Embed Size (px)
Citation preview

PNAG in B. subtilis biofilms
1
Identification of Poly-N-acetyl Glucosamine as a Major Polysaccharide Component of the Bacil- 1 lus Subtilis Biofilm Matrix* 2
3 Damien Roux 1,‡, Colette Cywes-Bentley1, Yi-Fan Zhang1,2, Stephanie Pons1, Melissa Konkol3, 4
Daniel Kearns3, Dustin J. Little4,5, P. Lynne Howell4,5, David Skurnik1,§,¶, Gerald B. Pier1,§,¶ 5 6 From the 1Division of Infectious Diseases, Department of Medicine, Brigham and Women's Hos- 7
pital, Harvard Medical School, Boston, MA 02115, and 8 the 2Harvard School of Dental Medicine, Boston, MA 02115, and 9
the 3Department of Biology, Indiana University, 405A Simon Hall, Bloomington, IN 47405, and 10 the 4Department of Biochemistry, University of Toronto, Toronto, Ontario, M5S 1A8, Canada, 11
and 12 the 5Program in Molecular Structure & Function, Research Institute, The Hospital for Sick Chil- 13
dren, Toronto, Ontario, M5G 1X8, Canada 14 15
*Running title: PNAG in B. subtilis biofilms 16 17
‡Present address: INSERM, IAME, UMR 1137, F-75018 Paris, France; University of Paris Dide- 18 rot, Sorbonne Paris Cité, F-75018 Paris, France 19
§Co-senior authors 20 21 ¶To whom correspondence may be addressed: Dr. David Skurnik, Division of Infectious Diseases, 22
Brigham and Women’s Hospital, 181 Longwood Ave., Boston, MA 02115. 23 E-mail: [email protected] 24
¶ or Dr. Gerald B. Pier, Division of Infectious Diseases, Brigham and Women’s Hospital, 181 25 Longwood Ave., Boston, MA 02115. E-mail: [email protected] 26
27 Keywords: 28
bacteria; bacterial adhesion; bacterial genetics; carbohydrate biosynthesis; carbohydrate structure; 29 glycobiology; glycosyltransferase; protein expression; protein structure 30 31 32 Background: Bacillus subtilis is a model organ- 33 ism for analyzing bacterial biofilms but the car- 34 bohydrate components are undescribed. 35 Results: Genes in the epsHIJK locus needed for 36 biofilm formation encode proteins synthesizing 37 the conserved bacterial polysaccharide, poly-N- 38 acetyl glucosamine (PNAG). 39 Conclusion: PNAG is a major carbohydrate 40 component of B. subtilis biofilms. 41 Significance: PNAG production is essential for 42 formation of B. subtilis biofilms. 43 44 ABSTRACT 45
Bacillus subtilis is intensively studied as a 46 model organism for the development of bac- 47 terial biofilms or pellicles. A key component is 48 currently undefined exopolysaccharides 49 (EPS) produced from proteins encoded by 50 genes within the eps locus. Within this locus 51 are four genes, epsHIJK, known to be essen- 52 tial for pellicle formation. We show they en- 53 code proteins synthesizing the broadly ex- 54 pressed microbial carbohydrate, poly-N- 55
acetylglucosamine (PNAG). PNAG was pre- 56 sent in both pellicle and planktonic wild-type 57 B. subtilis cells and in strains with deletions in 58 the epsA, B, C, D, E, F, G, L, M, N or O genes, 59 but not in strains deleted for epsH, I, J or K. 60 Cloning of the B. subtilis epsH, I, J, or K genes 61 into Escherichia coli with in-frame deletions 62 in the PNAG biosynthetic genes pgaA, B, C, 63 or D, respectively, restored PNAG production 64 in E. coli. Cloning the entire B. subtilis 65 epsHIJK locus into pga-deleted E. coli, 66 Klebsiella pneumoniae or alginate-negative 67 Pseudomonas aeruginosa restored or con- 68 ferred PNAG production. Bioinformatic and 69 structural predictions of the EpsHIJK pro- 70 teins suggest EpsH and EpsJ are glycosyl- 71 transferases (GT) with a GT-A fold, EpsI is a 72 GT with a GT-B fold, and EpsK is an α- 73 helical membrane transporter. B. subtilis, E. 74 coli, and pga-deleted E. coli carrying the 75 epsHIJK genes on a plasmid, were all suscep- 76 tible to opsonic killing by antibodies to 77 PNAG. The immunochemical and genetic da- 78
http://www.jbc.org/cgi/doi/10.1074/jbc.M115.648709The latest version is at JBC Papers in Press. Published on June 15, 2015 as Manuscript M115.648709
Copyright 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
2
ta identify the genes and proteins used by B. 79 subtilis to produce PNAG as a significant car- 80 bohydrate factor essential for pellicle for- 81 mation. 82 83
84 Many microbial organisms produce bio- 85
films, structurally complex multicellular com- 86 munities inside an extracellular matrix of varia- 87 ble factors that can include exopolysaccharides 88 (EPS),4 proteins and nucleic acids (1,2). Bio- 89 films are probably the most common structures 90 for microbial communities, as this state protects 91 against many environmental stresses such as an- 92 timicrobial factors (3). Biofilms are also in- 93 volved in pathogenesis of many infectious dis- 94 eases (1,4,5). Polysaccharides are often promi- 95 nent components of biofilms, but, like many fac- 96 tors in this structure, make a variable contribu- 97 tion depending on the microbial species, strain, 98 growth conditions and overall environment. In- 99 terestingly, numerous species have developed 100 diverse metabolic pathways for production of 101 EPS, alluding to the possibility that these sys- 102 tems have evolved independently. A common 103 EPS associated with microbial biofilm formation 104 is a polymeric β-1,6-linked-N-acetylglucosamine 105 (PNAG) structure which is highly conserved and 106 expressed by a range of bacterial, fungal and 107 protozoan microorganisms (6,7). PNAG was 108 first isolated and characterized from Staphylo- 109 coccus epidermidis (8) where it was referred to 110 as the polysaccharide intercellular adhesin 111 (PIA), then later shown to be produced by S. au- 112 reus (9,10). In these two species, PNAG is syn- 113 thesized by proteins encoded by 4 genes in the 114 ica operon (9-11), and ica-deleted, PNAG- 115 deficient strains were unable to produce biofilms 116 (9,11). However, PNAG-independent biofilm 117 formation in some Staphylococcal strains has al- 118 so been described (12). In general, depending on 119 the strain and culture conditions, PNAG is often 120 necessary, but not sufficient or essential, for bio- 121 film formation. 122
PNAG synthesis also occurs in various 123 Gram-negative organisms including Escherichia 124 coli (13), Acinetobacter baumannii (14) and 125 Burkholderia spp.(15). In these organisms 126 PNAG is synthesized by 4 proteins encoded by 127 genes in the pga operon. More recently, Cywes- 128 Bentley et al. (7) showed that a much wider 129 spectrum of microbes can synthesize PNAG, in- 130 cluding many human bacterial pathogens such as 131 Streptococcus pneumoniae, S. pyogenes, S. aga- 132 lactiae, Neisseria gonorrhoeae, N. meningitidis, 133
non-typable Haemophilus influenzae and Myco- 134 bacterium tuberculosis and eukaryotic organ- 135 isms such as fungal pathogens, Trichomonas 136 vaginalis and murine and human Plasmodia spp. 137 that cause malaria. Among many of these patho- 138 gens there are no readily identifiable genes ho- 139 mologous to those in the ica or pga loci. None- 140 theless, the broad conservation of PNAG synthe- 141 sis implies an important role in microbial biolo- 142 gy, perhaps related to growth phases involving 143 aggregation of microbes living in different envi- 144 ronments or protection from anti-microbial fac- 145 tors. 146
Bacillus subtilis is a spore-forming Gram- 147 positive bacillus wherein certain strains grown 148 under specific conditions develop complex bio- 149 films (16-18). A large amount of knowledge has 150 been accumulated that explains how the produc- 151 tion and composition of the B. subtilis biofilm is 152 regulated (17) although many aspects of this 153 process are not fully elucidated. Full biofilm 154 formation by some strains of B. subtilis such as 155 NCIB 3610 (3610) is dependent on 15 proteins 156 encoded by genes in the eps locus (epsA-epsO) 157 that are associated with the carbohydrate content 158 and complexity of a surface pellicle (16) in- 159 volved in the overall biofilm structure. The 160 composition and structure of polysaccharides 161 synthesized by the proteins within this complex 162 are not well described, but mutations in most of 163 the genes within the B. subtilis eps cluster lead 164 to loss of biofilm formation (19). Some genes, 165 such as epsE, have dual functions, affecting both 166 polysaccharide synthesis and flagella-based mo- 167 tility (20,21). 168
Given the ubiquity of PNAG synthesis 169 among a range of microbial organisms we eval- 170 uated B. subtilis biofilms for PNAG production 171 and further examined the B. subtilis eps locus for 172 genes potentially encoding PNAG biosynthetic 173 proteins. Both biofilm and planktonic cells pro- 174 duced PNAG, and within the available annotated 175 genome, we identified four genes, epsH-K, as 176 potentially being responsible for PNAG synthe- 177 sis. These genes are predicted to encode two 178 glycosyltransferases (GT) (epsH, epsJ), separat- 179 ed by another GT with potential EPS modifying 180 enzymatic activity (epsI), and a trans- 181 porter/facilitator of synthesis (epsK). Cloning of 182 the epsH-K genes into PNAG-deficient E. coli or 183 Klebsiella pneumoniae (Δpga), or alginate- 184 negative Pseudomonas aeruginosa, lead to syn- 185 thesis of a polymer immunochemically equiva- 186 lent to PNAG. Each of the four B. subtilis epsH, 187 I, J or K genes could individually trans- 188
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
3
