Embed Size (px)
Citation preview

Traffic 2011; 12: 1839–1849 © 2011 John Wiley & Sons A/S
doi:10.1111/j.1600-0854.2011.01271.x
Caenorhabditis elegans Numb Inhibits EndocyticRecycling by Binding TAT-1 AminophospholipidTranslocase
Lars Nilsson, Eva Jonsson and Simon Tuck∗
Umea Center for Molecular Medicine, Umea University,SE-901 87 Umea, Sweden*Corresponding author: Simon Tuck,[email protected]
Numb regulates endocytosis in many metazoans, but
the mechanism by which it functions is not completely
understood. Here we report that the Caenorhabditis ele-
gans Numb ortholog, NUM-1A, a regulator of endocytic
recycling, binds the C isoform of transbilayer amphipath
transporter-1 (TAT-1), a P4 family adenosine triphos-
phatase and putative aminophospholipid translocase
that is required for proper endocytic trafficking. We
demonstrate that TAT-1 is differentially spliced during
development and that TAT-1C-specific splicing occurs in
the intestine where NUM-1A is known to function. NUM-
1A and TAT-1C colocalize in vivo. We have mapped the
binding site to an NXXF motif in TAT-1C. This motif is not
required for TAT-1C function but is required for NUM-1A’s
ability to inhibit recycling. We demonstrate that num-1A
and tat-1 defects are both suppressed by the loss of the
activity of PSSY-1, a phosphatidylserine (PS) synthase.
PS is mislocalized in intestinal cells with defects in tat-1
or num-1A function. We propose that NUM-1A inhibits
recycling by inhibiting TAT-1C’s ability to translocate PS
across the membranes of recycling endosomes.
Key words: C. elegans, endocytosis, numb, phos-
phatidylserine, recycling, tat-1
Received 2 March 2011, revised and accepted for
publication 19 August 2011, uncorrected mauscript
published online 22 August 2011, published online 14
September 2011
Numb regulates endosomal trafficking in both inverte-brates and vertebrates including mammals (1). Numbhas been shown to bind to the endosomal protein,α-adaptin and to proteins containing epsin homologydomains (EHD) (2–4). However, although the interactionswith EHD proteins and α-adaptin are important for asubset of Numb’s functions, neither type of interactionfully accounts for Numb’s activity in regulating trafficking.Numb orthologs from some species lack the NPF motifrequired for binding to EHD proteins but are neverthelessactive in regulating endocytosis (2,5). Mutations in both α-adaptin and numb in Drosophila cause defects in cell fatespecification events mediated by Notch signaling (3), buta mutant Numb protein lacking the ability to bind α-adaptinis nevertheless active (6).
NUM-1A, the Caenorhabditis elegans Numb ortholog,inhibits endocytic recycling (5). Elevated levels of NUM-1A cause enlargement of the endocytic recyclingcompartment (ERC), a phenotype closely resembling thatcaused by loss-of-function mutations in rme-1, whichencodes an EHD protein (5,7). Loss of num-1A activityon the other hand bypasses the requirement for rme-1 inrecycling implying that NUM-1A must act, at least in part,independently of RME-1 (5). Precisely how NUM-1A actsto regulate recycling in C. elegans, however, is not known.
P-type adenosine triphosphatases of the Drs2p fam-ily (P4-ATPases) are required for proper vesicle trans-port in fungi, including Saccharomyces cerevisiae (8–10)and Magnaporthe grisea (11), in the multicellular eukary-otic model organisms Arabidopsis thaliana (12) andC. elegans (13,14) and in cultured mouse mastocytomacells (15). P4-ATPases are multipass transmembrane pro-teins thought to act by promoting the translocation ofphosphatidylserine (PS) and phosphatidylethanolaminewithin membranes from one leaflet of the bilayer tothe other (16,17). Purified preparations of the yeast P4-ATPase, Drs2p, and of a related bovine protein, Atp8a2,catalyze the translocation of PS across membranes ofprotein-free liposomes (18,19), indicating that these pro-teins have intrinsic translocase activity. Translocation of PSfrom the exoplasmic to cytosolic leaflet is required for thegeneration and maintenance of lipid asymmetry observedwithin many membranes in eukaryotic cells (20–23). Themodulation of this asymmetry is thought to be impor-tant for correct vesicle transport, but very little is knownabout how the activities of P4-ATPases themselves areregulated in multicellular eukaryotes.
Mutations in the C. elegans P4-ATPase TAT-1 causedefects in endocytic recycling and cause enlargedendolysosomal organelles to accumulate within theintestine and hypodermis (13,14). TAT-1 localizes toendosomal membranes and requires the activity of achaperone CHAT-1 for function, but how the activity ofTAT-1 is modulated is not understood. tat-1 mutantsdisplay abnormal distribution of PS at the plasmamembrane and within coelomocytes (14,24,25).
We report here on experiments aimed at establishingthe molecular mechanism by which NUM-1A regulatesrecycling. Experiments are described that indicate thatNUM-1A binds directly to a specific isoform of TAT-1, TAT-1C, which we show is present in tissues and organellesin which NUM-1A is known to function. The effects ofaltering NUM-1A and TAT-1 levels on the transport of PS
www.traffic.dk 1839

Nilsson et al.
and its distribution within membranes are reported. Basedon our findings, we propose a model in which NUM-1Ainhibits recycling by downregulating TAT-1’s function as aPS translocase.
Results
NUM-1A binds to TAT-1C
To understand how num-1A regulates endocytic recyclingin C. elegans, we performed a yeast 2-hybrid (Y2H) screenfor proteins that could bind NUM-1A (see section onMaterial and Methods). From a screen of 8 × 105 yeastcolonies, we isolated a partial cDNA clone spanning the3′- end of the C isoform of the tat-1 gene (Figure 1A).The cDNA clone isolated contained an early in-frame stopcodon and encodes a protein with just 32 amino acids fromthe very C-terminal end of TAT-1C (Figure 1B). These 32amino acids were found to be sufficient for robust bindingto NUM-1A (Figure 1C). The interaction with TAT-1 is nota characteristic of PTB domain proteins per se becausethe PTB domain from DAB-1, a protein that regulates bothendocytosis and secretion in C. elegans (26,27), failed tointeract with TAT-1C in the Y2H assay (Figure 1C). PTBdomains of the class to which the NUM-1A PTB domainbelongs have been shown to mediate binding to tworelated motifs, NPXY (where Y is unphosphorylated andX is any amino acid) or NXXF (28). The part of TAT-1Cencoded by the Y2H clone we isolated contained anNXXF motif, NRAF (Figure 1A,B). Replacement of N orF with A abolished the ability of the TAT-1C polypeptideto bind to the NUM-1A PTB domain in the Y2H assay(Figure 1C), implying that the NXXF motif is necessaryfor the interaction. The TAT-1C and NUM-1A proteinswere also able to bind to one another in vitro (Figure 1D).Furthermore, mutation of the NXXF motif severelyreduced binding in vitro. Taken together, these resultsdemonstrate that the PTB domain of NUM-1A interactswith the C-terminal domain of TAT-1C via an NXXF motif.
tat-1C-specific splicing occurs in the intestine
NUM-1A labels the ERC in C. elegans intestinal cells andfunctions in these cells to regulate recycling (5). The tat-1promoter is active in the intestine (13,14), but previouswork has not established in which tissues, the differenttat-1 spliced isoforms are found. To determine whethertat-1C-specific splicing occurs in the intestine, we gen-erated worms carrying a green fluorescent protein (GFP)reporter, pVB496LN (Figure 1A), designed to detect splic-ing to the tat-1C-specific exon. Expression of the reporterwas seen in a subset of tissues in which the tat-1 pro-moter is active (13) including the intestine (Figure 2A–D).Expression was absent from other cells in which the tat-1promoter is active including hypodermal cells and neuronsin the ventral cord (Figure 2E–L). Thus, splicing of tat-1 isdifferentially regulated, and splicing to the tat-1C-specificexon occurs in the intestine.
In the intestine, elevated levels of NUM-1A cause enlargedERCs to accumulate, which are decorated by NUM-1A (5).
Previous studies have shown that TAT-1C protein labelsmultiple endosomal organelles including the ERC (13,14).TAT-1C protein also labeled the enlarged ERC organellespresent in worms harboring elevated levels of NUM-1A(Figure 2M,N).
