Embed Size (px)
Citation preview

Braman et al.: Zoysiagrass Susceptibility to
S. vicinus 301
POTENTIAL RESISTANCE IN ZOYSIAGRASSESTO TAWNY MOLE CRICKETS (ORTHOPTERA:
GRYLLOTALPIDAE)
S.K. B
RAMAN
1
, A.F. P
ENDLEY
1
, R.N. C
ARROW
2
AND
M.C. E
NGELKE
3
1
Department of Entomology and
2
Department of Crop &Soil Science, University of Georgia,
College of Agriculture and Environmental SciencesExperiment Stations, Georgia Station, Griffin, GA 30223
3
Texas A&M University, Research & Extension Ctr.,17360 Coit Road., Dallas, TX 75252
A
BSTRACT
Reduction in growth by tawny mole crickets,
Scapteriscus vicinus
Scudder, at den-sities equivalent to 15 adults per 0.09 m
2
varied among nine experimental and threecommercially available zoysiagrass (
Zoysia
Willd.) cultivars. Reductions in root dryweights after a four week infestation period were similar among all cultivars, and av-eraged 27.1% less than non-infested controls. Shoot dry weight reduction was mostsevere for DALZ 8516, DALZ 9006, and Meyer zoysia. The cultivars that retained thehighest percentage of their normal growth were DALZ 8502, DALZ 8514, DALZ 8701,and Emerald zoysia. Crickets fed on cultivars that were least damaged usually pro-duced the least number of eggs. However, when the most severely injured selection(DALZ 8516) served as the host, a similarly low number of eggs was observed. Crick-ets fed on Meyer zoysia and DALZ 8508 produced the greatest number of eggs.
Key Words: Host plant resistance,
Scapteriscus
spp., turfgrass
R
ESUMEN
La reducción del crecimiento de los grillotopos aleonados, Scapteriscus vicinusScudder, a densidades equivalentes a 15 adultos por 0.09 m2, varió en nueve culti-vares experimentales y tres comerciales de hierba zoysia (Zoysia Willd.). Las reduc-ciones en el peso seco de las raices fueron similares en todos los cultivares, luego deun período de 4 semanas infestación, y promediaron un 27.1% menos que los testigosno infestados. La reducción en el peso seco de los brotes fue mas severa en las zoysiasDALZ 8516, DALZ 9006 Y Meyer. Los cultivares que retuvieron el mas alto porcentajede su crecimiento normal fueron DALZ 8502, DALZ 8516, DALZ 8701 y Emerald. Losgrillos que se alimentaron de los cultivares menos dañados, usualmente produjeronmenor numero de huevos. Sin embargo, cuando el clon mas severamente danado(DALZ 8516) sirvió como hospedante, fue observado un numero de huevos similarm-ente bajo. Los grillos alimentados de las zoysias Meyer y DALZ 8508 prdujeron el
mayor número de huevos.
Mole crickets in the genus
Scapteriscus
have become the most serious pests of turf-grasses in the southeastern United States since their entry into this country, probablyvia the ballast of ships at approximately 1900 (Walker & Nickle 1981, Nickle & Cast-ner 1984, Walker 1984). The tawny mole cricket,
Scapteriscus vicinus
Scudder, is the
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

302
Florida Entomologist
77(3) September, 1994
more damaging of the two species common in Georgia. Management of these pests hasinvolved chemical, cultural, and classical biological control efforts (Walker 1984, Hud-son et al. 1988).
Laboratory and field screening has identified turfgrass genotypes that are rela-tively resistant, tolerant, or less preferred by various insects and mites (see reviewsby Reinert 1982, Quisenberry 1990). Limited research has focused on resistance tomole crickets and white grubs (Potter & Braman 1991). Resistant cultivars areneeded to provide a safe, economical control strategy for these serious turf pests.Herein, we report the results of a greenhouse study that evaluated nine experimentaland three commercially available zoysiagrass (
Zoysia
Willd.) cultivars for their sus-ceptibility to mole cricket injury and suitability as oviposition substrates.
M
ATERIALS
AND
M
ETHODS
Zoysiagrass plugs (4.6 x 4.6 x 6.3 cm) of 12 cultivars were transplanted into gran-ular calcinated clay (Turface, Applied Industrial Materials, Corp., Deerfield, IL) inplastic pots (15 cm diam) in a greenhouse. Pots were watered daily and fertilized onceper week with a solution containing 250 ppm NPK (Peters
20-20-20). Milorganitewas applied (2.25 gm per pot) once per month. Turf was cut weekly to a height of 5 cmExperimental cultivars included a range of leaf textures, colors, and growth rates(Carrow 1992). Six months after transplanting, plugs were transferred to PVC tubes(38 cm tall; 15 cm diam) containing fine sand (children’s play sand). These tubes werecovered at the bottom with plastic petri dish lids and were placed into wooden boxframes and equipped with drip irrigation. Watering and fertilization regimes weremaintained as before for one month before infestation with adult crickets.
Adult
S. vicinus
were collected in Tifton, GA during April, 1992 using a standardacoustic trap similar to that described by Walker (1982). Twenty-four treatments (12cultivars each infested with mole crickets and the same 12 cultivars non-infested)were arranged in a randomized complete block design with 7 replications (168 totaltubes). Two female and one male mole cricket were introduced into each of the 84tubes designated as infested treatments (252 total crickets were used). Each con-tainer was covered with 32-mesh saran screen (Chicopee Manufacturing Co., Gaines-ville, GA) to prevent escape of crickets. Non-infested cages were also covered withscreens to ensure equivalent light, temperature and humidity conditions. Water andfertilizer regimes were maintained as described previously. Greenhouse microplotswere destructively sampled after four weeks of exposure to crickets.
Top growth was clipped to a height of 5 cm two weeks after crickets were intro-duced into the PVC containers. Clippings were placed in paper bags, oven dried for 7days, and then weighed. Top growth dry weight was also recorded four weeks aftercricket infestation. Numbers of green shoots per 18 cm
2
, selected at random from the182 cm
2
surface of each tube, and root dry weights were determined at the termina-tion of the experiment.
Sand from each experimental PVC microplot (1343 kg total for all plots) was siftedto recover mole cricket adults and eggs. Adult survival and numbers of eggs were re-corded and compared among cultivars using the GLM procedure (SAS 1985). Num-bers of eggs per cultivar were also regressed against injury. Means were separatedfollowing a significant analysis of variance by a least significant difference test (Sokal& Rohlf 1981). Growth reductions of roots, shoots, and shoot density (expressed as apercentage of non-infested plants for each cultivar) were transformed using an arc-sine square root of the proportion before being subjected to analysis of variance andmean separation using a least significant difference test.
Braman et al.: Zoysiagrass Susceptibility to
S. vicinus 303
R
ESULTS
AND
D
ISCUSSION
Reduction in top growth two weeks after infestation was statistically similaramong all cultivars (F=1.50; df=11,66; P>0.05; Table 1). Growth of cricket-infestedplants averaged 67.9% of non-infested plants at that time. Reduction in shoot dryweights differed (F=4.17; df=11,66; P=0.0001) among cultivars four weeks after infes-tation. DALZ 8516 achieved only 10.1% of its normal growth when infested with molecrickets. DALZ 8502, however, maintained 55% of its normal growth even under thishigh infestation level. The pest density used in this study was equivalent to 15 adultcrickets per 0.09 m
2
(= 1 ft
2
). This represents more than 15 times the pest density re-quiring chemical intervention to protect turf. Total reduction in top growth for the en-tire four week period ranged from 35.7 to 74.8% (inverse of the extremes presented inTable 1).
Shoot density followed a pattern similar to that of clipped dry weights. Root weightreduction, however, was similar (F=1.45; df=11,66; P>0.05) among all cultivars eval-uated and averaged 72.9% of non-infested controls (Table 1). The majority of the dam-age observed was confined to the crown of each infested zoysiagrass plug. Dead turfwas first visible at the center of each plug and expanded outward with increasing timeof exposure.
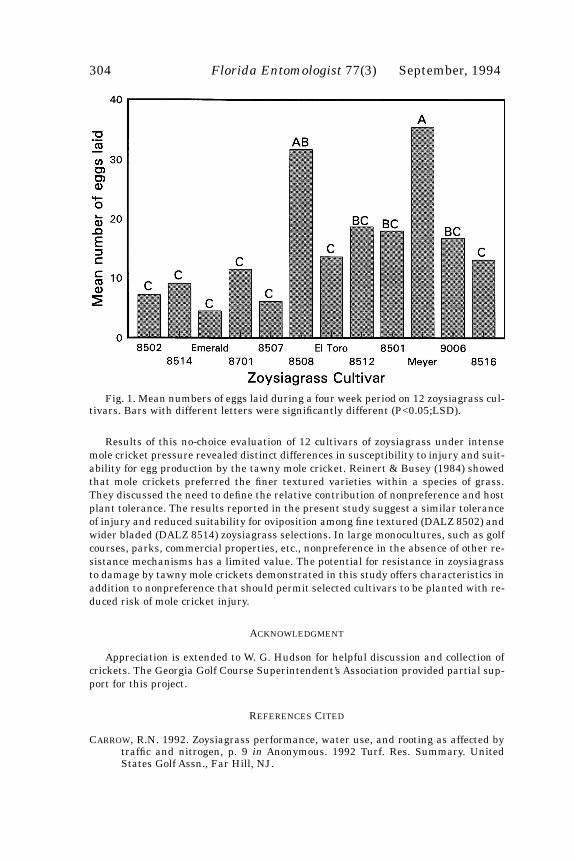
Adult survival at the termination of the four week exposure period was not signif-icantly affected by cultivar (F=1.58; df=11,66; P>0.05) and averaged 70.0% on all cul-tivars (data not given). Egg production during this time period, however, differed(F=2.63; df=11,66; P=0.003) among cultivars (Fig.1). Mean numbers of eggs per culti-var ranged from 4.5 to 35.4. In general, tubes containing cultivars that were leastdamaged (DALZ 8502, DALZ 8514, Emerald, DALZ 8701, and DALZ 8507) also con-tained lower numbers of eggs, except for DALZ 8516, which was the most severelydamaged, but contained relatively few eggs. Meyer zoysia and DALZ 8508 supportedthe greatest egg production. Regression of number of eggs per cultivar against injurywas significant (F=16.9; df=11,82; P=0.0001; r
2
=0.2), however, the low r
2
value indi-cated that plant injury explained little of the variation for numbers of eggs laid.
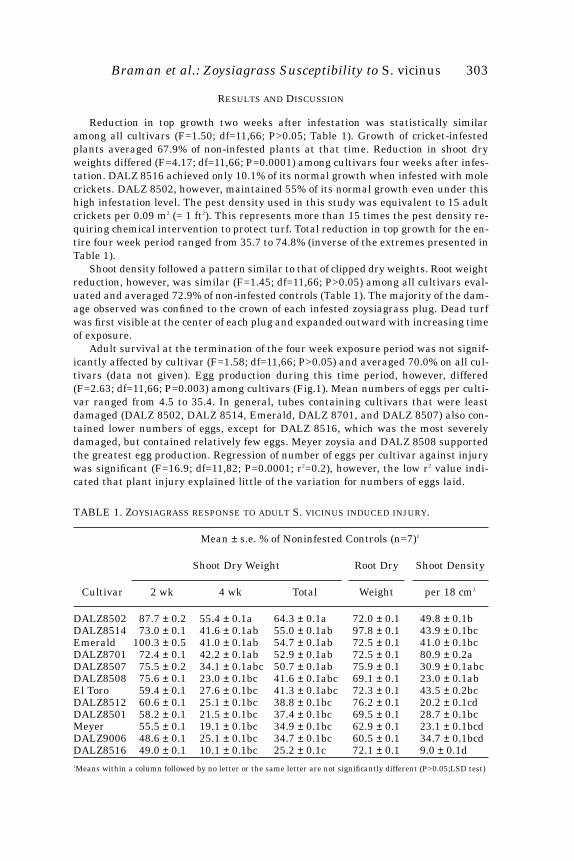
TABLE 1. Z
OYSIAGRASS
RESPONSE
TO
ADULT
S.
VICINUS
INDUCED
INJURY
.
Mean
±
s.e. % of Noninfested Controls (n=7)
1
Shoot Dry Weight Root Dry Shoot Density
Cultivar 2 wk 4 wk Total Weight per 18 cm
2
DALZ8502 87.7
±
0.2 55.4
±
0.1a 64.3
±
0.1a 72.0
±
0.1 49.8
±
0.1bDALZ8514 73.0
±
0.1 41.6
±
0.1ab 55.0
±
0.1ab 97.8
±
0.1 43.9
±
0.1bcEmerald 100.3
±
0.5 41.0
±
0.1ab 54.7
±
0.1ab 72.5
±
0.1 41.0
±
0.1bcDALZ8701 72.4
±
0.1 42.2
±
0.1ab 52.9
±
0.1ab 72.5
±
0.1 80.9
±
0.2aDALZ8507 75.5
±
0.2 34.1
±
0.1abc 50.7
±
0.1ab 75.9
±
0.1 30.9
±
0.1abcDALZ8508 75.6
±
0.1 23.0
±
0.1bc 41.6
±
0.1abc 69.1
±
0.1 23.0
±
0.1abEl Toro 59.4
±
0.1 27.6
±
0.1bc 41.3
±
0.1abc 72.3
±
0.1 43.5
±
0.2bcDALZ8512 60.6
±
0.1 25.1
±
0.1bc 38.8
±
0.1bc 76.2
±
0.1 20.2
±
0.1cdDALZ8501 58.2
±
0.1 21.5
±
0.1bc 37.4
±
0.1bc 69.5
±
0.1 28.7
±
0.1bcMeyer 55.5
±
0.1 19.1
±
0.1bc 34.9
±
0.1bc 62.9
±
0.1 23.1
±
0.1bcdDALZ9006 48.6
±
0.1 25.1
±
0.1bc 34.7
±
0.1bc 60.5
±
0.1 34.7
±
0.1bcdDALZ8516 49.0 ± 0.1 10.1 ± 0.1bc 25.2 ± 0.1c 72.1 ± 0.1 9.0 ± 0.1d1Means within a column followed by no letter or the same letter are not significantly different (P>0.05;LSD test)
304 Florida Entomologist 77(3) September, 1994
Results of this no-choice evaluation of 12 cultivars of zoysiagrass under intensemole cricket pressure revealed distinct differences in susceptibility to injury and suit-ability for egg production by the tawny mole cricket. Reinert & Busey (1984) showedthat mole crickets preferred the finer textured varieties within a species of grass.They discussed the need to define the relative contribution of nonpreference and hostplant tolerance. The results reported in the present study suggest a similar toleranceof injury and reduced suitability for oviposition among fine textured (DALZ 8502) andwider bladed (DALZ 8514) zoysiagrass selections. In large monocultures, such as golfcourses, parks, commercial properties, etc., nonpreference in the absence of other re-sistance mechanisms has a limited value. The potential for resistance in zoysiagrassto damage by tawny mole crickets demonstrated in this study offers characteristics inaddition to nonpreference that should permit selected cultivars to be planted with re-duced risk of mole cricket injury.
ACKNOWLEDGMENT
Appreciation is extended to W. G. Hudson for helpful discussion and collection ofcrickets. The Georgia Golf Course Superintendent’s Association provided partial sup-port for this project.
REFERENCES CITED
CARROW, R.N. 1992. Zoysiagrass performance, water use, and rooting as affected bytraffic and nitrogen, p. 9 in Anonymous. 1992 Turf. Res. Summary. UnitedStates Golf Assn., Far Hill, NJ.
Fig. 1. Mean numbers of eggs laid during a four week period on 12 zoysiagrass cul-tivars. Bars with different letters were significantly different (P<0.05;LSD).

Braman et al.: Zoysiagrass Susceptibility to S. vicinus 305
HUDSON, W. G., J. H. FRANK, AND J. L. CASTNER. 1988. Biological control of Scap-teriscus spp. mole crickets (Orthoptera: Gryllotalpidae) in Florida. Bull. Ento-mol. Soc. America 34: 192-198.
NICKLE, D.A., AND J. L. CASTNER. 1984. Introduced species of mole crickets in theUnited States, Puerto Rico, and the Virgin Islands (Orthoptera: Gryllotalpi-dae). Ann. Entomol. Soc. America 77: 450-465.
POTTER, D. A., AND S. K. BRAMAN. 1991. Ecology and management of turfgrass in-sects. Annu. Rev. Entomol. 36: 383-406.
QUISENBERRY, S. S. 1990. Plant resistance to insects and mites in forage and turfgrasses. Florida Entomol. 73: 411-421.
REINERT, J. A. 1982. A review of host resistance in turfgrasses to insects and acarineswith emphasis on the southern chinch bug, p. 3-12 in H.D. Niemczyk and B. G.Joyner [eds.]. Advances in turfgrass entomology. Hammer Graphics, Piqua,OH, 150 pp.
REINERT, J. A., AND P. BUSEY. 1984. Resistant varieties, p. 35-40 in T. J. Walker [ed.].Mole crickets in Florida. Florida Agric. Exp. Stn. Bull. 846.
SAS INSTITUTE INC. 1985. SAS Users Guide: Statistics, version 5 edition, Cary, NC,956 pp.
SOKAL, R. R., AND F. J. ROHLF. 1981. Biometry. Second edition. W. H. Freeman & Co.,San Francisco.
WALKER, T. J. 1982. Sound traps for sampling mole cricket flights (Orthoptera: Gryl-lotalpidae: Scapteriscus). Florida Entomol. 65:105-109.
WALKER, T. J. 1984. Mole crickets in Florida. Florida Agric. Exp. Stn. Bull. 846. 54 pp.WALKER, T.J., AND D.A. NICKLE. 1981. Introduction and spread of pest mole crickets:
Scapteriscus vicinus and Scapteriscus acletus reexamined. Ann. Entomol. Soc.
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
America 76:507-517.

