Embed Size (px)
Citation preview

Vol. 165, No. 3JOURNAL OF BACTERIOLOGY, Mar. 1986, p. 746-7550021-9193/86/030746-10$02.00/0Copyright C 1986, American Society for Microbiology
Negative Staining and Immunoelectron Microscopy of Adhesion-Deficient Mutants of Streptococcus salivarius Reveal that the
Adhesive Protein Antigens Are Separate Classes of CellSurface Fibril
ANTON H. WEERKAMP,1* PAULINE S. HANDLEY,2 ADA BAARS,3 AND JAN W. SLOT3Department of Oral Biology, Dental School, University of Groningen, 9713 AV Groningen, The Netherlands'; DepartmentofBacteriology and Virology, The Medical School, Manchester M13 9PT, England2; and Center for Electron Microscopy,
Medical School, University of Utrecht, 3511 HG Utrecht, The Netherlands3
Received 19 June 1985/Accepted 5 November 1985
The subcellular distribution of the cell wall-associated protein antigens of Streptococcus salivarius HB, whichare involved in specific adhesive properties of the cells, was studied. Mutants which had lost the adhesiveproperties and lacked the antigens at the cell surface were compared with the parent strain. Immunoelectronmicroscopy of cryosections of cells labeled with affinity-purified, specific antisera and colloidal gold-protein Acomplexes was used to locate the antigens. Antigen C (AgC), a glycoprotein involved in attachment to hostsurfaces, was mainly located in the fibrillar layer outside the cell wall. A smaller amount of label was also foundthroughout the cytoplasmic area in the form of small clusters of gold particles, which suggests a macromolec-ular association. Mutant HB-7, which lacks the wall-associated AgC, accumulated AgC reactivityintracellularly. Intracellular AgC was often found associated with isolated areas of increased electron density,but sometimes seemed to fill the entire interior of the cell. Antigen B (AgB), a protein responsible forinterbacterial coaggregation, was also located in the fibrillar layer, although its distribution differed from thatof the wall-associated AgC since AgB was found predominantly in the peripheral areas. A very small amountof label was also found in the cytoplasmic area as discrete gold particles. Mutant HB-V5, which lackswall-associated AgB, was not labeled in the fibrillar coat, but showed the same weak intracellular label as theparent strain. Immunolabeling with serum against AgD, another wall-associated protein but of unknownfunction, demonstrated its presence in the fibrillar layer of strain HB. Negatively stained preparations of wholecells of wild-type S. salivarius and mutants that had lost wall-associated AgB or AgC revealed that two classesof short fibrils are carried on the cell surface at the same time. AgB and AgC are probably located on separateclasses of short, protease-sensitive fibrils 91 and 72 nm in length, respectively. A third class of only verysparsely distributed short fibrils (63 nm) was observed on mutant HB-V51, which lacks both wall-associatedAgB and AgC antigens. The identity of these fibrils and whether they are present on the wild type are not clear.The function of long, protease-resistant fibrils of 178 nm, which are also present on the wild-type strain,remains unknown.
The involvement of surface appendages in the adhesion ofa wide variety of indigenous and pathogenic bacteria to hostsurfaces is well documented. Surface appendages whichmediate the attachment of streptococci to host surfaces areoften associated with a fibrillar layer outside the cell wall,historically termed the "fuzzy coat" on the basis of earlyobservations of thin sections of cells. This morphologicallyindistinct layer is present in such organisms as Streptococ-cus pyogenes (2), Streptococcus salivarius (5), Streptococ-cus mitior (11), and Streptococcus mutans (13). The pres-ence of proteinaceous compounds in this layer is indicatedby the observation that protease treatment removes thefuzzy coat (2). More recent studies on the morphology of thefuzzy coat of S. salivarius Lancefield group K cells (6)showed that this layer consists of densely packed fibrils witha loose, amorphous appearance. Similar observations weresubsequently made on strains of Streptococcus sanguis (7).However, little detailed knowledge is available on the finestructure and the chemical and functional components ofthese layers.The fuzzy coat of S. pyogenes contains the M protein, an
* Corresponding author.
antiphagocytic molecule (18) which also may be involved inthe adhesion of the bacteria to human tracheal epithelial cells(2). The fuzzy coat has furthermore been reported to containthe R and T proteins (8). M protein-like structures withreceptor activities for various serum proteins have beenobserved in these bacteria, as well as in group B, C, and Gstreptococci (10) among others.
S. salivarius K+ cells carry an elaborate fuzzy coat (5)consisting of two morphologically distinct types of fibrils (6)and at least two categories of specific functional, proteinace-ous compounds, or adhesins, mediating attachment to hostsurfaces and to various gram-negative oral bacteria, respec-tively (21, 23, 24). Two adhesins, the host attachment factor(antigen C; AgC) and the Veillonella-binding protein (AgB),respectively, were previously isolated from cell walls of S.salivarius HB and purified to apparent homogeneity, inaddition to a third cell wall protein antigen (AgD) whosefunction is not known (22). It would therefore be of interestto attempt the correlation of morphological and functionalstudies of the cell surface.
S. salivarius is a successful inhabitant of the human oralcavity which preferentially attaches to keratinized oral epi-thelial cells, reflecting its preference for the tonque dorsum(16). In addition, S. salivarius readily forms aggregates with
746
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
LOCALIZATION OF CELL SURFACE FIBRILLAR ANTIGENS
several oral anaerobic, gram-negative bacteria, which issuggested to enhance intergeneric ecological relationships inmulticellular microbial conglomerates (21).
In this paper we describe the localization of the S.salivarius adhesins by immunoelectron microscopy (IEM).We also present studies of the surface morphology of mutantstrains specifically lacking these adhesins, using the nega-tive-stain technique. A schematic picture of the cell surfacebased on these observations, and involving at least twofunctionally different fibril types, has been constructed.
MATERIALS AND METHODS
Bacterial strains and growth conditions. S. salivarius HBand mutant strains HB-7 and HB-V5 have been describedpreviously (23, 24). Mutant strain HB-V51 was prepared fromstrain HB-V5 by nitrosoguanidine mutagenesis according topreviously described procedures (23) and was unable toattach to buccal epithelial cells or to aggregate with saliva; inaddition it was unable to coaggregate with veillonellae.
Unless otherwise stated, the strains were grown in batchculture in Todd-Hewitt broth for 16 h at 37°C in air with 5%CO2.
