Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Plant Physiology and Biochemistry 60 (2012) 157e164
Contents lists available
Plant Physiology and Biochemistry
journal homepage: www.elsevier .com/locate/plaphy
Research article
Brassica juncea nitric oxide synthase like activity is stimulated by PKC activatorsand calcium suggesting modulation by PKC-like kinase
Pooja Saigal Talwar, Ravi Gupta, Arun Kumar Maurya 1, Renu Deswal*
Molecular Plant Physiology and Proteomics Laboratory, Department of Botany, University of Delhi, Delhi 110007, India
a r t i c l e i n f o
Article history:Received 8 June 2012Accepted 8 August 2012Available online 21 August 2012
Keywords:BjNOSlaL-NAMENBTNitric oxideNitric oxide synthasePhosphorylationProtein kinase-C
Abbreviations: 1,2-DG6, 1,2-dihexanoylglycerol;didecanoate; BH4, tetrahydrobiopterin; BjNOSla, Brasthase like activity; CaM, calmodulin; EGTA, ethylenNAME, N-nitro-L-Arg-methyl ester; NBT, nitro blue tPKC, protein kinase-C; PMA, phorbol 12-myristate ace* Corresponding author. Tel.: þ91 11 27667573; fax
E-mail address: [email protected] (R. Desw1 Present address: Department of Botany, Hans Raj
Delhi 110007, India.
0981-9428/$ e see front matter � 2012 Elsevier Mashttp://dx.doi.org/10.1016/j.plaphy.2012.08.005
a b s t r a c t
Nitric oxide (NO) is an important signaling molecule having varied physiological and regulatory roles inbiological systems. The fact that nitric oxide synthase (NOS) is responsible for NO generation in animals,prompted major search for a similar enzyme in plants. Arginine dependent NOS like activity (BjNOSla)was detected in Brassica juncea seedlings using oxyhemoglobin and citrulline assays. BjNOSla showed25% activation by NADPH (0.4 mM) and 40% by calcium (0.4 mM) but the activity was flavin mono-nucleotide (FMN), flavin dinucleotide (FAD) and calmodulin (CaM) independent. Pharmacologicalapproach using mammalian NOS inhibitors, NBT (300 mM) and L-NAME (5 mM), showed significantinhibition (100% and 67% respectively) supporting that the BjNOSla operates via the oxidative pathway.Most of the BjNOSla activity (80%) was confined to shoot while root showed only 20% activity. Locali-zation studies by NADPH-diaphorase and DAF-2DA staining showed the presence of BjNOSla in guardcells. Kinetic analysis showed positive cooperativity with calcium as reflected by a decreased Km (w13%)and almost two fold increase in Vmax. PMA (438 nM), a kinase activator, activated BjNOSla w1.9 foldwhile its inactive analog 4aPDD was ineffective. Calcium and PMA activated the enzyme to w3 folds.Interestingly, 1,2-DG6 (2.5 mM) and PS (1 mM) with calcium activated the enzyme activity to w7 fold. Asignificant inhibition of BjNOSla by PKC inhibitors-staurosporine (w90%) and calphostin-C (w40%),further supports involvement of PKC-like kinase. The activity was also enhanced by abiotic stressconditions (7e46%). All these findings suggest that BjNOSla generates NO via oxidative pathway and isprobably regulated by phosphorylation.
� 2012 Elsevier Masson SAS. All rights reserved.
1. Introduction
NO is involved as a signaling and regulatorymolecule in an arrayof physiological processes including seed germination, root devel-opment, stomatal movements, flowering, senescence and pro-grammed cell death [1]. Besides, NO evolution during differentabiotic and biotic stress conditions (such as temperature, salinity,drought, heavy metal stress and pathogen attack) is well estab-lished [1]. Furthermore, NO also functions as a protective moleculeagainst reactive oxygen species (ROS) [1].
4aPDD, 4a-phorbol 12,13-sica juncea nitric oxide syn-e glycol tetraacetic acid; L-etrazolium; NO, nitric oxide;tate; PS, phosphatidylserine.: þ91 11 27667829.al).College, University of Delhi,
son SAS. All rights reserved.
NO is produced by both enzymatic and non-enzymatic reac-tions. In animals, enzymatic NO production is mediated by NADPHdependent Nitric Oxide synthase (NOS), which catalyzes theoxidation of L-arginine to L-citrulline and NO is evolved as a by-product. Animal NOS occur in three homodimeric isoforms-endothelial NOS, neuronal NOS and inducible NOS [2]. NOproduction in plants was first observed in herbicide treatedsoybean by Klepper in 1979 [3] but its role in plant defence wasestablished by Durner [4] and Delledonne [5] in 1998. The fact thatNO production in animals is mediated by NOS led to the search ofsimilar enzyme in plants. NOS like activity has been reported inbacteria [6], slime mold (Physarum polycephalum) [7], algae(Ostreococcus tauri, Ostreococcus lucimarinus) [8] gymnosperms(Taxus brevifolia) and higher plants like Nicotiana tabacum, Glycinemax, Pisum sativum, Mucuna hasszoo, Lupinus albus, Arabidopsisthaliana, Zea mays [9] and Chorispora bungeana [10].
NO generation in plants have been detected either by in planta(citrulline assay, oxyhemoglobin assay, Griess reaction, DAF dye,ESR, NO electrode and Mass spectrometry) or ex planta (chem-iluminescence and laser based infrared spectroscopy) assays [11].

