Embed Size (px)
Citation preview

Calcium store sensor stromal-interaction molecule1-dependent signaling plays an important role incervical cancer growth, migration, and angiogenesisYih-Fung Chena,1, Wen-Tai Chiua,1, Ying-Ting Chenb, Pey-Yun Lina, Huey-Jy Huanga, Cheng-Yang Chouc,Hsien-Chang Changb,d, Ming-Jer Tange, and Meng-Ru Shena,c,f,g,2
aDepartment of Pharmacology, bInstitute of Biomedical Engineering, cDepartment of Obstetrics and Gynecology, dMedical Device Innovation Center,eDepartment of Physiology, fAdvanced Optoelectronic Technology Center, and gInfectious Diseases and Signaling Research Center, National Cheng KungUniversity, Tainan 70101, Taiwan
Edited by Anjana Rao, Immune Disease Institute and La Jolla Institute for Allergy and Immunology, Boston, MA, and approved August 10, 2011 (received forreview February 28, 2011)
Store-operated Ca2+ entry (SOCE) is the principal Ca2+ entry mecha-nism in nonexcitable cells. Stromal-interaction molecule 1 (STIM1) isan endoplasmic reticulum Ca2+ sensor that triggers SOCE activation.However, the role of STIM1 in regulating cancer progression remainscontroversial and its clinical relevance is unclear. Here we show thatSTIM1-dependent signaling is important for cervical cancer cell pro-liferation, migration, and angiogenesis. STIM1 overexpression in tu-mor tissue is noted in 71% cases of early-stage cervical cancer. Intumor tissues, the level of STIM1 expression is significantly associ-ated with the risk of metastasis and survival. EGF-stimulated cancercell migration requires STIM1 expression and EGF increases the in-teraction between STIM1 and Orai1 in juxta-membrane areas, andthus induces Ca2+ influx. STIM1 involves the activation of Ca2+-reg-ulated protease calpain, as well as Ca
2+
-regulated cytoplasmic kinasePyk2, which regulate the focal-adhesion dynamics of migratory cer-vical cancer cells. Because of an increase of p21 protein levels anda decrease of Cdc25C protein levels, STIM1-silencing in cervical can-cer cells significantly inhibits cell proliferation by arresting the cellcycle at the S and G2/M phases. STIM1 also regulates the productionof VEGF in cervical cancer cells. Interference with STIM1 expressionor blockade of SOCE activity inhibits tumor angiogenesis andgrowth in animal models, confirming the crucial role of STIM1-me-diated Ca2+ influx in aggravating tumor development in vivo. Theseresults make STIM1-dependent signaling an attractive target fortherapeutic intervention.
In most types of cells, modulation of intracellular Ca2+ levelsprovides versatile and dynamic signaling that mediate various
cellular processes, such as proliferation, migration, and geneexpression (1). Dysregulation of Ca2+ signaling has been iden-tified in tumor progression (2). The store-operated calcium entry(SOCE) is a major Ca2+ entry in nonexcitable cells (3, 4). SOCE,by definition, is activated by Ca2+ efflux from the internal store.Two genes, STIM1 (stromal-interaction molecule 1) and Orai1,are responsible for SOCE activation (3). Once endoplasmicreticulum Ca2+ depleted, STIM1 proteins aggregate into multiplepuncta and translocate to the close proximity of plasma mem-branes (5). Orai1 molecule, an essential pore-forming componentof the SOCE channel, translocates to the same STIM1-containingstructures during store depletion and opens to mediate Ca2+
entry (6). The physiological function of STIM1 has mostly beenstudied in immune systems (7). STIM proteins are required forthe development and function of regulatory T cells (8) andSTIM1-deficiency causes several autoimmune diseases and my-opathy in human subjects and mouse models (9). The chromo-somal location of human STIM1 is in 11p15.5, a region relatedto pediatric malignancies, which makes STIM1 a considerablefactor in mediating cell growth and transformation (10). STIM1was first suggested as a tumor-suppressor gene to induce celldeath (11). STIM1 mRNA is widely expressed in different hu-man tissues, and strongly presented in lymphoid and myeloidcells (9). STIM1 knockdown accelerates the cell motility of
melanoma cells and is defined as an antimetastasis gene (12).However, Yang et al. (13) demonstrated that STIM1 or Orai1silencing inhibits the migration and metastasis of breast cancercells by suppressing focal adhesion turnover. In contrast, anotherstudy showed that Orai1 could regulate mammary tumorigenesisby STIM1-independent pathways (14). Several works demon-strate that STIM1 could mediate cell proliferation. STIM1knockdown decreases the protein levels of cyclin D1 in a hepa-toma cell line (15) and STIM1-silencing inhibits cell proliferationin endothelial cells by arresting the cell cycle at the G2/M phase(16). However, the cell cycle stopped at the G1/S checkpointafter STIM1 depletion in rat vascular smooth-muscle cells (17).Cervical cancer is strongly associated with infection by oncogenic
types of human papillomavirus, but only a small fraction of thoseinfected develop cancer, indicating that other factors contribute tothe progression of the disease (18). Although intensive studies havebeen carried out, the tumor biology of this disease is largely un-known and little is known about the role of Ca2+ dysregulationinvolved in cervical malignancy. Using themodel of cervical cancer,we studied the role of STIM1 in promoting cancer-malignantbehaviors and how these processes are remodeled in cancer cells.The results indicate that STIM1 is important for Ca2+ signaling,which is necessary for cervical cancer cell growth, migration, andangiogenesis.We thus suggest STIM1-mediatedCa2+ signaling as apotential therapeutic target.
