-
8/8/2019 Carbohydrate Metabolism 1
1/9
LEGEND :
= Clinical correlation;
= Step number (Glycolysis)
= Positive regulation
= Negative regulation
= increase/d
=decrease/d
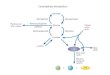
Glucose : C 6 H 12 O 6 Beginning or end of major pathways of
carbohydrate
metabolism Major form in which carbohydrates absorbed form GIT
is
presented to cells Major fuel for the brain Biomedical
importance
o Provide ATP in absence of oxygen allowing tissues to survive
anoxic episodes
o Heart muscle is adapted to aerobic performance glycolytic
pathway; poor survival under
conditions of ischemia or myocardialinfarction
hemolytic anemia: pyruvate kinase deficiencyfatigue:
phosphofructokinase deficiencycancer cachexia: lactate;
gluconeogenesis;
hypermetabolism lactic acidosis: impaired activity of
pyruvate
dehydrogenaseRela t ionsh ip of Glucose to Major pa thways
of
Carbohydra te Metabol i sm
GLYCOLYSIS (Harpers Illustrated Biochemistry Chapter 18) A.k.a.
Embden-Meyerhoff Pa thway A Catabolic Pathway (TCA = Amphibolic
Pathway) Used by all cells to extract energy from glucose Aerobic
or Anaerobic
A) Aerobic glycolysis Pyruvate will be oxidized to CO 2 ,H2 O,
and Energy (ATP);
B Anaerobic glycolysis
3 S tages of Glycolys i s
I. Pr imary S tage
D-Glucose + 2ATP D-fructose 1,6Bisphosphate +2ADP + 2H
A.k.a. Trapping stage trapping of glucose in formof Glucose
6-Phosphate (G6P) with the utilizationof ATP. Phosphorylation with
a negatively charged
PO 4 to Glucose forming G6P, glucose is preventedfrom moving
outside of the cell therefore trapping it inside the cell.
In von Gierke disease , aka Type I Glycogensosis(Glycogen
storage disease) deficiency of Glucose-6-Phosphatase (G6Pase) which
catalyzes thehydrolysis of G6P to D-Glucose, preventing G6P tobe
converted to D-glucose which can be
transported outside the cell. This results in theaccumulation in
the liver of excessive amounts of normal glycogen.
II. Spl i t t ing S tage
D-fructose 1 ,6-Bisphosphate 2 D-Glycera ldehyde 3-Phosphate
Splitting of FBP into two Tr iose phosphates(G3P)
III. Oxido- reduc tase Phosphory la t ion S tage
2D-G3P + 4ADP + 2Pi + 2H + 2Lacta te + 4ATP +2H 2 O
Earning stage formation of ATP
* Stage I and II are the investment stage (Investmentof 2
ATPs).
SUBJECT: BIOCHEMISTRY
TOPIC: CARBOHYDRATEMETABOLISM 1 (GLYCOLYSIS
&GLUCONEOGENESIS)
LECTURER: DR. UY
DATE: DECEMBER 2010
#
-
8/8/2019 Carbohydrate Metabolism 1
2/9
GLYCOLYSISSTEP 1 - Phosphory la t ion by
Hexokinase /Glucokinase ( I r revers ib le )- Trapping of
glucose by phosphorylation to G6P
DIFFERENCE BETWEEN HEXOKINASE AN DGLUCOKINASE
*This is wherethe 1 st ATP is invested.
Hexokinase :- Low Km for glucose relative to its concentration
in
blood. (high affinity for glucose)- Abundant in the Muscles
o Hexokinase immediately reacts even with alow concentration of
glucose.
o This rapid reaction of the enzyme isimportant for G6P to be
readily available in
the muscles.- Strongly inhib i ted by its product Glucose 6-
Phosha te (G6P) hence its reaction is not atequilibrium.o When
there is enough G6P in the muscle
Hexokinase is inhibited. (Low Vmax)- In liver, hexokinase is
saturated under normal
conditionGlucokinase (GK) :
- S 0.5 for glucose is higher than the Km for glucose of other
kinases
- less sens i t ive to product inhibition by G6P(G6P does not
inhibit Glucokinase )
- contribute to the capacity of the liver to bufferblood glucose
levels because of the following
GLUT4 (in the muscles and adipose tissues)insulin dependent
transporters
In DM type 1 patients, lack of insulin causesfatigue due to lack
of ATP caused by thelack of glucose transported inside.