P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164158
The combination of biochemical and molecular approach alongwith mutant analysis led to the identification of NO generatingenzyme AtNOS1 in A. thaliana [12]. Later, it was found to bea circularly permuted plastid GTPase and was renamed as NOassociated protein (AtNOA1) as it did not exhibit NOS like activity[13]. Recently, an L-arginine dependent NOS with 45% sequencehomology with human endothelial NOS was purified from an alga,O. tauri [8]. In spite of all these studies, purification of NOS from anyplant is still eluding researchers; therefore it is important tounderstand its biochemical properties including its regulation todevice a successful purification strategy.
Phosphorylation regulates NO mediated calcium release inguard cells of Arabidopsis [14] and Vicia faba [15]. The calciumrelease was reversibly suppressed by protein kinase antagoniststaurosporine and K252A but not by tyrosine kinase antagonistgenistein. Although, the role of protein phosphorylation in calciumrelease and regulating Kþ and Cl� channel is shown in Arabidopsisguard cells, but whether phosphorylation modulates NOS likeactivity is not known. There are few reports suggesting theinvolvement of calcium and protein kinases as upstream signal forNO synthesis in plants [16].
The aim of this studywas to detect and characterize the NOS likeactivity in Brassica juncea and its regulation.
2. Results
2.1. Optimization of NOS like activity by the oxyhemoglobin assay
B. juncea seedlings were homogenized in the extraction bufferas described in methods. For optimization of BjNOSla activityassay, freshly prepared oxyhemoglobin was used. BjNOSla activitywas assayed in 65 mM Hepes buffer, pH 7.4 and other reactioncomponents were varied independently to optimize the reactionconditions (Fig. 1AeD). Maximum activity was observed at 3 mML-arginine, the substrate (Fig. 1A). Oxyhemoglobin (10 mL) showed40% higher activity than control (Fig. 1B). To know the divalentcation requirement of BjNOSla, the assay was performed withvarying concentration of MgCl2 and CaCl2. MgCl2 (10 mM) showed
Fig. 1. Optimization of oxyhemoglobin assay for analyzing NOS like activity in B. juncea seeHepes buffer pH 7.4. NOS activity with (A) substrate L-arginine (0e4 mM), (B) oxyhemoglobiError bars represents standard deviation from ten biological replicates. Asterisk showed sig
w57% activation (Fig. 1C) while CaCl2 showed 40% increase in theBjNOSla at 0.4 mM (Fig. 1D). Respective optimized concentrationsof the assay components (3 mM L-arginine, 10 mL oxyhemoglobin,10 mM MgCl2 and 0.4 mM CaCl2) were used for further experi-mentation. The specific activity of rat neuronal NOS, used aspositive control, was calculated as w1.7 nmol min�1 mg�1 usingthis assay indicating good efficacy/efficiency of the oxyhemo-globin assay. The BjNOSla was completely abolished by 2 mMEGTA, suggesting that calcium is essential for the activity. Cofac-tors NADPH (0.4 mM) and BH4 (4 mM) led to 25% and 10% acti-vation respectively (data not shown). However, FMN (1e40 mM),FAD (1e40 mM) and CaM (0.5e1 mM) were ineffective (data notshown).
2.2. Mammalian NOS inhibitors inhibit NOS like activity
Inhibitors of mammalian NOS, NBT (300 mM) and L-NAME(5 mM) led to 100% and 67% inhibition of NOS like activity inB. juncea, suggesting similarity between mammalian NOS andBjNOSla (Fig. 2A and B). NBT was able to inhibit 100% activity evenat 1/10th concentration of L-NAME suggesting it to be amore potentinhibitor of BjNOSla.
2.3. BjNOSla is predominantly localized in the guard cells
To observe the effect of light on BjNOSla and its distribution indifferent parts, the activity was checked in etiolated and lightgrown seedlings in shoot and root. NOS like activity was predom-inantly (about 80%) localized in shoots (Fig. 3A). It was 15% and 20%higher in light grown shoots and roots respectively in comparisonto respective etiolated control (Fig. 3A).
NADPH-diaphorase co-localizes with the NOS [16], therefore, anattempt was made to localize the BjNOSla in the epidermal peel.Maximum NADPH-diaphorase staining was detected in the guardcells (Fig. 3B and C). To further confirm the localization of BjNOSlain the guard cells, DAF-2DA, a NO sensitive fluorophore was used.DAF-2DA showed strongest intensity with the substrate, L-arginine(Fig. 3I), further supporting the involvement of an argininedependent enzymatic source in NO biosynthesis.
dlings. The assay was performed by varying only one component at a time in 65 mMn (0e20 mL), (C) magnesium chloride (0e40 mM) and (D) calcium chloride (0e0.6 mM).nificant difference in comparison with control (Student’s t-test, p < 0.05).

Fig. 2. Effect of NOS inhibitors on BjNOSla. The activity was assayed with NOSinhibitors (A) NBT (1e300 mM) and (B) L-NAME (1e0.5 mM). Error bars representsstandard deviation from four biological replicates with each assay performed induplicates. Asterisk showed significant difference in comparison with control(Student’s t-test, p < 0.05).
P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164 159
L-NAME brought back the DAF-2DA fluorescence to the controllevel while NBT inhibited it further (Fig. 3J and K). These resultswere in consonance with the results of in vitro inhibition studies asdescribed in Fig. 2A and B.