complement E. coli strains deleted for the pgaA, 189 B, C or D genes, respectively. Also the epsH and 190 epsJ genes could partially complement E. coli 191 strains deleted for the pgaC or pgaA genes, re- 192 spectively. Extracts of both WT B. subtilis and 193 E. coli (Δpga) complemented with B. subtilis ep- 194 sHIJK contained PNAG immunoreactive, hex- 195 osamine-containing material that was destroyed 196 by treatment with both the PNAG-degrading en- 197 zyme Dispersin B (22) and by sodium periodate, 198 which can only hydrolyze polymeric hex- 199 osamines in a 1-6-linkage. Synthesis of PNAG 200 in E. coli from the epsH-K genes resulted in sus- 201 ceptibility of cells to killing in an opsonophago- 202 cytic assay using antibodies specific to PNAG, 203 indicative of functional conservation of PNAG 204 properties when B. subtilis gene products direct 205 synthesis of PNAG in E. coli. Overall, we identi- 206 fy the presence of PNAG in B. subtilis biofilms 207 and the genes within the eps locus that encoded 208 proteins that synthesize PNAG. B. subtilis lack- 209 ing epsH, I, J or K genes are unable to form bio- 210 films, indicating PNAG is essential for bio- 211 film/pellicle formation by this organism. 212
213 Experimental procedures 214
Bacterial Strains and plasmids 215 Bacterial strains (B. subtilis, E. coli, K. 216
pneumoniae and P. aeruginosa) and plasmids 217 used in this study are listed in Table 1. B. subtilis 218 was grown on minimal-medium glucose (MMG) 219 agar (23) or lysogeny broth (LB) agar (24) for 3- 220 5 days at RT, whereas the other bacterial strains 221 were grown overnight at 37 oC in LB or on LB 222 agar. 223 BLAST (basic local alignment search tool) 224 and structural analysis 225
Sequences of eps genes in B. subtilis strain 226 168 were obtained from the NCBI website (Ref- 227 erence Sequence: NZ_CM000487.1). Amino ac- 228 id sequences from four eps genes (epsH, epsI, 229 epsJ, epsK) were mapped on E. coli and S. aure- 230 us sequences using the basic local alignment 231 search tool (BLAST) from the NCBI website. 232 Amino acid sequences from the four pga genes 233 of E. coli IHE3034 were similarly mapped on S. 234 aureus sequences. The amino acid sequences of 235 B. subtilis eps H, I, J or K were analyzed by the 236 Protein Homology/analogY Recognition Engine 237 version 2 (PHYRE2) (25) for structural predic- 238 tions as to their function. 239 Strain construction in Gram-negative 240 bacteria 241
Deletions of individual pga genes in a clini- 242 cal isolate of an E. coli K1 strain from a child 243
with meningitis, designated E11 and kindly pro- 244 vided by Kwan Sik-Kim of Johns Hopkins Uni- 245 versity School of Medicine, and deletion of the 246 entire 4-gene pga locus in both E. coli E11 and 247 K. pneumoniae NTUH-K2044 (26) were con- 248 structed as previously described (27). Briefly, a 249 kanamycin resistance cassette flanked by FLP 250 recombinase recognition target sites and homol- 251 ogy arms to replace the DNA segments of inter- 252 est in-frame were generated by PCR with dele- 253 tion primers (Supplemental Table 1). Recombi- 254 nation within the targeted chromosomal se- 255 quences was mediated by the Red recombinase 256 encoded on pRdET (28), resulting in the re- 257 placement of the targeted sequence with a kan- 258 amycin-resistant cassette; all allelic replace- 259 ments were confirmed by PCR. Subsequently, 260 the kanamycin marker was removed, using the 261 FLP expression plasmid pCP20 (29). 262 Strain construction in B. subtilis 263
All constructs were either directly integrated 264 and resolved in the competent strain DK1042 265 (comIQ12L) or integrated in the competent strain 266 DS2569 (ΔpBS32), transferred to the 3610 267 background using SPP1-mediated generalized 268 phage transduction, and resolved (30). All 269 strains and plasmids used in this study are listed 270 in Table 1. Primer sequences are delineated in 271 Supplemental Table 1. 272
In-frame deletions. To generate the in 273 frame marker-less deletion constructs, each 274 plasmid was introduced by single cross-over in- 275 tegration at the restrictive temperature for plas- 276 mid replication (37 °C) using mls resistance as a 277 selection. To evict the plasmid, the strain was 278 incubated in 3 ml LB broth at a permissive tem- 279 perature for plasmid replication (22 °C) for 14 h. 280 Cells were then serially diluted and plated on LB 281 agar at 37 °C. Individual colonies were patched 282 onto LB agar plates and LB agar plates contain- 283 ing mls to identify mls sensitive colonies that 284 had evicted the plasmid. Chromosomal DNA 285 from colonies that had excised the plasmid was 286 purified and screened by PCR to determine 287 which isolate had retained the deletion allele. 288
To generate the ΔepsA in-frame marker-less 289 deletion construct, the region upstream of epsA 290 was amplified by PCR using the primer pair 291 3971/3972, digested with EcoRI and XhoI, and 292 the region downstream of epsA was amplified by 293 PCR using the primer pair 3973/3974 and di- 294 gested with XhoI and BamHI. The two frag- 295 ments were then simultaneously ligated into the 296 EcoRI and BamHI sites of pMiniMAD that car- 297
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
4
ries a temperature sensitive origin of replication 298 and an erythromycin resistance cassette to gen- 299 erate pMP201. 300
The same method was used to generate the 301 other eps gene in-frame marker-less deletion 302 constructs. Plasmids are detailed in Table 1 and 303 primers in Supplemental Table 1. 304 Genetic complementation 305 Wild-type, alginate over-producing mucoid 306 P. aeruginosa FRD1 and an alginate-deficient 307 strain (P. aeruginosa (Tn501::algF)) due to a 308 polar effect of the Tn501 insert in the algF gene 309 on alginate synthesis (31) were used to express 310 the B. subtilis epsHIJK locus.(29). 311
For complementation of the various muta- 312 tions, pga or eps genes from E. coli K1 strain 313 E11 or B. subtilis DS991, respectively, were 314 amplified by PCR with complementation pri- 315 mers (see Table 2) using chromosomal DNA as 316 a template. PCR products were cloned into the 317 broad-host-range vector pUCP18Tc as XbaI/SbfI 318 or SacI/XbaI fragments, and transformed into E. 319 coli Sm10 with selection on LB agar containing 320 tetracycline (10 mg/L). All constructs were con- 321 firmed by sequencing. E. coli and K. pneumoni- 322 ae strains were complemented with individual 323 plasmids by electroporation and selection on LB 324 agar supplemented with tetracycline (10 mg/L). 325 For P. aeruginosa, complementation plasmids 326 were conjugated from E. coli Sm10 as previous- 327 ly described (32). The complemented P. aeru- 328 ginosa strains were selected on LB agar contain- 329 ing irgasan (25 mg/L) and tetracycline (75 330 mg/L). 331 Immunochemical detection of PNAG on 332 microbial cells 333
To detect PNAG in B. subtilis biofilms, 334 strain DS991 was inoculated into 1 ml of MMG 335 broth in 12 well tissue culture plate and left for 1 336 week at room temperature (RT) in a humidified 337 environment. Bacterial biofilms were removed 338 with as little remaining media as possible and 339 placed onto glass slides in a demarcated well. 340 Samples were allowed to air dry before fixing 341 for 5 min at RT with ice-cold methanol, which 342 was removed by gently tipping the slide onto its 343 side and absorbing residual methanol with a tis- 344 sue. 345
Planktonic B. subtilis strains were grown in 346 MMG media at 25 oC for 3-10 days at RT 347 whereas other bacterial species were grown on 348 LB agar overnight at 37 ˚C then held at RT for 349 48 h to promote PNAG expression. Strains har- 350 boring one of the pUC vectors were grown on 351 LB agar plates with tetracycline (10 mg/L) under 352
the same conditions with protection from light. 353 Microbial samples were suspended in PBS, then 354 spotted onto microscope slides, air-dried, and 355 fixed for 1 min at RT with ice-cold methanol. 356 After washing, slides were reacted with control 357 or PNAG-specific MAbs directly conjugated to 358 Alexa Fluor 488 (AF488) at 5.2 µg/ml along 359 with nucleic acid stain, Syto 83 (Molecular 360 Probes) (7). After 2 h at RT or overnight at 4 °C, 361 slides were washed and evaluated by confocal 362 microscopy. For enzymatic and periodate treat- 363 ments, samples fixed to slides were incubated in 364 Tris-buffered saline (pH 6.4) containing either 365 50 µg Dispersin B/ml (digests PNAG) or 50 µg 366 chitinase/ml (no effect on PNAG) overnight at 367 37 °C or in 0.4 M periodate (destroys PNAG) for 368 2 h at 37 °C in a humidified environment. After 369 washing, cells were treated with the AF488 di- 370 rectly conjugated MAbs. 371 Extraction and detection of PNAG 372
B. subtilis DS991 and E. coli Δpga 373 (pUCP18Tc-epsHIJK) were grown as described 374 above on MMG agar or LB agar plates, respec- 375 tively, and cells used for extraction and immuno- 376 logic detection of PNAG by a slot blot as previ- 377 ously described (14) and chemical detection of 378 hexosamines as described (33). 379
B. subtilis DS991, B. subtilis DK2055 380 ΔepsK, E. coli WT and E. coli Δpga as a nega- 381 tive control were tested for intracellular PNAG 382 by a direct-binding ELISA using lysates of these 383 cells prepared by sonication. Briefly, cells from 384 blood agar plates grown overnight at 37 oC were 385 suspended in normal saline, washed and then 386 suspended in Tris-buffered saline containing 387 12.5 µg/ml Dispersin B to remove surface 388 PNAG during a 24 h 37 oC incubation step. Af- 389 ter washing, bacterial cells were suspended in 390 0.04 M phosphate buffer, pH 7.2 at an optical 391 density at 650 nm (OD650 nm) of 0.8, sonicated 392 with 10 sec bursts, 10 times, insoluble debris 393 removed by centrifugation and dilutions of the 394 lysate used to directly sensitize Immulon 4 395 ELISA plates. A standard curve was generated 396 using purified PNAG(0.015-0.1 µg/ml) to sensi- 397 tize ELISA wells in duplicate and samples 398 probed with 10 µg/ml MAb F598 followed by an 399 alkaline-phosphatase-conjugated goat antibody 400 to human IgG. After assay development with the 401 p-nitrophenyl phosphate substrate, A405 nm read- 402 ings were obtained, a standard curve of the puri- 403 fied PNAG concentration versus A405 nm calcu- 404 lated by linear regression and the amount of 405 PNAG in bacterial cell lysates calculated after 406 subtracting out the background from the nega- 407
