Embed Size (px)
Citation preview

Experimental and Molecular Pathology 88 (2010) 278–286
Contents lists available at ScienceDirect
Experimental and Molecular Pathology
j ourna l homepage: www.e lsev ie r.com/ locate /yexmp
Breast cancer cell surface annexin II induces cell migration and neoangiogenesis viatPA dependent plasmin generation
Meena Sharma a, Robert T. Ownbey b, Mahesh C. Sharma b,c,d,e,⁎a University of Pennsylvania, Philadelphia, PA 19102, USAb Department of Surgery, Drexel University College of Medicine, Philadelphia, PA 19102, USAc Department of Pathology, Drexel University College of Medicine, Philadelphia, PA 19102, USAd Department of Biochemistry, George Washington University, Washington, DC 20037, USAe Veterans Affairs Medical Center, Washington, DC 20422, USA
⁎ Corresponding author. Research Services (151A), V50 Irving street, NW, Washington, DC 20422, USA.
E-mail address: [email protected] (M.C. Sharm
0014-4800/$ – see front matter. Published by Elsevierdoi:10.1016/j.yexmp.2010.01.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 6 November 2009Available online 15 January 2010
Keywords:PlasminAnnexin IIBreast cancerNeoangiogenesistPA
Annexin II, an abundant phospholipids binding cell surface protein, binds tPA and functions as a regulator offibrinolysis. Annexin II also mediates angiogenesis and enhances tumor growth and metastasis. However, themechanism supporting this role is not known. Using human breast cancer model we show that invasivehuman breast cancer cells (MDA-MB231) synthesize annexin II and tissue plasminogen activator (tPA). Invitro both annexin II and tPA interacts which in turn convert zymogen plasminogen to reactive enzymeplasmin. Cell surface produced plasmin inhibited the migration of MDA-MB231 cells. Silencing of annexin IIgene in MDA-MB231 cells abolished tPA binding therefore inhibited tPA dependent plasmin generation.These annexin II suppressed MDA-MB231 cells showed reduced motility. Immunohistochemical analysis ofprediagnosed clinical specimens showed abundant secretion of tPA and expression of annexin II on thesurface of invasive human breast cancer cells which correlates with neovascularization of the tumor. Takentogether, these data indicate that annexin II may regulate localized plasmin generation in breast cancer. Thismay be an early event switching breast cancer from the prevascular phase to the vascular phase and thuscontributing to aggressive cancer with the possibility of metastasis. The data provide a mechanismexplaining the role of annexin II in breast cancer progression and suggest that annexin II may be an attractivetarget for therapeutic strategies aimed to inhibit angiogenesis and breast cancer.
eterans Affairs Medical Center,
a).
Inc.
Published by Elsevier Inc.
Introduction
Sprouting of new blood vessels and metastasis are two processesthat are central to the progression of cancer. As such, they havebecome important targets for the development of anti-cancer agents.The plasminogen/plasmin system, a serine protease pathway, isknown to trigger sprouting of new blood vessels therefore plays acritical role in breast cancer (Stephens et al., 1998). Analysis of theenzymes of the plasminogen/plasmin system clearly suggests thathigh urokinase-type plasminogen activator (uPA) in human breasttumors predicts poor survival (Janicke et al., 1989; Look et al., 2002)and is significantly associated with a high rate of relapse for patientswith breast cancer (Janicke et al., 1989). Despite the established roleof the plasminogen/plasmin system in breast cancer, the receptor(s)which regulates conversion of zymogen plasminogen to activeenzyme plasmin in breast cancer are poorly understood. In thephysiological system two differentmechanisms of plasmin generationhave been established. The first mechanism is involved in binding of
uPA to its receptor uPAR while the second mechanism convertsplasminogen to plasmin by binding of tissue type plasminogenactivator (tPA) to its receptor annexin II. This annexin II dependentplasmin exerts fibrinolytic activity that maintains continuous bloodflow in vessels by dissolving intravascular fibrin clots.
Annexin II is reported to be an endothelial cell receptor whichprovides binding sites for plasminogen and tPA for efficient plasmingeneration (Cesarman et al., 1994; Hajjar et al., 1994). A number ofreports have shown that cell surface annexin II regulates plasmingeneration (Diaz et al., 2004; Hajjar et al., 1994) which in turnfacilitates extracellular matrix (ECM) degradation, cell invasion(Brownstein et al., 2004; Diaz et al., 2004; Mignatti and Rifkin,1993), and migration (Tarui et al., 2002) and thus plays an importantrole in neoangiogenesis (Ling et al., 2004). There are speculations thatannexin II derived neoangiogenesis may be involved in metastasis,and tumor progression. This might explain why we and otherinvestigators have consistently found over expression of annexin IIin various cancers including breast cancer (Chuthapisith et al., 2009;Guedj et al., 2009; Ohno et al., 2009; Pei et al., 2007; Sharma et al.,2006a; Shiozawa et al., 2008; Syed et al., 2007; Zhong et al., 2009;Zimmermann et al., 2004). Overexpression of annexin II in cancerseems to correlate with advanced stage with poor prognosis and

279M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
metastasis (Ohno et al., 2009; Sharma et al., 2006a; Shiozawa et al.,2008; Wu et al., 2002).
Breast cancer growth and metastasis requires extensive neoangio-genesis. The fact is that neoangiogenesis is an independent and highlysignificant prognostic indicator for overall and relapse-free survival inpatients with early-stage breast carcinoma (Weidner et al., 1992).Angiostatin, a potent inhibitor of angiogenesis, was reported to inhibithuman breast cancer growth and metastasis in xenograft model(O'Reilly et al., 1996; Peyruchaud et al., 2003) suggesting that anti-angiogenic therapy could be a potential therapeutic approach forbreast cancer. Previously we have reported that angiostatin binds toendothelial cell surface annexin II (Tuszynski et al., 2002). Thesefindings, together with observations that angiostatin inhibits neoan-giogenesis dependent growth of breast cancer may argue thatangiostatin inhibits neoangiogenesis dependent breast cancer growthand metastasis by interfering with annexin II/tPA dependent plasmingeneration in tumor microenvironment.
tPA is secreted by endothelial cells (EC) and is a ligand for cellsurface annexin II. Emerging reports and clinical observations link tPAto neoplastic transformation and the invasive phenotype of highlyaggressive tumors such as glioblastoma (Goh et al., 2005), melanoma(Stack et al., 1999) pancreatic cancer (Diaz et al., 2002) and breastcancer (Grondahl-Hansen et al., 1990; Rella et al., 1993). Thefunctional role of tPA and its receptor annexin II in breast tumorbiology is not well-understood. Recent studies have providedevidence that showed association of tPA with the invasion of highlyaggressive MDA-MB-435s breast cancer cells (Chernicky et al., 2005).Since tPA plays a major role in plasmin generation, consistentlyplasmin has been linked to invasive and metastatic breast cancer(Stephens et al., 1998). Indeed, recent studies have clearly demon-strated over expression of annexin II in many cancer types and mayargue that annexin II functions as a regulator of neoangiogenesis invivo. However the precise mechanism by which annexin II is involvedin cancer progression and metastasis is still unclear.