2.4. BjNOSla shows positive cooperativity with calcium
The kinetic properties of BjNOSla were analyzed using bothoxyhemoglobin and citrulline assays. With oxyhemoglobin assay,the Km was 9.43 mM and Vmax was 2.43 pmol min�1 mg�1 withoutcalcium. However, Km decreased to 7.407 mMwhile Vmax increasedby 2 folds to 4.76 pmol min�1 mg�1 with calcium. Using radioactivecitrulline assay, MichaeliseMenten curve of L-[U-14C] arginineversus NOS like activity showed Km 14.28 mM and Vmax3 pmol min�1 mg�1 without calciumwhereas, a decrease in the Km
to 12.5 mM and an increase of 1.6 folds in Vmax to5.55 pmol min�1 mg�1 was observed with calcium (Table 1).Results suggest that calcium not only increased the affinity of thesubstrate for the enzyme as indicated by a decreased Km, but alsoenhanced the rate of the reaction by two fold. To further confirmthis positive cooperativity, Hill plots were plotted. The Hill coeffi-cient without calcium was calculated as 1 and 0.38 by oxyhemo-globin and citrulline assay respectively. However, the Hillcoefficient increased to 5.2 and 1.65 with calcium for oxyhemo-globin and citrulline assay respectively. The increase in coopera-tivity with calcium was almost 5 fold by both the assays.
2.5. PKC activators and calcium act synergistically to stimulate theNOS like activity
Earlier, NO mediated calcium release in the guard cell wasshown to be modulated by phosphorylation and only few reports
suggest the NOS modulation by phosphorylation in plants [17]. Inorder to evaluate the modulation of BjNOSla by PKC/PKC-likekinase, PKC activators (PMA, 1,2-DG6, a synthetic diacylglycerol andPS) were used. With calcium (0.4 mM) and PMA (438 nM), thestimulation of BjNOSla was 2.8 fold, whereas, 4aPDD, an inactiveanalog of PKC, was found to be ineffective (Fig. 4A). The stimulationof BjNOSla with 1,2-DG6 (1,2-dihexanoylglycerol, 2.5 mM), PS(phosphatidylserine, 1 mM) and calcium was w7 fold higher thancontrol (Fig. 4A). PMA, 1,2-DG6 and PS stimulated BjNOSla evenwithout calcium probably due to the residual calcium. Kinaseinhibitor staurosporine (20 nM) inhibited the activity by 90%(Fig. 4B) while calphostin-C (0.5 mM) inhibited it by 40% (Fig. 4C).
2.6. Abiotic stress conditions stimulate BjNOSla
To know whether NOS like enzyme contributes to NO produc-tion in response to environmental stress conditions, the BjNOSlaactivity was analyzed in different abiotic stress such as low andhigh temperature, salinity, drought and injury. LT (4 �C, 30 min)stimulated the activity by w20% (Fig. 5A) while heat treatment(50 �C, 5 h) showed 46% increase (Fig. 5B). The increase was rela-tively less (w7%) by salinity stress (0.5 MNaCl, 1 h, Fig. 5C). Droughtstress showed 35% and 22% higher activity at 30 min and 1 hrespectively (Fig. 5D). The injured seedlings showed 20% higheractivity after 30 min which remained elevated up to 1 h (Fig. 5E).Immediate effect of stress was a decline in NOS activity but acti-vationwas observed after 30minwhich continued to increase up to1 h except for HT where significant activation was observed after5 h.
These results suggest existence of NOS like activity in B. junceawhich seems somewhat similar to mammalian NOS. Furthermore,the BjNOSla showed positive cooperativity with calcium and isprobably regulated by phosphorylation.
3. Discussion
In animal system, it is a well established that NOS is mainlyresponsible for NO synthesis [18]. NO synthesis in plants is alsoknown since long [3] and reports suggest involvement of diverseenzymes such as putative NOS like enzyme, NR, Ni-NOR andxanthine oxidoreductase [18]. Search for animal like NOS led to theidentification of a unique AtNOS1 gene in Arabidopsiswhich did notexhibit NOS activity in vitro [13] and therefore was renamed asAtNOA1. Investigation revealed it to be a plastid GTPase, which ismainly responsible for the post-transcriptional regulation ofplastid-localized methylerythritol phosphate pathway enzyme [13]and ribosome assembly [18]. Recently, AtNOA1 has been shown toregulate physiological processes such as chlorophyll biosynthesisand Rubisco formation in rice [19]. AtNOA1 indeed modulates NOaccumulation in plants, possibly indirectly [20]. To date, no NOS hasbeen purified from higher plants except from some lower organ-isms such as slime mold (P. polycephalum) [7] and alga (O. tauri) [8]that have raised some hope for existence of NOS like activity/enzyme in plants.
The aim of this work was to characterize the arginine depen-dent NOS like activity in B. juncea, its modulation by variouspharmacological agents and to analyze its functional role in abioticstress. BjNOSla was detected by oxyhemoglobin and citrullineassays which was further confirmed using the mammalian NOSinhibitors.
Complete inhibition of BjNOSla by EGTA indicates that calciumplays critical role in regulation of its activity. The BjNOSla showeddependence on calcium as observed in Physarum [7], AtNOA1 [13],Lupinus [21], Pisum [22], maize [23], soybean [24], and Staphylo-coccus [25]. However, the NOS like activity from Flammulina

Table 1Showing Km, Vmax and Hill coefficient for BjNOSla, by oxyhemoglobin and citrullineassays.
Assay conditions Km (mM) Vmax (pmol min�1 mg�1) Hillcoefficient
Hb assay (�Ca) 9.43 � 103 2.43 1Hb assay (þCa) 7.4 � 103 4.76 5.2Citrulline assay (�Ca) 14.28 3 0.38Citrulline assay (þCa) 12.5 5.55 1.65
P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164160
velutipes [26] and O. tauri [7] was calcium independent. In contrastto NOS like activity reported in P. sativum, Arabidopsis, Z. mays [9]and C. bungeana [10], the BjNOSla was found to be independentof CaM, FAD and FMN. NOS like activity purified from F. velutipes[26] and O. tauri [8] was also CaM independent, supporting the CaMindependency of BjNOSla. Moreover, the NOS like activity analyzedin Brassica seedlings was comparable to the activity reported inother plants except Pisum, which showed almost 1000 times higheractivity (Table 2).