Preparation of antisera. Monospecific polyclonal antiseraagainst AgB, AgC, and AgD were prepared by immunoaf-finity chromatography of whole antiserum against S.salivarius HB cell walls (24) on columns of the purifiedantigens immobilized on Affigel-10 (Bio-Rad Laboratories).The antigens were isolated from lysates of cell wall from thewild-type strain and purified by a series of proceduresincluding ion-exchange and gel permeation chromatography,preparative isoelectric focusing, and affinity chromatogra-phy on concanavalin-Sepharose, as described in the accom-panying paper (27). The immunoaffinity columns (6 by 1.0cm) were preequilibrated with 0.1 M phosphate buffer (pH7.3) containing 0.5 M NaCl. After application of the serumsample (4 ml), the columns were eluted with the same bufferuntil no UV-absorbing material was detected in the eluate.Bound antibodies were subsequently released by eluting thecolumn with 0.1 M glycine hydrochloride buffer (pH 2.5).The pH of the eluate was quickly neutralized by the additionof solid Tris. The eluate was then either dialyzed and freezedried or concentrated by ultrafiltration using an Amicon cellequipped with a PM30 filter (Amicon Benelux, Oosterhout,The Netherlands) and stored at -20°C in the presence of 1%bovine serum albumin.The antisera were tested for purity by crossed immuno-
electrophoresis with crude cell wall digests of strain HB.Standard immunoelectrophoretic techniques in agarose gelsusing a Tris-Veronal buffer system (pH 8.6) were describedpreviously (22). Bromophenol blue was used as a marker forelectrophoresis in the first dimension.IEM. The subcellular location of antigens was tested using
IEM on thin sections (<100 nm) of frozen cells with asandwich labeling technique employing colloidal gold-complexed protein A, according to Geuze et al. (4). The cellswere fixed by mixing equal volumes of overnight culturesand a solution containing 0.1 M phosphate buffer (pH7.4)-4% (wt/vol) paraformaldehyde-0.4% (wt/vol) glutaral-dehyde. The bacteria were pelleted immediately, resus-pended in fixation solution containing 2% paraformaldehydeand 0.2% glutaraldehyde, and kept on ice for 1 to 2 h. Thefixed cells were then embedded in 10% (wt/vol) gelatin (15),and ultrathin cryosections were cut at -90°C and processedas described previously (4, 15).
All immunoincubations, washings, staining, and embed-
ding were performed at room temperature. The incubationswith antibodies and labels lasted 30 min; washes werecarried out with phosphate-buffered saline. Uniform proteinA-gold markers of 6 nm were used (17). After im-munolabeling, the sections were postfixed in 0.5% (wt/vol)glutaraldehyde, stained with uranylacetate, and embedded inmethylcellulose (20).
Negative staining. The bacteria were grown overnight inbrain heart infusion broth at 37°C, and the cells wereharvested and washed three times in distilled water. Thefinal suspension was used for negative staining with 1%methylamine tungstate as described previously (6). Fibrillengths were measured by drawing a line around the marginsof the fibrillar fringes on photographs of the cells. Twenty-seven micrographs of each strain, from three differentbatches, were measured in this way. Statistical analysis ofthe fibril lengths on wild-type and mutant strains was per-formed using the factor analysis of variance followed by acomparison of individual group means based on the least-significant-difference approach. Fibrillar lengths are not ab-solute lengths; each new measurement on a different batchdoes not give the exact measurement as tabulated, so thatstandard deviations are consistently quoted throughout thepaper.
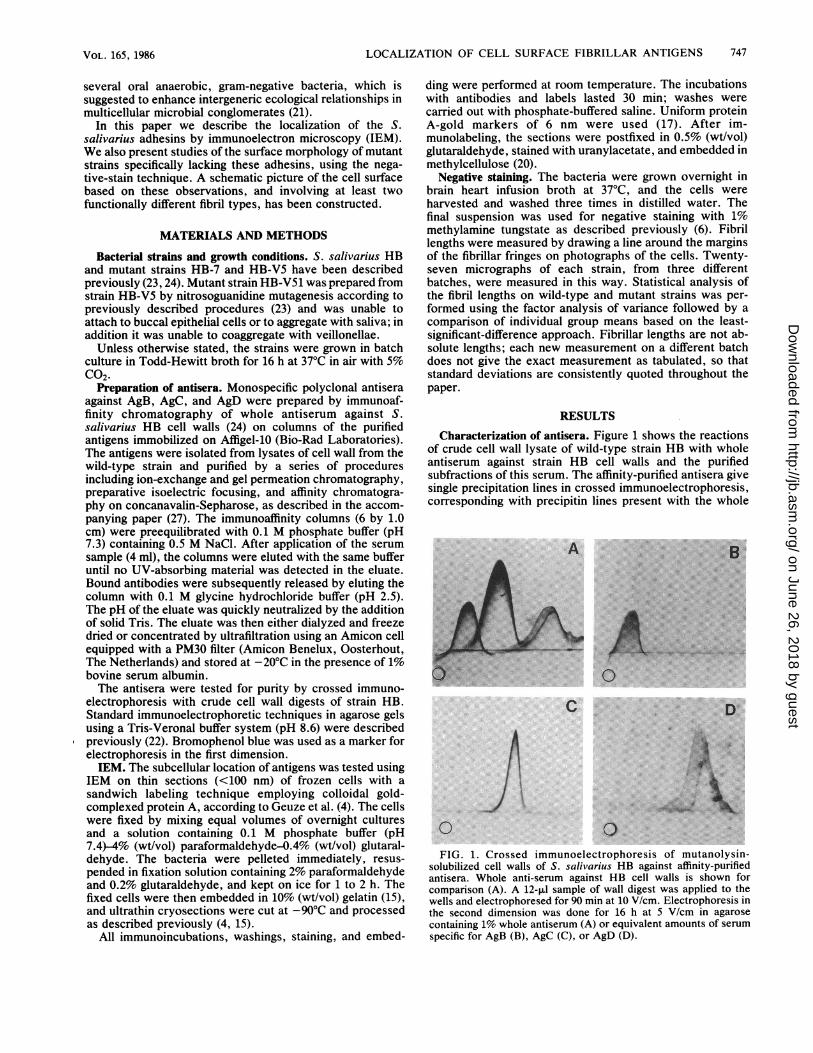
RESULTSCharacterization of antisera. Figure 1 shows the reactions
of crude cell wall lysate of wild-type strain HB with wholeantiserum against strain HB cell walls and the purifiedsubfractions of this serum. The affinity-purified antisera givesingle precipitation lines in crossed immunoelectrophoresis,corresponding with precipitin lines present with the whole
.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~: ..0........::..:.. :.:: ..
