Embed Size (px)
Citation preview

CARBOHYDRATES
1. In digestion of carbohydrates from the food:
a) participate enzymes of stomach mucosa (F)
b) participate enzyme formed in pancreas (T)
c) glycosidic bond is hydrolyzed (T)
d) starch is broken by amylase into free glucose (F)
2. Which statement about digestion of carbohydrates is correct:
a) starch is broken by amylase into disaccharide maltose (T)
b) water is required (T)
c) amylase is the most important enzyme (T)
d) amylase hydrolazes all carbohydrates form food (F)
3. Main source of glucose for newborn:
a) is compound present in fruits (F)
b) is compound with -1,4-glycosidic bond (T)
c) is lactate (F)
d) is compound which is hydrolyzed to glucose and fructose (F)
4. Product of the action of amylase into starch is:
a) compound which belongs to polysaccharides (F)
b) disaccharide maltose (T)
c) disaccharide glucose (F)
d) compound which is directly absorbed into blood (F)
5. Pyruvate:
a) is final product of complete oxidation of glucose in red blood cells (F)
b) is formed as final product of glucose oxidation in brain (F)
c) is formed as final product of glycolysis in aerobic conditions (T)
d) is compound synthesis of which proceeds in cytosol (T)
6. Substrate for alcoholic fermentation is:
a) monosaccaride glucose (T)
b) pyruvate (F)
c) compound which is oxidized by glycolysis (T)
d) compound which is oxidized by glycogenolysis (F)
7. Activation of glucose:
a) requires ATP as source of energy (T)
b) is conversion of glucose into glucose-6-phosphate (T)
c) is required for connection of glucose with glycolysis, glycogen synthesis and pentose phosphate
pathway (T)
d) in liver is catalyzed mainly by hexokinase with high affinity to glucose (F)
8. During activation of glucose:
a) glucose-1-phosphate is formed (F)
b) ALP i required (T)
c) in extrahepatic tissues glucokinase with high specificity is required (F)
d) in liver glucokinase with low affinity is required (T)
9. Glucokinase is enzyme which:
a) is responsible for phosphorylation of glucose (T)
b) is present in all tissues of human body (F)
c) can catalyze phosphorylation of all hexoses (F)
d) is not inhibited by product of reaction – glucose-6-phosphate (T)

10. Glucokinase:
a) is enzyme typical for liver tissue (T)
b) uses glucose as substrate (T)
c) is enzyme with high Km for glucose (T)
d) catalyzes phosphorylation of glucose when its concentration is low – during starvation (F)
11. When glucose is activated before entering metabolic processes:
a) ATP is used as source of energy (T)
b) depending on type of tissue either hexokinase or gucokinase is used (T)
c) enzyme present in cytosol is required (T)
d) glucose-6-phosphate is formed (T)
12. Glucose-6-phosphate is compound which:
a) is product of glucose activation (T)
b) is formed in reversible reaction (F)
c) in EHT is formed by high-affinity hexokinase (T)
d) in gluconeogenesis is changed by reversible reaction of glycolysisi into free glucose (F)
13. Glucokinase and hexokinase are enzymes which:
a) are responsible for glucose transport to the cells (F)
b) are responsible for activation (T)
c) both are present in all tissues of human body (F)
d) lead to synthesis of glucose-6-phosphate (T)
14. Enzyme hexokinase:
a) catalyzes one of four irreersible reactions of glycolysis (F)
b) as substrate uses glucose only (F)
c) in low glucose concentration is not active because of low Km for glucose (F)
d) is resonsible for glycogen degradation (F)
15. Glucose-6-phosphate enters metabolic processes:
a) which are located in mitochondria and cytosol (F)
b) as is for example glycolysis located in cytosol (T)
c) as is for example pentose phosphate pathway located in mitochondria (F)
d) as is for example synthesis of glycogen located in lysosomes (F)
16. Processes which utilize glucose-6-phosphate:
a) are processes function of which is synthesis of ATP only (F)
b) are processes which proceed only in aerobic conditions (T)
c) is for example glycolysis which allows synthesis of ATP also in anaerobic conditions (F)
d) is for example synthesis of glycogen (T)
17. Reaction of glucose with ATP:
a) is reaction of glucose activation before entering glycolysis (T)
b) is reaction of glucose activation for synthesis of glycogen (T)
c) in liver is catalyzed by glucokinase (T)
d) is reversible reaction (F)
18. Conversion of glucose-6-phosphate to fructose-6-phosphate:
a) is catalyzed by phosphoglucomutase (F)
b) is required for degradation of glucose by glycolysis (T)
c) is reversible reaction (T)
d) is important regulatory enzyme of glycolysis (F)
19. Change of glucose-6-phosphate to fructose-6-phosphate:
a) is catalyzed by phosphohexoisomerase (T)
b) is irreversible reaction (F)

c) is reaction important for glycogen synthesis (F)
d) is reaction of glycolysis (T)
20. Phosphofructokinase catalyzes reaction which:
a) is reversible (F)
b) leads to synthesis of fructose-6-phosphate (F)
c) is activated by ATP (F)
d) is activated during starvation (F)
21. Reaction catalyzed by phosphofructokinase:
a) leads to synthesis of fructose-1,6-bisphosphate (T)
b) is key regulatory step of glycolysis (T)
c) is irreversible (T)
d) is reaction activated by fructose-2,6-bisphosphate (T)
22. Key regulatory reaction of glycolysis is:
a) reaction of conversion of fructose-6-phosphate to fructose-1,6-bisphosphate (T)
b) reaction of fructose-6-phosphate synthesis (F)
c) reaction which is inhibited by ATP, NADH+H+ (T)
d) reaction which is activated by phosphorylation of the enzyme (F)
23. Which of following compounds influence activity of phosphofructokinase I:
a) ATP as inhibitor (T)
b) ADP, AMP, NAD as inhibitors (F)
c) citrate as activator (F)
d) fructose-2,6-bisphosphate which allows its reaction despite high concentration of ATP (T)
24. Inhibition of phosphofructokinase by ATP:
a) belongs to metabolic regulation of glycolysis (T)
b) can be removed by ADP (F)
c) can be removed by fructose-2,6-bisphosphate (T)
d) can be removad by compound which is foprmed in presence of glucagon (F)
25. Glyceraldehyde-3-phosphate and dihydroxyacetonephosphate are compounds which:
a) are formed in cytosol (T)
b) are intermediates of pentose phosphate pathway (F)
c) are formed form fuctose-6-phosphate (F)
d) can be interconverted by tiosephosphate isomerase (T)
26. Cleavage of fructose-1,6-bisphosphate in glycolysis:
a) is catalyzed by enzyme aldolase (T)
b) is hydrolytic reaction (F)
c) leads to synthesis of two phosphorylated aldotrioses (F)
d) leads to synthesis of glyceraldehyde phosphate and dihydroxyacetonephosphate (T)
27. Reaction of fructose-1,6-bisphosphate conversion in glycolysis:
a) is catalyzed by enzyme which belongs to lyases (T)
b) is catalyzed by phosphofructokinase (F)
c) leads to synthesis of glyceraldehyde-3-phosphate which is further metabolized in glycolysis (T)
d) is inhibited by ATP (F)
28. Triosephosphate isomerase catalyzes reaction:
a) in which water is required (F)
b) in which phosphorylated aldotriose is changed to phosphorylated ketotriose (T)
c) which is reversible (T)
d) which is required also for connection of alcoholic component of triacylglycerols to
gluconeogenesis (T)

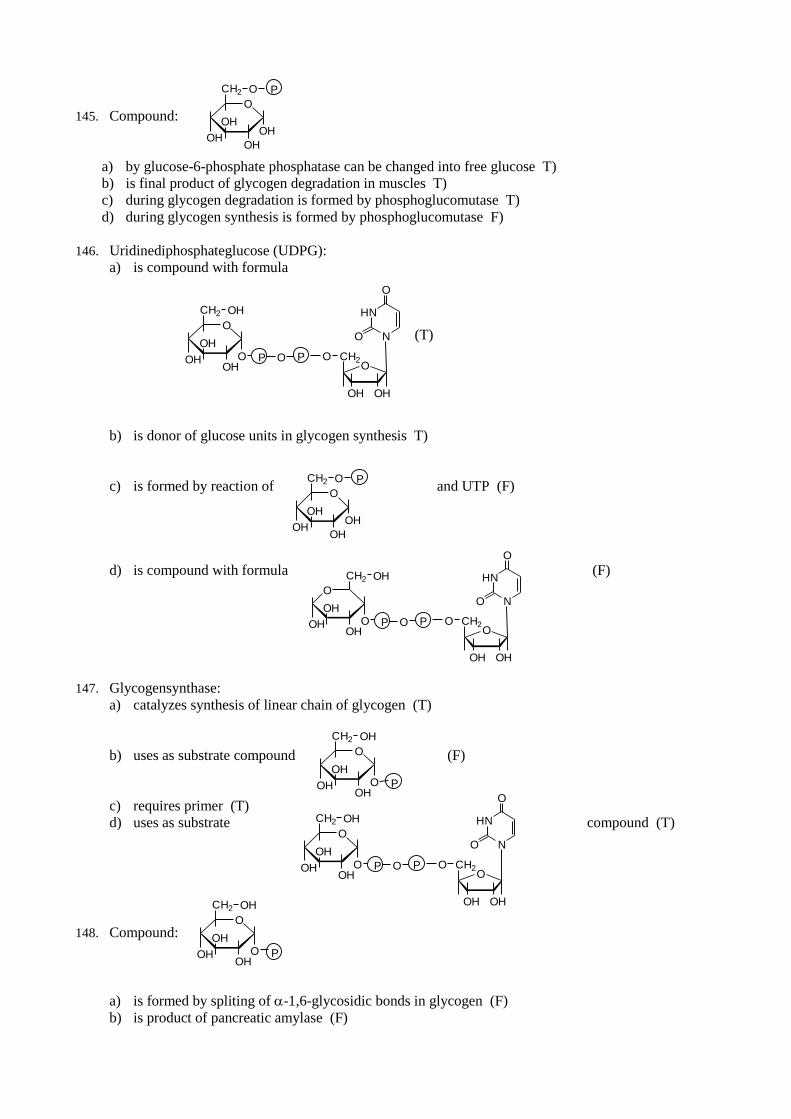
29. Intermediate of glycolysis which can be used for synthesis of triacylglycerols:
a) is dihydroxyacetonephosphate (T)
b) is pyruvate (F)
c) for synthesis of triacylglcerols is changed by its reduction (T)
d) is changed in reaction catalyzed by glycerolphosphate dehydrogenase (T)
30. Change of dihydroxyacetonephosphate to glycerolphosphate:
a) is reversible reaction (T)
b) is catalyzed by glycerophosphatedehydrogenase (T)
c) is not important for gluconeogenesis (F)
d) is important also for transport of reducing equivalents from cytosol into mitochondria – shuttles (T)
31. Dihydroxyacetonephosphate is changed to glycerolphosphate:
a) by reaction which is irreversible (F)
b) by oxidation catalyzed by glycerolphosphate dehydrogenase (F)
c) by reaction which is important for glycolysis (F)
d) by reaction which uses FADH2 as coenzyme (F)
32. Glycerolphosphate dehydrogenase is the enzyme which:
a) catylazes reversible reaction (T)
b) is important for gluconeogenesis (T)
c) is important for synthesis of fatty acids (F)
d) in mitochondria uses FAD as coenzyme (T)
33. Reaction catalyzed by glycerolphosphate dehydrogenase:
a) is irreversible (F)
b) in mitochondria uses NAD as coenzyme (F)
c) is important for synthesis of triacylglycerols (T)
d) does not proceed in cytosol (F)
34. During oxidation of glyceraldehyde-3-phosphate in glycolysis:
a) enzyme glycerolphosphate dehydrogenase is used (F)
b) intermediate of reaction contains thioester energy rich bond (F)
c) NADPH2 is formed (F)
d) coenzyme of reaction is FAD (F)
35. Product of reaction catalyzed by glyceraldehyde-3-phosphate dehydrogenase:
a) is compound with two energy rich bonds (F)
b) is compound with thioester energy rich bond (F)
c) is compound which is used for synthesis of ATP by substrate level phosphorylation (T)
d) is compound which is changed by glycerolkinase (F)
36. During oxidation of one glyceraldehyde-3-phosphate in aerobic conditions:
a) the same amount of ATP as in anaerobic conditions is formed (F)
b) 3 ATP by oxidative phosphorylation are formed (T)
c) 1 ATP by substrate level phosphorylation is formed (T)
d) totally 6 ATP are formed (F)
37. When glyceraldehyde-3-phosphate is oxidized in glycolysis:
a) ATP by oxidative phosphorylation is formd only in aerobic conditions (T)
b) in aerobic conditions by 1 more ATP is formed than in anaerobic conditions (F)
c) reaction allow synthesis of 1 ATP in both aerobic and anaerobic conditions (T)
d) in anaerobic conditions ATP is formed only by oxiodative phosphorylation
38. Reaction of conversion glyceraldehyde-3-phosphate –– 1,3-bis-phosphoglycerate ––
3-phosphoglycerate allows synthesis of:

a) the same amount of ATP is formed in both aerobic and anaerobic conditions (F)
b) 1 ATP by substrate level phosphorylation only in anaerobic conditions (F)
c) ATP only in aerobic conditions (F)
d) 3 ATP by oxidative phosphorylation in aerobic conditions (T)
39. Different asmounts of ATP by oxidation of glyceraldehyde-3-phosphate in aerobic and anaerobic
conditions are formed because:
a) in anaerobic conditions does not proceed terminal oxidation (T)
b) in anaerobic conditions does not proceed Krebs cycle (F)
c) in aerobic conditions ATP can be forme also by oxidative phosphorylation (T)
d) in anaerobic conditions reduced coenzyme transfers hydrogens to pyruvate and not to terminal
oxidation (T)
40. Glyceraldehyde-3-phosphate dehydrogenase is responsible for synthesis of:
a) 3-phosphoglycerate (F)
b) compound with acylphosphate energy rich bond (T)
c) compound which is changed by phosphoglycerate kinase (T)
d) compound which is used for synthesis of ATP by substrate level phosphorylation in all cells of
human body (T)
41. Conversion of 1,3-bisphosphoglycerate in glycolysis:
a) is catalyzed by glycerolkinase (F)
b) requires ATP as substrate (F)
c) is reaction of phosphorylation at the substrate level (T)
d) allows synthesis of 2 ATP per one glucose (T)
42. Substrate for phosphoglyceratekinase is compound which:
a) is 1,3-bisphosphoglycerate (T)
b) is formed by glycerolphosphate dehydrogenase (F)
c) contains enolphosphate energy rich bond (F)
d) allows synthesis of ATP by substrate level phosphorylation (T)
43. Phosphoglycerate kinase in glycolysis catalyzes conversion of:
a) 3-phosphoglycerate (F)
b) 1,3-bisphosphoglycerate (T)
c) compound which is product of glyceraldehyde-3-phosphate dehydrogenase (T)
d) compound which contains phosphoanhydride energy rich bond (F)
44. Phosphoglycerate kinase:
a) catalyzes reversible reaction (T)
b) catalyzes synthesis of 1,3-bisphosphoglycerate in glycolysis (F)
c) catalyzes synthesis of 1,3-bisphosphoglycerate in gluconeogenesis (T)
d) uses energy of acylphosphate energy rich bond for synthesis of ATP by substrate level
phosphorylation (T)
45. Phosphoglycerate mutase catalyzes::
a) reversible reaction (T)
b) conversion of 1,3-bisphosphoglycerate to 3-phosphoglycerate (F)
c) synthesis of 2-phosphoglycerate in glycolysis (T)
d) conversion of 2-phosphoglycerate in gluconeogenesis (T)
46. 1,3-bisphosphoglycerate:
a) is formed by glyceraldehyde-3-phosphate dehydrogenase in glycolysis (T)
b) is formd by irreversible reaction (F)
c) contains phosphate bound by acylphosphate energy rich bond to C1 (T)
d) contains phosphate boud by phosphoester bond to C3 (T)

47. 2-phosphoglycerate in glycolysis is changed:
a) by phosphoglycerate kinase (F)
b) by reversible reaction (T)
c) to phosphoenolpyruvate (T)
d) to compound with acylphosphate energy rich bond (F)
48. Conversion of 2-phosphoglycerate in glycolysis:
a) is catalyzed by enzyme enolase (T)
b) is irreversible reaction (F)
c) requires ATP (F)
d) leds to synthesis of compound which can be used for synthesis of ATP by substrate level
phosphorylation (T)
49. Compound formed by the action of enolase in glycolysis:
a) is enolpyruvate (F)
b) is substrate for pyruvate kinase (T)
c) contains enolphosphate energy rich bond (T)
d) is used for synthesis of ATP by substrate level phosphorylation only in anaerobic conditions (F)
50. Enolase is enzyme which:
a) catalyzes reversible reaction (T)
b) uses 2-phosphoglycerate as substrate (T)
c) leads to synthesis of phosphoenolpyruvate (T)
d) produces phosphoenolpyruvate only in anaerobic conditions (F)
51. Compound with enolphosphate energy rich bond:
a) is 1,3-bisphosphoglycerate (F)
b) in glycolysis is formed by pyruvate kinase (F)
c) in glycolysis is formed by reversible reaction (T)
d) is the only compound for synthesis of ATP by substrate level phosphorylation (F)
52. Enolphosphate energy rich bond contains:
a) compound which is product of enolase (T)
b) phosphoenolpyruvate (T)
c) compound conversion of which requires ATP (F)
d) compound which in gluconeogenesis is formed by pyruvate kinase (F)
53. Pyruvate kinase:
a) catalyzes irrevesible reaction (T)
b) catalyzes synthesis of phosphoenolpyruvate (F)
c) catalyzes synthesis of ATP in aerobic and anaerobic conditions (T)
d) is active in dephosphorylated form (T)
54. In regulation of pyruvate kinase activity:
a) ATP acts as inhibitor (T)
b) fructose-1,6-bisphosphate is activator (T)
c) glucagon inhibits the enzyme by phosphorylation (T)
d) insulin couses activation of the enzyme by its phosphorylation (F)
55. Further conversion of phosphoenolpyruvate in glycolysis:
a) is catalyzed by pyruvate kinase (T)
b) utilizes energy of enolphosphate bond for synthesis of ATP (T)
c) is inhibited by high concentration of ATP (T)
d) is activated during starvation by hormone glucagon (F)
56. In conversion of phosphoenolpyruvate in glycolysis:
a) enzyme enolase is used (F)

COOH
C OH
CH3
H
COOH
C OH
CH3
H
b) ATP is required as substrate (T)
c) GTP is formed (F)
d) enzyme which is active in dephosphorylated form is used (T)
57. Fructose-1,6-bisphosphate influences enzymes of glycolysis that:
a) inhibits phosphofructokinase I (F)
b) activates phosphofructokinase II (F)
c) activates reaction of phosphoenolpyruvate synthesis (F)
d) activates pyruvate kinase (T)
58. Pyruvate in glycolysis is synthesized by:
a) reversible reaction (F)
b) reaction which allows synthesis of ATP in anaerobic conditions (T)
c) reaction which is important also for gluconeogenesis (F)
d) reaction which is active in presence of insulin which causes dephosphorylation of the enzyme
responsible for the reaction (T)
59. In aerobic conditions in glycolysis:
a) 2 mol lactate are formed per 1 mol of glucose (F)
b) 2 mol of acetyl-CoA are formed per 1 mol of glucose (F)
c) 2 mol of CO2 are formed per 1 mol of glucose (F)
d) by reoxidation of reduced coenzyme formed by convesrion of 1 mol of glucose 6 ATP can be
formed (T)
60. Reaction C6H12O6 + 2 ADP + 2 P 2 + 2 ATP :
a) is summary reaction of complete glucose oxidation in aerobic conditions (F)
b) is summary reaction of aerobic glycolysis (F)
c) is typical for oxidation of glucose in brain (F)
d) is reaction which proceeds in lack of oxygen (T)
2 + 2 ATP : 61. Summary reaction C6H12O6 + 2 ADP + 2 P
a) is reaction of complete glucose oxidation on erythrocytes (T)
b) represents process located in cytosol (T)
c) represents process which allows synthesis of 2 ATP in reaction catalyzed by phosphoglycerate
kinase (T)
d) represents complete glucose oxidation in heart (F)
2 + 8 (6)ATP : 62. Reaction C6H12O6 + 8 (6) ADP + 8 (6)P
a) is summary reaction of complete glucose oxidation in aerobic conditions (F)
b) is reaction of complete glucose oxidation in brain (F)
c) represents the process which allows synthesis of 6 ATP by oxidative phosphorylation (T)
d) represents the process which allows synthesis of 2 ATP by substrate level phosphorylation (T)
63. Reaction C6H12O6 + 8 (6) ADP + 8 (6)P 2 + 8 (6)ATP :
COOH
C O
CH3
COOH
C O
CH3

a) is summary reaction of the process which proceeds in cytosol (T)
b) represent process in which ATP is formed by substrate level and oxidative phosphorylation (T)
c) represents process which allows synthesis of 6 ATP by reoxidation of NADH2 (T)
d) leds to synthesis of the product further conversion of which is located in mitochondria (T)
64. Process which can be expressed by summary reaction C6H12O6 + 6 O2 6 CO2 + 6 H2O + 38 ATP:
a) means summary reaction of aerobic glycolysis (F)
b) involves processes which proceed in cytosol and mitochondria (T)
c) requires processes: glycolysis, oxidative decarboxylation of pyruvate and Krebs cycle (T)
d) is characteristic for glucose oxidation in red blood cells (F)
65. Process expressed by summary reaction C6H12O6 + 6 O2 6 CO2 + 6 H2O + 38 ATP:
a) means complete glucose oxidation in aerobic conditions (T)
b) allows synthesis of 8 ATP in aerobic glycolysis (T)
c) allows synthesis of 6 ATP in reaction catalyzed by pyruvate dehydrogenase (T)
d) leads to synthesis of 6 CO2 vin Krebs cycle (F)
66. Oxidation of glucose which is expressed by summary reactio
C6H12O6 + 6 O2 6 CO2 + 6 H2O + 38 ATP:
a) is oxidation of glucose typical for intensively working muslce (F)
b) allows synthesis of 2 ATP by phosphorylation at the substrate level (F)
c) allows synthesis of 12 ATP in Krebs cycle (F)
d) represents mode of glucose oxidation for example during heart attack (F)
67. During complete aerobic oxidation of glucose:
a) 2 mol of pyruvate are formed (F)
b) 2 CO2 are formed in the process which requires thiaminepyrophosphate (T)
c) 24 ATP are formed in Krebs cycle (T)
d) by process of oxidative phosphorylation totally 24 ATP are formed (F)
68. Complete aerobic oxidation of glucose:
a) leds to synthesis of 6 ATP by oxidative phosphorylation in glycolysis (T)
b) leads to synthesis of 4 CO2 in Krebs cycle (T)
c) leds to synthesis of 6 ATP by substrate level phosphorylation (T)
d) requires thiaminepyrophosphate for the reaction located in cytosol (F)
69. is compound:
a) which is formed by reduction of glyceraldehyde-3-phosphate (F)
b) which is formed by the action of glycerolphosphate dehydrogenase (T)
c) synthesis of which requires NADH2 (T)
d) can be used for gluconeogenesis (T)
70. Compound of the formula :
a) is subtrate of glycerophospate dehydrogenase (T)
b) is product of glycerolphosphate dehydrogenase (T)
c) is formed in reaction important for connection of alcoholic component of triacylglycerol to
gluconeogenesis (T)
d) in adipose tissue can be formed by glycerolkinase (F)
PCH2
HC
O
OH
CH2 OH
PCH2
HC
O
OH
CH2 OH
COOH
C O
CH3
COOH
C OH
CH3
H + NAD+ + NADH2

ATP ADP
COOH
C O
CH2 OH
H P
COOH
C O
CH2
P~COOH
C O
CH3
ADP ATP
C
HC
O
OH
CH2 O P
H
C
C
O
OH
CH2 O P
O P
H
~
71. Reacrion:
a) is catalyzed by lactate dehydrogenase (T)
b) in given direction is important for utilization of lactate in gluconeogenesis (T)
c) is component of Cori,s cycle (T)
d) in given direction is required for reoxidation of NADH2 in aerobic conditions (F)
72. Reaction platí:
a) proceeds in anaerobic conditions (T)
b) is irreversible reaction (F)
c) proceeds for example in red blood cells (T)
d) leads to synthesis of compound which can cause acidosis (T)
73. Reaction:
a) in liver is catalyzed by glucokinase (T)
b) belongs to reversible reactions of glycolysis (F)
c) is reaction of gluconeogenesis (F)
d) leads to synthesis of compound which is substrate of phosphohexoisomerase (T)
74. Compound :
a) is product of glycerolphosphate dehydrogenase (F)
b) is compound with acylphosphate energy rich bond (T)
c) is formed in glycolysis from substrate (T)
d) is substrate of reaction which produces (F)
75. Reaction:
a) is catalyzed by pyruvate kinase (T)
b) allows synthesis of ATP in anaerobic conditions (T)
c) is activated by fructose 1,6-bisphosphate (T)
COOH
C O
CH3
COOH
C OH
CH3
H + NAD++ NADH2
O
OHOH
OH
OH
CH2 OH P
O
OHOH
OH
OH
CH2 O

