Embed Size (px)
Citation preview

Ca2+ and NSF differentially regulate disassembly of SNARE complexes on early
endosomes.
Qing Yan1, Wei Sun1, James A. McNew2, Thomas A. Vida3, and Andrew J. Bean1,4
1Department of Neurobiology and Anatomy, University of Texas Medical School,
Houston, TX 77030 USA, 2Department of Biochemistry and Cell Biology, Rice
University, Houston, TX 77005, 3Department of Microbiology And Molecular Genetics,
University of Texas Medical School, Houston, TX 77030
4To whom correspondence should be addressed
Andrew Bean
UTHSC Dept. NBA
6431 Fannin Street MSB 7.208
Houston, TX 77030 USA
Ph: 713-500-5614
Email: [email protected]
Running Title: Regulation of Endosomal SNARE Complexes
Key Words: hrs, SNAP-25, syntaxin, VAMP, SNARE
Abbreviations used in this paper: Hrs, hepatocyte responsive serum phosphoprotein; SNAP-25, synaptosomal associated protein of 25kDa; VAMP, vesicle associated membrane protein; NSF, N-ethylmalemide sensitive factor; SNARE, soluble N-ethylmaleimide-sensitive factor attachment protein receptor; UIM, ubiquitin interacting motif; GST, glutathione-S-transferase; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; PBS, phosphate buffered saline; EEA-1, early endosomal antigen-1; TeTx, tetanus toxin; PI(3)P, phosphatidylinositol 3-phosphate; DMEM, Dulbecco’s minimal essential medium.
JBC Papers in Press. Published on February 9, 2004 as Manuscript M400093200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Summary
The endosome-associated protein hrs inhibits the homotypic fusion of early
endosomes. A helical region of hrs containing a Q-SNARE motif mediates this effect as
well as its endosomal membrane association via SNAP-25, an endosomal receptor for
hrs. Hrs inhibits formation of an early endosomal SNARE complex by displacing
VAMP-2 from the complex, suggesting a mechanism by which hrs inhibits early
endosome fusion. We examined the regulation of endosomal SNARE complexes to
probe how hrs may function as a negative-regulator. We show that while NSF
dissociates the VAMP-2/SNAP-25/syntaxin 13 complex, it has no effect on the hrs-
containing complex. Whereas Ca2+ dissociates the hrs-containing, but not VAMP-2-
containing SNARE complex. This is the first demonstration of differential regulation of
R/Q-SNARE and all Q-SNARE containing SNARE complexes. Ca2+ also reverses the
hrs-induced inhibition of early endosome fusion in a tetanus toxin-sensitive manner and
removes hrs from early endosomal membranes. Moreover, hrs inhibition of endosome
fusion and its endosomal localization are sensitive to bafilomycin implying a role for
luminal Ca2+. Thus, hrs may bind a SNARE protein on early endosomal membranes
negatively regulating trans-SNARE pairing and endosomal fusion. The release of Ca2+
from the endosome lumen dissociates hrs, allowing a VAMP-2-containing complex to
form enabling fusion.
2
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Introduction
Endocytosis is a fundamental process essential for all eukaryotic cells. It functions
in nutrient uptake, regulation of the protein and lipid composition in the plasma
membrane, and modulation of cellular responses by affecting exocytosis and receptor
signaling. Molecules transit through the endocytic pathway by passing from one
compartment to another through a series of membrane fission and fusion reactions. The
ultimate role of this pathway is to allow sorting of molecules to be recycled from those to
be degraded. To function correctly, this system must regulate both the sorting events and
the fusion events that promote proper targeting of internalized small molecules, lipids,
and proteins.
Protein machinery is required for fusion of biological membranes. Interactions
among SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor)
proteins(1-4) associated with donor membranes (e.g.,VAMP/synaptobrevin) and acceptor
membranes (e.g., syntaxin and SNAP-25) are thought to be essential for fusion(1-5).
SNAREs are sufficient for membrane fusion in artificial membranes suggesting that they
form the core membrane fusion machinery(6). SNAREs form cytoplasmic helical bundles
that bridge two membranes (trans-SNARE complex) to enable membrane fusion(1,4,7).
SNAREs are characterized by a helical “SNARE” motif that contains a glutamine (Q) or
arginine (R ) at its center(4,8). Botulinum and tetanus toxins are zinc endoproteases that
cleave the SNAREs, inhibit the formation of SNARE complexes, and block fusion(1,2,9).
Once membrane fusion has occurred, the cytoplasmic adapter protein α-SNAP binds to
the SNARE complex and recruits NSF (N-ethylmalemide sensitive factor) from the
cytoplasm(1,2,7). The ATPase activity of NSF dissociates the cis-SNARE complex
3
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

allowing the proteins to be available for trans-pairing and subsequent fusion
events(1,2,7).
The trans-SNARE complex may be the catalyst for membrane fusion although
regulatory events or molecules may influence this process. For example, intraorganellar
Ca2+ release from the yeast vacuole, mammalian endosome, or nuclear vesicles is
required for fusion events involving those compartments(10-13). A model that has
emerged from these studies is that an unknown event triggers Ca2+ release from the
organelle. This local pool of Ca2+ is required for the fusion event although the nature of
its effector is unclear. Evidence from studies of homotypic vacuole fusion suggest that
calmodulin may be a Ca2+ target as it binds to vacuoles upon Ca2+ release and appears to
promote bilayer mixing of vacuoles and endosomes(11-13) although the effect of
calmodulin may be via regulation of SNARE complexes(14,15).
The endosome-associated protein hrs has been shown to bind the Q-SNARE
SNAP-25 and its homologs SNAP-23 and SNAP-29(16), and inhibits the homotypic
fusion of early endosomes(17). A heptad-repeat region of hrs containing a Q-SNARE
motif mediates this effect as well as its endosomal membrane association(17). SNAP-25
is an endosomal receptor for hrs, and hrs inhibits the formation of an early endosomal
SNARE complex by disallowing VAMP incorporation into the complex(17). Hrs is also
likely involved in cargo sorting at the level of the early/late endosome by recruiting
sorting or signaling components to the endosomal membrane. Therefore, Hrs may bind
to SNAP-25 using its Q-SNARE domain and inhibit endosomal fusion while it is
involved in cargo sorting or endosome motility using NH2-terminal VHS (Vps27, Hrs,
STAM, a domain found in a number of proteins involved in trafficking that, in some
4
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