BjNOSla was significantly inhibited by mammalian NOS inhibi-tors. Nitro group (-NO2) of NBT and L-NAME competitively interactswith the active site of the NOS and therefore probably does notallow the binding of the substrate, L-arginine, leading to inhibitionof NOS activity. Inhibition of BjNOSla bymammalian NOS inhibitorsalso indicates oxidative nature of the enzyme.
After ascertaining the existence of NOS like activity in B. juncea,its localization was investigated in shoot and root which showedhigher activity in shoots than roots.
As the activity was higher in shoot, its localizationwas analyzedin the epidermal peels using NADPH-diaphorase staining. NO levelswere higher in the guard cells, supporting the earlier observationsof NO being a key regulator of stomatal closure in Arabidopsis [17]and V. faba [15]. It was reconfirmed using the fluorescent probeDAF-2DA which also showed NOS localization in guard cells con-firming that guard cells are the major sites of NO production.
Fig. 3. Localization studies of BjNOSla in Brassica seedlings. (A) The activity was tested inepidermal peel showing localization of BjNOSla in the guard cells. (DeG) bright field microscL-arginine (control), presence of L-arginine, L-NAME and NBT respectively.
However, detection of BjNOSla in roots could indicate theinvolvement of NO in signal transduction pathways. Previously, therole of NO during root initiation and development has been re-ported in pea [27], soybean [28], tomato [29], Arabidopsis [12],Cucumis sativus [30], maize [31] and Lupinus [21].
Kinetic analysis of BjNOSla using citrulline assay showed a Km of12.5 mM with calcium which is comparable to the mammalian (ratnNOS) NOS Km (8.4 mM) [32], Arabidopsis (12.5 mM) [12] and alga(O. tauri) Km (12.6 � 5 mM) [8]. The Hill coefficient for oxyhemo-globin and citrulline assay was 5.2 and 1.65 respectively. Thebinding of a ligand to a macromolecule is often enhanced if thereare already other ligands present on the same macromolecule. Avalue higher than 1 indicates that there is positive cooperativity forthe substrate resulting in higher affinity for the other ligand
shoot and root grown in light and dark. (BeC) NADPH-diaphorase activity staining ofopic images and (HeK) fluorescent microscopic images of epidermal peel in absence of
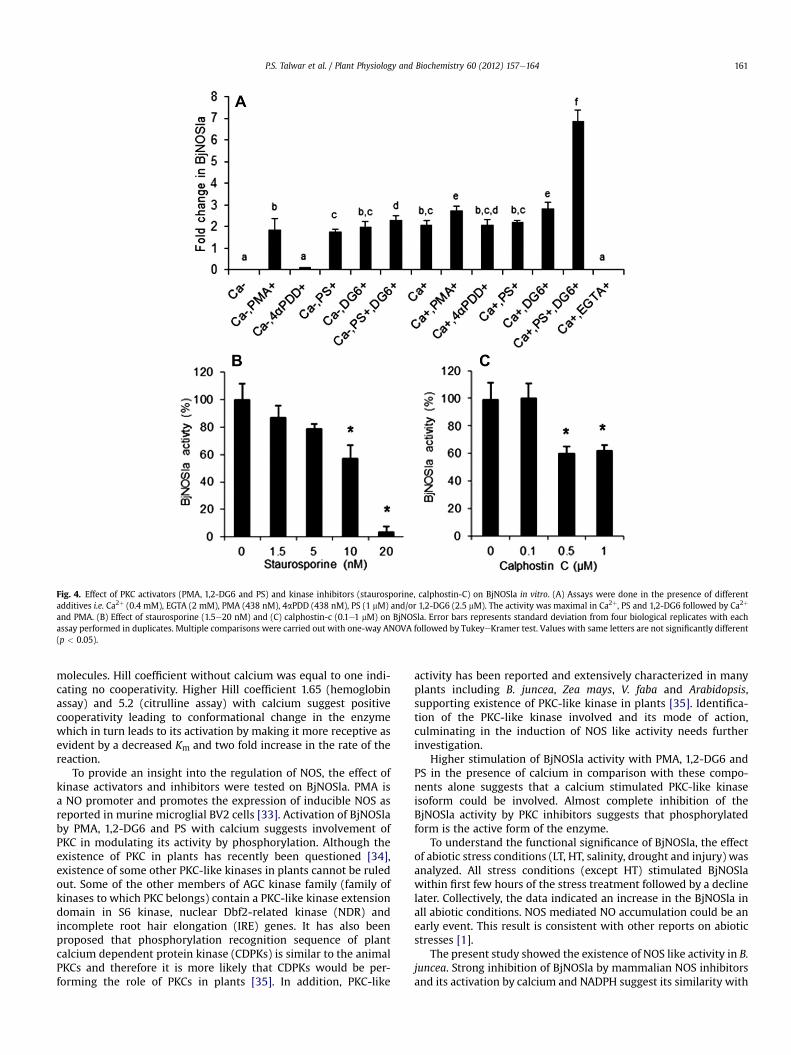
Fig. 4. Effect of PKC activators (PMA, 1,2-DG6 and PS) and kinase inhibitors (staurosporine, calphostin-C) on BjNOSla in vitro. (A) Assays were done in the presence of differentadditives i.e. Ca2þ (0.4 mM), EGTA (2 mM), PMA (438 nM), 4aPDD (438 nM), PS (1 mM) and/or 1,2-DG6 (2.5 mM). The activity was maximal in Ca2þ, PS and 1,2-DG6 followed by Ca2þ
and PMA. (B) Effect of staurosporine (1.5e20 nM) and (C) calphostin-c (0.1e1 mM) on BjNOSla. Error bars represents standard deviation from four biological replicates with eachassay performed in duplicates. Multiple comparisons were carried out with one-way ANOVA followed by TukeyeKramer test. Values with same letters are not significantly different(p < 0.05).