>~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.t . .SA....0
C D...
BEs i.Ct x | ... n .. .a lE:
AsSw fL.:: ,. ^
... :.:..:.;....wH-:...0::3k::.0FIG. 1. Crossed immunoelectrophoresis of mutanolysin-
solubilized cell walls of S. salivarius HB against affinity-purifiedantisera. Whole anti-serum against HB cell walls is shown forcomparison (A). A 12-,u sample of wall digest was applied to thewells and electrophoresed for 90 min at 10 V/cm. Electrophoresis inthe second dimension was done for 16 h at 5 V/cm in agarosecontaining 1% whole antiserum (A) or equivalent amounts of serumspecific for AgB (B), AgC (C), or AgD (D).
747VOL. 165, 1986
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
748 WEERKAMP ET AL. J. BACTERIOL.
* | *1 _ u .;. w0t0.o x+S~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~...f
*,l E t',i, " '.',,s,t,,''.,;R,'S;-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~II*iIIl
I I | I l * ~~~~~~~~~~~~~~~~~~~~~~~~.._-t5, /C-t > iNtw20X>,~~~~~~~~~~~.FIG.2.UltrathincryosectionsLi_Fof Ssai rus HB nietyimnlbldwt nieumaantACadpoenAclodlgl
nrh.Tpmlliit tntqen h-6n ni itce.-(AadR ersnttv rnrton ftodfeen utrs hwii o A n
high (B) intracellular labeling. Bar, 0.5 ,um.
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 165, 1986 LOCALIZATION OF CELL SURFACE FIBRILLAR ANTIGENS
B
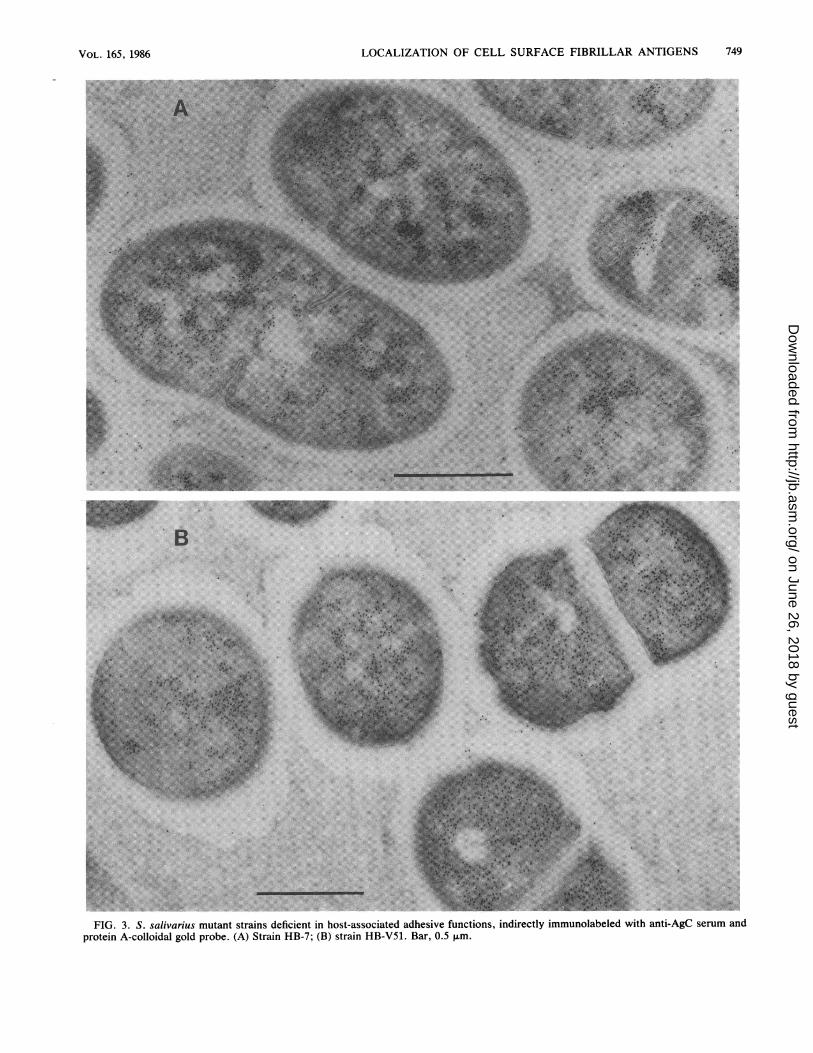
FIG. 3. S. salivarius mutant strains deficient in host-associated adhesive functions, indirectly immunolabeled with anti-AgC serum andprotein A-colloidal gold probe. (A) Strain HB-7; (B) strain HB-V51. Bar, 0.5 ,um.
749
....
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
750 WEERKAMP ET AL.
A
B
,,N _ .iN:.. ,,. Si ?
,tijSi,5R,09 dX2 te, + ,~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
...
Ut
A.M.
.............