C
HC
O
OH
CH2 O P
O P~C
HC
O
OH
CH2 O P
H NAD+
NADH+H+
P
d) is activated by glucagon during starvation (F)
76. Which of following enzymes catalyzes reaction:
a) triosephosphate isomerase (F)
b) glyceraldehyd-3-phosphate dehydrogenase (T)
c) phosphoglycerate mutase (F)
d) enzyme coenzyme of which allows synthesis of 3 ATP (T)
77. Phosphorylation at the substrate level by means of utilization of acylphosphate energy rich bond:
a) requires: oxidation of glyceldehyde-3-phosdphate and hydrolytic cleavage
of 1,3-bisphosphoglycerate (F)
b) requires enzymes glyceraldehyde-3-phosphate dehydrogenase and phosphoglycerate kinase (T)
c) is reversible reaction (T)
d) allows synthesis of 2 mol of ATP per 1 mol of glucose (T)
78. Phosphorylation at the substrate level by means of utilization of enolphosphate energy rich bond:
a) requires enzymes enolase and pyruvate kinase (T)
b) requires dehydrogenaztion of pyruvate and conversion of phosphoenolpyruvate to pyruvate (F)
c) allows synthesis of ATP only in anaerobic conditions (F)
d) is important reaction for synthesis of ATP in erthrocytes (T)
79. Anaerobic glycolysis:
a) is the only possibility for ATP synthesis in red blood cells (T)
b) is the process which allows synthesis of ATP by oxidative phosphorylation (F)
c) is main source of energy for all cells of human body (F)
d) leads to synthesis of 6 ATP per 1 glucose (F)
80. Aerobic glycolysis:
a) is the only source of energy for brain (F)
b) leads to synthesis of acetyl-CoA (F)
c) is the process which proceeds in cytosol of all cells of human body (F)
d) leads to synthesis of compound which is further metabolized in mitochondria (T)
81. Which of statements about aerobic glycolysis are correct:
a) leads to synthesis of 2 mol of pyruvate per 1 mol of glucose (T)
b) produces by substrate level phsohprylation the same amount of ATP as anaerobic glycolysis (T)
c) its intermediate can be used for synthesis of triacylglycerols (T)
d) is activated by phosphrylation of enzymes in presence of glucagon (F)
82. In metabolic regulation of glycolysis participate:
a) ADP as acivator pf phosphofructokinase (T)
b) glucose-6-phosphate as inhibitor of hexokinase (T)
c) fructose-1,6-bisphosphate as activator of pyruvate kinase (T)
d) ATP and NADH2 as inhibitors of phosphofructokinase (T)
83. NADH2 formed in reaction catalyzed by glyceraldehyde-3-phosphate dehydrogenase can be used for
synthesis of ATP by oxidative phosphorylation:
a) after its transfer into mitochondria by diffusion (F)
b) after its transfer into mitochondria by active transport (F)
c) by transfer of hydrogens into mitochondria by malate/glutamate shuttle (F)

d) by transfer of hydrogens into mitochondria by glycerolphosphate/dihydroxyacetone phosphate
shuttle (T)
84. Which of statements about transport of reduciong equivalents from cytosol into mitochondria is
correct:
a) is important because mitochondrial membrane is impermeable for NADH2 (T)
b) is important for utilization of reduced coenzymes formed by oxidation of fatty acids (F)
c) malate/aspartate shuttle allows synthesis of 3 ATP (T)
d) glycerolphosphate/dihydroxyacetonephosphate allows synthesis of 1 ATP more than
malate/aspartate shuttle (F)
85. Which of following reactions of glycolysis are irreversible:
a) reaction of fructose-1,6-bisphosphate synthesis (T)
b) reaction catalyzed by pyruvate kinase (T)
c) reaction of fructose-6-phosphate synthesis (F)
d) reaction of substrate level phosphorylation which uses phosphoenolpyruvare (T)
86. In the process of aerobic glycolysis.
a) activation of glucose requires 2 ATP (T)
b) by substrate level phosphorylation 4 ATP are formed (T)
c) lactate as final product is formed (F)
d) compound with thioester energy rich bond is formed as final product (F)
87. During aerobic glycolysis:
a) 2 CO2 are formed (F)
b) 2 ATP are formed by substrate level phosphorylation (F)
c) 6 ATP can be formed by reoxidation of NADH2 in mitochondria (T)
d) 12 ATP can be formed per 1 glucose (F)
88. Reaction typical for anaerobic glycolysis is:
a) conversion of pyruvate to lactate (T)
b) reaction which is required for regeneratzion of oxidized form of NAD (T)
c) reaction catalyzed by pyruvate kinase (F)
d) reaction typical for oxidation of glucose in brain (F)
89. Reaction of conversion of pyruvate to lactate:
a) is catalyzed by lactate dehydrogenase (T)
b) proceeds in cytosol (T)
c) is reversible reaction (T)
d) is final step of the process by which red blood cells gain energy (T)
90. Lactate dehydrogenase catalyzes reaction:
a) which uses pyruvate as substrate (T)
b) synthesis of pyruvate from lactate for gluconeogenesis in liver (T)
c) of transfer of hydrogens from NADH2 to pyruvate in good oxygen supply (F)
d) which is regulatory step of glycolysis in red blood cells (F)
91. Anaerobic glycolysis produces less ATP than aerobic glycolysis because:
a) in anaerobic conditions does not proceed Krebs cycle (F)
b) in aerobic conditions ATP is formd also by oxidative phosphorylation (T)
c) in aerobic conditions ATP is formd also by oxidation of NADH2 (T)
d) in anaerobic conditions lactate is formed (T)
92. Conversion of pyruvate to lactate:
a) is typical for anaerobic glycolysis (T)
b) in anaerobic conditions is required because NADH2 cannot be oxidized in terminal oxidation (T)
c) is final reaction of glycolysis in red blood cells (T)

d) is irreversible (F)
93. Production of lactate in working muscle::
a) is catalyzed by lactate dehydrogenase (F)
b) is consequence of relative lack of oxygen during work (T)
c) is ireversible reaction (F)
d) in higher concentrations can cause acidosis (T)
94. Red blood cells gain energy in metabolic process which:
a) is the only source of energy in anaerobic conditions (T)
b) produces lactate as final product (T)
c) leads to synthesis of compound which in red blood cells cannot be matabolized (T)
d) leads to synthesis of compound which can be used for gluconeogenesis (T)
95. Source of energy for red blood cells can be:
a) only glucose (T)
b) fatty acids during starvation (F)
c) ketone bodies during starvation (F)
d) compound which in red blood cells is oxidized by anaerobic glycolysis (T)
96. Conversion of lactate to pyruvate:
a) is catalyzed by lactate dehydrogenase (T)
b) proceeds in tissues in lack of oxygen (F)
c) allows utilization of lactate in gluconeogenesis (T)
d) uses coenzyme which allows synthesis of 3 ATP by oxidative phosphorylation (T)
97. Cori,s cycle:
a) connects anaerobic glycolysis in muscles and gluconeogenesis in liver (T)
b) utilizes lactate formed in liver vfor synthesis of ATP in heart (F)
c) in liver utilizes lactate formed in tissues in anaerobic conditions (T)
d) in the liver requires enzyme lactate dehydrogenase (T)
98. Compound with acylphosphate energy rich bond in glycolysis:
a) is formed by glycerolphosphate dehydrogenase (F)
b) is formed by reaction which requires FAD as coenzyme (F)
c) contains two phosphate residues bound by energy rich bond (F)
d) is used for synthesis of ATP by substrate level phosphorylation using enzyme glycerolkinase (F)
99. Which of statments about intermediates of glycolysis with energy rich bonds are true:
a) it is for example 1,3-bisphosphoglycerate (T)
b) it is enolpyruvate (F)
c) all are formed by irreversible reactions (F)
d) process of ATP synthesis using energy of energy rich bonds is called substrate level
phosphorylation (T)
100. Intermediates of glycolysis with energy rich bond:
a) are totally three (F)
b) are compounds formed in reversible reactions of glycolysis (T)
c) are compound used for synthesis of ATP in both aerobic and anaerobic conditions (T)
d) are used for synthesis of ATP in reversible reactions of glycolysis (F)
101. In the process of anaerobic glycolysis:
a) cells can form 2 ATP by oxidation of one glucose (T)
b) lactate is formed during reoxidation of NADH2 (T)
c) for activation of glucose 1 ATP is spent (F)
d) is important synthesis of compounds with energy rich bonds (T)

102. NADH2 in anaerobic glycolysis is oxidized by:
a) reaction in which lactate is formed (T)
b) reaction which proceeds in cytosol (T)
c) reaction of substrate level phsohorylation (F)
d) synthesis of compound which can be used for gluconeogenesis (T)
103. Which cells of human body use glucose as the only source of energy:
a) brain tissue (F)
b) heart cells (F)
c) red blood cells which by glucose oxidation gain 6 ATP (F)
d) red blood cells which can form ATP only by substrate level phosphorylation (T)
104. Lactate:
a) is formed as final product of glycolysis in brain (F)
b) is formed in higher concentrations in lack of oxygen (T)
c) in heart can be used for ATP synthesis (T)
d) as final product of metabolism is excreted by kidneys (F)
105. Which of following tissues is almost completely dependent on glucose as source of energy:
a) brain which during stavation can use also ketone bodies (T)
b) red blood cell which during starvation oxidize also fatty acids (F)
c) heart because requires large amounts of ATP (F)
d) brain becasue does not contain mitochondria (F)
106. Which statements about regulation of glycolysis are correct:
a) key regulatory enzyme is reaction of fructose-1,6-bisphosphate synthesis (T)
b) high concentrations of ATP and NADH2 inhibit phosphofructokinase and pyruvate kinase (T)
c) glucagon by phosdphorylation of tandem enzyme increases concentration of
fructose-2,6-bisphosphate inhibits phosphofructokinase (F)
d) insulin by dephosphorylation of pyruvate kinase causes its activation (T)
107. Gluconeogenesis is the process which:
a) proceeds only in liver (F)
b) leads to synthesis of glucose form glycogen (F)
c) allows synthesis of glucose mainly form amino acids (T)
d) is activated by insulin (F)
108. Gluconeogenesis:
a) is the process of glucose synthesis from fatty acids (F)
b) proceeds in liver and kidneys (T)
c) is not important because during starvation glucose is synthesized by glycogen degradation (F)
d) is activated by glucocorticoids (T)
109. For glucose synthesis by gluconeogenesis can be used:
a) lactate (T)
b) amino acids – for example alanine (T)
c) alcoholic component of triacylglycerols (T)
d) amino acids which can be changed to acetyl-CoA (F)
110. Organism can use for glucose synthesis by gluconeogenesis:
a) fatty acids (F)
b) ketone bodies (F)
c) glucogenic amino acids (T)
d) amino acid leucine (F)
111. Pyruvate carboxylase is the enzyme which:
a) is located in mitochondria (T)

b) as substrate uses compound formed form lactate (T)
c) requires GTP as source of energy (F)
d) is activated by phosphorylation in presence of glucagon (F)
112. Phosphoenolpyruvate carboxykinase:
a) uses pyruvate as substrate (F)
b) catalyzes reaction of glycolysis (F)
c) requires biotin as coenzyme (F)
d) requires ATP as source of energy (F)
113. Phosphoenolpyruvate carboxykinase is the enzyme which:
a) leads to synthesis of phosphoenolpyruvate (T)
b) catalyzes conversion of phosphoenolpyruvate in gluconeogenesis (F)
c) requires GTP as source energy (T)
d) uses oxaloacetate as substrate (T)
114. For synthesis of free glucose in the process of gluconeogenesis is responsible enzyme:
a) glucose-6-phosphate phosphatase (T)
b) which requires water as one of substrate of reaction (F)
c) present in liver and kidneys (T)
d) which participates also in glycogen degradation in muscles (F)
115. Conversion of glucose-6-phopsphate into free glucose catalyzes enzyme:
a) present only in liver (F)
b) glucokinase (F)
c) required for glucose synthesis for irreversibility of reaction catalyzed by glucokinase (T)
d) which is required only for gluconeogenesis (F)
116. Substrate for phosphoenolpyruvate carboxykinase is:
a) oxaloacetate (T)
b) product of pyruvate dehydrogenase (F)
c) compound which is formed form fatty acids (F)
d) compound for synthesis of which GTP is required (F)
117. Enzyme of gluconeogenesis which produces compound with enolphosphate energy rich bond:
a) is pyruvate kinase (F)
b) is enolase (F)
c) uses as substrate oxaloacetate (T)
d) requires specific source of energy GTP (T)
118. Reaction:
a) proceeds in cytosol (F)
b) is activated by acetyl-CoA (T)
c) is catalyzed by pyruvatekinase (F)
d) is activated by insulin durong starvation (F)
119. Pyruvate carboxylase:
a) catalyzes reaction
(T)
COOH
C O
CH2COOHC
O
CH3
COOH
+ ATP + CO 2 + ADP + P i
COOH
C O
CH2
COOH
C O
CH3COOH
+ ATP + CO 2 + ADP + P i

b) catalyzes reaction which proceeds in liver and kidneys (T)
c) is located in mitochondria (T)
d) is induced by cortisol (T)
120. Synthesis of phosphoenolpyruvate from oxaloacetate proceeds:
a) by reaction (T)
b) by reaction catalyzed by phosphoenolpyruvatecarboxykinase (T)
c) by reaction which is inhibited during starvation (F)
d) in the process which allows utilization of lactate and glucogenic amino acids for glucose synthesis
(T)
121. Phosphoenolpyruvate carboxykinase:
a) catalyzes reaction
(F)
b) is located in cytosol and mitochondria (T)
c) is enzyme activity of which is increased in hyperglyceamia (F)
d) is important enzyme for synthesis of glucose by gluconeogenesis (T)
122. Gluconeogenesis:
a) is important for glucose synthesis during starvation (T)
b) is the process in which irreversible reactions of glycolysis are bypassed by specific enzymes (T)
c) is process which allows utilization of ketone bodies for glucose synthesis (F)
d) is the process activated by hormone formed in -cells of Langerhans islets of pancreas (T)
123. During long starvation:
a) liver glycogen is main source of blood glucose (F)
b) organism uses fatty acids for gluconeogenesis (F)
c) gluconeogenesis is activated by insulin (F)
d) main substrate for gluconeogenesis are glucogenic amino acids (T)
124. During long starvation:
a) level of glucose does not decrease to zero because glucose can be formed by gluconeogenesis (T)
b) cortisol increases degradation of tissue protein and induces enzymes of gluconeogenesis in liver
(T)
c) source of energy for brain are ketone bodies and glucose (T)
d) insulin is the main activator of gluconeogenesis (F)
125. Process of gluconeogenesis:
a) proceeds in liver, kidneys and skeletal muscles (F)
b) requires specific source of energy – CTP (F)
c) for the brain is not important because brain durng starvation uses fatty acids and ketone bodies (F)
d) requires specific enzymes for example – pyruvate carboxylase, phosphenolpyruvate
carboxykinasee and two phosphatases (T)
126. Which of following enzymes are required for glucose synthesis by gluconeogenesis:
a) pyruvate carboxylase (T)
b) pyruvate kinase (F)
c) glucose-6-phosphate phosphatase (T)
d) glucokinase (F)
COOH
C O
CH2
P~
HOOC C O
CH2
COOH
+ GTP + GDP + CO2
COOH
C O
CH2
P~COOH
C O
CH3
+ CO2 + GTP + GDP + P i

127. Acetyl-CoA cannot be for gluconeogenesis because:
a) its conversion to pyruvate is not possible (T)
b) reaction catalyzed by pyruvate dehydrogenase is not reversible (T)
c) reaction catalyzed by pyruvate kinase is not reversible (F)
d) during starvation is not formed (F)
128. Connection of proteins with gluconeogenesis:
a) is not important because glucose is synthesized only from lactate (F)
b) requires proteases mainly in liver (F)
c) requires lactate dehydrogenase (F)
d) requires reactions by which amino acids are changed to keto acids (T)
129. Part of triacylglycerols which can be used for gluconeogenesis:
a) is glycerol (T)
b) is glycerol formed by hydrolysis of TAG by pancreatic lipase (F)
c) is chain of fatty acid (F)
d) is compound which is activated by glycerol kinase (T)
130. Connection of glycerol with process of gluconeogenesis requires:
a) enzyme glycerolkinase in adipose tissue (F)
b) enzyme glycerolkinase in liver (T)
c) enzyme which changes glycerolphosphate to dihydroxyacetonephosphate (T)
d) ATP as source of energy (T)
131. Gluconeogenesis is influenced by:
a) hormone of -cells of Langerhans islets which increases level of cAMP and by phosphorylation
activates enzymes of gluconeogenesis (F)
b) hormone of - cells of Langerhans islets which causes of enzymes of glycolysis and by this way
activation of gluconeogenesis (F)
c) cortisol which after bindimg to cytosoloc receptor increases production of enzymes required for
synthesis of glucose by gluconeogenesis (T)
d) hormone secreted from pancreas during starvation which causes phosphorylation of pyruvate
kinase and by this way allows utilization of phosphoenolpyruvate in gluconeogenesis (T)
132. Glygogen is compound:
a) which contains -1,4 and -1,6-glycosidic bonds (T)
b) which storage of glucose mainly in brain (F)
c) for synthesis of which UTP is required (T)
d) degradation of which is activated by glucagon (T)
133. Intermediate of glycogen synthesis is:
a) compound formed by phosphoglucomutase (T)
b) compound formed form glucose-6-phosphate and UTP (F)
c) compound which contains phosphoanhydride energy rich bond (T)
d) compound which is formed also as intermediate during digestion of glycogen in intestine (F)
134. UDP-glucose is compound which:
a) is formed from glucose-1-phosphate and UTP (T)
b) is substrate for glycogephosphorylase (F)
c) is formed in both synthesis and degradation of glycogen (F)
d) is required also during conversion of glucose to galactose (F)
135. Glucose-1-phosphate during glycogen synthesis:
a) is formed by glucokinase (F)
b) is formed by reversible reaction (T)
c) is formed by reaction which requires UTP (F)
d) is formed by the same enzyme as glucose-1-phosphate during glycogen degradation (F)

136. lucose-1-phosphate in metabolism of glycogen:
a) is intermediate of glycogen synthesis (T)
b) during degradation is formed by glycogenphosphorylase (T)
c) during synthesis of glycogen is formed by phosphoglucomutase (T)
d) during degradation of glycogen is changed by phosphoglucomutase (T)
137. Further conversion of glucose-1-phosphate in glycogen synthesis:
a) requires ATP as source of energy (F)
b) is catalyzed by phosphoglucomutase (F)
c) is catalyzed by UDPG-phosphorylase (T)
d) leads to synthesis of compound which is substrate for glycogen synthase (T)
138. Further conversion of glucose-1-phosphate in glycogen degradation:
a) requires ATP as source of energy (F)
b) is catalyzed by phosphoglucomutase (T)
c) is catalyzed by UDPG-phosphorylase (F)
d) leads to synthesis of compound which is substrate for glucose-6-phosphate phosphatase (T)
139. Synthesis of UDP-glucose:
a) requires glucose-6-phosphate and UTP as substrates (F)
b) catalyzes enzyme UDPG-phosphorylase (T)
c) is reaction of glucose activation in glycoge synthesis (T)
d) is reversible reaction (F)
140. UDP-glucose is compound:
a) which is formed by reaction of glucose-1-phosphate and UTP (T)
b) which is substrate for glycogensynthase (T)
c) which is formed during glycogen degradation (F)
d) which is substrate for synthesis of -1,6-glycosidic bond (F)
141. Glycogensynthase catalyzes synthesis of:
a) -1,4-glycosidic bond (T)
b) bond which during degradation is broken by hydrolysis (F)
c) bond which in the liver is broken by glycogenphosphorylase (T)
d) bond which is responsible for branched structure of glycogen (F)
142. -1,4 glycosidic bond:
a) is responsible for branched structure of glycogen (F)
b) is formed by action of glycogenphosphorylase (F)
c) is bond for synthesis of which UDP-glucose is required (T)
d) is bond which is broken by glygogenphosphorylase into glucose-1-phosphate (T)
143. Glycogensynthase is the enzyme which:
a) uses glucose-1-phosphate as substrate (F)
b) is responsible for synthesis of -1,6-glycosidic bond (F)
c) is activated by dephosphorylation in presence of insulin (T)
d) is main regulatory enzyme of glycogen synthesis (T)
144. Compound:
a) is substrate for UDPG-phosphorylase (T)
b) is substrate for glycogensynthase (F)
c) is product of digestion of glycogen by amylase (F)
d) is product of glycogen degradation in liver (T)
O
OH OOH
OH
CH2 OH
P
P
O
OHOH
OH
OH
CH2 O
O
OH OOH
OH
CH2 OH
P
145. Compound:
a) by glucose-6-phosphate phosphatase can be changed into free glucose T)
b) is final product of glycogen degradation in muscles T)
c) during glycogen degradation is formed by phosphoglucomutase T)
d) during glycogen synthesis is formed by phosphoglucomutase F)
146. Uridinediphosphateglucose (UDPG):
a) is compound with formula
(T)
b) is donor of glucose units in glycogen synthesis T)
c) is formed by reaction of and UTP (F)
d) is compound with formula (F)
147. Glycogensynthase:
a) catalyzes synthesis of linear chain of glycogen (T)
b) uses as substrate compound (F)
c) requires primer (T)
d) uses as substrate compound (T)
148. Compound:
a) is formed by spliting of -1,6-glycosidic bonds in glycogen (F)
b) is product of pancreatic amylase (F)
P
O
OHOH
OH
OH
CH2 O
O
OH OOH
OH
CH2 OH
P O P
HN
O
O N
O
OHOH
CH2O
O
OH OOH
OH
P O P
HN
O
O N
O
OHOH
CH2O
CH2 OH
O
OH OOH
OH
CH2 OH
P O P
HN
O
O N
O
OHOH
CH2O
O
OH OOH
OH
CH2 OH
P

c) is formed by spliting of -1,4-glycosidic bonds by glycogenphosphorylase (T)
d) in glycogen synthesis is changed to UDP-glucose by UDPG-phosphorylase (T)
149. Compound
a) is product of maltase (F)
b) is disaccharide maltose (T)
c) is product of glycogenphosphorylase (F)
d) is formed during glycogen hydrolysis in intestine (T)
150. Compound:
a) is product of glycogen degradation by pancreatic amylase (F)
b) is product of glycogen phosphorylase (F)
c) is formed by glucose-6-phosphate phosphatase during glycogen degradation in muscles (F)
d) is product of glycogen degradatioon by debranching enzyme (T)
151. Reaction:
a) is activated by glucagon and adrenaline (T)
b) is catalyzed by ATP-ase (F)
c) is reaction of synthesis of 2nd messenger cAMP (F)
d) leads to synthesis of compound which by phosphorylation activates glycogenphosphorylase (T)
152. -1,6 glycosidic bond:
a) is formed by glycogensynthase (F)
b) is bond for synthesis of which UDP-gucose as substrate is required (F)
c) during degradation of glycogen is broken by glycogenphosphorylase (F)
d) during degradation of glycogen is broken into free glucose (T)
153. Activity glycogensynthase in the liver decreases:
a) mainly hormine glucagon (T)
b) hormone formed by -cells of Langerhans islets (F)
c) hormone which in its mechanism of action increases level of cAMP (T)
d) hormone which is secreted in low glucose concentration in blood (T)
154. Activity of glycogensynthase in liver increases:
a) hormone of -cells of Langerhans islets (T)
b) hormone which causes dephosphorylation of glycogensynthase (T)
c) hormone which is secreted during starvation and by phosphorylation changes enzyme into active
form (F)
d) hormoe which is directly transported into liver cell (F)
155. Glycogenphosphorylase is the enzyme which:
O
OHOH
OH
CH2 OH
O
O
OHOH
OH
CH2 OH
O
OH OHOH
OH
CH2 OH
N
N N
N
NH2
O
OH OH
CH2 O P
N
N N
N
NH2
O
OH O
CH2 O
P
O P O P
PPi

a) causes hydrolysis of glycogen (F)
b) breaks -1,4 glycosidic bond (T)
c) produces glucose-1-phosphate (T)
d) in muscle is activated by glucagon (F)
156. Glycogenphosphorylase catalyzes:
a) synthesis of glucose-1-phosphate (T)
b) reaction which requires H3PO4 (T)
c) spliting of -1,4-glycosidic bond (T)
d) reversible reaction (F)
157. Glucose-1-phosphate:
a) is product of glycogenphosphorylase (T)
b) is formed by spliting of -1,6-glycosidic bond (F)
c) is intermediate of glycogen synthesis (T)
d) is intermediate of glycogen degradation (T)
158. Glucose-6-phosphate:
a) is intermediate of glycogen synthesis (T)
b) in glycogen degradation is formed by phosphoglucomutase (T)
c) is formed by degradation of -1,6-glycosidic bond (F)
d) is final product of glycogen degradation in skeletal muscles (T)
159. Free glucose during glycogen degradation can be formed:
a) by hydrolysis of -1,4-glycosidic bond (F)
b) by glycogenphosphorylase (F)
c) by debranching enzyme (T)
d) in both liver and skeletal muscle (T)
160. Activation of glycogenphosphorylase mediates:
a) ATP by activation of proteinkinase (F)
b) cAMP (T)
c) compound which is formed by cytosolic enzyme (F)
d) compound which is inactivated by adenylcyclase (F)
161. Regulatory enzymes of glycogen metabolism:
a) are glycogensynthase and glycogenphosphorylase (T)
b) are regulated by covalent modification (T)
c) both are active in dephosphorylated form (F)
d) are enzymes phosphorylation of which is activated by cAMP (T)
162. Glycogensynthase and glycogen phosphorylase are enzymes which:
a) are regulated by phosphorylation and dephosphorylation (T)
b) are phosphorylated in presence of cAMP (T)
c) both are activated during starvation (F)
d) both participate in regulation of glycogen degradation and glycolysis (F)
163. Glycogensynthasephosphorylasekinase (GSPK) is enzyme which:
a) is active in dephosphorylated form (F)
b) is responsible for phosphorylation of glycogensynthase and glycogenphosphorylase (T)
c) in presence of cAMP is in active phosphorylated form (T)
d) causes phosphorylation of glycogenphosphorylase and by this way activation of glycogen
degradation (T)
164. Comparing glycogen degradation in liver and in skeletal muscles we can say:
a) final product of glycogen degradation in muscles is glucose-6-phosphate (T)
b) degradation of glycogen in liver is activated by glucagon (T)

c) degradation of glycogen in muscles is activated by Ca2+ ions (T)
d) degradation of glycogen in both – liver and skeletal muscles can increase glucose level in blood
(F)
165. Pentose phosphate pathway:
a) is important for ATP synthesis (F)
b) proceeds in cytosol (T)
c) is required for ribosephosphate synthesis (T)
d) is required for synthesis of NADPH2 (T)
166. In pentose phosphate pathway:
a) glucose is reduced (F)
b) pentoses like deoxyribose are formed (F)
c) NADPH2 is formed in reaction of glucose-6-phosphate synthesis (F)
d) NADPH2 is formed in reaction of glucose-6-phosphate conversion (T)
167. Ribose-5-phosphate is compound which:
a) is formed in pentose phosphate pathway (T)
b) is formed in every metabolic process of glucose oxidation (F)
c) is formed in the process located in cytosol (T)
d) requires NAD for its synthesis (F)
168. Reaction:
a) is locatzed in cytosol (T)
b) is catalyzed by lactonase (F)
c) is important for energy metabolism (F)
d) is reaction of metabolic pathway where glucose is changed to pentoses (T)je lokalizovaná v cytozole
(T)
169. Glucose-6-phosphate dehydrogenase:
a) catalyzes reaction:
(F)
b) is cytosolic enzyme (T)
c) contains as coenzyme derivative of B2 vitamin (F)
d) leads to synthesis of coenzyme required for fatty acid synthesis (T)
170. Reaction:
P
O
OHOH
OH
OH
CH2 O
+ NADP+
C
C OH
C HHO
C OHH
CH
CH2
O
O
O P
H
+ NADPH2
P
O
OHOH
OH
OH
CH2 O
+ NAD+
C
C OH
C HHO
C OHH
CH
CH2
O
O
O P
H
+ NADH2
C
C OH
C HHO
C OHH
CH
CH2
O
O
O P
H
C
C OH
C HHO
C OHH
CH
CH2
O
O P
H
OH
OH
+ H2O

a) is catalyzed by lactonase (T)
b) belongs to hydrolytic reactions (T)
c) leds to synthesis of 6-phosphogluconate (T)
d) is reaction of the cycle which is important for synthesis of NADPH2 (T)
171. Reaction:
a) Is located in cytosol (T)
b) is reaction of glucose oxidation by glycolysis (F)
c) is catalyzed by epimerase (F)
d) is catalyzed by isomerase (T)
172. Thiaminepyrophosphate in pentose phosphate patway is required for:
a) glucose-6-phosphate dehydrogenase (F)
b) synthesis of ribose-5-phosphate (F)
c) transketolase (T)
d) reaction in which sedoheptulose-7-phosphate is formed (T)
173. NADPH+H+:
a) is formed by 6-phosphogluconate dehydrogenase (T)
b) is during synthesis of 6-phosphogluconate (T)
c) is important coenzyme for synthesis of cholesterol (T)
d) is important coenzyme for oxidation of fatty acids (F)
174. Conversion of glucose-6-phosphate to 6-phosphogluconate:
a) is reaction of glycolysis (F)
b) requires NADPH2 (F)
c) is important for enetgy metabolism (F)
d) allows production of coenzyme required for fatty acids synthesis (T)
175. Glucose-6-phosphate dehydrogenase:
a) catalyzes synthesis of 6-phosphogluconate (T)
b) catalyzes reaction of pentose phosphate pathway (T)
c) is source of hydrogens for terminal oxidation (F)
d) uses as coenzyme NADP (T)
176. Reaction of conversion of 6-phosphogluconate in pentose phosphate pathway:
a) is catalyzed by 6-phosphogluconate isomerase (F)
b) requires NAD as coenzyme (F)
c) leads to synthesis of ribulose-5-phosphate (T)
d) allows synthesis of coenzyme which is required for fatty acid synthesis (T)
177. Reaction of pentose phosphate pathway in which ribulose-5-phosphate is formed:
a) is catalyzed by glucose-6-phosphate dehydrogenase (F)
b) proceeds in mitochondria (F)
c) uses NADP as coenzyme (T)
d) uses 6-phosphogluconate as substrate (T)
CH2 OH
C O
C OHH
CH
CH2 O P
OH
C
C OH
C OHH
CH
CH2 O P
OH
H
O
H