305
MATING FREQUENCY OF THE PAPAYA FRUIT FLY(DIPTERA: TEPHRITIDAE) WITH AND WITHOUT HOST FRUIT
P
ETER
J. L
ANDOLT
Insect Attractants, Behavior, and Basic BiologyResearch Laboratory, Agricultural Research Service
U.S. Department of Agriculture, Gainesville, FL 32604
A
BSTRACT
Frequency of mating of female papaya fruit flies,
Toxotrypana curvicauda
Gers-taecker, was affected by the availability of host fruit for oviposition. Mature femalesheld for 5 days with males in cages in the laboratory without host fruit nearly alwaysmated once. Most mature females held in cages for 5 days with both males and imma-ture papaya fruit mated more than once. Remating by females held with fruit oc-curred occasionally before oviposition, indicating that an increased propensity toremate may not be due to sperm depletion. Mature males kept for 5 days with femalesmated up to 10 times (x=5.6+0.6).
Key Words: Insecta,
Toxotrypana curvicauda,
oviposition, pheromone, kairomone, re-mating
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

306
Florida Entomologist
77(3) September, 1994
R
ESUMEN
La frecuencia de apareamiento de hembras de la mosca frutera de la papaya, Tox-otrypana curvicauda Gerstaecker, fué afectada por la disponibilidad del fruto hos-pedante para la ovoposición. En el laboratorio las hembras maduras mantenidas enjaulas durante 5 días, con machos y sin el fruto hospedante, casi siempre se aparearonuna sola vez. La mayoría de las hembras maduras mantenidas en jaulas durante 5días con machos y frutos no maduros de papaya se apareó mas de una vez. Nuevosapareamientos de hembras mantenidas con frutos tuvieron lugar ocasionalmente an-tes de la ovoposición, indicando que el aumento en la propensión al reapareamiento nodebe ser el resultado del agotamiento del esperma. Los machos maduros mantenidosdurante 5 días con hembras se apareron hasta 10 veces (x=5.6
±
0.6)
Key Words: Insecta, oviposition, pheromone, kairomone, remating
The frequency of mating in tephritid fruit flies is an important aspect of their sex-ual behavior. It is relevant to the development of those pest control programs basedin part on sexual interactions. For example, sex attractants developed for femalesmay be more effective for species that remate frequently and may then repeatedly re-spond to male sex pheromone. Also, female mating frequency directly affects the effi-cacy of the sterile male insect technique for control and eradication of tephritid fruitflies.
Frequency of mating varies among females of species of fruit-infesting tephritids.Females of the apple maggot fly,
Rhagoletis pomonella
(Walsh), mate frequently, as of-ten as weekly, possibly to maintain fertility levels (Prokopy & Roitberg 1984). Tropicalspecies of fruit flies that lek, such as
Ceratitis capitata
Wiedemann (Nakagawa et al1971), and
Anastrepha suspensa
(Loew) (Sivinski & Heath 1988), are thought to mateusually once, with rematings due either to insemination failures at first matings or tosperm depletion following extensive oviposition. Landolt & Hendrichs (1983) reportedthat female papaya fruit flies,
Toxotrypana curvicauda
Gerstaecker, usually mateonce, based on observations of flies in a field cage. This might be expected because oftheir relatively short reproductive lifespan and limited egg production.
Studies of female papaya fruit fly responses to male sex pheromone, however, dem-onstrated that females are attracted to males even after mating (Landolt & Heath1988, 1990). Such sex attraction by mated females may be to locate a potential secondmate or may be a strategy to locate host fruit, since males in papaya fields normallycall from immature fruit (Landolt et al. 1992). Because of this apparent discrepancyand the limited current data on
T. curvicauda
mating frequency, the subject was re-addressed. We report here the frequency of mating of both sexes of the papaya fruitfly in the laboratory, and effects of the presence of papaya fruit for oviposition on fe-male mating frequency.
M
ATERIALS
AND
M
ETHODS
Papaya fruit flies were obtained as mature larvae in infested fruit collected in com-mercial papaya plantings in the Redlands area of Dade County and in SarasotaCounty, Florida. Larvae emerging from fruit pupated in sterilized potting soil. Papercannisters (250 ml) of pupae in potting soil were kept in the laboratory in screenedcages for adult emergence. Newly emerged flies were sorted by sex daily and were

Landolt: Papaya Fruit Fly Mating Frequency
307
kept in screened cages with cups of sugar water and inverted water bottles on the cagetops. The laboratory was at 22
°
C and 50
±
10% RH, with overhead fluorescent light-ing. Lights were on a 14:10 (L:D) cycle, with lights on at 0600 hours (E.S.T.) and offat 2000 hours (E.S.T.).
Mating frequency was determined for mature female papaya flies held with imma-ture papaya fruit (preferred oviposition sites), for mature female flies without fruit,and for mature male flies without fruit. For the first experiment, reproductively ma-ture females (> 6 days old, Landolt 1984) were held one per cage from 0730 hours ona Monday to 1700 hours on the following Friday. Each morning at 0800 hours, a malewas placed in each cage and observations were made at 30 min intervals until 1700hours when the male was removed. Because matings on average take 90 min (Landolt& Hendrichs 1983), it was assumed that none would be missed if observations weremade every 30 min. At each 30 min observation time, records were made of matingand oviposition (for females held with fruit). Females without fruit generally do notattempt to oviposit (Landolt & Reed 1990) and no oviposition attempts were seen inthis study by females held without fruit (egg deposition or probing with the ovipositoron the cage). After flies mated, the male was replaced. Each cage included a small cupof sugar water on cotton within the cage and an inverted water bottle on the cage top.Twenty-six females were tested in this way without fruit; 10 during one week, 10 dur-ing a second week and 6 during a third week. This experiment, using the same proto-col, was also conducted for 34 mature females held with males and papaya fruit (as 2sets of 6, 1 set of 12, and 1 set of 10 during 4 different weeks). Papaya fruit used weregreen immature fruit 5 - 7 cm in diam and 7 - 11 cm in length. During the replicatesconducted to determine mating frequency of females without fruit, papaya was alsoexcluded from the laboratory to avoid host odor effects on mating behavior.
To assess mating frequency of males, a similar protocol was followed. For five con-secutive days, at 0800 hours, a mature unmated female was placed in each cage con-taining one male. Observations were made each 30 minutes, until 1700 hours, whenthe female was removed. The female was replaced after each copulation, with anothermature unmated female. This was done for 21 mature (>4 days old) males, all heldwithout papaya fruit (as 2 sets of 6 and 1 set of 5 during 3 different weeks).
Mean mating frequencies for females held with and without papaya fruit werecompared using Student’s t-test.
R
ESULTS
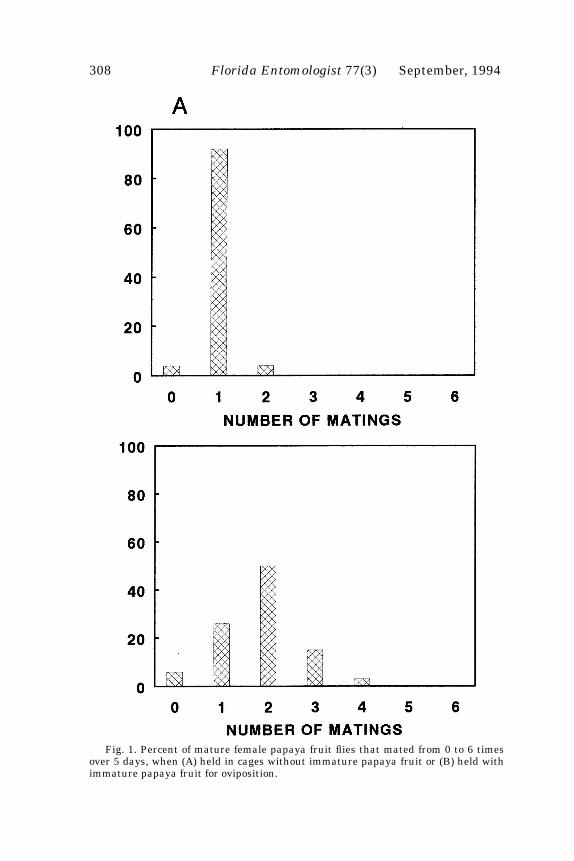
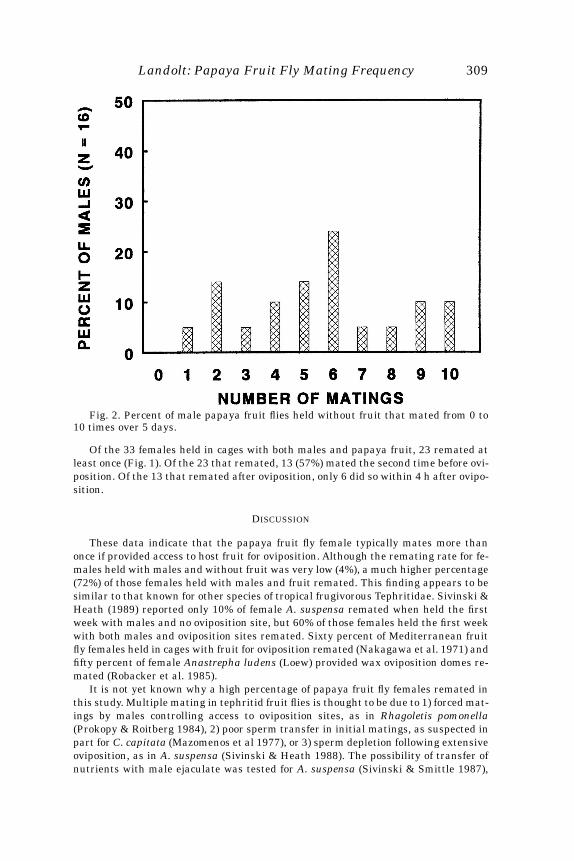
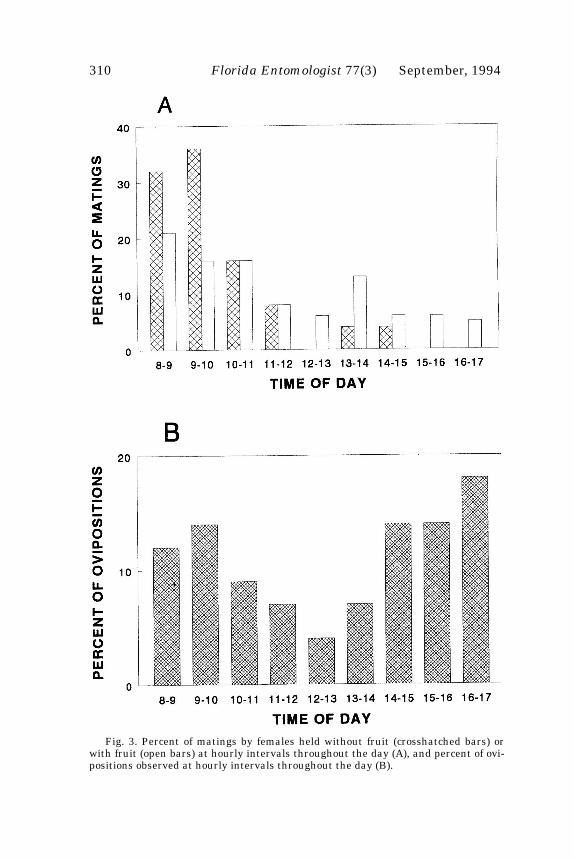
Nearly all female papaya fruit flies held in cages without fruit mated only onceduring the 5-day test period (Fig. 1). Mean number of matings per female (+ SE) was1.00 + 0.06. Females held in cages with papaya fruit mated up to 4 times, with mostfemales mating more than once (Fig. 1). Mean number of matings per female was 1.82+ 0.15. Females without fruit mated significantly less than did females with fruit(t=4.63, df=58, p<0.0001). Males mated up to 10 times over the course of the 5-day testperiod, with nearly all mating multiple times (Fig. 2). Mean number of matings permale was 5.60 + 0.6.
Matings by females held without fruit were predominantly during the morninghours, while matings of females held with fruit were more dispersed throughout theday (Fig. 3). Also, 24 of the 26 females held with males and without fruit mated on thefirst day of the experiment, and 31 of the 33 females held with males and with fruitmated on the first day of the experiment. The pattern of ovipositions observed duringthese tests appeared to be bimodal, with peaks of activity in early morning and againin late afternoon (Fig. 3).
308
Florida Entomologist
77(3) September, 1994
Fig. 1. Percent of mature female papaya fruit flies that mated from 0 to 6 timesover 5 days, when (A) held in cages without immature papaya fruit or (B) held withimmature papaya fruit for oviposition.
Landolt: Papaya Fruit Fly Mating Frequency
309
Of the 33 females held in cages with both males and papaya fruit, 23 remated atleast once (Fig. 1). Of the 23 that remated, 13 (57%) mated the second time before ovi-position. Of the 13 that remated after oviposition, only 6 did so within 4 h after ovipo-sition.
D
ISCUSSION
These data indicate that the papaya fruit fly female typically mates more thanonce if provided access to host fruit for oviposition. Although the remating rate for fe-males held with males and without fruit was very low (4%), a much higher percentage(72%) of those females held with males and fruit remated. This finding appears to besimilar to that known for other species of tropical frugivorous Tephritidae. Sivinski &Heath (1989) reported only 10% of female
A. suspensa
remated when held the firstweek with males and no oviposition site, but 60% of those females held the first weekwith both males and oviposition sites remated. Sixty percent of Mediterranean fruitfly females held in cages with fruit for oviposition remated (Nakagawa et al. 1971) andfifty percent of female
Anastrepha ludens
(Loew) provided wax oviposition domes re-mated (Robacker et al. 1985).
It is not yet known why a high percentage of papaya fruit fly females remated inthis study. Multiple mating in tephritid fruit flies is thought to be due to 1) forced mat-ings by males controlling access to oviposition sites, as in
Rhagoletis pomonella
(Prokopy & Roitberg 1984), 2) poor sperm transfer in initial matings, as suspected inpart for
C. capitata
(Mazomenos et al 1977), or 3) sperm depletion following extensiveoviposition, as in
A. suspensa
(Sivinski & Heath 1988). The possibility of transfer ofnutrients with male ejaculate was tested for
A. suspensa
(Sivinski & Smittle 1987),
Fig. 2. Percent of male papaya fruit flies held without fruit that mated from 0 to10 times over 5 days.
310
Florida Entomologist
77(3) September, 1994
Fig. 3. Percent of matings by females held without fruit (crosshatched bars) orwith fruit (open bars) at hourly intervals throughout the day (A), and percent of ovi-positions observed at hourly intervals throughout the day (B).

Landolt: Papaya Fruit Fly Mating Frequency
311
but with no evidence of a significant male contribution. In the case of the papaya fruitfly, there is no clear indication which of these explanations may account for the ob-served multiple matings.
The male papaya fruit fly in the field defends fruit from other males and courts ar-riving females, conceivably controlling female access to oviposition sites. However, fe-males appear capable of resisting courting males, making it unlikely that multiplematings by females are the result of forced copulations by males. The observed in-creased remating rates of females held with fruit, compared to those held withoutfruit, indicate that poor sperm transfer during first matings was not a problem con-tributing to multiple mating. Insemination failures at first matings would be expectedto result in rematings with or without available oviposition sites. In other studies oftephritid mating frequency, remating appeared to clearly relate to extended periods ofoviposition and was attributed to sperm depletion (Sivinski & Heath 1988; Cunning-ham et al 1971; Robacker et al 1985, Tzanakakis et al 1968). Remating by female pa-paya fruit flies clearly was associated with access to oviposition sites (immaturepapaya fruit) and oviposition activity, suggesting a similar need to replace depletedsperm stocks used during oviposition. However, papaya fruit fly females are relativelyshort lived (Landolt & Hendrichs 1983) and deposit limited numbers of eggs (Knab &Yothers 1914) compared to other frugivorous fruit flies. It is not known if males aresimilarly limited in numbers of sperm they can deliver during insemination, thus re-quiring female remating following oviposition. An additional possibility is that femalepapaya fruit flies arriving at good oviposition sites occupied by males may mate or re-mate as an expression of mate choice, since a male at the site has demonstrated suc-cess both in host location and territorial defense. Such success may involve heritabletraits in the male that are beneficial to the female’s offspring.
Mated female papaya fruit flies may respond to male pheromone (Landolt & Heath1988, 1990) both to locate potential oviposition sites and prospective mates. Previousstudies indicated that enhanced attraction by mated females occurred to a combina-tion of host odor and male pheromone (Landolt & Reed 1990, Landolt et al 1992). Itwas suggested that such behavior may aid females attempting to locate host fruit, byorienting to males calling from such fruit. If ovipositing female
T. curvicauda
need toremate following oviposition, such a strategy ensures the availability of both fruit anda mate simultaneously for an arriving female.
Male papaya fruit flies in this study mated three times more often than femalesduring the same 5-day time period. This is not a measure of male lifetime fertility,since males may live for considerably longer periods of time, but is an indication oftheir potential relative to females for polygamy. These results are similar to those re-ported for male
C. capitata
by Nakagawa et al (1971), with males mating every sev-eral days at every opportunity provided.
A
CKNOWLEDGMENTS
Technical assistance was provided by K. M. Davis-Hernandez. J. Burns and S.Sapp kindly provided access to their papaya groves for collection of fruit and papayafruit flies.
R
EFERENCES
C
ITED
C
UNNINGHAM
, R. T., G. F. F
ARIAS
, S. N
AKAGAWA
,
AND
D. L. C
HAMBERS
. 1971. Repro-duction in the Mediterranean fruit fly: Depletion of stored sperm in females.Ann. Entomol. Soc. America 64:312-313.

312
Florida Entomologist
77(3) September, 1994
K
NAB
, F.,
AND
W. W. Y
OTHERS
. 1914. The papaya fruit fly. J. Agric. Res. 2: 447-453.L
ANDOLT
, P. J. 1984. Reproductive maturation and premating period of the papayafruit fly,
Toxotrypana curvicauda
(Diptera: Tephritidae). Florida Entomol. 67:240-244.
L
ANDOLT
, P. J.,
AND
R. R. H
EATH
. 1988. Effects of age, mating and time of day on be-havioral responses of female papaya fruit fly,
Toxotrypana curvicauda
Gers-taecker (Diptera: Tephritidae) to synthetic pheromone. Environ. Entomol. 17:47-51.
L
ANDOLT
, P. J.,
AND
R. R. H
EATH
. 1990. Effects of pheromone release rate and time ofday on catches of male and female papaya fruit flies on pheromone-baited fruitmodel traps. J. Econ. Entomol. 83: 2040-2043.
L
ANDOLT
, P. J.,
AND
J. H
ENDRICHS
. 1983. Reproductive behavior of the papaya fruitfly,
Toxotrypana curvicauda
Gerstaecker (Diptera: Tephritidae). Ann. Entomol.Soc. America 76: 413-417.
L
ANDOLT
, P. J.,
AND
H. C. R
EED
. 1990. Behavior of the papaya fruit fly (Diptera: Te-phritidae): host finding and oviposition. Environ. Entomol. 19: 1305-1310.
L
ANDOLT
, P. J., H. C. R
EED
,
AND
R. R. H
EATH
. 1992. Attraction of female papaya fruitfly (Diptera: Tephritidae) to male pheromone and host fruit. Environ. Entomol.21: 1154-1159.
M
AZOMENOS
, B., J. L. N
ATION
, W. J. C
OLEMAN
, K. C. D
ENNIS
,
AND
R. E
SPONDA
. 1977.Reproduction in Caribbean fruit flies: Comparisons between a laboratorystrain and a wild strain. Florida Entomol. 60: 139-144.
N
AKAGAWA
, S., G. J. F
ARIAS
, D. S
UDA
, R. T. C
UNNINGHAM
,
AND
D. L. C
HAMBERS
. 1971.Reproduction in the Mediterranean fruit fly: frequency of mating in the labora-tory. Ann. Entomol. Soc. America 64: 949-950.
P
ROKOPY
, R. J.,
AND
B. D. R
OITBERG
. 1984. Foraging behavior of true fruit flies. Amer-ican Scientist. 72: 41-49.
R
OBACKER
, D. C., S. J. I
NGLE
,
AND
W. G. H
ART
. 1985. Mating frequency and responseto male-produced pheromone by virgin and mated females of the Mexican fruitfly. Southwest. Entomol. 10: 215-221.
S
IVINSKI
, J.,
AND
R. R. H
EATH
. 1988. Effects of oviposition on remating, response topheromones and longevity in the female Caribbean fruit fly,
Anastrepha sus-pensa
(Diptera: Tephritidae). Ann. Entomol. Soc. America 81: 1021-1024.S
IVINSKI
, J.,
AND
B. S
MITTLE
. 1987. Male transfer of materials to mates in the Carib-bean fruit fly,
Anastrepha suspensa
(Diptera: Tephritidae). Florida Entomol.70:233-238.
T
ZANAKAKIS
, M. E., J. A. TSITSIPIS, AND A. P. ECONOMOPOULOS. 1968. Frequency ofmating in females of the olive fruit fly under laboratory conditions. J. Econ. En-tomol. 61:1309-1312.

