phase
Half-life of Cremaining
(k) k1 k2 Phase 1 Phase 2
day�1 % day
Appling None 0.039 0.004 0.002 10.3 4.44 347Troup None 0.060 0.005 0.002 10.7 3.05 347Cecil None 0.027 0.003 0.002 6.10 3.45 347Decatur None 0.035 0.006 0.003 7.70 3.60 231Sucarnoochee None 0.019 0.010 0.008 11.3 7.47 87Linker None 0.038 0.013 0.007 11.5 5.11 99Hartsells None 0.037 0.013 0.008 11.6 5.51 87Dothan None 0.040 0.012 0.006 9.70 4.16 116Maytag None 0.045 0.018 0.009 11.6 4.27 77Colbert None 0.044 0.011 0.005 7.40 2.90 139Appling 1 0.150 0.008 0.001 20.5 1.32 693Troup 1 0.078 0.009 0.002 17.7 3.45 347Cecil 1 0.089 0.016 0.003 26.6 3.94 231Decatur 1 0.095 0.017 0.003 21.4 3.12 231Sucarnoochee 1 0.096 0.022 0.038 20.8 2.73 18Linker 1 0.075 0.024 0.007 19.5 4.21 99Hartsells 1 0.100 0.032 0.005 22.0 3.45 139Dothan 1 0.067 0.019 0.006 14.6 3.70 116Maytag 1 0.082 0.033 0.009 18.7 3.48 77Colbert 1 0.108 0.063 0.015 22.5 2.10 46Appling 2 0.082 0.01 0.002 25.2 4.49 347Troup 2 0.042 0.009 0.004 18.6 6.89 173Cecil 2 0.050 0.008 0.003 16.1 5.07 231Decatur 2 0.055 0.014 0.005 16.8 5.01 139Sucarnoochee 2 0.052 0.015 0.006 15.9 5.33 116Linker 2 0.082 0.020 0.004 16.2 2.73 173Hartsells 2 0.085 0.030 0.007 21.3 3.43 99Dothan 2 0.038 0.017 0.010 13.9 6.45 69Maytag 2 0.045 0.041 0.035 22.1 8.55 20Colbert 2 0.053 0.031 0.016 18.5 3.80 43
a The decomposition rate at phase 1 and phase 2 are represented by k1 and k2, respectively.
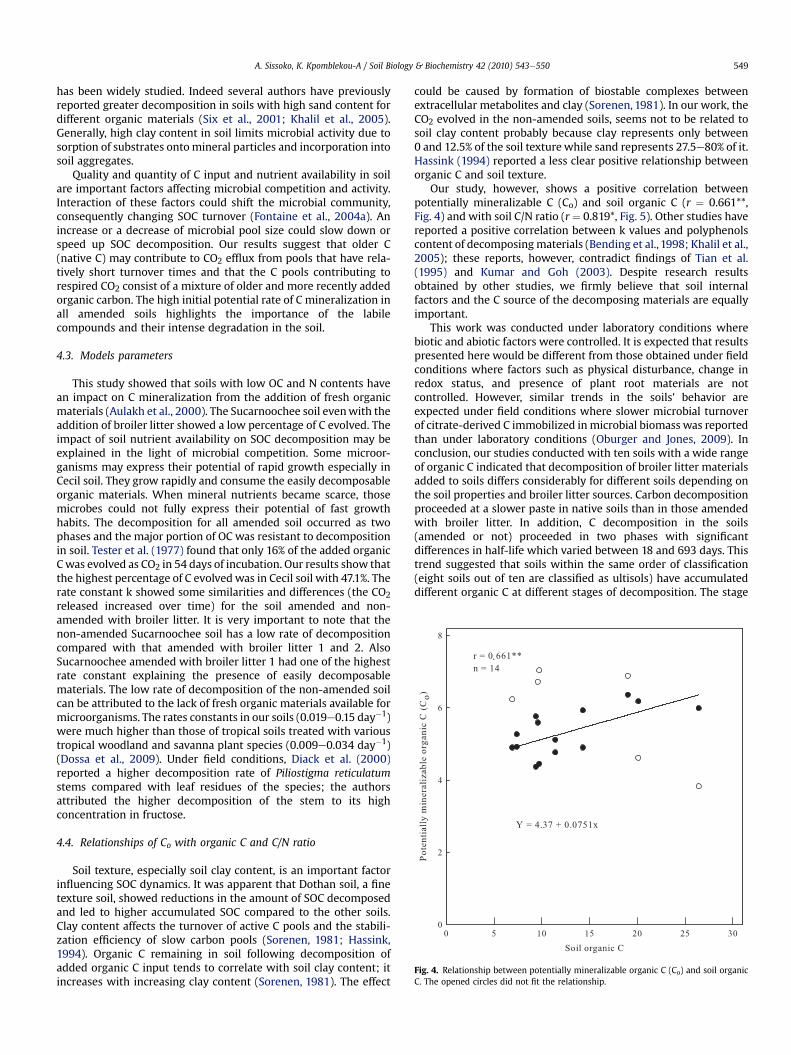
Fig. 4. Relationship between potentially mineralizable organic C (Co) and soil organicC. The opened circles did not fit the relationship.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550 549
has been widely studied. Indeed several authors have previouslyreported greater decomposition in soils with high sand content fordifferent organic materials (Six et al., 2001; Khalil et al., 2005).Generally, high clay content in soil limits microbial activity due tosorption of substrates ontomineral particles and incorporation intosoil aggregates.
