Embed Size (px)
Citation preview

Original Contributions Am J Otolaryngol 10:371-375.1989
Calcium Gradients in Inner Ear Endolymph
ALEC N. SALT, PHD, NAOKI INAMURA, MD, RUEDIGER THALMANN, MD, AND ARTI VORA, BS
Recent studies suggest that endolymphatic hydrops resulting from the ablation of the endolymphatic duct and sac in guinea pigs may be caused by a disturbance of endolymph calcium homeostasis. A similar disturbance of calcium homeostasis could represent the underlying cause of Menibre’s disease. In this study, we mapped the calcium concentra- tions and electrical potentials throughout the endolymphatic system in normal guinea pigs. Large concentration differences exist between different compartments, including a more than twofold increase along the length of the cochlea. The electrochemical potential for calcium (the force driving passive longitudinal calcium movement) was calculated for all the endolymphatic compartments. The results show that endolymph is extremely in- homogenous with respect to calcium potentials. On the basis of these potentials, it appears that calcium is transported into endolymph in the cochlea and out of endolymph in the saccule and utricle. The possibility that endolymphatic hydrops arises from a disturbance in longitudinal flow of calcium, rather than in longitudinal volume flow, is considered. AM J OTOLARYNGOL 10:371-375. 0 1989 by W.B. Saunders Company. Key words: calcium, cochlea, endolymph, hydrops.
Bosher and Warren1 first reported the extremely low calcium content of cochlear endolymph rela- tive to other extracellular fluids. Using flame pho- tometry, they measured the total calcium content of endolymph in the rat at 2.3 x lo-’ mol/L, while with ion selective microelectrodes, the ion- ized calcium content was found to be 3.0 x 1O-5
mol/L. These results were not significantly differ- ent, demonstrating that virtually all endolymph calcium is in ionized form. This low calcium level has since been confirmed by ion-selective micro- electrode methods in two studies: Ikeda et al2 re- ported a concentration of 2.2 x 10e5 mol/L in the basal turn of albino guinea pigs and Ninoyu and Meyer zum Gottesberge3 measured a level of 2.7 x
10m5 mol/L in the third turn of pigmented guinea pigs.
A significant disturbance of endolymphatic Ca++ after hydrops was induced by ablation of the endolymphatic sac and duct has been reported to occur in guinea pigs. This contrasts with the previous demonstration that the concentrations of many electrolytes (K, Na, and Cl) are unchanged in the endolymph of animals with hydrops.4 Us-
Received May 12, 1989, from the Department of Otolaryn- gology, Washington University School of Medicine, 517 S Eu- clid Ave, St Louis, MO 63110. Accepted for publication June 15, 1989.
Supported by program project grant DC 00384. Address correspondence and reprint requests to Alec N.
Salt, PhD, Department of Otolaryngology, Washington Univer- sity School of Medicine, 517 S Euclid Ave, St. Louis, MO 63110.
0 1989 by W.B. Saunders Company. 0196-0709/89/1006-0009$5.00/O
ing laser microprobe mass analysis, Meyer zum Gottesberge and Kaufmann’ demonstrated that the intracellular Ca+ + levels of secretory cells of the semicircular canals and utricle increased by up to two orders of magnitude. In a subsequent study, Ninoyu and Meyer zum Gottesberge’ re- ported the Ca+ + content of the cochlear en- dolymph to be increased by up to a factor of 10 in hydropic ears. The degree of Ca+ + increase was well-correlated with the degree of suppression of endocochlear potential. It has been suggested that the endolymphatic sac may play a major role in the homeostasis of Ca+ + in the inner ear.5 Distur- bance of the endolymphatic Ca+ + level is pre- sumed to be involved in the generation of en- dolymphatic hydrops, either by creating an os- motic load’ or by disturbing the homeostasis of endolymph potassium content.7
These recent data are more consistent with the hypothesis that endolymphatic hydrops results from a disturbance of Caf + homeostasis rather than from a disturbance of the volume flow of endolymph between the cochlea and endolym- phatic sac, as is generally believed. This view is also supported by recent studies which have shown that the longitudinal flow rate of en- dolymph is exceedingly 10w.~-‘~ Therefore, longi- tudinal movements of ions may take place more rapidly when driven by prevailing electrical and concentration gradients rather than by the longi- tudinal bulk flow of fluid.