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida
313
ANNOTATED CHECKLISTOF THE BARK AND AMBROSIA BEETLES
(COLEOPTERA: PLATYPODIDAE AND SCOLYTIDAE)OF TROPICAL SOUTHERN FLORIDA
T
HOMAS
H. A
TKINSON
1,3
AND
S
TEWART
B. P
ECK
2
1
Entomology and Nematology Department, University of FloridaGainesville, FL USA 32611
2
Department of Biology, Carleton UniversityOttawa, Ontario, Canada K1S 5B6
A
BSTRACT
The fauna of Scolytidae and Platypodidae is reviewed for tropical southern Florida(Collier, Broward, Dade, and Monroe Counties). The family Platypodidae is repre-sented by 3 species, all in the genus
Platypus
. The family Scolytidae includes 83 spe-cies in 37 genera in the region. This total includes 20 species considered immigrantsto the area. Three species previously reported from the region,
Cryptocarenus spatu-latus
Wood,
Xyleborus xylographus
(Say), and
Araptus politus
(Blandford), probablydo not occur there. Feeding habits, mating systems, hosts, and distributions are sum-marized for all species included.
Key Words: Taxonomy, distributions, ecology, hosts, introduced species
R
ESUMEN
Se revisa la fauna de las familias Scolytidae y Platypodidae de la region tropicaldel sur de Florida (condados de Collier, Broward, Dade y Monroe). La familia Platypo-didae está representada por 3 especies, todas en el genero
Platypus
. La familia Scoly-tidae esta representada por 83 especies en 37 generos. Este total incluye 20 especieslas cuales se consideran inmigrantes al area. Tres especies previamente reportadas dela region,
Cryptopcarenus spatulatus
Wood,
Xyleborus xylographus
(Say) y
Araptuspolitus
(Blandford), probablemente no existen allí. Habitos alimenticios, sistemas de
apareamiento y distribuciones se resumen para todas las especies incluidas.
Bark and ambrosia beetles (Coleoptera: Scolytidae and Platypodidae) constitute adiverse group of beetles that bore in a variety of woody tissues and are well repre-sented in most temperate and tropical forests. Best known are species of
Dendrocto-nus, Ips
, and
Scolytus
that are primary pests of conifers in high-latitude forests,although most of these are atypical of the group as a whole. All species breed in live,stressed, or newly-killed host material. Adults excavate a system of galleries withinwhich eggs are laid and immature development occurs. The greater part of the life cy-cle takes place within host tissues, except for dispersal between hosts. A fascinatingrange of patterns of host plant utilization (tissues consumed, relationships with fun-gal symbionts, and host specificity) and mating behaviors exists within this basic life
3
Current address: Dow Elanco, 13355 Noel Rd., Suite 1045 Dallas, TX 75240-6604
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

314
Florida Entomologist
77(3) September, 1994
history paradigm. Most species leave a persistent physical record of their life historyand activities in the woody tissues of their hosts that can be interpreted by a studentof the group. As a consequence, one can easily tabulate several important biologicalparameters for a large proportion of the species in an area, including those that havenot been studied in detail.
The taxonomy of the group is well-known (Wood 1982, Wood & Bright 1993) for theUnited States as a whole, but there is little information available from southern Flor-ida, largely due to its remoteness. Some collecting was done near the turn of the cen-tury by E. A. Schwarz and A.D. Hopkins. S. L. Wood collected there in the 1950’s.Besides ourselves, more recent collectors of note include D.E. Bright, M. A. Deyrup, R.H. Turnbow, E. G. Riley and R. A. Anderson. Bright’s (1985) checklist of the Caribbeanhighlighted the paucity of available information from that region. No student of thegroup has ever collected extensively on any of the Greater Antilles.
The physical, historical, and floristic geography of southern Florida has been re-viewed by Peck (1989). The area is geologically recent and has a subtropical climate.It has had direct and continuous land contact with temperate areas, but has neverhad any land connections with any tropical areas that would serve as source areas forits current biota. Most of the Neotropical flora of southern Florida has apparently dis-persed there by wind, water, or birds from the Bahamas and Greater Antilles (Tom-linson 1980).
This checklist represents a convergence of 2 separate projects, a faunal study ofthe entire group for the southeastern United States by the first author and a biogeo-graphic study of the insect fauna of tropical southern Florida by the second author(Peck 1989). The purpose of this paper is to document the scolytid and platypodidfauna of tropical southern Florida for a subsequent ecological and biogeographicalanalysis.
M
ETHODS
The second author has collected extensively in the region and accumulated largenumbers of specimens of bark and ambrosia beetles as part of a large scale project onthe biogeography of the insect fauna of tropical southern Florida (Peck 1989). Sam-pling was done with flight intercept traps that were left in place over extended periodsfor 5 years in areas of natural vegetation on the Keys and mainland (sites describedin Peck 1989). Over 2,500 specimens of Scolytidae and Platypodidae were collected.Most specimens and representatives of all species collected by the second author weredeposited in the Canadian National Collection, the Canadian Museum of Nature, andthe S.B. Peck collection (all in Ottawa, Ontario). Some representative specimens weredeposited in the Florida State Collection of Arthropods, Gainesville, Florida.
The first author has been involved in a faunal study of the Scolytidae and Platy-podidae of the southeastern United States for several years. A total of 1,250 speci-mens (other than those collected by the second author) were examined from southernFlorida, as well as an additional 3,000 specimens of the same species from other partsof the United States, Canada, the Caribbean, and Latin America. Scolytidae andPlatypodidae were examined by the first author from the following collections (abbre-viations used are from Arnett & Samuelson (1986)): Archbold Biological Station, LakePlacid, Florida (ABSC); Canadian Museum of Nature Collection, Ottawa, Ontario(CMNC); E. G. Riley private collection, College Station, Texas (EGRC), Department ofForestry and Environmental Science, State University of New York, Syracuse, NewYork (DFEC), Florida State Collection of Arthropods, Gainesville, Florida (FSCA),Henry and Ann Howden collection, Ottawa, Ontario, (CHAH), Museum of Compara-

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida
315
tive Zoology, Cambridge, Massachusetts (MCZC), R.H. Turnbow private collection, Ft.Rucker, Alabama (RHTC), S.L. Wood private collection, Provo, Utah (SLWC); T.H. At-kinson private collection, Riverside, California (THAC), University of Georgia at Ath-ens Georgia (UGCA), and U. S.. National Museum of Natural History, Washington,D.C. (USNM).
In addition to data taken from museum specimens, the relevant literature on theScolytidae and Platypodidae was critically reviewed by the first author for specific dis-tribution records in southern Florida, as well as host records and distribution recordsfor the entire ranges of the species included.
A
DVENTIVE
S
PECIES
Exactly what constitutes an adventive species and unambiguous criteria by whichone might judge particular cases are matters of considerable debate (Frank & McCoy1990, 1992, Whitehead & Wheeler 1990). Whitehead & Wheeler (1990) argued that asa general criterion one should consider species to be adventive if newly reported (ordetected) in the United States. They recognized that southern Florida had not beenwell-collected historically and that new records for Antillean species did not necessar-ily imply that these species were non-native. Frank & McCoy followed this criterionliterally in their recent list of new immigrants into Florida (1992) (i.e., new recordmeans new arrival), although they also acknowledged the lack of complete historicalinformation on the insect fauna of tropical southern Florida.
Part of the problem is semantic (Frank & McCoy 1990). Many terms have beenused inconsistently in the literature to indicate natural occurrence, occurrencethrough human intervention (accidental or not), and various types of restricted dis-tributions. Frank & McCoy (1990) restricted the term “introduced” to mean speciesdeliberately brought into a new area. They used the term “immigrant” to refer to spe-cies that have arrived and become established without intentional human activities.Their definition is deficient in an important sense in that it does not distinguish be-tween the accidental establishment of species through human agency and the move-ment and establishment of species into historically new areas by natural means ofdispersal, passive or active. To further complicate matters, there is no reason to sup-pose that the process of accumulation of the biota of southern Florida through “natu-ral means” has come to a stop.
This distinction is not trivial for southern Florida. Given that southern peninsularFlorida (including the Keys) has only become emergent in Wisconsonian and recentgeological periods (Tomlinson 1980, Peck 1989) and has never had any land connec-tions to the Greater Antilles, virtually any species with an Antillean or Neotropicaldistribution currently found in Florida is immigrant by this definition. Nonetheless,most of these occur there naturally and should be considered native. These would notbe considered “indigenous” as defined by Frank & McCoy (1990). The same would alsobe true of any Neartic or Holarctic species occurring naturally in the Florida penin-sula as well, although it has been available for colonization by terrestrial organismssince the mid-late Tertiary and Pleistocene. Some Nearctic and Holartic species haveonly recently been reported from the peninsula (e.g., Deyrup & Atkinson 1987, Atkin-son et al. 1991), yet no one would think to call these “immigrants”.
To avoid confusion, we consistently use the terms “adventive” and “immigrant” torefer to species that do not naturally occur in southern Florida. None of the adventivespecies known from the area have been deliberately introduced in the sense of Frank& McCoy (1990). We use the term “native” to refer to any species which we believe oc-curs naturally in southern Florida whether “indigenous” (
sensu
Frank & McCoy 1990)

316
Florida Entomologist
77(3) September, 1994
or “immigrant” in the sense of having dispersed to Florida as part of a natural dis-persal process not mediated by deliberate or accidental human activity.
There is no evidence that any Neotropical species of Scolytidae or Platypodidaehas been introduced, intentionally or accidentally, into Florida in modern times. Suchan event might have occurred more than 150 years ago and passed unnoticed (i.e., be-fore the publication dates of works treating the area). Neotropical adventives may berecognized by any combination of the following criteria: (1) the main part of the spe-cies’ historical distribution does not include the Caribbean; (2) strict association witha non-native host plant; (3) clear association with disturbed habitats and / or absencefrom natural communities; and (4) documented invasion and / or subsequent spread.Several species that originated in the Old World tropics have become established inthis century in the United States and have shown an historical pattern of distributionconsistent with spread from a single point, even when the original invasion and colo-nization passed unremarked. Examples include
Xylosandrus germanus
(Blandford)(Bright 1968, Chapin & Oliver 1986, Staines 1984, Weber & MacPherson 1982),
X.compactus
(Eichhoff) (Dixon & Woodruff 1982, Ngoan et al. 1976),
X. crassiusculus
(Motschulsky) (Anderson 1974, Chapin & Oliver 1986, Deyrup & Atkinson 1987, Ko-vach & Gorsuch 1985, Atkinson et al. 1991), and
Xyleborus atratus
(Eichhoff) (Atkin-son et al. 1990, 1991). In all of these cases, either the historical pattern of spread canbe documented from collection records (
Xylosandrus
spp.) or else a “novel species”known from another continent is detected over a wide area almost simultaneously(
Xyleborus atratus
).We maintain that any species that is currently found in southern Florida and in
the Bahamas and / or the Greater Antilles should be considered native (although al-most certainly immigrant) unless there is evidence to the contrary. There are 24 spe-cies in our area that have this distribution pattern. By the same token, anyNeotropical species whose modern range includes the Gulf Coast of the southeasternUnited States and Texas and is continuously distributed into Mexico and CentralAmerica should also be considered native (12 species, some of which are also widelydistributed in the eastern United States and / or the Caribbean).
By our restricted criteria, we consider 20 species of Scolytidae found in southernFlorida to be immigrants, mostly from Africa or Asia. Frank & McCoy (1992), by a me-chanical application of the rule of “recent record means new immigrant ”, listed 3other species as immigrants (
Pseudothysanoes securigerus
(Blackman)
, Theoborussolitariceps
Schedl
,
and
Trischidias exigua
Wood) that we consider natives. Wood(1977, 1982) listed an additional 14 Neotropical species as immigrants with distribu-tions that fit one of the 2 patterns described above.
Cryptocarenus heveae
(Hagedorn)
,C. seriatus
(Eggers)
, Xyleborus volvulus
(F.)
, Xylosandrus zimmermanni
(Hopkins),and
Corthylus spinifer
Schwarz have ranges that include the Antilles and South Flor-ida.
Hypothenemus eruditus
Westwood
, H. seriatus
(Eichhoff)
, Xyleborus affinis
Eich-hoff
,
and
X. ferrugineus
(F.) are continuously distributed along the Gulf Coast intoMexico and Central America. In contrast, other Neotropical species with similar dis-tributions were treated as native.
Dendrosinus bourreriae
Schwarz
, Chaetophloeusinsularis
(Blackman)
, Pycnarthrum hispidum
(Ferrari)
, Scolytodes schwarzi
(Hop-kins)
,
and
Ambrosiodmus lecontei
Hopkins are found in the Antilles and South Flor-ida.
Cnesinus strigicollis
LeConte is continuously distributed along the Gulf Coastthrough Texas and into Mexico. In neither publication were the criteria explained bywhich this conclusion was reached. All of these putative “Neotropical exotics” are hostgeneralists (polyphagous) and many reproduce by inbred polygyny. As noted by Wood(1977), this is true of many species recently established in North America, but it doesnot constitute
a priori
evidence that a particular species is non-native.

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida
317
E
XCLUDED
S
PECIES
Three species previously reported from southern Florida,
Cryptocarenus spatula-tus
Wood,
Xyleborus xylographus
(Say), and
Araptus politus
(Blandford), probably donot occur there.
Cryptocarenus spatulatus
and
Araptus politus
were intercepted com-ing into Miami from Latin America (Wood 1982), but no specimens of either specieshave ever been collected from local populations.
Araptus politus
is a host specialistand breeds in seeds of
Mucuna
spp. (Leguminosae) which are not known to occur inFlorida.
Xyleborus xylographus
was cited by Bright (1968) and Wood (1982) from KeyWest (presumably from the same specimen(s) in the Canadian National Collection).This species breeds in oaks and no other authentic specimens have ever been found inpeninsular Florida despite extensive collecting by the first author in northern Florida.The only species of oak that occurs in southern Florida is
Quercus virginiana
L., foundonly in Dade County near Miami and on Key Largo (Long & Lakela 1971, Tomlinson1980). The “Key West” material most likely represents an error in labelling or an in-terception.
O
RGANIZATION
OF
C
HECKLIST
The sequence of subfamilies and tribes follows Wood & Bright (1992). Species arearranged alphabetically within genera, and genera alphabetically within tribes. Ab-breviations for feeding habits (host tissues consumed), degree of host specificity, andmating system are listed in parenthesis for each species. Host information and overalldistribution patterns are also included. Abbreviations are used to indicate: (1)
Feed-ing Habits
: ph, phloem feeding (= true bark beetles) (phloeophagy); xm, feeding on ec-tosymbiotic fungi (=ambrosia beetles) (xylomycetophagy); xy, direct feeding onsapwood (xylophagy); my, pith of twigs and branches (myelophagy); sp, seeds (sper-matophagy); myc, non-ambrosial fungi (mycophagy); (2)
Host Specificity
: mo, re-stricted to hosts of a single species or genus (monophagy); ol, restricted to hosts of asingle family (or to a very limited number of hosts) (oligophagy); po, not host specific(polyphagy); (3)
Mating Systems
: mg, monogyny; hpg, harem polygyny (including big-amy); ipg, inbred polygyny (mating terminology follows Kirkendall 1983). A questionmark (?) after any abbreviation indicates an absence of data.
Collection localities are listed by county for the four southern Florida counties inthe following sequence: Collier, Broward, Dade and Monroe. Each locality is followedby a parenthetical reference to the collection(s) in which specimens are deposited (ab-breviations listed in Methods) or a literature reference. Complete collection data arenot included because of space limitations, but are available from the authors. Speci-mens were examined by the first author for all localities for which a collection is des-ignated.
A
NNOTATED
C
HECKLIST
OF
THE
P
LATYPODIDAE
AND
S
COLYTIDAE
OF
S
OUTHERN
F
LORIDA
Family Platypodidae
Platypus compositus
Say. (xm, po, mg). Found in large trunks and stumps of mosthardwoods within its range, also bald cypress,
Taxodium distichum
(L.) Rich.. Widelydistributed in the Neotropics, apparently adventive to Africa. Southeastern U.S. from

318
Florida Entomologist
77(3) September, 1994
Texas eastward.
Collier:
Royal Palm Hammock St. Park (USNM);
Dade:
ParadiseKey (USNM);
Monroe:
Big Pine Key (CMNC); Sugarloaf Key (CMNC).
Platypus flavicornis
(F.). (xm, mo, mg). Found in trunks and stumps of all speciesof
Pinus
(Pinaceae) within its range. Southeastern U.S., from eastern Texas.
Dade:Homestead (FSCA); Opa-locka (FSCA).
Platypus parallelus (F.). (xm, po, mg). Breeds in large diameter host material ofvirtually any woody plant within its range. It has also been reported from palmtrunks. Widely distributed in the Neotropics. In the U.S. found only in southern Flor-ida and southern Texas. Broward: Fort Lauderdale(FSCA); Collier: Collier Semi-nole St. Park (FSCA); Marco Island (FSCA); 8 mi SE Naples, (USNM); Dade:Everglades Natl. Park, Royal Palm Hammock (CMNC); Homestead (FSCA); 6 mi NHomestead, (USNM); Miami (FSCA); Miami Beach (FSCA); Perrine (FSCA); West Mi-ami (FSCA); Monroe: Big Pine Key (EGRC); Big Torch Key (CMNC); Cape Sable(FSCA); John Pennekamp St. Park (ABSC); Key Largo (USNM, RHTC); Key West(FSCA); No Name Key (EGRC); Plantation Key (FSCA); Stock Island (FSCA, USNM).
Family ScolytidaeSubfamily HylesininaeTribe Hylastini
Hylastes salebrosus Eichhoff. (ph, mo, mg). Found in roots, stumps, or branches incontact with the soil of most Pines within its range. Southeastern U.S., from Texaseastward. Dade: “Biscayne Bay” (Wood 1982).
Hylastes tenuis Eichhoff. (ph, mo, mg). Found in roots, stumps or branches in con-tact with the soil of most Pines within its range. Southeastern U.S., from Texas east-ward, also known from Hispaniola. Southwestern U.S. to central Mexico in montaneregions. Monroe: Key West (USNM).
Tribe Tomicini
Dendroctonus terebrans (Olivier). (ph, mo, mg). Found in stumps, large roots, andthe lower portion of large pines. Southeastern U.S. from eastern Texas to Atlantic sea-board. Broward: Fort Lauderdale (Wood 1982); Dade: Kendall (FSCA); Opa-locka(FSCA).
Tribe Bothrosternini
Cnesinus strigicollis LeConte. (my, po, mg). Very polyphagous, breeding in twigs ofa wide variety of plant species. Southeastern U.S. from eastern Texas to lower Atlan-tic seaboard. In Mexico along Gulf Coast to Yucatan Peninsula. Dade: Elliot Key(ABSC); Miami (FSCA); Deering Estate (CMNC); Matheson Hammock (CMNC);Monroe: Key Largo (UGCA, CMNC, Wood 1982); N. Key Largo (ABSC); PlantationKey (FSCA, ABSC).
Pagiocerus frontalis (F.) (sp, mo, mg). Breeds in fleshy seeds of several genera ofLauraceae, notably Persea. Widely distributed in lowland Neotropical areas. In U.S.found along lower Gulf Coast and lower Atlantic seaboard to North Carolina. It hasnot been collected in the Keys but native and exotic species of Persea (avocado) do oc-cur there (Long & Lakela 1971, Tomlinson 1980). Collier: Monroe Sta. (ABSC);Dade: Chekika St. Rec. Area, 50 km SW Miami (CMNC); Miami, Deering Estate(CMNC).