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
5
tive control. Samples of cells taken before and 408 after Dispersin B treatment were probed as de- 409 scribed above for immunochemical detection of 410 PNAG n microbial cells. 411 Opsonophagocytic assay 412
Opsonophagocytic killing (OPK) of B. sub- 413 tilis DS991, E. coli WT, E. coli (Δpga) and E. 414 coli (Δpga) (pUCP18Tc-epsHIJK) was carried 415 out as described previously (34) except differen- 416 tiated human promyelocytic HL60 cells were 417 used as the phagocyte source (35). The percent 418 killing mediated by antibodies in immune sera 419 raised to a conjugate of nine residues of β-1-6- 420 linked glucosamine and tetanus toxoid (TT; 421 9GlcNH2-TT vaccine) (36) was calculated by 422 dividing the colony-forming units (cfu) in the 423 test sera by those in the corresponding dilution 424 of the non-immune control serum. 425 426 RESULTS 427 Detection of PNAG in B. subtilis biofilms/ 428 pellicles 429
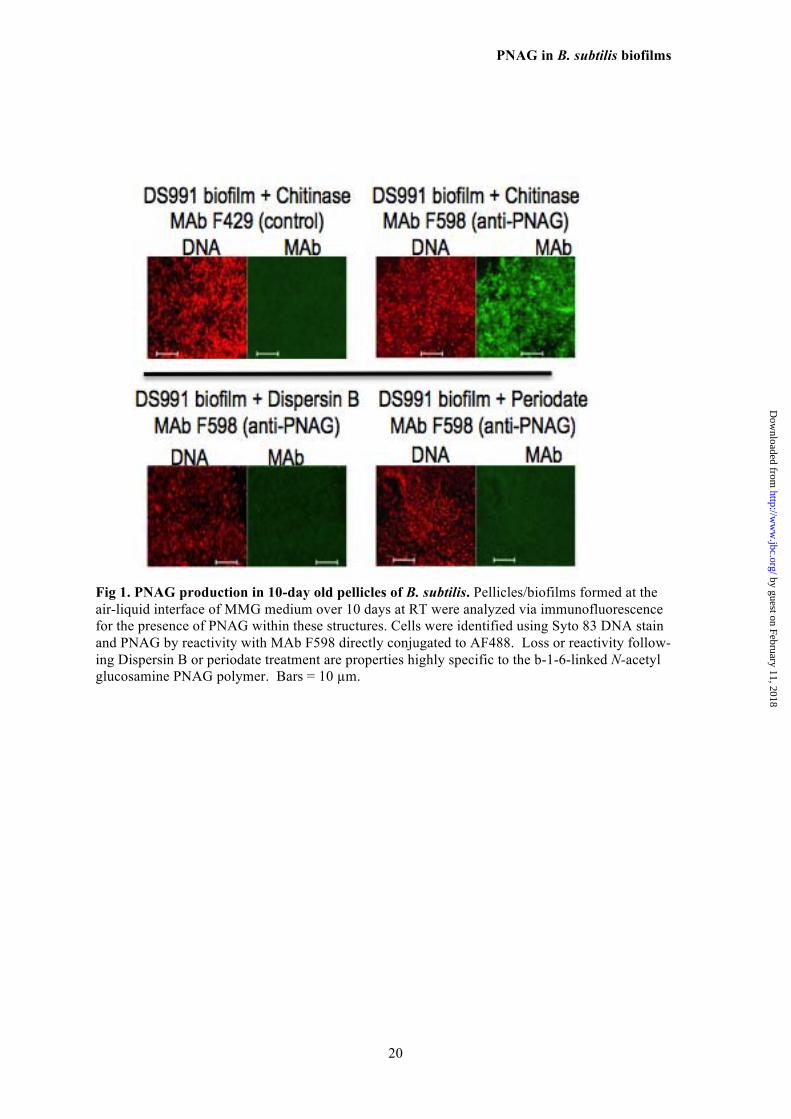
B. subtilis pellicles formed over 10 days of 430 growth at the air-liquid interface in MMG medi- 431 um were reacted with control MAb F429 or 432 MAb F598 to PNAG, both directly conjugated to 433 AF488 and visualized for immunofluorescence 434 by confocal microscopy. Bacilli embedded in a 435 strongly immunoreactive matrix of PNAG were 436 readily observed, and binding to MAb F598 was 437 lost after treating the B. subtilis biofilms with the 438 PNAG-degrading enzyme Dispersin B or with 439 PNAG-hydrolyzing sodium periodate (Fig. 1). 440 Expression of PNAG by WT and eps- 441 mutant B. subtilis 442
Planktonic cells of B. subtilis that overpro- 443 duced EPS (due to mutation of the master re- 444 pressor SinR) and also defective for the biofilm 445 organizing protein TasA (DS991) were also pos- 446 itive for PNAG expression by immunofluores- 447 cence microscopy (Fig. 2A) and immunoreactiv- 448 ity was lost following treatment with Dispersin 449 B or periodate (Fig. 2A). When PNAG produc- 450 tion was evaluated in B. subtilis strains with in- 451 frame deletions in all 15 identified eps genes 452 (Table 1), most of which are unable to form bio- 453 films, only disruption of the epsH, I, J and K 454 genes led to loss of PNAG production (Fig. 2B). 455 The epsH and epsJ genes are annotated as puta- 456 tive GTs and show similarity to proteins encod- 457 ed by S. aureus icaA and E. coli pgaC (Table 2). 458 The epsI gene is currently annotated as a puta- 459 tive polysaccharide pyruvyl transferase (Table 460 2) but as shown below, its structural predictions 461
suggest other functions. The epsK gene is cur- 462 rently annotated as a putative extracellular ma- 463 trix component exporter similar to the wzx pro- 464 teins in E. coli (Table 3). Two other eps genes 465 (epsE and epsF) are annotated as putative GTs 466 but deletions in these genes did not lead to loss 467 of PNAG production (Fig. 2B). Similarly, an 468 epsG deletion mutant, known to be deficient in 469 biofilm production like the epsH mutant (17), 470 was still able to produce PNAG (Fig. 2B). As 471 with other microbial species, PNAG production 472 appears to be necessary, but not sufficient, for 473 full biofilm formation in B. subtilis. 474 Bioinformatic analysis of B. subtilis 475 PNAG biosynthetic eps genes and proteins 476
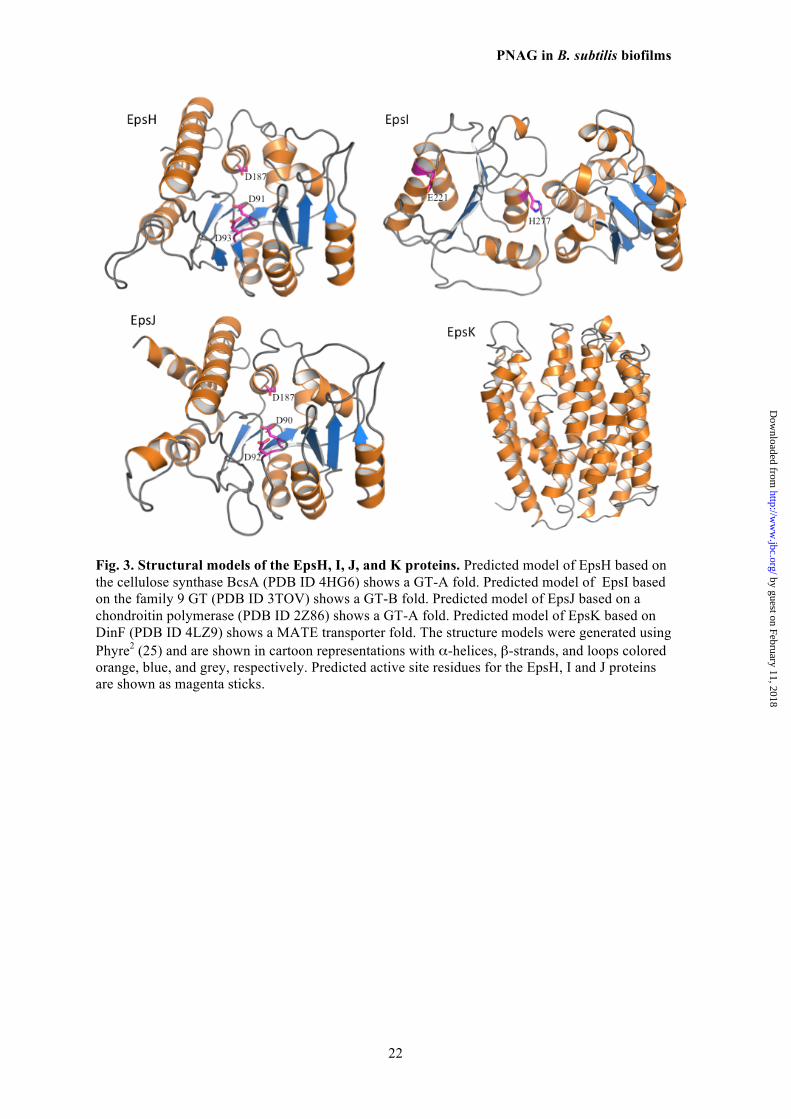
The above results, along with bioinformatic 477 analysis of the B. subtilis eps locus, indicated the 478 genes required for PNAG production, epsH, I, J 479 and K, could comprise a four-gene locus some- 480 what similar to the S. aureus icaADBC and E. 481 coli pgaABCD loci encoding the PNAG biosyn- 482 thetic proteins in these two organisms (Table 3) 483 (11,13). To obtain a more precise idea of the 484 functions of the EpsH, I, J and K proteins, as 485 well as other proteins within the eps locus, the 486 predicted protein structures were analyzed using 487 the PHYRE2 server (Fig. 3) (25). Overall, the 488 entire eps locus contains genes encoding pro- 489 teins known to be involved in polysaccharide 490 synthesis using either the Wzx/Wzy or ABC- 491 transporter pathways for lipopolysaccharides, 492 capsules, colonic acid, and S-layer components 493 (37-39). Both EpsH and EpsJ have nearly identi- 494 cal predicted structures (Fig. 3) with homology 495 to the GT domains of IcaA and PgaC proteins 496 used in S. aureus and E. coli for PNAG synthe- 497 sis, respectively, (13,40) as well as the BcsA 498 protein used for cellulose (β-1-4-linked glu- 499 cose) synthesis (41). With 60-70% sequence 500 coverage and 15-22% sequence identity, EpsH 501 and EpsJ are predicted with 100% confidence to 502 be UDP-N-acetylglucosamine transferases with a 503 GT-A fold. They are not predicted to have any 504 transmembrane domains, and therefore would be 505 unable to translocate the synthesized polymer 506 across the membrane. 507
BLAST analysis showed that EpsK belongs 508 to the polysaccharide biosynthesis protein fami- 509 ly, pfam01943. Members of this family are inte- 510 gral membrane proteins that encode for multi an- 511 timicrobial extrusion protein (MATE)-like 512 transporters, such as the O-antigen flippase Wzx. 513 EpsK was predicted to be an α-helical mem- 514 brane transporter with 14 transmembrane helices 515 using Phyre2 and various transmembrane do- 516
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
6
main prediction servers, respectively (42) (Fig. 517 3). Phyre2 predicts EpsK to be structurally simi- 518 lar to DinF, a member of the MATE family, 519 covering 84% of the sequence with 10-12% 520 amino acid identity and 100% confidence of the 521 fold prediction. 522
The analysis of the EpsI protein (Fig. 3) in- 523 dicated it was not related structurally to the 524 PNAG deacetylases IcaB or Pga (43,44), or any 525 members of the family 4 carbohydrate esterases 526 (deacetylases), but rather is predicted to be a cy- 527 toplasmic GT with homology to genes in lipo- 528 polysaccharide biosynthesis. There were no pre- 529 dicted signal sequences or transmembrane heli- 530 ces but almost complete coverage (80-90%) of 531 the EpsI sequence with 95% confidence in fold 532 prediction to known lipopolysaccharide transfer- 533 ases. 534 Functional equivalence of B. subtilis 535 EpsH, I, J and K proteins to E. coli PNAG 536 biosynthetic proteins 537
In order to determine if each of these four B. 538 subtilis eps genes encoded proteins could replace 539 those encoded by the pga operon, we construct- 540 ed individual, in-frame mutations in each of the 541 four E. coli pga genes, pgaA, B, C, or D, and 542 complemented each of these different mutants 543 with an eps-related gene judged to most likely 544 encode a protein of potential similar function. As 545 controls, we also complemented some of the E. 546 coli mutants with a mismatched gene from B. 547 subtilis. The WT E. coli strain produced PNAG 548 and deletion of any of the four individual pga 549 genes abolished PNAG production (Table 3). 550 Complementation with an empty vector did not 551 restore the phenotype in any strain with an in- 552 frame pga mutation. Complementation of the E. 553 coli-pga-mutant strains with individual clones of 554 B. subtilis epsH, epsI, epsJ or epsK resulted in 555 restoration of PNAG production in each E. coli 556 mutant strain with the gene from the B. subtilis 557 locus judged most likely to be a functional 558 equivalent (Table 3). Interestingly, we found 559 that PNAG production in the E. coli ∆pgaC 560 strain could be restored by complementation 561 with either the B. subtilis epsJ gene and, to a 562 lesser extent, the epsH gene (Table 3). Similar- 563 ly, we could achieve phenotypic complementa- 564 tion of PNAG production in the E. coli ΔpgaA 565 mutant strain most strongly with the B. subtilis 566 epsH gene and, to a lesser extent, the epsJ gene. 567 The E. coli pgaB mutant was efficiently com- 568 plemented with the B. subtilis epsI gene and the 569 E. coli pgaD mutant was complemented with the 570 B. subtilis epsK gene (Table 3). Cloning of the 571
B. subtilis epsI and epsK genes into the E. coli 572 ΔpgaA mutant did not result in PNAG produc- 573 tion. Finally, when the entire pga locus was de- 574 leted from either E. coli or K. pneumoniae, the 575 loss of PNAG production could be restored in 576 both strains by complementation with the entire 577 B subtilis epsHIJK locus in pUCP18Tc-epsHIJK 578 (Table 3). 579
We also analyzed the B. subtilis DS991 WT 580 and ΔepsK strains as well as WT E. coli for the 581 presence of intracellular PNAG. WT E. coli had 582 a low, but detectable, level of intracellular 583 PNAG (Table 4). Both WT and ΔepsK B. sub- 584 tilis had detectable intracellular PNAG (Table 585 4), with the B. subtilis ΔepsK strain having about 586 60% more intracellular PNAG, indicating that in 587 the absence of the predicted EpsK transporter, 588 there was accumulation of the PNAG polysac- 589 charide inside the B. subtilis ΔepsK cells. 590
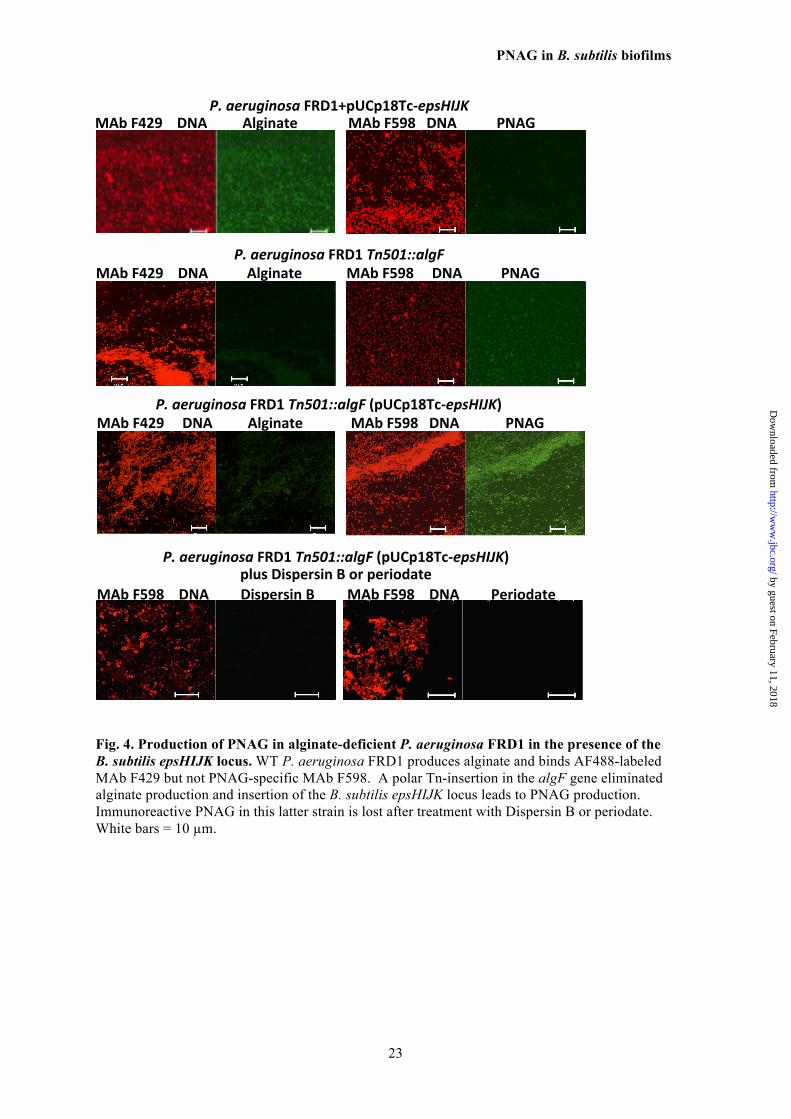