The purpose of this study was to measure the longitudinal electrical and chemical gradients for Ca++ so that the direction of longitudinal Ca+ +
371

372 CALCIUM GRADIENTS IN ENDOLYMPH
flows (driven by forces other than volume flow) could be assessed. Direct current potential and Ca++ concentration were measured in all four turns of the guinea pig cochlea, the saccule and the utricle. Equivalent measurements have been
recently reported for the semicircular canal and endolymphatic sac.3
METHODS
Pigmented guinea pigs (250 to 350 g) were anesthe- tized by the intraperitoneal administration of 35 mg/kg sodium pentobarbital. Supplementary anesthetic doses and gallamine triethiodide (muscle relaxant] were given intravenously. Animals were artificially venti- lated through a tracheal cannula. In each case, the co- chlea was exposed by a ventrolateral approach to the auditory bulla, leaving the middle ear function intact.
the electrode tip within the spiral ligament before in- sertion into endolymph. After measurement in each turn, the electrode response was checked in one stan- dard solution, allowing the stability of electrodes to be monitored. The series of measurements (all four turns) was then repeated, after which the electrode was re- calibrated. The measured Ca+ + concentrations were not significantly different between the two measure- ments (paired t test; n = 40), so the data from both measurements were combined as a single statistic.
In a second series of experiments, the Ca+ + levels of the utricle and saccule were measured. Access to the vestibular labyrinth was gained by carefully removing the stapes. After recording the perilymph composition, electrodes were inserted into endolymph through the membranous tissues. In these experiments, only one measurement was taken at each location because there was a tendency for the endolymphatic compartment to collapse during subsequent penetrations.
Endolymphatic Ca+ + was measured using double- barreled, ion-selective microelectrodes. Double- barreled electrodes were pulled from borosilicate glass, and one barrel (the ion-sensitive barrel) was silanized with dimethyldichlorosilane (Sigma, St Louis, MO) and filled with 500 mmol/L CaCl,. The other barrel (the po- tential or reference barrel) was filled with 500 mmol/L NaCl. Fluka 21048 Ca+ + exchanger (Fluka Chemical Corporation, Ronkonkoma, NY) was then drawn into the tip of the ion barrel. The tip diameter of the elec- trodes was 1 to 2 pm. Electrodes were calibrated in EGTA-buffered standard solutions, as described by Per- shadsingh and McDonald,‘l using a constant back- ground of 150 mmol/L KCl. Electrode responses were close to linear throughout the range in which they were calibrated (10e6 to low3 mmol/L), as previously reported.” We found that Ca+ + buffering of the stan- dards was not essential and, for concentrations above 10~~ mmol/L, extremely similar results were obtained using nonbuffered standard solutions. The mean slope of the electrode response was 26.9 ? 4.8 mV/decade change in Ca+ + (-c SD; n = 20).