ResultsTumor Expression Level of STIM1 Is Associated with Clinical Outcome.To clarify whether STIM1 expression has clinical significance intumor progression, 24 cases of early-stage cervical cancer withpair tissues of carcinoma and adjacent nonneoplastic epitheliawere analyzed by immunoblotting (Fig. 1A). Compared with thatof nonneoplastic tissue, the level of STIM1 expression in tumortissues was elevated in 71% cases (Fig. 1B). The immunofluo-rescent stainings confirmed that cervical cancer tissues clearlyexpressed STIM1 protein, whereas STIM1 protein was rarelydetected in the adjacent normal or nonneoplastic cervical epi-thelia (Fig. 1C). More importantly, the level of STIM1 expres-sion in tumor tissues was closely correlated with tumor size (Fig.1D) (linear fit, R = 0.76, P < 0.001), an important indicator ofhuman cervical carcinoma progression in vivo (19). In addition,
Author contributions: Y.-F.C., W.-T.C., Y.-T.C., H.-J.H., C.-Y.C., H.-C.C., M.-J.T., and M.-R.S.designed research; Y.-F.C., W.-T.C., Y.-T.C., P.-Y.L., H.-J.H., and M.-R.S. performed research;C.-Y.C. contributed new reagents/analytic tools; Y.-F.C., W.-T.C., Y.-T.C., P.-Y.L., H.-J.H.,C.-Y.C., H.-C.C., and M.-R.S. analyzed data; and Y.-F.C., W.-T.C., and M.-R.S. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1Y.-F.C. and W.-T.C. contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1103315108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1103315108 PNAS | September 13, 2011 | vol. 108 | no. 37 | 15225e15230
CELL
BIOLO
GY
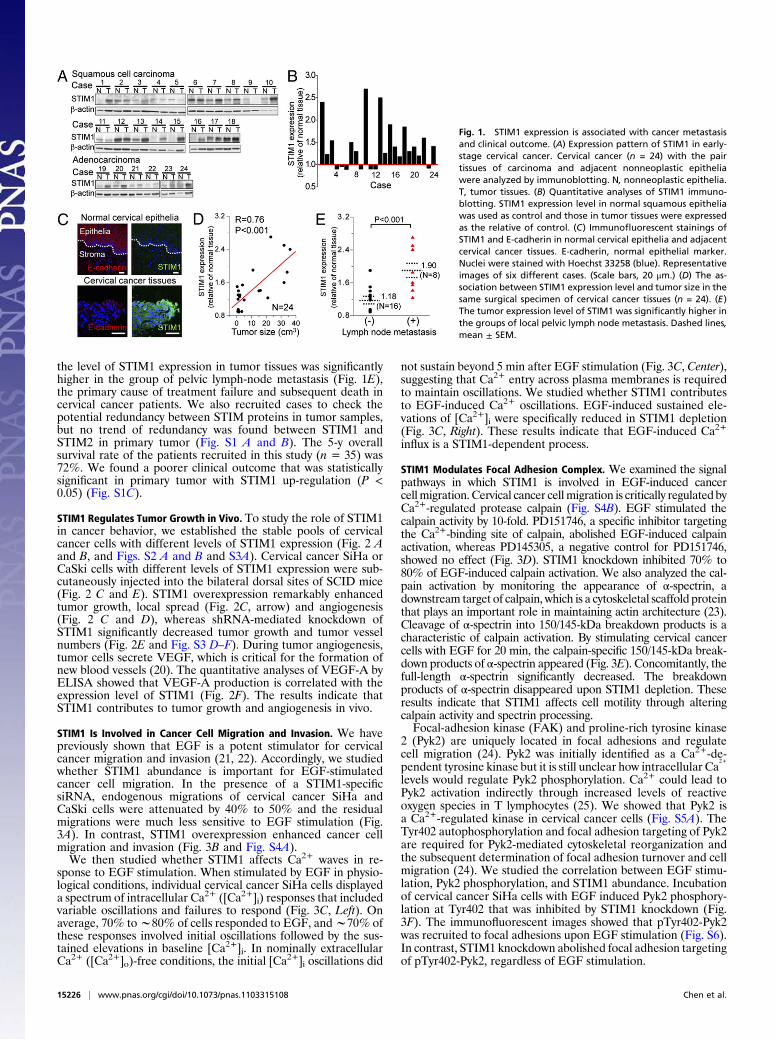
the level of STIM1 expression in tumor tissues was significantlyhigher in the group of pelvic lymph-node metastasis (Fig. 1E),the primary cause of treatment failure and subsequent death incervical cancer patients. We also recruited cases to check thepotential redundancy between STIM proteins in tumor samples,but no trend of redundancy was found between STIM1 andSTIM2 in primary tumor (Fig. S1 A and B). The 5-y overallsurvival rate of the patients recruited in this study (n = 35) was72%. We found a poorer clinical outcome that was statisticallysignificant in primary tumor with STIM1 up-regulation (P <0.05) (Fig. S1C).
STIM1 Regulates Tumor Growth in Vivo. To study the role of STIM1in cancer behavior, we established the stable pools of cervicalcancer cells with different levels of STIM1 expression (Fig. 2 Aand B, and Figs. S2 A and B and S3A). Cervical cancer SiHa orCaSki cells with different levels of STIM1 expression were sub-cutaneously injected into the bilateral dorsal sites of SCID mice(Fig. 2 C and E). STIM1 overexpression remarkably enhancedtumor growth, local spread (Fig. 2C, arrow) and angiogenesis(Fig. 2 C and D), whereas shRNA-mediated knockdown ofSTIM1 significantly decreased tumor growth and tumor vesselnumbers (Fig. 2E and Fig. S3 D–F). During tumor angiogenesis,tumor cells secrete VEGF, which is critical for the formation ofnew blood vessels (20). The quantitative analyses of VEGF-A byELISA showed that VEGF-A production is correlated with theexpression level of STIM1 (Fig. 2F). The results indicate thatSTIM1 contributes to tumor growth and angiogenesis in vivo.