cases, binds to GGA proteins), FYVE (Fab 1, YotB, Vac1, EEA-1, a dual zinc finger
domain found in a number of proteins involved in trafficking some of which bind to
PI3P, phosphatidylinositol 3-phosphate) or UIM (Ubiquitin Interacting Motif, a domain
found in a number of proteins that binds ubiquitin with low affinity) domains (17 and
references therein) or via other protein interactions. Interestingly, the binding of hrs to
SNAP-25 is negatively regulated by Ca2+ such that Ca2+inhbits the binding of hrs and
SNAP-25(18,19). In the present study, we have examined the regulation of hrs- and
VAMP-2-containing endosomal SNARE complexes and find that while NSF regulates
the dissociation of the VAMP-2/SNAP-25/syntaxin 13 complex, it has no effect on the
hrs-containing complex. Conversely, Ca2+ dissociates the hrs-containing, but not VAMP-
2-containing SNARE complex. Ca2+ removes hrs from early endosomal membranes at
concentrations similar to those that dissociate the hrs-containing SNARE complex.
Similar Ca2+ concentrations reverse the hrs-induced inhibition of early endosome fusion
in a tetanus toxin-sensitive manner. Moreover, the sensitivity of hrs inhibition of
endosome fusion to bafilomycin implies a role for luminal Ca2+ release in the dissociation
of hrs from SNAP-25/early endosomal membranes. These data suggest that hrs provides
a Ca2+-dependent negative influence on early endosome fusion and that a VAMP-2-
containing SNARE complex can form to enable fusion after the dissociation of hrs.
5
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Materials and Methods
Materials.
Hrs was expressed in insect cells as previously described(16). Syntaxin 13(20)
and VAMP-2 were expressed in E. coli as previously described(17,20-23). The light
chain of Tetanus toxin (a kind gift from Dr. Heiner Niemann) was expressed in E. coli as
described(9,24). Antibodies were obtained from the following sources: hrs (Alexsis
Biochemicals, San Diego, CA), EEA1 (Affinity BioReagents, Golden, CO), syntaxin 13
(a kind gift of Dr. Rytis Prekeris), VAMP-2 (Synaptic Systems, Germany), 6XHIS
(Sigma Chemicals St Louis, MO), SNAP-25 (Sternberger Monoclonals, Inc., Lutherville,
MD).
Early endosome fusion
The homotypic fusion of early endosomes was measured as previously
described(17). Effects of various molecules were assessed after incubation on ice for 15
min with the donor and acceptor membranes, cytosol, and ATP. The reactions were then
transferred to 37oC for 60 min. Free Ca2+ concentrations (0, 0.003, 0.03, 0.1, 0.3, 1, 10
and 30 mM) and 1 mM of Cu2+, Ba2+, and Mn2+ were calculated in the presence of 2mM
EGTA using the WebMaxCalc program
(http://www.stanford.edu/~cpatton/webmaxcS.htm)(18). TeTx treatment was performed
by isolating the early endosomes as described above and resuspending the vesicles with
varying concentrations of TeTx (5 to 6400 nM in a final volume of 30 µl)(9,24) followed
by incubation at 37oC for 30 or 60 min. Membranes were collected by centrifugation
(100,000 xg for 10 min) and resuspended in 100µl homogenization buffer for use in the
fusion reactions. For bafilomycin treatment, cells were incubated with DMEM
6
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