In this report we investigated if tPA interaction on breast cell surfaceannexin II is amolecular switch to regulate localized plasmin generationin vitro and neoangiogenesis in vivo. Taken together, this research mayprovide a clue for the annexin II-dependent mechanism of neoangio-genesis and its dependent breast cancer progression and metastasis.Annexin II may be a potential target for the development of effectivetherapeutic strategies to inhibit angiogenesis and breast cancer.
Materials and methods
Human Lys-plasminogen, plasmin and recombinant tPA waspurchased from Calbiochem, (La Jolla, CA). Lysine–Sepharose and ε-aminocaproic acid (ε-ACA)were procured from Sigma (St. Louis, MO).Chromatography and electrophoretic reagents were procured fromBioRad, (Richmond, CA). Anti-plasminogen monoclonal antibody waspurchased from Enzyme Research Inc. (Chicago, IL). Anti-tPA andAnti-CD105 monoclonal antibodies were purchased from AmericanDiagnostica (Stamford, CT) and Neomarkers (Fremont, CA) respec-tively. Antibodies to annexin II were generated in our lab as reportedearlier (Sharma et al., 2006b). Chromozyme PL was purchased fromRoche Molecular Biochemicals (IN). Immunohistochemistry reagentswere procured from DAKO Corporation (CA). All other chemicals usedin this study were of analytical grade.
Cell lines maintenance
Invasive andmetastatic human breast cancer cell lineMDA-MB231which is known to cause tumors in athymic mice and the ER positivenoninvasive breast cancer cell line MCF-7 were obtained from ATCC(Rockville, MD). These cell lines weremaintained in RPMI 1640mediacontaining either 10% fetal calf serum (FCS) or serum free medium(0.1% BSA supplemented with L-glutamine and antibiotics) (Sharma
et al., 2006a). Cell cultures were maintained in plastic flasks andincubated at 37 °C in a humidified chamber containing 5% CO2.
Immunohistochemistry
Previously diagnosed paraffin-embedded human breast cancersamples were obtained from the tumor bank of the Department ofPathology, Drexel University College of Medicine. Serial 4 μm sectionswere prepared on albumin-coated slides. Sections were deparaffi-nized and incubated in 3% H2O2 for 10 min to block endogenousperoxidase activity. Nonspecific protein binding was blocked with 3%bovine serum albumin (BSA)/PBS for 1 h. Sections were incubatedwith monoclonal antibodies overnight at room temperature. Stainingwas visualized using a DAKO kit with diaminobenzidine (DAB) as achromogen followed by nuclear counterstaining with hematoxylinaccording to our published protocol (Sharma et al., 2006a). For eachimmunohistochemical staining, we performed additional stainingwithout primary antibody in parallel and considered it as a control.The immunoreactive pattern was carefully recorded with the help ofour pathologist and photographed.
Immunoprecipitation and Western blot analysis
Cells grown in 12well-plates, rinsed in ice-cold phosphate-bufferedsaline (PBS), pH7.2, and lysed in 67 mmol/L Tris, pH 6.8, and 2% sodiumdodecyl sulfate on ice for total extracts or in buffer A [50 mmol/L Tris,150 mmol/L NaCl, pH 7.4, 0.1% Triton X-100 plus protease andphosphatase inhibitors (200 mmol/L Pefabloc, 1 mmol/L aprotinin,20 mmol/L leupeptin, 0.1 mmol/L sodium vanadate, 1 mmol/L sodiumfluoride, and 10 mmol/L sodium pyrophosphate)] for immunoprecip-itation. Protein concentration was measured using Bradford assay (Bio-Rad Laboratories, Hercules, CA). Immunoprecipitation were performedusing 0.4 to 1 mg of cellular proteins incubated with anti-annexin IImonoclonal antibody (0.5 μg/ml) overnight at 4 °C. The immunecomplex was precipitated with protein A Sepharose beads. Theimmunocatcher system was used to purify immunoglobulin complexby catching the resins containing immobilized Protein A. Resins werewashed three times and directly incubated with SDS sample dilutionbuffer and heated at 90 °C for 4 min. Immune complex was resolvedon 12% SDS-PAGE, transferred to nitrocellulose membrane, incubatedwith ant-tPA antibodies and analyzed by Western blot analysis. Afterincubation with the appropriate primary antibody, species-specificsecondary antibodies conjugated to horseradish peroxidase were usedand the membrane was developed by ECL as we have reportedpreviously (Sharma et al., 2006a,b; Tuszynski et al., 2002).
tPA binding and plasmin generation
Binding of tPA with breast cancer cell (MDA-MB 231) surface wasstudied by analyzing the capacity of cell bound tPA to activateplasminogen to plasmin. Human breast cancer cells were cultured,detached and counted. About 5000–7000 cells were incubated with300 units of recombinant human tPA ligand for 30 min on ice in PBS toallow tPA to bind cell surface annexin II. Cells were washed three timeswith chilled PBS to remove unbound tPA. Breast cancer cells with boundtPA were incubated with plasminogen and specific chromogenicsubstrate chromozyme PL in a total volume of 100 μl to determineplasminogen activation activity. tPA dependent plasmin generationwasrecorded at 405 nm for 40 min in 96-well-plate reader according to ourpublished protocol (Sharma et al., 2006a,b). Plasmin generated in thereactionmixture hydrolyzes the chromozyme PL forming a yellow colorof p-nitroaniline that absorbs light at 405 nm. The color absorbancewasmeasured by an automated 96-well plate reader (Bio-TeK Inc., VT). Thecolorimetric change is a direct measurement of plasmin generation.Plasminogen incubated with chromozyme PL was considered asnegative control indicative of auto degradation of plasminogen to

280 M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
plasmin during the reaction process. In a parallel experiment an equalamount of tPA unbound to cells were used as control. Absorbance ofcontrolwas subtracted fromexperimental readings. Datawere analyzedby non-linear regression algorithms using GraphPad Prizm software.
Annexin II gene silencing
Antisense phosphorothionate oligonucleotides (ODNs) and senseODNs (control) directed to 5′–3′coding sequence of the humanannexin II mRNA were designed and manufactured by Biognostic,Germany (www.biognostik.com). Three potential sites were identi-fied in annexin II gene and antisense oligonucleotides weresynthesized against these regions (5′CAA CAT GTC CAC TTC ACT - 3′location: 923-940 bases of total sequence) (5′ - CAT TTT CCA GGT CTCCTT - 3′ location: 882-899 bases of total sequence) (5′ - TCA TCC ACACCT TTG GTC - 3′ location: 277–294 bases of total sequence). Theantisense oligonucleotides were synthesized as 18-mer targetedannexin II gene sequences. The corresponding controls sequencewas also synthesized as 18 mer (5′GGA TTT ACC TAT TGC TGG 3′) (5′ACT ACG ACC TAC GTG AC 3′) (5′GGA TTT ACC TAT TGC TGG 3′).Control antisenses were used in all parallel experiments. Hereafter,the antisense and sense annexin II oligonucleotides will be referred toas RNAi ANX II and Control ANX II respectively. MDA-MB231 cellswere grown in 96-well plates in triplicates and transfected with RNAiANX II and control ANX II by adding into the culture medium.