STIM1 Is Involved in Cancer Cell Migration and Invasion. We havepreviously shown that EGF is a potent stimulator for cervicalcancer migration and invasion (21, 22). Accordingly, we studiedwhether STIM1 abundance is important for EGF-stimulatedcancer cell migration. In the presence of a STIM1-specificsiRNA, endogenous migrations of cervical cancer SiHa andCaSki cells were attenuated by 40% to 50% and the residualmigrations were much less sensitive to EGF stimulation (Fig.3A). In contrast, STIM1 overexpression enhanced cancer cellmigration and invasion (Fig. 3B and Fig. S4A).We then studied whether STIM1 affects Ca2+ waves in re-
sponse to EGF stimulation. When stimulated by EGF in physio-logical conditions, individual cervical cancer SiHa cells displayeda spectrum of intracellular Ca2+ ([Ca2+]i) responses that includedvariable oscillations and failures to respond (Fig. 3C, Left). Onaverage, 70% tow80% of cells responded to EGF, andw70% ofthese responses involved initial oscillations followed by the sus-tained elevations in baseline [Ca2+]i. In nominally extracellularCa2+ ([Ca2+]o)-free conditions, the initial [Ca
2+]i oscillations did
not sustain beyond 5 min after EGF stimulation (Fig. 3C, Center),suggesting that Ca2+ entry across plasma membranes is requiredto maintain oscillations. We studied whether STIM1 contributesto EGF-induced Ca2+ oscillations. EGF-induced sustained ele-vations of [Ca2+]i were specifically reduced in STIM1 depletion(Fig. 3C, Right). These results indicate that EGF-induced Ca2+
influx is a STIM1-dependent process.
STIM1 Modulates Focal Adhesion Complex. We examined the signalpathways in which STIM1 is involved in EGF-induced cancercellmigration. Cervical cancer cellmigration is critically regulated byCa2+-regulated protease calpain (Fig. S4B). EGF stimulated thecalpain activity by 10-fold. PD151746, a specific inhibitor targetingthe Ca2+-binding site of calpain, abolished EGF-induced calpainactivation, whereas PD145305, a negative control for PD151746,showed no effect (Fig. 3D). STIM1 knockdown inhibited 70% to80% of EGF-induced calpain activation. We also analyzed the cal-pain activation by monitoring the appearance of α-spectrin, adownstream target of calpain, which is a cytoskeletal scaffold proteinthat plays an important role in maintaining actin architecture (23).Cleavage of α-spectrin into 150/145-kDa breakdown products is acharacteristic of calpain activation. By stimulating cervical cancercells with EGF for 20 min, the calpain-specific 150/145-kDa break-down products of α-spectrin appeared (Fig. 3E). Concomitantly, thefull-length α-spectrin significantly decreased. The breakdownproducts of α-spectrin disappeared upon STIM1 depletion. Theseresults indicate that STIM1 affects cell motility through alteringcalpain activity and spectrin processing.Focal-adhesion kinase (FAK) and proline-rich tyrosine kinase
2 (Pyk2) are uniquely located in focal adhesions and regulatecell migration (24). Pyk2 was initially identified as a Ca2+-de-pendent tyrosine kinase but it is still unclear how intracellular Ca
2+
levels would regulate Pyk2 phosphorylation. Ca2+ could lead toPyk2 activation indirectly through increased levels of reactiveoxygen species in T lymphocytes (25). We showed that Pyk2 isa Ca2+-regulated kinase in cervical cancer cells (Fig. S5A). TheTyr402 autophosphorylation and focal adhesion targeting of Pyk2are required for Pyk2-mediated cytoskeletal reorganization andthe subsequent determination of focal adhesion turnover and cellmigration (24). We studied the correlation between EGF stimu-lation, Pyk2 phosphorylation, and STIM1 abundance. Incubationof cervical cancer SiHa cells with EGF induced Pyk2 phosphory-lation at Tyr402 that was inhibited by STIM1 knockdown (Fig.3F). The immunofluorescent images showed that pTyr402-Pyk2was recruited to focal adhesions upon EGF stimulation (Fig. S6).In contrast, STIM1 knockdown abolished focal adhesion targetingof pTyr402-Pyk2, regardless of EGF stimulation.
Fig. 1. STIM1 expression is associated with cancer metastasisand clinical outcome. (A) Expression pattern of STIM1 in early-stage cervical cancer. Cervical cancer (n = 24) with the pairtissues of carcinoma and adjacent nonneoplastic epitheliawere analyzed by immunoblotting. N, nonneoplastic epithelia.T, tumor tissues. (B) Quantitative analyses of STIM1 immuno-blotting. STIM1 expression level in normal squamous epitheliawas used as control and those in tumor tissues were expressedas the relative of control. (C) Immunofluorescent stainings ofSTIM1 and E-cadherin in normal cervical epithelia and adjacentcervical cancer tissues. E-cadherin, normal epithelial marker.Nuclei were stained with Hoechst 33258 (blue). Representativeimages of six different cases. (Scale bars, 20 μm.) (D) The as-sociation between STIM1 expression level and tumor size in thesame surgical specimen of cervical cancer tissues (n = 24). (E)The tumor expression level of STIM1 was significantly higher inthe groups of local pelvic lymph node metastasis. Dashed lines,mean ± SEM.
15226 | www.pnas.org/cgi/doi/10.1073/pnas.1103315108 Chen et al.

We further studied the focal-adhesion dynamics by immu-nostaining with FAK, a major component of the focal-adhesioncomplex (Fig. 3G). In control cells, FAK staining showeda punctate pattern of focal adhesion and EGF induced smallperipheral adhesions. In contrast, STIM1 knockdown cells dis-played the quiescent cell morphology, with large focal adhesionsaround the cell periphery regardless of EGF stimulation. Theeffect of STIM1 siRNA on focal adhesion turnover was rescuedby the subsequent transient expression of STIM1 cDNA insiRNA-treated cells (Fig. S7), suggesting that STIM1 is impor-tant for focal adhesion turnover. The immunoblotting alsoshowed that EGF increased the levels of pTyr397-FAK, whichwas significantly inhibited by STIM1 knockdown (Fig. S5B).
EGF Enhances the Interaction Between STIM1 and Orai1. It has beenshown that STIM1 is essential for the regulation of Orai1, a porecomponent of the SOC channel (6). The immunoprecipitationand living cell images showed that the interaction betweenSTIM1 and Orai1 was enhanced by EGF stimulation (Fig. 4A–C). Although cervical cancer SiHa cells express Orai2, Orai3,TRPC1, and TRPC6 (Fig. 4A and Fig. S2C), STIM1 interactswith none of them with or without EGF stimulation (Fig. 4 A,D, and E).