RESULTS
In the first series of experiments (n = lo), endolym- phatic Ca+ + levels were measured in all four turns of the cochlea. Access was achieved by thinning the otic capsule with a flap knife and making a small fenestra through the thinned bone with a fine pick. The typical diameter of fenestrae was approximately 30 Frn, so that damage to the underlying spiral ligament was minimal. Fenestrae were made in all four turns in a randomized sequence. The experimental procedure involved cali- brating the electrode, then recording the perilymph and endolymph composition in each turn in a randomized sequence. Perilymph composition was measured with
Electrical potentials and Caf + concentrations
recorded in the four cochlear turns, the saccule and the utricle, are summarized in Table 1. These results show that the Ca+ + content throughout
the endolymphatic system is markedly lower than in the perilymph. On the other hand, endolymph is clearly not homogeneous with respect to Ca+ + , and significant differences are observed in differ- ent locations. Even within the cochlea it can be
seen that the endolymph Ca+ + content more than doubles from base to apex. One possible explana- tion of the measured Ca+ + gradient is that it arises as a direct result of the endolymphatic po-
tential (EP) gradient which is known to exist from base to apex in the cochlea. Higher positive po-
tentials could result in greater Ca+ + efflux and correspondingly lower Ca+ + levels. Figure 1 shows the relationship between the individual EP levels and endolymph Ca+ + concentrations re- corded in this study. If Ca+ + was distributed pas-
sively in cochlear endolymph, a tenfold change in Ca++ should result from an EP difference of 31 mV (based on the Nernst equation]. The regres- sion line corresponding to the data shows a slope
of 73 mV/decade, demonstrating that the Ca+ + gradient is less than that expected on the basis of the potential gradient. Thus, the gradient of Ca’ +
TABLE 1. Calcium Content of Inner Ear Fluids
COCHLEA
Perilymph Ca (mmol/L)
Endolymph Ca (pmol/L)
Potential (mV)
UTRICLE SACCULE I II III lv
0.80 0.86 1.36 1.27 1.57 1.29 (0.40) (0.20) (0.53) (0.40) (0.57) (0.44)
133 17.1 23.7 31.7 40.6 (17) (E) (4.1) (5.6) (4.4) (9.3)
(E) 91.2 83.0 76.8 69.1
(2.9) (1.61 (4.31 (4.3)
n 5 10 10 10
NOTE. The values are the mean (SD in parentheses) of the number of experiments shown (n).
10 10
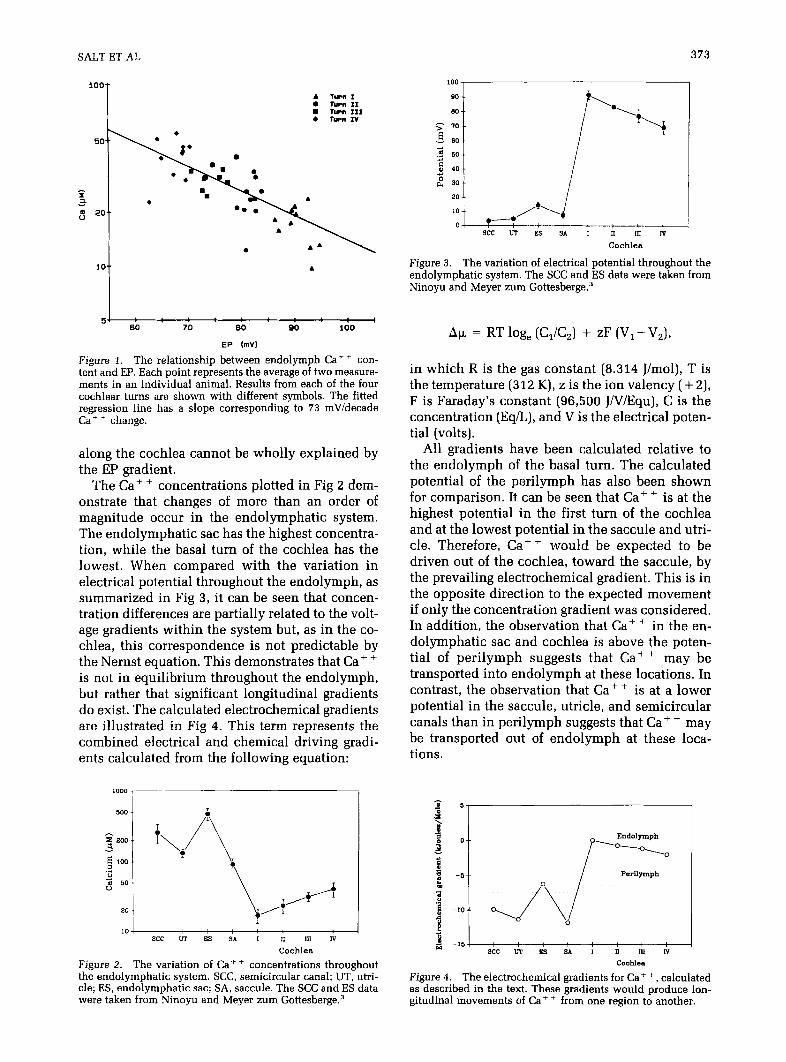
SALT ET AL 373
j; t . .