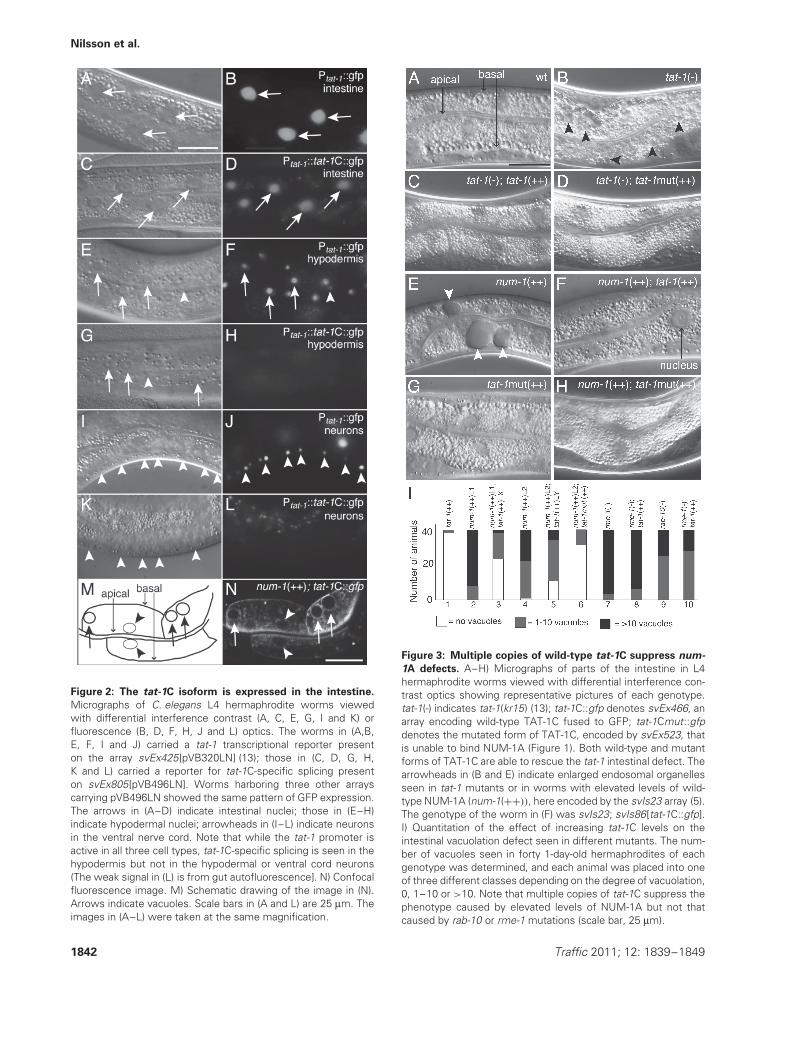
Multiple copies of tat-1C suppress num-1A defects
If NUM-1A acted to inhibit recycling by inhibiting TAT-1C function, then elevated levels of TAT-1C might beexpected to suppress defects caused by increased levelsof NUM-1A. In agreement with this prediction, transgenearrays containing multiple copies of a wild-type tat-1CcDNA strongly suppressed the accumulation of abnor-mal recycling endosomes caused by multiple copies ofnum-1A (Figure 3E,F,I). Increased copy number of a geneencoding a TAT-1C mutant with a mutated NXXF motifalso suppressed the accumulation of abnormal recyclingendosomes caused multiple copies of num-1A. Moreover,it did so more efficiently than increased copy number ofthe wild-type tat-1C gene (Figure 3E,G,H,I). Increasingtat-1C gene copy number did not affect the expressionof NUM-1A::GFP (Figure S1), suggesting that the extracopies of tat-1C do not suppress the accumulation ofabnormal recycling endosomes caused multiple copies ofnum-1A by affecting NUM-1A protein accumulation or sta-bility. The mutant form of TAT-1C also efficiently rescuestat-1 defects (Figure 3B–D). Thus, while NUM-1A requiresbinding to TAT-1C to function, the converse is not true.
Increasing tat-1C gene copy number had no effect onthe phenotype of rab-10 loss of function (Figure 3I,Figure S2A,B), which is morphologically very similar to thatcaused by increased num-1A gene copy number (5,29).Thus, elevating tat-1C gene copy number does notsuppress vacuolation nondiscriminately. In the case ofrme-1 loss of function, which also causes defectsmorphologically similar to those caused by increasednum-1A gene copy number (7), multiple copies of tat-1C modified the phenotype in a complex fashion. Whilethe number of vacuoles was not decreased (Figure 3I),the vacuoles were generally smaller than that in rme-1single mutants. Furthermore, some of the vacuoles had adifferent appearance from those in rme-1 single mutant:in particular, rather than having uniform, nonrefractile,aqueous interiors, they were filled with small, refractilevesicles similar to those normally found in the cytoplasmof intestinal cells. This effect was most marked in rme-1 animals in which the mutant form of TAT-1C wasexpressed (Figure S2D,E).
NUM-1A modulates intestinal transport of PS
In tat-1 mutants, when PS-binding proteins such asAnnexin V and LactC2-GFP are administered extracel-lularly, they ectopically label the plasma membraneof many cells (14,24,25). These observations stronglysuggest that TAT-1 is required to prevent the accumu-lation of PS in the outer leaflet of the plasma mem-brane. Within intestinal cells, TAT-1 protein is present
1840 Traffic 2011; 12: 1839–1849

Caenorhabditis elegans NUM-1A Binds to TAT-1
Figure 1: NUM-1A interacts with TAT-1C. A) The exons in the three tat-1 splice forms. Boxes represent exons, which are numbered1–19; lines represent introns. Filled regions denote coding regions. The stop codons in the three different splice forms are indicated.The tat-1 cDNA sequences present in the Y2H clone and other DNA constructs used in the Y2H assay are shown below on theright. The part of tat-1C encoding the NRAF motif is indicated. B) TAT-1C protein domain structure. The filled black boxes representpredicted transmembrane domains. The position of the ATPase catalytic site is indicated by a line below. The position of the NRAFmotif is indicated. C) Y2H analysis of the interaction between TAT-1C and NUM-1A. Eight different yeast cultures transformed withthe indicated plasmids were streaked onto a selective plate and allowed to grow for 3 days. NUM-1 indicates the NUM-1A-GAL4 DNABD fusion protein. The TAT-1 sequences present in the constructs encoding the different TAT-1-GAL4 AD fusion proteins are givenin (A). The plasmids pVB375LN and pVB376LN encode mutant TAT-1 fragments with the indicated amino acid sequence changes inthe NRAF motif. Fragments of p53 and SV40 large T antigen were used for the positive control. ‘Vector’ indicates empty vector andserves as a negative control. DAB-1 indicates the DAB-1 PTB domain-GAL4 BD fusion protein. D) Autoradiograph of sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel demonstrating that NUM-1A PTB domains and TAT-1C also interact in vitro.35S-methinionine-labeled NUM-1A PTB domain was incubated with the GST::TAT-1 fusion protein or with GST alone. Binding wasdetected between NUM-1A and wild-type GST::TAT-1C fusion protein (lane 2) but not between NUM-1A and mutant TAT-1C (lane 3) orbetween NUM-1A and GST (lane 4). In lane 1, NUM-1A protein alone was added. Below is a coomassie-stained SDS-PAGE gel showingthe input of the different GST proteins before mixing and subsequent pull-down with NUM-1A protein.
Traffic 2011; 12: 1839–1849 1841
Nilsson et al.
Figure 2: The tat-1C isoform is expressed in the intestine.
Micrographs of C. elegans L4 hermaphrodite worms viewedwith differential interference contrast (A, C, E, G, I and K) orfluorescence (B, D, F, H, J and L) optics. The worms in (A,B,E, F, I and J) carried a tat-1 transcriptional reporter presenton the array svEx425[pVB320LN] (13); those in (C, D, G, H,K and L) carried a reporter for tat-1C-specific splicing presenton svEx805[pVB496LN]. Worms harboring three other arrayscarrying pVB496LN showed the same pattern of GFP expression.The arrows in (A–D) indicate intestinal nuclei; those in (E–H)indicate hypodermal nuclei; arrowheads in (I–L) indicate neuronsin the ventral nerve cord. Note that while the tat-1 promoter isactive in all three cell types, tat-1C-specific splicing is seen in thehypodermis but not in the hypodermal or ventral cord neurons(The weak signal in (L) is from gut autofluorescence]. N) Confocalfluorescence image. M) Schematic drawing of the image in (N).Arrows indicate vacuoles. Scale bars in (A and L) are 25 μm. Theimages in (A–L) were taken at the same magnification.