Fig. 2. STIM1 is involved in tumor growth and angiogenesis. (A) STIM1knockdown by shRNA in cervical cancer SiHa and CaSki cell lines. (B) Estab-lishment of STIM1-overexpressed cervical cancer cell lines. EndogenousSTIM1 and exogenous STIM1 were differentiated by immunoblotting withSTIM1 (Left) and EGFP (Right), respectively. (C and D) STIM1 overexpressionenhances tumor angiogenesis and growth. Bilateral dorsal sites of SCID micewere subcutaneously inoculated with mock-transfected (Control) or STIM1-overexpressed cervical cancer cells. Representative tumor xenografts (C), themean tumor vessel numbers and mean tumor weight (D) 21 d after in-oculation. Arrowhead, local spread of tumor mass. (Scale bars, 0.5 cm.)Columns, mean ± SEM (n = 6), *P < 0.01. (E) STIM1 knockdown attenuatestumor growth and angiogenesis. Representative tumor xenografts (Left),mean tumor vessel numbers (Top Right), and mean tumor weight (BottomRight) 15 d after inoculation of control shRNA- or shSTIM1-transfected cer-vical cancer cells. Columns, mean ± SEM (n = 6), *P < 0.01. (Scale bar, 1 cm.)(F) STIM1 regulates VEGF-A production. The VEGF-A secretion in variousstable pools of cervical cancer cells were measured by ELISA. *P < 0.01,compared with wild type. Columns, mean ± SEM (n = 5).
Fig. 3. STIM1 modulates the focal-adhesion complex. (A and B) STIM1 is in-volved in cervical cancer cell migration. Columns, mean ± SEM from at leastfour different experiments. EGF, 100 ng/mL epidermal growth factor. (C) EGF-induced Ca2+ influx is a STIM1-dependent process. Gray lines, [Ca2+]i oscil-lations of individual cervical cancer SiHa cells. Black lines, the mean trace of[Ca2+]i oscillations. (D and E) STIM1 is necessary for EGF-stimulated calpainactivation. (D) Quantitative fluorescent analyses of intracellular calpain ac-tivities, measured with a fluorogenic membrane-permeable calpain substratet-Boc-LM-CMAC. PD151746 (50 μM), a specific inhibitor targeting Ca2+-bindingsite of calpain; PD145305 (50 μM), a negative control for PD151746. Columns,mean ± SEM from at least 100 cells. (E) STIM1 siRNA abolishes EGF-inducedcleavage of α-spectrin. Arrow and arrowhead, the full-length (280 kDa) andcalpain-digested cleaved form (150/145 kDa) of α-spectrin, respectively. (F)STIM1 affects EGF-stimulated Pyk2 activation. (Left) Representative immu-noblots from at least three different experiments. (Right) Densitometricquantification of Pyk2 phosphorylation (Tyr402) levels. Points, mean ± SEM;*P < 0.01, compared with control group. (G) STIM1 modulates the focal ad-hesion turnover. (Left) Representative images showing focal adhesions. FAK(green), focal-adhesion marker. (Right) Quantitative analyses of focal adhe-sion size. Focal-adhesion sizes were quantified by measuring the area of FAKstaining. Columns, mean ± SEM from at least 20 individual cells of three dif-ferent experiments. N.S., nonsignificant. (Scale bars, 10 μm.)
Chen et al. PNAS | September 13, 2011 | vol. 108 | no. 37 | 15227
CELL
BIOLO
GY
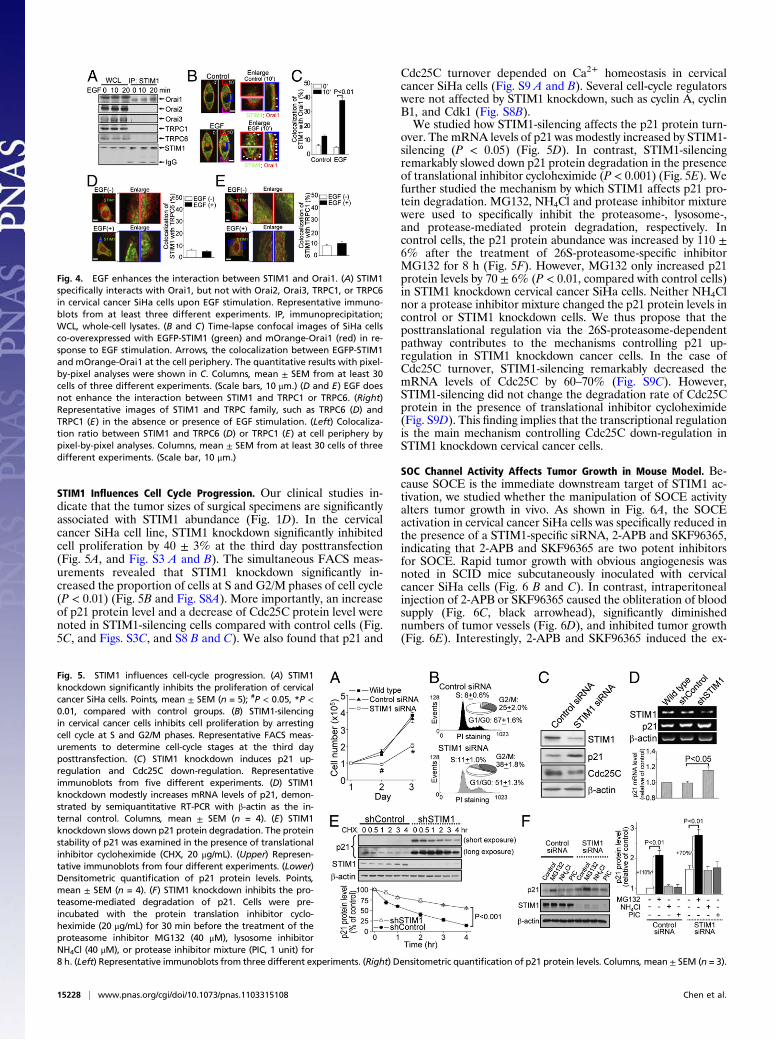
STIM1 Influences Cell Cycle Progression. Our clinical studies in-dicate that the tumor sizes of surgical specimens are significantlyassociated with STIM1 abundance (Fig. 1D). In the cervicalcancer SiHa cell line, STIM1 knockdown significantly inhibitedcell proliferation by 40 ± 3% at the third day posttransfection(Fig. 5A, and Fig. S3 A and B). The simultaneous FACS meas-urements revealed that STIM1 knockdown significantly in-creased the proportion of cells at S and G2/M phases of cell cycle(P < 0.01) (Fig. 5B and Fig. S8A). More importantly, an increaseof p21 protein level and a decrease of Cdc25C protein level werenoted in STIM1-silencing cells compared with control cells (Fig.5C, and Figs. S3C, and S8 B and C). We also found that p21 and
Cdc25C turnover depended on Ca2+ homeostasis in cervicalcancer SiHa cells (Fig. S9 A and B). Several cell-cycle regulatorswere not affected by STIM1 knockdown, such as cyclin A, cyclinB1, and Cdk1 (Fig. S8B).We studied how STIM1-silencing affects the p21 protein turn-