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida319
Tribe Phloeotribini
Phloeotribus texanus Schaeffer. (ph, mo, mg). Breeds in branches of Celtis spp (Ul-maceae). Southeastern U.S. from Texas eastward; lowland regions of Mexico. Collier:Collier Seminole St. Park (ABSC); Dade: “Biscayne” (Wood 1982).
Tribe Phloeosinini
Dendrosinus bourreriae Schwarz (xy, po, mg). Reported from branches of severalunrelated families of tropical hardwoods. Known only from southern Florida andGreater Antilles. This species has not been collected from the mainland althoughsome of its reported hosts are found in tropical hardwood hammocks there (Long &Lakela 1971, Tomlinson 1980). Monroe: Key Largo (FSCA); Stock Island (FSCA).
Phloeosinus taxodii taxodii Blackman. (ph, mo, mg). Breeds in branches of bald cy-press, Taxodium distichum (Taxodiaceae). Southeastern U.S. from eastern Texas. Adifferent subspecies, P. taxodii taxodiicolens Wood, is found throughout Mexico inMontezuma cypress, Taxodium mucronatum Ten. Collier: Naples, 8 mi SE (USNM).
Tribe Hypoborini
Chaetophloeus insularis (Blackman). (ph, mo, mg). Breeds in branches of nativeand exotic Manilkara spp. (Sapotacae). Southern Florida and Greater Antilles. Thisspecies has not been collected from the mainland although its native host, Manilkarabahamensis (Baker) Lam. & Meeuse, is found in there and an exotic host, M. zapota(L.) Royen is grown widely in southern Florida (Long & Lakela 1971, Tomlinson1980). Monroe: Big Pine Key, Cactus Hammock (CMNC); Big Torch Key (CMNC);Cudjoe Key (CMNC); Fat Deer Key (CMNC); Key West (Wood 1982); Lower Mate-cumbe Key (Wood 1982); No Name Key (CMNC); Sugarloaf Key (CMNC, Wood 1982).
Subfamily ScolytinaeTribe Scolytini
Cnemonyx ficus (Schwarz). (ph, mo?, mg). The type series was reportedly collectedfrom Ficus (Moraceae), probably due to an error in identification. The true host is themanchineel tree, Hippomane mancinella L. (Euphorbiaceae) (Atkinson 1993). Thisspecies breeds in larger branches (> 3 cm diameter) and trunks of its host tree. Thehost occurs throughout the Keys and on the mainland near Cape Sable, although it isnot abundant (Long & Lakela 1971, Tomlinson 1980). Southern Florida, Bahamas,Virgin Islands. Monroe: Big Pine Key (CMNC, THAC); Key West (MCZC, Wood1982).
Cnemonyx vagabundus Wood. (ph, mo?, mg). The type series was reportedly col-lected from Piscidia piscipula (Leguminosae), probably due to an error in identifica-tion. The true host is the manchineel tree, Hippomane mancinella L. (Euphorbiaceae)(Atkinson 1993). This species breeds in smaller branches (< 3 cm diameter) andtrunks of its host tree. Southern Florida, Puerto Rico and Panama. Monroe: Big PineKey (CMNC, THAC); Key West (Wood 1982).
Tribe Ctenophorini
Pycnarthrum hispidum (Ferrari). (ph, mo, mg). Branches of native and exotic figs(Ficus spp.) (Moraceae). Widely distributed in lowland Neotropical areas. In the U.S.

320 Florida Entomologist 77(3) September, 1994
known only from southern Florida and southern Texas. Broward: 1 mi N Andytown,(RHTC); Dade: “Biscayne” (MCZC); Coconut Grove (MCZC); Homestead (SLWC); Mi-ami (FSCA); Monroe: Key Largo (UGCA); Plantation Key (FSCA); Stock Isl. (FSCA);Sugarloaf Key (SLWC).
Scolytodes schwarzi (Hopkins). (ph, mo, hpg). Breeds in shaded-out branches ofliving Ficus spp. (Moraceae). Based on personal experience (THA), this species doesnot breed in cut branches. Southern Florida, Greater Antilles, lowland regions of Mex-ico. Broward: 1 mi N Andytown (RHTC); Collier: Corkscrew Swamp Sanctuary(ABSC); Dade: “Biscayne” (MCZC); Deering Estate Park (CMNC); Matheson Ham-mock (ABSC); Miami (Wood 1982); Monroe: Big Pine Key (CMNC); Everglades Natl.Park (Wood 1982); Key Largo (ABSC, Wood 1982); Plantation Key (Wood 1982); Sug-arloaf Key (SLWC).
Tribe Micracini
Micracis swainei Blackman. (xy, po, hpg). Reported from branches of a variety ofhardwood species. It is commonly found in willow (Salix) in Florida. SoutheasternU.S. from eastern Texas, lowland regions of Mexico and Central America. Dade: 6 miN. Homestead (USNM).
Micracisella nanula (LeConte). (my, po, mg). Breeds in twigs of a variety of trees,shrubs, and vines. Southeastern U.S. from Texas eastward. A closely related species,M. opacithorax Schedl (possibly conspecific) is found in southern Texas and the GulfCoast of Mexico. Dade: “Biscayne” (DFEC, MCZC); Homestead (Wood 1982); Miami(Wood 1982); Monroe: Big Pine Key (ABSC, Wood 1982); Key Largo (Wood 1982); KeyVaca, Marathon (Wood 1982); Key West (Wood 1982, Blackman 1928); Missouri Key(Wood 1982); N. Key Largo (ABSC); Sugarloaf Key (CMNC, Wood 1982).
Pseudothysanoes securigerus (Blackman). (?,?,?). Nothing is known about the hostsor feeding habits of this species. Species of Pseudothysanoes may be either phloem orsap-wood borers. All studied species are bigynous. Southern Florida, Hispaniola.Dade: Everglades Natl. Park, Long Pine Key (CMNC); Monroe: Big Pine Key(CMNC); No Name Key (CMNC).
Thysanoes fimbricornis LeConte. (xy, po, hp). Breeds in branches of a variety ofhardwood species. Southeastern U.S. from eastern Texas. Found in lowland areas ofeastern and southeastern Mexico. Dade: “Biscayne” (Blackman 1928).
Tribe Ipini
Ips avulsus (Eichhoff). (ph, mo, hp). Breeds in branches and crowns of all pineswithin its range. Southeastern U.S. from eastern Texas. Dade: “Biscayne” (MCZC);Miami (MCZC).
Ips calligraphus (Germar). (ph, mo, hp). Breeds in trunks and large branches of allpines within its range. Eastern U.S. and southeastern Canada, Montane regions ofthe southwestern U.S., Mexico, Guatemala and Honduras. Cuba and Hispaniola.Dade: Everglades Natl. Park, Paradise Key (Hopping 1965b); Monroe: Key Largo(Hopping 1965b); Key West (Hopping 1965b).
Ips grandicollis (Eichhoff). (ph, mo, hpg). Breeds in branches and trunks of allpines within its range. Eastern U.S. and southeastern Canada, Montane regions ofthe southwestern U.S., Mexico, Guatemala and Honduras. Cuba and Hispaniola.Dade: Biscayne (MCZC); Everglades Natl. Park, Long Pine Key (CMNC); EvergladesNatl. Park, Paradise Key (Hopping 1965a); Homestead (Hopping 1965a); Monroe:Big Pine Key (CMNC).

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida321
Tribe Dryocoetini
Coccotrypes advena (Blandford). (sp, po, ipg). Breeds in seeds of a variety of plants.Widely distributed in Old World tropics, adventive in New World. Dade: Coral Gables(FSCA). This species was intercepted in imported seeds and may not actually be es-tablished in Florida.
Coccotrypes carpophagus (Hornung). (sp, ol, ipg). Breeds in seeds of palms, espe-cially Sabal palmetto (Walt.) Lodd. ex Schultes and Washingtonia robusta Wendl.Widely distributed in Old World tropics, adventive in New World. Dade: CoconutGrove (Wood 1982); Deering Estate Park (CMNC); Everglades Natl. Park, Royal PalmHammock (CMNC); Homestead (ABSC); Matheson Hammock (CMNC); Monroe: BigPine Key (CMNC); Big Torch Key (CMNC); Cudjoe Key (CMNC); Fat Deer Key(CMNC); Grassy Key (ABSC); Key Largo (CMNC); Key Vaca, Marathon (CMNC,MCZC); No Name Key (CMNC); Plantation Key (ABSC); Sugarloaf Key (CMNC).
Coccotrypes cyperi (Beeson). (sp-ph, po, ipg). Extremely polyphagous. Breeds inphloem and seeds of many hosts. Widely distributed in Old World tropics, adventivein New World. Dade: Deering Estate Park (CMNC); Homestead (ABSC); Miami(FSCA); Monroe: Big Pine Key (CMNC); Fat Deer Key (CMNC).
Coccotrypes dactyliperda (F.). (sp, ol, ipg). Breeds in seeds of palms, especiallyPhoenix spp. Widely distributed in Old World tropics, adventive in New World. Mon-roe: Sugarloaf Key (CMNC).
Coccotrypes distinctus (Motschulsky). (sp, ol, ipg). Breeds in seeds of palms, espe-cially Phoenix spp., Sabal palmetto, and Washingtonia robusta. Widely distributed inOld World tropics, adventive in New World. Collier: Collier Seminole St. Park(ABSC); Dade: Chekika State Rec. Area (CMNC); Coconut Grove (Wood 1982); Deer-ing Estate Park (CMNC); Everglades Natl. Park, Long Pine Key (CMNC); FairchildTropical Gardens (FSCA); Homestead (ABSC); Matheson Hammock (CMNC); Miami(FSCA); Miami Beach (Wood 1982); Old Cutler Hammock (CMNC); Monroe: Planta-tion Key (ABSC); Stock Isl. (ABSC, CHAH, CMNC).
Coccotrypes rhizophorae (Hopkins). (sp, ol, ipg). Hosts: (Rhizophoraceae) Breeds inexpanding prop roots and propagules of its host, Rhizophora mangle L. All other spe-cies of Coccotrypes found in the New World are believed to be immigrants from Africaor Asia. C. rhizophorae is a special case and possible exception because it may haveimmigrated to the New World without human aid or intervention. Coccotrypes rhizo-phorae is also known from southeastern Asia in Rhizophora spp. Given that this bee-tle breeds in the fruits and developing seedlings of red mangrove which are capableof floating long distances in salt water, it is entirely plausible that this insect was dis-persed to the New World by infested host propagules, just as its host presumably dis-persed to the New World by ocean currents (Tomlinson 1980). Collier: EvergladesCity (USNM); Dade: Homestead (Wood 1982); Miami (MCZC, THAC); Miami, Deer-ing Estate (CMNC); Monroe: Key Largo (ABSC, USNM); Key West (Wood 1982).
Coccotrypes robustus Eichhoff (sp, ?, ipg). Reported from seeds of Euterpe sp. (Pal-mae). No host records from Florida. This species is known only from southern Floridaand the Greater Antilles. It is presumably of Old World origin as are all other speciesin this genus. Dade: Deering Estate Park (CMNC); Monroe: Big Pine Key (CMNC);Big Torch Key (CMNC); Cudjoe Key (CMNC); Fat Deer Key (CMNC); Key Vaca(CMNC); No Name Key (CMNC); Sugarloaf Key (THAC).
Coccotrypes vulgaris (Eggers). (sp, po, ipg). Breeds in seeds of many hosts. Widelydistributed in Old World tropics, immigrant in New World. Dade: Everglades Natl.Park, Long Pine Key (CMNC).
Dendrocranulus carbonarius (Hopkins). (my, ol?, mo?). All species of Dendrocran-ulus with known habits breed in the stems of cucurbit vines. Presumably D. carbon-

322 Florida Entomologist 77(3) September, 1994
arius does so as well. The original host record from Annona is almost certainly due toan error in labelling. Found in southern Florida and Caribbean Islands. This specieshas not been collected in Florida since its original description. Dade: “Biscayne Bay”(Wood 1982).
Tribe Crypturgini
Crypturgus alutaceus Schwarz. (ph, mo, mg). Breeds in phloem of dead or dyingpines (Pinaceae). Southeastern U.S. from eastern Texas to Atlantic seaboard. Collier:Collier Seminole St. Park (Wood 1982); Monroe: Big Pine Key (Wood 1982).
Tribe Xyleborini
Ambrosiodmus devexulus (Wood). (xm, po, ipg). In wide variety of hosts. Dade:Homestead (Wood 1982).
Ambrosiodmus lecontei Hopkins. (xm, po, ipg). Wide variety of hosts. Usuallyfound in branches or other small diameter material. Peninsular Florida and Carib-bean. Dade: “Biscayne” (MCZC); Everglades Natl. Park, Long Pine Key (CMNC); Ev-erglades Natl. Park, Royal Palm Hammock (CMNC); Homestead (FSCA, Wood 1982);Miami (FSCA, Wood 1982); North Miami (FSCA); Monroe: Key Largo (CMNC, Wood1982); Key Vaca (CMNC); Key West (Wood 1982); No Name Key (CMNC); SugarloafKey (CMNC).
Ambrosiodmus obliquus (LeConte). (xm, po, ipg). Wide variety of hosts. Widely dis-tributed in lowland Neotropical areas. Southeastern U.S. Dade: Everglades Natl.Park, Long Pine Key (CMNC); Everglades Natl. Park, Royal Palm Hammock(CMNC); Homestead (Bright 1968); Monroe: Cudjoe Key (CMNC); No Name Key(CMNC); Sugarloaf Key (CMNC).
Dryocoetoides sp. (xm, po?, ipg). Hosts unknown. A single specimen was collectedin a flight-intercept trap. It may represent an undescribed species because it does notagree with any species of this genus previously reported from the Caribbean or Me-soamerica and is not of any species found in the USNM or SLWC. Monroe: Big TorchKey (CMNC).
Premnobius cavipennis Eichhoff. (xm, po, ipg). Breeds in trunks of a wide varietyof hosts. Native to Africa, widely distributed in lowland Neotropical areas. In the U.S.,found in southern Florida only. Collier: Collier Semenole St. Park (Bright 1968);Dade: Chekika State Rec. Area (CMNC); Everglades Natl. Park, Long Pine Key(CMNC); Everglades Natl. Park, Royal Palm Hammock (CMNC); Homestead (Bright1968); Miami (Wood 1982); Monroe: Big Pine Key (CMNC); Flamingo Camp, Ever-glades (ABSC); Key Largo (CMNC); No Name Key (CMNC); Sugarloaf Key (CMNC).
Theoborus solitariceps Schedl. (xm, po, ipg). Found in branches of a wide varietyof hosts. Widely distributed in lowland Neotropical areas including the Caribbean.Known only from a single collection in Florida. Dade: Deering Estate Park (CMNC).
Xyleborinus gracilis (Eichhoff). (xm, po, ipg). Breeds in a wide variety of hosts.Lowland Neotropical areas. In U.S. known from Florida, Louisiana and North Caro-lina (Bright 1987). Dade: Biscayne Bay (Wood 1982); Everglades Natl. Park, LongPine Key (CMNC).
Xyleborinus saxeseni (Ratzeburg). (xm, po, ipg). Breeds in virtually any woodyplant within its range, including conifers. Eurasian species, immigrant in the NewWorld. Dade: Deering Estate Park (CMNC); Everglades Natl. Park, Long Pine Key(CMNC).

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida323
Xyleborus affinis (Eichhoff). (xm, po, ipg). Breeds in trunks of most woody plantswithin its range. Widely distributed in lowland Neotropical areas, southeastern U.S.Dade: Chekika State Rec. Area (CMNC); Deering Estate Park (CMNC); EvergladesNatl. Park, Long Pine Key (CMNC); Everglades Natl. Park, Royal Palm Hammock(CMNC); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); Cudjoe Key(CMNC); Key Largo (CMNC, RHTC); Key West (MCZC); Sugarloaf Key (CMNC).
Xyleborus ferrugineus (F.). (xm, po, ipg). Breeds in trunks of most woody plants inits range. Widely distributed in lowland neotropical areas, eastern U.S., southeasternCanada. Dade: Chekika State Rec. Area (CMNC); Coconut Grove (MCZC); DeeringEstate Park (CMNC); Everglades Natl. Park, Long Pine Key (CMNC); EvergladesNatl. Park, Royal Palm Hammock (CMNC); Monroe: Big Pine Key (CMNC); BigTorch Key (CMNC); Cudjoe Key (CMNC); Fat Deer Key (CMNC); Key Largo (CMNC,RHTC); Key Vaca (CMNC); No Name Key (CMNC); Sugarloaf Key (CMNC).
Xyleborus pubescens (Zimmermann). (xm, mo, ipg). Breeds in trunks of pines (Pi-naceae). This is one of the few species of this genus that is host specific. SoutheasternU.S. Collier: Collier Seminole St. Park (Wood 1982); Dade: Chekika State Rec. Area(CMNC); Dade Co. (Wood 1982); Everglades Natl. Park, Long Pine Key (CMNC); Par-adise Key (Wood 1982); Monroe: Big Pine Key (CMNC); Key Largo (UGCA); KeyWest (Bright 1968).
Xyleborus volvulus (F.). (xm, po, ipg). Breeds in trunks of most woody plants in itsrange. Widely distributed in lowland Neotropical areas. In U.S. only in southern Flor-ida and southern Texas. Broward: Ft. Lauderdale (FSCA); Collier: Naples (FSCA,Bright 1968); Dade: Biscayne Bay (Bright 1968); Chekika State Rec. Area (CMNC);Coconut Grove (MCZC); Coral Gables (FSCA); Deering Estate Park (CMNC); Ever-glades Natl. Park, Long Pine Key (CMNC); Everglades Natl. Park, Royal Palm Ham-mock (CMNC); Homestead (FSCA); Miami (FSCA); Miami Beach (FSCA); Perrine(FSCA); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); Cudjoe Key(CMNC); Fat Deer Key (CMNC); Key Largo (CMNC, FSCA, RHTC); Key Vaca(CMNC); Key West (Bright 1968); Loggerhead Key (FSCA); No Name Key (CMNC);Plantation Key (FSCA); Sugarloaf Key (CMNC).
Xyleborus xylographus (Say). (xm, mo, ipg). This species breeds in Oaks (Quercussp.) (Fagaceae). The record from Key West (Bright1968, Wood 1982) is either based ona misidentification or an interception since this species is not known to breed any-where in peninsular Florida. (T.H.A., unpublished).
Xylosandrus compactus (Eichhoff). (xm, mo, ipg). Breeds in twigs of a wide varietyof living trees, shrubs, and vines resulting in the death of the twig. This species sel-dom breeds in cut material. Native to southeastern Asia. Found along Gulf Coast toTexas. Broward: Ft. Lauderdale (Wood 1982); Collier: Collier Seminole St. Park(ABSC); Copeland (ABSC); Dade: Chekika State Rec. Area (CMNC); Coconut Grove(Wood 1982); Deering Estate Park (CMNC); Everglades Natl. Park, Long Pine Key(CMNC); Everglades Natl. Park, Royal Palm Hammock (CMNC); Miami (Wood 1982);Naranja (FSCA); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); CudjoeKey (CMNC); Key Largo (CMNC, Wood 1982); N Key Largo (ABSC); No Name Key(CMNC); Plantation Key (ABSC); Sugarloaf Key (CMNC).
Xylosandrus crassiusculus (Motschulsky). (xm, po, ipg). Breeds in stems of a widevariety of trees and shrubs. This species frequently breeds in living stems, especiallynear ground level. Secondary fungal invasions of old galleries may result in death ofthe plant. Native to Asia. First detected in coastal South Carolina, now found west toTexas (Atkinson et al. 1991). Collier: Collier Seminole St. Park (Deyrup & Atkinson1987); Dade: Chekika State Rec. Area (CMNC); Deering Estate Park (CMNC); Ever-