P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164 161
molecules. Hill coefficient without calcium was equal to one indi-cating no cooperativity. Higher Hill coefficient 1.65 (hemoglobinassay) and 5.2 (citrulline assay) with calcium suggest positivecooperativity leading to conformational change in the enzymewhich in turn leads to its activation by making it more receptive asevident by a decreased Km and two fold increase in the rate of thereaction.
To provide an insight into the regulation of NOS, the effect ofkinase activators and inhibitors were tested on BjNOSla. PMA isa NO promoter and promotes the expression of inducible NOS asreported in murine microglial BV2 cells [33]. Activation of BjNOSlaby PMA, 1,2-DG6 and PS with calcium suggests involvement ofPKC in modulating its activity by phosphorylation. Although theexistence of PKC in plants has recently been questioned [34],existence of some other PKC-like kinases in plants cannot be ruledout. Some of the other members of AGC kinase family (family ofkinases to which PKC belongs) contain a PKC-like kinase extensiondomain in S6 kinase, nuclear Dbf2-related kinase (NDR) andincomplete root hair elongation (IRE) genes. It has also beenproposed that phosphorylation recognition sequence of plantcalcium dependent protein kinase (CDPKs) is similar to the animalPKCs and therefore it is more likely that CDPKs would be per-forming the role of PKCs in plants [35]. In addition, PKC-like
activity has been reported and extensively characterized in manyplants including B. juncea, Zea mays, V. faba and Arabidopsis,supporting existence of PKC-like kinase in plants [35]. Identifica-tion of the PKC-like kinase involved and its mode of action,culminating in the induction of NOS like activity needs furtherinvestigation.
Higher stimulation of BjNOSla activity with PMA, 1,2-DG6 andPS in the presence of calcium in comparison with these compo-nents alone suggests that a calcium stimulated PKC-like kinaseisoform could be involved. Almost complete inhibition of theBjNOSla activity by PKC inhibitors suggests that phosphorylatedform is the active form of the enzyme.
To understand the functional significance of BjNOSla, the effectof abiotic stress conditions (LT, HT, salinity, drought and injury) wasanalyzed. All stress conditions (except HT) stimulated BjNOSlawithin first few hours of the stress treatment followed by a declinelater. Collectively, the data indicated an increase in the BjNOSla inall abiotic conditions. NOS mediated NO accumulation could be anearly event. This result is consistent with other reports on abioticstresses [1].
The present study showed the existence of NOS like activity in B.juncea. Strong inhibition of BjNOSla by mammalian NOS inhibitorsand its activation by calcium and NADPH suggest its similarity with

Table 2A comparative account of specific activity of NOS in different organisms.
Species Specific activity(pmol min�1 mg�1
crude protein)
Km value (mM) Reference
Rat nNOS 8.4 [32]
AngiospermsNicotiana tobaccum 0.5 N.A. [4]Lupinus albus 150 N.A. [21]Pisum sativum 5610 N.A. [22]Zea mays 3.36 (Leaves) N.A. [23]
3.88 (Root tips)Glycine max 4.5 N.A. [24]Arabidopsis 3.6 12.5 [12,38]Brassica juncea 3e13 12.5
AlgaeOstreococcus tauri 12 � 5 [8]
BacteriaStaphylococcus aureus 25.2 � 103 13.4 [25]Nocardia 150 5.7 [39]
FungiFlammulina velutipes 0.33 [26]Phycomyces blakesleeanus 1000e10,000 N.A. [40]
Slime moldPhysarum polycephalum 244 � 47 � 103
(Isoform A)66.7 � 14.5 [7]
253 � 47 � 103
(Isoform B)26.8 � 9.7
N.A. e not available.
P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164162
mammalian NOS. Its activation by PMA, 1,2-DG6 and PS (PKC acti-vators), and inhibition by staurosporine and calphostin (PKCinhibitors) indicate that the phosphorylated form is the active formof the enzyme. Further, for the first time role of phosphorylation in
Fig. 5. Effect of abiotic stress conditions on BjNOSla. Treatments were given for 0.25e24 h. (stress: seedlings were placed in a vial containing buffer in a water bath maintained at 50 �C.(D) Dehydration: induction of water deficit was achieved by drying the roots of the seedlingwas pricked by a needle at 4e5 places and then placed the seedlings in B.O.D. at 25 � 2 �C inAsterisk showed significant difference in comparison with control (Student’s t-test, p < 0.0
regulation of NOS like activity in plants is reported. Furthermore,the activation of BjNOSla by different abiotic stress indicates thatNOS like activity could be responsible for NO evolution duringstress conditions (Fig. 6).
4. Methods
4.1. Plant material and growth conditions
B. juncea var. varuna seeds were obtained from National SeedCorporation, Indian Agricultural Research Institute, New Delhi,India. Seeds were treatedwith ethanol for 20 s followed bywashingthrice with distilled water and soaked overnight. These were platedin wet germination paper rolls and transferred to B.O.D incubator(25 � 2 �C with 260 mmol m�2 s�1 light for 16 h/8 h light/darkphotoperiod) after keeping for 1 d in dark.