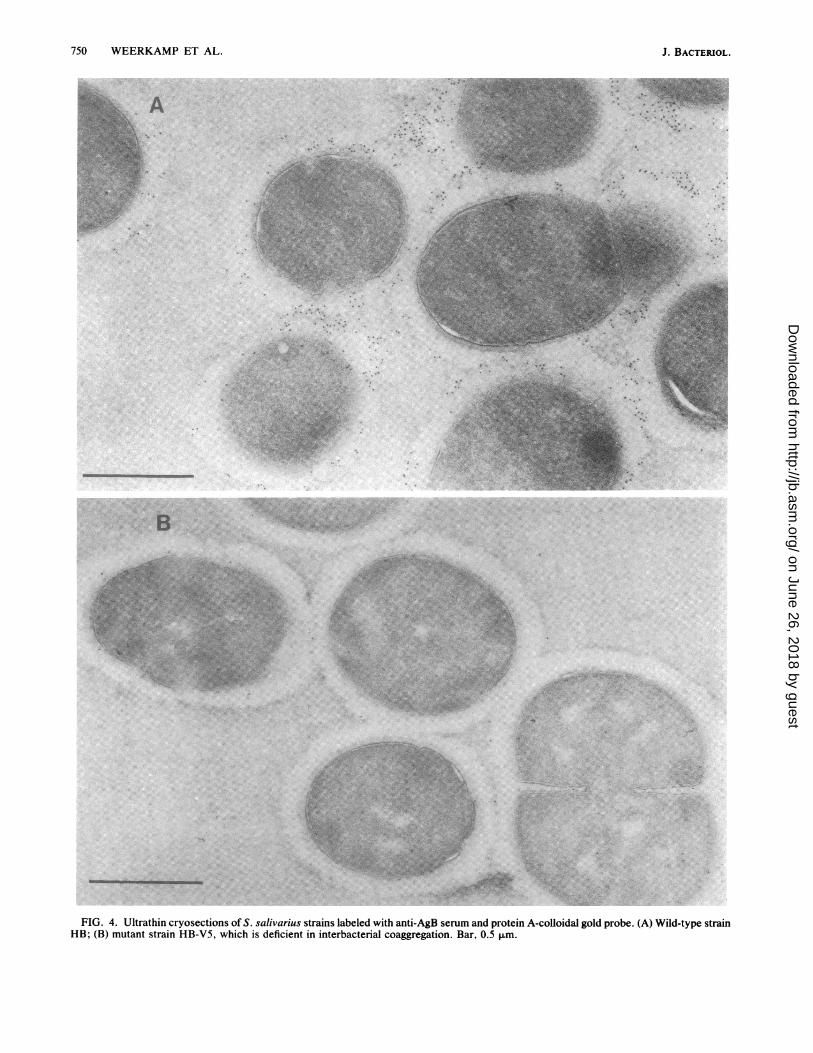
FIG. 4. Ultrathin cryosections of S. salivarius strains labeled with anti-AgB serum and protein A-colloidal gold probe. (A) Wild-type strainHB; (B) mutant strain HB-V5, which is deficient in interbacterial coaggregation. Bar, 0.5 ,um.
J. BACTERIOL.
I
11
II
e.,
1.
'A,
.*.
:
.Vl.-i .,-
. ;P.;.w..:P.M.R
FNE.".-';N'l"
*...'Mki -.
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
LOCALIZATION OF CELL SURFACE FIBRILLAR ANTIGENS
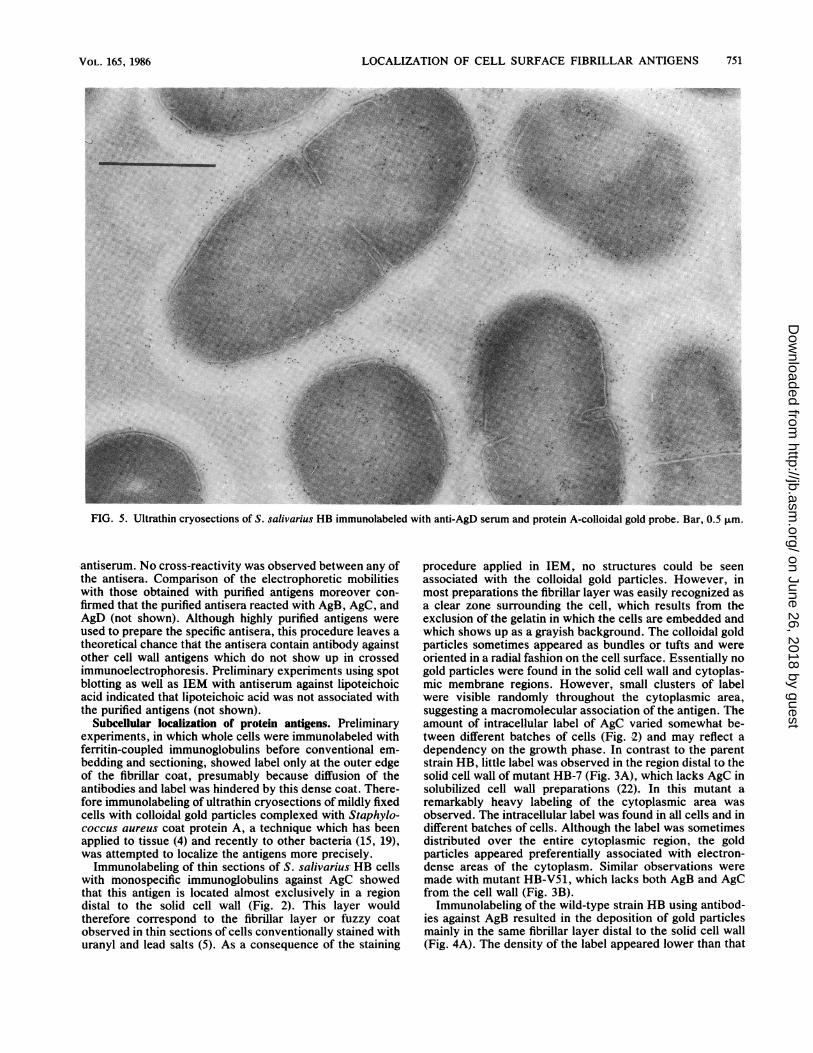
salivarius HB immunolabeled with anti-AgD serum and protein A-colloidal gold probe. Bar, 0.5 ,um.
antiserum. No cross-reactivity was observed between any ofthe antisera. Comparison of the electrophoretic mobilitieswith those obtained with purified antigens moreover con-
firmed that the purified antisera reacted with AgB, AgC, andAgD (not shown). Although highly purified antigens wereused to prepare the specific antisera, this procedure leaves atheoretical chance that the antisera contain antibody againstother cell wall antigens which do not show up in crossedimmunoelectrophoresis. Preliminary experiments using spotblotting as well as IEM with antiserum against lipoteichoicacid indicated that lipoteichoic acid was not associated withthe purified antigens (not shown).
Subcellular localization of protein antigens. Preliminaryexperiments, in which whole cells were immunolabeled withferritin-coupled immunoglobulins before conventional em-bedding and sectioning, showed label only at the outer edgeof the fibrillar coat, presumably because diffusion of theantibodies and label was hindered by this dense coat. There-fore immunolabeling of ultrathin cryosections of mildly fixedcells with colloidal gold particles complexed with Staphylo-coccus aureus coat protein A, a technique which has beenapplied to tissue (4) and recently to other bacteria (15, 19),was attempted to localize the antigens more precisely.