178. Increased production of ketone bodies in diabetes is caused by:
a) increased activity of Krebs cycle (F)
b) increased concentration of lactate in blood (F)
c) increased concetration of cortisol (F)
d) increased oxidation of lipids (T)
179. Which of following hormones participate in regulation of blood glucose:
a) adrenaline that increases glucose level (T)
b) all hormones of adrenal cortex (F)
c) insulin that decreases glucose level (T)
d) glucocorticoids that increase glucose level (T)
180. Glucocorticoids are hormones which:
a) are secreted from pancreas (F)
b) increase degradation of glycogen (F)
c) decrease rate of glycolysis (F)
d) increase level of cAMP (F)
181. Insulin is the hormone which:
a) decreases activity of pyruvate kinase (F)
b) increases rate of glycolysis (T)
c) activates synthesis of fatty acids (T)
d) stimulates gluconeogenesis (F)
182. Epinephrine (adrenaline) is the hormone which:
a) is secreted from adrenal cortex (F)
b) activates gluconeogenesis in liver (F)
c) activates glycogenolysis in skeletal muscle (T)
d) causes decrease of cAMP (F)
183. In regulation of blood glucose level participate:
a) thyroid hormone (T)
b) calcitonine (F)
c) glucagon (T)
d) hormone secreted during stress (T)
184. Insulin:
a) is secreted from -cells (F)
b) is secreted if blood glucose level is decreased (F)
c) is required for glucose transport into the cells (T)
d) decreases glucose oxidation in cells (F)
185. When level of glucose in blood increases:
a) glucagon is secreted (F)
b) fatty acids from adipose tissue are released (F)
c) glycogen in liver is broken (F)
d) gluconeogenesis is inhibited by insulin (T)
186. Insulin decreases blood glucose level by:
a) stimulation of gluconeogenesis (F)
b) activation of glucose conversion into fatty acids (T)
c) stimulation of glucose transport into the cells (T)
d) activation of fatty acid degradation in adipose tissue (F)
187. Insulin participates in regulation of blood glucose level by:
a) increased glucose transport into the cells (T)

COOH
C
O
H3C
b) activation of glycogen synthesis (T)
c) inhibition of fatty acid synthesis (F)
d) inhibition of gluconeogenesis (T)
188. Glucocorticoids:
a) are hormones formed in adrenal medulla (F)
b) increase level of cAMP (F)
c) increase synthesis of gluconeogenetic enzymes (T)
d) stimulate degradation of proteins in extrahepatic tissues (T)
189. Glucagon is the hormone which:
a) is secreted when plasma glucose level is low (T)
b) increases plasma membrane permeability for glucose (F)
c) activates gluconeogenesis (T)
d) activates synthesis of fatty acids from glucose (F)
190. Increased glucose concetration in blood causes:
a) increased osmolarity of blood (T)
b) decrease of pH – acidosis (F)
c) glycosuria (T)
d) decreased insulin secretion (F)
KREBS CYCLE
191. Acetyl-CoA is compound which:
a) is formed mainly from amino acids (F)
b) can be formed form glucose (T)
c) is formed only in cytosol (F)
d) cannot be used for gluconeogensis (T)
192. Which offollowing statements about acetyl-CoA is correct:
a) acetyl-CoA from glucose is formed in cytosol (F)
b) synthesis of acetyl-CoA from pyruvate requires thaiminepyrophosphate (T)
c) synthesis of acetyl-CoA from pyruvate is irreversible reaction (T)
d) utilization of acetyl-CoA is possible only in mitochondria (F)
193. Acetyl-CoA can be used:
a) for synthesis of ATP in mitochondria (T)
b) for synthesis of fatty acids in cytosol (T)
c) for synthesis of ketone bodies in cytosol (F)
d) for synthesis of cholesterol in mitochondria (F)
194. Acetyl-CoA:
a) can be oxidized in mitochondria and cytosol (F)
b) for synthesis of fatty acids is used in cytosol (T)
c) can be formed by pyruvate dehydrogenase (T)
d) can be changed into ketone bodies (T)
195. Conversion:
a) requires this compound as substrate F)
b) is catalyzed by transaminase (F)
c) is inhibited by ATP (T)
COOH
CH2
CO
COOHCOOH
CH2
CHO COOH
CH2
COOH

d) proceeds in mitochondria (T)
196. Citrate synthase:
a) is located in inner mitochondial membrane (F)
b) is located in cytosol (F)
c) catalyzes condensation of acetyl-CoA and citrate (F)
d) produces compound which is utilized in mitochondria only (F)
197. Citrate synthase catalyzes reaction which:
a) proceeds in mitochondrial matrix (T)
b) uses as substrate acetyl-CoA (T)
c) is activated by ATP (F)
d) produces compound which can be used for synthesis of fatty acids (T)
198. Which of statements about regulatory role of citrate synthase in Krebs cycle are correct:
a) it is one of two regulatory enzymes of Krebs cycle (T)
b) enzyme is inhibited by citrate (T)
c) main regulator of citrate synthase is insulin (F)
d) enzyme is inhibited by ATP and NADH+H+ (T)
199. Citrate synthase in regulation of Krebs cycle:
a) is activated by citrate (F)
b) is inhibited by ATP and NAD (F)
c) is activated by ATP (F)
d) is not influenced by hormones (T)
200. Aconitase is enzyme which:
a) is responsible for conversion of citrate to isocitrate (T)
b) uses NAD as coenzyme (F)
c) is important regulatory enzyme of Krebs cycle (F)
d) catalyzes reversible reaction (T)
201. Reaction catalyzed by aconitase:
a) allows conversion of citrate to isocitrate (T)
b) as substrate uses citrate (T)
c) leads to synthesis of compound for conversion of which NAD is required (T)
d) is located in outer mitochondrial membrane (F)
202. Isomere of citrate in Krebs cycle in formed in reaction which:
a) is catalyzed by citrate synthase (F)
b) is catalyzed by citrate isomerase (F)
c) is catalyzed by acinitase (T)
d) does not require any coenzyme (T)
203. Reaction catalyzed by isocitrate dehydrogenase:
a) can proceed in both mitochondria and cytosol (T)
b) proceeds only in aerobic conditions (T)
c) uses NAD+ as coenzyme (T)
d) is inhibited by ADP and NADH2 (F)
204. Isocitrate dehydrogenase is enzyme which:
a) is present only in mitochondria (F)
b) is key regulatory enzyme of Krebs cycle (T)
c) catalyzes synthesis of isocitrate (F)
d) allows synthesis of 3 ATP by oxidative phosphorylation (T)
205. Reaction of conversion of isocitrate in Krebs cycle:

a) is catalyzed by isocitrate dehydrogenase (T)
b) requires FAD as coenzyme (F)
c) is activated by ATP (F)
d) allows synthesis of ATP by phosdphorylation at the substrate level (F)
206. Conversion
a) is catalyzed by ketoglutarate dehydrogenase (F)
b) requires NAD as coenzyme (T)
c) proceeds in mitochondria and cytosol (T)
d) allows synthesis of 3 ATP by oxidative phosphorylation (T)
207. -ketoglutarate in Krebs cycle is formed:
a) by action of isocitrate dehydrogenase (T)
b) by the action of aconitase (F)
c) by reaction which requires thiaminepyrophosphate (F)
d) in reaction which is key regulatory step of Krebs cycle (T)
208. -ketoglutarate dehydrogenase:
a) is enzyme of Krebs cycle which catalyzes reversible reaction (F)
b) requires 5 coenzymes (T)
c) is responsible for synthesis of compound with thioester energy rich bond (T)
d) is enzyme coenzyme of which allows synthesis of 4 ATP (F)
209. -ketoglutarate dehydrogenase:
a) catalyzes oxidative decarboxylation of -ketoglutarate (T)
b) catalyzes synthesis of compound with thioester energy rich bond (T)
c) enables synthesis of 2 ATP in respiratory chain (F)
d) requires thiamine pyrophosphate and lipoate as coenzymes (T)
210. Enzyme which catalyzes convesrion of -ketoglutarate to succinyl-CoA:
a) is -ketoglutarate dehydrogenase (T)
b) is key regulatory enzyme of Krebs cycle (F)
c) requires coenzymes thiaminepyrophosphate, lipoate, CoA, NAD and FAD (T)
d) produces compound which can be used for synthesis of ATP by substrate level phosphorylation (T)
211. Succinyl-CoA:
a) is formed in reaction which reauires thiaminepyrophosphate, lipoate, CoA, NAD and FAD (T)
b) contains energy rich thioester bond (T)
c) can be used for ATP synthesis by substrate level phosphorylation also in anaerobic conditions (F)
d) is important for utilization of ketone bodies (T)
212. Conversion of succinyl-CoA to succinate:
a) is reaction where by substrate level phosphorylation GTP is formed (T)
b) is catalyzed by succinate dehydrogenase (F)
c) allows synthesis of ATP only in aerobic conditions (T)
d) requires thiaminepyrophosphate as coenzyme (F)
213. Substrate level phosphorylation in Krebs cycle:
a) is reaction of conversion of succinyl-CoA to succinate (T)
b) is catalyzed by succinylthiokinase (T)
c) uses energy of thioester energy rich bond for synthesis of GTP (T)
d) as it is substrate level phosphorylation proceeds also in anaerobic conditions (F)
COOH
C
CH2
CH2
COOH
O
COOH
CH2
HC COOH
CH
COOH
HO

214. Reaction of substrate level phosphorylation in Krebs cycle:
a) changes succinate to succinyl-CoA (F)
b) produces GTP by the enzyme thiolase (T)
c) requires enzyme succinylthiokinase (T)
d) produces compound which is source of energy in proteosynthesis (T)
215. Further conversion of succinate in Krebs cycle:
a) requires FAD as coenzyme (T)
b) leads to synthesis of fumarate (T)
c) allows synthesis of 4 ATP by oxidative phosphorylation (F)
d) is possible only in aerobic conditions (T)
216. Succinate dehydrogenase in Krebs cycle:
a) is responsible for conversion of succinyl-CoA (F)
b) uses as substrate product of succinylthiokonase (T)
c) uses as coenzyme compound derived form vitamin B2 (T)
d) uses as coenzyme compound which allows synthesis of 2 ATP by oxidative phosphorylatiion (T)
217. Conversion of fumarate in Krebs cycle:
a) leads to synzhesis of unsaturated dicarboxylic acid (F)
b) requires NAD as coenzyme (F)
c) is performed by adition of water (T)
d) is catalyzed by malic enzyme (F)
218. Fumarase is enzyme which:
a) uses as substrate trans-isomere of unsatuated dicarboxylic acid (T)
b) uses as substrate compound which is product of succinylthiokinase (F)
c) catalyzes reaction in which ATP is not formed (T)
d) leads to synthesis of malate (T)
219. Conversion
a) is catalyzed by malate dehydrogenase (F)
b) is located in mitochondria (T)
c) requires NAD as coenzyme (F)
d) is hydrogenation (F)
220. Malate dehydrogenase is the enzyme which:
a) catalyzes reaction important only for Krebs cycle (F)
b) uses as coenzyme NAD (T)
c) allows synthesis of 3 ATP by reoxidation of its coenzyme in terminal oxidation (T)
d) uses product of fumarase as substrate (T)
221. Reaction cataylzed by malatedehydrogenase:
a) proceeds in mitochondria and cytosol (T)
b) is component of the shuttle by which 3 ATP are formed in mitochondria (T)
c) can proceed also in anaerobic conditions (F)
d) proceeds in all cells of human body (F)
222. Regulators of isocitrate dehydrogenase are:
a) citrate which inhibits the enzyme (F)
b) ADP which activates the enzyme (T)
c) ATP and NADH2 which inhibit the enzyme (T)
d) Glukagon which by phosphorylation inhibits the enzyme (F)
COOH
CH
CH
COOH COOH
CHHO
CH2
COOH

223. Isocitrate dehydrogenase in regulation of Krebs cycle:
a) represents key regulatory enzyme (T)
b) is activate by product of reaction – ketoglutarate (F)
c) is activated in good energy state (F)
d) is inhibited by ATP which causes change of the enzyme into inactive dimeric form (T)
224. By oxidation of acetyl-CoA in Krebs cycle:
a) ATP is formed only in aerobic conditions (T)
b) CO2 is formed in reaction of ketoglutarate synthesis (T)
c) reoxidation of NADH2 allows synthesis of 9 ATP (T)
d) 2 ATP can be formed in reaction of fumarate synthesis (T)
225. Oxidation of acetyl-CoA in Krebs cycle:
a) allows synthesis of 2 ATP by substrate level phsohprylation (F)
b) leads to synthesis of FADH2 in reaction catalyzed by fumarase (F)
c) allows synthesis of 11 ATO by oxidative phosphorylation (T)
d) leds to synthesis of 3 NADH2 (T)
226. Reaction of substrate level phosphorylation in Krebs cycle:
a) uses succinate as coenzyme (F)
b) uses as substrate compound with energy rich thioester bond (F)
c) proceeds in mitochondria only (T)
d) is catalyzed by isocitrate dehydrogenase (F)
227. Reaction of Krebs cycle where substrate level phosphorylation occurs:
a) proceeds in cytosol (F)
b) leads to synthesis of GTP (T)
c) uses as substrate compound which can be formed from amino acids – methionine, valine and
isoleucine (T)
d) proceeds only in aerobic conditions (T)
228. Reoxidation of reduced coenzymes formed in Krebs cycle allows synthesis of:
a) 12 ATP by oxidative phosphorylation (F)
b) 2 ATP from coenzym formed by succinate dehydrogenase (T)
c) 3 ATP from coenzyme of isocitrate dehydrogenase (T)
d) 1 ATP duringh convesrion of oxaloacetate to citrate (F)
229. NADH+H+ in Krebs cycle is formed:
a) in three reactions celkove v troch reakciách (T)
b) in reaction of succinyl-CoA synthesis (T)
c) in reaction of succinyl-CoA conversion (F)
d) in reaction of isocitrate conversion (T)
230. Coenzyme by oxidation of which 3 ATP can be formed in Krebs cycle is formed:
a) in reaction of isocitrate synthesis (F)
b) in reaction of succinate conversion (F)
c) in four reactions (F)
d) in reaction of oxaloacetate synthesis (T)
231. Coenzyme by oxidation of which 2 ATP can be formed in Krebs cycle is synthesized:
a) in reaction of ketoglutarate convesrion (F)
b) in reaction of succinate synthesis (F)
c) in reaction of fumarate synthesis (T)
d) in reaction of succinate conversion (T)
232. Enzymes of Krebs cycle:
a) are part of outer mitochondrial membrane (F)

b) are responsible for synthesis of acetyl-CoA (F)
c) are required only for ATP synthesis (F)
d) in anaerobic conditions they cannot act (T)
233. Enzyme complex required for conversion of pyruvate to acetyl-CoA:
a) is located in cytosol (F)
b) allows synthesis of 3 ATP (T)
c) is present in all cells of human body (F)
d) is required for utilization of pyruvate in gluconeogenesis (F)
234. Conversion of pyruvate into acetyl-CoA:
a) is catalyzed by pyruvate dehydrogenase (T)
b) is catalyzed by enzyme which consists of three subunits (T)
c) requires 5 coenzymes including biotin (F)
d) is irreversible reaction (T)
235. Pyruvate dehydrogenase catalyzes reaction which:
a) proceeds in mitochondria (T)
b) is required for complete oxidation of glucose (T)
c) allows conversion of glucose into fatty acids (T)
d) requires thiaminepyrophosphate as one of the coenzymes (T)
236. Lipoic acid in pyruvate dehydrogenase complex:
a) directly participates in decarboxylation of pyruvate (F)
b) participates in oxidation of acetaldehyde (T)
c) is reduced (T)
d) is oxidized by transfer of hydrogens to NAD (F)
237. Compound required for utilization of ketone bodies in extrahepatic tissues:
a) is formed in glycolysis (F)
b) is formed by isocitrate dehydrogenase (F)
c) is compound with thioester energy rich bond (T)
d) is palmitoyl-CoA (F)
238. In regulation of Krebs cycle participates:
a) ATP which activates citrate synthase (F)
b) ATP and NADH2 which inhibit isocitrate dehydrogenase (T)
c) glucagon which inhibits all enzymes of Krebs cycle (F)
d) ADP which changes isocitrate dehydrogenase into active ocamer (T)
239. Citrate synthase and isocitrate dehydrogenase are enzymes which:
a) belong to regulatory enzymes of Krebs cycle (T)
b) are located in mitochondria (T)
c) are regulated by ratio ATP/ADP (T)
d) are regulated by energy state of the cell (T)
240. High concentrations of ATP:
a) inhibit all enzymes of Krebs cycle (F)
b) inhibit phosphofructokinase I in glycolysis (T)
c) inhibit isocitrate dehydrogenase in Krebs cycle (T)
d) activate glucokinase in glycolysis (F)
LIPIDS
241. Cerebrosides:
a) contain sphingosine as alcoholic component (T)
b) contain fatty acid bound by –CO–NH– bond (T)

c) contain carbohydrate bound to alcohol by ester bond (F)
d) contain ologosaccharide (F)
242. Cerebrosides contain in their structure:
a) the same alcohol as lecithines (F)
b) fatty acid boun by bond –O–CO– (F)
c) pentoses (F)
d) carbohydrate bound to NH2 goup of sphingosine (F)
243. Ceramide is compound which:
a) belong to phospholipids (F)
b) is basic component of acylglycerols (F)
c) contains fatty acid bound by peptide bond (T)
d) is component of glycolipids (T)
244. Lipids with carbohydrate component:
a) are phospholipids (F)
b) contain ceramide (T)
c) contain fatty acid bound by peptide bond (T)
d) are for example cerebrosides and lecithines (F)
245. Sphingosine:
a) is alcohol present in all phospholipids (F)
b) is unsaturated C18 aminoalcohol (T)
c) is alcoholic component of acylglycerols (F)
d) contain NH2 group to which fatty acid is bound (T)
246. C18 unsaturated aminoalcohol:
a) is inositol (F)
b) is component of sphingomyelins (T)
c) is component of gangliosides (T)
d) is alcoholic component of lipids in adipose tissue (F)
247. Compound:
a) is sphingomyelin (F)
b) belongs to phospholipids (F)
c) is main component of adipose tissue (F)
d) does not exist (T)
248. Sphingomyelins are lipids which:
a) belong to phospholipids (T)
b) are present in nervous tissue (T)
c) contain two phosphate residues bound to OH groups of sphingosine (F)
d) contain choline as non-lipidic component (T)
249. Phosphatidylinositols contain in their molecula:
a) two fatty acids (T)
b) phosphate bound to 1st or 2nd carbon (F)
c) cyclic alcohol inositol (T)
d) choline (F)
250. Lecithines:
CH3 (CH2)12 CH
O
NH2
CH CH2CH CH
C O
R
O CH2 CH2 N
CH3
CH3
CH3PO
O-
O

a) contain two fatty acids bond to OH groups of glycerol (T)
b) contain serine as non-lipidic component (F)
c) contain polar and non-polar part in their molecules (T)
d) are important component of plasma membranes (T)
251. Fatty acid in sphingomyelins is bound:
a) to phosphate (F)
b) by –CO–NH– bond (T)
c) by the same bond as fatty acids are bound in acylglycerols (F)
d) by the bond which is present for example in glugagon (T)
252. Alcoholic component of cerebrosides:
a) is sphingosine (T)
b) is formed form palmitoyl-CoA and serine (T)
c) binds fatty acid to its OH group (F)
d) is glycerol (F)
253. During activation of palmitic acid:
a) palmitoyl-CoA and ATP are substrate (F)
b) as intermediate acyladenylate is formed (T)
c) two energy rich bonds are spent (T)
d) enzyme present in inner mitochondrial membrane is required (T)
254. Activation of fatty acids:
a) is required for synthesis of fatty acids (F)
b) is catalyzed by acyl-CoA synthase (T)
c) requires GTP as source of energy (F)
d) ako zdroj energie využíva ATP, ktorý sa pri aktivácii VKK štiepi na ADP (F)
255. Activation of fatty acids can be catalyzed by:
a) enzyme system which uses ATP as source of energy (T)
b) fatty acid synthase (F)
c) acyl-CoA synthase (T)
d) CoA transferase in case of short chain fatty acids (T)
256. During activation of fatty acids before -oxidation:
a) ATP is broken into ADP (F)
b) compound with acylphosphate energy rich bond is formed as intermediate (T)
c) fatty acid is changed to acyl-CoA (T) väzby v ATP (F)
d) two ebergy rich bonds are spent (T)
257. Activated fatty acids are transported into mitochondria by help of:
a) CoA (F)
b) ATP (F)
c) active transport (F)
d) compound to which fatty acid is bound by ester bond (T)
258. For transport of activated fatty acid into mitochondria is required compound which:
a) contains OH group (T)
b) contains CH3 groups (T)
c) cannot be synthesized in human body (F)
d) binds fatty acid by help of carnitine acyltransferase I (T)
259. Carnitine is compound which:
a) is peptide required for transport of fatty acids (F)
b) is required for tranport of fatty acids in blood (F)
c) is synthesized form two essential amino acids (T)

d) binds fatty acid by peptide bond (F)
260. Carnitineacyltransferase I:
a) is located in cytosol (F)
b) uses carnitine and fatty acid as substrates (F)
c) catalyzes transport of carnitine into mitochondria (F)
d) is activated by malonyl-CoA (F)
261. Carnitineacyltransferase I is enzyme which:
a) is present in outer side of inner mitochondrial membrane (T)
b) uses acyl-CoA and carnitine as substrates (T)
c) catalyzes transfer of fatty acid residue to COOH group of carnitine (F)
d) is inhibited by malonyl-CoA (T)
262. Carnitineacyltransferase II is enzyme which:
a) is located in mitochondrial matrix (F)
b) catalyzes transfer of fatty acid residue form acylcarnitine to CoA (T)
c) participates in transport of activated fatty acid into mitochondria (T)
d) requires ATP for its activity (F)
263. Trnaport of activated fatty acids into place of -oxidation:
a) is not important because fatty acid are activated in mitochondria (F)
b) requires NAD as coenzyme (F)
c) requires carnitine (T)
d) is inhibited by malonyl-CoA (T)
264. Activated fatty acid is transported into mitochondria by:
a) active transport (F)
b) help of citrate (F)
c) CoA (F)
d) system which requires carnitineacyltransferase I and carnitineacyltransferase II (T)
265. Compound:
a) is carnitine (T)
b) requires methionine for its synthesis (T)
c) is required for transport of activated fatty acids for mitochondria into cytosol (F)
d) by reaction with acyl-CoA gives acylcarnitine (T)
266. Reaction of -oxidation which allows synthesis of 2 ATP by oxidative phosphorylation:
a) is reaction of conversion of acyl-CoA to enoyl-CoA (T)
b) uses coenzyxme derived form vitamion B2 (T)
c) is catalyzed by enoyl-CoA hydratase (F)
d) is reaction in which substrate fopr ketothiolase is formed (F)
267. Adition of water in -oxidation:
a) leads to synthesis of -hydroxyacyl-CoA (T)
b) is catalyzed by enoyl-CoA hydratase (T)
c) uses product of acyl-CoA dehydrogenase as substrate (T)
d) allows synthesis of 3 ATP by oxidative phosphorylation (F)
268. Reaction of -oxidation which allows synthesis of 3 ATP by oxidative phosphorylation is:
a) reaction of enoyl-CoA synthesis (F)
b) reaction of enoyl-CoA conversion (F)
c) reaction of convesrion of product of enoyl-CoA hydratase (T)
d) reaction which leads to synthesis of -ketoacyl-CoA (T)
H3C N CH2
CH3
CH3
C
H
OH
CH2 COOH
++

269. Synthesis of 3 ATP from coenzyme formed in -oxidatiopn allows:
a) reaction catalyzed by acyl-CoA dehydrogenase (F)
b) reaction of conversion of enoyl-CoA to -hydroxyacyl-CoA (F)
c) reaction of -keto-acyl-CoA synthesis (T)
d) of enoyl-CoA synthesis (F)
270. Dehydrogenation reactions in -oxidation are catalyzed by:
a) acyl-CoA dehydrogenase with FAD as coenzyme (T)
b) enzyme which produces enoyl-CoA (T)
c) enoly-CoA hydratase (F)
d) -hydroxyacyl-CoA dehydrogenase with NADF as coenzyme (T)
271. During oxidation of palmitic acid in -oxidation:
a) cycle of -oxidation proceeds 7 times (T)
b) 7 NADH+H+, 7 FADH2 and 8 acetyl-CoA are formed (T)
c) by oxidation of acetyl-CoA formed in -oxidation 88 ATP are formed (F)
d) ATP is formed in mitochondria (T)
272. During complete degradation of palmotoyl-CoA in -oxidation are formed:
a) 96ATP by oxidation of acetyl-CoA in Krebs cycle (T)
b) 21 ATP from coenzyme of acyl-CoA dehydrogenase (F)
c) 14 ATP in reaction of acetyl-CoA synthesis (F)
d) 21 ATP form coenzyme of -ketoacylthiolase (F)
273. During -oxidation of fatty acids with odd number of carbons as the last product is formed:
a) propionyl-CoA (T)
b) malonyl-CoA (F)
c) compound for conversion of which vitamin B12 is required (T)
d) compound which is changed to succinyl-CoA (T)
274. During oxidation of stearic acid in -oxidation:
a) 9 acetyl-CoA are formed (T)
b) cycle of -oxidation proceeds 9 times (F)
c) by reoxidation of coenzymes formed in -oxidation 35 ATP are formed (F)
d) comúpound which can be used for gluconeogenesis is formed (F)
275. Complete oxidation of stearic acid allows:
a) synthesis of 146 ATP (T)
b) synthesis of 129 ATP (F)
c) synthzesis of 40 ATP vy reoxidation of coenzymes formed in -oxidation (T)
d) synthesis of less ATP than oxidation of myristic acid (F)
276. Process of oxidation of myristic acid:
a) produces 7 NADH2 and 7 FADH2 (F)
b) leads to synthesis of 7 acetyl-CoA (T)
c) allows synthesis of 112 ATP (T)
d) for activation of myristic acid requires 1 energy rich bond (F)
277. Acetyl-CoA in -oxidation is formed:
a) from substrate acyl-CoA (F)
b) in reaction which uses NAD as coenzyme (F)
c) form substrate wghich is product of enoyl-CoA hydratase (F)
d) in reaction which uses water as one of substrates (F)
278. Synthesis of acetyl-CoA in -oxidation:

CH2
C
COOH
CH2
HO
COOH
COOH
a) is catalyzed by -ketothiolase (T)
b) requires CoA as one of substrates of reaction (T)
c) requires -hydroxyacyl-CoA as substrate (F)
d) can proceed only in aerobic conditions (T)
279. Acyl CoA dehydrogenase:
a) catalyzes reaction
(F)
b) uses as coenzyme FAD (T)
c) catalyzes conversion of katalyzuje reakciu premeny
(F)
d) catalyzes reaction which allows synthesis of 3 ATP (F)
280. Compound
a) by oxidation in Krebs cycle allows synthesis of 12 ATP (T)
b) during -oxidation of product of -ketoacylthiolase (T)
c) for fatty acids synthesis is transported from mitochondria into cytosol in the form of compound
(T)
d) is substrate for ketone bodies synthesis (T)
281. Reaction:
a) is reaction of fatty acid activation (T)
b) proceeds in inner mitochondrial membrane (F)
c) requires energy of one energy rich bond (F)
d) leds to synthesis of product which is transported into mitochondria by help of carnitine (T)
282. EnoylCoA hydratase:
a) catalyzes reaction of conversion of enoyl-CoA into (T)
b) leads to synthesis of product which is changed by reaction
(T)
c) uses FAD as coenzyme (F)
d) uses as substrate product of acyl-CoA dehydrogenase (T)
283. -oxidation of fatty acids:
a) is important source of energy in myocardium (T)
R CH CH2 C
O
S-CoA
R C CH2 C
O
S-CoAOOHFAD FADH2
R CH2 CH2 CO
S-CoA
R CH CH2 C
O
S-CoAOH
na
CH3 C
O
~S CoA
R COOH + HS CoA + ATP R C
O
~S CoA + AMP + PPi
Mg2+
, K+
R CH CH2 C
O
S-CoAOH
R CH CH2 C
O
S-CoA
R C CH2 C
O
S-CoAOOHNAD NADH2

b) proceeds also in anaerobic conditions (F)
c) requires reduced form of NAD (F)
d) is activated by insulin (F)
284. -oxidation of fatty acids:
a) requires aerobic conditions (T)
b) is located in endoplasmic reticulum (F)
c) is main source of energy for brain (F)
d) requires reqoxidation of reduced coenzymes in terminal oxidation (T)
285. Acetyl-CoA in synthesis of fatty acids is formed in cytosol:
a) from acylcarnitine (F)
b) from palmitate (F)
c) from citrate (T)
d) from compound which is formed in mitochondria (T)
286. Acetyl-Co required for fatty acids synthesis is formed:
a) in cytosol by pyruvate dehydrogenase (F)
b) in mitochondria from where is transported into cytosol in the form of citrate (T)
c) in cytosol by the enzyme ATP-ciatrate lyase (T)
d) in cytosol by the process of -oxidation (F)
287. Citrate synthase and ATP-citrate lyase are enzymes which:
a) both are required for fatty acid synthesis (T)
b) both are present in mitochondria (F)
c) both are present in cytosol (F)
d) both are activated by ATP (F)
288. ATP citrate lyase and citrate synthase are enzymes which:
a) participate in oxidation of fatty acids (F)
b) are required for transport of acetyl-CoA form mitochondria into cytosol (T)
c) are required for transport of fatty acids into mitochondria (F)
d) are inhibited by ATP (F)
289. Carboxylation of acetyl-CoA to malonyl-CoA:
a) uses substrates acetyl-CoA and CO2 (T)
b) uses pyridoxalphosphate as coenzyme (F)
c) proceeds in mitochondria (F)
d) is catalyzed by enzyme which is active in dephosphorylated form (T)
290. Synthesis of malonyl-CoA in synthesis of fatty acids:
a) proceeds in cytosol (T)
b) is regulatory step in fatty acids synthesis (T)
c) is catalyzed by acetyl-CoA carboxylase (T)
d) is activated by glucagon during starvation (F)
291. -hydroxyacyl-ACP in fatty acid synthesis is formed:
a) from acetyl-CoA and malonyl-CoA as substrates (F)
b) form -ketoacyl-ACP by oxidation (F)
c) by the action of condensing enzyme (F)
d) by addition of water (F)
292. Synthesis of -hydroxyacyl-ACP requires:
a) substrate -ketoacyl-ACP (T)
b) coenzyme formed in pentose phosphate pathway (T)
c) coenzyme FADH2 (F)
d) -ketoacylreductase (T)

CH2 COOH
CHO COOH
CH2 COOH
293. Regulatory step in fatty acid synthesis is:
a) reaction which requires biotine (T)
b) reaction in which methylmalonyl-CoA is formed (F)
c) reaction where acetyl-CoA is carboxylated (T)
d) reaction which is catalyzed by citrate synthase (F)
294. Enzmye which plays key role in regulation of fatty acid synthesis is:
a) acetyl-CoA carboxylase (T)
b) enzyme which requires ATP (T)
c) enzyme by action of which malonyl-CoA is formed (T)
d) enzyme which is activated during starvation (F)
295. Reaction:
a) is the first step in fatty acid synthesis (T)
b) is catalyzed by acetyl-CoA carboxylase which is active in dephosphorylated form (T)
c) uses thiamine as coenzyme (F)
d) is activated by citrate (T)
296. Synthesis of fatty acids:
a) proceeds in mitochondria (F)
b) uses as substrate CH3-CO~S-CoA which is formed by oxidative decarboxylation of pyruvate in
cytosol (F)
c) requires NADH2 (F)
d) uses for transport of CH3-CO~SCoA from mitochondria into cytosol
(T)
297. Synthesis of fatty acids is the process about which we can say:
a) proceeds in cytosol (T)
b) is activated by insulin (T)
c) actetyl-CoA as substrate for fatty aci synthesis is formed in cytosol by reaction
(T)
d) reduced coenzymes required for fatty aci synthesis are formed mainly in glycolysis and Krebs
cycle (F)
298. Reaction of fatty acid synthesis:
a) uses coenzyme formed in pentose phosphate pathway (T)
b) is catalyzes by enoyl-CoA hydratase (F)
c) is reaction of mitoíchondrial elongation of fatty acids (T)
d) is activated during starvation by glucagon (F)
299. Malic enzyme is enzyme which:
a) is responsible for synthesis of malate (F)
b) is responsible for convesrion of malate to pyruvate (T)
CH3 CO S CoA CH2 COHOOC S CoA+ HCO3- + ATP + ADP + P i~ ~
CH2 COOH
CHO COOH
CH2 COOH
+ ATP
HS-CoA
CH3 CO S CoA~ +
COOH
CO
CH2
COOH
CH3 (CH2)12 CH CH CO S ACP~ + NADPH+H+
CH3 (CH2)14 CO S ACP~ + NADP+

c) is required for gluconeogenesis (F)
d) is component of shuttle for transport of reducing equivalents into mitochondria (F)
300. Malonyl-CoA is compound which:
a) is formed form acetyl-CoA (T)
b) is formed by acetyl-CoAS carboxylase (T)
c) is formed in reaction which requires biotine (T)
d) regulates of process of -oxidation by activation of carnitineacyltransferase I (F)
301. Acetyl-CoA carboxylase catalyzes reaction which:
a) is responsible for acyl-CoA synthesis (F)
b) requires pyridoxalphosphate as coenzyme (F)
c) participates in fatty acid synthesis (T)
d) is activated by insulin (T)
302. Reaction of malonyl-CoA synthesis:
a) is regulatory step of fatty acid synthesis (T)
b) is activated by citrate (T)
c) requires biotin (T)
d) is inhibited by phosphorylation of enzyme in presence of glucagon (T)
303. Acetyl-CoA carboxylase:
a) participates in degradation of fatty acids (F)
b) is located in cytosol (T)
c) leads to synthesis of malonyl-CoA (T)
d) is inhibited by citrate (F)
304. Acetyl-CoA carboxylase is enzyme which:
a) participates in synthesis of fatty acids (T)
b) is inhibited by malonyl-CoA (T)
c) uses citrate as substrate (F)
d) is activated by phosphorylation in presence of insulin (F)
305. Source of hydrogens for reduction reactions of fatty acid synthesis is:
a) coenzym containing nicotinamide (T)
b) coenzyme NAD (F)
c) coenzyme formed by malic enzyme (T)
d) coenzyme which is also donor of hydrogens for terminal oxidation (F)
306. Condensing enzyme in fatty acid synthesis catalyzes reaction which:
a) uses as substrates 2 mol of malonyl-CoA (F)
b) leads to synthesis of -ketoacyl-ACP (T)
c) is main regulatory step of fatty acid synthesis (F)
d) is located in mitochondria (F)
307. -ketoacylreductase:
a) is enzyme of fatty acid oxidation (F)
b) uses as coenzyme compound formed in cytosol (T)
c) is responsible for synthesis of enoyl-ACP (F)
d) is enzyme activated during starvation (F)
308. -hydroxyacyl-ACP is changed to enoyl-ACP:
a) by dehydrataion (F)
b) by oxidation (F)
c) by reaction which requires water as substrate (F)
d) by reaction which requires NADPH2 as coenzyme (F)

309. Fatty acid synthase:
a) is multienzyme complex located in cytosol (T)
b) consists of three catalytic subunits (F)
c) synthesizes fatty acids with nmaximal length 16 carbons (T)
d) is main regulatory enzyme of fatty aciod synthesis (F)
310. Thioesterase is the enzyme which:
a) is component of fatty acid synthase (T)
b) is component of multienzyme complex located in cytosol (T)
c) is responsible for cleavage of -keto-acyl-CoA (F)
d) is enzyme of -oxidation (F)
311. Elongation of fatty acids:
a) proceeds in cytosol (F)
b) proceeds in mitochondria and smooth endoplasmic reticulum (T)
c) is required because fatty acid synthase cannot synthesize fatty acids longer than 16 carbons (T)
d) uses NADH2 and FADH2 as coenzymes (F)
312. Desaturation of fatty acids:
a) can change stearic acid into oleic acid (T)
b) proceeds in mitochondria (F)
c) uses specific cytochrome – cyt b5 (T)
d) allows synthesis of essential fatty acids (T)
313. NADPH2 in the process of synthesis of fatty acids:
a) is formed for example by glucose-6-phosphate dehydrogenase (T)
b) can be formed in cytosol by oxidation of isocitrate (T)
c) is required for condensing enzyme (F)
d) is required for conversion of enoyl-ACP into acyl-ACP (T)
314. Which of following reactions can be source of NADPH2 for synthesis of fatty acids:
a) mitochondrial malate dehydrogenase (F)
b) cytosolic malic enzyme (T)
c) 6-phosphogluconate dehydrogenase (T)
d) HMG-CoA reductase (F)
315. Fatty acid synthesis is regulated by:
a) insulin which causes dephosphorylation of acetyl-CoA carboxylase and its inhibition (F)
b) insulin which causes dephosphorylation of acetyl-CoA carboxylase and its activation (T)
c) citrate which acts as direct activator of acetyl-CoA carboxylase (T)
d) glucagon which during starvation activates synthesis of fatty acids (F)
316. Which statements about role of hormones in regulation of fatty acid synthesis are correct:
a) glucagon directly activates ATP-citrate lyase (F)
b) insulin causes dephosphorylation of acetyl-CoA carboxylase and thus its activation (T)
c) hormone secreted during starvation inhibits fatty acid synthesis by change acetyl-CoA carboxylase
into inactive – phosphorylated form (T)
d) adrenaline causes phosphorylation of acetyl-CoA carboxylase and activates fatty acid synthesis
during stress (F)
317. Which statements about role of glucagon in regulation of fatty acids synthesis are correct:
a) glucagon secreted during starvation stimulates transport of glucose into the cell and by this way
inderectly activates fatty acid synthesis (F)
b) glucagon increases synthesis of cAMP and activates acetyl-CoA carboxylase by its
phosphorylation (F)
c) glucagon increases synthesis of cAMP and causes inhibition of acetyl-CoA carboxylase by its
dephosphorylation (F)

CH2
CH
O
O
CH2 O
CO R1
CO R2
CO R3
d) glucagon increases cAMP and activates ATP-citrate lyase by its phosphorylation (F)
318. Which statements about regulatrion of fatty acid synthesis are correct:
a) citrate act as activator of acetyl-CoA carboxylase (T)
b) adrenaline activates fatty acid synthesis (F)
c) palmitoyl-CoA acts like inhibitor of acetyl-CoA carboxylase (T)
d) main regulatory enzyme of fatty aciod synthesis – acetyl-CoA carboxylase is activated by insulin
(T)
319. Which of processes activated by insulin is important for conversion of glucose into fatty acids:
a) activation of hormone sensitive lipase (F)
b) activation of acetyl-CoA (T)
c) activation of glycolysis by increased synthesis of fructose-2,6-bisphosphate (T)
d) activation of ATP-citrate lyase (T)
320. is compound which:
a) represents storage form of energy in liver (F)
b) in lumen of intestine is broken by lipoprotein lipase (F)
c) is formed in adipose tissue (T)
d) in adipose tissue is hyrolyzed by hormone sensitive lipase (T)
321. Which cells of human body synhesize and store triacylglycerols:
a) all cells without exception (F)
b) liver cells (F)
c) adipose tissue cells (T)
d) enterocytes (F)
322. For triacylglycerol synthesis in is used:
a) free glycerol (F)
b) active form of glycerol – glycerolphosphate (T)
c) CTP (F)
d) activated fatty acids in the form of acylcarnitine (F)
323. Alcoholic component of triacylglycerols in adipose tissue:
a) enters synthesis like free glycerol (F)
b) enters synthesis like glycerol-phosphate (T)
c) can be formed form free gylcerol by glycerolkinase (F)
d) can be formed form intermediate of glycolysis dihydroxyacetone phosphate (T)
324. In synthesis of triacylglycerols:
a) fatty acids in their active form – acyl-CoAS are required (T)
b) enzyme phosphatidate phosphatase is required (T)
c) phosphatidic acid as intermediate is formed (T)
d) source of alcoholic component in liver can be free glycerol (T)
325. Hormone sensitive lipase is the enzyme which:
a) is responsible for hydrolysis of TAG from food (F)
b) is responsible for hydrolysis of TAG in adipose tissue (T)
c) is activated during stavation by glucagon (T)
d) in presence of insulin is in inactive – phosphorylated form (F)
326. Degradation of triacylglycerols in adipose tissue:
a) is catalyzed by lipoprotein lipase (F)
b) is catalyzed by enzyme which cleaves TAG into fatty acids and glycerol (T)

HO
c) is inhibited by insulin which changes hormonesensitive lipase into inactive – dephosphorylated
form (T)
d) is activated during stress by adrenalin which changes hormone sensitive lipase into active –
phosphorylated form (T)
327. In synthesis of lecithins:
a) non-lipidic component – choline is formed form serine (T)
b) choline is activated by choline kinase (T)
c) CTP is required as source of energy (T)
d) CDP-choline is formed as intermediate (T)
328. CDP-diacylglycerol is compound which:
a) is intermediate of synthesis of glycolipids (F)
b) is formed form phosphatidic acid and CTP (T)
c) by reaction with inositol gives phosphatidylinositols (T)
d) contains phosphoanhydride energy rich bond (T)
329. Synthesis of cholesterol in human body:
a) is not important because cholesterol in taken by food (F)
b) proceeds in cytosol (T)
c) uses malonyl-CoA as substrate (F)
d) is activated by glucagon during starvation (F)
330. In the process of cholesterol synthesis:
a) acetoacetyl-CoA is formed as intermediate (T)
b) by action of HMG-CoA reductase mevalonate is formed (T)
c) reduced coenzyme formed in Krebs cycle is required (F)
d) ATP is required as source of energy (T)
331. is compound which:
a) is precursor of biologically important coompounds (bile acids, vitamin D, steroid hormones (T)
b) is synthesized in the process part of which is the reactio
(T)
c) is synthesized in cytosol from acetyl-CoA (T)
d) requires NADPH2 in the process of synthesis (T)
332. HMG-CoA during cholesterol synthesis is changed:
a) to mevalonate (T)
b) by HMG-CoA reductase (T)
c) by enzyme present in cytosol (T)
d) by enzyme which is active in dephosphorylated form (T)
333. Mevalonic acid is compound which:
a) contains two COOH groups (F)
b) is formed by oxidation of HMG-CoA (F)
c) is formed by reaction which uses NADPH2 as coenzyme (T)
CH3 C
OH
CH2 C
O
S CoACH2 COOH
CH3 C
OH
CH2 CH2
CH2 COOH
OH
HS-CoA
2 NADPH2 2 NADP+

d) is formed in reaction which is key regulatory step of cholesterol synthesis (T)
334. Mevalonic acid in cholesterol synthesis:
a) is formed from immediate precursor acetoacetyl-CoA (F)
b) is called also active isoprene (F)
c) is formed in cytosol (T)
d) is formed by HMG-CoA reductase (T)
335. HMG-CoA reductase is enzyxme which:
a) is responsible for synthesis of HMG-CoA (F)
b) catalyzes decarboxylation of HMG-CoA (F)
c) is main regulatory enzyme of cholesterol synthesis (T)
d) is activated by high concentration of cholesterol (F)
336. Which of statements about regulation of cholesterol synthesis are correct:
a) cholesterol inhibits reaction of mevalonic acid synthesis (T)
b) cholesterol activates HMG-CoA reductase (F)
c) regulatory enzyme is active in dephosphorylated form (T)
d) hormone glucagon causes inhibition of cholesterol synthesis by phosphorylation of HMG-CoA
reductase (T)
337. Chylomicrons:
a) are lipoproteins formed in the liver (F)
b) from enterocytes are secreted into lymph (T)
c) in blood are degraded by lipoprotein lipase (T)
d) contain apoprotein C (T)
338. Apoprotein A:
a) is important component of HDL (T)
b) acts like activator of lipoprotein lipase (F)
c) is formed by liver cells (T)
d) is activator of LCAT (T)
339. Apoprotein B:
a) is main protein component of all lipoproteins (F)
b) is activator of liporotein lipase (F)
c) is main protein component of HDL (F)
d) is important for binding of LDL to specific receptors (T)
340. Chylomicrons are lipoproteins which:
a) are formed in intestine (T)
b) are responsible for transport of TAG from liver to cells of EHT (F)
c) are degraded by lipoprotein lipase (T)
d) require apoprotein C for their degradation (T)
341. VLDL are lipoproteins which:
a) are formed in liver (T)
b) transport mainly cholesterol (F)
c) contains around 90% of lipids (T)
d) in blood are degraded by lipoprotein lipase (T)
342. VLDL:
a) are lipoprotein transporting exogenous triacylglycerols (F)
b) are lipoproteins with the lowest density (F)
c) contain around 40% of proteins (F)
d) in blood are changed to LDL by lipoprotein lipase (T)

343. Apoprotein C:
a) is main component of LDL (F)
b) is component of HDL where is required for action of LCAT (F)
c) is activator of lipoprotein lipase (T)
d) is required for catabolism of VLDL (T)
344. Lipoprotein lipase is the enzyme which:
a) is responsible for degradation of TAG from food (F)
b) is typical for hepatocytes (F)
c) is responsible for degradation of cholesterol in chylomicrons (F)
d) catalyzes hydrolysis of TAG in adipose tissue during starvation (F)
345. Lipoprotein lipase:
a) is enzyme bound to external surface of endothelial cells in capillaries (T)
b) is responsible for conversion of LDL to VLDL (F)
c) is responsible for degradation of chylomicrons (T)
d) in adipose tissue is activated by insulin (T)
346. Lipoprotein lipase is enzyme which:
a) is in high activities in intestine (F)
b) hydrolyzes tAG into glycerol and fatty acids (T)
c) is activated by apoprotein B (F)
d) is responsible for conversion of VLDL to LDL (T)
347. LDL are lipoproteins which:
a) are formed in intestine (F)
b) are formed in blood during degradation of vLDL by lipoprotein lipase (T)
c) are formed in liver by degradation of chylomicrons (F)
d) in high concentrations represent risk factor for evelopment of atherosclerosis (T)
348. LDL:
a) are formed in blood from HDL (F)
b) are responsible for transport of cholesterol from cells to the liver (F)
c) contain apoprotein B as main protein component (T)
d) are degraded in blood by lipoprotein lipase (F)
349. HDL are lipoproteins which:
a) are formed ih hepatocytes (T)
b) contain the highest amount of proteins (T)
c) are responsible for transport of cholesterol into blood vessel wall (F)
d) are source of apoprotein C for VLDL and chylomicrons (T)
350. HDL are lipoproteins which:
a) are formed in adipocytes (F)
b) contain the higest amount of phospholipids (T)
c) contains enzyme LCAT (T)
d) transport cholesterol from cells to the liver (T)
351. Which of statements about HDL are correct:
a) they are formed by cells of adipose tissue (F)
b) they are fored by liver in their nascent form (T)
c) they decrease risk of atherosclerosis (T)
d) they are metabolized by lipoprotein lipase (F)
352. Lecithine-cholesterol acyltransferase (LCAT) is the enzyme which:
a) is important for metabolism of chylomicrons (F)
b) is important for reverse cholesterol transport by HDL (T)

c) is activated by apoprotein A (T)
d) hydrolyzes cholesterol esters (F)
353. Lecithine-cholesterol acyltransferase (LCAT):
a) is formed by liver (T)
b) is required for transport of LDL into the cells of EHT (F)
c) catalyzes synthesis of cholesterolô esters (T)
d) uses as substrate lecithins which are components of plasma membranes (F)
354. Which of following mechanisms participate in regulation of cholesterol content in cells:
a) inhibition of LCAT by cholesterol (F)
b) inhibition of HMG-CoA reductase by high cholesterol cioncentration (T)
c) inhibitionn of LDL receptor synthesis by lyck of cholesterol (F)
d) activation of synthesis of receptors for lDL in low cholesterol concentration (T)
355. When intracellular concentration of cholesterol increases:
a) activity of HMG-CoA reductase increases (F)
b) synthesis of receptors for LDL is inhibited (T)
c) transport of HDL to the cells increases (F)
d) oxidation of cholesterol is activated (F)
NUCLEOTIDES
356. Nucleotides are compounds which:
a) are main component of membranes (F)
b) are used as sources of energy – for example UTP in gluconeogenesis (F)
c) participate in regulation of metabolic processes (T)
d) organism takes mainly form fodd (F)
357. Nucleotides in human body:
a) are required in activation of substrates (T)
b) are importnat in transfer of genetic information (T)
c) are component of coenzymes – NAD, FAD (T)
d) are are important sorces of energy – for example GTP for proteosynthesis (T)
358. Source of ribosephosphate in synthesis of purine nucleotides de novo is:
a) ribose-5-phosphate (F)
b) PRPP (T)
c) compound formed by reaction of ribose-5-phosphate and ATP (T)
d) compound formed by PRPP-amidotransferase (F)
359. PRPP is compound which:
a) is formed by PRPP synthase (T)
b) contains two phosphates bound to 5th carbon of ribose (F)
c) is required for synthesis of IMP by HGPRT (T)
d) is substrate for PRPP amidotransferase in de novo synthesis of purine nucleotides (T)
360. PRDP-amidotransferase catalyzes reaction which:
a) uses as substrates PRPP and essential aminoacid (F)
b) leads to synthesis of phosphoribosylamine (T)
c) is main regulatory step in purine nucleotide synthesis de novo (T)
d) is inhibited by final products – AMP, GMP and IMP (T)
361. Reaction of phosphoribosylamine synthesis:
a) requires PRPP and glutamine as substrates (T)
b) is catalyzed by pRPP-aminotransferase (F)
c) is inhibited by AMP, CMP and UMP (F)
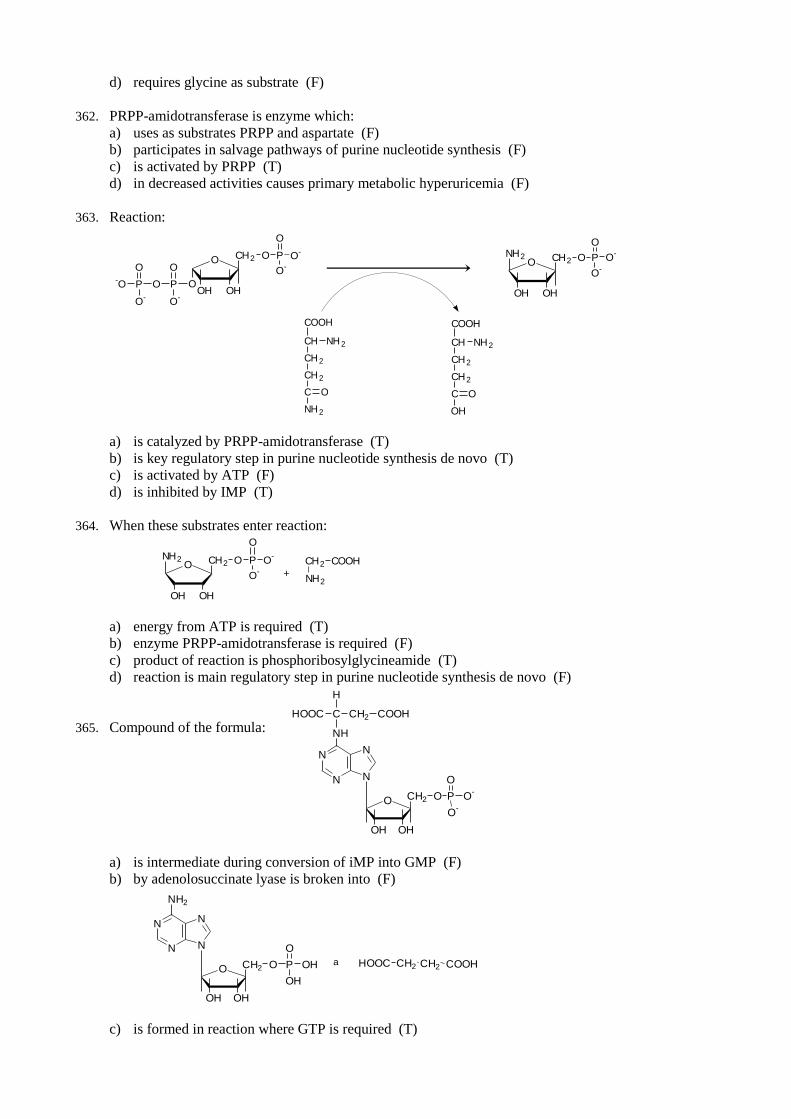
d) requires glycine as substrate (F)
362. PRPP-amidotransferase is enzyme which:
a) uses as substrates PRPP and aspartate (F)
b) participates in salvage pathways of purine nucleotide synthesis (F)
c) is activated by PRPP (T)
d) in decreased activities causes primary metabolic hyperuricemia (F)
363. Reaction:
a) is catalyzed by PRPP-amidotransferase (T)
b) is key regulatory step in purine nucleotide synthesis de novo (T)
c) is activated by ATP (F)
d) is inhibited by IMP (T)
364. When these substrates enter reaction:
a) energy from ATP is required (T)
b) enzyme PRPP-amidotransferase is required (F)
c) product of reaction is phosphoribosylglycineamide (T)
d) reaction is main regulatory step in purine nucleotide synthesis de novo (F)
365. Compound of the formula:
a) is intermediate during conversion of iMP into GMP (F)
b) by adenolosuccinate lyase is broken into (F)
c) is formed in reaction where GTP is required (T)
O
OH OHO
CH2 O P O-
O-
O
O P
O-
O
P-O
O-
OO
OH OH
NH2 CH2 O P O-
O-
O
NH2CH
COOH
CH2
CH2
C
NH2
O
NH2CH
COOH
CH2
CH2
C
OH
O
O
OH OH
NH2 CH2 O P O-
O-
O
NH2
CH2 COOH
+
N
N N
N
NH
O
OH OH
CH2 O P O-
O-
O
CHOOC CH2 COOH
H
N
N N
N
NH2
O
OH OH
CH2 O P OH
OH
O
COOHCH2CH2HOOCa