Antisense ODNs are actively taken up by cells, partially throughfluid-phase endocytosis and possibly also through the putative receptor
Fig. 1. Expressions and interactions of annexin II and tPA in human breast cancer cells. Manalyzed by immunoblot analysis. Fig. 2 displays selective expression of annexin II andimmunoprecipitated (IP) with anti-tPA antibodies and the immune complex was electrophoIP pellet, lane 3 supernatant and lane 4 is IgG negative control). Panel D: cell lysate was imelectrophoresed and immunoblotted with anti-tPA (Panel C, Lane 1 positive control, lane 2 Ipellet in lane 2 of both gels suggest interaction of tPA and annexin II.
protein p80 that facilitates the cellular uptake of negatively chargedmolecules like ODNs or heparin. Fluorescein (FITC)-labeled phosphor-othioate ODNs were used to monitor cellular uptake and distribution(data not shown). Labeled antisense ODNs met the same standards ofpurity and stability as antisenseproducts.Whereas the cellularuptake ofantisense ODNs may be enhanced through various cationic lipids, mostof the cationic lipids are cytotoxic, and the treatmentmust be limited to6 to 8 h. In contrast, the half-life of antisense ODNs in serum containingculture media is >48 h. Therefore, adding the antisense ODNs to theculturemedium for the full duration of the experiment ismore effectivefor experiments with a longer time frame. Details of the transfectionprocess are provided below.
TransfectionTo optimize transfection efficiencies MDA-MB231 cells were
incubated with 3 different concentrations of RNAi ANX II and controlANX II (0.5, 2.0 and 4.0 µmol/L) for 12, 24, 48, and 72 h to determinethe rate of transfection. Our optimizing experiments showed that theuptake of RNAi ANX II was maximal with 0.5 µmol/L and afterincubation for 48 h. At this time point, annexin II protein wassuppressed >90% (see results). Therefore we used these optimumconditions to suppress annexin II gene to determine tPA bindings, it'sdependent plasminogen activation and role in cell migration.
Scratch wound healing assay for cell migrationCell migration was assayed as described (Sharma et al., 2004).
Briefly, about8000 cellswere seededandgrown to confluency in96well
DA-MB231cell lysate (20 μg) was separated by SDS-PAGE and protein expression wastPA in invasive breast cancer cell line MDA-MB231 (Fig. 2A and B). Cell lysate wasresed and immunoblotted with anti-annexin II (Panel 2C, Lane 1 positive control, lane 2munoprecipitated (IP) with anti-annexin II antibodies and the immune complex wasP pellet, lane 3 supernatant and lane 4 is IgG negative control). Co-immunoprecipitated
281M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
plates. A wound was created in monolayer of cells by scratching with asterile pipette tip. The remainingadherent cellswerewashed twicewithPBS. Media was replaced with 2% serum-supplemented medium. Cellmotility was measured by counting the number of cells repopulated inthe cleared area. Ten different fields (20X) were viewed to countmigrating cells. Data were plotted using GraphPad Prizm software.
Boyden chamber migration assay
The measurement of cell migration was performed as described by(Terranova et al., 1986). Chemotaxis cell migration assay wasperformed using a 96 well plate fitted with 8 μm pore Boydenmigration chambers (CHEMICON, Inc. cat # ECM510). Briefly, cellsstarved in serum free medium were harvested and plated at a densityof 2.5×104 cells in 100 μl without chemo-attractant in a Boydenchamber (upper well) in a 96 well plate. This assay is based on theBoyden chamber principle. The bottom well was filled (100 μl) withmedium containing 10% fetal bovine serum (FBS). Test reagents(plasminogen, plasmin, angiostatin, ε-aminocaproic acid, or antibo-dies) were added to the upper chamber and incubated for 24 h at37 °C in a humidified chamber (5%CO). After 24 h, the plate was
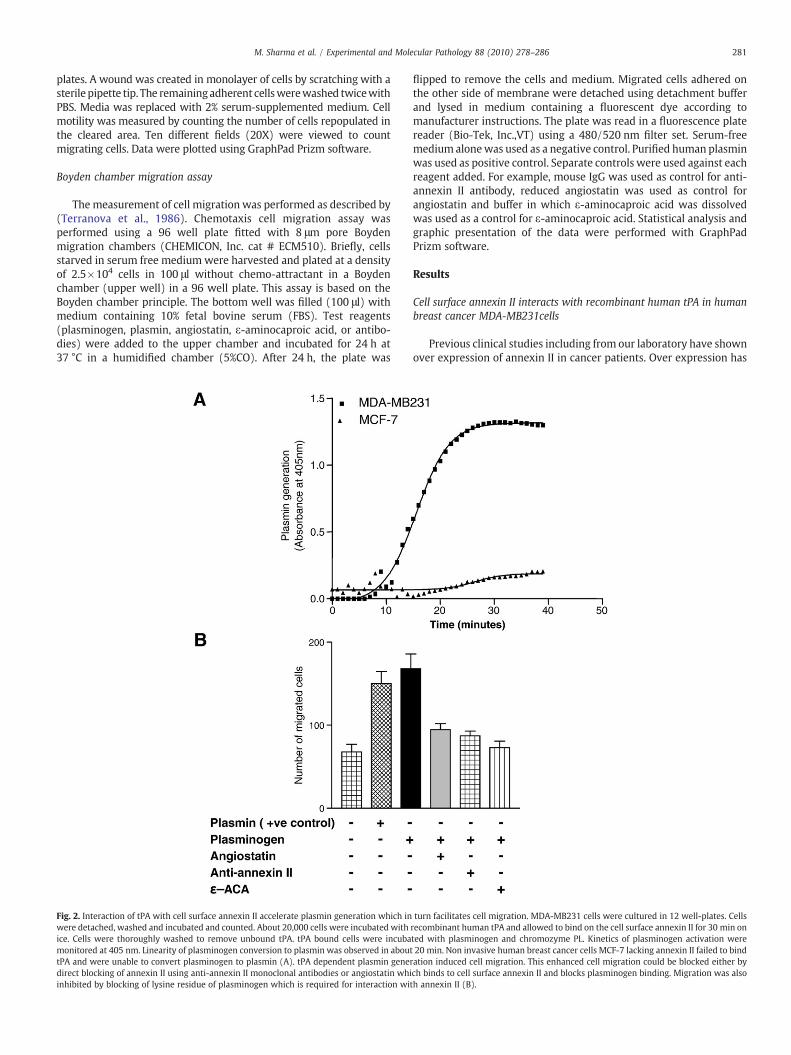
Fig. 2. Interaction of tPA with cell surface annexin II accelerate plasmin generation which inwere detached, washed and incubated and counted. About 20,000 cells were incubated withice. Cells were thoroughly washed to remove unbound tPA. tPA bound cells were incubamonitored at 405 nm. Linearity of plasminogen conversion to plasmin was observed in abouttPA and were unable to convert plasminogen to plasmin (A). tPA dependent plasmin genedirect blocking of annexin II using anti-annexin II monoclonal antibodies or angiostatin whiinhibited by blocking of lysine residue of plasminogen which is required for interaction wi
flipped to remove the cells and medium. Migrated cells adhered onthe other side of membrane were detached using detachment bufferand lysed in medium containing a fluorescent dye according tomanufacturer instructions. The plate was read in a fluorescence platereader (Bio-Tek, Inc.,VT) using a 480/520 nm filter set. Serum-freemedium alonewas used as a negative control. Purified human plasminwas used as positive control. Separate controls were used against eachreagent added. For example, mouse IgG was used as control for anti-annexin II antibody, reduced angiostatin was used as control forangiostatin and buffer in which ε-aminocaproic acid was dissolvedwas used as a control for ε-aminocaproic acid. Statistical analysis andgraphic presentation of the data were performed with GraphPadPrizm software.