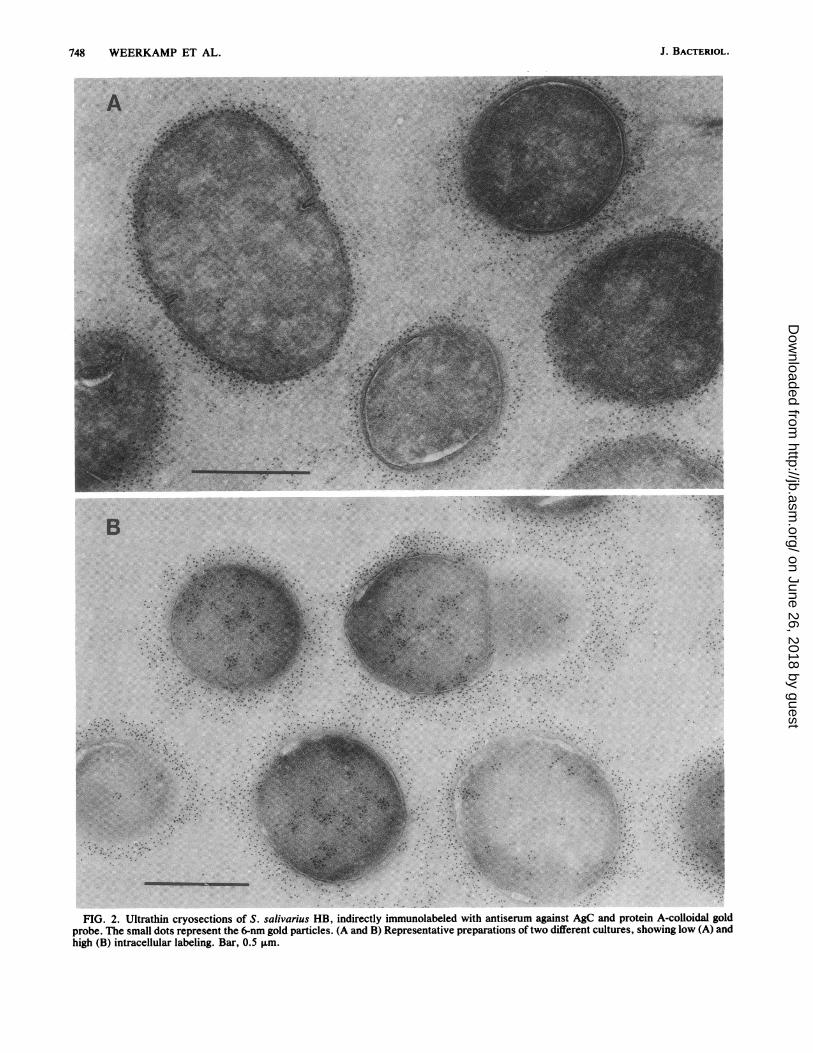
Immunolabeling of thin sections of S. salivarius HB cellswith monospecific immunoglobulins against AgC showedthat this antigen is located almost exclusively in a regiondistal to the solid cell wall (Fig. 2). This layer wouldtherefore correspond to the fibrillar layer or fuzzy coatobserved in thin sections of cells conventionally stained withuranyl and lead salts (5). As a consequence of the staining
procedure applied in IEM, no structures could be seenassociated with the colloidal gold particles. However, inmost preparations the fibrillar layer was easily recognized asa clear zone surrounding the cell, which results from theexclusion of the gelatin in which the cells are embedded andwhich shows up as a grayish background. The colloidal goldparticles sometimes appeared as bundles or tufts and wereoriented in a radial fashion on the cell surface. Essentially nogold particles were found in the solid cell wall and cytoplas-mic membrane regions. However, small clusters of labelwere visible randomly throughout the cytoplasmic area,suggesting a macromolecular association of the antigen. Theamount of intracellular label of AgC varied somewhat be-tween different batches of cells (Fig. 2) and may reflect adependency on the growth phase. In contrast to the parentstrain HB, little label was observed in the region distal to thesolid cell wall of mutant HB-7 (Fig. 3A), which lacks AgC insolubilized cell wall preparations (22). In this mutant aremarkably heavy labeling of the cytoplasmic area wasobserved. The intracellular label was found in all cells and indifferent batches of cells. Although the label was sometimesdistributed over the entire cytoplasmic region, the goldparticles appeared preferentially associated with electron-dense areas of the cytoplasm. Similar observations weremade with mutant HB-V51, which lacks both AgB and AgCfrom the cell wall (Fig. 3B).
Immunolabeling of the wild-type strain HB using antibod-ies against AgB resulted in the deposition of gold particlesmainly in the same fibrillar layer distal to the solid cell wall(Fig. 4A). The density of the label appeared lower than that
FIG. 5. Ultrathin cryosections of S.
VOL. 165, 1986 751
............. ......
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
752 WEERKAMP ET AL.
|||lt^=,x4',̂ B
..w~..
- .s '.m _ >,e'J!e..t
with.glbuaed. Bar 0.1
J. BACTERIOL.
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

LOCALIZATION OF CELL SURFACE FIBRILLAR ANTIGENS
BA
D
\A'4.KL14FIG. 7. Diagrammatic representation of the surface structures
observed by negative staining on the wild-type strain HB and itsmutants. (A) Strain HB carries long (178 + 11 nm) and short (91 ±5 nm) fibrils. (B) Strain HB-VS carries long (166 ± 11 nm) and short(72 ± 3 nm) fibrils. (C) Strain HB-7 carries only 90 ± 4-nm fibrilswhich are less dense than those of either strain HB or HB-V5. (D)Strain HB-V51 carries very sparse, short (63 ± 7 nm) fibrils only.
observed with AgC, more irregularly distributed, and pref-erentially located at the peripheral fringe of the fibrillar coat.Little label was observed in the solid cell wall, membrane,and cytoplasmic area. In contrast to AgC, the intracellularAgB label showed up as discrete gold particles and not asclusters. Mutant HB-V5, which specifically lacks wall-associated AgB (24), did not show label in the fibrillar layerupon treatment with antibodies against AgB (Fig. 4B). StrainHB-V5 reacted similarly to the wild-type strain when anti-serum against AgC was applied. Only a weak labeling,similar to that found in the wild-type strain, was found in thecytoplasmic area of the mutant.
S. salivarius HB was also treated with antibodies againstAgD (Fig. 5). AgD appeared to reside mainly in the fibrillarlayer, but label was also found in the cytoplasmic area,similar to AgC, and presumably also in the solid cell walL Anegative control could not be carried out with AgD becauseno mutants were available which specifically lack this anti-gen.