GLUT2 (in liver and pancreas)
GKRP attaches to glucokinase in its inactive form in thenucleus
forming a complex with GK.
With the increase in glucose concentration in the cell,release
of GK by GKRP into the cytosol is promoted converting GK to its
active form. While the increase of Fructose 6-Phosphate (F6P)
signals the inactivation of GKback into the nucleus with GKRP
therefore inhibiting itsactivity.
M ATURITY-ONSET D IABETES OF THE YOUNG (MODY)Type 2
An autosomal dominant disorder involving mutations in the
glucokinase (GCK) gene.
Patients have progressive hyperglycemia that is
usuallyasymptomatic at diagnosis and is usually managedwi th d ie t
a lone .
STEP 2 I somer isa t ion by Phosphoglucoseisomerise
(Reversible)
1
2
Stage I and !! :Investmentphase
Stage III : Energygeneration phase
-
8/8/2019 Carbohydrate Metabolism 1
3/9
-
8/8/2019 Carbohydrate Metabolism 1
4/9
ALDOLASE A ( found in RBCs and musc le )
- Absence of the enzyme (aldolase) may causenonspherocytic
hemolytic anemia.
*most/almost all of glycolytic enzyme deficienciesmanifest
hemolytic anemia because this causes thepumps in the membrane of
RBC do not functionproperly. This allows the RBC to be exposed
tooxidative properties which causes hemolysis or
swelling of the RBC.
*In this disease however the hemolytic anemia isnonspherocytic
which means the RBCs are not
spherical or sphere-shaped like most hemolyticanemia.
TRIOSE PHOSPHATE ISOMERASE (TPI)
- Patients with TPI deficiency have neonatal hemolyticanemia and
progressive neurologic involvement
- Progressive hypotonia with eventual diaphragmparalysis and
cardiomyopathy.
STEP 5 Most DHAP are conver ted to G3Pwhich i s u t i l i zed in
the g lycoly t ic pa thway.
*because most DHAP is converted to G3P there should be2
molecules of G3P converted to Pyruvate.
STEP 6 ox ida t ion by G3P dehydrogenase
As previously mentioned most of the DHAP will beconverted to
G3P, which G3P is the one utilized in theformation of Pyruvate,
therefore from everything beginning in step 5 happens in pair.
In this step, G3P is converted to a highenergy compound 1 ,3
-Bisphosphoglycera te (1 ,3BPG) by Glycera ldehyde 3-phosphate
dehydrogenase(G3PDH or GAPDH) the reduction of NAD+ to NADH+H
+which will form (3)ATP* in the ETC or utilized by the
pyruva te lac ta te lac ta te
dehydrogenase
9
10
4
5
x 26
x 2
7
8
-
8/8/2019 Carbohydrate Metabolism 1
5/9
STEP 7 1 st subs t ra te leve l Phosphory la t ion
byPhosphoglycera te k inase (PGK) (and ADP ATP)and format ion of 2
,3BPG byproduct .
At this step,
1,3BPG is converted to 3-Phosphoglycera te by transferring of
the phosphate to ADP forming ATP. This is the first substrate level
phosphorylation in Glycolysis. 2ATPsshould be formed because there
are 2 molecules of 1,3BPG from the splitting. At this point of
glycolysis the2ATPs invested in steps 1 and 3 has already been
regained.
There is also the presence of an enzyme mutase ,abundant in the
RBC, which may form a byproduct 2,3BPG from 1,3BPG . 2,3BPG shifts
the oxygen dissociation curveto the right releasing more oxygen to
the tissues. 2,3BPGcan then be converted to 3-Phosphoglycera te
(3PG) with the use of the enzyme Phosphatase .