over. The mRNA levels of p21 was modestly increased by STIM1-silencing (P < 0.05) (Fig. 5D). In contrast, STIM1-silencingremarkably slowed down p21 protein degradation in the presenceof translational inhibitor cycloheximide (P < 0.001) (Fig. 5E). Wefurther studied the mechanism by which STIM1 affects p21 pro-tein degradation. MG132, NH4Cl and protease inhibitor mixturewere used to specifically inhibit the proteasome-, lysosome-,and protease-mediated protein degradation, respectively. Incontrol cells, the p21 protein abundance was increased by 110 ±6% after the treatment of 26S-proteasome-specific inhibitorMG132 for 8 h (Fig. 5F). However, MG132 only increased p21protein levels by 70 ± 6% (P < 0.01, compared with control cells)in STIM1 knockdown cervical cancer SiHa cells. Neither NH4Clnor a protease inhibitor mixture changed the p21 protein levels incontrol or STIM1 knockdown cells. We thus propose that theposttranslational regulation via the 26S-proteasome-dependentpathway contributes to the mechanisms controlling p21 up-regulation in STIM1 knockdown cancer cells. In the case ofCdc25C turnover, STIM1-silencing remarkably decreased themRNA levels of Cdc25C by 60–70% (Fig. S9C). However,STIM1-silencing did not change the degradation rate of Cdc25Cprotein in the presence of translational inhibitor cycloheximide(Fig. S9D). This finding implies that the transcriptional regulationis the main mechanism controlling Cdc25C down-regulation inSTIM1 knockdown cervical cancer cells.
SOC Channel Activity Affects Tumor Growth in Mouse Model. Be-cause SOCE is the immediate downstream target of STIM1 ac-tivation, we studied whether the manipulation of SOCE activityalters tumor growth in vivo. As shown in Fig. 6A, the SOCEactivation in cervical cancer SiHa cells was specifically reduced inthe presence of a STIM1-specific siRNA, 2-APB and SKF96365,indicating that 2-APB and SKF96365 are two potent inhibitorsfor SOCE. Rapid tumor growth with obvious angiogenesis wasnoted in SCID mice subcutaneously inoculated with cervicalcancer SiHa cells (Fig. 6 B and C). In contrast, intraperitonealinjection of 2-APB or SKF96365 caused the obliteration of bloodsupply (Fig. 6C, black arrowhead), significantly diminishednumbers of tumor vessels (Fig. 6D), and inhibited tumor growth(Fig. 6E). Interestingly, 2-APB and SKF96365 induced the ex-
Fig. 4. EGF enhances the interaction between STIM1 and Orai1. (A) STIM1specifically interacts with Orai1, but not with Orai2, Orai3, TRPC1, or TRPC6in cervical cancer SiHa cells upon EGF stimulation. Representative immuno-blots from at least three different experiments. IP, immunoprecipitation;WCL, whole-cell lysates. (B and C) Time-lapse confocal images of SiHa cellsco-overexpressed with EGFP-STIM1 (green) and mOrange-Orai1 (red) in re-sponse to EGF stimulation. Arrows, the colocalization between EGFP-STIM1and mOrange-Orai1 at the cell periphery. The quantitative results with pixel-by-pixel analyses were shown in C. Columns, mean ± SEM from at least 30cells of three different experiments. (Scale bars, 10 μm.) (D and E) EGF doesnot enhance the interaction between STIM1 and TRPC1 or TRPC6. (Right)Representative images of STIM1 and TRPC family, such as TRPC6 (D) andTRPC1 (E) in the absence or presence of EGF stimulation. (Left) Colocaliza-tion ratio between STIM1 and TRPC6 (D) or TRPC1 (E) at cell periphery bypixel-by-pixel analyses. Columns, mean ± SEM from at least 30 cells of threedifferent experiments. (Scale bar, 10 μm.)
Fig. 5. STIM1 influences cell-cycle progression. (A) STIM1knockdown significantly inhibits the proliferation of cervicalcancer SiHa cells. Points, mean ± SEM (n = 5); #P < 0.05, *P <0.01, compared with control groups. (B) STIM1-silencingin cervical cancer cells inhibits cell proliferation by arrestingcell cycle at S and G2/M phases. Representative FACS meas-urements to determine cell-cycle stages at the third dayposttransfection. (C) STIM1 knockdown induces p21 up-regulation and Cdc25C down-regulation. Representativeimmunoblots from five different experiments. (D) STIM1knockdown modestly increases mRNA levels of p21, demon-strated by semiquantitative RT-PCR with β-actin as the in-ternal control. Columns, mean ± SEM (n = 4). (E) STIM1knockdown slows down p21 protein degradation. The proteinstability of p21 was examined in the presence of translationalinhibitor cycloheximide (CHX, 20 μg/mL). (Upper) Represen-tative immunoblots from four different experiments. (Lower)Densitometric quantification of p21 protein levels. Points,mean ± SEM (n = 4). (F) STIM1 knockdown inhibits the pro-teasome-mediated degradation of p21. Cells were pre-incubated with the protein translation inhibitor cyclo-heximide (20 μg/mL) for 30 min before the treatment of theproteasome inhibitor MG132 (40 μM), lysosome inhibitorNH4Cl (40 μM), or protease inhibitor mixture (PIC, 1 unit) for8 h. (Left) Representative immunoblots from three different experiments. (Right) Densitometric quantification of p21 protein levels. Columns,mean ± SEM (n = 3).
15228 | www.pnas.org/cgi/doi/10.1073/pnas.1103315108 Chen et al.