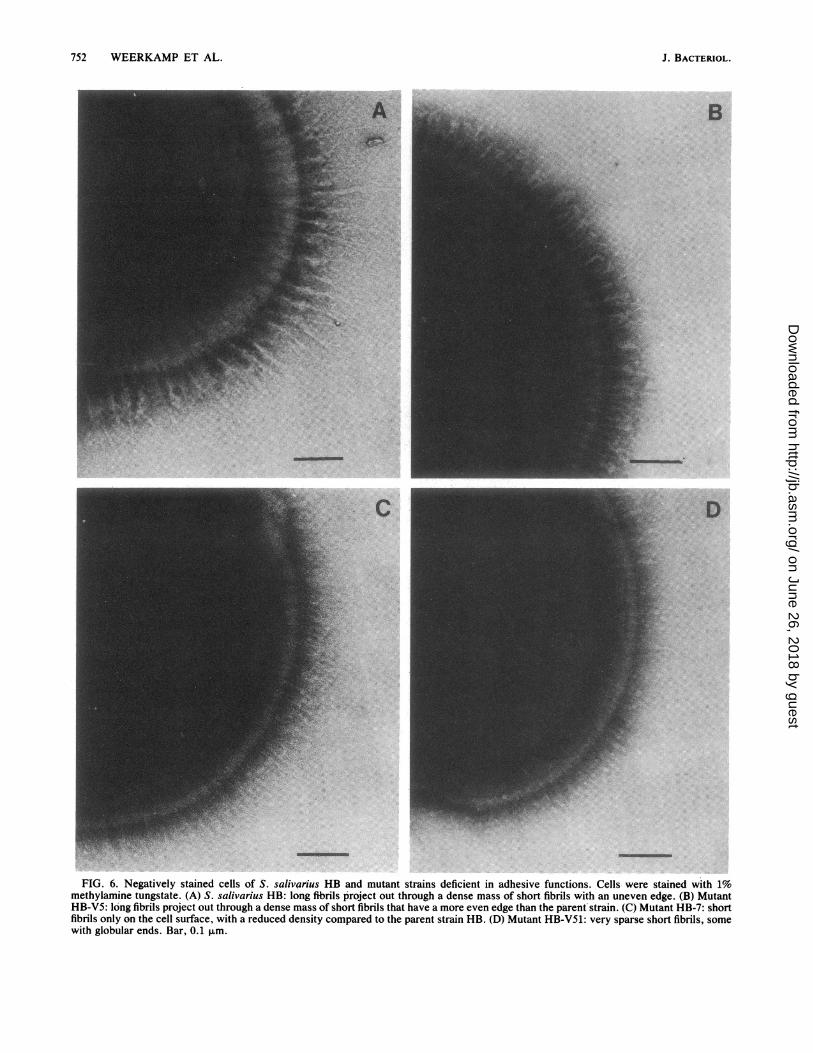
Negative staining. Whole cells of the wild-type strain HBand mutant strains were observed by negative staining in theelectron microscope to compare the structural compositionof the cell surfaces with the presence of particular proteinantigens (Fig. 6). As shown previously (6), negativelystained cells of strain HB apparently carry two types offibrils on the cell surface (Fig. 6A). These can be distin-
guished from each other on the basis of their length, density,and protease sensitivity (6). The long, slender fibrils have amean length of 178 ± 11 nm, are protease resistant, and aremore sparsely distributed than the short fibrils. The shorter,more globular, densily packed fibrils (91 ± 5 nm) have anuneven outside edge (Fig. 6A) and are protease sensitive (6).Surface structure analysis of the adhesion-deficient mutantsu'sing negative staining has revealed that the shorter fibrilsconsist of two or three separate fibril classes that can bedistinguished from each other on the basis of length (Fig. 7).Mutant HB-V5 appeared to be very similar to strain HB,
as it carries both long and short fibrils (Fig. 6B). The longfibrils (166 ± 11 nm) are not significantly shorter than thoseon strain HB (178 ± 11 nm). However, the short fibrils areonly 72 ± 3 nm long and are significantly shorter by 19 nm (P< 0.05) than the 91 + 5-nm fibrils on strain HB. Thisdifference in length suggests either that the 91-nm, fibrilshave lost length or that the 72-nm fibrils represent a distinctclass of fibrils obscured in the parent strain. These shorterfibrils have no measurable width when on the cell surfaceand have globular ends (Fig. 6B).Mutant HB-7 carries only short fibrils with a length of 90
± 4 nm and has completely lost the longer, more sparselydistributed fibrils. These 90-nm fibrils are not significantlyshorter than the 91-nm fibrils on the parent strain HB.Mutant HB-V51 carries very sparsely distributed fibrils
which are 63 ± 7 nm long and have globular ends. The 178-,91-, and 72-nm-long fibril classes have been lost (Fig. 6D).These 63-nm fibrils are significantly shorter (P < 0.05) thanthe 72-nm class of short fibrils and could represent a thirdstructural and functional class of short fibrils, obscured inboth strains HB-7 and HB-V5. Figure 8 is a compositediagram of all the classes of fibrils probably found on thewild-type strain HB.
DISCUSSIONCell walls of S. salivarius HB and most other Lancefield
group K strains of this species contain three majorproteinaceous components (22), at least two of which nowappear to be associated with different structural classes offibril on the surface of the cells. These fibrils form a denselypacked, amorphous layer with an almost confluent outeredge (6), representing the characteristic fuzzy coat of S.salivarius K+ cells. A similar layer is also found in a varietyof other streptococci (2, 4, 11, 13). A remarkable finding ofthis and our previous work (6, 22, 26) is that the fuzzy coat
178 ..............................
FIG. 8. Composite diagram to show the lengths and possible functions of some of the fibril classes on S. salivarius HB. Long, 178-nmfibrils: protease resistant; function(s) unknown. Short, 91-nm fibrils: protease sensitive; coaggregation adhesit AgB. Short, 72-nm fibrils:protease sensitive; -host-associated adhesin AgC. Short, 63-nm fibrils- protease sensitive; identity and function(s) unknown; actual presenceon strain HB not established.