Figure 3: Multiple copies of wild-type tat-1C suppress num-
1A defects. A–H) Micrographs of parts of the intestine in L4hermaphrodite worms viewed with differential interference con-trast optics showing representative pictures of each genotype.tat-1(-) indicates tat-1(kr15) (13); tat-1C::gfp denotes svEx466, anarray encoding wild-type TAT-1C fused to GFP; tat-1Cmut::gfpdenotes the mutated form of TAT-1C, encoded by svEx523, thatis unable to bind NUM-1A (Figure 1). Both wild-type and mutantforms of TAT-1C are able to rescue the tat-1 intestinal defect. Thearrowheads in (B and E) indicate enlarged endosomal organellesseen in tat-1 mutants or in worms with elevated levels of wild-type NUM-1A (num-1(++)), here encoded by the svIs23 array (5).The genotype of the worm in (F) was svIs23; svIs86[tat-1C::gfp].I) Quantitation of the effect of increasing tat-1C levels on theintestinal vacuolation defect seen in different mutants. The num-ber of vacuoles seen in forty 1-day-old hermaphrodites of eachgenotype was determined, and each animal was placed into oneof three different classes depending on the degree of vacuolation,0, 1–10 or >10. Note that multiple copies of tat-1C suppress thephenotype caused by elevated levels of NUM-1A but not thatcaused by rab-10 or rme-1 mutations (scale bar, 25 μm).
1842 Traffic 2011; 12: 1839–1849
Caenorhabditis elegans NUM-1A Binds to TAT-1
on many if not all endosomal membranes and alsostrongly decorates the apical plasma membrane, whichabuts the lumen of the intestine (13,14). To exam-ine directly whether TAT-1 can affect the transport ofPS across the apical membranes of intestinal cells,we examined the uptake of a fluorescently labeledPS derivative, 1-oleoyl-2-{6-[(7-nitro-2-1,3-benzoxadiazol-4-yl)amino]hexanoyl}-sn-glycero-3-phosphoserine (NDB-PS),into the intestine in wild-type and tat-1 mutant animals.When wild-type worms were fed NDB-PS, the dye rapidlyand efficiently labeled organelles within the intestinalcells (Figure 4B,I). Labeling was severely reduced in tat-1 mutants (Figure 4C,I). Thus, in the intestine, at least,TAT-1 does not solely prevent the movement of PS from
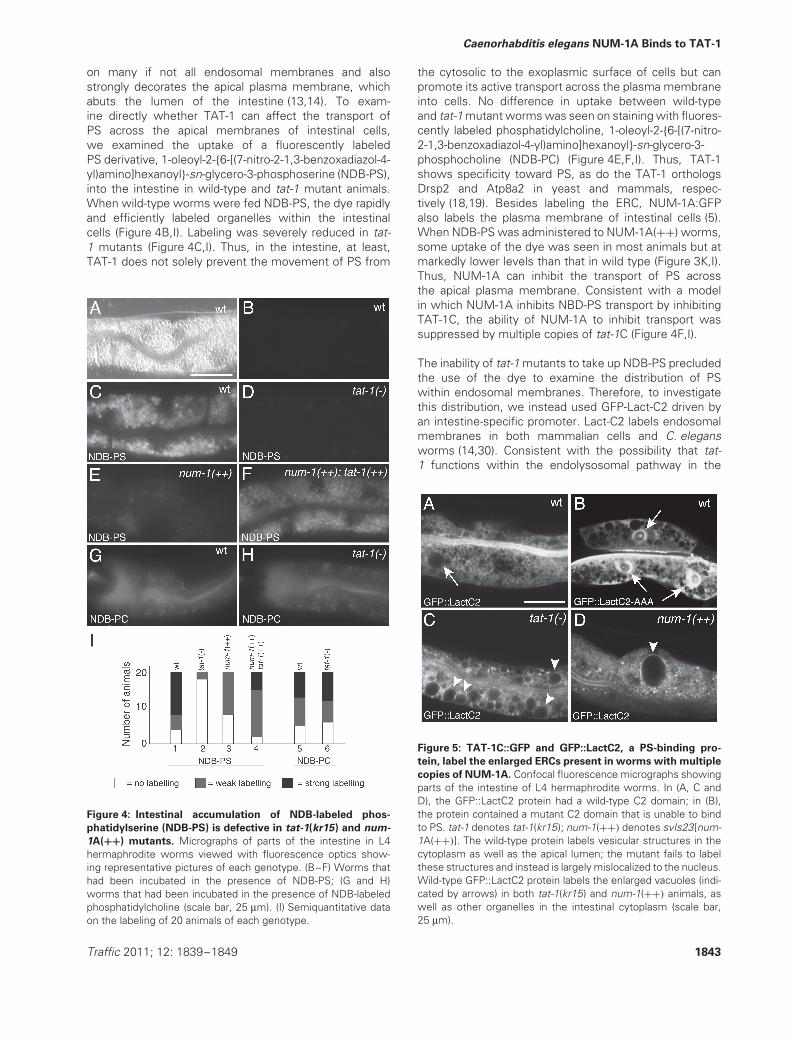
Figure 4: Intestinal accumulation of NDB-labeled phos-
phatidylserine (NDB-PS) is defective in tat-1(kr15) and num-
1A(++) mutants. Micrographs of parts of the intestine in L4hermaphrodite worms viewed with fluorescence optics show-ing representative pictures of each genotype. (B–F) Worms thathad been incubated in the presence of NDB-PS; (G and H)worms that had been incubated in the presence of NDB-labeledphosphatidylcholine (scale bar, 25 μm). (I) Semiquantitative dataon the labeling of 20 animals of each genotype.
the cytosolic to the exoplasmic surface of cells but canpromote its active transport across the plasma membraneinto cells. No difference in uptake between wild-typeand tat-1 mutant worms was seen on staining with fluores-cently labeled phosphatidylcholine, 1-oleoyl-2-{6-[(7-nitro-2-1,3-benzoxadiazol-4-yl)amino]hexanoyl}-sn-glycero-3-phosphocholine (NDB-PC) (Figure 4E,F,I). Thus, TAT-1shows specificity toward PS, as do the TAT-1 orthologsDrsp2 and Atp8a2 in yeast and mammals, respec-tively (18,19). Besides labeling the ERC, NUM-1A:GFPalso labels the plasma membrane of intestinal cells (5).When NDB-PS was administered to NUM-1A(++) worms,some uptake of the dye was seen in most animals but atmarkedly lower levels than that in wild type (Figure 3K,I).Thus, NUM-1A can inhibit the transport of PS acrossthe apical plasma membrane. Consistent with a modelin which NUM-1A inhibits NBD-PS transport by inhibitingTAT-1C, the ability of NUM-1A to inhibit transport wassuppressed by multiple copies of tat-1C (Figure 4F,I).
The inability of tat-1 mutants to take up NDB-PS precludedthe use of the dye to examine the distribution of PSwithin endosomal membranes. Therefore, to investigatethis distribution, we instead used GFP-Lact-C2 driven byan intestine-specific promoter. Lact-C2 labels endosomalmembranes in both mammalian cells and C. elegansworms (14,30). Consistent with the possibility that tat-1 functions within the endolysosomal pathway in the
Figure 5: TAT-1C::GFP and GFP::LactC2, a PS-binding pro-
tein, label the enlarged ERCs present in worms with multiple
copies of NUM-1A. Confocal fluorescence micrographs showingparts of the intestine of L4 hermaphrodite worms. In (A, C andD), the GFP::LactC2 protein had a wild-type C2 domain; in (B),the protein contained a mutant C2 domain that is unable to bindto PS. tat-1 denotes tat-1(kr15); num-1(++) denotes svIs23[num-1A(++)]. The wild-type protein labels vesicular structures in thecytoplasm as well as the apical lumen; the mutant fails to labelthese structures and instead is largely mislocalized to the nucleus.Wild-type GFP::LactC2 protein labels the enlarged vacuoles (indi-cated by arrows) in both tat-1(kr15) and num-1(++) animals, aswell as other organelles in the intestinal cytoplasm (scale bar,25 μm).