containing 1% BSA and 10mM Ca2+ for 60 min. Subsequently, cells were labeled with
fluorescent EGF (15 min at 37oC) in the presence of bafilomycin (100 nM). After
labeling, the cells were washed homogenized, and endosomes were isolated in the
presence of bafilomycin. Endosomes were incubated with hrs (180nM) in the presence
of bafilomycin (15min on ice). Bafilomycin was removed from some membranes by
dilution with 10 volumes of reaction buffer and all membranes were subjected to
centrifugation followed by resuspension of endosomes in reaction buffer. Reactions were
incubated (37oC for 60min) in the presence (hrs+bafilomycin) or absence (hrs-
bafilomycin) of bafilomycin.
SNARE complex disassembly
A constant amount of GST-syntaxin 13 (2 µg/reaction) bound to glutathione
agarose was incubated with constant amounts of SNAP 25 (1 µg), VAMP-2 (1 µg) or Hrs
(2 µg) in the presence or absence of NSF/αSNAP (0.25 mg/ml), MgATP or ATPγS (0.5
mM) or EDTA in PBS binding buffer to a final reaction volume of 50µl. In addition, the
hrs-containing complex was incubated in 50 µM free Ca2+. After incubation (4 °C for 60
min), samples were washed three times with binding buffer, boiled in SDS sample buffer,
and separated by SDS-PAGE. Immunoblot analysis was performed using anti-hrs and
anti-VAMP-2 antibodies followed by appropriate 125I-secondary antibodies. Parallel
experiments in which hrs- or VAMP-2-containing SNARE complexes were preformed
prior to incubation with NSF/αSNAP or Ca2+ yielded similar results (Figure 1B).
Immunoreactive bands were visualized and quantitated using phosphorimaging. The
experiment shown is representative of six experiments.
7
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Endosomal membrane binding
Endosomal membranes were purified from HeLa cells by centrifugation on a
discontinuous sucrose gradient(17). One 10 cm plate (approx 80% confluent) was
scraped in homogenization buffer (20 mM HEPES 7.4, 0.25 M sucrose, 2 mM EGTA, 2
mM EDTA, 0.1 mM DTT, 0.4 ml) and passed through 30 G needle 30 times. The
resulting lysate was subjected to centrifugation (100,000 xg 10 min) and the pellet was
resuspended in homogenization buffer (0.17 ml) then mixed with 61% sucrose to a final
concentration of 46% sucrose (0.5 ml total). The 46% sucrose cushion was overlaid with
two additional layers of sucrose 35% (0.65 ml) and 30% (0.45 ml) and then with
homogenization buffer (0.4 ml). The gradients were subjected to centrifugation in a
Beckman TLS55 rotor (124,000 xg for 60 min) and the early endosomes (interface
between 30-35% sucrose) were collected. A constant amount of HIS-tagged hrs (180
nM) was added to reactions containing purified endosomal membranes (as in a fusion
reaction), along with various concentration of free Ca2+ (0, 0.003, 0.03, 0.1, 0.3, 1, 3, 10
and 30 mM) and 1 mM of Cu2+, Ba2+ and Mn2+. Reactions were incubated at 37oC for
60 min and stopped by subjecting them to centrifugation (100,000 xg for 10 min). The
amount of HIS-hrs was determined in pellet and supernatant fractions by quantitative
western blotting using anti-HIS antibodies and 125I-conjugated secondary antibodies. The
experiment shown is representative of eight experiments.
8
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Results
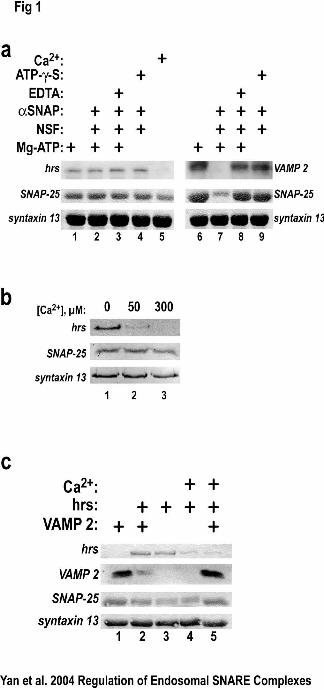
Ca2+ but not NSF dissociates an hrs-containing SNARE complex
Since Ca2+ inhibits hrs from binding to SNAP-25(18,19), we tested the possibility
that Ca2+ might prevent the formation of the recently identified SNARE complex
containing hrs, syntaxin 13, and SNAP-25 (17). In the presence of free Ca2+ (50 µM), <
10% of hrs bound to a complex of SNAP-25 and syntaxin 13 (Fig. 1A, lane 5) compared
to the binding in the absence of Ca2+ (Figure 1A, lanes 1-4). Even in the presence of
αSNAP and NSF, two proteins known to disassemble SNARE complexes, hrs was
incorporated into a complex with SNAP-25 and syntaxin 13 (Fig. 1A, lane 2). In contrast,
NSF and αSNAP dissociated the VAMP-2-containing SNARE complex (Fig. 1A, lane
7). The effect of NSF and αSNAP required Mg-ATP since the VAMP-2-containing
complex did not dissociate when EDTA was present (Fig. 1, lane 8). Moreover, the
ATPase activity of NSF was required for SNARE complex dissociation because the
complex did not dissociate in the presence of the non-hydrolyzable ATP analog, ATP-γ-
S, (Fig. 1, lane 9). Parallel experiments in which hrs- or VAMP-2 SNARE complexes
were preformed prior to incubation with Ca2+ or NSF/αSNAP yielded similar results
(Figure 1B and data not shown). The half maximal concentration of Ca2+ required to
inhibit hrs binding to the SNAP-25/syntaxin 13 complex was approximately 20µM (Fig
1B). These results suggested that Ca2+ could dissociate a SNARE complex containing
hrs, SNAP-25 and syntaxin while NSF and α-SNAP have no effect on the formation or
dissociation of this complex.
9
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Ca2+ dissociates an hrs-containing SNARE complex allowing VAMP-2 binding.
The effect of Ca2+ dissociation of hrs on the ability of VAMP-2 to enter a SNARE
complex was tested. When VAMP-2 and hrs alone were added to binding reactions with
SNAP-25 and syntaxin 13, they were both able to form SNARE complexes (Fig. 1C,
lanes 1 and 3, respectively). However, when VAMP-2 and hrs were added together,
>80% of VAMP-2 was not bound (Fig. 1C, lane 2), consistent with our previous
studies(23). Free calcium inclusion resulted in a loss of hrs binding (Fig. 1C, lane 4).
However, when VAMP-2 was included with hrs and Ca2+, VAMP-2 binding was
comparable to control (Fig. 1C, lane 5 compared to lane 1). This result demonstrates that
Ca2+ has no effect on formation/dissociation of a VAMP-2/SNAP-25/syntaxin 13
SNARE complex, and that in the presence of Ca2+ VAMP-2 is able to displace hrs for
SNAP-25/syntaxin 13 binding.
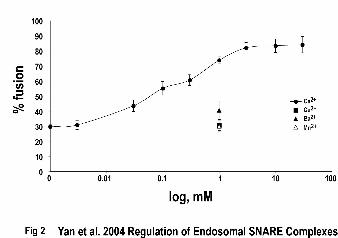
Ca2+ relieves the hrs-dependent inhibition of early endosome fusion
The ability of Ca2+ to displace hrs from the SNARE complex and allow complex
assembly with VAMP-2, suggests that calcium might reverse the inhibition of early
endosome fusion produced by hrs. We previously developed an assay for endosome
fusion in which different populations of HeLa cells engage in receptor-mediated
endocytosis of EGF linked to either Alexa 488 or tetramethylrhodamine (TMR), allowing
isolation of donor and acceptor pools of endosomes. These compartments are used in
fusion reactions that are analyzed by examining resonance energy transfer between the
fluorophores to detect content mixing. This assay is dependent on temperature, time,
energy, and cytosol(17). Fusion reactions containing 180 nM hrs were inhibited
10
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