753VOL. 165, 1986
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

754 WEERKAMP ET AL.
appears to be composed of biochemically, antigenically, andstructurally different fibrils which carry distinct adhesivefunctions. Each fibril class could represent single moleculesof the respective protein antigen.The fibrillar nature of one of the antigens, AgC, was
corroborated by the recent demonstration that this com-pound contains a high level of a-helix configuration (26). Inan accompanying paper (27) we furthermore demonstratethat purified AgB and AgC have fibrillar configurations whenobserved in the electron microscope. AgC, which is aglycoprotein containing about 42% by weight of hexose andhexosamine, was shown to be involved in adhesion ofbacteria to host surfaces and aggregation with saliva (22).AgB, which contains little carbohydrate, mediates coaggre-gation of the bacteria with oral Veillonella strains andvarious other gram-negative bacteria (21, 24). The biologicalfunctions of the third protein antigen, AgD, are so far notknown. The presence of antigenically and functionally dif-ferent surface appendages on the same cell was alreadydemonstrated for oral strains of Actinomyces viscosus (1),but these structures appear to be very different from thefibrils studied here and would be classified as fimbriaeaccording to the nomenclature applied by Handley andco-workers (6). In gram-negative bacteria, different pili-associated adhesins may also be present simultaneously (9).Obviously such a situation will broaden the opportunities ofa bacterium to colonize a host.A comparison of the surface structures on negatively
stained cells of parent and mutant strains and the localizationof adhesins by IEM allow us to draw a schematic picture ofthe cell surface of S. salivarius K+ cells (Fig. 8). For avariety ofreasons, it is postulated that AgC is represented bythe 72-nm class of fibrils present on the cells and not by thelong 178-nm fibrils. First, protease-treated cells still carrythe long fibrils, have lost the short fibrils (6), and havesimultaneously lost the host-associated adhesive function(23). Second, mutant HB-V5, which has normal amounts ofwall-associated AgC, carries a 72-nm fibrillar fringe that issignificantly shorter than that observed on both the wild-typestrain HB and strain HB-7 (91 nm). Third, IEM showed thatAgC label did not extend beyond the outside edge formed bythe short fibrils. In addition, we show in an acconmpanyingpaper (27) that free, isolated AgC is only 87 nm long and isprotease sensitive (22). It is therefore probable that inaddition to losing the long fibrils, mutant HB-7 has also lostthe shorter 72-nm class of fibrils. This loss is masked by thecontinued presence of the longer 90-nm fibrils responsiblefor coaggregation. This hypothesis is supported by ruthe-nium red-stained sections of strain HB-7 organisms whichshow a marked reduction in density of the short fibrils(unpublished data) consistent with the loss both of longfibrils and of the short 72-nm class of fibrils. The nature andpossible function of the protease-resistant long (178-nm)fibrils and the reason for their simultaneous absence inmutants HB-7 and HB-V51 remain unknown.AgB is apparently represented by the longer type of short
fibrils (91 nm), since these are still present in mutant HB-7but not in strain HB-V5. This conclusion is supported by theIEM data, which suggest that AgB locates slightly moreperipherally than AgC. It was noted previously that coag-gregation reactions associated with AgB were less sensitiveto protease treatment than reactions associated with AgC(22). Although most fibrils are finally removed from the cellsby protease treatment, as judged by negative-stain electronmicroscopy, the short (72-nm) fibrils were lost more easilythan the 91-nm fibrils (unpublished data). Thus, even upon
prolonged protease treatment an amount of functionallyactive adhesin sufficient to cause the coaggregation reactionis still present on the cells.The observation that the sparsely distributed fibrils on
mutant HB-V51 are significantly shorter than those onparent strain HB-V5 could, in a similar way, indicate thepresence of yet another structural and functional class ofprotease-sensitive fibrils, obscured in both strains HB andHB-V5. A possible candidate for the 63-nm fibril class wouldbe AgD, since cell walls of mutant HB-V51 contain amountsof this antigen comparable to those found in wild-type strainHB (unpublished data). Alternatively, the 63-ntn fibrils mayrepresent structurally incomplete and functionally inactiveremnants of one of the other fibril classes. The isolation ofmutants specifically lacking AgD should permit the identifi-cation of this compound.
Fibrils morphologically similar to those found in S.salivarius K+ have been observed on a variety of other oralstreptococci (3, 7). Cell wall-associated protein antigens,some apparently involved in adhesion, are also present inthese bacteria (12, 14). It therefore seems not unlikely that asimilar mechanismn of carrying separate functions on special-ized fibrils has evolved in the other species as well. Such amechanism mnay facilitate the differential expression of ad-hesive fuhctions such as observed with S. salivarius HBunder different growth conditions (25). However, the recentreport by Fives-Taylor and Thompson (3) on a collection ofmutants of S. sanguis FW213 which were defective in cellsurface fibrils is in favor of a model in which activitiesmediated by different gene products may be associated withthe same fibrils.A second important finding is that the genetic changes
leading to the selective disappearance from the cell surfaceof either AgB or AgC seem to be expressed at differentbiochemical levels. Mutants defective in wall-associatedAgC accutnulated AgC reactivity intracellularly, whereas nosuch accumulation was observed with AgB. Mutants defec-tive in AgB, on the other hand, did not accumulate antigensin the cytoplasm. Unfortunately, very little is known aboutthe genetics and the mechanisms of synthesis, assembly, andexcretion of fibrillar proteins in gram-positive bacteria thatco'uld help to explain the observations. The mutation instrain HB-7 may have led to the blocking or absence of aputative membrane receptor--mediated secretory apparatus,as was recently suggested for the secretion of alkalinephosphatase in a strain of Bacillus licheniformis (19). Theaccumulation of intracellular alkaline phosphatase in anonsecretory mutant of this strain was demonstrated bycolloidal gold immnunolabeling, analogous to our observa-tions on S. salivarius. Mutants of this type could be veryhelpful in elucidating the complex mechanisms of fibrilsynthesis and assembly.
ACKNOWLEDGMENTS
We thank Alan Gibbs of the Department of Community Health,The Medical School, Manchester University, for the statisticalanalysis and Henny van der Mei, Department of Oral Biology,Groningen University, for assistance in preparing the specificantisera.
LITERATURE CITED1. Cisai, J. D., E. L. Barsumian, S. H. Cuvel, A. E. Vatter, A. L.
Sandberg, and R. P. Siraganian. 1981. Detection and localiza-tion of a lectin on Actinomyces viscosus T14V by monoclonalantibodies. J. Immunol. 127:1318-1322.