Finally, we found that the predicted MATE 591 function of the B. subtilis EpsK protein had 592 similarity to PelG and PslK in P. aeruginosa 593 used for the synthesis of the PEL and PSL poly- 594 saccharides, respectively. P. aeruginosa, unlike 595 E. coli and K. pneumoniae, does not normally 596 synthesize PNAG, so we introduced the 597 pUCP18Tc-epsHIJK plasmid into WT and algF- 598 interrupted (Tn::algF) P. aeruginosa strain 599 FRD1 to ascertain if PNAG could be synthe- 600 sized. We did not detect PNAG synthesis in ei- 601 ther the WT P. aeruginosa strain or the strain 602 carrying pUCP18Tc-epsHIJK, but did show al- 603 ginate production by virtue of binding of MAb 604 F429 to these bacterial cells (Fig. 4). We hy- 605 pothesized that it might be problematic to pro- 606 duce or detect low level PNAG production in 607 WT FRD1 P. aeruginosa if this organism is ex- 608 pressing both the positively-charged PNAG and 609 negatively-charged alginate in the same cell. 610 We thus cloned pUCP18Tc-epsHIJK into P. ae- 611 ruginosa FRD1 (Tn501::algF) wherein the Tn 612 insertion has a polar effect on the alginate bio- 613 synthetic locus, leading to loss of alginate pro- 614 duction, as verified by loss of binding of MAb 615 F429 to cells of this strain (Fig. 4). Addition of 616 the pUCP18Tc-epsHIJK plasmid resulted in ex- 617 pression of PNAG on the recombinant P. aeru- 618 ginosa FRD1 (Tn501::algF) cells as detected by 619 immunofluorescence analysis of binding of 620 MAb F598 to PNAG. This binding was lost af- 621 ter treatment of the recombinant P. aeruginosa 622 strains with Dispersin B and periodate, indica- 623 tive of PNAG synthesis in alginate-negative P. 624
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
7
aeruginosa by proteins encoded by the B. sub- 625 tilis epsHIJK genes. 626 Detection of PNAG by WT and recombi- 627 nant B. subtilis and E. coli strains 628
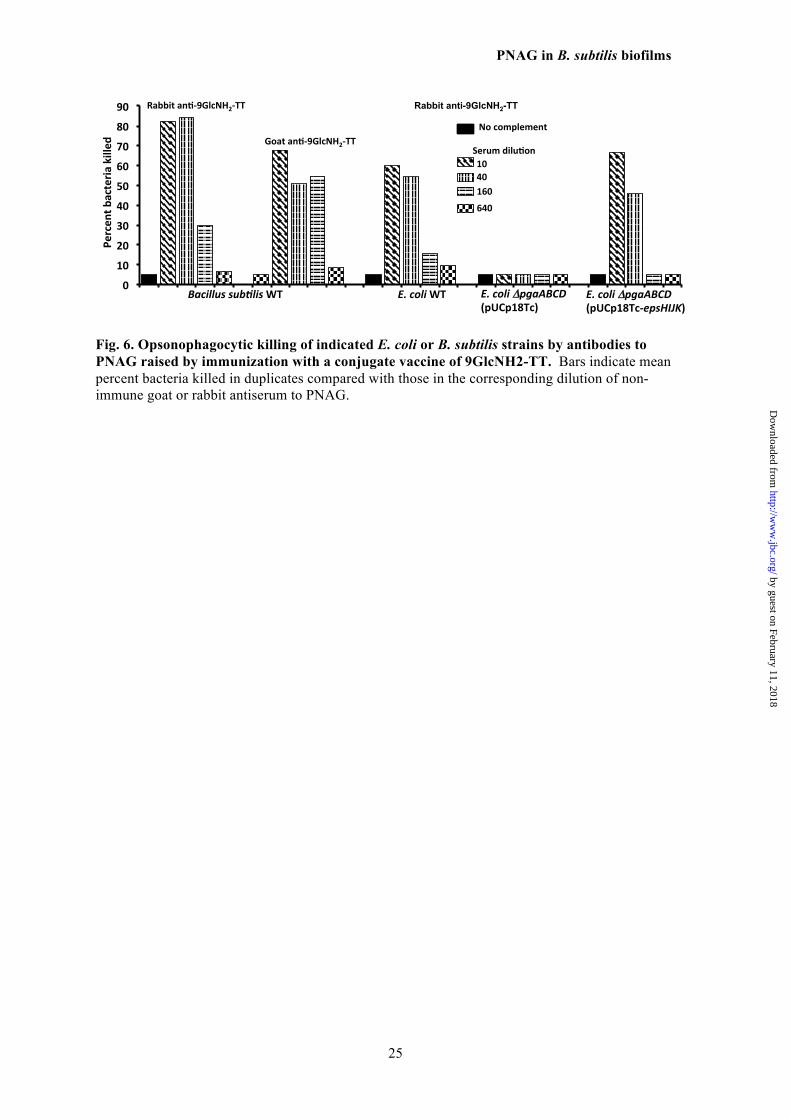
To confirm that PNAG was produced by 629 WT B. subtilis but not B. subtilis ΔepsH or 630 ΔepsJ, and by WT E. coli E11 and E. coli E11 631 (Δpga) (pUCP18Tc-epsHIJK) but not E. coli 632 (Δpga), we extracted surface material from cells 633 as described (14) and used slot-blots for immu- 634 nologic detection of extracted antigen. As shown 635 in Fig. 5, WT B. subtilis as well as WT E. coli 636 E11 produced immunoreactive PNAG while the 637 B. subtilis ΔepsH and ΔepsJ strains did not, nor 638 did E. coli (Δpga). Complementation of the 639 PNAG-deficient E. coli (Δpga) strain with the B. 640 subtilis epsHIJK genes restored detectable 641 PNAG production. Confirmation that the immu- 642 noreactive material was likely PNAG was ob- 643 tained by treating extracts with Dispersin B or 644 periodate, which both destroy PNAG. Such 645 treated materials had no reactivity with the MAb 646 to PNAG in a slot-blot assay (Fig 5). Finally, 647 chemical analysis of the extracts indicated hex- 648 osamine was only detectable in the PNAG- 649 positive extracts. Attempts to further purify 650 PNAG to a degree sufficient for analysis by 651 NMR were unsuccessful. This is consistent with 652 almost all prior publications indicating that 653 methods have not yet been developed to purify 654 PNAG sufficiently for NMR analysis in the ab- 655 sence of hyper-expression of the biosynthetic 656 proteins in organisms like S. aureus, E. coli or A. 657 baumannii (13,14,45,46). In organisms such as 658 S. epidermidis (8) and Vibrio parahemolyticus 659 (47), isolation of PNAG-related, small- 660 molecular weight fragments has been achieved 661 only by use of natural hyper-producers of bio- 662 films as sources of the initial extracts. 663 Opsonic killing mediated by antibody to 664 PNAG 665
The susceptibility of the B. subtilis and E. 666 coli strains to OPK was tested in an assay using 667 antibodies raised in either a rabbit or goat to the 668 9GlcNH2-TT vaccine (36). Antibodies in both of 669 these antisera readily mediated OPK of WT B. 670 subtilis and WT E. coli (Fig. 6). Deletion of the 671 pga locus in E. coli resulted in no effect of anti- 672 body to PNAG on cell survival in an OPK assay, 673 whereas introduction of the pUCP18Tc-epsHIJK 674 plasmid into pga-deleted E. coli restored the 675 susceptibility to OPK. Thus, B. subtilis EpsH, I, 676 J, and K proteins produced an antigen in E. coli 677
functionally equivalent to native PNAG from 678 this organism. 679
680 DISCUSSION 681
B. subtilis has served as one of the major 682 model organisms for scrutinizing biofilm for- 683 mation and multi-cellular activity in bacterial 684 communities (18,48). Many of the factors, genes 685 and conditions needed for producing and modu- 686 lating formation of biofilms by this organism are 687 well studied (49). Components of the B. subtilis 688 biofilm include EPS and polymeric substances 689 such as poly–DL-glutamic acid and proteins in- 690 cluding TapA, TasA and BslA (48). However, 691 the chemistry of the biofilm EPS constituents are 692 not defined, and no definitive polysaccharide 693 structures have previously been identified. Here 694 we found that within the 15 gene eps cluster of 695 B. subtilis there was a 4-gene locus encompass- 696 ing epsHIJK that encodes proteins that can syn- 697 thesize either the conserved bacterial surface 698 polysaccharide PNAG, a common component of 699 microbial biofilms, or an antigenically cross- 700 reactive material. In the B. subtilis biofilm, 701 PNAG, or a related structure, likely serves as a 702 scaffold as well as an anchoring substrate for the 703 other components in the biofilm, which requires 704 gene products from the eps locus other than ep- 705 sHIJK for full matrix formation, as many of the 706 eps gene products are needed to establish the 707 biofilm phenotype (48). 708
To synthesize PNAG using the B. subtilis 709 EpsH, I, J and K proteins we speculate that 710 EpsH is an undecaprenyl (UnDP) priming trans- 711 ferase which makes UnDP-3-O-acyl N- 712 acetylglucosamine. EpsI could either be modify- 713 ing this first GlcNAc or possibly adding on an- 714 other sugar monomer and also providing a 715 deacetylase function. EpsJ is potentially the 716 poly-GlcNAc transferase that is needed for 717 long chain extension to the lipid linker of the 718 UDP-N-acetylglucosamine precursor to synthe- 719 size the PNAG-like molecule, and EpsK either 720 facilitates the activity of EpsJ and/or transports 721 the polysaccharide out of the cell. 722
In regard to EpsK, it is unclear how an 723 α-helical membrane transporter from a Gram- 724 positive organism would function in PNAG syn- 725 thesis and transport in E. coli. Using BLAST 726 analysis we noted that EpsK is a member of the 727 Wzx family of proteins (Table 2) which show 728 overall little conservation in their primary amino 729 acid sequences, but Wzx proteins can inter- 730 changeably export different polysaccharides 731 containing N-acetylglucosamine or N- 732
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
8
acetylgalactosamine as the initiating sugar (50). 733 Therefore, as a member of the Wzx family of 734 proteins it is entirely plausible that EpsK is 735 transporting the PNAG polymer out of the cell in 736 both B. subtilis and E. coli. 737
These structural inferences of the EpsHIJK 738 proteins are consistent with the predictions that 739 synthesis of PNAG, alginate, and cellulose, all 740 components of microbial biofilms, is mediated 741 by proteins with similar hallmarks and functions 742 but not necessarily with conserved architectures 743 (41,51). Moreover, it appears that some of the 744 proteins, in spite of little overall amino acid se- 745 quence identity, nonetheless maintain sufficient- 746 ly conserved and interchangeable functions, as 747 documented here for the B. subtilis and E. coli 748 PNAG biosynthetic proteins. 749
We found that deletion of these genes in B. 750 subtilis disrupted PNAG production as detected 751 by loss of immunoreactivity with MAb F598. 752 Cloning either the epsHIJK genes individually 753 into E. coli strains with in-frame deletions in the 754 endogenous pgaA, B, C, or D genes, or as a 4- 755 gene cluster into pgaABCD-deleted E. coli or K. 756 pneumoniae, resulted in production of immuno- 757 chemically-detectable PNAG, as did cloning the 758 epsHIJK genes into alginate-deficient P. aeru- 759 ginosa. Expression of epsHIJK in E. coli mu- 760 tants deficient in pga genes could result in either 761 production of authentic PNAG or a material that 762 is a component of another E. coli factor, such as 763 an LPS or a glycoprotein. We also found that the 764 epsH and epsJ genes can both complement the 765 E. coli pgaA and the pgaC genes, yet in both of 766 these organisms loss of any one of these genes 767 results in a PNAG-negative phenotype. It ap- 768 pears there is no cross-complementation within 769 the host strain. While the structural similarity of 770 EpsH and EpsJ explains, in part, the ability of 771 each to provide some cross-complementation, it 772 seems that they likely have some additional, 773 non-shared specific functions within B. subtilis 774 that are provided by other proteins when they are 775 expressed in E. coli. 776
While PNAG itself, or a structure containing 777 a PNAG component, appears to be an EPS ele- 778 ment involved in B subtilis biofilm formation, it 779 is likely other EPS molecules are also needed for 780 full pellicle formation. Studies of the eps locus 781 indicate deletion of numerous other genes also 782 disrupts biofilm formation, and some of these 783 appear to be classic EPS biosynthetic genes 784 (17,18,48). The EpsM, N and O proteins are 785 predicted to be an acetyltransferase (EpsM), an 786 amino transferase/sugar dehydratase (EpsN), and 787