approximately 70% compared to control reactions (Fig. 2). However, when Ca2+ was
added to these reactions in the range of 3 nM to 30 mM, fusion became more efficient in
a concentration-dependent fashion reaching a maximum of ~84% of control reactions
(Fig. 2). This effect had a half-maximal value of ~90 µM Ca2+ and was specific to
calcium as other divalent cations such as Cu2+, Ba2+, and Mn2+ (all at 1 mM) were unable
to relieve the inhibition (Fig. 2). Although not shown, Cu2+, Ba2+, and Mn2+ did not
prevent hrs from binding in SNARE complexes formed in vitro as performed earlier
(Figs. 1 and 2). The concentration of calcium required and the cation specificity
necessary for relieving the hrs-dependent inhibition of early endosome fusion correlated
well with the block in formation of the hrs/SNAP-25/syntaxin 13 SNARE complex. This
provided evidence to suggest that calcium might function to dissociate a “nonfusogenic”
SNARE complex (containing hrs) and allow a “fusogenic” SNARE complex (containing
VAMP-2) to assemble on early endosomes.
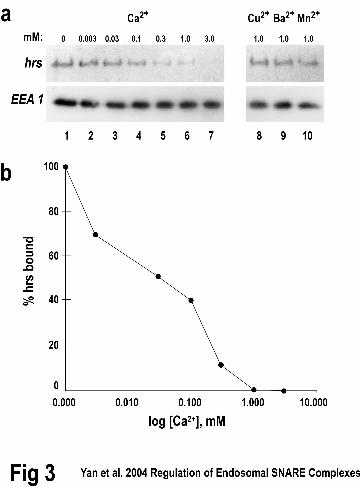
Calcium blocks hrs from binding to early endosomes
To further examine the notion that calcium might block binding of hrs to SNARE
complexes on membranes, we examined the effect of divalent cations on the binding of
hrs to purified early endosomes. When hrs was added (180 nM) to early endosomes
(purified as described in Material and Methods) in the presence of increasing Ca2+, a
concentration-dependent decrease of hrs binding was observed (Fig. 3, lanes 1-7). The
half-maximal value for calcium in this inhibition was ~30 µM. As seen previously (Fig.
3), Cu2+, Ba2+, and Mn2+ had no effect (Fig. 3, lanes 8-10). Importantly, Ca2+ did not
affect the binding of an early endosomal protein, EEA1, suggesting specificity in its
11
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

effect on hrs (Fig. 3, lanes 1-7). The inhibition of hrs binding to early endosomes and the
SNAP-25/syntaxin 13 SNARE complex suggested that Ca2+ might regulate how hrs
interacts with a receptor on membranes such as SNAP-25(17). SNAP-25 is considered to
be a strictly neuroendocrine or neuronal protein (although see (25)). Our use of HeLa
cells for not only purified early endosomes, but also homotypic endosome fusion assays,
and the dependence of these assays on SNAP-25 as suggested by their sensitivity to
Botulinum Toxin E, suggests that SNAP-25 resides in this non-neuronal cell. To
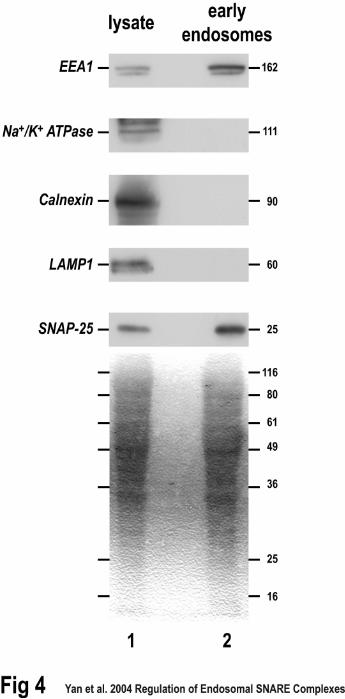
demonstrate that this is indeed the case, we attempted to detect SNAP-25 in Hela cell
lysates and purified early endosomes. As expected, markers for the ER (calnexin), plasma
membrane (Na/K ATPase), lysosomes (LAMP1), and early endosomes (EEA1) were
detectable in crude lysates from HeLa cells (Fig. 4, lane 1). The HeLa cell lysate also
contained SNAP-25 (Fig. 4, lane 1). Most importantly, purified membranes from HeLa
cells containing the early endosomal marker, EEA1, also contained SNAP-25 (Fig. 4,
lane 2). These membranes did not contain detectable amounts of the Na/K ATPase,
calnexin, or LAMP1, suggesting the lack of plasma membrane, ER, and lysosomes,
respectively (Fig. 4. lane 2). This demonstrated the membranes were not contaminated
with other organelles and allowed us to conclude that SNAP-25 was indeed located on
early endosomes from HeLa cells.
VAMP-2 is required for calcium to efficiently relieve hrs inhibition of early
endosome fusion
Certain bacterial toxins are valuable tools for dissecting the molecular details of
SNARE-dependent membrane fusion. These toxins are proteases with highly specific
12
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