2. Ellen, R. P., and R. J. Gibbons. 1972. M protein-associated
J. BACTERIOL.
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

LOCALIZATION OF CELL SURFACE FIBRILLAR ANTIGENS
adherence of Streptococcus pyogenes to epithelial surfaces:prerequisite for virulence. Infect. Immun. 5:826-830.
3. Fives-Taylor, P. M., and D. W. Thompson. 1985. Surfaceproperties of Streptococcus sanguis FW213 mutants nonadher-ent to saliva-coated hydroxyapatite. Infect. Immun. 47:752-759.
4. Geuze, H. J., J. W. Slot, P. A. van der Ley, R. C. T. Scheffer,and J. M. Griffeth. 1981. Use of colloidal gold particles indouble-labeling immunoelectron microscopy of ultrathin frozentissue sections. J. Cell. Biol. 89:653-665.
5. Gibbons, R. J., J. van Houte, and W. F. Liljemark. 1972.Parameters that affect the adherence of Streptococcus salivariusto oral epithelial surfaces. J. Dent. Res. 51:424-435.
6. Handley, P. S., P. L. Carter, and J. Fielding. 1984. Streptococ-cus salivarius strains carry either fibrils or fimbriae on the cellsurface. J. Bacteriol. 157:64-72.
7. Handley, P. S., P. L. Carter, J. E. Wyatt, and L. M. Hesketh.1985. Surface structures (peritrichous fibrils and tufts of fibrils)found on Streptococcus sanguis strains may be related to theirability to coaggregate with other oral genera. Infect. Immun.47:217-227.
8. Johnson, R. H., W. A. Simpson, J. B. Dale, I. Ofek, and E. H.Beachey. 1980. Lipoteichoic acid-binding and biological proper-ties of T protein of group A Streptococcus. Infect. Immun.29:791-798.
9. Jones, G. W., and R. E. Isaacson. 1983. Proteinaceous bacterialadhesins and receptors. Crit. Rev. Microbiol. 10:229-260.
10. Kronvall, G., L. Bjorck, E. Myhre, and L. Wannamaker. 1979.Binding of aggregated 032-microglobulin, IgG, IgA and fibrinogento group A, C, and G streptococci with special reference tostreptococcal M protein, p. 74-76. In M. J. Parker (ed.),Pathogenic streptococci. Reedbooks Ltd., Chertsey, England.
11. Liljemark, W. F., and R. J. Gibbons. 1972. Proportional distri-bution and relative adherence of Streptococcus miteor (mitis) onvarious surfaces in the human oral cavity. Infect. Immun.6:852-859.
12. Nagata, K., M. Nakao, S. Shibata, S. Shizuhuishi, R. Nakamura,and A. Tsunemitsu. 1983. Purification and characterization ofgalactose-philic component present on the cell surface of Strep-tococcus sanguis ATCC 10557. J. Periodontol. 54:163-172.
13. Nalbandian, J., M. L. Freedman, J. M. Tanzer, and S. M.Lovelace. 1974. Ultrastructure of mutants of Streptococcusmutans, with reference to agglutination, adhesion, and extracel-lular polysaccharide. Infect. Immun. 10:1170-1179.
14. Russell, R. R. B. 1979. Wall-associated protein antigens ofStreptococcus mutans. J. Gen. Microbiol. 114:109-115.
15. Schilstra, M. J., J. W. Slot, P. H. van der Meide, G. Posthuma,
A. F. M. Cremers, and L. Bosch. 1984. Immunocytochemicallocalization of the elongation factor Tu in Escherichia coli cells.FEBS Lett. 165:175-179.
16. Sklavounou, A., and G. R. Germaine. 1980. Adherence of oralstreptococci to keratinized and nonkeratinized human oral epi-thelial cells. Infect. Immun. 27:686-689.
17. Slot, J. W., and H. J. Geuze. 1985. A new method of preparinggold probes for multiple-labeling cytochemistry. Eur. J. CellBiol. 38:87-93.
18. Swanson, J., K. C. Hsu, and E. C. Gotschlich. 1969. Electronmicroscopic studies on streptococci. I. M antigen. J. Exp. Med.130:1063-1091.
19. Tinglu, G., A. Ghosh, and B. K. Ghosh. 1984. Subcellularlocalization of alkaline phosphatase in Bacillus licheniformis749/c by immunoelectron microscopy with colloidal gold. J.Bacteriol. 159:668-677.
20. Tokuyasu, K. T. 1980. Immunochemistry on ultrathin frozensqctions. Histochem. J. 12:381-403.
21. Weerkamp, A. H. 1985. Coaggregation of Streptococcussalivarius with gram-negative oral bacteria: mechanism andecological significance, p. 177-183. In S. Mergenhagen and B.Rosan (ed.), Molecular basis for oral microbial adhesion. Amer-ican Society for Microbiology, Washington, D.C.
22. Weerkamp, A. H., and T. Jacobs. 1982. Cell wall-associatedprotein antigens of Streptococcus salivarius: purification, prop-erties, and function in adherence. Infect. Immun. 38:233-242.
23. Weerkamp, A. H., and B. C. McBride. 1980. Characterization ofthe adherence properties of Streptococcus salivarius. Infect.Immun. 29:459-468.
24. Weerkamp, A. H., and B. C. McBride. 1981. Identification of aStreptococcus salivarius cell wall component mediating coag-gregation with Veillonella alcalescens Vl. Infect. Immun.32:723-730.
25. Weerkamp, A. H., H. C. van der Mei, D. P. E. Engelen, andC. E. A. de Windt. 1984. Adhesion receptors (adhesins) of oralstreptococci, p. 85-97. In J. M. ten Cate, S. A. Leach, and J.Arends (ed.), Bacterial adhesion and preventive dentistry. IRLPress, Oxford, England.
26. Weerkamp, A. H., H. C. van der Mei, and R. S. Liem. 1984.Adhesive cell wall-associated glycoprotein of Streptococcussalivarius (K') is a cell surface fibril. FEMS Microbiol. Lett.23:163-166.
27. Weerkamp, A. H., H. C. van der Mei, and R. S. Liem. 1986.Structural properties of fibrillar proteins isolated from the cellsurface and cytoplasm of Streptococcus salivarius (K') cellsand nonadhesive mutants. J. Bacteriol. 165:756-762.
755VOL. 165, 1986
on June 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