NH2
CHHOOC CH2 CH2 C NH2
O
d) is formed by enzyme which is inhibited by AMP (T)
366. Further conversion of this compound in synthesis of
purine nucleotides:
a) requires (T)
b) leads to synthesis of AMP (F)
c) requires ATP as source of energy (T)
d) is inhibited in deficiency of tetrahydrofolate (F)
367. Synthesis of purine nucleotides de novo is the process which:
a) uses ATP as main source of energy (T)
b) requires glutamine as source of two nitrogen atoms in purine heterocycle (T)
c) can be decreased in folic acid deficiency (T)
d) proceeds only in dividing cells (F)
368. For conversion of IMP to AMP:
a) aspartate is required (T)
b) adenylosuccinate is fornmed as intermediate (T)
c) GTP is used as source of energy (T)
d) adenylosuccinate lyase is required (T)
369. Source of energy in conversion of IMP into AMP is:
a) ATP (F)
b) GTP (T)
c) CTP (F)
d) compound formed by substrate level phosphorylation in Krebs cycle (T)
370. Source of NH2 group in AMP:
a) is non-essential amino acid (T)
b) is aspartate (T)
c) is glutamate (F)
d) is compound required also for pyrimidine nucleotide synthesis (T)
371. In conversion of IMP into GMP:
a) aspartate is required as source of NH2 group (F)
b) GTP is required as source of energy (F)
c) ATP is required as source of energy (T)
d) IMP dehydrogenase is required (T)
372. GMP in de novo synthesis of purine nucleotides is formed:
a) from immediate precursor IMP (F)
b) from immediate precursor XMP (T)
c) by reaction which requires glutamine as source of NH2 group (T)
d) by reaction which requires GTP as source of energy (F)
373. For synthesis of GMP from IMP:
a) IMP dehydrogenase is required (T)
b) xanthine oxidase is required (F)
c) glutamate is required as source of NH2 group (F)
HN
NH
N
O
O N
O
OH
CH2
OH
O P O-
O
O-

d) ATP is required as source of energy (T)
374. Source of NH2 group in synthesis of GMP de novo is:
a) aspartate (F)
b) glutamate (F)
c) essential amino acid (F)
d) compound which is also source of NH2 group fpr synthesis of AMP (F)
375. Source of energy in conversion of IMP into GMP is:
a) ATP (T)
b) GTP (F)
c) compound which is also required for synthesis of adenylosuccinate (F)
d) compound which is formed by oxidative phosphorylation in mitochondria (T)
376. Nitrogen atoms in purine heterocycle:
a) all come from glycine (F)
b) come from THF (F)
c) two come for glutamine (T)
d) one comes form aspartate (T)
377. Which of statements about origin of atoms in purine heterocycle are correct:
a) glycine is source of two carbons in purine ring (T)
b) glycine is source of one nitrogen in purine ring (T)
c) aspartate is source of two nitrogens in purine rong (F)
d) tetrahydrofolate is source of two carbons (T)
378. Main regulatory enzyme of purine nucleotide syntheisi de novo is:
a) PRPP amidotransferase (T)
b) enzyme which uses PRPP and glutamine as substrates (T)
c) IMP dehydrogenase (F)
d) enzyme which is activated by AMP (F)
379. Key regulatory enzyme of purine nucleotide synthesis de novo is:
a) PRPP aminotransferase (F)
b) enzyme which produces phosphoribosylamine (T)
c) enzyme which is activated by ATP (F)
d) enzyme which is inhibited by AMP, GMP and IMP (T)
380. Activity of PRPP-amidotransferase is influenced by:
a) AMP and GMP as inhibitors (T)
b) GTP and ATP as activators (F)
c) PRPP as activator (T)
d) IMP as inhibitor (T)
381. Conversion of IMP into AMP is inhibited by:
a) IMP (F)
b) AMP (T)
c) compound for synthesis of which GTP is required (T)
d) product of adenylosuccinate lyase (T)
382. Conversion of IMP into GMP is inhibited by:
a) ATP (F)
b) GMP (T)
c) compound which is product of IMP dehydrogenase (F)
d) compound for synthesis of which glutamine is required (T)
383. Synthesis of GMP from IMP is inhibited by:

+ CO2 + 2 ATP
NH2CH
COOH
CH2
CH2
C NH2
O
NH2
COO P
NH2
CNH
CH
CH2
C
O
O
HO
COOH
a) product of IMP dehydrogenase (F)
b) product of GMP synthase (T)
c) compound for synthesis of which GTP is required (F)
d) compound which can be also formed by HGPRT (T)
384. Source of NH2 group for carbamoylphosphate synthesis in synthesis of pyrimidine nucleotides is:
a) free NH3 (F)
b) glutamate (F)
c) glutamine (T)
d) compound formed in reaction whic requires ATP (T)
385. Synthesis of carbamoylphosphate in de novo synthesis of pyrimidine nucleotides:
a) proceeds in cytosol (T)
b) is catalyzed by carbamoylsynthase II (F)
c) is activated by ATP (T)
d) is main regulatory step of pyrimidine nucleotide synthesis in eucaryots (T)
386. Carbamoylphosphate in synthesis of pyrimidine nucleotides is formed:
a) in mitochondria (F)
b) by carbamoylphosphate synthase II (T)
c) in reaction which uses glutamine as source of NH2 group (T)
d) in reaction which is inhibited by ATP (F)
387. Reaction of condensation of aspartate with carbamoylphosphate:
a) is reaction of pyrimidine nucleotide synthesis (T)
b) proceeds in cytosol (T)
c) is catalyzed by aspartate transcarbamoylase (T)
d) is main regulatory enzyme of synthesis of pyrimidine nucleotides in eucaryots (F)
388. Aspartate transcarbamoylase is the enzyme which:
a) requires ATP as source of energy (F)
b) catalyzes reaction of ammonia detoxification in urea cycle (F)
c) uses as substrate carbamolyphosphate which is formed in mitochondria (F)
d) uses as substrate aspartate which can be formed by AST (T)
389. Orotate is compound which:
a) is formed from orotidine monophosphate (OMP) (F)
b) is product of dihydroorotase (F)
c) by decarboxylation produces uracil (F)
d) is intermediate of degradation of pyrimidine nucleotides (F)
390. Reaction:
a) leads to synthesis of compound: (T)
b) proceeds in cytosol (T)
c) is required for purine nucleotide synthesis (F)
d) is main regulatory step in pyrimidine nucleotide synthesis in eucaryots (T)
391. is compound which:

a) is formed by aspartate transcarbamoylase (T)
b) requires for synthesis essential amino acid (F)
c) is synthesizsd in reaction which is the main regulatory step of pyrimidine nucleotide synthesis in
eucaryots (F)
d) by dihyroorotase is changed into (F)
392. Reaction of conversion:
a) is catalyzed by thymidylate synthase (T)
b) requires methyl-THF as source of CH3 group (F)
c) is characteristic for red blood cells (F)
d) can be decreased by inhibitors of dihydrofolate reductase – folate analogs (T)
393. UMP in synthesis of pyrimidine nucleotides de novo:
a) is formed by reaction of uracil with PRPP (F)
b) is formed by decarboxylation of OMP (T)
c) is formed from immediate precursor dihydroorotate (F)
d) is changed to thymine nucleotide by receiving CH3 group form methionine (F)
394. For synthesis of cytidine nucleotides:
a) conversion of UMP into UTP is required (T)
b) energy is not required (F)
c) source of NH2 group isglycine (F)
d) source of energy is GMP (F)
395. CTP is compound which:
a) in de novo synthesis of pyrimidine nucleotides is formed from UTP (T)
b) requires aspartate as source of NH2 group (F)
c) requires glutamate as source of NH2 group (F)
d) is inhibitor of carbamoylphosphate synthase II (T)
396. Thymine nucleotides:
a) are formed from immediate precursor dUMP (T)
b) contain methyl group source of which is methionine (F)
c) require tetrahydrofolate for their synthesis (T)
d) are nuceotides synthesis of which is important for dividing cells (F)
397. Synthesis of thymine nucleotides:
a) requires methylene tetrahydrofolate as source of CH3 group (T)
b) is catalyzed by the enzyme thymidylate synthase (F)
c) is important mainly for red blood cells (F)
d) can be decreased by analogs of folic acid which inhibit dihydrofolate reductase (T)
398. Main regulatory enzyme of pyrimidine nucleotide synthesis in eucaryotic cells is:
CH
C
HNC
O
O NH
COOH
HN
O
O N
O
H OH
CH2 O P OH
OH
O
HN
O
O N
O
H OH
CH2 O P OH
OH
O
CH3
na

a) enzyme located in cytosol (T)
b) carbamoylphosphate synthase II (T)
c) enzyme which uses as substrates glutamine, CO2 and ATP (T)
d) enzyme which is inhibited by UTP (T)
399. Amino acid glutamine in synthesis of nucleotidesis required:
a) as source of NH2 group in AMP and GMP (F)
b) as source of NH2 group in CMP and TMP (F)
c) as source of two nitrogen atoms in purine heterocycle (T)
d) for production of carbamoylphosphate in synthesis of pyrimidine nucleotides (T)
400. Aspartate in synthesis of nucleotides is required:
a) for conversion of IMP to AMP (T)
b) as source of one nitrogen atom in purine heterocycle (T)
c) as source of carbon atoms in purine heterocycle (F)
d) as source of NH2 group in synthesis of CMP (F)
401. Conversion of nucleotides into deoxynucleotides:
a) proceeds at the level of nucleosidemonophosphates (F)
b) is catalyzed by ribonucleotide reductase (T)
c) requires oxidized form of thioredoxine (F)
d) requires NADPH2 for regeneration of reduced form of thioredoxine (T)
402. Ribonukleotidreductase is enzyme which:
a) catalyzes reaction of conversion of nucleosidediphosphates to deoxynucleosidediphosphates (T)
b) uses thioredoxine as source of hydrogens (F)
c) is inhibited by dATP (T)
d) requires coenzyme formed Krebs cycle (F)
403. In AMP degradation:
a) enzyme 5,-nucleotidase is required (T)
b) by the action of 5,-nucleotidase adenosine is formed (T)
c) by AMP deaminase AMP can be changed to IMP (T)
d) uric acid is formed as final product (T)
404. 5,-nucleotidase is enzyme which:
a) catalyzes conversion of AMP to adenosine (T)
b) catalyzes conversion of IMP to inosine (T)
c) participates in degradation of nucleosides (F)
d) leads to conversion of AMP to guanosine (F)
405. Adenosine is compound which:
a) is formed from AMPby 5,-nucleotidase (T)
b) is changed to hypoxanthine by adenosine deaminase (F)
c) can be changed to AMP by adenosine kinase (T)
d) belong to purine nucleotides (F)
406. Adenosine is changed to inosine:
a) in reaction catalyzed by adenosine deaminase (T)
b) in reaction catalyzed by adenosine kinase (F)
c) in reaction which requires water as one of substrates of reaction (T)
d) by phosphorolytic cleavage (F)
407. During degradation of purine nucleotides:
a) IMP is formed by AMP-deaminase (T)
b) conversion of IMP requires 5-nucleotidase (T)
c) purine nucleoside phosphorylase changes nucleotides into nucleosides (F)

d) final product of degradation is formed by xanthine oxidase (T)
408. Inosine during degradation of purine nucleotides:
a) can be formed by adenosine deaminase (T)
b) can be formed from IMP by 5,-nucleotidase (T)
c) can be changed to hypoxanthine by xanthine oxidase (F)
d) can be changed to adenosine by purine nucleoside phosphorylase (F)
409. Purine nucleoside phosphorylase is the enzyme which:
a) uses all purine nucleosides as substrate (F)
b) catalyzes synthesis of purine nucleosides form nucleotides (F)
c) leads to synthesis of hypoxanthine from inosine (T)
d) by degradation of guanosine produces guanine and ribose-1-phosphate (T)
410. Hypoxanthine is compound which:
a) is formed as intermediate during degradation of GMP (F)
b) is product by 5-nucleotidase (F)
c) in degradation of nucleotides is changed by xanthine oxidase (T)
d) can be chaged to IMP in reaction catalyzed by HGPRT (T)
411. Xanthine oxidase is enzyme which:
a) uses hypoxanthine as substrate (T)
b) uses xanthine as substrate (T)
c) synthesizes xanthine as product (T)
d) can be inhibited by allopurinol (T)
412. Final product of degradation of purine nucletides is:
a) uric acid (T)
b) compound which is also púroduct of ammonia detoxification in humans (F)
c) compound which can be used in salvage pathways (F)
d) compound which is excreted out of the body by lungs (F)
413. Uric acid is compound which:
a) final product of degradation of all nucleotides (F)
b) is formed by xanthine oxidase (T)
c) is bad soluble in water (T)
d) in high concentrations precipitates in joints causing gout (T)
414. Allopurinol is compound which:
a) is competitive inhibitor of xanthine oxidase (T)
b) inhibits reaction by which hypoxanthine is converted into xanthine and uric acid (T)
c) is used in therapy of hyperuricemia (T)
d) increases solubility of uric acid in water (F)
415. Allopurinol is used in therapy of hyperuricemia because:
a) decreases production of uric acid by inhibition of xanthine oxidase (T)
b) stimulates conversion of uric acid to water soluble compound (F)
c) activates salvage pathways of synthesis of purine nucleotides (F)
d) inhibits activity of all enzymes of purine nucleotide degradation and thus decreases synthesis of
uric acid (F)
416. In the process of GMP degradation:
a) 5,-nukleotidase is required (T)
b) purine nucleoside phosphorylase catalyzes conversion of guanosine to guanine (T)
c) xanthine is formed as intermediate (T)
d) uric acid is formed by the action of xanthine oxidase (T)

417. When these substrate enter reaction:
a) IMP can be formed by 5'-nuceotidase (F)
b) inosine is formed by deamination (F)
c) adenosine is formed by 5'-nucleotidase (T)
d) depending on the enzyme either IMP of adenosine can be formed (T)
418. Reaction:
+ H3PO4
a) is catalyzed by adenosine deaminase (F)
b) is catalyzed by purinenucleoside phosphorylase (T)
c) leads to synthesis of guanine and ribose-1-phosphate (T)
d) can be activated in primary metabolic hyperuricemia (F)
419. Reaction:
+ H2O + O2 + H2O2
a) is reaction of degradation of pyrimidine nucleotides (F)
b) is catalyzed by xanthine oxidase (T)
c) can be inhibited by allopurinol (T)
d) does not proceed in human cells (F)
420. Compound of the formula:
a) is final product of purine nucleotide degradation (T)
b) can be used as substrate for synthesis of purine nucleotides by salvage pathways (F)
c) is excreted out of the body by bile (F)
d) in higher concentration can precipitate in joints and urinary tract (T)
421. During CMP degradation:
a) -alanine is formed as final product (T)
b) compound which is component of proteins is fored as final products (F)
c) 5,-nucleotidase is required (T)
d) coenzyme formed in pentose phosphate pathway is required (T)
422. Degradation of thymine nucleotides:
a) requires 5,-nucleotidase (T)
N
N N
N
NH2
O
OH OHH
CH2 O P O-
O-
OH2O+
HN
N
N
O
H2N N
O
OH OH
CH2 OH
N
N NH
N
OH
N
N NH
N
OH
HO
N
N NH
N
OH
HO
OH

b) requires NADH2 (F)
c) leads to synthesis of -aminoisobutyrate as final product (T)
d) leads to synthesis of comúpound which in higher concentrations causes gout (F)
423. -alanine is formed:
a) by degradation of all nucleotides (F)
b) by breakdown of ureidopropionate during degradation of CMP (T)
c) by breakdown of ureidoisobutyrate during degradation of UMP (F)
d) as final product of nucleotides which are present in RNA (T)
424. -aminoisobutyrate is formed as final product of degradation of:
a) UMP (F)
b) TMP (T)
c) nucleotide which contains deoxyribose as carbohydrate component (T)
d) nucleotide for synthesis of which thetrahydrofolate is required (T)
425. Synthesis of AMP from adenosine:
a) is reaction of salvage pathways of synthesis of purine nucleotides (T)
b) needs less energy than synthesis of AMP by de novo pathway (T)
c) is catalyzed by adenine phosphoribosyltransferase (F)
d) requires ATP (T)
426. Adenosine kinase catalyzes reaction which:
a) uses adenosine and ATP as substrates (T)
b) as source of energy uses GTP (F)
c) spends 1 ATP for AMP synthesis (T)
d) requires PRPP (F)
427. GMP in salvage pathways is formed:
a) from guanine and PRPP as substrates (T)
b) by HGPRT (T)
c) by 5,-nucleotidase (F)
d) by enzyme lower activity of which causes primary metabolic hyperuricemia (T)
428. IMP in salvage pathways is formed:
a) from hypoxanthine as substrate (T)
b) by reaction where donor of ribosephosphate is PRDP (T)
c) from innosine (F)
d) by reaction catalyzed by HGPRT (T)
429. Hypoxanthine-guanine phosphoribosyltransferase (HGPRT) is enzyme:
a) which uses as substrate purine bases (T)
b) responsible for synthesis of GMP by salvage pathway (T)
c) activity of which is higher in Lesh-Nyhan syndrome (F)
d) decreased activity of which couses primary metabolic hyperuricemia (T)
430. HGPRT in salvage pathways of nucleotide synthesis:
a) catalyzes conversion of purine nucleosides to nucleotides (F)
b) is responsible for synthesis of AMP (F)
c) uses adenosine as substrate (F)
d) catalyzes synthesis of uric acid (F)
431. Conversion of hypoxanthine to IMP:
a) is catalyzed by xanthine oxidase (F)
b) requires ATP as source of energy (F)
c) in human cells does not proceed because IMP is formed only by de novo pathway (F)
d) is increased in primary metabolic hyperuricemia (F)

432. Adenosine kinase catalyzes reaction which:
a) uses as substrates
(T)
b) uses as substrates (F)
c) leads to synthesis (T)
d) requires GTP as source of energy (F)
433. In salvage pathways of nucleotide synthesis participate:
a) adenosine kinase which uses ATP as source of energy (T)
b) enzyme which requires ATP for conversion of guanine into guanosine (F)
c) enzyme which requires PRDP for conversion of uracil into UMP (T)
d) uridine/cytidine kinase which changes pyrimidine nucleosides into nucleotides (T)
434. Primary metabolic hyperuricemia:
a) is caused by decreased activity of PRPP-amidotransferase (F)
b) is caused by increased destruction of cells (F)
c) is caused by decreased activity of enzyme which catalyzes conversion of IMP to GMP (F)
d) is state when excretion of uric acid by urine is decreased (F)
435. Causes of primary metabolic hyperuricemia can be:
a) decreased activityx of adenosine deaminase (F)
b) decreased activity of HGPRT (T)
c) decreased activity of enzyme responsible for uric acid synthesis (F)
d) defect in regulation of enzyme which produces phosphoribosylamine (F)
436. Causes of secondary metabolic hyperuricemia can be:
a) increased activity of pRPP-amidotransferase (F)
b) decreased activity of salvage pathways of synthesis (F)
c) increased destruction of cells (F)
d) defect in excretion of uric acid (T)
437. What are differences between metabolic and renal hyperuricemia:
a) in metabolic hyperuricemia concentration of usic acid in blood is higher while in renal is decreased
(F)
b) in metabolic hyperuricemia production of uric acid is increased while in renal is decreased (F)
c) in renal hyperuricemia excretion of uric acid is decreased while in metabolic is normal or higher
(T)
d) they are not differences between both kinds of hyperuricemia (F)
N
N N
N
NH2
O
OH OH
CH2 OH
N
N N
N
NH2
O
OH OH
CH2 O P O
O-
O
P O-
O-
O
P O
O-
O
a
N
N N
N
NH2
O
OH OH
CH2 O P O-
O-
O
N
N NH
N
NH2
O
OH OHO
CH2 O P O-
O-
O
O P
O-
O
P-O
O-
Oa

438. In primary renal hyperuricemia:
a) ecretion of uric acid by urine is increased (F)
b) activity of HGPRT is decreased (F)
c) excretion of uric acid by tubular secretion is decreased (T)
d) clearance of creatinine is in physiological range (T)
439. Primary renal hyperuricemia is result of:
a) decreased glomerular filtration (F)
b) defect in tubular secretion of uric acid (T)
c) defect of kidneys when excretion of all compounds is lower (F)
d) defect of HGPRT in kidneys (F)
440. Secondary renal hyperuricemia:
a) is state when concentration of uric acid in blood and urine is higher (F)
b) is caused by defect of glomerular filtration (T)
c) is coused by defect in tubular secretion (F)
d) develops in kidney diseases when clearance of creatinine is decreased (T)
BIOLOGIC OXIDATIONS
441. Cells gain energy required for their functions:
a) by anaerobic oxidation of carbohydrates, lipids and proteins (F)
b) by all oxidation reduction reactions (F)
c) by oxidation of ATP and ADP (F)
d) by the processes called intermediary metabolism (T)
442. Living organism:
a) uses as direct source of energy energy rich compounds mainly ATP (T)
b) gains energy in oxidation-reduction processes (T)
c) produces ATP only in aerobic conditions (F)
d) can produce ATP in glycolysis also in anaerobic conditions (T)
443. Human organism:
a) takes as source of energy diretly energy rich compounds like ATP (F)
b) uses directly energy of glucose for all processes which require energy (F)
c) energy of foodstuffs converts to energy in ATP by reduction of substrates (F)
d) energy of foodstuffs converts to energy in ATP by oxidation of substrates (T)
444. In cells of human organism:
a) ATP is formed only in presence of oxygen (F)
b) ATP is formed mainly in lack of oxygen (F)
c) ATP is formed by processes which are located in mitochondria and cytosol (T)
d) ATP can be formed by process of substrate level phosphorylation (T)
445. Cells can gain energy:
a) by oxidation of main sorces of energy for example ATP (F)
b) by oxidation of carbohydrates, lipids and proteins (T)
c) only in aerobic conditions (F)
d) in cytosol by the process called oxidative phosphorylation (F)
446. Oxidation of compounds in the cells:
a) leads to synthesis of the same products as oxidation in vitro (T)
b) proceeds mainly by dehydrogenation (T)
c) releases enrgy mainly in the form of heat (F)
d) released energy uses for synthesis of energy rich compounds (T)