324 Florida Entomologist 77(3) September, 1994
glades Natl. Park, Long Pine Key (CMNC); Everglades Natl. Park, Royal Palm Ham-mock (CMNC); Old Cutler Hammock (CMNC).
Xylosandrus zimmermanni (Hopkins). (xm, po, ipg). Breeds in small diameter ma-terial. Not known to attack living hosts. Lowland Neotropical areas, southern Florida.Dade: Biscayne Bay (Bright 1968); Everglades Natl. Park, Long Pine Key (CMNC);Everglades Natl. Park, Long Pine Key (CMNC).
Tribe Cryphalini
Cryptocarenus heveae (Hagedorn). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Widely distributed in lowland Neotropical areas, southern Florida.Collier: Collier Seminole St. Park (ABSC, Wood 1982); Dade: Everglades Natl. Park,Long Pine Key (CMNC).
Cryptocarenus seriatus Eggers. (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Widely distributed in lowland Neotropical areas, peninsular Florida.Collier: Collier Seminole St. Park (ABSC, Wood 1982)); Copeland (ABSC); Faka-hatchee Strand (ABSC); Ochopee (Wood 1982); Dade: “Biscayne” (Wood 1982); Deer-ing Estate Park (CMNC); Entrance Everglades Natl. Park (RHTC); Everglades Natl.Park, Long Pine Key (CMNC); Hialeah (FSCA); Miami (FSCA); Paradise Key (Wood1982); Monroe: Big Pine Key (Wood 1982); Big Torch Key (CMNC); Fat Deer Key(CMNC); Grassy Key (Wood 1982); Key Largo (Wood 1982); Key Vaca (Wood 1982);Key West (Wood 1982); Lower Matecumbe Key (Wood 1982); Stock Island (FSCA).
Cryptocarenus spatulatus Wood. (xm, po, ipg). The record from Miami (Wood 1982)is based on an interception from Peru. There is no indication at present that it is ac-tually established in Florida.
Hypocryphalus mangiferae Schedl. (ph, mo, mg). Breeds in branches of mango(Mangifera indica L.) (Anacardiaceae). Native to southern Asia, widely established intropical areas of the world where its host is grown, including southern Florida. Dade:Homestead (Wood 1982); Perrine (Wood 1954).
Hypothenemus areccae (Hornung). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Immigrant from Old World tropics. Dade: Everglades Natl. Park,Long Pine Key (CMNC); Homestead (FSCA).
Hypothenemus birmanus (Eichhoff). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Immigrant from Old World Tropics. Broward: Andytown, 1 mi N(RHTC); Hollywood (ABSC); Collier: Collier Seminole St. Park (ABSC, Wood 1982);Fakahatchee Strand (ABSC); Dade: Chekika State Rec. Area (CMNC); Deering Es-tate Park (CMNC); Everglades Natl. Park, Long Pine Key (CMNC); Everglades Natl.Park, Royal Palm Hammock (CMNC); Homestead (ABSC, Wood 1982); MathesonHammock (THAC, Wood 1982); Miami (Wood 1982); Perrine (Wood 1982); Monroe:Big Pine Key (CMNC); Cudjoe Key (CMNC); Key Largo (UGCA, CMNC, Wood 1982);Key Vaca (CMNC); Lower Matecumbe Key (ABSC); N. Key Largo (ABSC); No NameKey (CMNC); Sugarloaf Key (CMNC).
Hypothenemus brunneus (Hopkins). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Immigrant from Old World tropics. Collier: Copeland (ABSC);Dade: “Biscayne” (MCZC); Everglades Natl. Park, Long Pine Key (CMN
C); Homestead (Wood 1982); Miami (Wood 1982); Monroe: Key Largo (Wood1982); Key West (Wood 1982); Matecumbe Key (Wood 1982); Plantation Key (ABSC,FSCA); Sugarloaf Key (Wood 1982).
Hypothenemus californicus Hopkins. (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Immigrant from Old World tropics. Dade: Homestead (Wood 1982);

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida325
Perrine (Wood 1982); Monroe: Key Largo (Wood 1982); Key Vaca (Wood 1982); KeyWest (Wood 1982); Matecumbe Key (Wood 1982); Plantation Key (Wood 1982).
Hypothenemus columbi Hopkins. (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Immigrant from Old World tropics. Dade: Homestead (Wood 1982);Perrine (Wood 1982).
Hypothenemus crudiae (Panzer). (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Immigrant from Old World tropics. Broward: Davie (FSCA); Collier:Collier Seminole St. Park (ABSC); Monroe Sta. (ABSC); Dade: Chekika State Rec.Area (CMNC); Elliot Key (ABSC); Everglades Natl. Park, Long Pine Key (CMNC); Hi-aleah (FSCA); Homestead (FSCA); Monroe: Big Pine Key (CMNC); Big Torch Key(CMNC); Cudjoe Key (CMNC); Fat Deer Key (CMNC); Key Largo (CMNC, ABSC); NoName Key (ABSC); Sugarloaf Key (CMNC).
Hypothenemus dissimilis (Zimmermann). (my, po, ipg). Breeds in pith of twigs ofa wide variety of hosts. Eastern U.S. Dade: “Biscayne Bay” (Wood 1982).
Hypothenemus eruditus Westwood. (ph-my, po, ipg). Breeds in pith or phloem oftwigs and branches of a wide variety of hosts. Widely distributed in lowland Neotro-pical areas, eastern U.S. Broward: Andytown, 1 mi N (RHTC); Collier: Collier Sem-inole St. Park (ABSC); Copeland (ABSC); Dade: Chekika State Rec. Area (CMNC);Deering Estate Park (CMNC); Elliot Key (ABSC); Everglades Natl. Park, Long PineKey (CMNC); Everglades Natl. Park, Royal Palm Hammock (CMNC); Homestead(ABSC); Old Cutler Hammock (CMNC); Monroe: Big Pine Key (CMNC); Big TorchKey (CMNC); Cudjoe Key (CMNC); Fat Deer Key (CMNC); Islamorada (UGCA); KeyLargo (CMNC); Key Vaca (CMNC); No Name Key (CMNC); Upper Matecumbe Key(CMNC).
Hypothenemus gossypii (Hopkins). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Mexico, Cuba, southern Florida. Dade: Everglades Natl. Park, LongPine Key (CMNC, THAC); Homestead (Wood 1982); Monroe: Key Largo (Wood 1982);Key West (Wood 1982); Long Key (Wood 1982); Matecumbe Key (Wood 1982); Planta-tion Key (Wood 1982).
Hypothenemus hirsutus (Wood). (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Known from southern Florida only. Dade: Elliott Key (ABSC); Mon-roe: Big Pine Key (CMNC, Wood 1982); Big Torch Key (CMNC); Cudjoe Key (CMNC);Fat Deer Key (CMNC); Grassy Key (Wood 1982); Key Largo (Wood 1982); Key Vaca(Wood 1982); Key West (Wood 1982); Matecumbe Key (Wood 1982); No Name Key(ABSC, CMNC); Plantation Key (Wood 1982); Sugarloaf Key (CMNC, Wood 1982).
Hypothenemus interstitialis (Hopkins). (my, po, ipg). Breeds in pith of twigs of awide variety of hosts. Widely distributed in lowland Neotropical areas, southeasternU.S. Collier: Collier Seminole St. Park (ABSC); Fakahatchee Strand (ABSC); Mon-roe Station (ABSC); Dade: Deering Estate Park (CMNC); Everglades Natl. Park,Long Pine Key (CMNC); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); KeyLargo (ABSC, CMNC, UGCA); Key Vaca (CMNC); No Name Key (ABSC); SugarloafKey (CMNC).
Hypothenemus javanus (Eggers). (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Immigrant from Old World. Broward: Delray Beach (Wood 1982); Hol-lywood (ABSC); Collier: Collier Seminole St. Park (Wood 1982); Dade: Chekika StateRec. Area (CMNC); Coconut Grove (Wood 1982); Deering Estate Park (CMNC); Ever-glades Natl. Park, Long Pine Key (CMNC); Everglades Natl. Park, Royal Palm Ham-mock (CMNC); Homestead (Wood 1982); Miami (Wood 1982); Paradise Key (Wood1982); Perrine (Wood 1982); Monroe: Key Largo (CMNC, Wood 1982); Key Vaca(CMNC); N. Key Largo (ABSC); No Name Key (CMNC); Sugarloaf Key (CMNC).

326 Florida Entomologist 77(3) September, 1994
Hypothenemus miles (LeConte). (my?, ?, ipg). Hosts unknown. Gulf Coast fromFlorida to Texas. Dade: Everglades Natl. Park, Long Pine Key (CMNC); Monroe:Sugarloaf Key (CMNC).
Hypothenemus obscurus (F.). (my, po, ipg). Breeds in pith of twigs of a wide varietyof hosts. Immigrant from Old World. Dade: Homestead (FSCA); Miami (Wood 1982);Monroe: Fat Deer Key (CMNC); Key Vaca (CMNC); No Name Key (CMNC); Sugar-loaf Key (CMNC).
Hypothenemus pubescens (Hopkins). (my, ol, ipg). Breeds in stems of grasses. Mex-ico, Puerto Rico, southern Florida, southern Texas. Dade: Everglades Natl. Park,Long Pine Key (CMNC); Monroe: Cudjoe Key (CMNC); Fat Deer Key (CMNC); KeyLargo, Pennekamp St. Park (CMNC); Key Vaca (Wood 1982); Key West (Wood 1982);Missouri Key (Wood 1982).
Hypothenemus seriatus (Eichhoff). (my, po, ipg). Breeds in pith of twigs of a widevariety of hosts. Widely distributed on lowland Neotropical areas, eastern U.S. Bro-ward: Davie (FSCA); Ft. Lauderdale (USNM); Dade: Everglades Natl. Park, LongPine Key (CMNC); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); CudjoeKey (CMNC, FSCA); Fat Deer Key (CMNC); Islamorada (UGCA); Key Largo (UGCA,CMNC); Key West (FSCA); No Name Key (CMNC); Stock Isl. (CMNC, FSCA); Sugar-loaf Key (CMNC); Windley Key (CHAH).
Hypothenemus setosus (Eichhoff). (my, po, ipg). Breeds in pith of twigs of a wide va-riety of hosts. Native to Old World, southern Florida. Dade: Miami (Wood 1982).
Hypothenemus squamosus (Hopkins). (my, po, ipg). Breeds in pith of twigs of awide variety of hosts. Lowland areas of Mexico, Central America, and the Antilles.Southern Florida. Monroe: Key Largo (FSCA, Wood 1982); Matecumbe Key (Wood1982).
Hypothenemus sp. (my, po, ipg). Breeds in pith of twigs of a wide variety of hosts.Southern Florida, Cuba. Collier: 1 mi W Ochopee (THAC); Dade: Everglades Natl.Park, Long Pine Key (CMNC); Homestead (ABSC); Monroe: Big Pine Key (CMNC);Key Largo (CMNC); Key Vaca (CMNC). This species does not match any included forNorth and Central America (Wood1982) or from the Caribbean (Bright 1985). It wascompared to all available types in the U.S. National Museum of Natural History (in-cluding types of Hopkins) and the S.L. Wood collection. It may be a previously unde-tected and undescribed native or Antillean species or may have been introduced fromthe Old World.
Scolytogenes knabi (Hopkins). (ph-my, po?, mg). Breeds in pith of twigs of a widevariety of hosts. Lowland Neotropical areas, Peninsular Florida. Collier: CollierSeminole St. Park (ABSC, ABSC); Dade: Miami (Wood 1982); Chekika State Rec.Area (CMNC); Monroe: Crawl Key (RHTC); Fat Deer Key (CMNC); Plantation Key(Wood 1982); Sugarloaf Key (Wood 1982).
Trischidias atoma (Hopkins). (myc, ?, ipg). Breeds in fungus-infested twigs andbranches. Southeastern U.S. Collier: Copeland (ABSC); Dade: Everglades Natl.Park, Long Pine Key (CMNC); Monroe: No Name Key (CMNC); Sugarloaf Key(CMNC).
Trischidias exigua Wood. (myc, ?, ipg). Breeds in carbonaceous fruiting bodies ofascomycete fungi on branches (Deyrup 1987). Known from southern Florida and theYucatan Peninsula. Dade: Everglades Natl. Park, Long Pine Key (CMNC); Old Cut-ler Hammock (CMNC); Monroe: Big Pine Key (CMNC); Big Torch Key (CMNC); KeyLargo (CMNC); Sugarloaf Key (CMNC).
Trischidias minutissima Wood. (myc, ?, ipg). Found in fungus pustules on aerialroots of red mangrove. Known only from Keys. Monroe: Sugarloaf Key (SLWC).

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida327
Trischidias striatus Atkinson. (?, ?, ?). Hosts unknown. Known only from Keys.Dade: Everglades Natl. Park, Long Pine Key(CMNC); Monroe: Big Pine Key(CMNC); Big Torch Key (CMNC); Cudjoe Key (CMNC); Key Largo (CMNC); No NameKey (CMNC); Sugarloaf Key (CMNC).
Tribe Corthylini
Araptus dentifrons Wood. (my, mo, mg). In stems of milkweed vine (Sarcostemmaclausum (Jacq.) R. & S.) (Asclepiadaceae). Known from several localities in Mexico,southern Texas and southern Florida. Collier: Fakahatchee Strand (ABSC); MonroeStation (ABSC).
Araptus politus (Blandford). This species was intercepted in Miami in “bird seed”but there is no evidence that it has become established (Wood 1982). It breeds in thelarge seeds of Mucuna spp. (Leguminosae) which are not known to occur in Florida.
Corthylus papulans Eichhoff (= spinifer Schwarz). (xm, po, mg). Breeds inbranches of a wide variety of hosts. Widely distributed in lowland neotropical areas,peninsular Florida. Broward: Andytown, 1 mi N (RHTC); Collier: Copeland (ABSC);Ochopee Trail Lake campsite (UGCA); Dade: Everglades Natl. Park, Long Pine Key(CMNC); Miami (FSCA, Wood 1982); Paradise Key (Wood 1982); Monroe: Key Largo(UGCA); Ramrod Key (Wood 1982); Sugarloaf Key (CMNC).
Monarthrum mali (Fitch). (xm, po, hpg). Breeds in trunks of a variety of hardwoodspecies. Widely distributed in eastern North America. Collier: Collier Seminole St.Park (Wood 1982).
Pityoborus comatus (Zimmermann). (xm, mo, mg). In shaded-out branches of liv-ing pines. Southeastern U.S., Bahamas. Dade: “Biscayne Bay” (Wood 1982); Ever-glades Natl. Park, Long Pine Key (UGCA); Monroe: Big Pine Key (Wood 1982).Pityophthorus annectens LeConte. (ph, mo, hpg). In branches of pines. SoutheasternU.S. Montane areas of southwestern U.S., Mexico, Central America, and Caribbean.Dade: Homestead (Bright 1981); Monroe: Big Pine Key (Bright 1981).
Pityophthorus borrichiae Wood. (ph, mo, hpg). Stems of Borrichia arborescens (L.)DC., B. frutescens (L.) DC. (Compositae). Known only from the Keys, although bothhosts are found on the mainland as well (Long & Lakela 1971, Tomlinson 1980). Mon-roe: Key Largo (SLWC).
Pityophthorus concentralis Eichhoff. (ph, mo, hpg). Branches and trunks of Me-topium toxiferum (L.) Krug & Urban (Anacardiaceae). Southern Florida, Cuba. Col-lier: Royal Palm Hammock St. Park (Bright 1981); Dade: “Biscayne Bay” (MCZC,Wood 1982); Deering Estate Park (CMNC); Everglades Natl. Park, Long Pine Key(CMNC); Everglades Natl. Park, Royal Palm Hammock (CMNC); Paradise Key(Bright 1981); Monroe: Big Pine Key (CMNC, Bright 1981); Big Torch Key (CMNC);Fat Deer Key (CMNC); Everglades Natl. Pk., Flamingo (RHTC); Key Largo (THAC,UGCA); Key Vaca (Wood 1982, Bright 1981); Key West (Wood 1982); No Name Key(ABSC, CMNC); Plantation Key (Wood 1982, Bright 1981); Stock Isl. (Wood 1982);Sugarloaf Key (CMNC, Bright 1981, Wood 1982).
Pityophthorus crinalis Blackman. (ph, mo, hpg). Toxicodendron radicans (L.)Kuntze (Anacardiaceae). This species has not been collected in the Keys although itshost is found there (Long & Lakela 1971). Southeastern U.S. Collier: Collier Semi-nole St. Park (ABSC); Dade: S. Miami, Deering Estate Park (CMNC).
Pityophthorus lautus Eichhoff. (ph, mo, hpg). Toxicodendron radicans (Anacardi-aceae). Widely distributed in eastern North America, possibly a species complex.Specimens referred to Pityophtorus lautus have been collected in several unrelated

328 Florida Entomologist 77(3) September, 1994
hosts in the eastern United States including Cercis canadensis L. (Leguminosae),Acer saccharinum L. (Aceraceae), Sassafras albidum (Nutt.) Nees. (Lauraceae), Ul-mus rubra Muhl. (Ulmaceae), Pinus strobus L. (Pinaceae), as well as several hosts inthe Anacardiaceae. At first glance this might appear to be a polyphagous species ex-cept that it does not normally occur in any hosts other than those mentioned aboveand their close relatives. All specimens referable to this species collected in peninsu-lar Florida have only been collected in poison ivy, despite the fact that some of theseother hosts also occur in the same areas (T.H. A., unpublished data). It may occur inthe Keys as well as on the mainland since poison ivy is found on the islands (Long &Lakela 1971. Collier: Copeland (ABSC).
Pityophthorus pulicarius (Zimmermann). (ph-my, mo, hpg). In phloem and pith ofbranches and twigs of pines (Pinaceae). Southeastern U.S. Collier: Royal Palm Ham-mock St. Park (Bright 1981); Dade: Biscayne (MCZC); Everglades Natl. Park, LongPine Key (CMNC); Homestead (Bright 1981); Paradise Key (Bright 1981); Monroe:Big Pine Key (ABSC, CMNC, Bright 1981).
Pityopthorus pecki Atkinson. (ph?, ?, hpg?). Hosts unknown. Known only from theKeys. Monroe: Big Pine Key (CMNC); No Name Key (CMNC).
ACKNOWLEDGMENTS
Travel by the first author to visit the Museum of Comparative Zoology was sup-ported by an Ernst Mayr Grant from that institution and to Provo, Utah to visit theS. L. Wood collection by a grant from the American Philosophical Society. Field workof S. Peck in southern Florida was partially supported by research grants to S. B.Peck from the Natural Sciences and Engineering Research Council of Canada. Jaram-ila Peck greatly aided in making collections under most unpleasant conditions of heat,humidity, and mosquitoes in the hardwood hammock forests. This is Florida Agricul-tural Experiment Station Journal Series No. R-03178.
REFERENCES CITED
ANDERSON, D. M. 1974. First record of Xyleborus semiopacus in the continentalUnited States (Coleoptera, Scolytidae). U.S. Dept. Agric. Coop. Econ. Ins. Rept.24(45-48): 863-864.
ARNETT, R., AND P.A. SAMUELSON. 1986. The Insect and Spider Collections of theWorld. Brill/ Flora & Fauna Publications. Gainesville.
ATKINSON, T.H. 1993. Rediscovery of 2 neotropical bark beetles (Coleoptera: Sco-lytidae) from southern Florida. Coleopterists Bull. (in press).
ATKINSON, T.H., R.J. RABAGLIA, AND D.E. BRIGHT. 1990. Newly detected exotic spe-cies of Xyleborus (Scolytidae) in eastern North America, with a revised key tospecies. Canadian Entomol. 122: 93-104.
ATKINSON, T. H., R. J. RABAGLIA, S. J. PECK, AND J. L. FOLTZ. 1991. New records ofScolytidae and Platypodidae from the U.S. and Bahamas. Coleopterists Bull.45: 152-164.
BLACKMAN, M. W. 1928. Notes on Micracinae with description of twelve new species.New York St. Coll. For., Syracuse, Tech. Pub. 25: 185-208.
BRIGHT, D.E. 1968. Review of the tribe Xyleborini in America north of Mexico (Co-leoptera: Scolytidae). Canadian Entomol. 100: 1288-1323.
BRIGHT, D.E. 1981. A taxonomic monograph of the genus Pityophthorus Eichhoff inNorth America (Coleoptera: Scolytidae). Entomol. Soc. Canada Mem. 118: 1-378.
BRIGHT, D.E. 1985. Studies on West Indian Scolytidae (Coleoptera) 3. Checklist of Sc-olytidae of the West Indies, with descriptions of new species and taxonomicnotes. Entomol. Arb. Mus. Frey 33/34: 169-187.