travasation of tumor feeding vessels (Fig. 6C, blue arrowhead),implying the damage of blood vessels. These results indicate thatblockade of SOCE activity causes tumor growth regression.
DiscussionThis study highlights the novel role of Ca2+ store-sensor STIM1in tumor malignant behavior. We show that STIM1 is importantfor cervical cancer cell proliferation, migration, and angiogene-sis. This conclusion is supported by the following evidence. (i)STIM1-silencing in cervical cancer cells significantly inhibits cellproliferation by arresting the cell cycle at the S and G2/M phases.(ii) STIM1 overexpression enhances the invasive migration ofcervical cancer cells, whereas STIM1 knockdown attenuates it.(iii) STIM1 expression regulates VEGF-A secretion from cancercells. (iv) The animal model confirms that tumor abundance ofSTIM1 is linked with tumor growth, angiogenesis and local in-vasion. (v) The tumor expression level of STIM1 is closely as-sociated with the clinical outcome of early-stage cervical cancer.To the best of our knowledge, this study is unique in showing themultiple functions of STIM1 in tumor biology, as evidenced bycell-line studies, animal models, and clinical sample analyses.Our clinical studies indicate that more than 70% cases of early-
stage cervical cancer display STIM1 overexpression. High abun-dance of tumor STIM1 indicates the high risk of metastasis. Westudied how STIM1 overexpression leads to the enhancement ofcancer cell migration and proposed that STIM1 affects cellmotility through altering calpain activity and spectrin processing
based on the following evidence. (i) Calpain activity is involved incervical cancer cell migration. (ii) STIM1 affects calpain activationand spectrin processing. (iii) STIM1 is important for focal adhe-sion turnover. We also highlight the molecular identity of EGF-induced Ca2+ influx in cervical cancer cells. STIM1 is the mo-lecular linker from endoplasmic reticulum Ca2+ store depletion tothe plasma membrane SOCE (5, 26). STIM1 can interact withvarious plasma membrane Ca2+ channels, such as Orai proteinsand the transient-receptor potential channel family, to form thefunctional pore-subunit of the SOC channel (27). The living cellimaging and immunoprecipitation demonstrate that EGF stim-ulates the aggregation and translocation of STIM1 toward theproximity of the plasma membrane, where STIM1 specificallyinteracts with Orai1 to mediate SOCE.The findings ofYang et al. (13) show the emerging importance of
STIM1 in tumor biology. Yang et al. proposed that blockingSTIM1-mediating Ca2+ influx impairs focal adhesion turnover,which can be rescued by the small GTPases Ras and Rac. Ourfindings suggest that STIM1 is the key regulator of EGF-inducedCa2+ influx that is necessary for the activation of Ca2+-regulatedprotease calpain and tyrosine kinase Pyk2. The activation of cal-pain and Pyk2 regulates multiple signaling events crucial for thefocal adhesion turnover and the locomotion of cervical cancer cells.The pathological significance of tumor STIM1 overexpression canbe proposed to benefit the locomotion of cancer cells. In animalstudies, we carefully dissected the tumor implants and un-expectedly found that, in addition to blocking tumor growth, theSOCE inhibitors, such as 2-APB and SKF96365, obliterated tumorfeeding vessels. We thus suggest that both tumor cells and tumorvessels are possible targets of these drugs. In the study ofYang et al.(13), a single inhibitor (SKF96365) at a higher concentration wasshown to inhibit lung metastasis. However, the information on theantiangiogenic effect of SKF96365 is not available in their work.It has been reported that STIM1 mediates SOCE in endo-
thelial and vascular smooth-muscle cells, and STIM1 knockdownelicits cell-cycle arrest (16, 17). Our results show that STIM1-silencing in cervical cancer cells leads to a decrease in pro-liferation by arresting the cell cycle at the S and G2/M phases.Several proteins alter G2/M checkpoint progression, such as p21,Cdc25C, and the Cdk1 (Cdc2)-cyclin B complex (28). Cdk1 canbe dephosphorylated and activated by Cdc25C, leading to cellmitosis (29). In contrast, p21 inhibits the activity of Cdk1 andcauses cell-cycle arrest (30). STIM1-silencing cervical cancercells exhibited an increase of p21 protein levels and a decrease ofCdc25C protein. Compared with modest changes in p21 mRNAlevels, the protein stability of p21 was remarkably increased inSTIM1 knockdown cells, suggesting that STIM1 regulates p21expression mainly through the posttranslational modification.STIM1 is also involved in angiogenesis through its effect onVEGF production from cancer cells. Further study needs toexplore how STIM1 regulates VEGF production in detail. Takentogether, the molecular mechanism by which STIM1 affectscancer malignant behaviors involves the regulation of Ca2+ sig-naling essential for cancer cell proliferation, migration, and tu-mor angiogenesis. These results make the STIM1-mediatedsignaling an attractive target for therapeutic intervention.
Materials and MethodsCell Cultures, Transfection, RNA Interference. Cultures of human cervical cancercell lines (SiHa and CaSki) were prepared as described previously (21). Stablepools of cervical cancer cells overexpressing human STIM1 were used in thisstudy. The full-length cDNA of EGFP-STIM1, mOrange-STIM1, and mOrange-Orai1 were subcloned into the eukaryotic expression vector pCMV6-XL5 (31).EGFP- ormOrange-STIM1 cDNA plasmids were transfected into cervical cancerSiHa cells by Lipofectamine 2000 (Invitrogen) and cells with STIM1 over-expression were selected by G418 (Sigma-Aldrich). FACSAria cell sorter (BDBiosciences) was used to isolate the stable pools of cells overexpressing humanSTIM1. For RNAi-mediated STIM1 knockdown, siRNAs or shRNAs were used(Table S1). Cells were introducedwith siRNAor shRNAwith Lipofetamine 2000(Invitrogen) or by electroporation. The details of RNAi targeting human STIM1and RNAi rescue are listed in SI Materials and Methods.