Results
Cell surface annexin II interacts with recombinant human tPA in humanbreast cancer MDA-MB231cells
Previous clinical studies including from our laboratory have shownover expression of annexin II in cancer patients. Over expression has
turn facilitates cell migration. MDA-MB231 cells were cultured in 12 well-plates. Cellsrecombinant human tPA and allowed to bind on the cell surface annexin II for 30 min onted with plasminogen and chromozyme PL. Kinetics of plasminogen activation were20 min. Non invasive human breast cancer cells MCF-7 lacking annexin II failed to bind
ration induced cell migration. This enhanced cell migration could be blocked either bych binds to cell surface annexin II and blocks plasminogen binding. Migration was alsoth annexin II (B).

Fig. 3. Annexin II gene silencing inhibits tPA dependent plasmin generation. Threedifferent antisense oligos were designed and synthesized as 16-mer dithionatedoligonucleotide to silence annexin II gene. Immunoblot analysis suggests that two outof three antisense oligos were highly potent and almost completely silenced annexin IIexpression (A). β-actin was used as a loading control to confirm equal protein loading.Quantitative analysis of band densities showed more than 90% inhibition of annexin IIexpression as compared to β-actin loading control (B). Annexin II-silenced cells failedto bind tPA and significantly inhibited plasminogen activation (C).
282 M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
been correlated well with poor clinical outcomewith advanced stagesof cancers (Brichory et al., 2001; Roseman et al., 1994; Sharma et al.,2006a; Sharma and Sharma, 2007; Singh, 2007; Syed et al., 2007; Wuet al., 2002; Zimmermann et al., 2004 ). Because tPA is a ligand forannexin II and known to activate cell surface bound plasminogen toplasmin, we first tested whether annexin II and tPA proteins aresynthesized by human breast cancer cells. Immunoblot analysisindicated selective expression of annexin II and tPA in highly invasiveMDA-MB231cells but not in non invasive MCF-7 cells (Fig. 1A and B).Next we investigated whether tPA synthesized by MDA-MB231 cellsinteracts with annexin II. To determine interaction, we immunopre-cipitated an MDA-MB231 cell lysate with anti-tPA monoclonalantibody, and the immune complex was resolved on SDS-PAGE andtransferred onto nitrocellulose membranes. Bound proteins wereidentified by anti-annexin II monoclonal antibody by immunoblot-ting. Immunoblot analysis identified tPA bound protein as annexin IIindicating that tPA synthesized by MDA-MB231 cells binds to annexinII (Fig. 1C). These results were confirmed by a converse experimentwhere lysate was immunoprecipitated by anti-annexin II monoclonalantibody and bound protein was recognized by anti-tPA antibodyconfirming that tPA binds to annexin II (Fig. 1D).
tPA binding to cell surface annexin II is required for plasminogenactivation in breast cancer
The assembly of tPA and plasminogen on the cell surface annexin IIfacilitates conversion of inactive plasminogen to reactive enzymeplasmin. To test whether interaction of tPA and annexin II in MDA-MB231 cells are physiologically active, we assayed plasminogenactivation in tPA bound live cells. MDA-MB231 cells were incubatedwith recombinant tPA and analyzed for cell surface binding. UnboundtPA was removed by washing. In parallel experiments non-invasiveMCF-7 cells that lack annexin II expression were used as control.Specific tPA binding was determined by its ability to convertplasminogen to plasmin. Indeed, tPA bound to the MDA-MB231 cellsurface and was able to activate plasminogen into plasmin in a time-dependent manner. As expected, MCF-7 cells lacking annexin IIexpression therefore tPA were unable to bind and consequently failedto activate plasminogen (Fig. 2A). To further confirm if cell surfaceinteraction of annexin II and tPA is critical to activate plasminogen, wesilenced annexin II gene in MDA-MB231 cells (Fig. 3A).These annexinII silenced cells were analyzed for their ability to bind tPA and theircapacity to activate plasminogen. Results shown in Fig. 3C indicatethat annexin II silenced MDA-MB231 cells were unable to bind tPA.Therefore activation of plasminogen was significantly inhibited. Incontrast, wild type cells bound with recombinant tPA that activatedplasminogen to plasmin efficiently (Fig. 3C). Collectively, these resultsstrongly suggest that MDA-MB231 cell surface annexin II is requiredfor tPA binding and is required for efficient plasmin generation.