cleavage recognition sites(9,24). For example, botulinum toxin E (BoNT/E) is a zinc
endoprotease that cleaves the C-terminal 26 amino acids of SNAP-25 and thereby blocks
membrane fusion that requires a four helical SNARE complex containing SNAP-25(9).
Likewise, tetanus toxin (TeTx) cleaves the C-terminal 41 amino acids of VAMP-2(24)
and blocks membrane fusion that requires a four helical SNARE complex containing
VAMP-2(1,7,9).
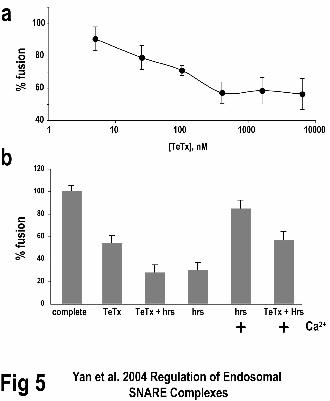
Pretreatment of early endosomes with TeTx inhibited membrane fusion in a dose
dependent manner (Fig. 5a). Treatment of early endosomal membranes resulted in ~54%
maximal fusion efficiency (Fig. 5a and b). The half-maximal value of TeTx for this
inhibition was ~80 nM comparable to the concentrations required to inhibit
neurotransmitter secretion. This result suggested that a TeTx substrate,
presumablyVAMP-2, played a role in early endosome fusion. The combination of TeTx
and hrs was no more efficacious than hrs alone in inhibiting early endosome fusion (Fig.
5b or Fig. 3). When Ca2+ was added along with hrs the fusion efficiency increased to
~85% (Fig. 5b and Fig. 2). However, if membranes were first treated with TeTx and then
incubated in the presence of hrs and free Ca2+, the fusion efficiency was just ~56% (Fig.
5b). This suggested that a TeTx substrate, presumably VAMP-2, was in part required for
calcium to reverse the hrs-dependent inhibition of early endosome fusion, accounting for
a loss of ~30% (85%-56%; Fig. 5b).
Release of calcium from luminal stores affects hrs function and localization
An experiment was performed to test if release of Ca2+ from luminal stores
could reverse hrs-dependent inhibition of early endosome fusion. The steady-state level
13
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

of Ca2+ in HeLa cells is estimated to be about 100 nM(26), which is below the
concentration that affects hrs-mediated block in early endosome fusion. During isolation,
endosomes lose their Ca2+ in 5-10 min through a vacuolar-type ATPase(27). To
overcome this, we loaded HeLa cells with extracellular calcium in the presence of
bafilomycin, an inhibitor of vacuolar-type ATPases(28,29). Bafilomycin was maintained
at a constant level during the lysis and isolation of endosomal membranes used in the
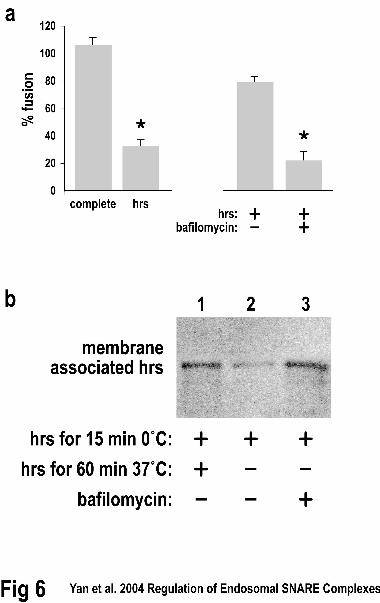
fusion assay, which would block release of luminal Ca2+. In control cells hrs inhibited
early endosome fusion by ~70% when it was present throughout the fusion reaction (Fig
6a. left side). In the continued presence of bafilomycin, fusion was inhibited by ~80%
when hrs was incubated with the endosomes and washed out prior to the beginning of
fusion reactions (Fig. 6a, right side). However, in the absence of bafilomycin, the effect
of hrs addition and removal prior to the fusion reactions was significantly decreased
(74.5 vs. 20.7% of control, Fig. 6a, right side). This nearly four-fold increase suggested
that luminal Ca2+ loss though a bafilomycin-sensitive V-ATPase can decrease the hrs-
dependent inhibition of early endosome fusion. To examine whether Ca2+ release from
luminal stores could dissociate hrs from endosomal membranes we performed a similar
experiment and quantitatively examined the amount of hrs present on the resulting
endosomes. Similar amounts of hrs were found on control endosomes and those in which
bafilomycin was present throughout the fusion reaction (Fig. 6b, lanes 1 and 3).
However, in the absence of bafilomycin the amount of hrs on the endosomal membranes
decreased to ~15% of control values (Fig. 6b, lane 2). These data suggest that luminal
Ca2+ can both remove hrs from endosomal membranes and reverse the inhibition of
endosomal fusion produced by hrs.
14
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Discussion
The SNARE hypothesis offers a molecular explanation for how membranes can
overcome the energy barrier for fusion. Pairing of SNARE proteins on opposing
membranes is required for, and may underlie the specificity of, the fusion reaction(1,7).
Regulation of SNARE complex formation/dissociation is critical for the control of
whether, and when, membrane fusion occurs. However, the control of SNARE complex
assembly/disassembly is poorly understood. The endosomal SNAREs, SNAP-25 and
syntaxin 13, exist in at least two distinct heterotrimeric complexes: one is fusogenic and
includes VAMP-2 and the other is nonfusogenic and includes hrs(17,20). The hrs-
containing complex appears to predominate in vitro and prevents formation of the
VAMP-2-containing complex(17). This suggests a mechanism by which hrs can inhibit
endosome fusion. We have examined the regulation of the hrs- and VAMP-2-containing
complexes and find that while the VAMP-2-containing complex is dissociated by the
action of the NSF ATPase, NSF does not dissociate the hrs-containing complex. In
contrast, Ca2+ dissociates the hrs-containing complex without affecting the VAMP-2-
containing complex. The micromolar concentrations of Ca2+ required for the dissociation
of the hrs-containing complex are similar to those that remove hrs from endosomal
membranes and that reverse the hrs-induced inhibition of VAMP-2-dependent homotypic
endosome fusion. This is the first demonstration of differential regulation of R/Q-SNARE
and all Q-SNARE containing SNARE complexes. Furthermore, these data suggest a
mechanism by which the regulation of hrs- and VAMP-2-containing complexes allows
for the regulation of homotypic endosome fusion. Thus, hrs binding to SNAP-25 on
15
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