Quality and quantity of C input and nutrient availability in soilare important factors affecting microbial competition and activity.Interaction of these factors could shift the microbial community,consequently changing SOC turnover (Fontaine et al., 2004a). Anincrease or a decrease of microbial pool size could slow down orspeed up SOC decomposition. Our results suggest that older C(native C) may contribute to CO2 efflux from pools that have rela-tively short turnover times and that the C pools contributing torespired CO2 consist of a mixture of older and more recently addedorganic carbon. The high initial potential rate of C mineralization inall amended soils highlights the importance of the labilecompounds and their intense degradation in the soil.
4.3. Models parameters
This study showed that soils with low OC and N contents havean impact on C mineralization from the addition of fresh organicmaterials (Aulakh et al., 2000). The Sucarnoochee soil evenwith theaddition of broiler litter showed a low percentage of C evolved. Theimpact of soil nutrient availability on SOC decomposition may beexplained in the light of microbial competition. Some microor-ganisms may express their potential of rapid growth especially inCecil soil. They grow rapidly and consume the easily decomposableorganic materials. When mineral nutrients became scarce, thosemicrobes could not fully express their potential of fast growthhabits. The decomposition for all amended soil occurred as twophases and the major portion of OC was resistant to decompositionin soil. Tester et al. (1977) found that only 16% of the added organicCwas evolved as CO2 in 54 days of incubation. Our results show thatthe highest percentage of C evolvedwas in Cecil soil with 47.1%. Therate constant k showed some similarities and differences (the CO2released increased over time) for the soil amended and non-amended with broiler litter. It is very important to note that thenon-amended Sucarnoochee soil has a low rate of decompositioncompared with that amended with broiler litter 1 and 2. AlsoSucarnoochee amended with broiler litter 1 had one of the highestrate constant explaining the presence of easily decomposablematerials. The low rate of decomposition of the non-amended soilcan be attributed to the lack of fresh organic materials available formicroorganisms. The rates constants in our soils (0.019e0.15 day�1)were much higher than those of tropical soils treated with varioustropical woodland and savanna plant species (0.009e0.034 day�1)(Dossa et al., 2009). Under field conditions, Diack et al. (2000)reported a higher decomposition rate of Piliostigma reticulatumstems compared with leaf residues of the species; the authorsattributed the higher decomposition of the stem to its highconcentration in fructose.
4.4. Relationships of Co with organic C and C/N ratio
Soil texture, especially soil clay content, is an important factorinfluencing SOC dynamics. It was apparent that Dothan soil, a finetexture soil, showed reductions in the amount of SOC decomposedand led to higher accumulated SOC compared to the other soils.Clay content affects the turnover of active C pools and the stabili-zation efficiency of slow carbon pools (Sorenen, 1981; Hassink,1994). Organic C remaining in soil following decomposition ofadded organic C input tends to correlate with soil clay content; itincreases with increasing clay content (Sorenen, 1981). The effect
could be caused by formation of biostable complexes betweenextracellular metabolites and clay (Sorenen, 1981). In our work, theCO2 evolved in the non-amended soils, seems not to be related tosoil clay content probably because clay represents only between0 and 12.5% of the soil texture while sand represents 27.5e80% of it.Hassink (1994) reported a less clear positive relationship betweenorganic C and soil texture.