a GT with a GT-B fold (EpsO). The EpsE pro- 788 tein is involved in both motility and biofilm 789 formation (21) acting as a clutch of the flagella 790 cellular motility apparatus to inhibit movement, 791 but also having a predicted GT structure. 792
Recently, Elsholz et al. (52) reported that the 793 B. subtilis EPS serves as a positive regulator of 794 its own synthesis by binding to the extracellular 795 portion of a receptor encoded by the epsA gene 796 which interacts with a tyrosine kinase encoded 797 by epsB and inhibits EpsB autophosphorylation. 798 Dephosphorylated EpsB is associated with en- 799 hanced EPS production. In an epsH mutant una- 800 ble to produce biofilm and, as shown here, 801 PNAG, a his-tagged recombinant EpsB protein 802 was highly phosphorylated due to lack of EPS 803 production. We found neither EpsA nor EpsB 804 were needed for synthesis of immunoreactive 805 PNAG in B. subtilis. This is consistent with the 806 finding from Elsholz et al. that PNAG-related 807 material prepared using a modified method from 808 Mack et al. (8) that yields small molecular 809 weight PIA had no effect on EpsB autophos- 810 phorylation. Given that the epsH mutant was not 811 producing a factor binding to the EpsA extracel- 812 lular domain it appears that other EPS compo- 813 nents either depend on EpsH for synthesis or 814 PNAG is needed to facilitate the inhibition of 815 EpsB autophosporylation. However, the actual 816 factors regulating B. subtilis EPS production via 817 EpsA and EpsB are currently uncharacterized in 818 regards to specific chemical properties. 819
One final aspect to consider is that the poly- 820 saccharide synthesized by the B. subtilis EpsH, I, 821 J and K proteins may not be PNAG but a molec- 822 ular entity with sufficient β-1-6-linked N- 823 acetylglucosamine in it to make it immunoreac- 824 tive with antibodies to PNAG but also contain- 825 ing other components. MAb F598 binding to 826 PNAG requires a minimum of seven β-1-6- 827 linked N-acetylglucosamine residues (36,53), in- 828 dicating a minimal PNAG-related constituent 829 present in the material synthesized by the EpsH, 830 I, J and K proteins. We do note that every mi- 831 crobial strain wherein we or others have identi- 832 fied a Dispersin B and periodate sensitive, hex- 833 osamine-containing antigen that reacts with an- 834 tibodies to native PNAG, and wherein the reac- 835 tive material has been isolated, turns out to be 836 chemically-verified PNAG (14,45,54-56). Fur- 837 thermore, only 1-6-linked hexosamines, and no 838 other possible amino-sugar linkages, are sensi- 839 tive to periodate and only β-1-6-linked N-acetyl 840 glucosamine can be digested by Dispersin B 841
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
9
(22). Also, for every microbe for which genes 842 encoding PNAG biosynthetic proteins have been 843 identified and deleted, there is loss of antibody 844 reactivity with material on the cell surface due to 845 gene loss. Thus, we consider it highly unlikely 846 that the material produced by the EpsH, I, J and 847 K proteins is not PNAG, but, at a minimum, it is 848 at least a PNAG-containing molecular structure. 849
Overall, our results show PNAG, or a close- 850 ly related entity, is a component of the B. subtilis 851 biofilm matrix synthesized by the EpsH, I, J and 852 K proteins. These proteins can also be expressed 853 and are functional in Gram-negative organisms 854 including E. coli, K. pneumoniae, and P. aeru- 855 ginosa. However, it is possible that the EpsH, I, 856 J and K proteins are also required for producing 857 other polysaccharide components of the biofilm 858 matrix, inasmuch as PNAG does not appear to 859 be the only carbohydrate entity in this organ- 860 ism’s biofilm (48). However, no carbohydrate 861 component other than PNAG identified here has 862 been fully characterized as being synthesized by 863 proteins encoded by genes within the B. subtilis 864 eps locus, leaving open the question as to the 865 chemical composition of additional eps- 866 dependent polymeric carbohydrates present in 867 this structure. Analysis of EPS composition 868 would undoubtedly advance the understanding 869 of the B subtilis biofilm-formation process, but it 870 should be noted that published methods used to 871 isolate EPS (52) would contain very little native 872 PNAG, as this molecule is poorly soluble at neu- 873 tral pH, particularly after alcohol precipitation 874 (14,45). Thus, in the absence of an appropriate 875 method, researchers have not been able to purify 876 PNAG to obtain an appreciable yield for detailed 877 chemical structure determination. We did find, 878 however, that extracts from the WT B. subtilis 879 cells and recombinant E. coli cells carrying 880 pUCP18Tc-epsHIJK plasmid contained immu- 881 noreactive, Dispersin B and periodate sensitive 882 hexosamine-containing material present. Thus, 883 the analysis of the products of the B. subtilis ep- 884 sHIJK locus described here are fully consistent 885 with the production by WT B. subtilis of PNAG, 886 which is likely a necessary component of this 887 organism’s biofilm based on the genetic data 888 showing the essentiality of epsH, I J and K gene 889 products for production of this structure (19). 890
891 Acknowledgments 892
This work was supported by NIH grants 893 EY016104 and AI057159 (a component of 894 award U54A1057159) to GBP, NIH grant 895 GM093030 to DBK, the AXA Research Fund 896
and the Fondation pour la Recherche Médicale 897 (FRM) to DR, awards from the William Ran- 898 dolph Hearst Fund to DR and the Hood Founda- 899 tion to DS and by a grant from the Canadian In- 900 stitutes of Health Research (CIHR, #43998) to 901 PLH, who is also the recipient of a Canada Re- 902 search Chair. D.J.L. has been supported in part 903 by graduate scholarships from the University of 904 Toronto, the Ontario Graduate Scholarship Pro- 905 gram, and CIHR. 906
907 Conflict of interest 908
Gerald B. Pier is an inventor of intellectual 909 properties [human monoclonal antibody to 910 PNAG and PNAG vaccines] that are licensed by 911 Brigham and Women’s Hospital to Alopexx 912 Vaccine, LLC, and Alopexx Pharmaceuticals, 913 LLC, entities in which GBP also holds equity. 914 As an inventor of intellectual properties, GBP 915 also has the right to receive a share of licensing- 916 related income (royalties, fees) through Brigham 917 and Women’s Hospital from Alopexx Pharma- 918 ceuticals, LLC, and Alopexx Vaccine, LLC. 919 GBP’s interests were reviewed and are managed 920 by the Brigham and Women’s Hospital and 921 Partners Healthcare in accordance with their 922 conflict of interest policies. 923
Colette Cywes-Bentley and David Skurnik 924 are inventors of intellectual properties [use of 925 human monoclonal antibody to PNAG and use 926 of PNAG vaccines] that are licensed by Brigham 927 and Women’s Hospital to Alopexx Vaccine, 928 LLC, and Alopexx Pharmaceuticals, LLC. As 929 inventors of intellectual properties, CC-B and 930 DK also have the right to receive a share of li- 931 censing-related income (royalties, fees) through 932 Brigham and Women’s Hospital from Alopexx 933 Pharmaceuticals, LLC, and Alopexx Vaccine, 934 LLC. 935
936 Author contributions 937
DR performed experiments, analyzed data, 938 contributed to the study concept and wrote the 939 manuscript. CCB, YFZ, SP, MK, DK, DJL, 940 PLH, and DS performed experiments, analyzed 941 data, provided reagents, edited the manuscript 942 and contributed to the study concept. GBP su- 943 pervised the project, developed the study con- 944 cept, analyzed data and wrote the manuscript. 945
946
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
10
References 947 1. Hall-Stoodley, L., and Stoodley, P. (2009) Evolving concepts in biofilm infections. Cell 948
Microbiol 11, 1034-1043 949 2. Epstein, A. K., Pokroy, B., Seminara, A., and Aizenberg, J. (2011) Bacterial biofilm shows 950
persistent resistance to liquid wetting and gas penetration. Proc Natl Acad Sci U S A 108, 995- 951 1000 952
3. McDougald, D., Rice, S. A., Barraud, N., Steinberg, P. D., and Kjelleberg, S. (2012) Should we 953 stay or should we go: mechanisms and ecological consequences for biofilm dispersal. Nat Rev 954 Microbiol 10, 39-50 955
4. Harriott, M. M., and Noverr, M. C. (2011) Importance of Candida-bacterial polymicrobial 956 biofilms in disease. Trends Microbiol 19, 557-563 957
5. Bakaletz, L. O. (2012) Bacterial biofilms in the upper airway - evidence for role in pathology and 958 implications for treatment of otitis media. Paediatr Respir Rev 13, 154-159 959
6. Mack, D., Davies, A. P., Harris, L. G., Knobloch, J. K., and Rohde, H. (2009) Staphylococcus 960 epidermidis Biofilms: Functional Molecules, Relation to Virulence, and Vaccine Potential. Top 961 Curr Chem 288, 157-182 962
7. Cywes-Bentley, C., Skurnik, D., Zaidi, T., Roux, D., Deoliveira, R. B., Garrett, W. S., Lu, X., 963 O'Malley, J., Kinzel, K., Zaidi, T., Rey, A., Perrin, C., Fichorova, R. N., Kayatani, A. K., Maira- 964 Litran, T., Gening, M. L., Tsvetkov, Y. E., Nifantiev, N. E., Bakaletz, L. O., Pelton, S. I., 965 Golenbock, D. T., and Pier, G. B. (2013) Antibody to a conserved antigenic target is protective 966 against diverse prokaryotic and eukaryotic pathogens. Proc Natl Acad Sci U S A 110, E2209- 967 2218 968
8. Mack, D., Fischer, W., Krokotsch, A., Leopold, K., Hartmann, R., Egge, H., and Laufs, R. (1996) 969 The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a 970 linear β-1,6-linked glucosaminoglycan: purification and structural analysis. J Bacteriol 178, 175- 971 183 972