early endosomal membranes negatively regulates trans-SNARE pairing and once Ca2+
dissociates hrs; a VAMP-2-containing complex forms, allowing fusion to occur.
The ATPase activity of NSF has been shown to dissociate SNARE complex
protein interactions(1,3,6,7), and recently for dissociating other protein complexes(30-33)
suggesting a conserved function as a dissociating factor. NSF binds indirectly to syntaxin
family members via interactions with αSNAP and in the presence of Mg/ATP disrupts
syntaxin/SNAP-25/VAMP complexes. The simplest explanation for the NSF resistance
of the hrs-containing complex is steric hindrance. Perhaps hrs, being relatively larger than
the SNAREs, blocks syntaxin from binding to aSNAP and NSF. Steric hindrance would
likely not block calcium from dissociating an hrs/SNAP-25/syntaxin 13 complex. The
effect of Ca2+ is specific as other divalent cations such as barium, copper, and manganese
have no effect on the hrs-containing complex. The role of Ca2+ in fusion of organelle
membranes has been recognized for years although there are many possible effector
proteins to which Ca2+ binds (e.g. (1,7,11-13,34), making an understanding of the
mechanism of Ca2+ action unclear. Ca2+ has been suggested to play a role in
conformational/structural changes in proteins, actions that could lead to protein complex
assembly or dissociation.
SNAP-25 has been recently shown to have a role in early endosome fusion(17) in
accord with its presence on endosome membranes(37). However, SNAP-25 has been
almost exclusively associated with a function in exocytosis. Moreover, SNAP-25 has
been thought to be present solely in neuronal and neuroendocrine tissues and cell lines
[although see(25)]. We have presented evidence that SNAP-25 is present in HeLa cell
lysate and on purified early endosomes isolated from HeLa cells, a cervical tumor derived
16
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

line. The presence of SNAP-25 on endosomes in these cells is a further suggestion that
SNAP-25 may be present in various cells from different lineages where it may be
involved in membrane fusion events [e.g. (25)].
The mechanism of the membrane association of hrs has been the subject of some
debate [(17) and references therein]. While the FYVE domain may provide a link to the
membrane though an interaction with PI3P(38), the Q-SNARE domain of hrs interacts
with SNAP-25 and SNAP-25 can act as a saturable binding site for hrs on endosomal
membranes(17). Since the FYVE domain may partially penetrate membranes and
regulate residence time(38) it is possible that SNAP-25 is the protein receptor and that the
FYVE-lipid interaction may be a regulatory influence for hrs binding to endosomes. Ca2+
dissociates hrs from early endosomal membranes at concentrations similar to that
required for dissociation of the hrs-SNAP-25 proteins complex. How relevant are these
Ca2+ concentrations for membrane fusion in the endocytic pathway? The luminal
concentration of Ca2+ in endosomes has been estimated at 1mM(35), 360µM in the Golgi,
350µM in the ER(26), and 2mM in yeast vacuole(11). Resting Ca2+ in HeLa cells is
approximately 100nM creating a large driving force for Ca2+, and the concentration of
Ca2+ at the endosome surface directly outside of the pore could likely reach 100µM or
greater(36). Ca2+ in the micromolar range can reverse the effect of chelators and support
endosome fusion(12,13). For homotypic vacuolar fusion, 100µM Ca2+ in the medium is
required to establish the normal luminal Ca2+ concentration and support fusion(11).
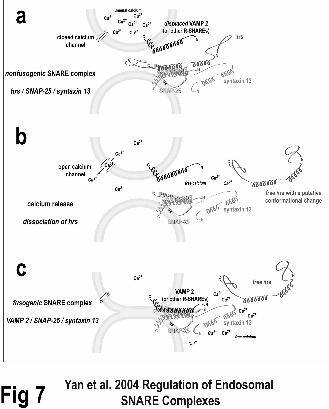
These data are consistent with a model in which luminal Ca2+ release, triggered by an
unknown mechanism, results in >100µM Ca2+ in discrete domains on the endosome
17
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

membrane, resulting in dissociation of hrs from SNAP-25, and allowing the VAMP-2-
SNAP-25-syntaxin13 complex to form that enables fusion (Figure 7).
Although tetanus toxin light chain inhibits the fusion of early endosomes in a
saturable and concentration dependent manner, the maximal inhibition obtained is
approximately 50%. In this regard, hrs can completely inhibit the formation of the
syntaxin 13– SNAP-25–VAMP-2 complex and it does not completely inhibit early
endosome fusion. Moreover, BoNT/E inhibits early endosome fusion to the same extent
as hrs. The incomplete inhibition of endosome fusion may be related to the inability of
tetanus toxin to affect previously formed trans-SNARE complexes. Alternatively, these
data may suggest the involvement of multiple R-SNAREs(39,40) in early endosomal
fusion.
Hrs inhibits early endosome fusion in a Ca2+-reversable manner. The
concentrations of Ca2+ required for the reversal of fusion, the dissociation of hrs from the
SNAP-25-syntaxin 13 complex, and the removal of hrs from endosomal membranes, are
consistent. If VAMP-2 is present when Ca2+-induces the removal of hrs from the SNAP-
25-syntaxin 13 complex then VAMP-2 will enter the SNARE complex, even in the
presence of Ca2+, and can support fusion. These data suggest differential regulation of
hrs- and VAMP-2-containing early endosomal SNARE complexes and suggest a
biochemical pathway by which luminal Ca2+ may regulate endosome fusion.
18
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure Legends
Figure 1. Calcium, but not NSF, dissociates a SNARE complex containing hrs. A.
GST-syntaxin 13 (2 µg) was bound to glutathione-agarose and incubated (4 °C for 1h)
with constant amounts of SNAP-25 (1 µg), VAMP-2 (1 µg) or hrs (2 µg) in the presence
or absence of Mg-ATP, NSF/αSNAP, EDTA, Mg-ATP-γ-S or Ca2+ as indicated. B. A
protein complex composed of hrs, SNAP-25, and syntaxin 13 was preformed (60 min at
4oC, lanes 1-3). After complex formation, either 2 mM EGTA (control, lane 1), 50µM
free Ca2+ (lane 2), or 300µM free Ca2+ (lane 3) was added and incubations were
continued (60 min at 4oC). All binding reactions were washed three times, boiled in SDS
sample buffer, and separated by SDS-PAGE. Hrs and VAMP 2 were detected using
specific antibodies, 125I-secondary antibodies, and quantified using phosphorimaging.
SNAP-25 and syntaxin 13 were detected using Ponceau S. The results are representative
of six such experiments. C. Calcium does not dissociate a SNARE complex containing
VAMP-2. Protein complexes with the same amounts of GST-syntaxin 13, SNAP-25, hrs,
of VAMP-2 were formed as in panel A. Free Ca2+ (50 µM) was added to the reactions in
lanes 4 and 5. Immunoblot analysis was used to detect hrs and VAMP-2 with specific
primary antibodies, 125I-secondary antibodies, and then phosphorimaging. SNAP-25 and
syntaxin 13 were detected using Ponceau S. The results are representative of three such
experiments.
Figure 2. Calcium relieves the hrs-dependent inhibition of early endosome fusion.
Separate cultures of HeLa cells were treated with EGF linked to either Alexa488 or
19
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