4.2. Protein extraction and estimation
Seven days old B. juncea seedlings were homogenized in Hepesbuffer (65 mM, pH 7.4) containing sucrose (514 mM), EDTA(0.16 mM), DTT (1.6 mM), PMSF (0.5 mM) and protease inhibitorcocktail (Sigma, 5 mL g�1). The extract was centrifuged at12,500 rpm for 25min at 4 �C. Supernatant was passed through twolayeredmuslin cloth andwas incubatedwith AG50-WX8 resin (Naþ
form, BioRad) for 1 h with intermittent shaking. Resin was allowedto settle and the extract was used for activity assay. Proteins werequantified using Bradford’s method [36].
4.3. Nitric oxide synthase activity assay
Both oxyhemoglobin and citrulline assays were employed toanalyze the NOS activity in B. juncea. For oxyhemoglobin assay,fresh oxyhemoglobin was prepared every day [28]. Hemoglobin
A) Low temperature stress: seedlings were incubated in ice cold buffer at 4 �C. (B) Heat(C) Salinity stress was provided by submerging the roots of the seedlings in 0.5 M NaCl.using a tissue paper and incubating them in B.O.D. at 25 � 2 �C. (E) Injury: the seedlingbuffer solution. Error bars represent standard deviation from three biological replicates.5).

Fig. 6. BjNOSla is phosphorylated. Calcium, NADPH, light, PMA, PS, 1,2-DG6 and abioticstress activate BjNOSla while the PKC inhibitors-staurosporine and calphostin inhibit it.
P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164 163
(25 mg) was suspended in 1 mL of TriseHCl buffer (10 mM, pH 7.0)and incubatedwith sodium dithionite (20mgmL�1) for 30min. Thesolution was passed through sephadex G-25 (10 mL) column fordesalting. Aliquots (1 mL) were eluted using 10 mM Tris containing0.1 M NaCl, at a flow rate of 1 mL min�1 and the fraction havingmaximum oxyhemoglobin was used for the assay [21]. For opti-mization, the assay was performed in 65 mM Hepes with varyingconcentrations of the substrate (L-arginine, 1e4 mM) and otherreaction mixture components like oxyhemoglobin (2.5e20 mL),CaCl2 (0.1e0.6 mM) and MgCl2 (5e40 mM). BjNOSla was alsomonitored in presence of NADPH (0.1e1 mM), calmodulin (CaM,0.5e1 mM), FAD (1e40 mM), FMN (1e40 mM) and BH4 (4e12 mM). Allexperiments were performed with ten biological replicates, witheach experiment carried out in duplicates. NOS like activity wasexpressed as pmol min�1 mg�1 protein and the results areexpressed as percent activity. Km, Vmax and Hill coefficient werecalculated by plotting MichaeliseMenten curve (v versus s),LineweavereBurk plot (1/v versus 1/s) and Hill plot [log v/(Vmax � v) versus log s]. Rat neuronal NOS was used as positivecontrol.
Citrulline assay was performed according to Cueto et al. withslight modifications [21]. The assay was performed using 20 mL ofthe crude extract, 0.2 mM L-arginine, 0.3 mM CaCl2 and 0.05e0.45 mCi of L-[U-14C] arginine. The reaction was terminated usingAG50WX-8, suspended in Milli Q water (1:1 ratio). The supernatantwas added to 3 mL scintillation cocktail-W and quantified by liquidscintillation counting (Beckman Coulter, Model LS 6000). Fourbiological replicates were performed with each assay performed induplicates.
4.4. Nitric oxide detection by fluorescent microscopy
In vivo detection of BjNOSla was performed in epidermal peelusing NADPH-diaphorase [21] and DAF-2DA staining [37].Epidermal peels were gently removed from the abaxial surface ofthe 4 weeks old Brassica leaves. For NADPH-diaphorase staining,epidermal peels were fixed with 4% p-formaldehyde and 1%glutaraldehyde in 0.1 M phosphate buffered saline pH 7.4 and thenincubated with 4% p-formaldehyde for 2 h. These peels weremounted in 20% glycerol followed by washed thrice for 10 mineach in 10 mM Tris buffer pH 7.6 and was suspended in 5% TritonX-100 in Tris buffer pH 7.6. The peels were incubated in dark for2 h at 37 �C in a solution containing NBT (0.25 mg mL�1), NADPH(1 mg mL�1) and Triton X-100 (0.5%) in 0.1 M Tris pH 7.6 [21].Peels were mounted on slides using glycerine and were observedunder light microscope (40� magnification). In the control set,NADPH was omitted. Experiments were repeated thrice induplicates.
DAF-2DA staining was performed according to Foissner et al.[37]. In brief, epidermal peels were placed in a petri dish containing
5 mL of 10 mM Tris pH 7.2 and 10 mM DAF-2DA. Epidermal peelswere maintained in dark for 1 h. The peels were removed andwashed with buffer (10 mM Tris, pH 7.2) for 15 min and then with1mM L-arginine for 30min for 25 �C. L-NAME (5mM, 30min, 25 �C)and NBT (300 mM), were added to the buffer after washing theprobe. Fluorescent microscope (Zeiss, Oberkochen, Germany) wasused for analysis and sections were excited with UV light. Dyeemissions were recorded using a 505e530 nm band pass filter.Autofluorescence of chloroplasts was captured using a 585 nm longpass filter. Images were processed and captured by Zeiss Micro-systems camera and analyzed using Zeiss LSM510 software.Experiments were carried out three times in duplicates.