9. McKenney, D., Pouliot, K. L., Wang, Y., Murthy, V., Ulrich, M., Doring, G., Lee, J. C., 973 Goldmann, D. A., and Pier, G. B. (1999) Broadly protective vaccine for Staphylococcus aureus 974 based on an in vivo-expressed antigen. Science 284, 1523-1527 975
10. Cramton, S. E., Gerke, C., Schnell, N. F., Nichols, W. W., and Gotz, F. (1999) The intercellular 976 adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. 977 Infect Immun 67, 5427-5433 978
11. Heilmann, C., Schweitzer, O., Gerke, C., Vanittanakom, N., Mack, D., and Gotz, F. (1996) 979 Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. 980 Mol Microbiol 20, 1083-1091 981
12. Foulston, L., Elsholz, A. K., DeFrancesco, A. S., and Losick, R. (2014) The extracellular matrix 982 of Staphylococcus aureus biofilms comprises cytoplasmic proteins that associate with the cell 983 surface in response to decreasing pH. MBio 5, e01667-01614 984
13. Wang, X., Preston, J. F., 3rd, and Romeo, T. (2004) The pgaABCD locus of Escherichia coli 985 promotes the synthesis of a polysaccharide adhesin required for biofilm formation. J Bacteriol 986 186, 2724-2734 987
14. Choi, A. H. K., Slamti, L., Avci, F. Y., Pier, G. B., and Maira-Litrán, T. (2009) The pgaABCD 988 locus of Acinetobacter baumannii encodes the production of poly-β-1-6-N-acetyl glucosamine 989 which is critical for biofilm formation. J Bacteriol 191, 5953-5963 990
15. Skurnik, D., Davis, M. R., Jr., Benedetti, D., Moravec, K. L., Cywes-Bentley, C., Roux, D., 991 Traficante, D. C., Walsh, R. L., Maira-Litran, T., Cassidy, S. K., Hermos, C. R., Martin, T. R., 992 Thakkallapalli, E. L., Vargas, S. O., McAdam, A. J., Lieberman, T. D., Kishony, R., Lipuma, J. 993 J., Pier, G. B., Goldberg, J. B., and Priebe, G. P. (2012) Targeting pan-resistant bacteria with 994 antibodies to a broadly conserved surface polysaccharide expressed during infection. J Infect Dis 995 205, 1709-1718 996
16. Branda, S. S., Chu, F., Kearns, D. B., Losick, R., and Kolter, R. (2006) A major protein 997 component of the Bacillus subtilis biofilm matrix. Mol Microbiol 59, 1229-1238 998
17. Marvasi, M., Visscher, P. T., and Casillas Martinez, L. (2010) Exopolymeric substances (EPS) 999 from Bacillus subtilis: polymers and genes encoding their synthesis. FEMS Microbiol Lett 313, 1000 1-9 1001
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
11
18. Shank, E. A., and Kolter, R. (2011) Extracellular signaling and multicellularity in Bacillus 1002 subtilis. Curr Opin Microbiol 14, 741-747 1003
19. Nagorska, K., Ostrowski, A., Hinc, K., Holland, I. B., and Obuchowski, M. (2010) Importance of 1004 eps genes from Bacillus subtilis in biofilm formation and swarming. J Appl Genet 51, 369-381 1005
20. Blair, K. M., Turner, L., Winkelman, J. T., Berg, H. C., and Kearns, D. B. (2008) A molecular 1006 clutch disables flagella in the Bacillus subtilis biofilm. Science 320, 1636-1638 1007
21. Guttenplan, S. B., Blair, K. M., and Kearns, D. B. (2010) The EpsE flagellar clutch is 1008 bifunctional and synergizes with EPS biosynthesis to promote Bacillus subtilis biofilm 1009 formation. PLoS Genet 6, e1001243 1010
22. Ramasubbu, N., Thomas, L. M., Ragunath, C., and Kaplan, J. B. (2005) Structural analysis of 1011 dispersin B, a biofilm-releasing glycoside hydrolase from the periodontopathogen Actinobacillus 1012 actinomycetemcomitans. J Mol Biol 349, 475-486 1013
23. Martinez, A., Ramirez, O. T., and Valle, F. (1997) Improvement of culture conditions to 1014 overproduce beta-galactosidase from Escherichia coli in Bacillus subtilis. Appl Microbiol 1015 Biotechnol 47, 40-45 1016
24. Bertani, G. (2004) Lysogeny at mid-twentieth century: P1, P2, and other experimental systems. J 1017 Bacteriol 186, 595-600 1018
25. Kelley, L. A., and Sternberg, M. J. (2009) Protein structure prediction on the Web: a case study 1019 using the Phyre server. Nat Protoc 4, 363-371 1020
26. Hsieh, P.-F., Lin, T.-L., Yang, F.-L., Wu, M.-C., Pan, Y.-J., Wu, S.-H., and Wang, J.-T. (2012) 1021 Lipopolysaccharide O1 antigen contributes to the virulence in Klebsiella pneumoniae causing 1022 pyogenic liver abscess. PLoS ONE 7, e33155 1023
27. Lu, X., Skurnik, D., Pozzi, C., Roux, D., Cywes-Bentley, C., Ritchie, J. M., Munera, D., Gening, 1024 M. L., Tsvetkov, Y. E., Nifantiev, N. E., Waldor, M. K., and Pier, G. B. (2014) A poly-n- 1025 acetylglucosamine-Shiga toxin broad-spectrum conjugate vaccine for Shiga toxin-producing 1026 Escherichia coli. MBio 5, e00974-00914-e00974-00914 1027
28. Chuang, C. H., Wang, Y. H., Chang, H. J., Chen, H. L., Huang, Y. C., Lin, T. Y., Ozer, E. A., 1028 Allen, J. P., Hauser, A. R., and Chiu, C. H. (2013) Shanghai fever: a distinct Pseudomonas 1029 aeruginosa enteric disease. Gut 63, 736-743 1030
29. Cherepanov, P. P., and Wackernagel, W. (1995) Gene disruption in Escherichia coli: TcR and 1031 KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. 1032 Gene 158, 9-14 1033
30. Yasbin, R. E., and Young, F. E. (1974) Transduction in Bacillus subtilis by bacteriophage SPP1. 1034 Journal of virology 14, 1343-1348 1035
31. Franklin, M. J., and Ohman, D. E. (1993) Identification of algF in the alginate biosynthetic gene 1036 cluster of Pseudomonas aeruginosa which is required for alginate acetylation. J Bacteriol 175, 1037 5057-5065 1038
32. Skurnik, D., Roux, D., Cattoir, V., Danilchanka, O., Lu, X., Yoder-Himes, D., Han, K., Guillard, 1039 T., Jiang, D., Gaultier, C., Guerin, F., Aschard, H., Leclercq, R., Mekalanos, J. J., Lory, S., and 1040 Pier, G. B. (2013) Enhanced in vivo fitness of Pseudomonas aeruginosa carbapenem-resistant 1041 oprD mutants revealed through In-seq analysis. Proc Natl Acad Sci U S A 110, 20747-20752 1042
33. Smith, R. L., and Gilkerson, E. (1979) Quantitation of glycosaminoglycan hexosamine using 3- 1043 methyl-2- benzothiazolone hydrazone hydrochloride. Anal Biochem 98, 478-480 1044
34. Maira-Litran, T., Kropec, A., Goldmann, D. A., and Pier, G. B. (2005) Comparative opsonic and 1045 protective activities of Staphylococcus aureus conjugate vaccines containing native or 1046 deacetylated Staphylococcal Poly-N-acetyl-beta-(1-6)-glucosamine. Infect Immun 73, 6752-6762 1047
35. Romero-Steiner, S., Frasch, C. E., Carlone, G., Fleck, R. A., Goldblatt, D., and Nahm, M. H. 1048 (2006) Use of opsonophagocytosis for serological evaluation of pneumococcal vaccines. Clin 1049 Vaccine Immunol 13, 165-169 1050
36. Gening, M. L., Maira-Litran, T., Kropec, A., Skurnik, D., Grout, M., Tsvetkov, Y. E., Nifantiev, 1051 N. E., and Pier, G. B. (2010) Synthetic β-(1->6)-linked N-acetylated and nonacetylated 1052 oligoglucosamines used to produce conjugate vaccines for bacterial pathogens. Infect Immun 78, 1053 764-772 1054
37. Valvano, M. A. (2011) Common themes in glycoconjugate assembly using the biogenesis of O- 1055 antigen lipopolysaccharide as a model system. Biochemistry (Mosc) 76, 729-735 1056
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
12
38. Willis, L. M., and Whitfield, C. (2013) Structure, biosynthesis, and function of bacterial capsular 1057 polysaccharides synthesized by ABC transporter-dependent pathways. Carbohydr Res 378, 35-44 1058
39. Whitfield, C., and Paiment, A. (2003) Biosynthesis and assembly of Group 1 capsular 1059 polysaccharides in Escherichia coli and related extracellular polysaccharides in other bacteria. 1060 Carbohydr Res 338, 2491-2502 1061
40. Gerke, C., Kraft, A., Sussmuth, R., Schweitzer, O., and Gotz, F. (1998) Characterization of the 1062 N-acetylglucosaminyltransferase activity involved in the biosynthesis of the Staphylococcus 1063 epidermidis polysaccharide intercellular adhesin. J Biol Chem 273, 18586-18593 1064
41. Morgan, J. L., Strumillo, J., and Zimmer, J. (2013) Crystallographic snapshot of cellulose 1065 synthesis and membrane translocation. Nature 493, 181-186 1066
42. Alibert-Franco, S., Pradines, B., Mahamoud, A., Davin-Regli, A., and Pages, J. M. (2009) Efflux 1067 mechanism, an attractive target to combat multidrug resistant Plasmodium falciparum and 1068 Pseudomonas aeruginosa. Curr Med Chem 16, 301-317 1069
43. Little, D. J., Bamford, N. C., Pokrovskaya, V., Robinson, H., Nitz, M., and Howell, P. L. (2014) 1070 Structural basis for the De-N-acetylation of Poly-beta-1,6-N-acetyl-d-glucosamine in gram- 1071 positive bacteria. J Biol Chem 289, 35907-35917 1072
44. Little, D. J., Poloczek, J., Whitney, J. C., Robinson, H., Nitz, M., and Howell, P. L. (2012) The 1073 structure- and metal-dependent activity of Escherichia coli PgaB provides insight into the partial 1074 de-N-acetylation of poly-beta-1,6-N-acetyl-D-glucosamine. J Biol Chem 287, 31126-31137 1075
45. Maira-Litran, T., Kropec, A., Abeygunawardana, C., Joyce, J., Mark, G., 3rd, Goldmann, D. A., 1076 and Pier, G. B. (2002) Immunochemical properties of the staphylococcal poly-N- 1077 acetylglucosamine surface polysaccharide. Infect Immun 70, 4433-4440 1078
46. Conover, M. S., Sloan, G. P., Love, C. F., Sukumar, N., and Deora, R. (2010) The Bps 1079 polysaccharide of Bordetella pertussis promotes colonization and biofilm formation in the nose 1080 by functioning as an adhesin. Mol Microbiol 77, 1439-1455 1081
47. Ye, L., Zheng, X., and Zheng, H. (2014) Effect of sypQ gene on poly-N-acetylglucosamine 1082 biosynthesis in Vibrio parahaemolyticus and its role in infection process. Glycobiology 24, 351- 1083 358 1084
48. Vlamakis, H., Chai, Y., Beauregard, P., Losick, R., and Kolter, R. (2013) Sticking together: 1085 building a biofilm the Bacillus subtilis way. Nat Rev Microbiol 11, 157-168 1086
49. Romero, D. (2013) Bacterial determinants of the social behavior of Bacillus subtilis. Res 1087 Microbiol 164, 788-798 1088
50. Marolda, C. L., Vicarioli, J., and Valvano, M. A. (2004) Wzx proteins involved in biosynthesis 1089 of O antigen function in association with the first sugar of the O-specific lipopolysaccharide 1090 subunit. Microbiology 150, 4095-4105 1091