*2,3BPG or diphosphyglycerate/DPG is also likely anintermediate
of the reaction.
* However, this pathway does not produce ATP.
*PGK is inhibited by arsenate
Arsenic tox ic i ty arsenate competes with theinorganic
phosphate (P i) hydrolyzing 3PG to 2PGwithout ATP formation
STEP 8 I somer iza t ion by Phosphoglycera temutase
3-Phosphoglycerate is isomerized to 2-Phosphoglycerate
STEP 9 Dehydra t ion ca ta lyzed by Enolase
2-Phosphoglycera te removes H 2 O to form a high energy
phosphate compound phosphoenol pyruva te (PEP)
*enol compounds=high energy compounds
Enolase is inhibited by Flouride ; used to inhibitglycolysis in
blood samples used for measuring glucose.
STEP 10 2 nd Subs t ra te leve l phosphory la t
ion(Irreversible)
At this step, phosphate is transferred from PEP to ADP bypyruva
te k inase(PK) forming 2 molecules of ATP perglucose. Since the
invested ATPs were already regained,
this 2 nd substrate level phosphorylation is the first gaining
of ATP from glucose. PK is allosterically activated byFBP (product
of the rate limiting step page2)
Pyruvate Kinase Def ic iency & Hemoly t icAnemia
Lack of ATP affects ion pumps especially Na + /K + ATPase.
causing the cells to swell & lyse
During starvation , g lucagon is increased. This stimulates the
activity of adenyly l cyc lase which converts ATP tocAMP. cAMP then
activates PKA. PKA phosphorylatesPyruvate kinase(PK) turning it
into its inactive form.Therefore, with decreased glucose glycolysis
decreaseproduction of pyruvate with the inactivation of PK.
*kinases are inactive when phosphorylated similar to PFK-2
During Anaerobic glycolysis pyruva te forms Lacta tecatalyzed by
the enzyme Lacta te dehydrogenaseutilizing with it. NADH+H + from
the activity of G3Pdehydrogenase
2 ATP consumpt ion with theenzymesHexokinase/Glucokinase
andPhosphofructoki nase-1 (PFK-1).
2NADH+2H + p roduced fromG3P dehydrogenase .
4ATPproduced from(2) Phospho-
glyceratekinase (PGK)and (2) (PK Pyruvatekinase )
In anaerobicpathway, 2NADH
-
8/8/2019 Carbohydrate Metabolism 1
6/9
3 IRREVERSIBLE STEPS OF GLYCOLYSIS WHICH ARE ALSOREGULATORY
STEPS
*All are kinase enzymes.
NADH Generated by Glycolysis has to be oxidized back toNAD+.
This is done by the enzyme LactateDehydrogenase(anaerobic) &
Substrate Shuttles(aerobic).
In Aerobic glycolysis, NADH produced by G3PDH needs tobe
transported into the mitochondria for ATP synthesis in the ETC,
however, NADH is NOT permeable to the innermitochondrial membrane.
By utilizing substrates (Malateand Aspartate) permeable to the
inner mitochondrialmembrane, NADH is made available to the ETC in
themitochondria.
OAA + NADH Malate Dehydrogenase Malate + NAD+
Malatedehydrogenase (MDH) Oxaloace ta te OAA Mala te
which is also permeable to the inner mitochondrialmembrane then
exits the mitochondria and converted back
to OAA by AST.
Fate of the PYRUVATE
Carboxylation of Pyruvate toOxaloace ta te (OAA) iscatalyzed by
Pyruvatecarboxylase .
(Oxidative) Decarboxylation of Pyruvate to Acety l CoA
iscatalyzed be Pyruvate deydrogenase complex along with
the reduction of NAD +. Acetyl CoA will proceed to TCA cycle
.The Pyruvate dehydrogenase complex is a multienzymecomplex which
consists of Pyruvate Dehydrogenase,Dihydrolypoyl Tranacetylase,
Dihydrolipoyl Dehydrogenase.