Annexin II mediated plasmin generation is crucial for cell migration
Cell motility is critically involved in metastasis, neoangiogenesisand tumor invasion (Stack et al., 1999). It has been reported thatplasmin is a strong serine protease and known to hydrolyze ECM andbasement membrane (BM) proteins and thereby facilitates cellularmigration, invasion and angiogenesis (Bajou et al., 2001; Diaz et al.,2002; Pepper, 2001; Tarui et al., 2002). Several studies wereperformed to determine the ability of annexin II to influence breastcancer cell migration. First we used a Boyden chamber assay modeland tested if annexin II dependent plasmin generation is required forMDA-MB231 cell migration. The results of this experiment revealedthat when cells were incubated with plasminogen they activatedplasminogen to plasmin which in turn degraded ECM proteins andinvaded across the membrane (Fig. 2B). This cell migration wassignificantly blocked by anti-annexin II antibody (Fig. 2B) suggestingthat annexin II is involved in plasmin generation and its dependent
cell migration. Additional experiments were performed to confirmthat annexin II-dependent plasmin generation plays a critical rolein breast cancer cell migration. Previously we have shown that
283M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
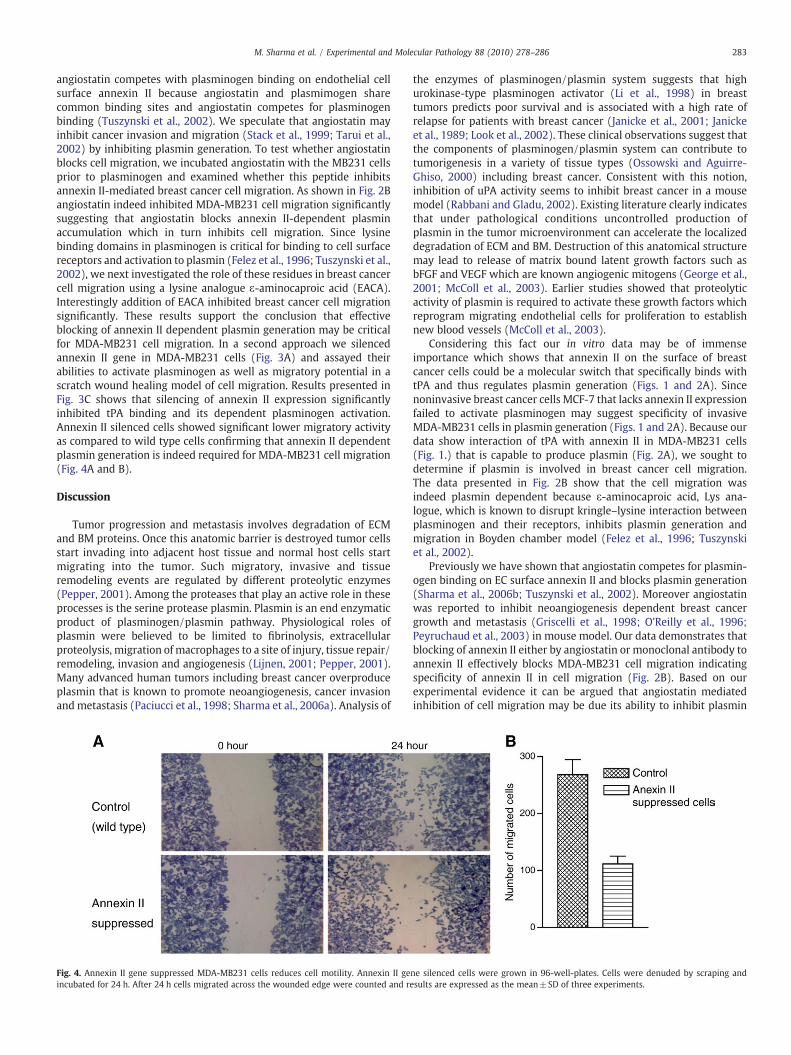
angiostatin competes with plasminogen binding on endothelial cellsurface annexin II because angiostatin and plasmimogen sharecommon binding sites and angiostatin competes for plasminogenbinding (Tuszynski et al., 2002). We speculate that angiostatin mayinhibit cancer invasion and migration (Stack et al., 1999; Tarui et al.,2002) by inhibiting plasmin generation. To test whether angiostatinblocks cell migration, we incubated angiostatin with the MB231 cellsprior to plasminogen and examined whether this peptide inhibitsannexin II-mediated breast cancer cell migration. As shown in Fig. 2Bangiostatin indeed inhibited MDA-MB231 cell migration significantlysuggesting that angiostatin blocks annexin II-dependent plasminaccumulation which in turn inhibits cell migration. Since lysinebinding domains in plasminogen is critical for binding to cell surfacereceptors and activation to plasmin (Felez et al., 1996; Tuszynski et al.,2002), we next investigated the role of these residues in breast cancercell migration using a lysine analogue ε-aminocaproic acid (EACA).Interestingly addition of EACA inhibited breast cancer cell migrationsignificantly. These results support the conclusion that effectiveblocking of annexin II dependent plasmin generation may be criticalfor MDA-MB231 cell migration. In a second approach we silencedannexin II gene in MDA-MB231 cells (Fig. 3A) and assayed theirabilities to activate plasminogen as well as migratory potential in ascratch wound healing model of cell migration. Results presented inFig. 3C shows that silencing of annexin II expression significantlyinhibited tPA binding and its dependent plasminogen activation.Annexin II silenced cells showed significant lower migratory activityas compared to wild type cells confirming that annexin II dependentplasmin generation is indeed required for MDA-MB231 cell migration(Fig. 4A and B).
Discussion
Tumor progression and metastasis involves degradation of ECMand BM proteins. Once this anatomic barrier is destroyed tumor cellsstart invading into adjacent host tissue and normal host cells startmigrating into the tumor. Such migratory, invasive and tissueremodeling events are regulated by different proteolytic enzymes(Pepper, 2001). Among the proteases that play an active role in theseprocesses is the serine protease plasmin. Plasmin is an end enzymaticproduct of plasminogen/plasmin pathway. Physiological roles ofplasmin were believed to be limited to fibrinolysis, extracellularproteolysis, migration of macrophages to a site of injury, tissue repair/remodeling, invasion and angiogenesis (Lijnen, 2001; Pepper, 2001).Many advanced human tumors including breast cancer overproduceplasmin that is known to promote neoangiogenesis, cancer invasionandmetastasis (Paciucci et al., 1998; Sharma et al., 2006a). Analysis of
Fig. 4. Annexin II gene suppressed MDA-MB231 cells reduces cell motility. Annexin II geincubated for 24 h. After 24 h cells migrated across the wounded edge were counted and re
the enzymes of plasminogen/plasmin system suggests that highurokinase-type plasminogen activator (Li et al., 1998) in breasttumors predicts poor survival and is associated with a high rate ofrelapse for patients with breast cancer (Janicke et al., 2001; Janickeet al., 1989; Look et al., 2002). These clinical observations suggest thatthe components of plasminogen/plasmin system can contribute totumorigenesis in a variety of tissue types (Ossowski and Aguirre-Ghiso, 2000) including breast cancer. Consistent with this notion,inhibition of uPA activity seems to inhibit breast cancer in a mousemodel (Rabbani and Gladu, 2002). Existing literature clearly indicatesthat under pathological conditions uncontrolled production ofplasmin in the tumor microenvironment can accelerate the localizeddegradation of ECM and BM. Destruction of this anatomical structuremay lead to release of matrix bound latent growth factors such asbFGF and VEGF which are known angiogenic mitogens (George et al.,2001; McColl et al., 2003). Earlier studies showed that proteolyticactivity of plasmin is required to activate these growth factors whichreprogram migrating endothelial cells for proliferation to establishnew blood vessels (McColl et al., 2003).
Considering this fact our in vitro data may be of immenseimportance which shows that annexin II on the surface of breastcancer cells could be a molecular switch that specifically binds withtPA and thus regulates plasmin generation (Figs. 1 and 2A). Sincenoninvasive breast cancer cells MCF-7 that lacks annexin II expressionfailed to activate plasminogen may suggest specificity of invasiveMDA-MB231 cells in plasmin generation (Figs. 1 and 2A). Because ourdata show interaction of tPA with annexin II in MDA-MB231 cells(Fig. 1.) that is capable to produce plasmin (Fig. 2A), we sought todetermine if plasmin is involved in breast cancer cell migration.The data presented in Fig. 2B show that the cell migration wasindeed plasmin dependent because ε-aminocaproic acid, Lys ana-logue, which is known to disrupt kringle–lysine interaction betweenplasminogen and their receptors, inhibits plasmin generation andmigration in Boyden chamber model (Felez et al., 1996; Tuszynskiet al., 2002).