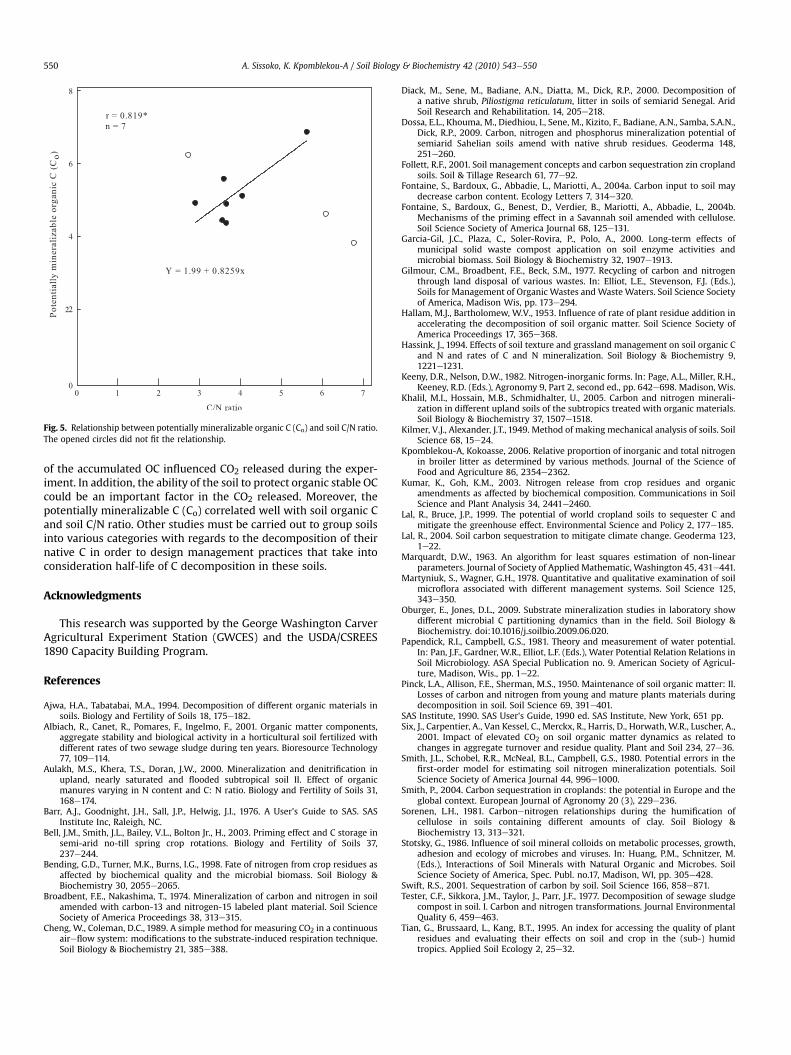
Our study, however, shows a positive correlation betweenpotentially mineralizable C (Co) and soil organic C (r ¼ 0.661**,Fig. 4) and with soil C/N ratio (r ¼ 0.819*, Fig. 5). Other studies havereported a positive correlation between k values and polyphenolscontent of decomposingmaterials (Bending et al., 1998; Khalil et al.,2005); these reports, however, contradict findings of Tian et al.(1995) and Kumar and Goh (2003). Despite research resultsobtained by other studies, we firmly believe that soil internalfactors and the C source of the decomposing materials are equallyimportant.
This work was conducted under laboratory conditions wherebiotic and abiotic factors were controlled. It is expected that resultspresented here would be different from those obtained under fieldconditions where factors such as physical disturbance, change inredox status, and presence of plant root materials are notcontrolled. However, similar trends in the soils' behavior areexpected under field conditions where slower microbial turnoverof citrate-derived C immobilized inmicrobial biomass was reportedthan under laboratory conditions (Oburger and Jones, 2009). Inconclusion, our studies conducted with ten soils with a wide rangeof organic C indicated that decomposition of broiler litter materialsadded to soils differs considerably for different soils depending onthe soil properties and broiler litter sources. Carbon decompositionproceeded at a slower paste in native soils than in those amendedwith broiler litter. In addition, C decomposition in the soils(amended or not) proceeded in two phases with significantdifferences in half-life which varied between 18 and 693 days. Thistrend suggested that soils within the same order of classification(eight soils out of ten are classified as ultisols) have accumulateddifferent organic C at different stages of decomposition. The stage
2
Fig. 5. Relationship between potentially mineralizable organic C (Co) and soil C/N ratio.The opened circles did not fit the relationship.