51. Whitney, J. C., and Howell, P. L. (2013) Synthase-dependent exopolysaccharide secretion in 1092 Gram-negative bacteria. Trends Microbiol 21, 63-72 1093
52. Elsholz, A. K., Wacker, S. A., and Losick, R. (2014) Self-regulation of exopolysaccharide 1094 production in Bacillus subtilis by a tyrosine kinase. Genes Dev 28, 1710-1720 1095
53. Gening, M. L., Tsvetkov, Y. E., Pier, G. B., and Nifantiev, N. E. (2007) Synthesis of β-(1->6)- 1096 linked glucosamine oligosaccharides corresponding to fragments of the bacterial surface 1097 polysaccharide poly-N-acetylglucosamine. Carbohydr Res 342, 567-575 1098
54. Kaplan, J. B., Velliyagounder, K., Ragunath, C., Rohde, H., Mack, D., Knobloch, J. K., and 1099 Ramasubbu, N. (2004) Genes involved in the synthesis and degradation of matrix polysaccharide 1100 in Actinobacillus actinomycetemcomitans and Actinobacillus pleuropneumoniae biofilms. J 1101 Bacteriol 186, 8213-8220 1102
55. Izano, E. A., Sadovskaya, I., Vinogradov, E., Mulks, M. H., Velliyagounder, K., Ragunath, C., 1103 Kher, W. B., Ramasubbu, N., Jabbouri, S., Perry, M. B., and Kaplan, J. B. (2007) Poly-N- 1104 acetylglucosamine mediates biofilm formation and antibiotic resistance in Actinobacillus 1105 pleuropneumoniae. Microb Pathog 43, 1-9 1106
56. Bobrov, A. G., Kirillina, O., Forman, S., Mack, D., and Perry, R. D. (2008) Insights into Yersinia 1107 pestis biofilm development: topology and co-interaction of Hms inner membrane proteins 1108 involved in exopolysaccharide production. Environ Microbiol 10, 1419-1432 1109
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
13
57. Simon, R., O'Connell, M., Labes, M., and Puhler, A. (1986) Plasmid vectors for the genetic 1110 analysis and manipulation of rhizobia and other gram-negative bacteria. Methods Enzymol 118, 1111 640-659 1112
58. Kearns, D. B., and Losick, R. (2003) Swarming motility in undomesticated Bacillus subtilis. Mol 1113 Microbiol 49, 581-590 1114
59. Pozsgai, E. R., Blair, K. M., and Kearns, D. B. (2012) Modified mariner transposons for random 1115 inducible-expression insertions and transcriptional reporter fusion insertions in Bacillus subtilis. 1116 Appl Environ Microbiol 78, 778-785 1117
60. Konkol, M. A., Blair, K. M., and Kearns, D. B. (2013) Plasmid-encoded ComI inhibits 1118 competence in the ancestral 3610 strain of Bacillus subtilis. J Bacteriol 195, 4085-4093 1119
61. Muyrers, J. P., Zhang, Y., Testa, G., and Stewart, A. F. (1999) Rapid modification of bacterial 1120 artificial chromosomes by ET-recombination. Nucleic Acids Res 27, 1555-1557 1121
62. Doublet, B., Douard, G., Targant, H., Meunier, D., Madec, J. Y., and Cloeckaert, A. (2008) 1122 Antibiotic marker modifications of lambda Red and FLP helper plasmids, pKD46 and pCP20, for 1123 inactivation of chromosomal genes using PCR products in multidrug-resistant strains. J 1124 Microbiol Methods 75, 359-361 1125
63. Urban, T. A., Griffith, A., Torok, A. M., Smolkin, M. E., Burns, J. L., and Goldberg, J. B. (2004) 1126 Contribution of Burkholderia cenocepacia flagella to infectivity and inflammation. Infect Immun 1127 72, 5126-5134 1128
64. Patrick, J. E., and Kearns, D. B. (2008) MinJ (YvjD) is a topological determinant of cell division 1129 in Bacillus subtilis. Mol Microbiol 70, 1166-1179 1130
1131 Footnotes 1132 4The abbreviations used are: 1133
EPS exopolysaccharides 1134 PNAG poly-N-acetyl glucosamine 1135 PIA polysaccharide intercellular adhesin 1136 GT glycosyltransferase 1137 MMG minimal-medium glucose 1138 LB lysogeny broth 1139 OPK Opsonophagocytic killing 1140 MATE multi antimicrobial extrusion protein 1141 UnDP undecaprenyl 1142
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
14
Figure Legends 1143 Fig 1. PNAG production in 10-day old pellicles of B. subtilis. Pellicles/biofilms formed at the 1144 air-liquid interface of MMG medium over 10 days at RT were analyzed via immunofluorescence 1145 for the presence of PNAG within these structures. Cells were identified using Syto 83 DNA stain 1146 and PNAG by reactivity with MAb F598 directly conjugated to AF488. Loss or reactivity follow- 1147 ing Dispersin B or periodate treatment are properties highly specific to the β-1-6-linked N-acetyl 1148 glucosamine PNAG polymer. Bars = 10 µm. 1149 Fig. 2. Demonstration of PNAG production by B. subtilis WT and Tn-insertion strains by 1150 immunofluorescence microscopy. A. Parental strain B. subtilis DS991 failed to react with MAb 1151 F429 to P. aeruginosa alginate after chitinase treatment but did bind MAb F598 to PNAG. Bind- 1152 ing was lost after treatment of cells with Dispersin B or sodium periodate. B. Binding of MAb 1153 F429 (control) or F598 to PNAG to indicate B. subtilis strain with a Tn insertion in the indicated 1154 gene in the eps locus (Δeps). DNA stained with Syto 83 to visualize cells, MAbs conjugated di- 1155 rected to Alexa Fluor 488 (green). White bars = 10 µm. 1156 Fig. 3. Structural models of the EpsH, I, J, and K proteins. Predicted model of EpsH based on 1157 the cellulose synthase BcsA (PDB ID 4HG6) shows a GT-A fold. Predicted model of EpsI based 1158 on the family 9 GT (PDB ID 3TOV) shows a GT-B fold. Predicted model of EpsJ based on a 1159 chondroitin polymerase (PDB ID 2Z86) shows a GT-A fold. Predicted model of EpsK based on 1160 DinF (PDB ID 4LZ9) shows a MATE transporter fold. The structure models were generated using 1161 Phyre2 (25) and are shown in cartoon representations with α-helices, β-strands, and loops colored 1162 orange, blue, and grey, respectively. Predicted active site residues for the EpsH, I and J proteins 1163 are shown as magenta sticks. 1164 Fig. 4. Production of PNAG in alginate-deficient P. aeruginosa FRD1 in the presence of the 1165 B. subtilis epsHIJK locus. WT P. aeruginosa FRD1 produces alginate and binds AF488-labeled 1166 MAb F429 but not PNAG-specific MAb F598. A polar Tn-insertion in the algF gene eliminated 1167 alginate production and insertion of the B. subtilis epsHIJK locus leads to PNAG production. 1168 Immunoreactive PNAG in this latter strain is lost after treatment with Dispersin B or periodate. 1169 White bars = 10 µm. 1170 Fig. 5. Detection of PNAG in extracts of E. coli and B. subtilis strains. Extracts of the indicat- 1171 ed E. coli or B. subtilis strain were immobilized on membranes and probed with MAb F598 to 1172 PNAG followed by anti-human IgG conjugated to HRP. Both WT E. coli and E. coli Δpga carry- 1173 ing the cloned B. subtilis epsHIJK genes expressed PNAG whereas the Δpga strain did not con- 1174 tain detectable PNAG. Similarly, extracts of cells of B. subtilis DS991 had detectable PNAG but 1175 none was present in the strains lacking either the epsH or epsJ genes. Extracts with detectable 1176 PNAG lost reactivity with MAb F598 following Dispersin B or periodate treatment, and con- 1177 tained detectable hexosamine. N/A- not applicable. 1178 Fig. 6. Opsonophagocytic killing of indicated E. coli or B. subtilis strains by antibodies to 1179 PNAG raised by immunization with a conjugate vaccine of 9GlcNH2-TT. Bars indicate mean 1180 percent bacteria killed in duplicates compared with those in the corresponding dilution of non- 1181 immune goat or rabbit antiserum to PNAG. 1182
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
15
Table 1. Strains and plasmids used in this study 1183 Strain or plasmid Relevant characteristics Source or 1184 reference 1185 E. coli sm10 thi-1 relA1 thi-1 thr leu tonA lacY 1186 supE recA RP4-2-Tc::Mu Kmr (57) 1187 B. subtilis 3610 Undomesticated wild strain (58) 1188 B. subtilis 168 (DS991) sinR::kan tasA::Tn10 spec (59) 1189 B. subtilis DK1042 comIQ12L (60) 1190 B. subtilis DS2569 ΔpBS32 (60) 1191 B. subtilis 168 (DK1943) ΔepsAi comIQ12L This study 1192 B. subtilis 168 (DK1806) ΔepsB comIQ12L This study 1193 B. subtilis 168 (DK1807) ΔepsC comIQ12L This study 1194 B. subtilis 168 (DS4248) ΔepsD This study 1195 B. subtilis 168 (DS2152) ΔepsE (20) 1196 B. subtilis 168 (DS4164) ΔepsF This study 1197 B. subtilis 168 (DS7499) ΔepsG This study 1198 B. subtilis 168 (DS6776) ΔepsH This study 1199 B. subtilis 168 (DK1758) ΔepsI comIQ12L This study 1200 B. subtilis 168 (DS4166) ΔepsJ This study 1201 B. subtilis 168 (DK2055) ΔepsK comIQ12L This study 1202 B. subtilis 168 (DS7432) ΔepsL This study 1203 B. subtilis 168 (DS4901) ΔepsM This study 1204 B. subtilis 168 (DS4900) ΔepsN This study 1205 B. subtilis 168 (DK1759) ΔepsO comIQ12L This study 1206 1207 E. coli E11 K1 capsule type; clinical isolate K. Sik-Kim 1208 E. coli E11 ∆pga E. coli E11 with in-frame deletion of Baltimore, MD 1209
entire pga locus 1210 E. coli E11 ∆pgaA E. coli E11 in-frame deletion of pgaA This study 1211 E. coli E11 ∆pgaB E. coli E11 in-frame deletion of pgaB This study 1212 E. coli E11 ∆pgaC E. coli E11 in-frame deletion of pgaC This study 1213 E. coli E11 ∆pgaD E. coli E11 in-frame deletion of pgaD This study 1214 1215 K. pneumoniae K2 capsule type; clinical isolate 1216 K. pneumoniae ∆pga K. pneumoniae K2 with in-frame deletion of 1217
entire pga locus This study 1218 P. aeruginosa WT strain FRD1 (31) 1219 P. aeruginosa Tn::algF FRD1 Tn501::algF, alginate-deficient (31) 1220 Plasmid 1221 pRED/ET Red/ET expression plasmid (61) 1222 pCP20 Helper plasmid; FLP+, temperature-sensitive, 1223
AmpR, CmR (62) 1224 pUCP18Tc Broad-host-range vector 1225 pUCP18 derivative; TetR (63) 1226 pUCP18Tc-pga TetR, pgaABCD (pga) locus from E. coli E11 This study 1227 cloned into pUCP18Tc at XbaI–SbfI sites 1228 pUCP18Tc-pgaA TetR, pgaA from E. coli E11 cloned into This study 1229 pUCP18Tc at XbaI–SbfI sites 1230 pUCP18Tc-pgaB TetR, pgaB from E. coli E11 cloned into This study 1231 pUCP18Tc at XbaI–SbfI sites 1232