Traffic 2011; 12: 1839–1849 1843

Nilsson et al.
intestine to promote the transfer of PS across themembranes of endolysosomal organelles, cytoplasmicallyexpressed GFP-Lact-C2 labeled the enlarged endocyticorganelles seen in tat-1 mutants (Figure 5C). GFP-Lact-C2also labeled the enlarged ERCs found in NUM-1A(++)
animals (Figure 5D), indicating that PS is present in theouter membranes of the enlarged ERC organelles seen inthese animals. Mutant Lact-C2 protein that is unable tobind to PS (30) was mislocalized: it was present mostly inthe nucleus (Figure 5B).
PSSY-1, a phosphatidylserine synthase, is required
both for the expression of tat-1 endosomal defects
and for NUM-1A to inhibit recycling
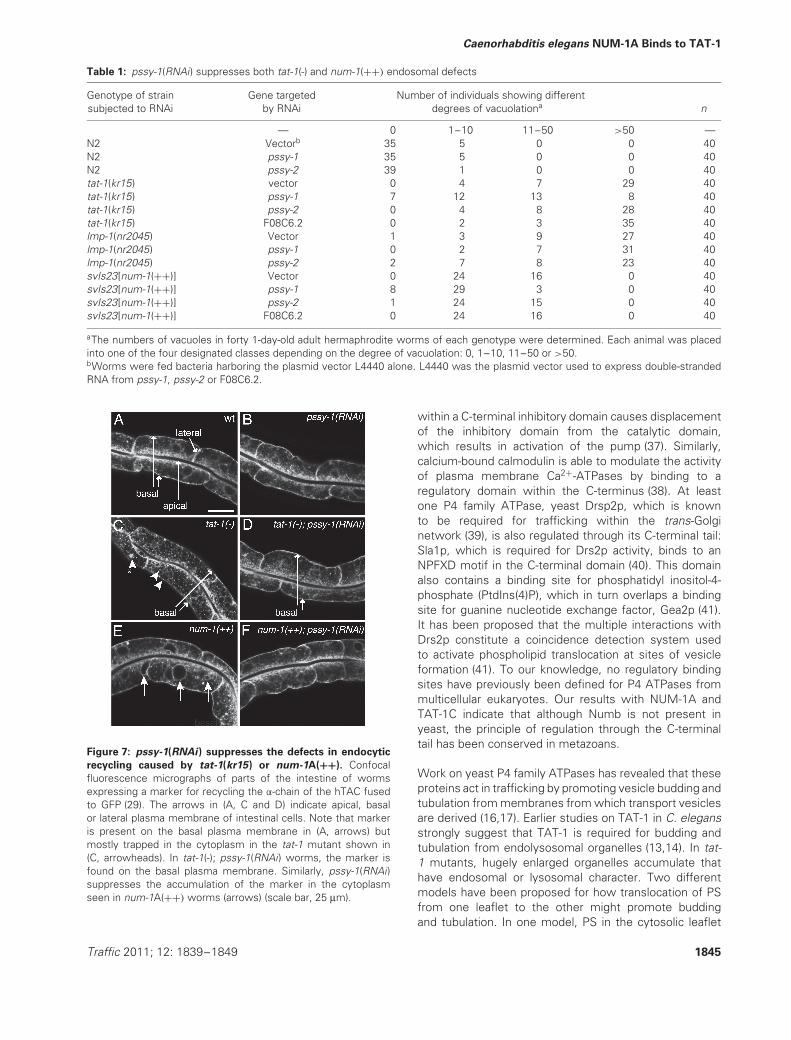
In mammalian cells, the synthesis of PS is catalyzed bytwo different PS synthases, PSS1 and PSS2. Two PSsynthases have also been identified in the C. elegansgenome, pssy-1 and pssy-2. RNAi of either gene in anotherwise wild-type genetic background did not give riseto any discernible developmental or endocytic defects(Figure 6B, Table 1). However, pssy-1(RNAi) [but notpssy-2(RNAi)] very efficiently suppressed the intestinaldefects seen in tat-1 mutants (Figure 6C,D,G,H, Table 1).In particular, whereas the intestines of tat-1(kr15) mutants
Figure 6: pssy-1(RNAi) suppresses the vacuolated intestine
phenotype caused by tat-1(kr15) or num-1A(++). Micrographsof parts of the intestine in L4 hermaphrodite worms viewed withdifferential interference contrast optics. The worms in (B, D, Fand H) were raised on HT115 bacteria expressing pssy-1 dsRNA.Those in (A, C, E and G) are negative controls in which the wormswere fed HT115 bacteria having only the empty L4440 RNAivector plasmid (scale bar, 25 μm).
were completely filled with vacuoles, those of tat-1(kr15);pssy-1(RNAi) animals showed almost no vacuolation. Incontrast, RNAi of the sole predicted phosphatidylcholinesynthase gene in C. elegans, F08C6.2, did not suppress(Table 1). Importantly, pssy-1(RNAi) did not suppress thevesicle phenotype caused by other C. elegans endosomalmutants (Figure 6E,F, Table 1). Thus, the suppressionis not the result of a general suppression of vesicleformation. Consistent with a model in which NUM-1A inhibits recycling by affecting TAT-1C on ERCorganelles, RNAi of pssy-1 strongly suppressed num-1A(++) intestinal vacuolation (Figure 6G,H).
To examine whether pssy-1(RNAi) could suppress theeffects of tat-1 and num-1A on endocytic recycling,we examined whether pssy-1(RNAi) could suppress thedefects seen in the recycling of the α-chain of the humanIL-2 receptor (hTAC) fused to GFP (a well-establishedmarker for recycling (29)). In worms with multiple copies ofnum-1A, or those mutant for tat-1, recycling of hTAC::GFPback to the plasma membrane is reduced (5,13). In bothcases, pssy-1(RNAi) efficiently suppressed the recyclingdefect (Figure 7).
Discussion
Endocytosis is of vital importance to eukaryotic cells fortheir ability to take up nutrients from their environment andfor the proper subcellular distribution of cell membraneproteins and lipids (34,35). Endocytic recycling is criticalfor the maintenance of proper plasma membranecomposition and for the rapid transport of a subsetof internalized proteins and lipids back to the plasmamembrane (36). It is thought that, in many cells, as muchas 50% of proteins, lipids and fluids taken into the cell arerecycled back to the cell surface within a few minutes (36).We and others have previously demonstrated that Numbprotein is an important regulator of recycling in bothmammalian cells and C. elegans and that it is enriched onrecycling endosomes (4,5). In this manuscript, we havepresented evidence that the C. elegans Numb ortholog,NUM-1A, functions to regulate recycling by modulating theability of the P4 family ATPase, TAT-1, to translocate PS.
The P4 subfamily of ATPases belong to the largerfamily of P-type ATPases of which five subfamilies,P1–P5, are recognized (9). Members of the P1, P2and P3 families transport mono- or divalent cationsacross cell membranes; both the function and regulationof P-type ATPases in these subfamilies have beenwell characterized. It is known that the activities ofthese types of P-type ATPases are frequently tightlyregulated, particularly at the posttranslational level (9).In the light of our findings with NUM-1A and TAT-1C,it is noteworthy that P2 and P3 subfamily membersgenerally contain regulatory domains in the cytosolic tailswhose functions can themselves be modulated (37,38).For example, phosphorylation of the plant H+ ATPase
1844 Traffic 2011; 12: 1839–1849
Caenorhabditis elegans NUM-1A Binds to TAT-1
Table 1: pssy-1(RNAi) suppresses both tat-1(-) and num-1(++) endosomal defects
Genotype of strainsubjected to RNAi
Gene targetedby RNAi
Number of individuals showing differentdegrees of vacuolationa n
— 0 1–10 11–50 >50 —N2 Vectorb 35 5 0 0 40N2 pssy-1 35 5 0 0 40N2 pssy-2 39 1 0 0 40tat-1(kr15) vector 0 4 7 29 40tat-1(kr15) pssy-1 7 12 13 8 40tat-1(kr15) pssy-2 0 4 8 28 40tat-1(kr15) F08C6.2 0 2 3 35 40lmp-1(nr2045) Vector 1 3 9 27 40lmp-1(nr2045) pssy-1 0 2 7 31 40lmp-1(nr2045) pssy-2 2 7 8 23 40svIs23[num-1(++)] Vector 0 24 16 0 40svIs23[num-1(++)] pssy-1 8 29 3 0 40svIs23[num-1(++)] pssy-2 1 24 15 0 40svIs23[num-1(++)] F08C6.2 0 24 16 0 40
aThe numbers of vacuoles in forty 1-day-old adult hermaphrodite worms of each genotype were determined. Each animal was placedinto one of the four designated classes depending on the degree of vacuolation: 0, 1–10, 11–50 or >50.bWorms were fed bacteria harboring the plasmid vector L4440 alone. L4440 was the plasmid vector used to express double-strandedRNA from pssy-1, pssy-2 or F08C6.2.