Previously we have shown that angiostatin competes for plasmin-ogen binding on EC surface annexin II and blocks plasmin generation(Sharma et al., 2006b; Tuszynski et al., 2002). Moreover angiostatinwas reported to inhibit neoangiogenesis dependent breast cancergrowth and metastasis (Griscelli et al., 1998; O'Reilly et al., 1996;Peyruchaud et al., 2003) in mouse model. Our data demonstrates thatblocking of annexin II either by angiostatin or monoclonal antibody toannexin II effectively blocks MDA-MB231 cell migration indicatingspecificity of annexin II in cell migration (Fig. 2B). Based on ourexperimental evidence it can be argued that angiostatin mediatedinhibition of cell migration may be due its ability to inhibit plasmin
ne silenced cells were grown in 96-well-plates. Cells were denuded by scraping andsults are expressed as the mean±SD of three experiments.
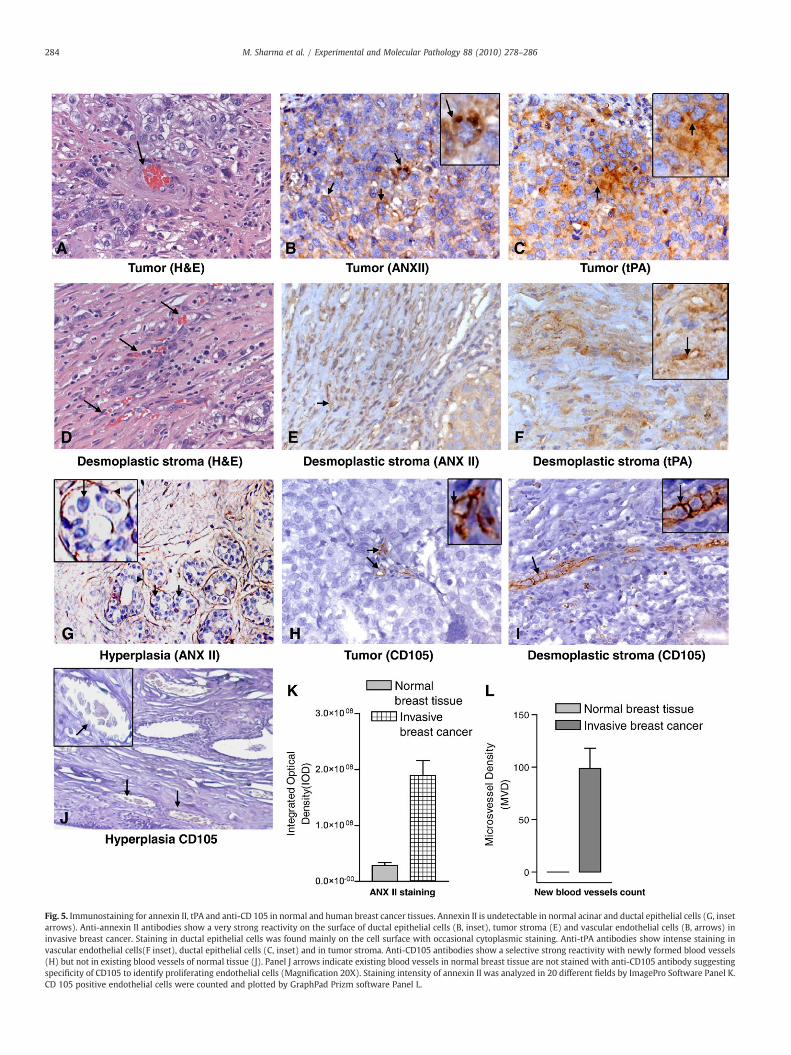
Fig. 5. Immunostaining for annexin II, tPA and anti-CD 105 in normal and human breast cancer tissues. Annexin II is undetectable in normal acinar and ductal epithelial cells (G, insetarrows). Anti-annexin II antibodies show a very strong reactivity on the surface of ductal epithelial cells (B, inset), tumor stroma (E) and vascular endothelial cells (B, arrows) ininvasive breast cancer. Staining in ductal epithelial cells was found mainly on the cell surface with occasional cytoplasmic staining. Anti-tPA antibodies show intense staining invascular endothelial cells(F inset), ductal epithelial cells (C, inset) and in tumor stroma. Anti-CD105 antibodies show a selective strong reactivity with newly formed blood vessels(H) but not in existing blood vessels of normal tissue (J). Panel J arrows indicate existing blood vessels in normal breast tissue are not stained with anti-CD105 antibody suggestingspecificity of CD105 to identify proliferating endothelial cells (Magnification 20X). Staining intensity of annexin II was analyzed in 20 different fields by ImagePro Software Panel K.CD 105 positive endothelial cells were counted and plotted by GraphPad Prizm software Panel L.
284 M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286

285M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
generation by blocking plasminogen/tPA binding on cell surfaceannexin II (Brownstein et al., 2004; Stack et al., 1999; Tuszynski et al.,2002). Silencing of annexin II gene in MDA-MB231 cells inhibits theirability to bind tPA and plasminogen activation (Fig. 3). Furthermorethese cells were less migratory as compared to wild type in a scratchwound healing model of migration assay (Fig. 4). Based on our data itis not surprising that Shiozawa et al. have found involvement ofannexin II in adhesion, migration and homing of prostate cancer cells(Shiozawa et al., 2008). Together, these in vitro and in vivoexperimental evidence links a potential role of annexin II in breastcancer.
It is possible that plasmin mediated destruction of the anatomicbarrier of ECM and BM could support EC /cancer cell migration (Taruiet al., 2002) that may be the key for new blood vessel formation tosupport tumor expansion andmetastasis (Ling et al., 2004;McColl et al.,2003; Semov et al., 2005).Our observations are further supported by thefacts that tumors failed to growonplasminogen(PLG-/-) geneknockoutmice and unable to metastasize because in absence of plasmin tumor'sability to fromnewbloodvesselswas significantly inhibited (Bajou et al.,2001; Perides et al., 2006). Consistently a number of prior articles havedescribed that direct inhibition of plasmin suppresses experimentalmetastasis, neoangiogenesis and tumor progression in Lewis LungCarcinoma (LLC) model (Bugge et al., 1997; Tanaka et al., 1982).
Taken together, our data provides strong evidence that annexin II-dependent plasmin generation is required for breast cancer cellmigration. It seems that annexin II is a regulatory switch for continuousplasmin generation in the tumor microenvironment and might explainthe presence of annexin II on the cell surface of many different types ofhuman cancers.