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
16
pUCP18Tc-pgaC TetR, pgaC from E. coli E11 cloned into This study 1233 pUCP18Tc at SacI–XbaI sites 1234 pUCP18Tc-pgaD TetR, pgaD from E. coli E11 cloned This study 1235 into pUCP18Tc at XbaI–SbfI sites 1236 pUCP18Tc-epsHIJK TetR, epsHIJK from B. subtilis DS991 cloned This study 1237 into pUCP18Tc at XbaI–SbfI sites 1238 pUCP18Tc-epsH TetR, epsH locus from B. subtilis DS991 cloned This study 1239 into pUCP18Tc at XbaI–SbfI sites 1240 pUCP18Tc-epsI TetR, epsI locus from B. subtilis DS991 cloned This study 1241 into pUCP18Tc at XbaI–SbfI sites 1242 pUCP18Tc-epsJ TetR, epsJ locus from B. subtilis DS991 cloned This study 1243 into pUCP18Tc at XbaI–SbfI sites 1244 pUCP18Tc-epsK TetR, epsK locus from B. subtilis DS991 cloned This study 1245 into pUCP18Tc at XbaI–SbfI sites 1246 pMiniMAD oriBsTs amp mls (64) 1247 pMP201 ΔepsA mls amp This study 1248 pMP202 ΔepsB mls amp This study 1249 pMP203 ΔepsC mls amp This study 1250 pKB90 ΔepsD mls amp This study 1251 pKB91 ΔepsF mls amp This study 1252 pSG52 ΔepsG mls amp This study 1253 pSG37 ΔepsH mls amp This study 1254 pMP204 ΔepsI mls amp This study 1255 pKB92 ΔepsJ mls amp This study 1256 pMP215 ΔepsK mls amp This study 1257 pSG53 ΔepsL mls amp This study 1258 pMP6 ΔepsM mls amp This study 1259 pMP5 ΔepsN mls amp This study 1260 pMP206 ΔepsO mls amp This study 1261 1262 Cassette 1263 FRT-PGK-gb2-neo-FRT PGK-gb2-neo cassette flanked by FRT sites Gene Bridges GmbH 1264
Heidelberg Ger- 1265 many 1266
1267
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
PNAG in B. subtilis biofilms
17
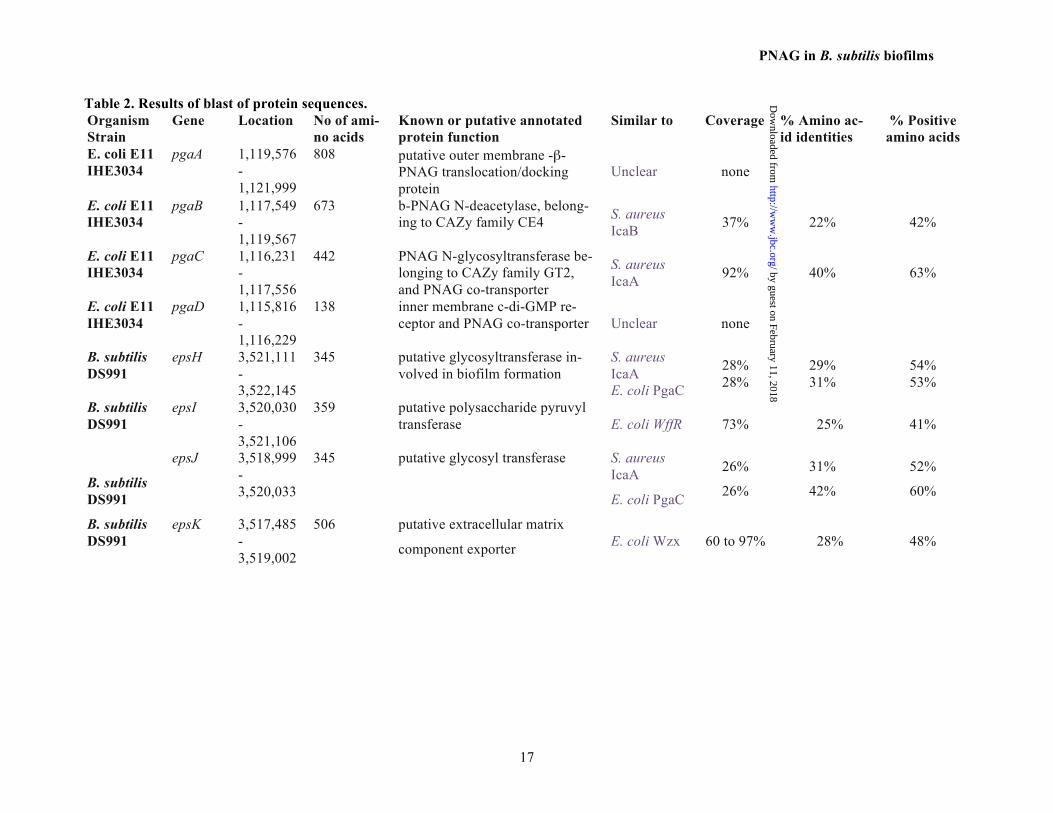
Table 2. Results of blast of protein sequences. Organism Strain
Gene Location No of ami-no acids
Known or putative annotated protein function
Similar to
Coverage % Amino ac-id identities
% Positive amino acids
E. coli E11 IHE3034
pgaA 1,119,576-1,121,999
808 putative outer membrane -β-PNAG translocation/docking protein
Unclear none
E. coli E11 IHE3034
pgaB 1,117,549-1,119,567
673 b-PNAG N-deacetylase, belong-ing to CAZy family CE4 S. aureus
IcaB 37% 22% 42%
E. coli E11 IHE3034
pgaC 1,116,231-1,117,556
442 PNAG N-glycosyltransferase be-longing to CAZy family GT2, and PNAG co-transporter
S. aureus IcaA 92% 40% 63%
E. coli E11 IHE3034
pgaD 1,115,816-1,116,229
138 inner membrane c-di-GMP re-ceptor and PNAG co-transporter Unclear none
B. subtilis DS991
epsH 3,521,111-3,522,145
345 putative glycosyltransferase in-volved in biofilm formation
S. aureus IcaA E. coli PgaC
28% 28%
29% 31%
54% 53%
B. subtilis DS991
epsI 3,520,030-3,521,106
359 putative polysaccharide pyruvyl transferase E. coli WffR 73% 25% 41%
B. subtilis DS991
epsJ 3,518,999-3,520,033
345 putative glycosyl transferase S. aureus IcaA
E. coli PgaC
26%
26%
31%
42%
52%
60%
B. subtilis DS991
epsK 3,517,485-3,519,002
506 putative extracellular matrix
component exporter E. coli Wzx 60 to 97% 28% 48%
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
18
Table 3. A. Summary for detection of PNAG synthesis by immunofluorescence in indicated strain
Strain Complemented with Control MAb
MAb to PNAG
E. coli K1 WT -a +++
E. coli K1 ∆pgaA - -
E. coli K1 ∆pgaB - -
E. coli K1 ∆pgaC - -
E. coli K1 ∆pgaD - -
E. coli K1 ∆pgaA (pUCP18Tc) - -
E. coli K1 ∆pgaB - -
E. coli K1 ∆pgaC - -
E. coli K1 ∆pgaD - -
E. coli K1 ∆pgaA (pUCP18Tc-pgaA) - +++
E. coli K1 ∆pgaB (pUCP18Tc-pgaB) - +++
E. coli K1 ∆pgaC (pUCP18Tc-pgaC) - +++
E. coli K1 ∆pgaD (pUCP18Tc-pgaD) - +++
E. coli K1 ∆pgaA (pUCP18Tc-epsH) - +++
E. coli K1 ∆pgaA (pUCP18Tc-epsJ) - +
E. coli K1 ∆pgaB (pUCP18Tc-epsI) - +++
E. coli K1 ∆pgaC (pUCP18Tc-epsH) - +
E. coli K1 ∆pgaC (pUCP18Tc-epsJ) - +++
E. coli K1 ∆pgaD (pUCP18Tc-epsK) - +++
E. coli K1 ∆pgaABCD (pUCP18Tc-epsHIJK) - +++
K. pneumoniae ∆pga (pUCP18Tc) - -
a Key: -, no immunofluorescence detected; +, 25-50% of WT levels of PNAG observed; ++, 51-80% of WT levels of PNAG observed; +++, >80% of WT levels of PNAG observed
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
19
Table 4. Levels of intracellular PNAG in WT and B. subtilis ΔepsK strain
Strain ng/108 CFU (mean ± SD)
B. subtilis DS991 (WT) 152±52 B. subtilis ΔepsK 242±64 E. coli E11 (WT) 6±1
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
PNAG in B. subtilis biofilms
20
Fig 1. PNAG production in 10-day old pellicles of B. subtilis. Pellicles/biofilms formed at the air-liquid interface of MMG medium over 10 days at RT were analyzed via immunofluorescence for the presence of PNAG within these structures. Cells were identified using Syto 83 DNA stain and PNAG by reactivity with MAb F598 directly conjugated to AF488. Loss or reactivity follow-ing Dispersin B or periodate treatment are properties highly specific to the b-1-6-linked N-acetyl glucosamine PNAG polymer. Bars = 10 µm.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
21
Fig. 2. Demonstration of PNAG production by B. subtilis WT and Tn-insertion strains by immunofluorescence microscopy. A. Parental strain B. subtilis DS991 failed to react with MAb F429 to P. aeruginosa alginate after chitinase treatment but did bind MAb F598 to PNAG. Bind-ing was lost after treatment of cells with Dispersin B or sodium periodate. B. Binding of MAb F429 (control) or F598 to PNAG to indicate B. subtilis strain with a Tn insertion in the indicated gene in the eps locus (Δeps). DNA stained with Syto 83 to visualize cells, MAbs conjugated di-rected to Alexa Fluor 488 (green). White bars = 10 µm.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
PNAG in B. subtilis biofilms
22
Fig. 3. Structural models of the EpsH, I, J, and K proteins. Predicted model of EpsH based on the cellulose synthase BcsA (PDB ID 4HG6) shows a GT-A fold. Predicted model of EpsI based on the family 9 GT (PDB ID 3TOV) shows a GT-B fold. Predicted model of EpsJ based on a chondroitin polymerase (PDB ID 2Z86) shows a GT-A fold. Predicted model of EpsK based on DinF (PDB ID 4LZ9) shows a MATE transporter fold. The structure models were generated using Phyre2 (25) and are shown in cartoon representations with α-helices, β-strands, and loops colored orange, blue, and grey, respectively. Predicted active site residues for the EpsH, I and J proteins are shown as magenta sticks.
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
PNAG in B. subtilis biofilms
23
Fig. 4. Production of PNAG in alginate-deficient P. aeruginosa FRD1 in the presence of the B. subtilis epsHIJK locus. WT P. aeruginosa FRD1 produces alginate and binds AF488-labeled MAb F429 but not PNAG-specific MAb F598. A polar Tn-insertion in the algF gene eliminated alginate production and insertion of the B. subtilis epsHIJK locus leads to PNAG production. Immunoreactive PNAG in this latter strain is lost after treatment with Dispersin B or periodate. White bars = 10 µm.
P.#aeruginosa#FRD1+pUCp18Tc-epsHIJK,MAb,F429,,,,DNA ,,Alginate,,,,,,,,,,,,,,MAb,F598,,,DNA ,,,,,,PNAG,
P.#aeruginosa#FRD1#Tn501::algF,,,, , , ,,,, ,,,MAb,F429,,,,DNA ,,,Alginate ,,,,MAb,F598,,,,,DNA,,, ,,,,,,,PNAG,
P.#aeruginosa#FRD1,Tn501::algF,(pUCp18Tc-epsHIJK),MAb,F429,,,,,DNA ,,,Alginate ,,,,,MAb,F598,,,DNA ,,,,,,,,PNAG,
P.#aeruginosa#FRD1,Tn501::algF,(pUCp18Tc-epsHIJK)##plus,Dispersin,B,or,periodate,
MAb,F598,,,,DNA ,Dispersin,B ,,,,MAb,F598,,,,DNA ,,,,Periodate,
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

PNAG in B. subtilis biofilms
24
Fig. 5. Detection of PNAG in extracts of E. coli and B. subtilis strains. Extracts of the indicat-ed E. coli or B. subtilis strain were immobilized on membranes and probed with MAb F598 to PNAG followed by anti-human IgG conjugated to HRP. Both WT E. coli and E. coli Δpga carry-ing the cloned B. subtilis epsHIJK genes expressed PNAG whereas the Δpga strain did not con-tain detectable PNAG. Similarly, extracts of cells of B. subtilis DS991 had detectable PNAG but none was present in the strains lacking either the epsH or epsJ genes. Extracts with detectable PNAG lost reactivity with MAb F598 following Dispersin B or periodate treatment, and con-tained detectable hexosamine. N/A- not applicable.
B.#sub'lis#E.#coli#WT#
#Δpga#
#+pepsHIJK#
WT################Pos############Pos##ΔepsH###########N/A###########N/A##ΔepsJ############Pos#############N/A#
Dispersin/periodate##sensi4vity#
E.#coli #B.#sub'lis#Hexosamine#
E.#coli #B.#sub'lis#####Pos #Pos#
#######Neg #Neg########Pos #Neg#
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from
PNAG in B. subtilis biofilms
25
Fig. 6. Opsonophagocytic killing of indicated E. coli or B. subtilis strains by antibodies to PNAG raised by immunization with a conjugate vaccine of 9GlcNH2-TT. Bars indicate mean percent bacteria killed in duplicates compared with those in the corresponding dilution of non-immune goat or rabbit antiserum to PNAG.
0"
10"
20"
30"
40"
50"
60"
70"
80"
90"
No"complement"
10"40"160"
640"
Percen
t"bacteria
"killed
"
Bacillus(sub*lis(WT"
Goat"an?@9GlcNH2@TT"
Rabbit"an?@9GlcNH2@TT"
E.(coli(WT" E.(coli(ΔpgaABCD"(pUCp18Tc)(
E.(coli(ΔpgaABCD((pUCp18Tc3epsHIJK)"
Serum"dilu?on"
Rabbit anti-9GlcNH2-TT
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from

Daniel Kearns, Dustin J. Little, P. Lynne Howell, David Skurnik and Gerald B. PierDamien Roux, Colette Cywes-Bentley, Yi-Fan Zhang, Stephanie Pons, Melissa Konkol,
of the Bacillus Subtilis Biofilm MatrixIdentification of Poly-N-acetyl Glucosamine as a Major Polysaccharide Component
published online June 15, 2015J. Biol. Chem.
10.1074/jbc.M115.648709Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2015/06/15/M115.648709.DC1
by guest on February 11, 2018http://w
ww
.jbc.org/D
ownloaded from





