I . AA
I:: 60 70 00 90 100
EP (mv)
Figure I. The relationship between endolymph Ca+ + con- tent and EP. Each point represents the average of two measure- ments in an individual animal. Results from each of the four cochlear turns are shown with different symbols. The fitted regression line has a slope corresponding to 73 mV/decade Ca + + change.
along the cochlea cannot be wholly explained by the EP gradient.
The Ca++ concentrations plotted in Fig 2 dem- onstrate that changes of more than an order of magnitude occur in the endolymphatic system. The endolymphatic sac has the highest concentra- tion, while the basal turn of the cochlea has the lowest. When compared with the variation in electrical potential throughout the endolymph, as summarized in Fig 3, it can be seen that concen- tration differences are partially related to the volt- age gradients within the system but, as in the co- chlea, this correspondence is not predictable by the Nernst equation. This demonstrates that Ca+ + is not in equilibrium throughout the endolymph, but rather that significant longitudinal gradients do exist. The calculated electrochemical gradients are illustrated in Fig 4. This term represents the combined electrical and chemical driving gradi- ents calculated from the following equation:
,000
104 : I I xc UT ES s* I D IU N
Cochlea
Figure 2. The variation of Ca++ concentrations throughout the endolymphatic system. SCC, semicircular canal; UT, utri- cle; ES, endolymphatic sac; SA, saccule. The SCC and ES data were taken from Ninoyu and Meyer zum Gottesberge.3
see u7 ES SA I II ill N
Cochlea
Figure 3. The variation of electrical potential throughout the endolymphatic system. The XC and ES data were taken from Ninoyu and Meyer zum Gottesberge.3
Au = RT log, (C,/C,) + zF (V, -V,),
in which R is the gas constant (8.314 J/mol), T is the temperature (312 K), z is the ion valency ( + 2), F is Faraday’s constant (96,500 JnTiEqu), C is the concentration (Eq/L), and V is the electrical poten- tial (volts).
All gradients have been calculated relative to the endolymph of the basal turn. The calculated potential of the perilymph has also been shown for comparison. It can be seen that Ca+ + is at the highest potential in the first turn of the cochlea and at the lowest potential in the saccule and utri- cle. Therefore, Cat + would be expected to be driven out of the cochlea, toward the saccule, by the prevailing electrochemical gradient. This is in the opposite direction to the expected movement if only the concentration gradient was considered. In addition, the observation that Caf + in the en- dolymphatic sac and cochlea is above the poten- tial of perilymph suggests that Ca+ + may be transported into endolymph at these locations. In contrast, the observation that Ca+ + is at a lower potential in the saccule, utricle, and semicircular canals than in perilymph suggests that Ca+ + may be transported out of endolymph at these loca- tions.
. d 5 o 0. O-0 Endolymph
‘0 s
5
& -6
-- /
Perilymph cl
a
j -so-- :
0, /\ 0 0
h ”
Figure 4. The electrochemical gradients for Ca+ +, calculated as described in the text. These gradients would produce lon- gitudinal movements of Ca+ + from one region to another.