447. Oxidation of compounds in vivo:
a) is catalyzed by hydrolases (F)
b) is made only by removal of hydrogens – dehydrogenation (F)
c) is for example reaction of synthesis of lactate from pyruvate (F) jedine odoberaním vodíkov –
dehydrogenáciou (F)
d) requires oxygen for oxidation of fatty acids (T)
448. Glucose is compound which:
a) is universal source of energy (T)
b) allows synthesis of ATP in pentose phosphate pathway (F)
c) is oxidized to CO2 and H2O in all cells of human organism (F)
d) requires oxygen for complete oxidation (T)
449. Glucose:
a) allows synthesis of ATP by its reduction in glycolysis (F)
b) allows synthesis of ATP mainly in cytosol (F)
c) is source of energy for all cells of human body (T)
d) allows synthesis of ATP in mitochondria and in cytosol (T)
450. For synthesis of ATP from glucose:
a) its activation is required (T)
b) metabolic processes located in cytosol and in mitochondria are required (T)
c) oxygen is required in all cells of human body (F)
d) substrate level phosphorylation in cytosol is required (T)
451. Oxidation of compounds in the cells can be performed by:
a) addition of electrons (F)
b) reaction of compounds with NADH2 (F)
c) enzymes which use FAD, NAD and NADP as coenzymes (T)
d) by reaction of compound with oxygen (T)
452. Oxidation of compounds in vivo can proceed:
a) by removal of hydrogens in reactions catalyzed for example by succinate dehydrogenase (T)
b) by removal of electrons (T)
c) by reaction with oxygen for example in hydroxylation reactions (T)
d) by dehydrogenation for example in conversion of lactate to pyruvate (T)
453. Reduction of compounds in the cells:
a) can be made by hydrogenation (T)
b) can proceed by reaction of compounds with oxygen (F)
c) can be made by removal of electrons (F)
d) can be made by addition of proton for example in reaction HCO3- + H+ (F)
454. Reduction of compounds is possible:
a) by uptake of electrons (T)
b) by dehydrogenation (F)
c) by hydrogenation for example in reaction of -hydroxybutyrate synthesis from acetoacetate (T)
d) by removal of electrons for example during terminal oxidation (F)
455. Compound is oxidized:
a) when receives electrons (F)
b) when loses electrons (T)
c) by reaction with for example NADP (T)
d) by reaction with oxygen (T)
456. Compound is reduced:
a) when receives electrons (T)

b) when loses electrons (F)
c) by reaction with for example FAD (F)
d) when receives hydrogen (T)
457. Dehydrogenation is reaction:
a) which is the most frequent mode of oxidation-reduction reactions (T)
b) is reaction of conversion of alcohol to aldehyde (T)
c) is reaction by which water is removed (F)
d) which is used in synthesis of cholesterol (F)
458. During dehydrogenation:
a) compounds are oxidized (T)
b) hydrogens can be transfered to coenzyme FAD (T)
c) hydrogens are transfered to coenzyme pyridoxalphosphate (F)
d) hydrogens are transfered to coenzyme NAD (T)
459. Dehydrogenation is:
a) reaction by which comounds are oxidized (T)
b) reaction by which compounds are reduced (F)
c) mode of oxidation of compounds for example in Krebs cycle (T)
d) for example conversion of pyruvate to lactate (F)
460. During oxidation reduction reactions energy is released:
a) by transfer of electrons from compound with lower oxidation-reduction potential to compound
with higher oxidation-reduction potential (T)
b) for example by transfer of hydrogens from FADH2 to CoQ in terminal oxidation (T)
c) for example by transfer of electrons from FMNH2 to FeS protein in terminal oxidation (T)
d) for example in reaction of glucose synthesis by gluconeogenesis (F)
461. Energy rich compounds are compound which:
a) are formed in both aerobic and anaerobic conditions (T)
b) contain bonds by hydrolysis of which energy more than 30 kJ.mol-1 is released (T)
c) are formed in mitochondria and endoplasmic reticulum (F)
d) are used for activation of substrates (T)
462. Energy rich bonds:
a) can be formed also in anaerobic conditions (T)
b) are synthesized only in mitochondria (F)
c) are bonds by hydrolysis of which energy more than 3 kJ/mol is released (F)
d) are totally four types (F)
463. Compounds with energy rich bonds:
a) are formed mainly by the process of oxidative phosphorylation (T)
b) all cen be used for synthesis of ATP by substrate level phosphorylation (F)
c) all contains phosphate (F)
d) may contain phosphoanhydride bond – for example ATP and ADP (T)
464. Which of following bonds belong to energy rich:
a) phosphodiester in NAD (F)
b) acylphosphate present in acyl-CoA (F)
c) acylphosphate present in 1,3-bisphosphoglycerate (T)
d) enolphosphate which is formed during glycolysis (T)
465. Bonds which belong to energy rich are:
a) acylphosphate present in aminoacyladenylate (T)
b) thioester present in acetyl-CoA (T)
c) guanidinium phosphate present in GTP (F)

d) guanidinium phosphate uses for regeneration of ATP in working muscles (T)
466. Phosphoanhydride energy rich bond:
a) is present in ATP (T)
b) is source of energy for Na+/K+ ATP-ase (T)
c) is present in DNA (F)
d) is present in CTP which is used for phospholipid synthesis (T)
467. Which of statements about phosphoanhydride energy rich bond are correct:
a) is present in ATP which can be synthesized in mitochondria (T)
b) is present in GTP which can be synthesized by substrate level phosphorylation in mitochondria (T)
c) is synthesized in mitochondria using substrate AMP a H3PO4 (F)
d) is present in UTP which is used for synthesis of glycogen (T)
468. Acylphosphate energy rich bond:
a) is present in 1,3-bisphosphoglycerate (T)
b) is used for synthesis of ATP by substrate level phosphorylation (T)
c) contains the highest amount of energy (F)
d) is present in intermediate of fatty aciod activation – acyladenylate (T)
469. Acylphosphate energy rich bond is bond which:
a) is present in 3-phosphoglycerate (F)
b) can be formed by reaction of fatty acids with ATP (T)
c) can be formed by enolase (F)
d) is used for synthesis of ATP by oxidative phosphorylation (F)
470. Enolphosphate energy rich bond:
a) is present in enolpyruvate (F)
b) contains the highest amount of energy (T)
c) is formed by reaction of pyruvate with ATP (F)
d) can be used for synthesis of ATP in anaerobic conditions (T)
471. Phosphoenolpyruvate:
a) contains acylphosphate energy rich bond (F)
b) can be used for synthesis of ATP by substrate level phosphorylation in mitochondria (F)
c) is formed by dehydrogenation reaction (F)
d) is compound for synthesis of which in gluconeogenesis GTP is required (T)
472. Compound with guanidiniumphosphate energy rich bond:
a) is creatine phosphate (T)
b) is formed in mucles during work (F)
c) is direct source of energy for muscle contraction (F)
d) can be used for synthesis of ATP by substrate level phosphorylation (F)
473. Guanidinium phosphate energy rich bond:
a) is present in GDP (F)
b) is present in compound which is storage form of energy in muscles (T)
c) is specific donor of energy for gluconeogenesis (F)
d) is formed by creatine kinase (T)
474. Thioester energy rich bond is present in:
a) common intermediate of metabolism of carbohydrates, lipids and amino acids (T)
b) compound which is product of activation of fatty acids (T)
c) acyladenylate (F)
d) S-acetyllipoate which is formed during oxidative decarboxylation of pyruvate (T)
475. Thioster bond is bond which:

a) belongs to energy rich bonds (T)
b) is formed by binding of fatty acids residues to phosphate in CoA (F)
c) is present in intermediate of glyceraldehyde-3-phosphate oxidation to 1,3-bisphosphoglycerate (T)
d) can be used for synthesis of ATP by substrate level phosphorylation in Krebs cycle (T)
476. Energy released by spliting of energy rich bonds can be used for:
a) activation of substrate for example in synthesis of glucose-6-phosphate (T)
b) active transport (T)
c) transport of Na+ onto the cells (F)
d) synthesis of ATP by oxidative phosphorylation (F)
477. Energy of guanidiniumphosphate energy rich bond can be used for:
a) synthesis of ATP by substrate level phosphorylation (F)
b) transport of compounds through membranes (F)
c) regeneration of ATP in muscles during work (T)
d) proteosynthesis (F)
478. Creatine phosphate is compound which:
a) contains guanidiniumphosphate energy rich bond (T)
b) is direct source of energy in heart (F)
c) is formed by creatine kinase (T)
d) can be used for ATP synthesis by creatine kinase (T)
479. Acetyl-CoA contains:
a) thioester energy rich bond (T)
b) adenine nucleotide (T)
c) bond which is used for synthesis of ATP by oxidative phosphorylation (F)
d) energy rich bond which is also present in product of fatty acid activation (T)
480. Which of following bonds belon to energy rich:
a) guanidiniumphosphate bond present in GTP (F)
b) phosphoanhydride bond present in ADP (T)
c) acylphosphate bond present in 1,3-bisphosphoglycerate (T)
d) thioester bond present in CoA (F)
481. Oxidoreductases are enzymes which:
a) catalyze oxidation of substrate (T)
b) catalyze reduction of substrates (T)
c) participate for example in synthesis of fatty acids (T)
d) are also component of respiratory chain (T)
482. Which of following enzymes belongs to oxidoreductases:
a) all enzymes of Krebs cycle (F)
b) all enzymes of terminal oxidation (T)
c) all enzymes of glycolysis (F)
d) enzymes which allow synthesis of ATP by substrate level phosphorylation (F)
483. Catalase is the enzyme which:
a) is located in cytosol (F)
b) is located in peroxisomes (T)
c) catalyzes reduction of hydrogen peroxide (T)
d) catalyzes oxidation of hydrogen peroxide (T)
484. H2O2:
a) can be formed by the action of aerobic dehydrogenases (T)
b) by catalase is reduced to O2 (F)
c) by catalase is reduced to H2O (T)

d) by peroxidase is reduced to H2O (T)
485. NAD is coenzyme which:
a) contains riboflavine (F)
b) contains adenine nuceotide (T)
c) binds two protons and two electrons during its reduction (F)
d) is carrier of hydrogens in terminal oxidation (F)
486. Monooxygenases are enzymes which:
a) belong to oxidoreductases (T)
b) catalyze binding of molecula of O2 into the substrate (F)
c) catalyze hydroxylation of substrates (T)
d) require NADPH2 and oxygen (T)
487. FAD is coenzyme which:
a) is derived for riboflavine (T)
b) contains adenine nucleotide (T)
c) during reduction binds one hydrogen and one electron (F)
d) transfers hydrogens to terminal oxidation directly to cytochrome a (F)
488. FAD:
a) is coenzyme of oxidoreductases (T)
b) is coenzyme of monooxygenases (F)
c) allows synthesis of 3 ATP by oxidative phosphorylation (F)
d) is coenzyme which during reduction binds two hydrogen atoms (T)
489. Monooxygenases:
a) catalyze binding of one oxygen atom into the substrate (T)
b) catalyze reduction of substrates (F)
c) are components of respiratory chain (F)
d) by hydroxylation of substrates they increase solubility of compounds in water (T)
490. Oxidation-reduction reactions in the cells:
a) can be catalyzed by dehydrogenases with coenzymes for example NAD or FAD (T)
b) can be catalyzed by peroxidase, which produces H2O2 (F)
c) can catalze binding of oxygen into substrate – for example monooxygenases and dioxygenases (T)
d) all are important for synthesis of ATP (F)
491. Which of components of terminal oxidation transfers hydrogens:
a) NAD+ (F)
b) FMN (T)
c) Green complex IV (F)
d) coenzyme Q (T)
492. Which of components of terminal oxidation transfers only electrons:
a) FMN (F)
b) CoQ (F)
c) cytochromes for example cyt b5 (F)
d) FeS protein (T)
493. NADH+H+ is coenzyme which:
a) is the coenzyme of monooxygenases (F)
b) transfers hydrogens to FMN in respiratory chain (T)
c) enables synthesis of 3 ATP by oxidative phosphorylation (T)
d) when is oxidized 6 protons are transported into mitochondrial matrix (F)
494. Carriers of hydrogens and electrons in terminal oxidation:

a) transfer electrons from system with higher oxidation-reduction potential to system with lower
oxidation-reduction potential (F)
b) are arranged in order:FMN, FeS-protein, cytochrome c, cytochrome b, coenzyme Q, cytochrome
a and cytochrome a3 (F)
c) transfer protons into mitochondrial matrix (F)
d) catalyze synthesis of ATP by oxidative phosphorylation (F)
495. Which of statements about carriers of electrons and hydrogens in terminal oxidation are correct:
a) coenzyme Q can take electrons from FADH2 throug Green complex II (T)
b) coenzyme Q during its reduction takes electron from FeS protein (T)
c) coenzyme Q during its reduction takes two H+ from mitochondrial matrix (T)
d) cytochrome c oxidase is responsible for transfer of electrons from FeS protein to
cytochrome c (F)
496. When hydrogens are transfered to terminal oxidation by NADH2:
a) FMN is the first acceptor (T)
b) 6 H+ are transported into intermembrane space (T)
c) 3 ATP can be formed (T)
d) coenzyme Q is the first acceptor (F)
497. Green complex I:
a) is present in outer mitochondrial membrane (F)
b) is NADH–ubiquinone reductase (T)
c) transfers two hydrogens from NADH2 to FeS–protein (F)
d) is important for reoxidation of FADH2 in terminal oxidation (F)
498. FeS protein in terminal oxidation:
a) takes electrons from FMNH2 (T)
b) transfers electrons directly to oxygen (F)
c) transfers electron and proton to coenzyme Q (F)
d) during reduction changes from Fe2+ into Fe3+ (F)
499. Terminal oxidation is the process which:
a) is located in inner mitochondrial membrane (T)
b) is used for reoxidation of coenzymes NAD and NADP2 (F)
c) produces gradient of protons which is used for synthesis of ATP (T)
d) is inhibited by compounds called uncouplers (F)
500. Coenzyme Q is compound which:
a) is part of Green complex II (F)
b) during reduction takes two hydrogens from FMNH2 (T) pri svojej redukcii prijíma dva atómy
vodíka z FMN (F)
c) is oxidized by transfer of electrons to cytochrome c and cytochrome b (T)
d) is the first acceptor of hydrogens when donor of them is FADH2 (T)
501. ATP/ADP translocase:
a) is component of respiratory chain (F)
b) is important because of impermeability of inner mitochondrial membrane for ATP and ADP (T)
c) is antiport transport system (T)
d) transports ATP out of mitochondria to cytosol (T)
502. ATP/ADP translocase is system which:
a) is present in inner mitochondrial membrane (T)
b) is responsible for synthesis of ATP by oxidative phosphorylation (F)
c) is responsible for active transport of ATP and ADP (F)
d) can be inhibited by compounds called uncouplers (F)

503. Cytochrome b in terminal oxidation:
a) transfers electrons to coenzyme Q durings its reduction (T)
b) takes electrons from coenzyme Q during its oxidation (T)
c) contains ions of iron and copper (F)
d) transfers electrons form FeS protein to coenzyme Q (F)
504. Ubiquinone-cytochrome c reduktase is system which:
a) is component of Green complex III (T)
b) is required in the process of oxidation of NADH2 in terminal oxidation (T)
c) is required in the process of oxidation of FADH2 in terminal oxdation (T)
d) is responsble for transfer of electrons to oxygen (F)
505. Cytochrome c oxidase:
a) is located in mitochondrial matrix (F)
b) contains ions of iron and copper (T)
c) transfers electrons to oxygen (T)
d) can be activated by CN- ions (F)
506. When FADH2 is donor of hydrogens to respiratory chain, then:
a) FMN is the first acceptor of hydrogens (F)
b) CoQ is the first acceptor of hydrogens (T)
c) 6 protons are transported to outer side of mitochondrial membrane (F)
d) 2 ATP can be formed (T)
507. Mitochondrial ATP-ase:
a) is present in mitochondrial matrix (F)
b) is enzyme of terminal oxidation (F)
c) contains F0 subunitwhich represents proton channel (T)
d) contains F1 subunit which synthesizes ATP form ADP and phosphate (T)
508. Mitochondrial ATP-ase is enzyme which:
a) consists of F1 and F0 subunits (T)
b) is responsible for hydrolysis of ATP (F)
c) uses energy of proton gradient for synthesis of aTP (T)
d) is responsible for transport of ATP into cytosol (F)
509. Uncouplers for example dinitrophenol:
a) inhibit process of terminal oxidation (F)
b) increase synthesis of ATP (F)
c) increase permeability of inner mitochondrial membrane for protons (T)
d) increase production of proton gradient (F)
510. Process of terminal oxidation:
a) is called also oxidative phosphorylation (F)
b) is inhibited by lack of oxygen (T)
c) is activated during hypoxia by lack of oxygen (F)
d) is activated by increased ratio ADP/ATP in mitochondria (T)
ACETYL-CoA + KETONE BODIES
511. Acetyl-CoA is compound which:
a) is product of aerobic glycolysis (F)
b) is common intermediate of metabolism of carbohydrates, lipids and amino acids (T)
c) can be formed from ketogenic amino acids (T)
d) can be used for synthesis of cholesterol in mitochondria (F)
512. Acetyl-CoA:

a) can be formed in mitochondria by -oxidation of fatty acids (T)
b) can be formed in mitochondria by pyruvate dehydrogenase (T)
c) can be formed in cytosol by utilization of ketone bodies (F)
d) can be formed in anaerobic conditions (F)
513. Acetyl-CoA can be utilized in the cells:
a) for oxidation in Krebs cycle in mitochondria (T)
b) for synthesis of fatty acids in cytosol (T)
c) for synthesis of ketone bodies in cytosol (F)
d) for synthesis of cholesterol in mitochondria (F)
514. Utilization of acetyl-CoA is possible:
a) in cytosol for synthesis of ATP (F)
b) for synthesis of mevalonate in cytosol (T)
c) for synthesis of ketone bodies in mitochondria (T)
d) for synthesis of glucose by gluconeogenesis (F)
515. Acetyl-CoA can be formed:
a) in aerobic conditions from pyruvate (T)
b) from pyruvate in reaction which requires TPP, lipoic acid, CoA, NAD and FMN (F)
c) from glucose by irreversible reaction (T)
d) during -oxidation by spliting of -ketoacyl-CoA (F)
516. Acetyl-CoA:
a) contains thioester energy rich bond (T)
b) can be formed in mitochondria and cytosol (T)
c) for synthesis of gfatty acids is used in cytosol where is transported in the form of citrate (T)
d) from fatty acids can be formed also in anaerobic conditions (F)
517. Utilization of acetyl-CoA for ketone bodies synthesis:
a) proceeds in cytosol (F)
b) requires enzymes acetoacetyl-CoA thiolase, HMG-CoA synthase and HMG-CoA lyase (T)
c) leds to synthesis of mevalonate as intermediate (F)
d) is activated by insulin (F)
518. Which of statements about utilization of acetyl-CoA are correct::
a) oxidation of acetyl-CoA in Krebs cycle allows synthesis of 12 ATP (T)
b) utilization of acetyl-CoA for cholesterol synthesis requires NADPH2 (T)
c) utilization of acetyl-CoA for fatty acid synthesis is activated during starvation (F)
d) Utilization of acetyl-CoA for ketone bodies synthesis proceeds only in liver (T)
519. Acetoacetyl-CoA:
a) can be formed as intermediate of -oxidation of fatty acids (T)
b) in intermediate of conversion of acetyl-CoA to ketone bodies (T)
c) can be formed during utilization of ketone bodies by succinyl-CoA transferase (T)
d) can be hydrolyzed into two acetyl-CoA (F)
520. Acetoacetyl-CoA is compound which:
a) is formed form two acetyl-CoA in tthe process of ketone bodies synthesis (T)
b) in synthesis of ketone bodies is formed by -ketothiolase (T)
c) in utilization of ketone bodies is formed by -ketothiolase (F)
d) in utilization of ketone bodies is changed by -ketothiolase (T)
521. Ketone bodies are compounds which:
a) are formed only in pathological conditions (F)
b) are synthesized from immediate precursor acetoacetyl-CoA (F)
c) what cannot be utilized in red blood cells (T)

d) what are utilized in all cells of human body (F)
522. Which of following compounds belong to ketone bodies:
a) all compounds with keto group (F)
b) -hydroxybutyrate (T)
c) -hydroxy--methylglutaryl-CoA (F)
d) compound formed from -hydroxy--methylglutaryl-CoA by HMG-CoA lyase (T)
523. Acetone is compound which:
a) belongs to ketone bodies (T)
b) is formed from acetoacetate by spontaneous decarboxylation (T)
c) can be utilized in extrahepateic tissues for synthesis of ALP (F)
d) causes acidosis when is in high concentration (F)
524. -hydroxy--methyl-glutaryl-CoA:
a) is formed from acetoacetate and acetyl-CoA (F)
b) is intermediate in ketone bodies synthesis (T)
c) in extrahepatic tissues can be changed to acetoacetate (F)
d) is product of -oxidation (F)
525. Under physiological conditions:
a) ketone bodies are utilized in brain (F)
b) ketone bodies are not formed (F)
c) ketone bodies are not excreted by urine (T)
d) myocardial cells can oxidize ketone bodies for ATP synthesis (T)
526. Acetoacetyl-CoA is broken by -ketothiolase:
a) during ketone bodies synthesis (F)
b) during ketone bodies utilization in extrahepatic tissues (T)
c) into 2 molecules of acetone (F)
d) into 2 molecules of acetyl-CoA (T)
527. Which of following compounds is required for utilization of ketone bodies:
a) -hydroxy- -methyl glutaryl-CoA (F)
b) acetyl-CoA (F)
c) succinyl-CoA (T)
d) intermediate of Krebs cycle with thioester energy rich bond (T)
528. Synthesis of ketone bodies:
a) proceeds in cytosol of liver cell (F)
b) proceeds also under phasiological conditions and ketone bodies serve as source of energy for
extrahepatic tissues (T)
c) is increased in diabetes (T)
d) in starvatuion is inhibited by glucaon (F)
529. Ketegenesis is the process which:
a) requires HMG-CoA lyase (T)
b) leads to synthesis of acetoacetate, -hydroxybutyrate and acetone (T)
c) uses acetyl-CoA as basic substrate (T)
d) during starvation and diabetes is activated by insulin (F)
530. Ketone bodies are compounds which:
a) are formed in skeletal muscle (F)
b) are formed only in pathologic onditions (F)
c) cannot be metabolized in liver (T)
d) can be oxidized in brain (T)

531. Acetoacetate is compound which:
a) is formed by HMG-CoA lyase during ketone bodies synthesis (T)
b) is formed by -hydroxybutyrate dehydrogenase durin extrahepatal utilization of ketone bodies (T)
c) in high concetrations can cause acidosis (T)
d) can be utilizaed in liver (F)
532. Acetoacetate is compound about which can be said:
a) belongs to ketone bodies (T)
b) it is compound which can be used for synthesis of ATP in all cells of human body (F)
c) for its synthesis 3 acetyl-CoA are used (T)
d) in extrahepatic tissues is activated ba reaction with ATP (F)
533. Process of ketone bodies synthesis:
a) requires acetyl-CoA as basic substrate (T)
b) requires enzyme thiolase (T)
c) requires enzyme HMG-CoA reductase (F)
d) is activated by high glucose concentration (F)
534. During process of ketone bodies synthesis:
a) acetoacetyl-CoA is formed from HMG-CoA (F)
b) HMG-CoA is synthesized form acetoacetyl-CoA and acetyl-CoA (T)
c) HMG-CoA is synthesized by HMG-CoA lyase (F)
d) acetone is formed by spontaneous decarboxylation of acetoacetate (T)
535. HMG-CoA is compound which:
a) is formed by reaction of acetoacetate with acetyl-CoA (F)
b) requires energy form ATP for conversion into acetoacetate (F)
c) is changed to acetone by HMG-CoA lyase (F)
d) as intermediate of ketone bodies synthesis i synthesized in mitochondria (T)
536. Hydroxybutyrate dehydrogenase:
a) is enzyme required in the process of ketone bodies synthesis (T)
b) is enzyme required for extrahepatal utilization of ketone bodies (T)
c) uses FAD as coenzyme (F)
d) is in high activities in myocardium (T)
537. -hydroxybutyrate:
a) can be used for synthesis of ATP in brain during starvation (T)
b) in the liver is formed by -hydroxybutyrate dehydrogenase (T)
c) can be decarboxylated to acetone (F)
d) requires NAD and hydroxybutyrate dehydrogenase for its utilization in extrahepetic tissues (T)
538. -hydroxybutyrate is compound which:
a) is synthesized in cytosol of liver cells (F)
b) contains keto goup and thus belongs to ketone bodies (F)
c) is formed by reduction of acetoacetate (T)
d) is formed in higher concentrations in patients with diabetes melllitus (T)
539. Which cells of human body cannot utilize ketone bodies:
a) red blood cells (T)
b) skeletal muscle (F)
c) liver (T)
d) brain cells (F)
540. Extrahepatal utilization of ketone bodies:
a) is important source of energy in heart (T)
b) requires conversion of acetoacetate into acetoacety-CoA by CoA transferase (T)

c) proceeds in anaerobic conditions (F)
d) is increased in diabetes (F)
541. Reaction of conversion of acetoacetate into acetoacetyl-CoA:
a) requires enzyme succinyl-CoA transferase (T)
b) requires ATP (F)
c) requires HMG-CoA (F)
d) is catalyzed by enzyme which is absent in liver (T)
542. Malonyl-CoA is compound which:
a) is formed as intermediate of fatty acid synthesis (T)
b) regulates transport of fatty acids into mitochodria and their oxidation (T)
c) activates carnitineacyltransferase I (F)
d) by accitavtion of fatty acid oxidation increases synthesis of ketone bodies (F)
543. Malonyl-CoA:
a) regulates process of ketogenesis by inhibition of transport of fatty acids into mitochondria and their
oxidation (T)
b) is formed in cytosol by acetyl-CoA carboxylase (T)
c) is formed by enzyme which is active in phosphorylated form (F)
d) is intermediate of fatty acid synthesis does not participate in regulation of their oxidation (F)
544. Increased synthesis of ketone bodies in diabetes is caused by:
a) increased concentration of glucose in blood (F)
b) increased oxidation of fatty acids (T)
c) increased activity of hormone senstive lipase due to absence of antilipolytic effect of insulin (T)
d) decreased utilization of acetyl-CoA in Krebs cycle (T)
545. Factors which stimulate synthesis of ketone bodies in diabetes and starvation are:
a) decreased lipolysis in adipose tissue (F)
b) decreased utilization of acetyl-CoA in fatty synthesis (T)
c) activation of enzymes of -oxidation by glucagon (F)
d) increades degradation of triacylglycerols in adipose tissue (T)
546. In regulation of ketone bodies synthesis participate:
a) inhibition of hormone sensitive lipase by insulin (T)
b) increased oxidation of acetyl-CoA in Krebs cycle (F)
c) activity of acetyl-CoA carboxylase which allows utilization of acetyl-CoA in fatty acid synthesis
(T)
d) regulation of -oxidation by insulin (F)
547. In activation of ketone bodies synthesis participate:
a) lipolysis in adipose tissue stimulated by change of hormone sensitive lipase into acctive –
phosphorylated form (T)
b) increased transport of acetyl-CoA into the liver (F)
c) decreased utilization of acetyl-CoA in fatty acid synthesis (T)
d) increased oxidation of fatty acids caused by increased activity of carnitine acyltransferase (T)
548. Increased concentration of ketone bodies causes:
a) Ketonemia (T)
b) ketonuria (F)
c) decrease of pH – acidosis (T)
d) diabetes mellitus (F)
549. Increased concentration of ketone bodies in blood in diabetes:
a) is caused by increased synthesis of ketone bodies in liver (T)
b) is caused by increased utilization of ketone bodies in brain (F)

c) is related to increased oxidation of fatty acids (T)
d) can cause metabolic acidosis which is compensated by hyperventilation (T)
550. Increased concetration of ketone bodies in blood:
a) is typical for diabetes mellitus I. type (T)
b) is found during starvation (T)
c) is caused by activation of ketogenesis in liver (T)
d) by increased proton concentration increases pH (F)
VITAMINS
551. Functions of riboflavine (vitamin B2) are:
a) is the coenzyme of glutamate dehydrogenase (F)
b) is the component of NAD+ (F)
c) is the coenzyme of transaminases (F)
d) is required for Krebs cycle (T)
552. Riboflavine is vitamin which:
a) is component of FMN (T)
b) is component of coenzyme of succinate dehydrogenase (T)
c) is required for oxidative decarboxylation of pyruvate (T)
d) participates in indirect deamination of aminoacids (F)
553. In amino acid metabolism vitamin B2 is required for:
a) deamination of glutamate (F)
b) transamination (F)
c) direct deamination of amino acids (T)
d) reaction catalyzed by aminoacid oxidases (T)
554. Vitamin B2 is the component of the coenzyme for:
a) transamination (F)
b) oxidative deamination of amino acids (T)
c) carboxylation of pyruvate (F)
d) oxidative decarboxylation of pyruvate (T)
555. During reduction molecula of FAD binds:
a) 2 hydrogens into adenine component (F)
b) 2 hydrogens into isoalloxazine (T)
c) 1 hydrogen + 1 electrone into nicotinamide (F)
d) 2 protons and two electrons into riboflavine (T)
556. Which of following vitamins is not required in oxidation-reduction reactions:
a) vitamin C (F)
b) vitamin D (T)
c) vitamins B6 (T)
d) folic acid (T)
557. Which of following vitamins are present in coenzymes of oxidation-reduction reactions:
a) folic acid (F)
b) nicotinamide (T)
c) riboflavine (T)
d) vitamin PP (T)
558. Thiamine pyrophosphate:
a) contains energy-rich bond (T)
b) is required for formation of acetyl-CoA from glucose (T)
c) is required for further oxidation of glucose (T)

d) is required for decarboxylation of amino acids (F)
559. Which of following enzymes requires thiamine pyrophosphate:
a) glutamate dehydrogenase (F)
b) transketolase (T)
c) lactate dehydrogenase (F)
d) pyruvate dehydrogenase (T)
560. In decarboxylation of amino acids participates:
a) biotin (F)
b) thiamine (F)
c) vitamin B6 (T)
d) para-amino benzoate (F)
561. Lipoate is the coenzyme required for:
a) oxidative decarboxylation of pyruvate (T)
b) synthesis of acetyl-CoA form glucose (T)
c) ketogutarate synthesis in Krebs cycle (F)
d) ketoglutarate conversion in Krebs cycle (T)
562. Lipoic acid:
a) the component of pyruvate dehydrogenase complex (T)
b) participate in one reaction of Krebs cycle (T)
c) is coenzyme for oxidative decarboxylation of -ketoglutarate (T)
d) is coenzyme for decarboxylation of amino acids (F)
563. Which of following vitamins is the component of the enzyme system for oxidative decarboxylation of
ketoglutarate in Krebs cycle:
a) ascorbate (F)
b) vitamin B2 (T)
c) vitamin B6 (F)
d) thiamine (T)
564. Which of following coenzymes is the coenzyme for oxidative decarboxylation of pyruvate:
a) lipoic acid (T)
b) thiamine pyrophosphate (T)
c) tetrahydrofolate (F)
d) coenzyme containing vitamin B2 (T)
565. NADPH2:
a) is coenzyme formed during oxidation of fatty acids (F)
b) is formed during oxidation of glucose in glycolysis (F)
c) is required for synthesis of cholesterol (T)
d) is required for hydroxylation processes (T)
566. Nicotinamide:
a) in the forme of cenzyme is the component of aminotransferases (F)
b) in the form of coenzyme participates in oxidation-reduction reactions (T)
c) is present in FAD (F)
d) in the form of coenzyme is required for synthesis of fatty acids (T)
567. Coenzyme form of nicotinamide:
a) is NADP (T)
b) is component of respiratory chain (F)
c) is required for glycolysis (T)
d) participates in form reactions of Krebs cycle (F)