(The succeeding discussion on Oxidative decarboxylation was
notdiscussed but it was in the PPT and is also found in the
Harperson chapter 18, page 153, The oxidation of pyruvate to
Acetyl-CoAis the irreversible route from glycolysis to the citric
acid cycle.)
During the conversion the intermediates do not dissociate
and remain bound to the components of the
multienzymecomplex.
MECHANISM: (OXIDATIVE CARBOXYLATIONOF PYRUVATE)
Pyruvate is decarboxylated to a hydroxyethyl (Acyl) bound to
thiamine diphosphate (TPP or TDP) with the enzrymePyruvate
dehydrogenase, forming Acyl-TPP. Acyl-TPP reactswith an oxidized
lipoamide (Lip-S 2 ) catalyzed be theDihydrolipoyl Transacetylase
component, forming Acyl-Lipoate/Acetyl lipoamide (in Harpers).
Acyl-lipoate will thenreacts with Coenzyme A(CoA-SH) forming Acetyl
CoA and
the reduced lipoamide. However lipoamide has to bereturned to
its original form which is reoxidized by theDihydrolipoyl
Dehydrogenase(flavoprotein componentcontaining an FAD, forming FADH
and Lip-S 2 . DihydrolipoylDehydrogenase is finally oxidized by NAD
+ for it to return toits orginal form flavoprotein containing
FAD.
-
8/8/2019 Carbohydrate Metabolism 1
7/9
-
8/8/2019 Carbohydrate Metabolism 1
8/9
2Lactate + 6ATP + 6H 2 O Glucose + 6ADP + 6P i + 4H +
Glucose can be immediately trapped by the muscle with theenzyme
Hexokinase. During exercise there will be anincreased amount of
lactate causing lact ic acidosis . Thiswill cause pain in the
muscles (commonly in the form of cramps). Because of muscles do not
have lac ta tedehydrogenase Lactate will be transported to the
liverwhere it can be converted to glucose. In the liver lac ta
tedehydrogenase can catalyze the oxidation of Lac ta te to
Pyruva te which requires 6ATP to form glucose(gluconeogenesis)
to be sent back to the muscle and other
tissues. Utilizing 6ATP makes gluconeogenesis an
expensivepathway.
Amino ac ids as subs t ra te for Gluconeogenes is
In the muscles Alanine t ransaminase or Alanineaminotransferase
(ALT) catalyze the transferring of anamino group from glu tamate to
pyruva te forming -ke toglu ta ra te and a lan ine . Through the
blood Alanine
travels to the liver where it is converted back to pyruvatewith
ALT. Aside from the 6 ATP required for pyruvate toundergo
gluconeogenesis, Ammonium needs to bereleased from glutamate
through urea cycle utilizing 4 ATP.This makes amino acids as
substrate more expensive (ATP).
2Ala + 10ATP + CO 2 Glucose + Urea + 10ADP + 10P i
Excessive amounts of ammonia loaded into the liver, up to a
point that not everything can be immediatelyconverted to urea can
cause comatose due to hepaticencephalopathy. This is common in
liver cirrhosiscaused by alcoholism.
GLUCOKINASE IN THE LIVER Regulates blood glucose after a meal
Promotes increased hepatic utilization of glucose
o Promotes Glycolysis not gluconeogenesisnor glycogenolysis.
Carboxyla t ion of Pyruva te is catalyzed by Pyruvatecarboxylase
by transferring the CO 2 attached to biotinforming Oxaloace ta te
(OAA).
Because OAA is not permeable to the mitochondrialmembrane it is
reduced by NADH to mala te .
In the cytosol, mala te is reoxidized to OAA with NAD +.
OAA will then be converted (decarboxylation andphosphorylation)
to Phosphoenol pyruva te (PEP ) byPEP carboxykinase (PEPCK) with
the cofactor GTP( GDP) .
The succeeding steps beginning from PEP are reversereactions of
glycolysis,except in FBPase &G6Pase.
ATP is again utilizedin the formation of 1,3-BPG.