These data raised the possibility that annexin II dependent plasmingeneration may be an endogenous regulator of neoangiogenesis (Linget al., 2004). To investigate this we analyzed human breast tissuesamples by immunohistochemical staining. Immunohistochemicalstudies revealed expression of ANX II mainly on the surface of invasivecancerous cells (Fig. 5B). In contrast, epithelial cells in normal ductswere completely negative (Fig. 5G). Consistent with annexin IIexpression, immunostaining of thin serial sections also revealed highlevels of tPA in tumor stroma (Fig. 5C). In agreement with our findingsearlier clinical studies have also noticed elevated levels of tPA in theplasma of human breast cancer (Grondahl-Hansen et al., 1990; Rellaet al., 1993), glioblastoma and pancreatic cancers (Aguilar et al., 2004;Goh et al., 2005). Careful investigation of immunostained sectionsrevealed collagenous stroma composed of fibroblasts, macrophages,lymphocytes and ECM indicating an inflammatory response andannexin II in desmoplastic stroma (Fig. 5E) suggesting chronicinflammation in the tumor microenvironment and may be indicativeof neoangiogenic activity. Next, we investigated neoangiogenic activityin the serial sections of same tissue using anti-CD105, selective markerfor sprouting neovessels (Gerber et al., 2006; Kumar et al., 1999).Staining pattern of CD105 revealed numerous sprouting neovessels intumor aswell indesmoplastic stromaconfirmingneoangiogenic activity(Fig. 5H and I). Quantitative analysis of neoangiogenic activity asmeasured by micro vascular density (MVD) appears to correlate withannexin II expression patterns (Fig. 5K and L).
Based on our in vitro and in vivo data, it is quite likely that over-expression of annexin II in human breast cancer regulates localplasmin generation which in turn degrades ECM and BM. Hydrolysisof matrix components by plasmin may release and activate matrixbound inactive angiogenic cytokine VEGF and may induce neoangio-genic activity (McColl et al., 2003). This could be an early event in thetumor microenvironment switching breast cancer from prevascularphase to the vascular phase (Ling et al., 2004) which is characterizedby aggressive phase with the possibility of metastasis. Consistent withour observations neoangiogenic activity reported to be a significantprognostic indicator in early stage of breast cancer (Weidner et al.,1992).
Collectively, the data reported hear and previously publishedreports strongly support the concept that annexin II on tumor/ECmaybe an angiogenic switch. This may be a potential reason why annexinII is being repeatedly identified as widely overexpressed protein innumber of cancers and strongly associated with prognostic signifi-cance in metastasis (Brichory et al., 2001; Chuthapisith et al., 2009;Guedj et al., 2009; Pei et al., 2007; Roseman et al., 1994; Sharma et al.,2006a; Sharma and Sharma, 2007; Shiozawa et al., 2008; Singh, 2007;Syed et al., 2007; Wu et al., 2002; Zhong et al., 2009; Zimmermannet al., 2004). Whether annexin II could be a valuable target in inhib-iting neoangiogenic activity and breast cancer in vivo is in interestingbut unanswered question that is currently under investigation in ourlaboratory.
Acknowledgments
This study was supported in part by grants from PennsylvaniaDepartment of Public Health and Department of Defense Idea award #W81XWH-07-1-0424 and Concept award # DAMD1703-1-0761 toMCS. We thank Chalon Saunders and Miriam Smith for technicalassistance.
References
Aguilar, S., et al., 2004. Tissue plasminogen activator in murine exocrine pancreas cancer:selective expression in ductal tumors and contribution to cancer progression. Am. J.Pathol. 165, 1129–1139.
Bajou, K., et al., 2001. The plasminogen activator inhibitor PAI-1 controls in vivo tumorvascularization by interaction with proteases, not vitronectin. Implications forantiangiogenic strategies. J. Cell Biol. 152, 777–784.
Brichory, F.M., et al., 2001. An immune response manifested by the common occurrenceof annexins I and II autoantibodies and high circulating levels of IL-6 in lung cancer.Proc. Natl. Acad. Sci. U. S. A. 98, 9824–9829.
Brownstein, C., et al., 2004. Annexin II mediates plasminogen-dependent matrix invasionby human monocytes: enhanced expression by macrophages. Blood 103, 317–324.
Bugge, T.H., et al., 1997. Growth and dissemination of Lewis lung carcinoma inplasminogen-deficient mice. Blood 90, 4522–4531.
Cesarman, G.M., et al., 1994. An endothelial cell receptor for plasminogen/tissueplasminogen activator (t-PA). II. Annexin II-mediated enhancement of t-PA-dependent plasminogen activation. J. Biol. Chem. 269, 21198–21203.
Chernicky, C.L., et al., 2005. Tissue-type plasminogen activator is upregulated inmetastaticbreast cancer cells exposed to insulin-like growth factor-I. Clin. Breast Cancer 6,340–348.
Chuthapisith, S., et al., 2009. Annexins in human breast cancer: possible predictors ofpathological response to neoadjuvant chemotherapy. Eur. J. Cancer 45, 1274–1281.
Diaz, V.M., et al., 2004. Specific interaction of tissue-type plasminogen activator (t-PA)with annexin II on the membrane of pancreatic cancer cells activates plasminogenand promotes invasion in vitro. Gut 53, 993–1000.
Diaz, V.M., et al., 2002. Tissue plasminogen activator is required for the growth, invasion,and angiogenesis of pancreatic tumor cells. Gastroenterology 122, 806–819.
Felez, J., et al., 1996. Characterization of cellular binding sites and interactive regionswithin reactants required for enhancement of plasminogen activation by tPA on thesurface of leukocytic cells. Thromb. Haemost. 76, 577–584.
George, S.J., et al., 2001. Plasmin-mediated fibroblast growth factor-2 mobilisationsupports smooth muscle cell proliferation in human saphenous vein. J. Vasc. Res. 38,492–501.
Gerber, S.A., et al., 2006. Preferential attachment of peritoneal tumor metastases toomental immune aggregates and possible role of a unique vascular microenviron-ment in metastatic survival and growth. Am. J. Pathol. 169, 1739–1752.
Goh, K.Y., et al., 2005. Tissue plasminogen activator expression in meningiomas andglioblastomas. Clin. Neurol. Neurosurg. 107, 296–300.
Griscelli, F., et al., 1998. Angiostatin gene transfer: inhibition of tumor growth in vivo byblockage of endothelial cell proliferation associatedwith amitosis arrest. Proc. Natl.Acad. Sci. U. S. A. 95, 6367–6372.
Grondahl-Hansen, J., et al., 1990. Tissue-type plasminogen activator in plasma frombreastcancer patients determined by enzyme-linked immunosorbent assay. Br. J. Cancer. 61,412–414.
Guedj, N., et al., 2009. Comparative protein expression profiles of hilar and peripheralhepatic cholangiocarcinomas. J. Hepatol. 51, 93–101.
Hajjar, K.A., et al., 1994. An endothelial cell receptor for plasminogen/tissueplasminogen activator. I. Identity with annexin II. J. Biol. Chem. 269, 21191–21197.
Janicke, F., et al., 2001. Randomized adjuvant chemotherapy trial in high-risk, lymphnode-negative breast cancer patients identified by urokinase-type plasminogen activatorand plasminogen activator inhibitor type 1. J. Natl. Cancer Inst. 93, 913–920.