568. NAD:
a) is coenzyme of for example ketoglutarate dehydrogenase (T)
b) contains nicotinamide (T)
c) is required for deamination of glutamate (T)
d) transfers hydrogens to terminal oxidation directly to CoQ (F)
569. In amino acid metabolism vitamin B6 is required for:
a) deamination of glutamate (F)
b) decarboxylation of glutamate (T)
c) conversion of serine into glycine (F)
d) deamination of serine (T)
570. Pyridoxalphosphate is the coenzyme in reaction where ketoglutarate is the substrate. It is the reaction
of:
a) decarboxylation of ketoglutarate (F)
b) reductive amination of ketoglutarate (F)
c) transamination (T)
d) reaction of synthesis of glutamate from ketoglutarate and alanine (T)
571. Pyridoxalphosphate:
a) the coenzyme for decarboxylation of ketoacids (F)
b) is coenzyme of deamination of serine (T)
c) is the coenzyme for oxidative deamination of amino acids (F)
d) is the coenzyme for transamination (T)
572. Indirect deamination of amino acids requires coenzymes derived from vitamins:
a) thiamine (F)
b) riboflavine (F)
c) nicotinamide (T)
d) pyridoxine (T)
573. Transfer of CH3 groups from methyl-tetrahydrofolate into homocysteine to regenerate methionine
requires vitamin:
a) B6 (F)
b) B12 (T)
c) derived from nicotinamide (F)
d) the same as in the reaction catalyzed by methylmalonyl-CoA mutase (T)
574. Vitamin B12 is required for:
a) conversion ofpropionyl-CoA into methyl-malonyl- CoA (F)
b) -oxidation of fatty acids (F)
c) conversion of methyl-malonyl-CoA into succinyl-CoA (T)
d) normal glucose oxidation (F)
575. Methylmalonyl-CoA is formed:
a) from propionyl-CoA by decarboxylation (F)
b) from succinyl-CoA by transfer of CH3 group (F)
c) from malonyl-CoA (F)
d) from propionyl-CoA in presence of biotine (T)
576. Methylmalonyl-CoA mutase:
a) produces methylmalonate (F)
b) is one step in conversion of propionyl-CoA into succinyl-CoA (T)
c) requires vitamin B12 as coenzyme (T)
d) participates in synthesis of fatty acids (F)
577. Methylmalonyl-CoA in the presence of vitamin B12 is converted into:

a) methylmalonate (F)
b) propionyl-CoA (F)
c) succinyl-CoA (T)
d) intermediate of Krebs cycle (T)
578. Which of following compounds can be source of one carbon groups:
a) serine (T)
b) glutamate (F)
c) histidine (T)
d) tryptophane (T)
579. Panthotenic acid is vitamin which:
a) belongs to water soluble vitamins (T)
b) is component of CoA (T)
c) is component of acylcarrier protein (ACP) (T)
d) is required for oxidation of glucose by glycolysis (F)
580. Coenzyme A:
a) contains panthotenic acid (T)
b) contains riboflavine (F)
c) contains adenine nucleotide (T)
d) cantains energy rich thioester bond (F)
581. Folic acid is vitamin which:
a) belong to vitamins of B group (T)
b) contains nicotinamide (F)
c) by folate reductase and dihydrofolate reductase changes into its coenzyme form (T)
d) in the form of tetrahydrofoloate is required for nucleotide synthesis (T)
582. Folic acid:
a) is vitamin coenzyme form of which is tetrahydrofolate (T)
b) is form of coenzyme is required for transfer of one carbon groups (T)
c) in form of coenzyme participate in conversion of serine into glycine (T)
d) in form of coenzyme is required for synthesis of purine nucleotides (T)
583. Tetrahydrofolate:
a) contains paraaminobenzoate (T)
b) contains glutamate (T)
c) participates in transfer of one-carbon groups (T)
d) is required for synthesis of thymine (T)
584. One carbon groups are used for synthesis of:
a) glucose by gluconeogenesis (F)
b) synthesis of thymine nucleotides (T)
c) synthesis of purine nucleotides de novo (T)
d) synthesis of purine nucleotides by salvage pathways (F)
585. Which of following reactions require biotin as a coenzyme:
a) synthesis of oxaloacetate from pyruvate (T)
b) synthesis of malonyl-CoA in fatty acid synthesis (T)
c) decarboxylation of pyruvate (F)
d) carboxylation of pyruvate (F)
586. Rate limiting reaction of fatty acid synthesis which requires biotin is the reaction:
a) catalyzed by acetyl-CoA carboxylase (T)
b) catalyzed by condensing enzyme (F)
c) catalyzed by -keto acyl reductase (F)

d) where malonyl-CoA is formed (T)
587. Biotin is required as coenzyme:
a) in the process of gluconeogenesis (T)
b) in synthesis of cholesterol (F)
c) in the process of fatty acid synthesis (T)
d) in reaction of oxidative decarboxylation of pyruvate (F)
588. Vitamin C (ascorbate) is required for:
a) decarboxylation of pyruvate (F)
b) deamination of amino acids (F)
c) synthesis of adrenaline (T)
d) synthesis of collagen (T)
589. Active form of vitamin B6:
a) is componund with formula (F)
b) is coenzyme of ALT which catalyzes transamination of glutamate and oxaloacetate (F)
c) is coenzyme of this reaction (T)
d) is coenzyme required for utilization of amino acids in gluconeogenesis (T)
MOLECULAR BIOLOGY
590. During process of replication:
a) DNA strands are seperated by DNA-ligase (F)
b) primase synthesizes DNA (F)
c) one strand serves as the template for the synthesis of complementary strand (T)
d) synthesis of DNA proceeds in direction 3 to 5 (F)
591. Synthesis of proteins:
a) requires aminoacyl-tRNA synthase (T)
b) requires GTP as the source of energy (T)
c) is located at ribosomes (T)
d) is proces which produces energy (F)
592. For replication are required:
a) primase (T)
b) nuclesidetriphosphates (F)
c) deoxynucleoside triphosphates (T)
d) DNA polymerase III (T)
593. Primase is the enzyme which:
a) is required for DNA synthesis (T)
b) uses deoxynucleosidetriphosphates as a substrates (F)
c) participates in the synthesis of proteins (F)
d) synthesizes chain of RNA (T)
594. During replication:
a) DNA is synthesized in 3-5direction (F)
N
OHCH2
CH3
C
HO
H O
COOH
CH
CH2
CH2
COOH
NH2
CO2
CH2
CH2
CH2
COOH
NH2

b) Okazaki fragments are joined by DNA polymerase (F)
c) primer is formed from deoxyribonucleotides (F)
d) DNA helicase forms double helix (F)
595. DNA polymerase III:
a) requires primer (T)
b) catalyzes synthesis of DNA in direction 5, to 3, (T)
c) has endonuclease activity (F)
d) is required for reparation of DNA (F)
596. Okazaki fragments:
a) are formed during transcription (F)
b) are formed during replication in lagging strand (T)
c) are joined by DNA helicase (F)
d) consist of ribo- and deoxynucleotides (T)
597. DNA polymerase I:
a) has exonuclese activity (T)
b) is the main enzyme for elongation of DNA (F)
c) during replication is required for removal of primer (T)
d) is important for reparation of DNA (T)
598. mRNA:
a) in its molecula nucleotides are connected by phosphoanhyride bond (F)
b) contains triplet of nucleotides called anticodon (F)
c) is formed in nucleus (T)
d) is formed by the process of translation (F)
599. Transcription is the process:
a) of synthesis of DNA according to DNA (F)
b) which requires template (T)
c) which is catalyzed by DNA-polymerase (F)
d) which requires deoxynucleoside monophosphates (F)
600. tRNA:
a) is required for transcription (F)
b) binds activated amino acid (T)
c) has triplet of nucleotides called codon (F)
d) terminal nucleotides are CCA (T)
601. Synthesis of RNA:
a) requires RNA-polymerase (T)
b) requires primase (F)
c) is called translation (F)
d) is called replication (F)
602. Promoter is:
a) enzyme responsible for replication (F)
b) initial codon in mRNA (F)
c) part of mRNA through mRNA bins to ribosome (F)
d) enzyme catalyzing formatiuon of peptide bond (F)
603. Transcription is the process:
a) of the synthesis of mRNA (T)
b) of the synthesis of tRNA (T)
c) located at ribosomes (F)
d) which requires RNA polymerase (T)

604. During process of transcription:
a) RNA is synthesized according to the both strands of DNA (F)
b) proteins are synthesized at ribosomes (F)
c) RNA polymerase catalyzes formation of phosphodiesteric bond (T)
d) DNA is synthesized according to the sequence of amino acids in proteins (F)
605. Write which statement about tRNA and mRNA is true:
a) mRNA is formed in nucleus and tRNA is formed at ribosomes (F)
b) mRNA contains anticodon and tRNA not (F)
c) tRNA binds activated amino acid (T)
d) mRNA codes for primary structure of proteins (T)
606. mRNA in eucaryotic cell:
a) is formed directly by transcription (F)
b) contains non-coding regions called exons (F)
c) has CCA as terminal nucleotides (F)
d) at 5, end has bound methylguanine (T)
607. Synthesis of RNA:
a) requires RNA-polymerase (T)
b) requires primer (F)
c) utilizes AMP, GMP, CMP and UMP as the substrates (F)
d) is called replication (F)
608. Introns:
a) are coding sequences in hnRNA (F)
b) are non-coding sequences in hnRNA (T)
c) are triplets of nucleotides in tRNA (F)
d) are removed by splicing (T)
609. tRNA:
a) is synthesized in ribosomes (F)
b) codes amino acid by its codon (F)
c) for binding of amino acid requires CTP as donor of energy (F)
d) binds amino acid at ribose of terminal nucleotide adenine (T)
610. When proteins are formed in ribosomes:
a) GTP is the main donor of energy (T)
b) amino acids are activated in presence of UTP (F)
c) tRNA binds amino acid at the anticodon site (F)
d) the process is called translation (T)
611. For synthesis of proteins is required:
a) ATP as the source of energy (T)
b) GTP as the source of energy (T)
c) aminoacyl-tRNA synthase (T)
d) DNA-polymerase (F)
612. Aminoacyl-t-RNA synthase:
a) is enzyme required for transcription (F)
b) is enzyme which activates aminoacid (T)
c) is responsible for peptide bond synthesis (F)
d) requires ATP as a source of energy (T)
613. Steroid hormones can influence gene expression by:
a) binding to membrane receptors (F)
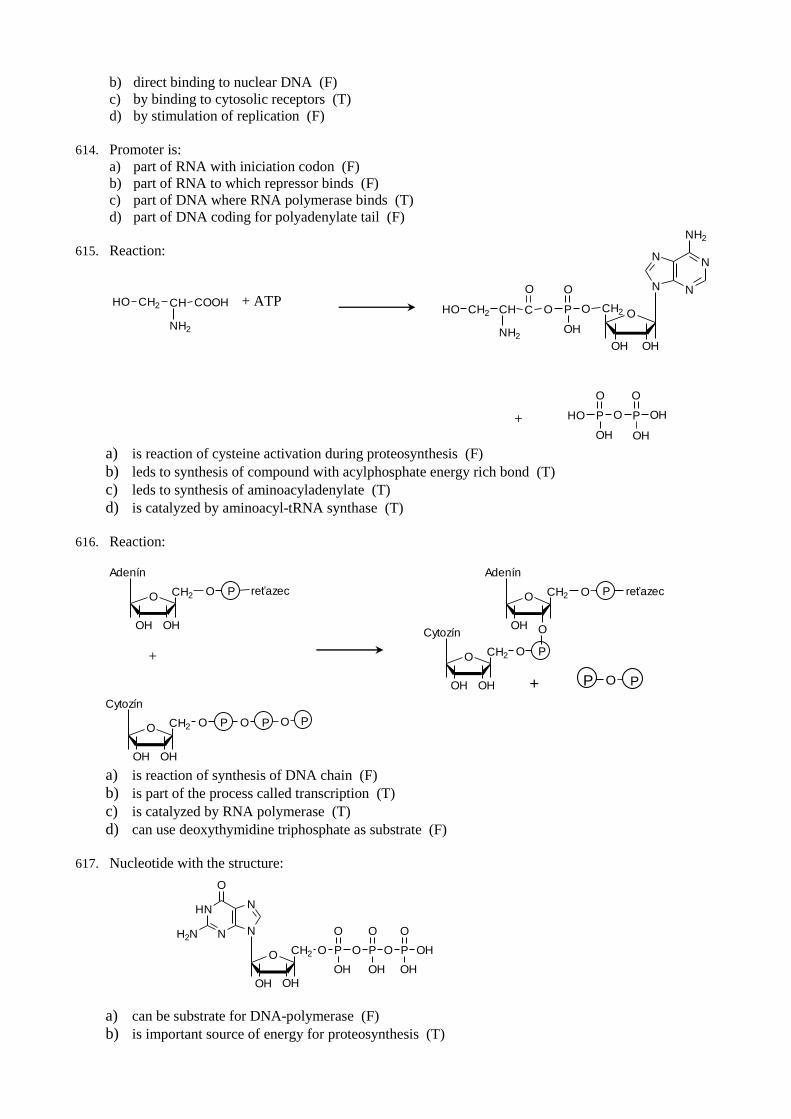
HO CH2 CH COOH
NH2
HO CH2 CH C
NH2
O
O
P O
OH
O
N
NN
N
NH2
O
OHOH
CH2
HO P O
OH
O
P OH
OH
O
Cytozín
O
OH OH
CH2 O P O P O P
Adenín
O
OH O
CH2 O P reťazec
Cytozín
O
OH OH
CH2 O P
P O P+
b) direct binding to nuclear DNA (F)
c) by binding to cytosolic receptors (T)
d) by stimulation of replication (F)
614. Promoter is:
a) part of RNA with iniciation codon (F)
b) part of RNA to which repressor binds (F)
c) part of DNA where RNA polymerase binds (T)
d) part of DNA coding for polyadenylate tail (F)
615. Reaction:
+ ATP
+
a) is reaction of cysteine activation during proteosynthesis (F)
b) leds to synthesis of compound with acylphosphate energy rich bond (T)
c) leds to synthesis of aminoacyladenylate (T)
d) is catalyzed by aminoacyl-tRNA synthase (T)
616. Reaction:
+
a) is reaction of synthesis of DNA chain (F)
b) is part of the process called transcription (T)
c) is catalyzed by RNA polymerase (T)
d) can use deoxythymidine triphosphate as substrate (F)
617. Nucleotide with the structure:
a) can be substrate for DNA-polymerase (F)
b) is important source of energy for proteosynthesis (T)
Adenín
O
OH OH
CH2 O P reťazec
HN
N
N
O
H2NN
O
OH
CH2
OH
O P
O
OH
O P OH
O
OH
O P
O
OH

c) is complementary to thymine nucleotide in DNA (F)
d) is one of substrates in synthesis of RNA (T)
618. Compound with the structure:
a) is substrate for DNA polymerase (T)
b) is substrate for synthesis of primer (F)
c) is complementary to adenine nucleotide in chain of DNA (T)
d) is complementary to uracil in synthesis of RNA (F)
619. Nucleotide with the structure:
a) can be component of rRNA (T)
b) is present in both DNA and RNA (F)
c) is present in tRNA and to ribose of it actvated amino acid is bound (T)
d) in form of polymer is peesent at 5, end of mRNA (F)
620. Bond present in this structure:
a) is 2'-5'-phosphodiester bond (F)
b) is in chain of RNA (T)
c) is broken by RNA-polymerase (F)
d) is formed during transcription (T)
REGULATIONS + RECEPTORS + HORMONES
621. Which of statements about hormones are correct:
a) all hormones are proteins (F)
b) hydrophobic hormones act using intracellular receptors (T)
c) protein hormones use cytosolic receptors (F)
d) hormones are secreted in the same amount during all the life (F)
622. Receptors for hormones:
a) are proteins (T)
HN
N
CH3
O
O
O
H
CH2
OH
O P
O
OH
O P OH
O
OH
O P
O
OH
N
N N
N
NH2
O
OH OH
CH2 O P OH
OH
O
Adenín
O
OH O
CH2 O
Cytozín
O
OOH
H2COPO
OH

b) can be located in cytosol (T)
c) can be located in nucleus (T)
d) all are located in plasma membrane (F)
623. Hydrophobic regulatory compounds can influence cellular processes by:
a) binding to plasma membrane receptors (F)
b) binding to cytosolic receptors (T)
c) binding to nuclear receptors (T)
d) stimulation of cAMP synthesis (F)
624. Cytosolic receptors:
a) are receptors for steroid hormoines (T)
b) can influence gene expression (T)
c) are activators of adenylate cyclase (F)
d) mediate phosphorylation of proteins (F)
625. Which of following receptors belongs to catalytic receptors:
a) glucocorticoid receptor (F)
b) glucagon receptors (F)
c) insulin receptor (T)
d) nicotinic receptor (F)
626. Gi-protein:
a) consists of three subunits (T)
b) in active state binds GDP to -subunit (F)
c) in inactive state binds GTP to -subunit (F)
d) its activation leads to activation of adenylate cyclase (F)
627. Gi-protein:
a) consists of four subunits (F)
b) in active state binds GDP to -subunit (F)
c) in inactive state binds GTP to -subunit (F)
d) its activation leads to activation of adenylate cyclase (F)
628. Adenylatecyclase:
a) breaks cyclic AMP (F)
b) synthesizes cyclic AMP from ADP (F)
c) is present in plasma membrane (T)
d) is present at membranes of endoplasmic reticulum (F)
629. Adenylate cyclase is the enzyme which:
a) is responsible for synthesis of cAMP from ATP (T)
b) is activated by -subunit of Gp protein (F)
c) is activated after glucagon action (T)
d) catalyzes phosphorylation of proteins (F)
630. Hydrophobic regulatory compounds can influence functions of the cells:
a) after their transport to the cells (T)
b) by binding to plasma membrane receptors (F)
c) by binding to intracellular receptors (T)
d) by activation of cAMP synthesis (F)
631. Hydrophobic compounds can influence cellular metabolism by:
a) means of the second messengers (F)
b) means of cytosolic receptors (T)
c) means of nuclear receptors (T)
d) means of induction or repression of gene transcription (T)

632. Protein hormones:
a) are transported through plasma membrane (F)
b) for tranfer of information use G-proteins (T)
c) use for their action the seccond messengers (T)
d) can be bound to cytosolic receptors (F)
633. Phosphodiesterase:
a) catalyzes synthesis of cAMP (F)
b) catalyzes degradation of cAMP (T)
c) is present in cytosol (T)
d) its activity is increased by insulin (T)
634. Gp-protein:
a) is component of plasma membrane (T)
b) in inactive state binds GDP to -subunit (T)
c) in active state binds GTP to -subunit (T)
d) in active state causes increased activity of phospholipase C (T)
635. -subunit of Gs-protein:
a) activates phospholipase C (F)
b) decreases concentration of cAMP (F)
c) binds GTP in active state (T)
d) can hydrolyze GTP (T)
636. -subunit of Gp-protein:
a) activates phospholipase C (T)
b) decreases concentration of IP3 (F)
c) binds GDP in active state (F)
d) can hydrolyze GTP (T)
637. -subunit of Gi-protein:
a) activates adenylate cyclase (F)
b) decreases concentration of cAMP (T)
c) binds GTP in active state (T)
d) can synthesize GTP (F)
638. Inositoltrisphosphate (IP3) is compound which:
a) is formed by cytosolic enzyme (F)
b) is formed by phospholipase C (T)
c) increases concentration of Ca2+ in cytosol (T)
d) is activator of proteinkinase A (F)
639. Receptor for insulin:
a) is present in cytosol (F)
b) is connected with GS protein (F)
c) belongs to catalytic receptors (T)
d) has activity of tyrosine kinase (T)
640. Phosphodiesterase is enzyme which:
a) is responsible for degradation of cAMP (T)
b) is activated by insulin (T)
c) is present in plasma membrane (F)
d) causes phosphorylation of proteins (F)
641. Acetylcholine is compound which:
a) is synthesized by choline acetyltransferase (T)

b) is neurotransmitter of preganglionic neurons of sympathetic nervous system (T)
c) is neurotransmitter of postganglionic neurons of parasympathetic nervous system (T)
d) is degraded by acetylcholine esterase into choline and acetyl-CoA (F)
642. Acetylcholine:
a) is synthesized from acetic acid and choline (F)
b) is synthesized by acetylcholine esterase (F)
c) is neurotransmitter at terminals of parasympathetic nervous system (T)
d) is hydrophobic compound (F)
643. Acetylcholine is inactivated:
a) in presynaptic neurons (F)
b) by acetylcholine esterase (T)
c) by hydrolytic reaction (T)
d) into acetyl-CoA and choline (F)
644. Acetylcholine esterase:
a) is enzyme present in presynaptic cell (F)
b) is responsible for synthesis of acetylcholine (F)
c) is responsible for degradation of cholinergic neurotransmitter (T)
d) by hydrolysis of acetylcholine leads to synthesis of choline and acetic aciod (T)
645. Nicotinic receptors are receptors which:
a) are connected with Gp protein (F)
b) belong to ionic channels (T)
c) are found at neuromuscular juctions (T)
d) are stimulated by acetylcholine (T)
646. Muscarinic receptors:
a) belong to cholinergic receptors (T)
b) are found at terminals of sympathetic nervous system (F)
c) are activated during stress (F)
d) are present for example in heart where they induce inhibition of myocardial contractility (T)
647. M1 receptors:
a) are cholinergic receptors (T)
b) are connected with Gp protein (T)
c) are responsible for stimulation of contractility of smooth muscles (T)
d) are activated by noradrenaline (F)
648. M2 receptorrs are receptors which:
a) are present in heart (T)
b) are activated by acetylcholine (T)
c) after activation cause decrease of cAMP (T)
d) are responsible for supression of frequency of heart beating (T)
649. Which of statements about cholinergic receptors are correct:
a) cholinergic receptors are found at terminals of parasympathetic nervous system (T)
b) muscarinic receptors activate contraction of skeletal muscles (F)
c) activation of nitotinic receptors causes depolarization of postsynaptic cell (T)
d) activation of M1 receptors increases concetration of IP3 (T)
650. Activation of M1 receptors causes:
a) increase of concentration of cAMP (F)
b) decrease of concentration of cAMP (F)
c) increase of Ca2+ concetration (T)
d) contraction of smooth muscles (T)