Atkinson & Peck:Bark and Ambrosia Beetles of South Florida329
BRIGHT, D.E. 1987. Notes on the occurrence of Xyleborinus gracilis (Eichhoff) in theUnited States (Coleoptera: Scolytidae). Coleopterists Bull. 41: 338.
CHAPIN, J. B., AND A. D. OLIVER. 1986. New records for Xylosandrus and Xyleborusspecies (Coleoptera: Scolytidae). Proc. Entomol. Soc. Washington 88: 680-683.
DEYRUP, M.A. 1987. Trischidias exigua Wood, new to the Untied States, with notes onthe biology of the genus (Coleoptera: Scolytidae). Coleopterists Bull. 41: 339-343.
DEYRUP, M.A., AND T.H. ATKINSON. 1987. New records of Scolytidae (Coleoptera) fromIndiana and Florida. Great Lakes Entomol. 20: 67-68.
DIXON, W. N., AND R. E. WOODRUFF. 1982. The black twig borer, Xylosandrus compac-tus (Eichhoff) (Coleoptera: Scolytidae). Fla. Dept. Agric. Consumer Serv., Div.Plant Indus. Entomol. Circ. No. 250, 2 p.
FRANK, J.H., AND E.D. MCCOY. 1990. Endemics and epidemics of shibboleths andother things causing chaos. Florida Entomol. 73: 1-9.
FRANK, J.H., AND E.D. MCCOY.1992. The immigration of insects to Florida, with a tab-ulation of records published since 1970. Florida Entomol. 75: 1-28.
HOPPING, G.R. 1965a. The North American species in Group IX of Ips DeGeer (Co-leoptera: Scolytidae). Canadian Entomol. 97: 422-434.
HOPPING, G.R.1965b. The North American species in Group X of Ips DeGeer (Co-leoptera: Scolytidae). Canadian Entomol. 97: 803-808.
KIRKENDALL, L.R. 1983. The evolution of mating systems in bark and ambrosia bee-tles (Coleoptera: Scolytidae and Platypodidae). Zool. J. Linnean Soc. 77: 293-352.
KOVACH, J., AND C. S. GORSUCH. 1985. Survey of ambrosia beetle species infestingSouth Carolina peach orchards and a taxonomic key for the most common spe-cies. J. Agric. Entomol. 2: 238-247.
LONG, R.W., AND O. LAKELA. 1971. A flora of tropical Florida: a manual of the seedplants and ferns of southern peninsular Florida. Univ. Miami Press, Coral Ga-bles, Fla.
NGOAN, N. D., R. C. WILKINSON, D. E. SHORT, C. S. MOSES, AND J. R. MANGOLD. 1976.Biology of an introduced ambrosia beetle, Xylosandrus compactus, in Florida.Ann. Entomol. Soc. America 69: 872-876.
PECK, S.B. 1989. A survey of insects of the Florida Keys: post-pleistocene land-bridgeislands: introduction. Florida Entomol. 72: 603-612.
STAINES, C.L. 1984. Distribution of Xylosandrus germanus (Blandford) (Coleoptera:Scolytidae) in Maryland. Proc. Entomol. Soc. Washington 86: 702.
TOMLINSON, P.B. 1980. The biology of trees native to tropical Florida. Harvard Univ.Printing Office, Allston, Mass.
WEBER, B.C., AND J. E. MACPHERSON. 1982. The distribution of Xylosandrus ger-manus in America north of Mexico (Coleoptera: Scolytidae). Great Lakes Ento-mol. 15: 171-174.
WHITEHEAD, D. R., AND A. G. WHEELER, JR. 1990. What is an immigrant arthropod.Ann. Entomol. Soc. America 83: 9-14.
WOOD, S.L. 1954. A revision of North American Cryphalini (Scolytidae: Coleoptera).Univ. Kansas Sci. Bull. 36: 959-1089.
WOOD, S.L. 1977. Introduced and exported American Scolytidae (Coleoptera). GreatBasin Natur. 37: 67-74.
WOOD, S.L. 1982. The bark and ambrosia beetles of North and Central America (Co-leoptera: Scolytidae), a taxonomic monograph. Great Basin Natur. Mem. 6: 1-1356.
WOOD, S.L., AND D.E. BRIGHT. 1992. A catalog of Scolytidae and Platypodidae (Co-leoptera), Part II: Taxonomic index. Great Basin Nat. Mem. 13: 1-1553 (2 vol-umes).

330
Florida Entomologist
77(3) September, 1994
USE OF A BIOTINYLATED DNA PROBE FOR DETECTION OF THE ASTER YELLOWS MYCOPLASMALIKE ORGANISM IN
DALBULUS
MAIDIS
AND
MACROSTELES
FASCIFRONS
(HOMOPTERA: CICADELLIDAE).
F
ERNANDO
E. V
EGA
,
1,2
R
OBERT
E. D
AVIS
,
3
E
LLEN
L. D
ALLY
,
3
P
EDRO
B
ARBOSA
1
, A
LEXANDER
H. P
URCELL
,
4
AND
I
NG
-M
ING
L
EE
3
1
Department of Entomology, University of Maryland, College Park, MD 20742.
2
Present address: Subtropical Insects Research Unit, U. S. Horticultural Research Laboratory, USDA, ARS, 2120 Camden Road, Orlando, FL 32803.
3
Molecular Plant Pathology Laboratory, USDA, ARS, Beltsville, MD 20705.
4
Department of Environmental Science, Policy and Management, Entomology Program, University of California, 201 Wellman Hall, Berkeley, CA 94720.
A
BSTRACT
A DNA probe was used to detect aster yellows mycoplasmalike organism acquisi-tion by the corn leafhopper,
Dalbulus
maidis
(DeLong & Wolcott), an insect that doesnot transmit aster yellows mycoplasmalike organism, as well as by
Macrosteles
fasci-frons
(Stål), a vector insect. Results show the effectiveness of the probe for pathogendetection in both the non-vector and the vector insect.
R
ESUMEN
Se utilizó una prueba de DNA para detectar la adquisición del organismo de mico-plasma de las estrias amarillas (AYMLO) por la chicharrita del maíz,
Dalbulus mai-dis
(DeLong & Wolcott), un insecto que no transmite AYMLO, así como de
Macrostelesfascifrons
(Stal) un insecto vector. Los resultados muestran la efectividad de la prueba
para detectar el patógeno en el insecto no vector y en el insecto vector.
Positive and negative effects have been reported in vector and non-vector insectsafter feeding on diseased plants (Vega 1992). In at least one case, an expansion of hostplant range occurred when a non-vector insect species fed upon a diseased host (Ma-ramorosch 1958). This system involved the aster yellows mycoplasmalike organism- an aster leafhopper [
Macrosteles
fascifrons
(Stål); Homoptera: Cicadellidae] (So-rensen & Sawyer 1989) transmitted a plant pathogen with host plants in over 40 dif-ferent families (McCoy et al. 1989) - and the corn leafhopper,
Dalbulus
maidis
(DeLong & Wolcott), (Homoptera: Cicadellidae).
D.
maidis
is a
Zea
spp. specialist (Nault & DeLong 1980) that cannot normally sur-vive on China asters [
Callistephus
chinensis
(L.) Nees] (Maramorosch 1958) nor doesit transmit the pathogen causing aster yellows (Maramorosch 1952). However, if
D.maidis
feeds on asters infected with the aster yellows mycoplasmalike organism, notonly can it survive on the plant, but after a minimum of three days on the plant it cansurvive on healthy asters and other non-host plants, such as carrots and rye, on which
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Vega et al.: Use of a Biotinylated DNA probe
331
it typically would not survive (Maramorosch 1960; Purcell 1988). This phenomenon iscalled conditioning (Maramorosch 1958).
Recently, Vega et al. (1993) confirmed the presence of the aster yellows mycoplas-malike organism in
D.
maidis
by using the polymerase chain reaction (PCR). The ob-jective of this study was to determine whether a biotinylated DNA probe could be usedto detect the mycoplasmalike organism in a vector and a non-vector insect.
M
ATERIALS
AND
M
ETHODS
Healthy and aster yellow-infected aster plants, as well as healthy and infected
M.fascifrons
, were provided by the fifth author. The non-vector insects (i.e.,
Dalbulusmaidis
) had been maintained in colonies at the University of Maryland greenhousefor 15-20 generations.
Pathogen Acquisition
Vector and non-vector insects were placed on asters infected with the ‘Severe’(SAY) strain of the aster yellows mycoplasmalike organism for various acquisition ac-cess periods (AAP). The AAP was followed by various incubation periods, duringwhich insects were held on uninfected asters in the case of
M.
fascifrons
, or on unin-fected corn (cv. Aristogold Bantam Evergreen) in the case of
D.
maidis
. When
D.
mai-dis
was allowed to feed on healthy aster plants, the insect died within 4 days(Maramorosch 1958). To reduce mortality of the non-vector insect after the AAP onSAY-infected asters, we held
D. maidis
on corn for the incubation period.
D.
maidis
was given an AAP of 7 days followed by incubation periods of 0, 7 or 14 days and anAAP of 14 days followed by incubation periods of 0 or 7 days.
M.
fascifrons
was givenan AAP of 7 days followed by incubation periods of 0, 7, or 14 days and an AAP of 14days followed by an incubation period of 0, 7, or 14 days. Negative controls consistedof insects that fed on corn (
D.
maidis
) or on aster (
M.
fascifrons
).
M.
fascifrons
AAPand incubation periods were conducted at the University of California, Berkeley.
D.maidis
AAP and incubation periods were conducted at the University of Marylandgreenhouse. Insects were frozen (-86
°
C) after each treatment combination until DNAanalysis.
DNA Extraction
Batches of five insects were separately placed in 12 ml glass tissue homogenizers(Bellco Biotechnology, Vineland, N.J.) and macerated several times in 400
µ
l of extrac-tion buffer (100 mM Tris, 50 mM EDTA, 500 mM NaCl, pH 8.0), 2
µ
l of ß-mercaptoeth-anol, and 20
µ
l of 20% sodium dodecyl sulfate (SDS). The solution was transferred toa 1.5 ml Eppendorf tube and re-macerated with a mini-pestle. After clarifying threetimes by centrifugation, nucleic acids were extracted using chloroform-isoamyl alco-hol and TE-saturated phenol. To precipitate nucleic acids, 2.5 volumes of cold absoluteethanol was added to samples followed by 30 min at -86
°
C, and 20 min of centrifuga-tion at 14,000 rpm. The DNA pellet was resuspended in 100
µ
l 6x saline sodium cit-rate (SSC; 1x= 0.9 M NaCl, 0.09 M sodium citrate, pH 7.0).
Filter Preparation
Resuspended DNA was denatured by adding to each 50
µ
l SSC, 3
µ
l of 2N NaOH,boiling for 10 min, cooling in ice and neutralizing with 3
µ
l of 2M TRIS and 1
µ
l 1.5

332
Florida Entomologist
77(3) September, 1994
M NaAc. Three microliters of undiluted and serial dilutions were spotted on nitrocel-lulose paper and baked at 80
°
C for 2 h under vacuum.
Filter Pre-hybridization
Filters were pre-hybridized with gentle rotation at 42
°
C for 2-4 h in presence of50% formamide, 5x SSC, 5X Denhardt’s solution (0.5 g Ficoll, 0.5 g polyvinylpyrroli-done, 0.5 g bovine serum albumin), 25 mM sodium phosphate and 0.5 mg/ml of dena-tured salmon sperm DNA.
Probe Hybridization and Visualization
Hybridization was conducted at 42
°
C using 5% Dextran sulfate, 45% formamide,5X SSC, 1X Denhardt’s solution, 20 mM sodium phosphate (pH 6.5), 0.2 mg
µ
l of de-natured salmon sperm DNA, and approximately 0.3 mg of biotinylated DNA probeper ml. The probe was prepared by cloning aster yellows mycoplasmalike organismDNA fragments into plasmid vectors pSP64 or pSP65 which were then amplified in
Escherichia
coli
strain JM83 (for a full description see Lee & Davis 1988). After 12-16h incubation, filters were rinsed twice for 3 min in 2X SSC with 0.1% SDS, 2 rinses of0.2X SSC with 0.1% SDS (both rinses at room temperature), and 2 rinses at 50
°
C for15 min using 0.16X SSC with 0.1% SDS. For signal detection, filters were rinsed for1 min in buffer 1 (0.1 M Tris-HCl, 0.15 M NaCl, pH 7.5), and blocked for 1 h at 64
°
Cin buffer 2 (3% bovine serum albumin in buffer 1). Filters were incubated with gentlerotation for 10 min using streptavidin-alkaline phosphatase diluted in buffer 1, fol-lowed by two 15 min rinses in buffer 1 and one 10 min rinse in buffer 3 (0.1 M Tris-HCl, 0.1 M NaCl, 0.05 M MgCl
2
, pH 9.5). To visualize reactions, filters were incubatedfor about 30 min in the dark using nitroblue tetrazolium and 5-bromo-4-chloro-in-dolylphosphate diluted in buffer 3. To stop reactions, filters were washed in termina-tion buffer (20 mM Tris, 0.5 mM EDTA, pH 7.5). A bluish color indicated a positivereaction, i.e., presence of the pathogen’s DNA.
R
ESULTS
AND
D
ISCUSSION
For
D.
maidis given a 7-day AAP, higher amounts of DNA were detected as the in-cubation period increased from 0 to 7, and then to 14 day (Table 1). There was a re-duction in the SAY DNA detected as the incubation period increased from 0 to 7 daysin insects given a 14-day AAP (Table 1). Similar reductions in aster yellows concen-tration have been reported in the alimentary canal of M. fascifrons by Sinha &Chiykowski (1967). SAY DNA was detected in M. fascifrons at all dilutions and in alltreatments except for the control and for insects given a 7 day AAP and no incubationperiod (Table 1).
Our results (Table 1) indicate that a biotinylated DNA probe can be used for thedetection of SAY MLO in vector and non-vector insects. Even though sensitive tech-niques such as PCR are now routinely used for pathogen detection, biotinylated DNAprobes are a sound alternative in cases where PCR technology is not available. In thisstudy, the use of a biotinylated DNA probe yielded information that suggests an in-crease in pathogen titer as the incubation period increased for non-vector insects witha 7 days AAP.
If widespread, the conditioning phenomenon could play an important role in theexpansion of insects host plant range. The availability of different pathogen detection
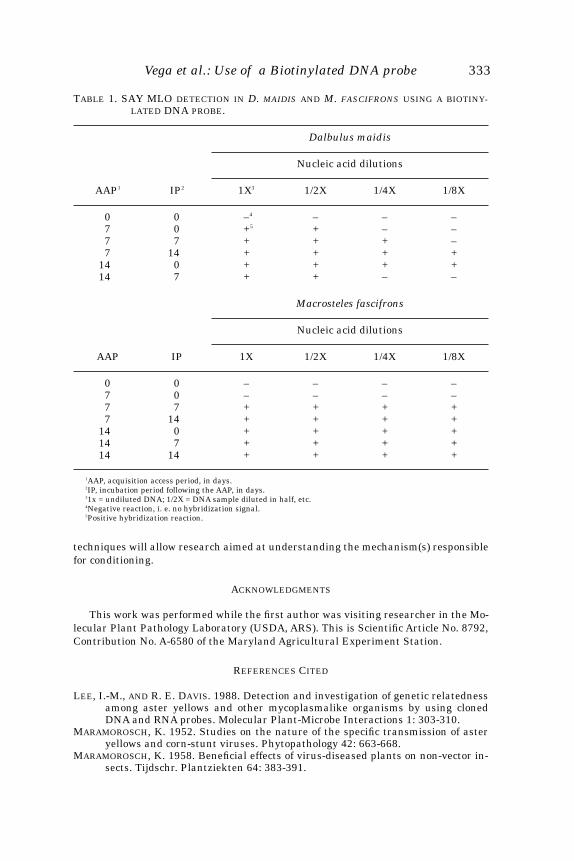
Vega et al.: Use of a Biotinylated DNA probe 333
techniques will allow research aimed at understanding the mechanism(s) responsiblefor conditioning.
ACKNOWLEDGMENTS
This work was performed while the first author was visiting researcher in the Mo-lecular Plant Pathology Laboratory (USDA, ARS). This is Scientific Article No. 8792,Contribution No. A-6580 of the Maryland Agricultural Experiment Station.
REFERENCES CITED
LEE, I.-M., AND R. E. DAVIS. 1988. Detection and investigation of genetic relatednessamong aster yellows and other mycoplasmalike organisms by using clonedDNA and RNA probes. Molecular Plant-Microbe Interactions 1: 303-310.
MARAMOROSCH, K. 1952. Studies on the nature of the specific transmission of asteryellows and corn-stunt viruses. Phytopathology 42: 663-668.
MARAMOROSCH, K. 1958. Beneficial effects of virus-diseased plants on non-vector in-sects. Tijdschr. Plantziekten 64: 383-391.
1AAP, acquisition access period, in days.2IP, incubation period following the AAP, in days.31x = undiluted DNA; 1/2X = DNA sample diluted in half, etc.4Negative reaction, i. e. no hybridization signal.5Positive hybridization reaction.
TABLE 1. SAY MLO DETECTION IN D. MAIDIS AND M. FASCIFRONS USING A BIOTINY-LATED DNA PROBE.
Dalbulus maidis
Nucleic acid dilutions
AAP1 IP2 1X3 1/2X 1/4X 1/8X
0 0 –4 – – –7 0 +5 + – –7 7 + + + –7 14 + + + +
14 0 + + + +14 7 + + – –
Macrosteles fascifrons
Nucleic acid dilutions
AAP IP 1X 1/2X 1/4X 1/8X
0 0 – – – –7 0 – – – –7 7 + + + +7 14 + + + +
14 0 + + + +14 7 + + + +14 14 + + + +

334 Florida Entomologist 77(3) September, 1994
MARAMOROSCH, K. 1960. Friendly viruses. Sci. American 203: 138-144.MCCOY, R. E., A. CAUDWELL, C. J. CHANG, T. A. CHEN, L. N. CHIYKOWSKI, M. T.
COUSIN, J. L. DALE, G. T. N. DE LEEUW, D. A. GOLINO, K. J. HACKETT, B. C.KIRKPATRICK, R. MARWITZ, H. PETZOLD, R. C. SINHA, M. SUGIURA, R. F. WHIT-COMB, I. L. YANG, B. M. ZHU, AND E. SEEMULLER. 1989. Plant diseases associ-ated with mycoplasmalike organisms, pp. 545-640 in R. F. Whitcomb & J.G.Tully [eds.], The Mycoplasmas, Vol. V, Academic Press, New York.
NAULT, L. R., AND D. M. DELONG. 1980. Evidence for co-evolution of leafhoppers inthe genus Dalbulus (Cicadellidae: Homoptera) with maize and its ancestors.Ann. Entomol. Soc. America 73: 349-353.
PURCELL, A. H. 1988. Increased survival of Dalbulus maidis, a specialist on maize, onnon-host plants infected with mollicute plant pathogens. Entomol. Exp. Appl.46: 187-196.
SINHA, R. C., AND L. N. CHIYKOWSKI. 1967. Initial and subsequent sites of aster yel-lows virus infection in a leafhopper vector. Virology 33: 702-708.
SORENSEN, J. T., AND S. M. SAWYER. 1989. Assessing the multivariate evolutionary re-sponses of phenological differentiation for sibling species: biosystematics in theMacrosteles fascifrons complex (Homoptera: Cicadellidae). Ann. Entomol. Soc.America 82: 250-261.
VEGA, F. E. 1992. The role of plant disease on the biology and ecology of vector andnon-vector insect herbivores. Ph.D. dissertation, Univ. of Maryland, CollegePark.
VEGA, F. E., R. E. DAVIS, E. DALLY, P. BARBOSA, A. H. PURCELL, AND I.-M. LEE. 1993.Detection of a plant pathogen in a nonvector insect by the polymerase chain re-
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
action. Phytopathology 83:621-624.