Because 2 moleculesis required in theformation of FBP 2pyruvate
must beutilized to form 1molecule of glucose.
After this step, 4 ATPand 2GTP has beenutilized by the
pathway.
2NADH will beutilized to convert 2moleculesof1,3BPG to form
2molecules of G3P.
PFK-1 is anirreversible enzymeof glycolysis
thereforegluconeogenesisneeds its ownenzyme FBPase.
The last step of gluconeogenesis is catalyzed by
Glucose-6-phosphatase(G6Pase) which dephosphorylates G6P
to Glucose . G6P is not found in the muscles
therefore,gluconeogenesis cannot occur in the muscle
von Gierkes Disease (mentioned in page 1 ) - type I glycogen
storage disease(GSD)- deficiency of glucose-6-phosphatase- severe
hypoglycemia causing lethargy,seizures and
brain damage- hepatomegaly, increased bleeding (clotting
factors
2,6,7,9,10 and platelets are affected) and growthretardation
Glucagon Maingluconeogenic
hormone
Similar to theprevious discussionin page 3 and 5,
-
8/8/2019 Carbohydrate Metabolism 1
9/9
Catecholamine increase secretion of Glucagon which issynthesized
from -pancreatic cells.
G LUCOSE IS S YNTHESIZED FROM M OS T AMINO ACIDS All amino acids
except leucine & lysine can supply
carbon for the net synthesis of glucose by
gluconeogenesis Pyruvate, oxaloacetate catabolic products of
amino
acids from which glucose synthesis can occur Anaplerotic
Reactions those that lead to net
synthesis of TCA intermediates
Hepat ic encephalopa thy utilization of aminoacids for
gluconeogenesis may cause production of excess urea which may lead
to an encephalopathyassociated with cirrhosis of the liver,
attributed to thepassage of toxic nitrogenous substances from
theportal to the systemic circulation; cerebralmanifestations may
include coma.
Gluconeogenic Ketogenic Both
GlycineSerineValineHistidineArginineCysteineProlineHydrodyprolineAlanineGlutamateGlutamineAspartateAsparaginesMethionine
LeucineLysine
ThreonineIsoleucinePhenylalanineTyrosineTryptophan
Glucose can be synthes ized f rom Propionyl -CoA(odd-numbered
carbon) Propionyl-CoA good precursor for gluconeogenesis as
it yields oxaloacetate by anaplerotic pathway(TCA cycle).
Triacylglycerol(TAG) when hydrolyzed yields 3 FAs &
glycerol (a substrate for gluconeogenesis)
o Because Glycerol kinase(found in adipose tissue) is absent in
the liver, TAG needs to be hydrolyzed
During starvation a great amount of ATP is used
forgluconeogenesis which may be taken from the oxidation of fatty
acids in the liver. This process however forms ketonebodies.Insu l
in
the only counter-regulatory hormone in gluconeogenesis main
glycolytic hormone action
o binds to receptoro receptor sends signal activating
phosphoprotein
phosphataseo inactive PFK-2 becomes active PFK-2 (Pi removed)o
active PFK-2 increases concentration of F-2,6 BPo F-2,6 BP binds to
PFK-1 & increases its activityo Rate of glycolysis is
increased
Alcohol Oxida t ion Inh ib i t s Gluconeogenes is
Due to excess NADH forcing the equilibrium of thereactions
catalyzed by lactate dehydrogenase and malatedehydrogenase
PEPCK Deficiency- rare but severe metabolic defect
- absence of the cytosolic form results in cerebra la t rophy ,
opt ic a t rophy , fatty infiltration of the liverand kidney and
intractable hypoglycemia (including lost of vision)
Fruc tose-1 ,6-Bisphosphatase Def ic iency
- Presents with neonatal hypoglycemia, along withacidosis,
irritability, tachycardia, dyspnea, hypotonia,moderate
hepatomegaly.
- typically only the liver enzyme is deficient, the
muscleactivity is normal
(Please refer to slide 82 of the PPT for the gluconeogenesis
summary)