Fig. 6. Store-operated Ca2+ entry affects tumor growth in vivo. (A) STIM1siRNA, 2-APB (20 μM), or SKF96365 (50 μM) inhibits SOCE of cervical cancerSiHa cells. To measure Ca2+ entry, SiHa cells loaded with Fura-2/AM (2 μM)were preincubated in Ca2+-free media plus 2 μM thapsigargin for 30 min todeplete the internal Ca2+ store. Each trace was averaged from at least 30single cells. (B–E) Blockade of SOCE retards tumor growth and angiogenesis.(B) Female SCID mice bearing tumor xenograft of SiHa cells were in-traperitoneally injected every 3 d (arrows) with control vehicle (n = 6),SKF96365 (2.5 mg/kg; n = 6), or 2-APB (50 μg/kg; n = 6) from the sixth daypostinoculation. Representative tumor xenografts (C), mean tumor vesselnumbers (D), and mean tumor weight (E) at the 15th day postinoculation.Arrows, rapid tumor growth with obvious angiogenesis. Black arrowheads,the obliteration of blood supply. Blue arrowheads, the extravasation of tu-mor feeding vessels. Columns, mean ± SEM (n = 6); *P < 0.01, compared withcontrol group. (Scale bars, 1 cm.)
Chen et al. PNAS | September 13, 2011 | vol. 108 | no. 37 | 15229
CELL
BIOLO
GY

Surgical Specimens. We collected the frozen tissues from the patients withearly stage (International Federation of Gynecology andObstetrics staging Ib)cervical cancer who were scheduled for radical hysterectomy and pelviclymphadenectomy at National Cheng Kung University Hospital, Taiwan. Thepatients with local pelvic lymph node metastases did not exhibit distantmetastases, such as paraaortic lymph nodes or distant organs. The collectionof surgical specimens was approved by the institutional review board ofNational Cheng Kung University Hospital.
Migration and Invasion Assay. In brief, cells were allowed to migrate acrossa membrane (8-μm pore) toward the medium containing 10 μg/mL fibronectinat 37 °C (21, 22). The invasive migration was done in the BD Matrigel invasionchamber (BD Biosciences) as an index of invasive activity of tumor cells (32).
VEGF-A Secretion Assay. Briefly, 2 × 106 cells were plated for 72 h and treatedwith 0.3 mM suramin for 3 h to release surface matrix-bound VEGF intoculture medium. The VEGF secretion in culture supernatants were quantifiedusing ELISA (R&D Systems) according to the manufacturer’s instructions.
Immunoblotting and Immunoprecipitation. The preparation of cell lysateand procedures of SDS/PAGE are described in detail in SI Materials andMethods. Bands in immunoblots were quantified using Vision WorksLSsoftware (UVP).
Immunofluorescence, Confocal Microscopy, and Image Analyses. The fluo-rophores were excited by laser at 405, 488, or 543 nm and detected bya scanning confocal microscope (FV-1000, Olympus). Cells with EGFP-STIM1 ormOrange-Orai1 overexpression were directly activated by laser at 488 or 543nm, respectively (33). Cells were maintained in phenol red-free medium at37 °C throughout the recording period. For the measurement of μ-calpainactivity, an artificial μ-calpain fluorescent substrate t-butoxycarbonyl-Leu-Met-chloromethylaminocoumarin (t-Boc-LM-CMAC; Invitrogen) was used(33). The fluorescent intensity of t-Boc-LM-CMAC hydrolytic product wasquantitatively analyzed as an index of μ-calpain activity of individual cell. Apixel-by-pixel analysis by FV-1000 software was used to assess the colocali-zation of STIM1with Oria1, TRPC1, or TRPC6 in confocal images. Focal-ad-hesion sizes were analyzed by dividing FAK staining pixels by the number offocal-adhesion patches.
Single Cell [Ca2+]i Measurement. [Ca2+]i was measured at 37 °C with the Fura-2fluorescence ratio method on a single-cell fluorimeter, as previously de-scribed (33). In brief, cells loaded with 2 μM Fura-2/acetoxymethyl ester(Fura-2/AM) were excited alternatively between 340 nm (I340) and 380 nm(I380) using the Polychrome IV monochromator (Till Photonics). The fluores-cence intensity of excitation at 510 nm was monitored to calculate [Ca2+]iusing the TILLvisION 4.0 program (Till Photonics).
Proliferation Assay and Cell Cycle Analysis. Cell proliferation was assessed bycell counting with Trypan blue exclusion (0.08%) to monitor viability. Cell-cycle stage was determined by FACS. Cellular DNA content was determinedafter propidium iodide staining. Cells were classified as in G0/G1, G2/M, and Sphase based on the fluorescence intensity and the cell cycle distribution wasanalyzed by the Cell Fit software (BD Biosciences).
RT-PCR. Total RNAs were isolated by RNeasy Mini Kit (Qiagen) and cDNA wasprepared with SuperScript II (Invitrogen). The primers targeting humanβ-actin, GAPDH, p21, Cdc25C, and STIM1 are described in SI Materialsand Methods.
Animal Models. For the inoculation of xenograft, the bilateral dorsal sites offemale BALB/c SCID mice aged 6 to 8 wk were subcutaneously injected with5 × 106 cervical cancer SiHa cells with different STIM1 expression level. Theanimal experiments were performed according to the ethical guidelines andapproved by the institutional ethical committee of National Cheng KungUniversity.
Statistics. All values were reported as mean ± SEM. Student’s paired t test,unpaired t test, or linear regression was used for statistical analyses. Survivaldata were assessed by Kaplan–Meier methods and differences were com-pared by the log-rank statistic. Differences between values were consideredsignificant when P < 0.05.
ACKNOWLEDGMENTS. We thank Dr. Liangyi Chen for STIM1 plamids, Ms.Yu-Ting Yen for help in animal studies and ELISA, Dr. Yi-Chuang E. Lin forEnglish editing, and Dr. Clive Ellory for critical discussion. This work wassupported in part by the National Science Council and Department ofHealth, Executive Yuan, Taiwan.
1. Berridge MJ, Bootman MD, Roderick HL (2003) Calcium signalling: Dynamics, ho-meostasis and remodelling. Nat Rev Mol Cell Biol 4:517e529.