tetramethyl-rhodamine (TMR) and early endosomes were prepared for use as donor and
acceptor membranes(17). Fusion assays using standard conditions were performed in the
presence of various concentrations of free Ca2+, Cu2+, Ba2+, or Mn2+ as indicated. The
fluorescent signal at 580 nm (acceptor emission) was measured as an indicator of content
mixing(17). The results are representative of three such experiments.
Figure 3. Calcium inhibits hrs binding to early endosomes. A. Early endosomes were
purified from HeLa cells (described in Materials and Methods). Binding reactions
contained purified early endosomes, recombinant 6Xhis-hrs (180 nM), and various
concentrations of divalent cations as indicated (lanes 2-10). After incubations (60 min at
37˚C) membranes were collected by centrifugation at 100,000 xg, and proteins were
separated by SDS-PAGE. 6XHIS-hrs was detected with a monoclonal antibody against
the HIS tag, and quantified using an 125I-secondary antibody and phosphorimaging. The
early endosomal marker, EEA1, was detected with chemiluminescence. B. Plot of the hrs
amount bound to early endosomes from data in panel A. The results shown are
representative of six experiments.
Figure 4. Early endosomes from HeLa cells contain SNAP-25. HeLa cell lysate (lane
1) and purified early endosomes (lane 2) were subjected to SDS-PAGE and proteins were
detected on western blots with specific antibodies using chemiluminescence. In addition
to SNAP-25, marker proteins for the ER (calnexin), plasma membrane (Na/K ATPase),
lysosomes (LAMP1), and early endosomes (EEA1) were detected. The bottom blot
shows Ponceau S staining before antibody detection.
20
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 5. Tetanus toxin prevents efficient fusion of early endosomes. A. Titration of
tetanus toxin (TeTx) concentrations on early endosome fusion assays. B. The effect of
Ca2+, hrs and TeTx on early endosome fusion assays. The complete reaction was
performed under standard conditions and all others were the complete reaction plus the
indicated reagents. The results shown are representative of three experiments.
Figure 6. Bafilomycin prevents efficient fusion of early endosomes. A. The complete
homotypic early endosome fusion reactions were performed under standard
conditions(17). Hrs addition prior to, and during, the fusion reaction resulted in a
decrease in fusion to 30.4% of control (left panel, right side). If bafilomycin was added
during the endosome labeling and isolation and excluded from the fusion reaction, the
prior addition of hrs produced only a 26.5% inhibition of early endosome fusion (right
panel, left side). In contrast, if bafilomycin was present throughout the endosome
labeling and isolation and remained present during the fusion reactions, hrs inhibited the
fusion by ~80% (right panel, right side). The results shown are representative of three
experiments. B. Ca2+ release from luminal stores can dissociate hrs from endosomal
membranes. Endosomes were isolated as in panel A and the amount of hrs on their
membranes was quantified by western blotting with 125I-secondary antibody and
phosphorimaging. Similar amounts of hrs were found on control endosomes and those in
which bafilomycin was present throughout the fusion reaction (lanes 1 and 3). In the
absence of bafilomycin the amount of hrs on the endosomal membranes decreased to
15+/-5% of control values. The results shown are representative of three experiments.
21
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 7. A model of how calcium may regulate the formation of fusogenic and
nonfusogenic SNARE complexes on early endosomes. (A) The concentration of
calcium near two early endosomes is below that amount required to release hrs from a Q-
SNARE complex with SNAP-25 and syntaxin 13. Fusion of the endosomes is blocked.
(B) An unknown trigger activates a calcium channel in an early endosome (one
endosome shown for simplicity). The concentration of calcium rises locally to the point
that causes hrs to dissociate from the Q-SNARE complex most likely due to a putative
conformational change. (C) Once calcium removes hrs, VAMP-2 (or other R-SNARES)
can then from a fusogenic SNARE complex with SNAP-25 and syntaxin 13, facilitating
fusion of early endosomes. This model is based in part from how calcium might regulate
vacuole fusion in yeast(11).
22
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgments The authors wish to thank Dr. Phyllis Hanson for helpful discussions, as well as Dr. Neal Waxham for comments on the manuscript. We thank Bill Evans and Yasmin Lotfi for technical assistance. These studies were supported in part by NIH MH58920.
23
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