A. Sissoko, K. Kpomblekou-A / Soil Biology & Biochemistry 42 (2010) 543e550550
of the accumulated OC influenced CO2 released during the exper-iment. In addition, the ability of the soil to protect organic stable OCcould be an important factor in the CO2 released. Moreover, thepotentially mineralizable C (Co) correlated well with soil organic Cand soil C/N ratio. Other studies must be carried out to group soilsinto various categories with regards to the decomposition of theirnative C in order to design management practices that take intoconsideration half-life of C decomposition in these soils.
Acknowledgments
This research was supported by the George Washington CarverAgricultural Experiment Station (GWCES) and the USDA/CSREES1890 Capacity Building Program.
References
Ajwa, H.A., Tabatabai, M.A., 1994. Decomposition of different organic materials insoils. Biology and Fertility of Soils 18, 175e182.
Albiach, R., Canet, R., Pomares, F., Ingelmo, F., 2001. Organic matter components,aggregate stability and biological activity in a horticultural soil fertilized withdifferent rates of two sewage sludge during ten years. Bioresource Technology77, 109e114.
Aulakh, M.S., Khera, T.S., Doran, J.W., 2000. Mineralization and denitrification inupland, nearly saturated and flooded subtropical soil II. Effect of organicmanures varying in N content and C: N ratio. Biology and Fertility of Soils 31,168e174.
Barr, A.J., Goodnight, J.H., Sall, J.P., Helwig, J.I., 1976. A User's Guide to SAS. SASInstitute Inc, Raleigh, NC.
Bell, J.M., Smith, J.L., Bailey, V.L., Bolton Jr., H., 2003. Priming effect and C storage insemi-arid no-till spring crop rotations. Biology and Fertility of Soils 37,237e244.
Bending, G.D., Turner, M.K., Burns, I.G., 1998. Fate of nitrogen from crop residues asaffected by biochemical quality and the microbial biomass. Soil Biology &Biochemistry 30, 2055e2065.
Broadbent, F.E., Nakashima, T., 1974. Mineralization of carbon and nitrogen in soilamended with carbon-13 and nitrogen-15 labeled plant material. Soil ScienceSociety of America Proceedings 38, 313e315.
Cheng, W., Coleman, D.C., 1989. A simple method for measuring CO2 in a continuousaireflow system: modifications to the substrate-induced respiration technique.Soil Biology & Biochemistry 21, 385e388.
Diack, M., Sene, M., Badiane, A.N., Diatta, M., Dick, R.P., 2000. Decomposition ofa native shrub, Piliostigma reticulatum, litter in soils of semiarid Senegal. AridSoil Research and Rehabilitation. 14, 205e218.
Dossa, E.L., Khouma, M., Diedhiou, I., Sene, M., Kizito, F., Badiane, A.N., Samba, S.A.N.,Dick, R.P., 2009. Carbon, nitrogen and phosphorus mineralization potential ofsemiarid Sahelian soils amend with native shrub residues. Geoderma 148,251e260.
Follett, R.F., 2001. Soil management concepts and carbon sequestration zin croplandsoils. Soil & Tillage Research 61, 77e92.
Fontaine, S., Bardoux, G., Abbadie, L., Mariotti, A., 2004a. Carbon input to soil maydecrease carbon content. Ecology Letters 7, 314e320.
Fontaine, S., Bardoux, G., Benest, D., Verdier, B., Mariotti, A., Abbadie, L., 2004b.Mechanisms of the priming effect in a Savannah soil amended with cellulose.Soil Science Society of America Journal 68, 125e131.
Garcia-Gil, J.C., Plaza, C., Soler-Rovira, P., Polo, A., 2000. Long-term effects ofmunicipal solid waste compost application on soil enzyme activities andmicrobial biomass. Soil Biology & Biochemistry 32, 1907e1913.
Gilmour, C.M., Broadbent, F.E., Beck, S.M., 1977. Recycling of carbon and nitrogenthrough land disposal of various wastes. In: Elliot, L.E., Stevenson, F.J. (Eds.),Soils for Management of Organic Wastes and Waste Waters. Soil Science Societyof America, Madison Wis, pp. 173e294.
Hallam, M.J., Bartholomew, W.V., 1953. Influence of rate of plant residue addition inaccelerating the decomposition of soil organic matter. Soil Science Society ofAmerica Proceedings 17, 365e368.