2. Roderick HL, Cook SJ (2008) Ca2+ signalling checkpoints in cancer: Remodelling Ca2+
for cancer cell proliferation and survival. Nat Rev Cancer 8:361e375.3. Parekh AB (2010) Store-operated CRAC channels: Function in health and disease. Nat
Rev Drug Discov 9:399e410.4. Putney JW, Jr. (2005) Capacitative calcium entry: Sensing the calcium stores. J Cell Biol
169:381e382.5. Zhang SL, et al. (2005) STIM1 is a Ca2+ sensor that activates CRAC channels and mi-
grates from the Ca2+ store to the plasma membrane. Nature 437:902e905.6. Park CY, et al. (2009) STIM1 clusters and activates CRAC channels via direct binding of
a cytosolic domain to Orai1. Cell 136:876e890.7. Di Capite J, Parekh AB (2009) CRAC channels and Ca2+ signaling in mast cells. Immunol
Rev 231(1):45e58.8. Oh-Hora M, et al. (2008) Dual functions for the endoplasmic reticulum calcium sensors
STIM1 and STIM2 in T cell activation and tolerance. Nat Immunol 9:432e443.9. Feske S (2009) ORAI1 and STIM1 deficiency in human and mice: Roles of store-oper-
ated Ca2+ entry in the immune system and beyond. Immunol Rev 231:189e209.10. Parker NJ, Begley CG, Smith PJ, Fox RM (1996) Molecular cloning of a novel human
gene (D11S4896E) at chromosomal region 11p15.5. Genomics 37:253e256.11. Sabbioni S, Barbanti-Brodano G, Croce CM, Negrini M (1997) GOK: A gene at 11p15
involved in rhabdomyosarcoma and rhabdoid tumor development. Cancer Res 57:4493e4497.
12. Suyama E, et al. (2004) Identification of metastasis-related genes in a mouse modelusing a library of randomized ribozymes. J Biol Chem 279:38083e38086.
13. Yang S, Zhang JJ, Huang XY (2009) Orai1 and STIM1 are critical for breast tumor cellmigration and metastasis. Cancer Cell 15(2):124e134.
14. Feng M, et al. (2010) Store-independent activation of Orai1 by SPCA2 in mammarytumors. Cell 143(1):84e98.
15. El Boustany C, et al. (2008) Capacitative calcium entry and transient receptor potentialcanonical 6 expression control human hepatoma cell proliferation. Hepatology 47:2068e2077.
16. Abdullaev IF, et al. (2008) Stim1 and Orai1 mediate CRAC currents and store-operatedcalcium entry important for endothelial cell proliferation. Circ Res 103:1289e1299.
17. Potier M, et al. (2009) Evidence for STIM1- and Orai1-dependent store-operatedcalcium influx through ICRAC in vascular smooth muscle cells: Role in proliferationand migration. FASEB J 23:2425e2437.
18. Woodman CB, Collins SI, Young LS (2007) The natural history of cervical HPV in-fection: Unresolved issues. Nat Rev Cancer 7(1):11e22.
19. Mayr NA, et al. (2002) Method and timing of tumor volume measurement for out-come prediction in cervical cancer using magnetic resonance imaging. Int J RadiatOncol Biol Phys 52(1):14e22.
20. Nagy JA, Dvorak AM, Dvorak HF (2007) VEGF-A and the induction of pathologicalangiogenesis. Annu Rev Pathol 2:251e275.
21. Shen MR, et al. (2006) Insulin-like growth factor 1 is a potent stimulator of cervicalcancer cell invasiveness and proliferation that is modulated by alphavbeta3 integrinsignaling. Carcinogenesis 27:962e971.
22. Chen YF, et al. (2009) Motor protein-dependent membrane trafficking of KCl co-transporter-4 is important for cancer cell invasion. Cancer Res 69:8585e8593.
23. Nicolas G, et al. (2002) Tyrosine phosphorylation regulates alpha II spectrin cleavageby calpain. Mol Cell Biol 22:3527e3536.
24. Mitra SK, Hanson DA, Schlaepfer DD (2005) Focal adhesion kinase: In command andcontrol of cell motility. Nat Rev Mol Cell Biol 6(1):56e68.
25. Lysechko TL, Cheung SM, Ostergaard HL (2010) Regulation of the tyrosine kinase Pyk2by calcium is through production of reactive oxygen species in cytotoxic T lympho-cytes. J Biol Chem 285:31174e31184.
26. Roos J, et al. (2005) STIM1, an essential and conserved component of store-operatedCa2+ channel function. J Cell Biol 169:435e445.
27. Yuan JP, Zeng W, Huang GN, Worley PF, Muallem S (2007) STIM1 heteromultimerizesTRPC channels to determine their function as store-operated channels. Nat Cell Biol 9:636e645.
28. Taylor WR, Stark GR (2001) Regulation of the G2/M transition by p53. Oncogene 20:1803e1815.
29. Hutchins JR, Dikovskaya D, Clarke PR (2003) Regulation of Cdc2/cyclin B activation inXenopus egg extracts via inhibitory phosphorylation of Cdc25C phosphatase by Ca(2+)/calmodulin-dependent protein [corrected] kinase II. Mol Biol Cell 14:4003e4014.
30. Abbas T, Dutta A (2009) p21 in cancer: Intricate networks and multiple activities. NatRev Cancer 9:400e414.
31. Xu P, et al. (2006) Aggregation of STIM1 underneath the plasma membrane inducesclustering of Orai1. Biochem Biophys Res Commun 350:969e976.
32. Albini A, et al. (1987) A rapid in vitro assay for quantitating the invasive potential oftumor cells. Cancer Res 47:3239e3245.
33. Chiu WT, Tang MJ, Jao HC, Shen MR (2008) Soft substrate up-regulates the interactionof STIM1 with store-operated Ca2+ channels that lead to normal epithelial cell apo-ptosis. Mol Biol Cell 19:2220e2230.
15230 | www.pnas.org/cgi/doi/10.1073/pnas.1103315108 Chen et al.






![Molecule-radiation and molecule-molecule …Introduction Molecular quantum electrodynamics (QED) [1,2] is the most suitable formalism for the description of the interaction of a radiation](https://img.pdfslide.us/doc/110x75/5ec9d9ac19d4756e1547e308/molecule-radiation-and-molecule-molecule-introduction-molecular-quantum-electrodynamics.jpg)






