References 1. Jahn, R., Lang, T., and Sudhof, T. C. (2003) Cell 112, 519-533
2. Lin, R. C., and Scheller, R. H. (2000) Annu Rev Cell Dev Biol 16, 19-49
3. Sollner, T., Bennett, M. K., Whiteheart, S. W., Scheller, R. H., and Rothman, J. E.
(1993) Cell 75, 409-418
4. Sutton, R. B., Fasshauer, D., Jahn, R., and Brunger, A. T. (1998) Nature 395, 347-
353
5. Chen, Y. A., Scales, S. J., and Scheller, R. H. (2001) Neuron 30, 161-170
6. Weber, T., Parlati, F., McNew, J. A., Johnston, R. J., Westermann, B., Sollner, T.
H., and Rothman, J. E. (2000) J Cell Biol 149, 1063-1072
7. Chen, Y. A., and Scheller, R. H. (2001) Nat Rev Mol Cell Biol 2, 98-106
8. Fasshauer, D., Sutton, R. B., Brunger, A. T., and Jahn, R. (1998) Proc Natl Acad
Sci U S A 95, 15781-15786
9. Hayashi, T., McMahon, H., Yamasaki, S., Binz, T., Hata, Y., Sudhof, T. C., and
Niemann, H. (1994) Embo J 13, 5051-5061
10. Sullivan, K. M., Busa, W. B., and Wilson, K. L. (1993) Cell 73, 1411-1422
11. Peters, C., and Mayer, A. (1998) Nature 396, 575-580
12. Holroyd, C., Kistner, U., Annaert, W., and Jahn, R. (1999) Mol Biol Cell 10,
3035-3044
13. Colombo, M. I., Beron, W., and Stahl, P. D. (1997) J Biol Chem 272, 7707-7712
14. Quetglas, S., Leveque, C., Miquelis, R., Sato, K., and Seagar, M. (2000) Proc
Natl Acad Sci U S A 97, 9695-9700
24
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

15. Quetglas, S., Iborra, C., Sasakawa, N., De Haro, L., Kumakura, K., Sato, K.,
Leveque, C., and Seagar, M. (2002) Embo J 21, 3970-3979
16. Tsujimoto, S., Pelto-Huikko, M., Aitola, M., Meister, B., Vik-Mo, E. O.,
Davanger, S., Scheller, R. H., and Bean, A. J. (1999) Eur J Neurosci 11, 3047-
3063
17. Sun, W., Yan, Q., Vida, T. A., and Bean, A. J. (2003) J Cell Biol 162, 125-137
18. Bean, A. J., Seifert, R., Chen, Y. A., Sacks, R., and Scheller, R. H. (1997) Nature
385, 826-829
19. Kwong, J., Roundabush, F. L., Hutton Moore, P., Montague, M., Oldham, W., Li,
Y., Chin, L. S., and Li, L. (2000) J Cell Sci 113 ( Pt 12), 2273-2284
20. Prekeris, R., Klumperman, J., Chen, Y. A., and Scheller, R. H. (1998) J Cell Biol
143, 957-971
21. Advani, R. J., Bae, H. R., Bock, J. B., Chao, D. S., Doung, Y. C., Prekeris, R.,
Yoo, J. S., and Scheller, R. H. (1998) J Biol Chem 273, 10317-10324
22. Ward, D. M., Pevsner, J., Scullion, M. A., Vaughn, M., and Kaplan, J. (2000) Mol
Biol Cell 11, 2327-2333
23. Tsujimoto, S., and Bean, A. J. (2000) J Biol Chem 275, 2938-2942
24. McMahon, H. T., Ushkaryov, Y. A., Edelmann, L., Link, E., Binz, T., Niemann,
H., Jahn, R., and Sudhof, T. C. (1993) Nature 364, 346-349
25. Zhao, C. M., Jacobsson, G., Chen, D., Hakanson, R., and Meister, B. (1997) Cell
Tissue Res 290, 539-551
25
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

26. Van Baelen, K., Vanoevelen, J., Callewaert, G., Parys, J. B., De Smedt, H.,
Raeymaekers, L., Rizzuto, R., Missiaen, L., and Wuytack, F. (2003) Biochem
Biophys Res Commun 306, 430-436
27. Gerasimenko, J. V., Tepikin, A. V., Petersen, O. H., and Gerasimenko, O. V.
(1998) Curr Biol 8, 1335-1338
28. Finbow, M. E., and Harrison, M. A. (1997) Biochem J 324 ( Pt 3), 697-712
29. Stevens, T. H., and Forgac, M. (1997) Annu Rev Cell Dev Biol 13, 779-808
30. Muller, J. M., Rabouille, C., Newman, R., Shorter, J., Freemont, P., Schiavo, G.,
Warren, G., and Shima, D. T. (1999) Nat Cell Biol 1, 335-340
31. Muller, J. M., Shorter, J., Newman, R., Deinhardt, K., Sagiv, Y., Elazar, Z.,
Warren, G., and Shima, D. T. (2002) J Cell Biol 157, 1161-1173
32. Hanley, J. G., Khatri, L., Hanson, P. I., and Ziff, E. B. (2002) Neuron 34, 53-67
33. Sagiv, Y., Legesse-Miller, A., Porat, A., and Elazar, Z. (2000) Embo J 19, 1494-
1504
34. Pryor, P. R., Mullock, B. M., Bright, N. A., Gray, S. R., and Luzio, J. P. (2000) J
Cell Biol 149, 1053-1062
35. Chemello, M. E., Aristimuno, O. C., Michelangeli, F., and Ruiz, M. C. (2002) J
Virol 76, 13083-13087
36. Smith, S. J., Buchanan, J., Osses, L. R., Charlton, M. P., and Augustine, G. J.
(1993) J Physiol 472, 573-593
37. Tao-Cheng, J. H., Du, J., and McBain, C. J. (2000) J Neurocytol 29, 67-77
38. Stahelin, R. V., Long, F., Diraviyam, K., Bruzik, K. S., Murray, D., and Cho, W.
(2002) J Biol Chem 277, 26379-26388
26
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

27
39. Martinez-Arca, S., Alberts, P., and Galli, T. (2000) Biol Cell 92, 449-453
40. Muzerelle, A., Alberts, P., Martinez-Arca, S., Jeannequin, O., Lafaye, P., Mazie,
J. C., Galli, T., and Gaspar, P. (2003) Neuroscience 122, 59-75
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from

Qing Yan, Wei Sun, James A. McNew, Thomas A. Vida and Andrew J. Beanendosomes
and NSF differentially regulate disassembly of SNARE complexes on early2+Ca
published online February 9, 2004J. Biol. Chem.
10.1074/jbc.M400093200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 1, 2018http://w
ww
.jbc.org/D
ownloaded from