4.5. NOS inhibitor, PKC inhibitors and PKC activators treatments
Effect of mammalian NOS inhibitors, NBT (10e300 mM) and L-NAME (1e5 mM) was observed on BjNOSla. EGTA (1e2 mM), PKCinhibitors, staurosporine (1.5e20 nM), calphostin-C (0.1e1 mM)were incubated with the extract for 5 min before measuring theactivity. Each experiment was carried out in duplicates with fourbiological replicates. NOS like activity was analyzed with PMA(438 nM), 4aPDD (438 nM), 1,2-DG6 (2.5 mM) and PS (1 mM) in vitro.
4.6. Abiotic stress treatments
The seedlings were subjected to different abiotic stress condi-tions for 0.25e24 h. Seedlings were incubated in ice cold Hepesbuffer at 4 �C for low temperature and 50 �C for high temperature.Dehydration was done by keeping seedlings on a tissue paper at25 �C and for salinity in 0.5 M NaCl. Injury stress was given to theseedlings by pricking at 4e5 places by sterilized needle. Experi-ments were carried out in three biological replicates with twotechnical replicates.
Acknowledgments
The researchworkwas partially supported by SAP program fromUniversity Grants Commission and Council of Scientific andIndustrial Research (CSIR), Government of India. PST acknowledgesCSIR for fellowship. We thank Prof. S. K. Bansal (Department ofBiochemistry, V. P. Patel Chest Institute, Delhi) for providing theliquid scintillation counting facility.
References
[1] M.H. Siddiqui, M.H. Al-Whaibi, M.O. Basalah, Role of nitric oxide in toleranceof plants to abiotic stress, Protoplasma 248 (2011) 447e455.
[2] I.D. Wilson, S.J. Neill, J. Hancock, Nitric oxide synthesis and signalling in plants,Plant Cell. Environ. 31 (2008) 622e631.
[3] L. Klepper, Nitric oxide (NO) and nitrogen dioxide (NO2) emissions fromherbicide treated soyabean plants, Atmos. Environ. 13 (1979) 537.
[4] J. Durner, D. Wendehenne, D.E. Klessig, Defence gene induction in tobacco bynitric oxide, cyclic GMP and cyclic ADP ribose, Proc. Natl. Acad. Sci. U.S.A. 95(1998) 10328e10333.
[5] M. Delledonne, Y. Xia, R.A. Dixon, C. Lamb, Nitric oxide functions as a signal inplant disease resistance, Nature 394 (1998) 585e588.
[6] B.R. Crane, J. Sudhamsu, B.A. Patel, Bacterial nitric oxide synthases, Annu. Rev.Biochem. 79 (2010) 445e470.
[7] M.S. Leitner, C. Bommassar, G. Golderer, P. Grobner, E.R. Werner, G. Werner-Felmayer, Physarum nitric oxide synthases: genomic structures and enzy-mology of recombinant proteins, Biochem. J. 418 (2009) 691e700.
[8] N. Foresi, N. Correa-Aragunde, G. Parisi, G. Calo, G. Salerno, L. Lamattina,Characterization of a nitric oxide synthase from the plant kingdom: NOgeneration from the green alga Ostreococcus tauri is light irradiance andgrowth phase dependant, Plant Cell 22 (2010) 3816e3830.
[9] L.A. del Rıoa, F.J. Corpasa, J.B. Barroso, Nitric oxide and nitric oxide synthaseactivity in plants, Phytochemistry 65 (2004) 783e792.
[10] Y. Liu, H. Jiang, Z. Zhao, L. An, Nitric oxide synthase like activity-dependentnitric oxide production protects against chilling-induced oxidative damagein Chorispora bungeana suspension cultured cells, Plant Physiol. Biochem. 48(2010) 936e944.

P.S. Talwar et al. / Plant Physiology and Biochemistry 60 (2012) 157e164164
[11] L.A.J. Mur, J. Mandon, S.M. Cristescu, F.J.M. Harren, E. Prats, Methods of nitricoxide detection in plants: a commentary, Plant Sci. 181 (2011) 509e519.
[12] F.Q. Guo, M. Okamoto, N.M. Crawford, Identification of a plant nitric oxidesynthase gene involved in hormonal signalling, Science 302 (2003) 100e103.
[13] M. Moreau, G.I. Lee, Y. Wang, B.R. Crane, D.F. Klessig, AtNOS/AtNOA1 isa functional Arabidopsis thaliana cGTPase and not a nitric-oxide synthase,J. Biol. Chem. 283 (2008) 32957e32967.
[14] J. Bright, R. Desikan, J.T. Hancock, I.S. Weir, S.J. Neill, ABA-induced NOgeneration and stomatal closure in Arabidopsis are dependent on H2O2synthesis, Plant J. 45 (2006) 113e122.
[15] X. Liu, Y.Q. Wang, S.W. Jia, C.H. Lou, S.Q. Zhang, Localization of NOS-likeprotein in guard cells of Vicia faba L. and its possible function, Chin. Sci.Bull. 52 (2007) 84e90.
[16] T.M. Dawson, D.S. Bredt, M. Fotuhi, P.M. Hwang, S.H. Snyder, Nitric oxidesynthase and neuronal NADPH diaphorase are identical in brain and periph-eral tissues, Proc. Natl. Acad. Sci. U.S.A. 88 (1991) 7797e7801.
[17] S. Neill, R. Barros, J. Bright, R. Desikan, J. Hancock, J. Harrison, P. Morris,D. Ribeiro, I. Wilson, Nitric oxide, stomatal closure, and abiotic stress, J. Exp.Bot. 59 (2008) 165e176.
[18] K.J. Gupta, A.R. Fernie, W.M. Kaiser, J.T. van Dongen, On the origin of nitricoxide, Trends Plant Sci. 16 (2011) 160e168.