Figure 7: pssy-1(RNAi) suppresses the defects in endocytic
recycling caused by tat-1(kr15) or num-1A(++). Confocalfluorescence micrographs of parts of the intestine of wormsexpressing a marker for recycling the α-chain of the hTAC fusedto GFP (29). The arrows in (A, C and D) indicate apical, basalor lateral plasma membrane of intestinal cells. Note that markeris present on the basal plasma membrane in (A, arrows) butmostly trapped in the cytoplasm in the tat-1 mutant shown in(C, arrowheads). In tat-1(-); pssy-1(RNAi) worms, the marker isfound on the basal plasma membrane. Similarly, pssy-1(RNAi)suppresses the accumulation of the marker in the cytoplasmseen in num-1A(++) worms (arrows) (scale bar, 25 μm).
within a C-terminal inhibitory domain causes displacementof the inhibitory domain from the catalytic domain,which results in activation of the pump (37). Similarly,calcium-bound calmodulin is able to modulate the activityof plasma membrane Ca2+-ATPases by binding to aregulatory domain within the C-terminus (38). At leastone P4 family ATPase, yeast Drsp2p, which is knownto be required for trafficking within the trans-Golginetwork (39), is also regulated through its C-terminal tail:Sla1p, which is required for Drs2p activity, binds to anNPFXD motif in the C-terminal domain (40). This domainalso contains a binding site for phosphatidyl inositol-4-phosphate (PtdIns(4)P), which in turn overlaps a bindingsite for guanine nucleotide exchange factor, Gea2p (41).It has been proposed that the multiple interactions withDrs2p constitute a coincidence detection system usedto activate phospholipid translocation at sites of vesicleformation (41). To our knowledge, no regulatory bindingsites have previously been defined for P4 ATPases frommulticellular eukaryotes. Our results with NUM-1A andTAT-1C indicate that although Numb is not present inyeast, the principle of regulation through the C-terminaltail has been conserved in metazoans.
Work on yeast P4 family ATPases has revealed that theseproteins act in trafficking by promoting vesicle budding andtubulation from membranes from which transport vesiclesare derived (16,17). Earlier studies on TAT-1 in C. elegansstrongly suggest that TAT-1 is required for budding andtubulation from endolysosomal organelles (13,14). In tat-1 mutants, hugely enlarged organelles accumulate thathave endosomal or lysosomal character. Two differentmodels have been proposed for how translocation of PSfrom one leaflet to the other might promote buddingand tubulation. In one model, PS in the cytosolic leaflet
Traffic 2011; 12: 1839–1849 1845

Nilsson et al.
helps recruit proteins that induce the membrane bendingrequired for the formation of incipient buds and tubules.In the other model, the asymmetric distribution of PS isitself proposed to help initiate bending of the membrane.Because the phosphoserine headgroup is considerablybroader than the lipid tail, the presence of a greaterconcentration of PS in one leaflet is suggested to leadto an increase in the length of the cytosolic leaflet relativeto the exoplasmic one. The resulting discrepancy leads tobending of the membrane in such a way that an incipientbud is formed. However, both of these models in theirpure forms are difficult to reconcile with what is knownabout TAT-1 and PS distribution in C. elegans. In wild-type animals, the plasma membrane of many, if not all,cells is composed of bilayers that are apparently highlyasymmetric with respect to their PS composition (24,25).Therefore, asymmetry of PS distribution alone does notseem to be sufficient for membrane bending. On the otherhand, two of the results we report here argue against amodel in which PS in the cytosolic leaflets of organellesinitiates budding solely by recruiting cytoplasmic proteinsto the inner (cytosolic) leaflet of the plasma membraneor the cytosolic leaflet of endolysosomal organelles. First,GFP-LactC2 expressed in the cytoplasm labels the plasmamembrane and the enlarged organelles in tat-1 mutants.Because GFP-LactC2 apparently does not pass throughmembranes (30), this result suggests that in tat-1 mutants,the cytosolic leaflets of both the plasma membraneand of the enlarged organelles still contain PS. Second,reducing pssy-1 function strongly suppresses rather thanenhances tat-1 mutant defects. These observations arebest reconciled by a model in which PS in the outer leafletof the plasma membrane, or in the lumenal (exoplasmic)leaflet of endosomal organelles, inhibits vesicle formation,i.e. an important function of TAT-1 in promoting traffickingis to exclude PS from exoplasmic leaflets. Rigoroustesting of this model in vivo, however, would requiremanipulating and measuring the concentration of PS in theexoplasmic leaflets of endosomal organelles in wild-type,experiments that are presently not technically feasible. Itis noteworthy, however, that in tat-1 mutants, enlargedorganelles accumulate in coelomocytes that appear tocontain PS in their exoplasmic leaflets (14).
Caenorhabditis elegans has six P4 ATPases, TAT-1 to TAT-6 (24,32). The tat-5 promoter is active in apparently alltissues but those of tat-1 to tat-4 all show varying degreesof tissue specificity (32). Our observation that the splicingof tat-1 is different in different tissues demonstrates anadditional layer of complexity in the regulation of P4ATPase expression in C. elegans. It is noteworthy thatmultiple splice forms have also been detected for tat-2,tat-4 and tat-5 (32). To date, however, of the different TATproteins, only those encoded by the tat-1 locus havebeen implicated in the regulation of vesicle-mediatedtrafficking (13,14,32,42). tat-2, tat-3 and tat-4 mutationsdo not cause the intestinal vacuolation seen in tat-1mutants (Figure S2). Nor do mutations in tat-2, tat-3 ortat-4 suppress or enhance the tat-1 intestinal vacuolation
defect (Figure S3). Thus, there is presently no evidencefor redundancy in P4 ATPase function in endolysosomaltrafficking in the intestine. This lack of redundancy standsin contrast to the situation in S. cerevisiae. This organismcontains five P4 family ATPases named Drs2p, Neo1p,Dnf1p, Dnf2p and Dnf3p (8,10). All implicated in transportevents within the endosomal or secretory pathways, butdefects are often seen only when two or more of theproteins are absent (39,43–45).
In previous work, we have shown that loss of num-1Afunction suppresses the recycling and enlarged endosomedefects caused by loss of rme-1 function (5). The presentfindings, together with these earlier observations, mighttherefore suggest a model in which RME-1 acts toregulate recycling by inhibiting NUM-1A activity in theERC. In turn, NUM-1A acts by regulating TAT-1, whichthen regulates the distribution of PS within the membraneof the ERC. While most of our results are consistent withthis linear model, one finding is not: multiple copies of tat-1C potently suppress endosomal defects associated withincreasing num-1A gene dosage, but the effect on the rme-1 phenotype is more complex. This apparent discrepancycould be explained if the effect of removing num-1Afunction when TAT-1C is expressed at normal levels isnot precisely the same as increasing TAT-1C expression.Perhaps when tat-1C gene dosage is increased, increasedlevels of NUM-1A can help to ensure that excess TAT-1C protein does not take on new functions. A secondpossibility is that RME-1 can itself affect the distributionof PS independent of both NUM-1A and TAT-1C. RME-1has been shown to be able to bind liposomes containingPS in vitro and to stimulate tubulation (46). While thistabulating function of RME-1 is likely to be its mainrole, it is possible that, by binding to it, RME-1 can alsoaffect PS distribution. RME-1 might, for example, counterthe action of scramblases that promote the movementof PS from the cytosolic to the exoplasmic leaflets ofbilayers surrounding the ERC. Because the loss of num-1A function suppresses rme-1(lf), for this model to becorrect, it is necessary to invoke the existence of a proteinthat functions to some extent in the absence of RME-1 tostimulate tubulation. Interestingly, even in the absence ofRME-1, AMPH-1 is able to bind to endosomal membranesin vivo and to stimulate the tubulation of liposomescontaining PS in vitro (46). It should be stressed, however,that other models can be envisaged for the regulatoryrelationships between num-1A, rme-1 and tat-1 that arealso consistent with our and others’ findings.