Janicke, F., et al., 1989. Urokinase-type plasminogen activator antigen and early relapsein breast cancer. Lancet 2, 1049.
Kumar, S., et al., 1999. Breast carcinoma: vascular density determined using CD105antibody correlates with tumor prognosis. Cancer Res. 59, 856–861.

286 M. Sharma et al. / Experimental and Molecular Pathology 88 (2010) 278–286
Li, H., et al., 1998. Adenovirus-mediated delivery of a uPA/uPAR antagonist suppressesangiogenesis-dependent tumor growth and dissemination in mice. Gene Ther. 5,1105–1113.
Lijnen, H.R., 2001. Plasmin and matrix metalloproteinases in vascular remodeling.Thromb. Haemost. 86, 324–333.
Ling, Q., et al., 2004. Annexin II regulates fibrin homeostasis and neoangiogenesis invivo. J. Clin. Invest. 113, 38–48.
Look, M.P., et al., 2002. Pooled analysis of prognostic impact of urokinase-typeplasminogen activator and its inhibitor PAI-1 in 8377 breast cancer patients. J. Natl.Cancer Inst. 94, 116–128.
McColl, B.K., et al., 2003. Plasmin activates the lymphangiogenic growth factors VEGF-Cand VEGF-D. J. Exp. Med. 198, 863–868.
Mignatti, P., Rifkin, D.B., 1993. Biology and biochemistry of proteinases in tumor invasion.Physiol. Rev. 73, 161–195.
O'Reilly, M.S., et al., 1996. Angiostatin induces and sustains dormancy of human primarytumors in mice. Nat. Med. 2, 689–692.
Ohno, Y., et al., 2009. Annexin II represents metastatic potential in clear-cell renal cellcarcinoma. Br. J. Cancer 101, 287–294.
Ossowski, L., Aguirre-Ghiso, J.A., 2000. Urokinase receptor and integrin partnership:coordination of signaling for cell adhesion, migration and growth. Curr. Opin. CellBiol. 12, 613–620.
Paciucci, R., et al., 1998. The plasminogen activator system in pancreas cancer: role of t-PA in the invasive potential in vitro. Oncogene 16, 625–633.
Pei, H., et al., 2007. Proteome analysis and tissue microarray for profiling proteinmarkers associated with lymph node metastasis in colorectal cancer. J. ProteomeRes. 6, 2495–2501.
Pepper, M.S., 2001. Extracellular proteolysis and angiogenesis. Thromb. Haemost. 86,346–355.
Perides, G., et al., 2006. The fibrinolytic system facilitates tumor cell migration across theblood-brain barrier in experimental melanoma brain metastasis. BMC Cancer 6, 56.
Peyruchaud, O., et al., 2003. Angiostatin inhibits bone metastasis formation in nudemice through a direct anti-osteoclastic activity. J. Biol. Chem. 278, 45826–45832.
Rabbani, S.A., Gladu, J., 2002. Urokinase receptor antibody can reduce tumor volume anddetect the presence of occult tumor metastases in vivo. Cancer Res. 62, 2390–2397.
Rella, C., et al., 1993. Tissue-type plasminogen activator as marker of functional steroidreceptors in human breast cancer. Thromb. Res. 69, 209–220.
Roseman, B.J., et al., 1994. Annexin II marks astrocytic brain tumors of high histologicgrade. Oncol. Res. 6, 561–567.
Semov, A., et al., 2005. Metastasis-associated protein S100A4 induces angiogenesisthrough interaction with Annexin II and accelerated plasmin formation. J. Biol.Chem. 280, 20833–20841.
Sharma, M., et al., 2006a. Angiogenesis Associated Protein Annexin II in Breast Cancer:Selective Expression in Invasive Breast Cancer and Contribution to Tumor Invasionand Progression. Exp. Mol. Pathol. 81, 146–156.
Sharma, M., et al., 2006b. Antibody-directed targeting of angiostatin's receptorannexin II inhibits Lewis Lung Carcinoma tumor growth via blocking ofplasminogen activation: Possible biochemical mechanism of angiostatin's action.Exp. Mol. Pathol. 81, 136–145.
Sharma, M.C., Sharma, M., 2007. The role of annexin II in angiogenesis and tumorprogression: a potential therapeutic target. Curr. Pharm. Des. 13, 3568–3575.
Sharma, M.R., et al., 2004. Angiostatin-induced inhibition of endothelial cellproliferation/apoptosis is associated with the down-regulation of cell cycleregulatory protein cdk5. J. Cell. Biochem. 91, 398–409.
Shiozawa, Y., et al., 2008. Annexin II/annexin II receptor axis regulates adhesion,migration, homing, and growth of prostate cancer. J. Cell. Biochem. 105, 370–380.
Singh, P., 2007. Role of Annexin-II in GI cancers: interaction with gastrins/progastrins.Cancer Lett. 252, 19–35.
Stack, M.S., et al., 1999. Angiostatin inhibits endothelial andmelanoma cellular invasionby blocking matrix-enhanced plasminogen activation. Biochem. J. 340, 77–84.
Stephens, R.W., et al., 1998. The urokinase plasminogen activator system as a target forprognostic studies in breast cancer. Breast Cancer Res. Treat. 52, 99–111.
Syed, S.P., et al., 2007. Angiostatin receptor annexin II in vascular tumors includingangiosarcoma. Hum. Pathol. 38, 508–513.
Tanaka, N., et al., 1982. Ultrastructural study of the effects of tranexamic acid andurokinase on metastasis of Lewis lung carcinoma. Br. J. Cancer 46, 428–435.
Tarui, T., et al., 2002. Plasmin-induced migration of endothelial cells. A potential targetfor the anti-angiogenic action of angiostatin. J. Biol. Chem. 277, 33564–33570.
Terranova, V.P., et al., 1986. Basement membrane and the invasive activity of metastatictumor cells. J. Natl. Cancer Inst. 77, 311–316.
Tuszynski, G.P., et al., 2002. Angiostatin binds to tyrosine kinase substrate annexinII through the lysine-binding domain in endothelial cells. Microvasc. Res. 64,448–462.
Weidner, N., et al., 1992. Tumor angiogenesis: a newsignificant and independent prognosticindicator in early-stage breast carcinoma. J. Natl. Cancer Inst. 84, 1875–1887.
Wu, W., et al., 2002. Identification and validation of metastasis-associated proteins inhead and neck cancer cell lines by two-dimensional electrophoresis and massspectrometry. Clin. Exp. Metastasis 19, 319–326.
Zhong, L.P., et al., 2009. Increased expression of Annexin A2 in oral squamous cellcarcinoma. Arch. Oral Biol. 54, 17–25.
Zimmermann, U., et al., 2004. Expression of annexin II in conventional renal cellcarcinoma is correlated with Fuhrman grade and clinical outcome. Virchows Arch.445, 368–374.





