Hassink, J., 1994. Effects of soil texture and grassland management on soil organic Cand N and rates of C and N mineralization. Soil Biology & Biochemistry 9,1221e1231.
Keeny, D.R., Nelson, D.W., 1982. Nitrogen-inorganic forms. In: Page, A.L., Miller, R.H.,Keeney, R.D. (Eds.), Agronomy 9, Part 2, second ed., pp. 642e698. Madison, Wis.
Khalil, M.I., Hossain, M.B., Schmidhalter, U., 2005. Carbon and nitrogen minerali-zation in different upland soils of the subtropics treated with organic materials.Soil Biology & Biochemistry 37, 1507e1518.
Kilmer, V.J., Alexander, J.T., 1949. Method of making mechanical analysis of soils. SoilScience 68, 15e24.
Kpomblekou-A, Kokoasse, 2006. Relative proportion of inorganic and total nitrogenin broiler litter as determined by various methods. Journal of the Science ofFood and Agriculture 86, 2354e2362.
Kumar, K., Goh, K.M., 2003. Nitrogen release from crop residues and organicamendments as affected by biochemical composition. Communications in SoilScience and Plant Analysis 34, 2441e2460.
Lal, R., Bruce, J.P., 1999. The potential of world cropland soils to sequester C andmitigate the greenhouse effect. Environmental Science and Policy 2, 177e185.
Lal, R., 2004. Soil carbon sequestration to mitigate climate change. Geoderma 123,1e22.
Marquardt, D.W., 1963. An algorithm for least squares estimation of non-linearparameters. Journal of Society of Applied Mathematic, Washington 45, 431e441.
Martyniuk, S., Wagner, G.H., 1978. Quantitative and qualitative examination of soilmicroflora associated with different management systems. Soil Science 125,343e350.
Oburger, E., Jones, D.L., 2009. Substrate mineralization studies in laboratory showdifferent microbial C partitioning dynamics than in the field. Soil Biology &Biochemistry. doi:10.1016/j.soilbio.2009.06.020.
Papendick, R.I., Campbell, G.S., 1981. Theory and measurement of water potential.In: Pan, J.F., Gardner, W.R., Elliot, L.F. (Eds.), Water Potential Relation Relations inSoil Microbiology. ASA Special Publication no. 9. American Society of Agricul-ture, Madison, Wis., pp. 1e22.
Pinck, L.A., Allison, F.E., Sherman, M.S., 1950. Maintenance of soil organic matter: II.Losses of carbon and nitrogen from young and mature plants materials duringdecomposition in soil. Soil Science 69, 391e401.
SAS Institute, 1990. SAS User's Guide, 1990 ed. SAS Institute, New York, 651 pp.Six, J., Carpentier, A., Van Kessel, C., Merckx, R., Harris, D., Horwath, W.R., Luscher, A.,
2001. Impact of elevated CO2 on soil organic matter dynamics as related tochanges in aggregate turnover and residue quality. Plant and Soil 234, 27e36.
Smith, J.L., Schobel, R.R., McNeal, B.L., Campbell, G.S., 1980. Potential errors in thefirst-order model for estimating soil nitrogen mineralization potentials. SoilScience Society of America Journal 44, 996e1000.
Smith, P., 2004. Carbon sequestration in croplands: the potential in Europe and theglobal context. European Journal of Agronomy 20 (3), 229e236.
Sorenen, L.H., 1981. Carbonenitrogen relationships during the humification ofcellulose in soils containing different amounts of clay. Soil Biology &Biochemistry 13, 313e321.
Stotsky, G., 1986. Influence of soil mineral colloids on metabolic processes, growth,adhesion and ecology of microbes and viruses. In: Huang, P.M., Schnitzer, M.(Eds.), Interactions of Soil Minerals with Natural Organic and Microbes. SoilScience Society of America, Spec. Publ. no.17, Madison, WI, pp. 305e428.
Swift, R.S., 2001. Sequestration of carbon by soil. Soil Science 166, 858e871.Tester, C.F., Sikkora, J.M., Taylor, J., Parr, J.F., 1977. Decomposition of sewage sludge
compost in soil. I. Carbon and nitrogen transformations. Journal EnvironmentalQuality 6, 459e463.
Tian, G., Brussaard, L., Kang, B.T., 1995. An index for accessing the quality of plantresidues and evaluating their effects on soil and crop in the (sub-) humidtropics. Applied Soil Ecology 2, 25e32.





