Loss-of-function mutations in num-1A or increasing num-1A dosage affect endocytic recycling, but no evidenceexists presently for a role for NUM-1A within otherendosomal organelles such as sorting endosomes (whererab-10 functions) (5). In contrast, loss of tat-1 functionaffects multiple endosomal compartments (13,14). Onepossibility is that different TAT-1 isoforms function indifferent endosomal organelles. The generation of the
1846 Traffic 2011; 12: 1839–1849

Caenorhabditis elegans NUM-1A Binds to TAT-1
C isoform, which is the only one containing a Numb-binding motif, allows TAT-1 function to be regulatedwithin membranes surrounding the ERC, where NUM-1A is present. It is possible that TAT-1 activity withinthe membranes of other organelles is not regulated.Alternatively, TAT-1 function in other organelles couldbe subject to different forms of regulation, independentof NUM-1A. Either of these models could explain whynum-1A loss-of-function mutations can suppress defectscaused by rme-1 (5) but not those in rab-10 (5), whichaffect sorting endosomes (29).
Materials and Methods
Culturing of C. elegans wormsStandard procedures were used for culturing C. elegans worms (31).All strains were grown at 20◦C unless otherwise stated. The wild-type C. elegans strain Bristol N2 and the following mutant alleleswere used: LG I, rab-10(dx2) (29); LG II, tat-4(tm1801) (32); unc-4(e120) (31); LG III, tat-3(tm1275) (32); tat-1(kr15) (13); LG IV, tat-2(tm1773) (32); LG V, rme-1(b1045) (7); tat-6(ok1984) (32); LG X, lmp-1(nr2045) (33). The transgenes used were pwIs112 (29), svIs23[num-1(+)
unc-4(+)], svIs24[num-1(+) unc- 4(+)], svIs86[Pvha-6::tat-1::gfp(+) rol-6(su1006)], svIs88[Pvha-6::tat-1::gfp rol-6(su1006)], svEx425[Ptat-1::GFP-NLS unc-4(+)] (13), svEx805[Ptat-1::tat-1(exon 15-19)::gfp-NLS unc-4(+)], svIs99[Pvha-6::gfp::Lact-C2] rol-6(su1006)], svIs104[Pnum-1::num-1::mCherry unc-4(+)], svIs107[Pvha-6::tat-1(mut)::gfp rol-6(su1006)] (thisstudy).
Lipid uptake assayTo investigate phospholipid uptake at the apical membrane of intestinalcells, L4 worms were soaked for 2 h at room temperature in 80 μg/mLsolutions in M9 of either 1-oleoyl-2-{6-[(7-nitro-2-1,3-benzoxadiazol-4-yl)amino]hexanoyl}-sn-glycero-3-phosphoserine (NDB-PS) or NDB-PC flu-orescent lipids (Avanti Polar Lipids Inc). For experiments with NBD-PS, theworms were destained for 45 min on normal nematode growth mediumplates with bacteria before observation. NDB-PC was found to degraderapidly inside the worm intestine. Therefore, for experiments with thisdye, the worms were not extensively destained but washed briefly in M9buffer immediately prior to examination.
Analysis of protein–protein interactionsThe Y2H screen was performed by interaction mating. The strains usedwere Y137 and AH109 (BD Biosciences), which are of opposite matingtypes. A 480 bp fragment spanning the entire NUM-1A PTB domain(amino acids 88-245) was fused to sequences encoding the Gal4 DNA-binding domain (BD) by subcloning the fragment into the yeast expressionvector, pGBK-T7 (BD Biosciences). The resulting plasmid, pVB362LN, wastransformed into AH109. A C. elegans cDNA library in the yeast expressionvector pACT (a kind gift from Z. Zhou and H. R. Horvitz) was transformedinto Y137. Yeast clones from the library were mated to the AH109-derivedstrain expressing the GAL4 DNA BD-NUM-1A PTB fusion protein, andthe products of the mating were plated onto medium lacking Trp, Leu,His and Ade to select for putative interaction partners. Positive cloneswere retested in an assay in which the ability to induce expression ofβ-galactosidase was measured. Binding to the PTB domain of DAB-1 wasalso determined. To test for interaction in vitro, equal amounts of thebacterially expressed fusion proteins GST::TAT-1C(WT), GST::TAT-1C(N->A) or GST protein alone were immobilized on magnetic GST particles(Promega, WI), washed and thereafter incubated with equal amounts of35S-methionine-labeled NUM-1A PTB domain (Promega, WI), generatedby in vitro translation using the Y2H-based vector pVB362LN as a template.The bound proteins were subjected to SDS-PAGE, and coprecipitated PTBdomain protein was detected by autoradiography.
DNA constructs
Y2H constructspVB362LN (NUM-1::BD) – Using the Est-clone yk565f1 as a template, aDNA fragment encoding the entire PTB domain of NUM-1A was amplifiedusing primers flanked by restriction enzyme sites and inserted betweenthe EcoRI and BamHI sites of Y2H vector pGBK-T7 (BD Biosciences).The resulting plasmid encodes the PTB domain of NUM-1A fused withthe DNA-BD of Gal4p. The primers used were 5′- ga gaa ttc ggt tcg aagact gaa cag-3′(numPTB.1-Eco) and 5′- ga gga tcc gat tact tic gct gat tgc-3′(numPTB.2-Bam) (the restriction sites added to the primers are underlined).
pVB368LN (DAB-1::BD) – A DNA fragment encoding the entire PTBdomain of DAB-1 was amplified using primer pairs flanked by restrictionenzyme sites and inserted into the EcoRI site of pGBK-T7. The primersused were
5′-ga gaa ttc tcg gag aac aag atg tgg aca- 3′ (dab1.F) and5′-gaga att cca agt cgt gaa gat cag- 3′ (dab1.R).
pVB372LN (TAT-1C::AD) – A ClaI fragment from the tat-1C EST cloneyk1228h04 (generously provided by Y. Kohara) was inserted into the ClaIsite of the Y2H vector pGAD-T7 (BD Biosciences). The resulting plasmidencodes the very C-terminal end of TAT-1C fused with the activationdomain of Gal4.
pVB373LN (TAT-1::AD) – A DNA fragment encoding the last 30 aminoacids of the TAT-1C isoform, including the NXXF motif, was amplified usingprimer pairs flanked by EcoRI sites and inserted into pGAD-T7. pVB375LNand pVB376LN contain mutant versions of this fragment generated by theuse of mutant upstream primers. The upstream, forward primers usedwere:
5′- gag gaa ttc cgg agc tcg tac acg aat cgc gcg ttc ata -3′ (tat-WT),5′- gag gaa ttc cgg agc tcg tac acg gct cgc gcg ttc ata -3′ (tat-N-A) and5′- gag gaa ttc cgg agc tcg tac acg aat cgc gcg gcc ata -3′ (tat-F-A);
the downstream, reverse primer used for all three fragments was:
5′- gag gaa ttc tca taa cct cgt ccc act acc atc-3′ (tat-EcoRev).
tat-1 expression constructspVB320LN (Pvha-6::tat-1::gfp) has been described previously (13).
pVB496LN (Ptat-1::tat-1(exon 15-19)::gfp-NLS) – The same 2.5 kb pro-moter fragment as that used in pVB320LN was fused with a 5.1 kbgenomic fragment spanning exons 15–19 of the tat-1 gene. The 2.5 kbpromoter was subcloned from pVB320LN into pPD49.26 using HindIII andBamHI. Thereafter, a 5.1 kb, polymerase chain reaction (PCR)-amplifiedgenomic fragment was inserted into the NheI site. In this manner, theopen-reading frame within exon 15 was fused in frame with the start ATGwithin exon 1a present in the promoter fragment. A DNA fragment encod-ing nuclear-localized GFP was amplified from pPD121.83 and inserted inframe at a SacI site within exon 17. The primers used to amplify exons15–19 were as follows:
5′-ggc gct agc cca cgt gaa ctt gct gta atg -3′(exon15.Nhe.F) and5′-gcc gct agc tta tcg tcc agt cgg ttt ttc tct agt-3′ (tat-1R.Nhe).
pVB378LN (Pvha-6::tat-1(mut)::gfp) – To generate a construct to express amutant form of TAT-1C, with a mutated NUM-1A-binding site, we used theplasmid pVB349LN (13) (which contains a wild-type tat-1C cDNA drivenby a vha-6 promoter fragment) as a template in overlap extension PCR.In this reaction, we used internal primers containing a nucleotide change
Traffic 2011; 12: 1839–1849 1847

Nilsson et al.
in the codon encoding the asparagine residue within the NRAF NUM-1A-interaction sequence. The sequence change results in an N to A amino acidsubstitution. The wild-type tat-1C fragment in pVB349LN was thereafterreplaced with the mutant version using flanking KpnI sites.