334
Florida Entomologist
77(3) September, 1994
LEPIDOPTEROUS PESTS OF COTTON AND THEIR PARASITOIDS
IN A DOUBLE-CROPPING ENVIRONMENT
F. C. T
INGLE
, E. R. M
ITCHELL
,
AND
J. R. M
C
L
AUGHLIN
Insect Attractants, Behavior, and Basic BiologyResearch Laboratory, Agricultural Research Service
U.S. Department of Agriculture, Gainesville, FL 32604
A
BSTRACT
Seasonal populations of
Spodoptera frugiperda, S. exigua, S. eridania
,
Heliothisvirescens
,
Helicoverpa zea
,
Pseudoplusia includens
, and their parasitoids, were mon-itored on late-season cotton in northcentral Florida in 1992. The cotton was plantedas a second crop, following corn, in one field and as a first crop in a second field inwhich cotton has not been grown for many years. At least twelve species of parasitoidsemerged from lepidopterous larvae collected from the cotton plants. The most com-mon parasitoids were
Cotesia marginiventris
,
Meteorus autographae
,
Cardiochiles
ni-griceps
,
Netelia
sayi
, and
Copidosoma truncatellum
. The native parasitoid thatshowed the most potential as a biological control agent was
C
.
marginiventris
, be-cause it attacks a broad range of pests including the
Heliothis
/
Helicoverpa
complex,
Spodoptera
spp., and
Pseudoplusia includens
.
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Tingle et al.: Cotton Pests and Parasitoids in Florida
335
Key Words: Cotton,
Heliothis
/
Helicoverpa
,
Spodoptera
, parasitoids
R
ESUMEN
Las poblaciones estacionales de
Spodoptera frugipereda
,
S. exigua
,
S. eridania
,
Heliothis virescens
,
Helicoverpa zea
,
Pseudoplusia includens
, y sus parasitoides, fu-eron muestreadas periodicamente en algodón tardío en la region noroccidental deFlorida en 1992. El algodón fue plantado en un campo como segundo cultivo, despuesde maíz, y en otra area donde no se había sembrado algodón durante muchos años. Almenos doce especies de parasitoides emergieron de las larvas de lepidopteros colecta-das de las plantas de algodón. Los parasitoides mas comunes fueron
Cotesia margin-iventris
,
Meteorus autographae
,
Cardiochiles nigriceps
,
Netelia sayi
, y
Copidosomatrucatellum
. El parasitoide nativo que mostró el mayor potencial como agente de con-trol biologico fué
C. marginiventris
, porque ataca un amplio rango de plagas, incluy-
endo el complejo
Heliothis
/
Spodoptera
,
Spodoptera
spp., y
Pseudoplusia includens
.
Cotton had not been grown commercially in northcentral Florida for over 50 yearsuntil a 60-ha field was planted following an early corn crop in Gilchrist County in1991. This crop was successful and more acreage was planted in 1992. Because cottonhad not been planted in this area for many years and particularly because it wasplanted as a second crop in a double-cropping environment, we chose to monitor pop-ulations of cotton pests and their parasitoids in cotton in 1992. Information on theparasitoids was needed to define the natural populations and to determine their po-tential value in integrated pest management programs.
M
ATERIALS
AND
M
ETHODS
The cooperating grower planted cotton in a 18-ha dryland field on June 6 as thefirst crop of the year and in a nearby 152-ha irrigated field July 11-18 following anearly corn crop that was harvested for silage. Two cotton varieties, Delta Pine 90 andHS 46, were planted in each of the two fields, which were separated by a highway andtree line. Information on insect pest infestations and damage levels was provided tothe grower who applied conventional insecticide treatments at his discretion. Approx-imately 52% of the insecticide applications involved synthetic pyrethroids. Other pes-ticide groups applied included carbamates, chlorinated bicyclic sulfites, organo-phosphates, and
Bacillus thuringiensis
. The dryland cotton received 12 applicationsof pesticide, and 10 applications were made in the irrigated field. Although some irri-gation was used, it was not a major factor that influenced the outcome of the study be-cause rainfall was sufficient throughout the growing season.
Cotton plants (stems, leaves, squares, blooms, and bolls) were checked for damageand lepidopterous eggs and larvae were counted twice weekly in each of the 2 variet-ies at 6 preselected locations in the dryland field and at 12 locations in the irrigatedfield (25 randomly selected plants per location). Different plants were checked on eachsample date. The eggs and larvae were collected by hand, returned to the laboratory,and reared individually in 33-ml plastic cups on artificial diet (Guy et al. 1985) foremergence of adults or adult parasitoids. Larval and egg collections were made fromJune 30 through October 16 in the dryland cotton. The larval and egg collections inthe irrigated field began on July 21 and continued through November 20. Parasitiza-tion levels were determined throughout the season based on the emergence of moths

336
Florida Entomologist
77(3) September, 1994
or parasitoids from collected eggs and larvae. Identification of pest and parasitoid spe-cies was verified by comparison with preserved specimens.
R
ESULTS
AND
D
ISCUSSION
The insect surveys and the emergence of adults from eggs and larvae collectedfrom the late-season cotton enabled us to monitor field populations (Fig. 1-3) of the fol-lowing pest species: fall armyworm (FAW),
Spodoptera frugiperda
(J.E. Smith); beetarmyworm (BAW),
S.
exigua
(Hübner); southern armyworm (SAW),
S.
eridania
(Cramer); tobacco budworm (TBW),
Heliothis
virescens
(F.); corn earworm (CEW),
Helicoverpa
zea
(Boddie); and soybean looper (SBL),
Pseudoplusia
includens
(Walker).Prior to September 1, the predominant pest species in the dryland cotton, as indi-
cated by the number of adults that emerged from collected larvae and eggs, was FAW(Figures 2A and 3A, respectively). Parasitoids could have been a major factor in pre-venting the emergence of adults from the TBW and CEW larvae found on the cottonplants (Figure 1B), because 50 to 100% of the field-collected TBW and CEW larvaewere parasitized during August. Also during August, 65 to 75% of SBL larvae wereparasitized (Fig. 1C), whereas less than 40% of the FAW and BAW larvae were para-sitized (Figure 1A). Parasitism of FAW and BAW larvae increased rapidly, reaching100% during the first week of September. At this time SBL became the most commonpest (Figures 1C and 2B) until the cotton matured. However, very few SBL eggs werefound on the cotton plants in either field, and no parasitoids emerged from any of thefield-collected lepidopteran eggs. Predation of larvae and eggs by beneficial insectswas not assessed.
The predominant pest species in the later-planted cotton in the irrigated field wasTBW, as indicated by adult emergence from larval and egg collections (Fig. 2D and3D), until outnumbered by FAW in late August (Fig. 2C and 3C). Subsequently, BAWlarvae predominated in September. As in the dryland cotton, parasitoids apparentlyprevented a substantial number of TBW and CEW larvae from reaching the adultstage. Parasitization of TBW and CEW larvae peaked at 80% during the first week ofSeptember and again at almost 70% during the first week of October (Fig. 1E). Para-sitization of FAW and BAW larvae did not exceed 50% throughout the season (Fig.1D).
SBL larval populations in the irrigated cotton increased in mid-September andpeaked two weeks later (Fig. 1F and 2D). Parasitism levels of the SBL larvae rangedfrom 20 to 50% during September and October. More FAW than BAW eggs were col-lected from cotton plants during October (Fig. 3C), but by late October more BAWadults emerged from the field-collected larvae than any other pest species (Fig. 2C).
No SAW larvae were found in either field, but some SAW egg masses were collectedfrom plants in the irrigated field in late October (Fig. 3C). Differences between the lar-val and egg collection data likely result from protection of the eggs and small larvaein the laboratory from insecticide applications, predators, parasitoids, heavy rains,and other environmental hazards. Most of the field-collected larvae of each pest spe-cies were half-grown or smaller.
At least 12 species of parasitoids emerged from the lepidopterous larvae collectedfrom the cotton plants. These included the following Hymenoptera:
Cardiochiles
nig-riceps
(Viereck) [Braconidae],
Chelonus
insularis
Cresson [Braconidae],
Copidosomatruncatellum
(Dalman) [Encyrtidae],
Cotesia marginiventris
(Cresson) [Braconidae],
Meteorus autographae
Muesebeck [Braconidae],
Netelia
sayi
(Cushman) [Ichneu-monidae],
Ophion
sp. [Ichneumonidae], and
Pristomerus
spinator
(F.) [Ichneu-
Tingle et al.: Cotton Pests and Parasitoids in Florida
337
monidae]. Four species of Diptera, all Tachinidae, were identified:
Archytasmarmoratus
(Townsend),
Eucelatoria
rubentis
(Coquillett),
Lespesia
archippivora
(Ri-ley), and
Winthemia rufopicta
(Bigot).
Fig. 1. Number of Spodoptera (FAW and BAW), Heliothis/Helicoverpa (TBW andCEW), and P. includens (SBL) counted per 100 cotton plants and percentage parasit-ism of field-collected larvae in each field.
338
Florida Entomologist
77(3) September, 1994
Average parasitism of FAW and BAW combined was 23% in each field although theaverage numbers of larvae counted per week were almost 5 times greater in the irri-gated field than in the dryland cotton. The number of FAW and BAW larvae per 100plants averaged 0.7
±
0.3 (June 30-October 16) in the dryland cotton and 3.4
±
0.9(July 21-November 20) in the irrigated cotton that followed an early corn crop. Mostof the parasitoids that emerged from the FAW and BAW larvae were C. marginiven-tris. This species made up 66% and 83%, respectively, of the parasitoids that emergedfrom larvae collected in the dryland and irrigated fields. The second most commonparasitoid in each field was M. autographae (10 and 15% of the parasitism in the dry-land and irrigated fields, respectively).
Parasitism of the TBW and CEW larvae combined averaged 65% per weekthroughout the season in the dryland cotton but only 29% in the later-planted irri-gated cotton. Also, the TBW and CEW larval population was almost 4 times greaterin the dryland cotton [5.9(x) ± 2.9(SEM) larvae per 100 plants] than in the irrigatedfield (1.5 ± 0.5 larvae per 100 plants).
Fig. 2. Percent of each pest species as determined by emergence of adults from lar-vae collected from cotton plants in each field (FAW = fall armyworm; BAW = beet ar-myworm; TBW = tobacco budworm; CEW = corn earworm; SBL = soybean looper).
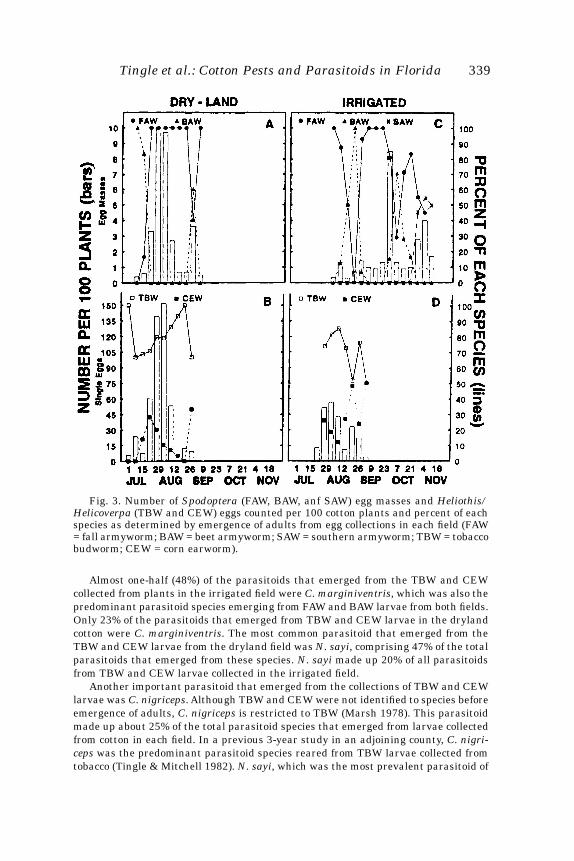
Tingle et al.: Cotton Pests and Parasitoids in Florida 339
Almost one-half (48%) of the parasitoids that emerged from the TBW and CEWcollected from plants in the irrigated field were C. marginiventris, which was also thepredominant parasitoid species emerging from FAW and BAW larvae from both fields.Only 23% of the parasitoids that emerged from TBW and CEW larvae in the drylandcotton were C. marginiventris. The most common parasitoid that emerged from theTBW and CEW larvae from the dryland field was N. sayi, comprising 47% of the totalparasitoids that emerged from these species. N. sayi made up 20% of all parasitoidsfrom TBW and CEW larvae collected in the irrigated field.
Another important parasitoid that emerged from the collections of TBW and CEWlarvae was C. nigriceps. Although TBW and CEW were not identified to species beforeemergence of adults, C. nigriceps is restricted to TBW (Marsh 1978). This parasitoidmade up about 25% of the total parasitoid species that emerged from larvae collectedfrom cotton in each field. In a previous 3-year study in an adjoining county, C. nigri-ceps was the predominant parasitoid species reared from TBW larvae collected fromtobacco (Tingle & Mitchell 1982). N. sayi, which was the most prevalent parasitoid of
Fig. 3. Number of Spodoptera (FAW, BAW, anf SAW) egg masses and Heliothis/Helicoverpa (TBW and CEW) eggs counted per 100 cotton plants and percent of eachspecies as determined by emergence of adults from egg collections in each field (FAW= fall armyworm; BAW = beet armyworm; SAW = southern armyworm; TBW = tobaccobudworm; CEW = corn earworm).

340 Florida Entomologist 77(3) September, 1994
TBW in postharvest tobacco (Tingle & Mitchell 1982), also emerged from TBW larvaecollected from cotton in the present study.
The weekly average percentage parasitism of SBL larvae collected from cottonduring the season was 49% in the dryland field and 39% in the irrigated field. The av-erage number of SBL larvae counted per 100 plants per week was [9.9(x) ± 4.6(SEM)]in the dry-land cotton and 4.2 ± 1.0 in the irrigated field. Although C. marginiventrisemerged from 58% of the parasitized SBL larvae collected in the irrigated field, 92%of parasitized SBL in the earlier planted cotton contained C. truncatellum. This spe-cies also emerged from 32% of the parasitized SBL larvae collected from cotton in theirrigated field.
C. truncatellum, a polyembryonic egg-larval parasitoid of several insect pests, hasa high reproductive potential and has been reported as the most prevalent parasitoidof SBL (Burleigh 1971). However, this parasitoid may not be a promising candidate inan integrated control program for SBL because Hunter & Stoner (1975) found thatSBL larvae parasitized by this species consumed 35% more food than unparasitizedlarvae. The increased food consumption of parasitized larvae could outweigh benefitsresulting from reduced SBL populations.
C. marginiventris, however, has potential as a biological control agent because itattacks a broad range of lepidopterous pests such as the Heliothis / Helicoverpa group,Spodoptera spp., and Pseudoplusia includens. This parasitoid frequently causes highmortality among these pests (e.g., Tingle et al. 1978; McCutcheon & Turnipseed 1981,Pair et al. 1982, 1986). Ashley (1979) reported that C. marginiventris is one of themost frequently recovered parasitoids from FAW larval collections. It is an importantnatural enemy of FAW because it parasitizes first- and second-instar larvae that diewhen they reach the fourth instar (Ashley et al. 1982).
Growers in northcentral Florida are experimenting with various crops and produc-tion schemes in an attempt to increase profitability on existing acreage. The conceptof producing a cash crop such as cotton following a crop of silage corn has received at-tention and appears promising. Of particular concern to growers pursuing this prac-tice has been the mix of insect pests encountered in cotton planted so late in thegrowing season, and also how the pest complex might be exacerbated by the early (i.e.,first) crop of corn.
This study indicates that the species of lepidopterous pests in dryland (early sea-son) and irrigated (late season, 2nd crop) cotton were virtually the same as was theparasitoid complex associated with these pests. Although several species of parasi-toids were recorded, C. marginiventris was the most prevalent and had the largest im-pact because of its propensity to attack a broad range of pests including the Heliothis/Helicoverpa complex, Spodoptera spp., and Pseudoplusia includens.
The data presented here suggest that parasitoids in general, and C. marginiven-tris in particular, could play a major role in control of most lepidopterous pests on cot-ton. Pest control strategies should foster the preservation of these and other naturalenemies via the selection and judicious use of pesticides that have minimal impact onthe natural enemies complex and, where possible, provide suitable habitats in andaround cotton fields to encourage parasitoid development and survival.
ACKNOWLEDGMENTS
We gratefully acknowledge the technical assistance of W. Copeland, J. Leach, J.Rye, R. Navasero-Ward, F. Adams, C. Green, and R. Hemphill of this laboratory. Also,we thank J.R. Ruberson (Dept. Entomol., Coastal Exp. Stn., Tifton, GA) and person-nel of the Division of plant Industry (DPI), Fla. Dept. of Agric., Gainesville, FL, for