374 CALCIUMGRADIENTSINENDOLYMPH
DISCUSSION
While it has been known for some time that the electrical potentials and ionic composition of co- chlear and vestibular endolymph are markedly different, it has only recently been documented that differences occur within the cochlea itself. Sterkers et a113s14 reported the existence of signif- icant gradients for EP, endolymph potassium, chloride, and osmolarity in the rat cochlea. A sim- ilar decrease of EP and endolymph K concentra- tion from base to apex was measured in guinea pigs. l5 While potassium and chloride appear to decrease from base to apex, our study demon- strates that the level of Ca+ + increases along the length of the cochlea by a factor of greater than 2. This increase in Ca+ + is not wholly explained by the EP gradient, and an electrochemical gradient exists that would tend to drive Ca+ + from base to apex. These data also support the view that Ca+ + must be transported into endolymph at least in the basal turn of the cochlea, as suggested by Bosher and Warren’ and Ikeda et al.’ The alterna- tive possibility, that endolymph Ca+ + is above the equilibrium potential with respect to peri- lymph because of Ca’ + entry from the saccule, is unlikely since the electrochemical gradient for Ca++ is clearly directed from the cochlea to the saccule.
Significant gradients in endolymph composi- tion would not be expected if high rates of en- dolymph flow occurred from one region to an- other. These data are therefore consistent with the recent finding that the rate of endolymph flow within the cochlea is extremely s10w.‘~ Therefore, it appears that endolymph homeostasis is domi- nated by local ion transport mechanisms, rather than by a process in which endolymph is secreted in the cochlea and resorbed by the endolympatic sac. It is not known whether the flows of Caf ’ induced by the prevailing electrochemical gradi- ents are significant with respect to Ca+ + turnover in each compartment, or whether they are impor- tant for normal function. Alternatively, the differ- ent concentrations may represent independent homeostatic mechanisms in different regions.
One interesting aspect of these data is the find- ing that the utricle and saccule have the lowest electrochemical potentials with respect to Ca+ + [well below that of perilymph), while those of the cochlea and endolymphatic sac are higher. This raises the question of the apparently differing re- quirements for Cat + between the auditory and vestibular portions of the inner ear. Both the sac- cular and utricular maculae have otoconia that contain calcium carbonate in the form of calcite. In contrast, the equivalent structure in the co- chlea, the tectorial membrane, does not have such
a mineral deposit. The otoconial Ca+ + is not static, but has a turnover with a half-time of ap- proximately 11 days in gerbils.16 Although it may be expected that those regions with otoconia would require Ca+ + transport into the en- dolymph, this does not appear to be the case. However, a distinction must be made between the importance of the Ca+ + electrochemical gradient and the Ca+ + concentration. For transport be- tween perilymph and endolymph, or for move- ments between endolymphatic compartments, Ca++ electrochemical potential is the most im- portant factor. In contrast, the solubility of otoco- nia will depend only on the concentration of Ca++, as they are immersed in solution at the endolymphatic potential, so there is no voltage gradient across them. The low electrochemical potential for Ca+ + is more likely the result of Ca++ uptake from endolymph in these regions or resorption of Ca+ + leached from otoconia [ie, processes involving the tissues bounding the en- dolymph) and may not be of direct importance to the otoconia.
Changes in endolymphatic Ca+ + content may play a role in a number of dysfunctional condi- tions that affect the inner ear, including Meniere’s disease. There is no doubt that Caf + homeostasis is disturbed in guinea pigs in which endolym- phatic hydrops is induced by the ablation of the endolymphatic duct and sac.5*6 In addition, it has been reported that degeneration of otoconia may occur in patients with Meniere’s disease.17 It is possible that the flow of Cat +, induced by the electrochemical gradient demonstrated here, could be important to normal function. The co- chlea could represent a source of Cat+ (Ca’ + transported into endolymph) and the saccule and utricle could represent sinks for Ca+ + (Caf + transported out of endolymph). This is analogous to the prevalent view that endolymph is secreted in the cochlea and resorbed by the endolymphatic sac, except that the flow is of Ca+ + rather than of fluid. This flow would be driven by an electro- chemical gradient rather than by hydrostatic pres- sure. However, it is also possible that the longitu- dinal movement of Ca+ + is insignificant in the normal ear and that measured levels represent the independent homeostasis of each compartment. Even if this is true, it is highly probable that a disturbance of the Ca+ + level in one region would result in nonphysiologic flows of Cat+ within the endolymph and the subsequent distur- bance of more than one region. The changes that occur during the development of hydrops in ex- perimental animals [and of Meniere’s disease in humans) are highly complex and may depend on interactions between disturbances of ions and electrical fields. Understanding these interactions

SALT ET AL 375
should lead to the development of new therapies for Meniere’s disease.