Constructs for protein expressionTo generate wild-type and mutant TAT-1C::GST fusion protein, wesubcloned XmaI to XhoI tat-1C fragments from pVB372LN and pVB388LN,respectively, into the vector pGex5x-1 (GE Healthcare).
RNAi constructsPlasmid constructs for the genes pssy-1 and pssy-2 for use in RNAi feedingexperiments were generated by inserting 1 kb cDNA fragments from eachof these two genes into the RNAi feeding vector L4440 (generouslyprovided by A. Fire) digested with EcoRI. The two cDNA fragments weregenerated by the one-step reverse transcription-polymerase chain reaction(RT-PCR) system (Invitrogen) from total RNA prepared from N2 wormsusing the gene-specific primers:
• 5′- gag gaa ttc atg gcc gac gac gtc tta ggg-3′ (pssy-1.F)• 5′- ctc gaa ttc cag ctc ggt tag caa cca aat -3′ (pssy-1.R)• 5′- gag gaa ttc tca ttc tcg gcg cac tcg gat-3′ (pssy-2.F)• 5′- ctc gaa ttc gtt gta cca atc gta tcc gc-3′ (pssy-2.R)
Lactadherin constructA fragment encoding the C2 domain of Bovine Lactadherin was amplifiedusing the plasmid GFP-Lact-C2 (Haematologic Technologies Inc) as atemplate. This fragment was fused at its 5′ end with sequences encodingGFP, using overlap extension PCR, and thereafter inserted into the KpnIsite of pVB343ML, a vector harboring the intestinal-specific promoterPvha-6(13). The upstream primer to amplify the C2 domain of lactadherinalso contained sequences corresponding to the 3′ end of gfp to facilitatethe subsequent fusion. The sequence of the upstream primer was 5′- atggat gaa cta ata caa atg cac tga acc cct agg cct g-3′(gfp-lactC2.F) and thatof the downstream primer was 5′- ctc ggt acc cta aca gcc cag cag ctccac-5′(lactC2.Kpn.R).
MicroscopyAnimals were mounted on 1 × M9, 2% agarose pads and examinedunder epiflourescence and/or differential interference contrast optics usingDMRB compound microscope (Leica) or by confocal microscopy using aLeica TCS SP2 AOBS confocal microscope. The worms were mounted ina drop of M9 solution containing 2.0 mM levimasole to inhibit movement.
Acknowledgments
We thank A. Ronnlund for excellent technical assistance and C. Anderssonfor help with experiments on pssy-1 and pssy-2. We are grateful tothe Caenorhabditis Genetics Center (which is funded by the NIH NationalCenter for Research Resources) for strains, to A. Fire for plasmids and to Y.Kohara for cDNAs. Ola Billing and Gautam Kao are gratefully acknowledgedfor comments on the manuscript. The work was supported by grants fromCancerfonden (09 0507) and Vetenskapsradet (K2009-67X-20441-03-3;to S. T.)
Supporting Information
Additional Supporting Information may be found in the online version ofthis article:
Figure S1: Multiple copies of tat-1 do not reduce NUM-1A expression,
and multiple copies of num-1A do not reduce TAT-1C expression.
Confocal fluorescence micrographs of parts of the intestines of wormswith num-1A(++) or tat-1C transgenes. svIs86 and svIs88 harbor multiple
copies of a transgene encoding wild-type TAT-1C fused to GFP. svIs104harbors multiple copies of a transgene encoding NUM-1A::mCherry.svIs107 harbors multiple copies of a transgene encoding mutant TAT-1C fused to GFP. Note that multiple copies of tat-1C do not cause areduction in the expression of NUM-1A::mCherry or vice versa. Notably,TAT-1C::GFP intestinal expression is frequently mosaic with certain cellshaving much higher or lower levels of expression [indicated by an arrowand an arrowhead, respectively, in (E)] than the average. However, highlevels of TAT-1C::GFP expression is not associated with lower levels ofNUM-1A::mCherry expression, and a low level of TAT-1C expression is notassociated with higher levels of NUM-1A::mCherry (scale bar, 25 μm).
Figure S2: The effect of multiple copies of tat-1C on phenotypes
caused by rme-1 or rab-10 loss of function. Micrographs of parts ofthe intestines of L4 hermaphrodites viewed with differential interferencecontrast optics. Note that the intestine of the animals depicted in (D) and(E) contain vacuoles that, unlike those in rme-1 single mutants, are filledwith refractile material (scale bar, 25 μm).
Figure S3: tat-2, tat-3, tat-4 or tat-6 single mutants do not show
the intestinal vacuolation seen in tat-1 mutants. Micrographs ofparts of the intestines of L4 hermaphrodites viewed with differentialinterference contrast optics. The arrows in B indicate vacuoles in thetat-1 single mutant. The mutants depicted are tat-1(kr15), tat-2(tm1773),tat-3(tm1275), tat-4(tm1801) and tat-6(ok1984) (scale bar, 25 μm).
Figure S4: tat-2, tat-3 or tat-4 mutations do not enhance or
suppress the intestinal vacoulation caused by loss of tat-1 function.
Micrographs of parts of the intestines of L4 hermaphrodites viewed withdifferential interference contrast optics. The genotypes were tat-1(kr15),tat-1(kr15); tat-2(tm1773), tat-3(tm1275) tat-1(kr15) and tat-1(kr15); tat-4(tm1801 scale bar, 25 μm).
Please note: Wiley-Blackwell are not responsible for the content orfunctionality of any supporting materials supplied by the authors.Any queries (other than missing material) should be directed to thecorresponding author for the article.
References
1. Gulino A, Di Marcotullio L, Screpanti I. The multiple functions ofNumb. Exp Cell Res 2009;316:900–906.
2. Salcini AE, Confalonieri S, Doria M, Santolini E, Tassi E, Minenkova O,Cesareni G, Pelicci PG, Di Fiore PP. Binding specificity and in vivotargets of the EH domain, a novel protein-protein interaction module.Genes Dev 1997;11:2239–2249.
3. Berdnik D, Torok T, Gonzalez-Gaitan M, Knoblich JA. The endocyticprotein alpha-Adaptin is required for numb-mediated asymmetric celldivision in Drosophila. Dev Cell 2002;3:221–231.
4. Smith CA, Dho SE, Donaldson J, Tepass U, McGlade CJ. The cell fatedeterminant numb interacts with EHD/Rme-1 family proteins and hasa role in endocytic recycling. Mol Biol Cell 2004;15:3698–3708.
5. Nilsson L, Conradt B, Ruaud AF, Chen CC, Hatzold J, Bessereau JL,Grant BD, Tuck S. Caenorhabditis elegans num-1 negatively regulatesendocytic recycling. Genetics 2008;179:375–387.
6. Tang H, Rompani SB, Atkins JB, Zhou Y, Osterwalder T, Zhong W.Numb proteins specify asymmetric cell fates via an endocytosis-and proteasome-independent pathway. Mol Cell Biol 2005;25:2899–2909.
7. Grant B, Zhang Y, Paupard MC, Lin SX, Hall DH, Hirsh D. Evidencethat RME-1, a conserved C. elegans EH-domain protein, functions inendocytic recycling. Nat Cell Biol 2001;3:573–579.
8. Graham TR. Flippases and vesicle-mediated protein transport. TrendsCell Biol 2004;14:670–677.
9. Kuhlbrandt W. Biology, structure and mechanism of P-type ATPases.Nat Rev 2004;5:282–295.
10. Pomorski T, Holthuis JC, Herrmann A, van Meer G. Tracking down lipidflippases and their biological functions. J Cell Sci 2004;117:805–813.
11. Gilbert MJ, Thornton CR, Wakley GE, Talbot NJ. A P-type ATPaserequired for rice blast disease and induction of host resistance. Nature2006;440:535–539.