651. Stimulation of muscarinic receptors in the heart causes:
a) decrease of contractility (T)
b) increase level of K+ ions in cytosol (T)
c) increase of phospholipase C activity (F)
d) decrease of cAMP level (T)
652. Concentration of IP3 and Ca2+ in the cells is increased after activation of:
a) nitotinic receptors (F)
b) M1 receptors (T)
c) all adrenergic receptors (F)
d) 1-receptors (T)
653. Adrenaline (epinephrine):
a) formed in adrenal medulla (T)
b) requires S-adenosylmethionine for the synthesis (T)
c) is inactivated by COMT (T)
d) requires S-adenosylmethionine for degradation (T)
654. The main degradation product of epinephrine is:
a) metanephrine (F)
b) norepinephrine (F)
c) vanilmandelic acid (T)
d) dopamine (F)
655. Acivation of nicotinic receptors:
a) activates adenylate cyclase (F)
b) inhibits adenylate cyclase (F)
c) increases Na+ in cytosol (T)
d) causes depolarization of postsynaptic cell (T)
656. Nerotransmitters of sympathetic nervous system are:
a) catecholamines (T)
b) products of tyrosine metabolism (T)
c) compounds of hydrophobic character (F)
d) compound which stimulate adrenergic receptors (T)
657. Adrenaline (epinephrine):
a) is formed in adrenal medulla (T)
b) requires S-adenosylmethionine for the synthesis (T)
c) is inactivated by COMT (T)
d) requires S-adenosylmethionine for degradation (T)
658. Monoaminooxidase is the enzyme which:
a) is responsible for noradrenaline synthesis (F)
b) is responsible for noradrenaline degradation (T)
c) is responsible for adrenaline degradation (T)
d) requires S-adenosylmethionine (F)
659. Degradation of catecholamines is catalyzed by:
a) tyrosine hydroxylase (F)
b) N-methyltransferase (F)
c) catechol-O-methyltransferase (T)
d) enzyme which requires S-adenosylmethionine as donor of methyl-group (T)
660. Eprinephrine and norepinephrine cause:
a) vasoconstriction by activation of M1 receptors (F)

b) contraction of myocardial muscle by stimulation of -receptors (T)
c) dilation of smooth muscle by stimulation of -receptors (T)
d) glycogenolysis by stimulation of 2-receptors (F)
661. Adrenergic receptors:
a) are activated by acetylcholine (F)
b) are present at terminals of sympathetic nervous system (T)
c) can be devided into M1 and M2 (F)
d) are connected with G-proteins (T)
662. Activation of 1-adrenergic receptors causes:
a) contraction of smooth muscle (T)
b) increase of concentration of IP3 (T)
c) inhibition of adenylate cyclase (F)
d) vasoconstriction for example in skin (T)
663. 1-receptors:
a) are activated by acetylcholine (F)
b) are connected with Gi protein (F)
c) stimulate contranction of skeletal muscles (F)
d) mediate increase of Ca2+ concentration (T)
664. 2-adrenergic receptors:
a) are activated by acetylcholine (F)
b) are connected with Gi protein (T)
c) after stimulation cause relaxation of smooth muscles (F)
d) after stimulation decrease concentration of cAMP (T)
665. Activation of 2-receptors causes:
a) activation of phospholipase C (F)
b) inhibition of adenylate cyclase (T)
c) activation of myocardial contractility (F)
d) activation of hormone sensitive lipase (F)
666. Adrenergic receptors:
a) are present in cytosol (F)
b) are connected with G-proteins (T)
c) are present at terminals of sympathetic nervous system (T)
d) can be divided into nicotinic and muscarinic (F)
667. -adrenergic receptors:
a) are activated by acetylcholine (F)
b) are connected with Gs-protein (T)
c) are present in heart where they stimulate contractility (T)
d) are present in smooth muscles where they cause increased contractility (F)
668. Stimulation of -adrenergic receptors causes:
a) increase of concentration of cAMP (T)
b) activation of phospholipase C (F)
c) increase of concentration of Ca2+ (F)
d) relaxation of smooth muscles (T)
669. Phospholipase C is enzyme which:
a) catalyzes synthesis of phosphotidylinositols (F)
b) is activated by stimulation of M1-receptors (T)
c) is activated by stimulation of 1-receptors (T)

d) is activated by stimulation of 2-receptors (F)
670. Which of following receptors is responsible for inhibition of adenylate cyclase:
a) nicotinic receptors (F)
b) M1-cholinergic receptors (F)
c) M2-cholinergic receptors (T)
d) 2-adrenergic receptors (T)
671. Concentration of cAMP is increased after stimulation of:
a) M1-cholinergic receptors (F)
b) M2-cholinergic receptors (F)
c) 2-adrenergic receptors (F)
d) -adrenergic receptors (T)
672. Glucocorticoids:
a) are formed in the adrenal medula (F)
b) can bind on cytoplasmic receptors (T)
c) bind only on the membrane receptors (F)
d) activate gluconeogenese in the liver (T)
673. Parathyroid hormone:
a) regulates level of kalium in blood (F)
b) its molecula is a polypeptide (T)
c) is formed in the parathyroid a glands (T)
d) is secreted when level of calcium ions in blood is low (F)
674. Aldosterone:
a) has a polypeptide structure (F)
b) is the main mineralocorticoid (T)
c) its secretion is regulated by the renin-angiotensin system (T)
d) inhibits reabsorption of sodium cations from primary urine in the kidney (F)
675. In adipose tissue:
a) insulin activates degradation of triacylglycerols (F)
b) glucagons activates degradation of triacylglycerols (T)
c) adrenalin inhibits degradation of triacylglycerols (F)
d) degradation of triacylglycerols is not regulated by hormones (F)
676. Peptide hormones:
a) pass through cell membranes (F)
b) some of them stimulate formation of a second messenger (T)
c) are epinephrine and thyroxine (F)
d) can transfer signals by means of G-proteins (T)
677. Insulin:
a) is a hormone of the hypothalamus (F)
b) has the structure of a polypeptide (T)
c) decreases level of glucose in blood (T)
d) binds on a cytosolic receptor (F)
678. Glucagon:
a) is formed in the beta cells of the pancreas (F)
b) activates degradation of glycogen in the liver (T)
c) stimulates formation of cAMP in cells (T)
d) is important for metabolism of carbohydrates (saccharides) (T)
679. Hormone receptors:

a) are often glycoproteins (T)
b) are always present in the cytosole (F)
c) can have protein kinase activity (T)
d) can consist of several subunits (T)
680. Glucocorticoids:
a) are secreted from the adrenal cortex (T)
b) activate glycogen degradation in muscle (F)
c) inhibit gluconeogenesis in the liver (F)
d) increase level of cAMP in cells (F)
681. Insulin:
a) increases blood glucose level (F)
b) decreases activity of pyruvate kinase (F)
c) inhibits gluconeogenesis (T)
d) stimulates hydrolysis of lipids (F)
682. Glucagon:
a) is formed in the alfa cells of the pancreas (T)
b) has no effect on a blood glucose level (F)
c) decreases level of cAMP in cells (F)
d) stimulates oxidation of glucose in cells (F)
683. Which of the following hormones is required for transport of glucose into skeletal muscle cells:
a) glucocorticoids (F)
b) insulin (T)
c) glucagon (F)
d) adrenaline (F)
684. We can say about renin-angiotensin system that:
a) renin is a hormone (F)
b) angiotensinogen is formed in the liver (T)
c) angiotensin II activates secretion of cortisol (F)
d) renin has the activity of an enzyme (T)
685. In fatty acid metabolism:
a) insulin activates splitting of triacylglycerols in adipose tissue (F)
b) glucagon inhibits synthesis of fatty acids (T)
c) insulin inhibits acetyl-CoA carboxylase (F)
d) aldosteron does not affect the synthesis of fatty acids (T)
686. Adrenaline (epinephrine):
a) is formed in adrenal medulla (T)
b) requires S-adenosylmethionine for the synthesis (T)
c) is inactivated by COMT (T)
d) requires S-adenosylmethionine for degradation (T)
687. Epinephrine:
a) requires for its synthesis thyrosine (T)
b) is formed in the adrenal cortex (F)
c) requires S-adenosylmethionine for its synthesis (T)
d) activates splitting of glycogen in skeletal muscle cells (T)
688. In the group of steroid hormones can be included:
a) aldosteron (T)
b) thyroxine (F)
c) adrenocorticotropic hormone (F)

d) cortisol (T)
689. On cytoplasmic receptor binds:
a) glucagon (F)
b) cortisol (T)
c) adrenalin (F)
d) hormones of the adrenal cortex (T)
690. The hormone with the structure of a peptide is:
a) estradiol (F)
b) glucagon (T)
c) epinephrine (F)
d) parathyroid hormone (T)
691. Which of the following hormones influence level of calcium in blood:
a) calcitonin (T)
b) parathyroid hormone (T)
c) thyroxine (F)
d) cortisol (F)
AMINO ACIDS
692. Among glucogenic amino acids belong:
a) cysteine, serine, alanine (T)
b) amino acid which is formed by transamination of pyruvate (T)
c) alanine, leucine (F)
d) glutamate, aspartate (T)
693. Glutamate:
a) is glucogenic amino acid (T)
b) by oxidative deamination is converted to 2-oxoglutarate by help of glutamate dehydroganase (T)
c) is important donor of amino group for synthesis of essential amino acids (F)
d) is part of tripeptide which is needed in oxidation-reduction reactions (T)
694. Oxidases of amino acids:
a) have coenzyme pyridoxal-5-phosphate (F)
b) belong to aerobic dehydrogenases (T)
c) have coenzymes FAD or FMN (T)
d) are located mainly in peroxisomes of hepatocytes (T)
695. Which of following amino acids enter Krebs cycle through succinyl-CoA:
a) glutamate (N)
b) methionine (S)
c) valine (S)
d) leucine (N)
696. Which of following amino acids enter Krebs cycle through ketoglutarate:
a) all glucogenic amino acids (N)
b) all ketogenic amino acids (N)
c) histidine (S)
d) arginine (S)
697. Glutamate dehydrogenase:
a) is located in cytosol of cell (F)
b) catalyzes reversible reaction located in mitochondria (T)
c) reduced form of its coenzyme gives in respiratory chain 3 ATP (T)

d) plays role in synthesis of non essential amino acids where is needed reductive amination of
oxoglutarate (T)
698. Glutamate dehydrogenaseis enzyme which:
a) catalyzes oxidative deamination of glutamate (T)
b) can be used for synthesis of glutamate (S)
c) uses FMN as coenzyme (N)
d) allows synthesis of 3 ATP by oxidative phosphorylation (S)
699. Glutamate dehydrogenase:
a) is enzyme present in mitochondria (S)
b) catalyzes synthesis of ketoglutarate from glutamate (N)
c) can use ketoglutarate as substrate (S)
d) participates in indirect deamination of amino acids (S)
700. Decarboxylation of glutamate:
a) requires FMN as the coenzyme (F)
b) leads to synthesis of gamma aminobutyrate (T)
c) is important for synthesis of phospholipids (F)
d) leads to synthesis of inhibitory neurotransmitter (T)
701. Indirect deamination of amino acids requires:
a) coenzyme pyridoxal-5-phosphate (in the first phase) (T)
b) coenzyme NAD (in the second phase) (T)
c) specific amino transferase (T)
d) acetoacetate (F)
702. Ammonia is transported from tissues to the liver in a form of:
a) urea (F)
b) uric acid (F)
c) glutamine (T)
d) compound which is formed by glutamine synthetase which requires ATP (T)
703. Serine is deaminated:
a) by serine dehydratase coenzyme of which is FMN (F)
b) by deamination which belong to direct deaminations (T)
c) by serine dehydratase coenzyme of which is active form of vitamine B6 (T)
d) and product of reaction is oxaloacetate (F)
704. Cysteine:
a) if is deaminated product is compound important for gluconeogenesis (T)
b) is substrate for formation of glutathion (T)
c) is deaminated by cysteine desulfhydrase and coenzyme is NAD (F)
d) product of deamination is 2-oxo-glutarate (F)
705. By decarboxylation:
a) of amino acid is formed primary amine (T)
b) are formed amines important in regulatory reactions (T)
c) of serine is formed histamine (F)
d) of amino acid with hydroxy group in molecule is formed amine important for synthesis of
phospholipids (T)
706. Coenzyme of decarboxylases of amino acids is:
a) active form of vitamine B5 (F)
b) active form of vitamine B6 (T)
c) coenzyme which is coenzyme of cysteine desulfhydrase (T)

d) coenzyme which is common for AST, ALT and also for decarboxylases of amino acids (T)
707. Essential amino acids:
a) must be taken by food (T)
b) are those which can be synthetised by organism (F)
c) are e.g. valine, leucine and isoleucine (T)
d) can be substrate for synthesis of conditionally essential amino acids (T)
708. Aspartate aminotransferase:
a) utilizes aspartate + 2-oxoglutarate as the substrates (T)
b) utilizes oxaloacetate + glutamate as the substrates (T)
c) is located in mitochondria (T)
d) requires NAD as coenzyme (F)
709. Ammonia:
a) is released from amino acids by amino acid oxidases (T)
b) causes increase of pH (T)
c) in the kidney is detoxified into urea (F)
d) in the blood is transported as glutamate (F)
710. Alanine aminotransferase:
a) utilizes alanine as substrate (T)
b) is part of glucose-alanine cycle (T)
c) has pyridoxal phosphate as the coenzyme (T)
d) is the enzyme typical for the liver tissue (T)
711. Glutamine:
a) is formed by decarboxylation of glutamate (F)
b) is intermediate of urea cycle (F)
c) is transport form of ammonia from tissues to the liver (T)
d) its synthesis from glutamate requires ATP (T)
712. Decarboxylation of glutamate:
a) requires NAD as the coenzyme (F)
b) leads to the synthesis of gamma aminobutyrate (T)
c) catalyzes synthesis of primary amine (T)
d) needs ATP as a source of energy (F)
713. Glutamate dehydrogenase:
a) is enzyme present in mitochondria (T)
b) catalyzes synthesis of 2-oxoglutarate (T)
c) catalyzes irreversible reaction (F)
d) participates in indirect deamination of amino acids (T)
714. Ammonia:
a) can be formed by serine dehydratase (T)
b) its synthesis by amino acid oxidases needs NAD (F)
c) is detoxified into uric acid in humans (F)
d) is transported in the blood as glutamate (F)
715. Decarboxylation of serine:
a) synthesizes pyruvate (F)
b) produces compound requires for synthesis of choline (T)
c) requires pyridoxalphosphate as coenzyme (T)
d) is important for phospholipids synthesis (T)
716. Ammonia:

a) is formed by glutamate dehydrogenase (T)
b) is toxic compound for living cells (T)
c) in kidney is released by glutaminase (T)
d) in cells binds to glutamate (T)
717. Pyruvate:
a) is compound formed from aspartate by its decarboxylation (F)
b) is product of ALT (T)
c) is substrate of ALT (T)
d) is formed from serine in the presence of FMN (F)
718. Alanine aminotransferase:
a) is enzyme typical for the liver tissue (T)
b) utilizes pyruvate as the substrate (T)
c) is part of malate shuttle (F)
d) uses thiamine as coenzymes (F)
719. Carbamoylphosphate:
a) brings to urea 2 NH2 groups (F)
b) is released from arginine (F)
c) during its synthesis ATP is formed (F)
d) is compound through which free ammonia enters urea cycle (T)
720. During urea cycle:
a) ammonia is detoxified to uric acid in humans (F)
b) aspartate enters by its reaction with ornithine (F)
c) fumarate is released from argininesuccinate (T)
d) 3 ATP are formed (F)
721. Urea (NH2)2CO:
a) in human is final product of purine nucleotides degradation (F)
b) is well soluble in water and easily cross membranes (T)
c) is compound which is synthetised in the liver (T)
d) in ureotelic organisms is final product of detoxification of ammonia (T)
722. Enzymes of urea cycle:
a) are inductive and their amount is increased during increased catabolism of proteins (T)
b) are costitutive and their amount is increased mainly during starvation (F)
c) their production is stimulated by glucocorticoids (T)
d) are comletely present only in the liver (T)
723. Universal acceptor of of amino group from majority of all amino acids can be:
a) oxaloacetate (F)
b) 2- oxoglutarate (T)
c) 5- carbon oxoacid which is intermediate of Krebs cycle (T)
d) oxoacic which is formed in metabolism of ketone bodies (T)
724. Pyruvate:
a) is formed from alanine by the action of AST (F)
b) is formed by deamination of serine (T)
c) in the liver is used for gluconeogenesis (T)
d) for its synthesis from cysteine NAD is required (F)
725. During urea cycle:
a) aspartate enters by its reaction with citrulline (T)
b) urea is formed by hydrolysis of arginine (T)
c) 3 ATP are formed (F)

NH2CH
COOH
CH2
CH2
CH2
NH
C
NH2
NH
+ H2O
NH2CH
COOH
CH2
CH2
CH2
NH2
+
NH2
C
NH2
O
d) argininosuccinate is brokeninto arginine (T)
726. Glutamine:
a) for its synthesis ATP is required (T)
b) is transport form of ammonia from tissues to the liver (T)
c) is product of glutamate decarboxylation (F)
d) in kidney tubular cells is hydrolyzed releasing ammonia (T)
727. Aminoacid which is donor of one amino group in urea:
a) after transamination with 2-oxoglutarate can enter Krebs cycle (T)
b) belongs to the neutral amino acids (F)
c) is amino acid which creates bigger part of carbon sceleton of pyrimidine ring (T)
d) is formed by transamination of pyruvate (F)
728. Ornithine transcarbamoylase:
a) is enzyme of urea cycle (T)
b) is enzyme locatedin cytosol of hepatocyte (F)
c) is enzyme located in mitochondria of periportal hepatocyte (T)
d) catalyzes production of arginine from ornithine and carbamoyl phosphate (F)
729. Arginase:
a) is located in cytosol of hepatocyte (T)
b) is hydrolase and cleveas arginine to ornithine and urea in cytosol of hepatocyte (T)
c) is hydrolase and belongs to the enzymes of Krebs cycle (F)
d) is enzyme required for synthesis of arginine (N)
730. Reaction
a) is catalyzed by hydrolytic enzyme (N)
b) is located in cytosol of liver cells (S)
c) uses as substrate compound for synthesis of which aspartate is required (S)
d) is reaction of urea cycle (S)
731. Reaction
a) proceeds in cytosol of periportal hepatocytes (S)
b) is catalyzed by arginase (S)
c) is reaction imúportant for detoxification of ammonia by uric acid synthesis (N)
d) is reversible reaction (N)
NH2CH
COOH
CH2
CH2
CH2
NH
C
NH2
N CH
CH2
COOH
COOH
NH2CH
COOH
CH2
CH2
CH2
NH
C
NH2
NH
CH
CH
COOH
COOH
+

NH2CH
COOH
CH2
CH2
COOH
+NH3
OC
COOH
CH2
CH2
COOH
+ H2O+ NAD++NADH+H
+
732. Compound
a) is substrate for synthesis of compound: (S)
b) is copmpound through which free ammonia enters urea cycle (S)
c) is synthesized by carbamoylphosphate synthase I in mitochondria of liver cells (S)
d) can be synthesized in cytosol by carbamoylphosphate synthase II (S)
733. Which of following statements about this reaction are correct
a) reaction is catalyzed by glutaminase (N)
b) it is reaction of indirect deamination of amino acids (N)
c) product of reaction represents non-toxic transport form of ammonia (S)
d) product of reaction is important also for regulation of acid-base balance in kidneys (S)
734. Compound
a) is amide of aspartic acid (F)
b) is the most important transport form of ammonia (T)
c) is formed by glutamine synthase (F)
d) is liver is important for regulation of acid-base balance (F)
735. Reaction:
a) is catalyzed by glutaminase (T)
b) requires ATP as source of energy (F)
c) is kidneys is important for regulation of pH of urine (T
d) produces free ammonia which is used for urea synthesis in liver (T
736. Reaction
a) is catalyzed by glutamate dehydrogenase (T)
b) is catalyzed by amino acid oxidases (F)
c) is important for synthesis of non-essential amino acids (T)
C
O
O
NH2
P OH
OH
O
NH2CH
COOH
CH2
CH2
COOH
+ NH3 + ATPNH2CH
COOH
CH2
CH2
CO
+
NH2
ADP + P
NH2CH
COOH
CH2
CH2
CO NH2
NH2CH
COOH
CH2
CH2
CO NH2
+ H2O
NH2CH
COOH
CH2
CH2
COOH
+ NH3
NH2
CH COOHCH2CH2CH2
NH
C
NH2
O

NH2CH
COOH
CH2
CH2 S CH3
ATPN
NN
N
NH2
O
OHOH
CH2
NH2CH
COOH
CH2
CH2 S
CH3
+
d) in high concentration of ammonia decreases amount of ketoglutarate in Krebs cycle which leads
to decreased ATP synthesis (T)
737. Which of statements about this reaction are correct:
a) reaction is catalyzed by glutamate dehydrogenase (T)
b) reaction is one step of indirect deamination of amino acids (T)
c) substrate of reaction is formed by transaminase (T)
d) reaction is located in cytosol (F)
738. Aspartate aminotransferase
a) catalyzes reversible reaction (T)
b) catalyzes reaction: (T)
c) is important for utilization of aspartate in gluconeogenesis (T)
d) uses as coenzyme (F)
739. Compound
a) is substrate of aspartate aminotransferase (T)
b) is product of aspartate aminotransferase (T)
c) can be synthesized from oxaloacetate (T)
d) is compound for synthesis of which pyridoxalphosphate is required (T)
740. S-adenosylmethionine:
a) is formed by reaction (T)
b) is used for creatine synthesis (T)
c) is source of methyl group in synthesis of adrenaline (T)
d) is source of methyl group in synthesis of thymine (F)
741. Decarboxylation of serine
a) leads to synthesis of -alanine (F)
b) requires pyridoxalphosphate as coenzyme (T)
c) is important for phospholipid synthesis (T)
OC
COOH
CH2
CH2
COOH
++NH2CH
COOH
CH2
COOH
NH2CH
COOH
CH2
CH2
COOH
OC
COOH
CH2
COOH
NH2CH
COOH
CH2
COOH
C
CH2 OH
NH3C
HO
OH
PLP
CO2
NH2
CHHOOC CH
OH
CH3
CH2
CH2
OH
NH2
NH2CH
COOH
CH2
CH2
COOH
+ NH3
OC
COOH
CH2
CH2
COOH
+ H2O + NAD+ + NADH+H
+

d) proceeds by reaction: (F)
742. Transamination of alanine:
a) requires amino acid oxidase (F)
b) requires paridoxalphosphate as coenzyme (T)
c) is important for utilization of alanine in gluconeogenesis (T)
d) proceeds by reaction
(T)
WATER + MINERALS, ACID-BASE BALANCE
743. Water:
a) represents 20% of body weight (F)
b) transport water soluble copounds like bilirubin (F)
c) is needed for thermoregulation (T)
d) its absorption in the kidney is stimulated by renin (F)
744. Hyperhydration can be caused by:
a) hypersecretion of vasopresin (T)
b) diabetes mellitus (F)
c) cardiac diseases (T)
d) diabetes insipidus (F)
745. Atrial natriuretic peptide:
a) is secreted in decrease of blood pressure (F)
b) increase cAMP content in adrenal coretx (F)
c) increases cGMP content in adrenal medulla (T)
d) decreases aldosterone secretion (T)
746. Angiotensin is the compound which:
a) stimulates vasopresin secretion (F)
b) inhibits aldosteron secretion (T)
c) is formed in blood (T)
d) causes vasodilation (F)
747. Sodium reabsorption in kidney is increased by:
a) cotisol (F)
b) vasopressin (F)
c) ACTH (F)
d) aldosteron (T)
748. Intracellular fluid:
a) has as main cation K+ (T)
b) represents 20% of body weight (T)
c) has as a main anion Cl– (F)
d) is hyperosmolar to ECF (F)
749. Aldosteron secretion is stimulated by:
a) angiotensin formed in juxtaglomerular cells (F)
COOH
CH NH2
CH3COOH(CH2)2
C O
COOH
+PLP
ALT
COOH
C O
CH3COOH(CH2)2
CH
COOH
+ NH2

b) renin formed in the liver (F)
c) angiotensin II (T)
d) ACTH (T)
750. Which of statement about water in the body are correct::
a) represents 60% of body weight (T)
b) is polar solvent (T)
c) its content is regulated by glucocorticoids (T)
d) its content is regulated by vasopresin (T)
751. Extracellular fluid:
a) represents 60% of body weight (F)
b) contains Na+ as main cation (T)
c) contains Cl– as main cation (F)
d) contains proteins in lower concntration than interstitial fluid (F)
752. Volume of water:
a) in extracellular fluid depends mainly on concentration of Na+ K+ (F)
b) in extracellular fluid depends mainly on concentration of Na+ and Cl– (T)
c) influences blood pressure (T)
d) is regulated by antidiuretic hormone (T)
753. Which of following factors participates in regulation of water and minerals metabolism:
a) hormone of adrenal medulla – aldosterone (F)
b) aldosterone which is secreted after activation of renin-angiotensine system (T)
c) aldosterone which increases excretion of Na+ (F)
d) vasopressin which increases absorption of water in collecting ducts in kidneys (T)
754. Dehydration may occur:
a) in diabetes mellitus (T)
b) in diabetes insippidus (T)
c) in increased production of antidiuretic hormone (F)
d) in overproduction of aldosterone (F)
755. What is the meaning of Na+ in the organism:
a) it is the main extracellular cation (T)
b) binds water in intracellular fluid (F)
c) influences membrane potentials and excitability of cells (T)
d) influences osmotic pressure of extracellular fluid (T)
756. Vasopresin:
a) is formed in anterion pituitary (F)
b) is secreted when plasma osmotic pressure is low (F)
c) is formed in hypothalamic cells (T)
d) increases sodium absorption in kidney (F)
757. Main extracellular cation::
a) is K+ , concetration of which in extracellular fluid is 135 – 145 mmol/l (F)
b) is cation content of which ios regulated by aldosteron (T)
c) is Na+ (T)
d) is cation which is also component of main buffer system in blood (T)
758. Sodium is cation which:
a) in extracellular fluid is in concentration 135 – 145 mmol/l (T)
b) together with K+ and Cl- participates in membrane potentials (T)
c) influences volume of extracellular fluid (T)

d) is is transorted into the cells by Na+K+-ATP-ase (F)
759. Which of statements about potassium are correct:
a) it is main extracellular cation (F)
b) its concentration in blood úplasma is 100 mmol/l (F)
c) together with Na+ participates ibn membrane potentials (T)
d) its content in body is regulated by hormone of adrenal cortex (T)
760. Potassium:
a) is main intracellular anion (F)
b) is exctreted by urine (T)
c) is transported into the cells by Na+,K+–ATPase (T)
d) is cation concentration of which influences excitability of the cells (T)
761. Hyperkaliemia occurs in:
a) increased aldosterone secretion (F)
b) catabolic state (T)
c) alkalosis (F)
d) acidosis (T)
762. Decreased potassium concentration::
a) can be caused by high secreation of aldosteron (T)
b) can be caused by lover secretion of renin (F)
c) can be caused by alkalosis (T)
d) is typical for catabolic state (F)
763. Which of following factors participates in regulation of K+:
a) steroid hormone formed in zona glomerulosa (T)
b) aldosterone sectretion of which is regulated by renin-angiotensin system (T)
c) mainly liver (F)
d) H+,K+-ATP–ase located in parietal cells (F)
764. What is range of pH compatible with life:
a) 7,35 - 7,45 (F)
b) 7,0 - 7,6 (F)
c) 6,8 - 7,8 (T)
d) pH is not important for functions of living system (F)
765. What is physiologic range of pH:
a) 6,8 - 7,8 (F)
b) 7,35 - 7,45(T)
c) 7,0 - 7,2 (F)
d) 7,65 - 7,85 (F)
766. Which of following buffers is the most important in blood:
a) bicarbonate buffer (T)
b) phosphate buffer (F)
c) glutamine (F)
d) biffer which is regenerated by kidneys (T)
767. Which of following systems are important in regulation of acid-base balance:
a) bicarbonate buffer in the blood (T)
b) phosphate buffer in the blood (F)
c) kidneys by ability excretion of H+ and regeneration of HCO3– (T)
d) lungs by production of CO2 (F)
768. Phosphate buffer:

a) consists of H3PO4 + Ca3PO4 (F)
b) consists of NaH2PO4- + Na2HPO4
2- (T)
c) is the most important buffer in the blood (F)
d) is important in urine where binds protons (T)
769. Phosphate buffer consists of:
a) H3PO4+ Ca3PO4 (F)
b) H3PO4+ Ca(OH)2 (F)
c) NaH2PO4+ Na2HPO4 (T)
d) H3PO4 + NaCl (F)
770. Carboanhydrase in tubular cells:
a) catalyzes spliting of H2CO3 (F)
b) synthesizes H2CO3 from H+and HCO3- (F)
c) is required for regeneration of HCO3- (T)
d) utilizes CO2 and H2O as the substrates (T)
771. Carboanhydrase is enzyme which:
a) catalyzes synthesis of H2CO3 in erythrocytes (T)
b) catalyzes synthesis of H2CO3 in kidney tubular cells (T)
c) catalyzes dissociation of H2CO3 into H+ + HCO3- (F)
d) catalyzes reversible reaction which in lungs produces CO2 (T)
772. Glutaminase:
a) synthesizes glutamine (F)
b) releases NH3 from glutamate (F)
c) participates in regulation of intracellular pH (F)
d) is present in kidney tubular cells (T)
773. Protons secreted by tubular cells into urine can be boud to:
a) glutamine (F)
b) H2PO4– (F)
c) ammonia (T)
d) HPO42– (T)
774. Glutamine in regulation of acid base balance:
a) releases ammonia which decreases acidity of urine (T)
b) regulates acid base balance in blood (N)
c) is the most important buffer in intracellular fluid (N)
d) binds ammonia in alkalosis (F)
775. In case of metabolic acidosis:
a) concentration of HCO3- is decreased (T)
b) acidosis is compensated by hyperventilation (T)
c) kidney excretes more H+ (T)
d) regeneration of HCO3- in kidneys is decreased (F)
776. Which of following compounds may cause acidosis in diabetics:
a) lactate (F)
b) acetone (F)
c) acetoacetate (T)
d) -hydroxybutyrate (T)
777. Respiratory acidosis is compensated by:
a) hyperventilation (F)
b) decreased regeneration of HCO3- (F)
c) increased formation of HCO3- in lungs (F)

d) increased formation of HCO3- in kidney (T)
778. Respiratory acidosis:
a) is caused by hypoventilation (T)
b) is compensated by hypoventilation (F)
c) is state when concentration of CO2 (H2CO3) is increased (T)
d) is compensated by liver (N)
b) is compensated by hyperventilation
c) kidney excretes more H+
d) production of HCO3- in kidney is increased
779. Metabolic acidosis is consequence of:
a) diabetes mellitus (T)
b) long starvation (T)
c) increased concentration of lactate (T)
d) increased ventilation (F)
780. Respiratory alkalosis:
a) is caused by hyperventilation (T)
b) can be caused by activation of respiratory center (T)
c) is compensated by pyperventilation (F)
d) is compensated by increased excretion of HCO3– by kidneys (T)