References
1. Bosher SK, Warren RL: Very low calcium content of co- chlear endolymph, an extracellular fluid. Nature 1978; 273:377-378 2. Ikeda K, Kusakari J, Takasaka T, et al: The Ca+ + activity
of cochlear endolymph of the guinea pig and the effect of in- hibitors. Hear Res 1987; 26:117-125
3. Ninoyu 0, Meyer zum Gottesberge AM: Ca+ + activity in the endolymphatic space. Arch Othorhinolaryngol 1986; 243:141-142 4. Konishi T, Salt AN, Kimura RS: Electrophysiological
studies of experimentally induced endolymphatic hydrops in guinea pigs, in Vosteen K-H, et al (eds): Meniere’s Disease Pathogenesis and Treatment. New York, Georg Thieme Verlag, 1981, pp 47-58 5. Meyer zum Gottesberge AM, Kaufmann R: Is an imbal-
anced calcium homeostasis responsible for the experimentally induced endolymphatic hydrops? Acta Otolaryngol (Stockh) 1986; 102:93-98
6. Ninoyu 0, Meyer zum Gottesberge AM: Changes in Ca+ + activity and DC potential in experimentally induced endolym- phatic hydrops. Arch Otorhinolaryngol 1986; 243:106-107
7. Thalmann R, Salt AN, DeMott J: Endolymph volume reg- ulation: Possible mechanisms, in Nadol JB, Jr (ed): Second In- ternational Symposium on M&-n&e’s Disease. Amsterdam, Kugler, 1989 (in press]
8. Salt AN, Thalmann R, Marcus DC, et al: Direct measure- ment of longitudinal endolymph flow rate in the guinea pig cochlea. Hear Res 1986; 23:141-151
9. Salt AN, Thalmann R: Interpretation of endolymph flow results. Hear Res 1988: 33:279-281 (letter) \ I
IO. Salt AN, Thalmann R: Rate of longitudinal flow of co- chlear endolvmnh, in Nadol TB, lr led): Second International Symposium on *M&n&e’s Disease. Amsterdam, Kugler, 1989 (in press)
11. Pershadsingh HA, McDonald JM: A high affinity cal- cium-stimulated magnesium-dependent adenosine triphos- phatase in rat adipocyte plasma membranes. J Biol Chem 1980; 255:4087-4093
12. Lanter F, Steiner RA, Ammann D, et al: Critical evalua- tion of the applicability of neutral carrier-based calcium selec- tive microelectrodes. Anal Chim Acta 1982; 135:51-59
13. Sterkers 0, Saumon G, Trans Ba Huy P, et al: Electro- chemical heterogeneity of the cochlear endolymph: Effect of acetazolamide. Am J Physiol 1984; 246:F47-F53
14. Sterkers 0, Ferrary E, Amiel C: Inter- and intracompart- mental osmotic gradients within the rat cochlea. Am J Physiol 1984; 247:F602-606
15. Salt AN, Thalmann R: Cochlear fluid dynamics, in John AF, Santos-Sacchi J (eds]: Physiology of the Ear. New York, Raven, 1988, pp 341-357
16. Preston RE, Johnsson LG, Hill JH, et al: Incorporation of radioactive calcium into otolithic membranes and middle ear ossicles of the gerbil. Acta Otolaryngol 1975; 80:269-275
17. Johnsson LG, Rouse RC, Wright CG, et al: Pathology of neuroepithelial suprastructures of the human inner ear, Am J Otolaryngol 1982; 3:77-90