1848 Traffic 2011; 12: 1839–1849

Caenorhabditis elegans NUM-1A Binds to TAT-1
12. Poulsen LR, Lopez-Marques RL, McDowell SC, Okkeri J, Licht D,Schulz A, Pomorski T, Harper JF, Palmgren MG. The Arabidopsis P4-ATPase ALA3 localizes to the golgi and requires a beta-subunit tofunction in lipid translocation and secretory vesicle formation. PlantCell 2008;20:658–676.
13. Ruaud AF, Nilsson L, Richard F, Larsen MK, Bessereau JL, Tuck S.The C. elegans P4-ATPase TAT-1 regulates lysosome biogenesis andendocytosis. Traffic 2009;10:88–100.
14. Chen B, Jiang Y, Zeng S, Yan J, Li X, Zhang Y, Zou W, Wang X.Endocytic sorting and recycling require membrane phosphatidylserineasymmetry maintained by TAT-1/CHAT-1. PLoS Genet 2010;6:e1001235.
15. Xu P, Okkeri J, Hanisch S, Hu RY, Xu Q, Pomorski TG, Ding XY.Identification of a novel mouse P4-ATPase family member highlyexpressed during spermatogenesis. J Cell Sci 2009;122:2866–2876.
16. Graham TR, Kozlov MM. Interplay of proteins and lipids in generatingmembrane curvature. Curr Opin Cell Biol 2010;22:430–436.
17. Lopez-Marques RL, Poulsen LR, Hanisch S, Meffert K, Buch-PedersenMJ, Jakobsen MK, Pomorski TG, Palmgren MG. Intracellular targetingsignals and lipid specificity determinants of the ALA/ALIS P4-ATPasecomplex reside in the catalytic ALA alpha-subunit. Mol Biol Cell2010;21:791–801.
18. Coleman JA, Kwok MC, Molday RS. Localization, purification, andfunctional reconstitution of the P4-ATPase Atp8a2, a phosphatidylser-ine flippase in photoreceptor disc membranes. J Biol Chem2009;284:32670–32679.
19. Zhou X, Graham TR. Reconstitution of phospholipid translocaseactivity with purified Drs2p, a type-IV P-type ATPase from buddingyeast. Proc Natl Acad Sci U S A 2009;106:16586–16591.
20. Auland ME, Roufogalis BD, Devaux PF, Zachowski A. Reconstitutionof ATP-dependent aminophospholipid translocation in proteolipo-somes. Proc Natl Acad Sci U S A 1994;91:10938–10942.
21. Tang X, Halleck MS, Schlegel RA, Williamson P. A subfamily of P-type ATPases with aminophospholipid transporting activity. Science1996;272:1495–1497.
22. Natarajan P, Wang J, Hua Z, Graham TR. Drs2p-coupled aminophos-pholipid translocase activity in yeast Golgi membranes and relationshipto in vivo function. Proc Natl Acad Sci U S A 2004;101:10614–10619.
23. Alder-Baerens N, Lisman Q, Luong L, Pomorski T, Holthuis JC. Lossof P4 ATPases Drs2p and Dnf3p disrupts aminophospholipid transportand asymmetry in yeast post-Golgi secretory vesicles. Mol Biol Cell2006;17:1632–1642.
24. Zullig S, Neukomm LJ, Jovanovic M, Charette SJ, Lyssenko NN,Halleck MS, Reutelingsperger CP, Schlegel RA, Hengartner MO.Aminophospholipid translocase TAT-1 promotes phosphatidylserineexposure during C. elegans apoptosis. Curr Biol 2007;17:994–999.
25. Darland-Ransom M, Wang X, Sun CL, Mapes J, Gengyo-Ando K,Mitani S, Xue D. Role of C. elegans TAT-1 protein in maintainingplasma membrane phosphatidylserine asymmetry. Science 2008;320:528–531.
26. Kamikura DM, Cooper JA. Clathrin interaction and subcellular localiza-tion of Ce-DAB-1, an adaptor for protein secretion in Caenorhabditiselegans. Traffic 2006;7:324–336.
27. Holmes A, Flett A, Coudreuse D, Korswagen HC, Pettitt J. C. elegansdisabled is required for cell-type specific endocytosis and isessential in animals lacking the AP-3 adaptor complex. J Cell Sci2007;120:2741–2751.
28. Uhlik MT, Temple B, Bencharit S, Kimple AJ, Siderovski DP, John-son GL. Structural and evolutionary division of phosphotyrosine bind-ing (PTB) domains. J Mol Biol 2005;345:1–20.
29. Chen CC, Schweinsberg PJ, Vashist S, Mareiniss DP, Lambie EJ,Grant BD. RAB-10 is required for endocytic recycling in theCaenorhabditis elegans intestine. Mol Biol Cell 2006;17:1286–1297.
30. Yeung T, Gilbert GE, Shi J, Silvius J, Kapus A, Grinstein S. Membranephosphatidylserine regulates surface charge and protein localization.Science 2008;319:210–213.
31. Brenner S. The genetics of Caenorhabditis elegans. Genetics1974;77:71–94.
32. Lyssenko NN, Miteva Y, Gilroy S, Hanna-Rose W, Schlegel RA. Anunexpectedly high degree of specialization and a widespreadinvolvement in sterol metabolism among the C. elegans putativeaminophospholipid translocases. BMC Dev Biol 2008;8:96.
33. Kostich M, Fire A, Fambrough DM. Identification and molecular-genetic characterization of a LAMP/CD68-like protein from Caenorhab-ditis elegans. J Cell Sci 2000;113:2595–2606.
34. Brodsky FM, Chen CY, Knuehl C, Towler MC, Wakeham DE. Biologi-cal basket weaving: formation and function of clathrin-coated vesicles.Annu Rev Cell Dev Biol 2001;17:517–568.
35. Polo S, Di Fiore PP. Endocytosis conducts the cell signaling orchestra.Cell 2006;124:897–900.
36. Maxfield FR, McGraw TE. Endocytic recycling. Nat Rev 2004;5:121–132.
37. Duby G, Boutry M. The plant plasma membrane proton pump ATPase:a highly regulated P-type ATPase with multiple physiological roles.Pflugers Arch 2009;457:645–655.
38. Di Leva F, Domi T, Fedrizzi L, Lim D, Carafoli E. The plasmamembrane Ca2+ ATPase of animal cells: structure, function andregulation. Arch Biochem Biophys 2008;476:65–74.
39. Gall WE, Geething NC, Hua Z, Ingram MF, Liu K, Chen SI, Graham TR.Drs2p-dependent formation of exocytic clathrin-coated vesiclesin vivo. Curr Biol 2002;12:1623–1627.
40. Liu K, Hua Z, Nepute JA, Graham TR. Yeast P4-ATPases Drs2p andDnf1p are essential cargos of the NPFXD/Sla1p endocytic pathway.Mol Biol Cell 2007;18:487–500.
41. Natarajan P, Liu K, Patil DV, Sciorra VA, Jackson CL, Graham TR.Regulation of a Golgi flippase by phosphoinositides and an ArfGEF.Nat Cell Biol 2009;11:1421–1426.
42. Seamen E, Blanchette JM, Han M. P-type ATPase TAT-2 negativelyregulates monomethyl branched-chain fatty acid mediated function inpost-embryonic growth and development in C. elegans. PLoS Genet2009;5:e1000589.
43. Chen CY, Ingram MF, Rosal PH, Graham TR. Role for Drs2p, a P-typeATPase and potential aminophospholipid translocase, in yeast lateGolgi function. J Cell Biol 1999;147:1223–1236.
44. Hua Z, Fatheddin P, Graham TR. An essential subfamily of Drs2p-related P-type ATPases is required for protein trafficking betweenGolgi complex and endosomal/vacuolar system. Mol Biol Cell2002;13:3162–3177.
45. Pomorski T, Lombardi R, Riezman H, Devaux PF, van Meer G,Holthuis JC. Drs2p-related P-type ATPases Dnf1p and Dnf2p arerequired for phospholipid translocation across the yeast plasmamembrane and serve a role in endocytosis. Mol Biol Cell2003;14:1240–1254.
46. Pant S, Sharma M, Patel K, Caplan S, Carr CM, Grant BD. AMPH-1/Amphiphysin/Bin1 functions with RME-1/Ehd1 in endocytic recy-cling. Nat Cell Biol 2009;11:1399–1410.
Traffic 2011; 12: 1839–1849 1849





