[19] Q. Yang, H. He, H. Li, H. Tian, J. Zhang, L. Zhai, J. Chen, H. Wu, G. Yi, Z.H. He,X. Peng, NOA1 functions in a temperature-dependent manner to regulatechlorophyll biosynthesis and Rubisco formation in rice, PLoS ONE 6 (2011).http://dx.doi.org/10.1371/journal.pone.0020015.
[20] J. Lozano-Juste, J. León, Enhanced abscisic acid-mediated responses in nia1-nia2noa1-2 triple mutant Impaired in NIA/NR- and AtNOA1-dependent nitricoxide biosynthesis in Arabidopsis, Plant Physiol. 152 (2010) 891e903.
[21] M. Cueto, H. Perera, R. Martin, M.L. Bentura, J. Rodrigo, S. Lamas, M.P. Golvano,Presence of nitric oxide synthase activity in roots and nodules of Lupinusalbus, FEBS Lett. 398 (1996) 159e164.
[22] J.B. Barrosso, F.J. Corpas, A. Carreras, L.M. Sandalio, R. Valderrama, J.M. Palma,J.A. Lupianez, L.A.D. Rio, Localization of nitric-oxide synthase in plant perox-isomes, J.Biol. Chem. 274 (1999) 36729e36733.
[23] E.A.J. Rebeiro, F.Q. Cunha, W.M.S.C. Tamashiro, I.S. Martins, Growth phase-dependent subcellular localization of nitric oxide synthase in maize cells,FEBS Lett. 445 (1999) 283e286.
[24] L.V. Modolo, F.Q. Cunha, M.R. Braga, I. Salgado, Nitric oxide synthase mediatedphytoalexin accumulation in soybean cotyledons in response to Diaporthephaseolorum sp meridionalis elicitor, Plant Physiol. 130 (2002) 1288e1297.
[25] I. Hong, Y.K. Kim, W.S. Choi, D.W. Seo, J.W. Yoon, J.W. Han, H.Y. Lee, H.W. Lee,Purification and characterization of nitric oxide synthase from Staphylococcusaureus, FEMS Microbiol. Lett. 222 (2003) 177e182.
[26] N. Song, C. Jeong, H. Choi, Identification of nitric oxide synthase in Flammulinavelutipes, Mycologia 92 (2000) 1027e1032.
[27] F.J. Corpas, J.B. Barroso, A. Carreras, Cellular and subcellular localization ofendogenous nitric oxide in young and senescent pea plants, Plant Physiol. 136(2004b) 2722e2733.
[28] X. Hu, S.J. Neill, Z. Tang, W. Cai, Nitric oxide mediates gravitropic bending insoybean roots, Plant Physiol. 137 (2005) 663e670.
[29] N. Correa-Aragunde, M. Graziano, C. Chevalier, L. Lamattina, Nitric oxidemodulates the expression of cell cycle regulatory genes during lateral rootformation in tomato, J. Exp. Bot. 57 (2006) 581e588.
[30] G.C. Pagnussat, M.L. Lanteri, M.C. Lombardo, L. Lamattina, Nitric oxide medi-ates the indole acetic acid induction activation of a mitogen-activated proteinkinase cascade involved in adventitious root development, Plant Physiol. 135(2004) 279e286.
[31] C.M.C.P. Gouvea, J.F. Souza, A.C.N. Magalhães, I.S. Martins, NO-releasingsubstances that induce growth elongation in maize root segments, PlantGrowth Regul. 21 (1997) 183e187.
[32] R.G. Knowles, M. Palacios, R.M.J. Palmer, S. Monacada, Kinetic characteristicsof nitric oxide synthase from rat brain, Biochem. J. 269 (1990) 207e210.
[33] I.O. Han, H.S. Kim, H.C. Kim, E.H. Joe, W.K. Kim, Synergistic expressionof inducible nitric oxide synthase by phorbol ester and interferon g is medi-ated through NF-kB and ERK in microglial cells, J. Neurosci. Res. 73 (2003)659e669.
[34] T. Munnik, C. Testerink, Plant phospholipid signaling: “in a nutshell”, J. LipidRes. (Suppl.) (April 2009) S260eS265.
[35] A. Sato, F. Gambale, I. Dreyer, N. Uozumi, Modulation of the Arabidopsis KAT1channel by an activator of protein kinase C in Xenopus laevis oocytes, FEBS J.277 (2010) 2318e2328.
[36] M.M. Bradford, A rapid and sensitive method for quantification of microgramquantities of protein utilizing the principle of protein-dye binding, Anal.Biochem. 42 (1976) 248e254.
[37] I. Foissner, D. Wendehenne, C. Langebartels, J. Durner, In vivo imagingof an elicitor-induced nitric oxide burst in tobacco, Plant J. 23 (2000)817e824.
[38] D. Zeidler, U. Zahringer, I. Gerber, I. Dubery, T. Hartung, W. Bors, P. Hutzler,J. Durner, Innate immunity in Arabidopsis thaliana: lipopolysaccharides acti-vate nitric oxide synthase (NOS) and induce defense genes, Proc. Natl. Acad.Sci. U.S.A. 101 (2004) 15811e15816.
[39] Y.J. Chen, J.P.N. Rosazza, A bacterial nitric oxide synthase from Nocardiaspecies, Biochem. Biophys. Res. 203 (1994) 1251e1258.
[40] J. Maier, R. Hacker, P. Rockel, H. Ninnemann, Role of nitric oxide synthase inthe light-induced development of sporangiophore in Phycomyces blakesleea-nus, Plant Physiol. 126 (2001) 1323e1330.