Tingle et al.: Cotton Pests and Parasitoids in Florida 341
providing preserved specimens to aid in identification of parasitoids. Mention of a pro-prietary product does not constitute an endorsement or recommendation for its use byUSDA.
REFERENCES CITED
ASHLEY, T. R. 1979. Classification and distribution of fall armyworm parasites. Flor-ida Entomol. 62: 114-123.
ASHLEY, T. R., V. H. WADDILL, E. R. MITCHELL, AND J. RYE, 1982. Impact of nativeparasites on the fall armyworm, Spodoptera frugiperida (Lepidoptera: Noctu-idae), in south Florida and the release of the exotic parasite, Eiphosoma vitti-cole (Hymenoptera: Ichneumonidae). Environ. Entomol. 11:833-837.
BURLEIGH, J. B. 1971. Parasites reared from the soybean looper in Louisiana 1968-9.J. Econ. Entomol. 64: 1550-1551.
GUY, R. H., N. C. LEPPLA, J. R. RYE, C. W. GREEN, S. L. BARRETTE, AND K. A. HOLLIEN.1985. Trichoplusia ni, pp. 487-494, in P. Singh and R. F. Moore [eds.] Handbookof Insect Rearing, Vol. 2. Elsevier Science Publishers B.V., Amsterdam.
HUNTER, K. W., JR., AND A. STONER. 1975. Copidosoma truncatellum: Effect of para-sitization on food consumption of larval Trichoplusia ni. Environ. Entomol. 4:381-382.
MARSH, P. M. 1978. The Braconid parasites (Hymenoptera) of Heliothis species (Lep-idoptera: Noctuidae). Proc. Entomol. Soc. Washington 80: 15-36.
MCCUTCHEON, G. S., AND S. G. TURNIPSEED. 1981. Parasites of lepidopterous larvaein insect resistant and susceptible soybeans in South Carolina. Environ. Ento-mol. 10: 69-74.
PAIR, S. D., M. L. LASTER, AND D. F. MARTIN. 1982. Parasitoids of Heliothis spp. (Lep-idoptera: Noctuidae) larvae in Mississippi associated with sesame interplant-ings in cotton, 1971-1974: Implications of host habitat interaction. Environ.Entomol. 11: 509-512.
PAIR, S. D., J. R. RAULSTON, A. N. SPARKS, AND P. B. MARTIN. 1986. Fall armyworm(Lepidoptera: Noctuidae) parasitoids: Differential spring distribution and inci-dence on corn and sorghum in the Southern United States and NortheasternMexico. Environ. Entomol. 15: 342-348.
TINGLE, F. C., T. R. ASHLEY, AND E. R. MITCHELL. 1978. Parasites of Spodoptera ex-igua, S. eridania (Lepidoptera: Noctuidae) and Herpetogramma bipunctalis(Lepidoptera:Pyralidae) collected from Amaranthus hybridus in field corn. En-tomophaga 23: 343-347.
TINGLE, F. C., AND E. R. MITCHELL. 1982. Effect of synthetic pheromone on parasiti-zation of Heliothis virescens (F.) (Lepidoptera: Noctuidae) in tobacco. Environ.Entomol. 11: 913-916.

342
Florida Entomologist
77(3) September, 1994
EGG MORPHOLOGY OF
ANASTREPHA OBLIQUA
AND SOME COMPARATIVE ASPECTS WITH EGGS OF
ANASTREPHA FRATERCULUS
(DIPTERA:TEPHRITIDAE)
T
ERESITA
M
URILLO
1
AND
L
UIS
F
ERNANDO
J
IRÓN
2
.
1
Laboratorio de Control de Calidad, Corporación PIPASA, San Antonio de Belén, Heredia, Costa Rica.
2
Museo de Insectos, Facultad de Agronomía,Universidad de Costa Rica, San José, Costa Rica.
A
BSTRACT
The egg morphology of
Anastrepha obliqua
(Macquart) is described utilizing bothlight and scanning electron microscopy. Other observations include oviposition depthby
A. obliqua
on mango and comparative analysis with
A. fraterculus
(Wiedemann)egg morphology.
A. obliqua
eggs bear a conspicuous structure at the anterior pole,which is called a respiratory horn. A discussion of this term and definitions given byother authors for similar structures found in other fruit fly eggs is given. We suggestthat the occurrence of the respiratory horn outside the mango fruit peel by
A. obliqua
may be a useful character for post-harvest inspection and mango fruit disinfestation.
Key Words: Mango, fruit flies, insect egg anatomy, insect respiration
R
ESUMEN
La morfología de
Anastrepha obliqua
(Macquart) es descrita por medio de micros-copia de luz y electrónica de barrido. Otras observaciones incluyen la profundidad deovoposición por
A. obliqua
en mangos y el analisis comparativo con la morfología delhuevo de
A. fraterculus
(Wiedemann). Los huevos de A. obliqua poseen una notable es-tructura en el polo anterior, que es llamada cuerno respiratorio. Se ofrece una discu-sion acerca de este termino y de definiciones dadas por otros autores para estructurassimilares en otros huevos de moscas fruteras. Sugerimos que la aparición del cuernorespiratorio fuera de la cáscara del mango en
A. obliqua
puede ser una caracteristica
util para la inspección post cosecha y la desinfección de los frutos.
The taxonomy of
Anastrepha
fruit flies is based primarily on adult forms; however,egg morphology also has taxonomic value, and more descriptive work needs to be done(Norrbom & Foote 1989). Emmart (1933) published descriptions of the eggs of
A.ludens
(Loew),
A. obliqua
(Macquart) (confused with
A. fraterculus
(Wiedemann)),
A. striata
Schiner and
A. serpentina
(Wiedemann). Sein (1933) reported that a portionof the egg of
A. obliqua
remains outside of the fruit peel after oviposition. Later,Lawrence (1979) described the eggs of
A. suspensa
(Loew). More recently, Norrbom(1985) described the general characters of the eggs of eight
Anastrepha
species, in-cluding
A. obliqua
. Additional descriptions of the eggs of
A. bistrigata
Bezzi (Steck &Malavasi 1988),
A. grandis
(Macquart) (Steck & Wharton 1988) and
A. ludens
(Car-roll & Wharton 1989) have appeared since then.
Based on the above mentioned descriptions, there is some confusion concerningthe morphology of
A. obliqua
eggs. Emmart (1933) described a twisted anterior end of
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Murillo & Jirón:
Anastrepha obliqua
Egg Morphology
343
the egg, hairs around the micropyle and on the posterior end, and a sculptured surfaceon the anterior (micropylar) end. Sein (1933) described a smooth surface over the en-tire egg. Norrbom (1985) described
A. obliqua
eggs as having an anterior lobe coveredby pore-like openings and a reticulated surface at the base of this lobe. The latter au-thor made these observations on ova dissected from preserved females and expressedsome reservations about the appearance of the egg after oviposition.
Here, we describe the morphology of the eggs of
A. obliqua
with light and scanningelectron microscopy (SEM). The position of the egg after oviposition into mango fruitis also described with SEM. We also make a comparative description with
A. fratercu-lus
egg morphology.
M
ATERIALS
AND
M
ETHODS
Adults of
A. obliqua
were reared at the laboratories of Organismo InternacionalRegional de Sanidad Agropecuaria (OIRSA) in Guadalupe (San José Prov., elev. 1200m.) obtained from field infested mangoes collected at Orotina (Alajuela Prov., elev. 200m.). About 300 of these adults (both sexes) were held in a 1 m
3
screened cage and al-lowed to oviposit into an artificial medium consisting of a 10 cm diam agar hemi-sphere coated with a beeswax-paraffin mixture. Thirty eggs were retrieved and placedin glycerin on depression slides and measured under a compound light microscope.General features were observed with light microscopy by mounting another thirtyeggs in Hoyers temporary mounting medium. Adults of
A. fraterculus
were obtainedfrom a laboratory colony (generation F23), and eggs were obtained by placing a pieceof black cloth externally on the upper wall of the colony cage for oviposition. Theywere treated as described above.
Eggs of both
Anastrepha
species were fixed for SEM in 2.5% glutaraldehyde and2.0% paraformaldehyde, in phosphate buffer (0.1 M, pH 7.4), post-fixed in 1% osmiumtetraoxide, dehydrated in graded alcohol, and critically point dried. Stub mountedspecimens were examined with a scanning electron microscope (Hitachi S-570). Prep-aration for cryofracture was carried out, as an intermediate step after alcoholic dehy-dration, using a substitution graded mixture up to 1:1 alcohol: DMSO (dimethylsulfoxide). An aluminum surface frozen by liquid nitrogen was the support for thesample, which was cryofractured by a stroke with a razor blade. After the SEM prep-aration process, we observed some problems keeping the egg in turgid form; however,the anterior pole maintained its shape, thus facilitating its observation.
Five mature mango fruits of Yellow, Mora and Tommy Atkins varieties were ex-posed to oviposition by about 300
A. obliqua
adults (both sexes) in a 1 m
3
screenedcage for twelve hours. Thirty 1 cm
3
pieces of fruit rind, each containing one egg, weretaken from each variety for microscopic examination and measurement of the exposedportion of the egg with a dissecting microscope. Oviposition depth into mango fruit by
A. obliqua
was calculated as the difference between the average egg length and thelength of each of the exposed respiratory horns. Ten pieces of each mango variety, eachcontaining one egg, were similarly treated for SEM as described above to examine theexposed egg horn after oviposition.
R
ESULTS
AND
D
ISCUSSION
Anastrepha obliqua
and
A. fraterculus
are closely related species belonging to the
fraterculus
subgroup (Norrbom 1985); however, their egg morphologies are clearly dif-ferent.

344
Florida Entomologist
77(3) September, 1994
Anastrepha obliqua
eggs observed by light microscopy are creamy white with theanterior end drawn into a respiratory horn and the posterior end bluntly rounded(Fig. 1). Average measurements of thirty
A. obliqua
eggs were: total length 1.447 mm(1.181-1.584 mm), greatest width 0.225 mm (0.202-0.259 mm) and respiratory hornlength 0.142 mm (0.086-0.245 mm).
The respiratory horn is a projection of the chorion and aeropyles are grouped onthe apex; this structure obviously functions in gaseous exchange. It has been observedthat eggs hatch after being in bubbling water for 48 hours (for mass rearing purposes)(Aagesen, personal communication). Thus, it seems possible that the egg’s respiratorysystem can function as a plastron. At the base of the respiratory horn, we observed amicropyle (Fig. 3). No hair-like structures were found as Emmart (1933) previouslyreported.
Our use of the term respiratory horn is made in the same sense as that of Hinton(1981) who used it to describe structures on eggs which are used in atmospheric res-piration. In the case of
A. obliqua
, this structure seems obviously to function in respi-ration, and the term respiratory horn seems preferable to other terms commonly usedto describe similar structures on tephritid eggs such as “pedicel” (Headrick & Goeden1990), “stalk” (Benjamin 1934, Cavender & Goeden 1984), or “lobe” and “appendage”(Norrbom 1985). Pedicel, for example, is the diminutive of the latin
ped
or
pedis
andmeans “little foot”; stalk refers to a lengthened part on which an organ grows or issupported; thus, neither of the former terms is appropriate to the structure seen here,and other terms are less descriptive of its apparent function.
Most of the egg’s surface is smooth with no obvious structures or “sculpturing” onit. However, it is reticulated near both ends and these areas have a granular texture(Fig. 4). The reticulation at the respiratory horn’s base is more strongly defined, butless polygonal than at the posterior end.
The entire chorion as well as the respiratory horn is cavernous; thus, there is acontinuous network of open spaces extending from the respiratory horn to the rest ofthe chorion over the entire egg body. The internal structures of the chorion and respi-ratory horn are shown in Fig. 5 and 6, respectively.
Bacteria were observed on recently oviposited eggs of
A. obliqua
. Previous obser-vations by Murillo et al. (1990) showed that some bacterial flora, which they called“indigenous microflora”, are associated with
A. obliqua
eggs just after oviposition.These bacterial species belong to the Enterobacteriaceae group, which also have beenreported on the eggs of other tephritid species (Drew & Lloyd 1989, Howard et al.1985).
Anastrepha fraterculus
eggs are elongated, creamy white, with a very short chori-onic extension anteriorly and bluntly rounded posteriorly (Fig. 2). The micropyle ismore conspicuous than that observed in
A. obliqua
, and it is located almost at the an-terior apex of the egg (Fig. 8). The eggs of
A. fraterculus
bear polygonal reticulationsover the anterior pole and subdistally to the posterior pole, and they are more clearlydefined than those of
A. obliqua
eggs. There are aeropyles on the borders of the retic-ulations on the anterior pole, surrounding the micropyle (Fig. 9).
Anastrepha obliqua
lay their eggs individually with the respiratory horn remain-ing at the fruit’s surface from the micropyle upward (Fig. 7). This suggests that the
A.obliqua
egg needs unobstructed air exchange with the atmosphere before the larvahatches. The angle of the respiratory horn relative to the fruit surface may change,but it remains exposed. Average oviposition depth was 1.34 mm in the three mangovarieties studied, and there were no statistically significant differences among them(1.347 mm oviposition depth for Yellow and Tommy Atkins, and 1.334 mm for Mora;SE 0.022, 0.032 and 0.040, respectively;
P
≤
0.05).
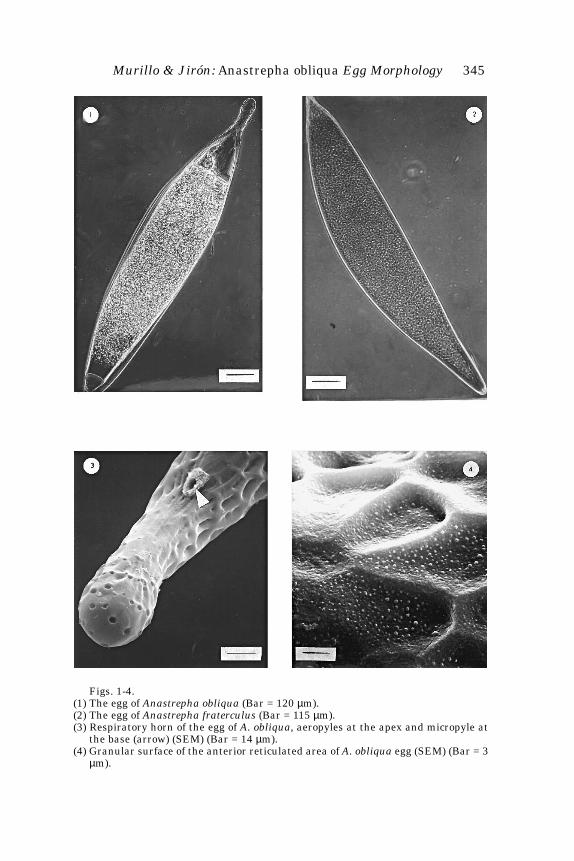
Murillo & Jirón:
Anastrepha obliqua
Egg Morphology
345
Figs. 1-4.(1) The egg of Anastrepha obliqua (Bar = 120 µm).(2) The egg of Anastrepha fraterculus (Bar = 115 µm).(3) Respiratory horn of the egg of A. obliqua, aeropyles at the apex and micropyle at
the base (arrow) (SEM) (Bar = 14 µm).(4) Granular surface of the anterior reticulated area of A. obliqua egg (SEM) (Bar = 3
µm).
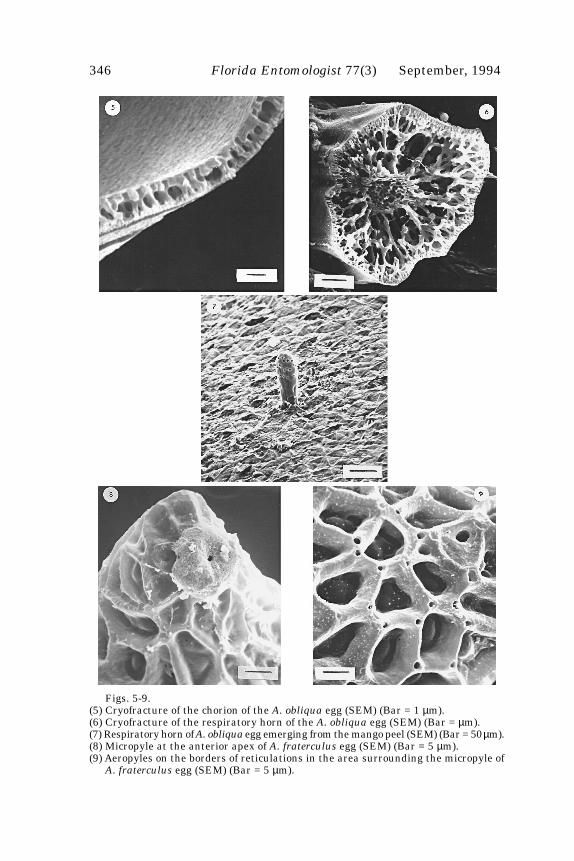
346
Florida Entomologist
77(3) September, 1994
Figs. 5-9.(5) Cryofracture of the chorion of the A. obliqua egg (SEM) (Bar = 1 µm).(6) Cryofracture of the respiratory horn of the A. obliqua egg (SEM) (Bar = µm).(7) Respiratory horn of A. obliqua egg emerging from the mango peel (SEM) (Bar = 50 µm).(8) Micropyle at the anterior apex of A. fraterculus egg (SEM) (Bar = 5 µm).(9) Aeropyles on the borders of reticulations in the area surrounding the micropyle of
A. fraterculus egg (SEM) (Bar = 5 µm).

Murillo & Jirón: Anastrepha obliqua Egg Morphology 347
Plastron-bearing respiratory horns have been reported in species belonging toseven dipteran families (Dryomyzidae, Sepsidae, Coelopidae, Sphaeroceridae, Droso-philidae, Cordiluridae and Muscidae) (Hinton 1981). However, so far there has beenno report of its occurrence in Tephritidae. Mouzaki & Margaritis (1987) suggestedthat the respiratory structures present in the family Drosophilidae have been re-placed in Tephritidae by: a) an area for respiration on the main body of the egg, b)later by the whole egg’s surface, and c) very limited areas on the main body and theanterior cup of the egg. However, at least in A. obliqua a respiratory horn is present,and the structures found in A. nigrifascia (Stone), A. pittieri (Caravallo) (Norrbom1985), Paracantha gentilis (Hering) (Headrick & Goeden 1990), P. cultaris (Coquillett)(Cavender & Goeden 1984), P. culta (Wiedemann), P. forficula (Benjamin), Toxotry-pana curvicauda Gerstaecker, and Zonosemata electa (Say) (Benjamin 1934) eggs alsomay serve a similar function.
In addition to a respiratory function, the egg horn of A. obliqua may play a role inthe recognition of infested hosts (fruits) by gravid females. Takasu and Hirose (1988)found that Ooencyrtus nezarae Ishii (Encyrtidae: Hymenoptera) utilizes the egg stalk(as they named it) as an external marker, which permits the female to recognize hostswhich are already infested. It is possible that A. obliqua females, before oviposition,also check suitable fruit for a physical marker (exposed horn), and as a parasitism in-dicator complementary to oviposition deterring pheromones (chemical marker)(Prokopy & Roitberg 1984). However, this function needs to be demonstrated.
After the larva hatches it migrates into internal fruit tissues. The respiratory hornmaintains its physical integrity attached to the fruit peel, unless it is broken off byrubbing or handling.
Perhaps the exposure of the A. obliqua respiratory horn, outside the mango fruitpeel, could be an aspect to consider when planning post-harvest technology for freshfruits. There may be value in developing a chemical, physical or biological method todetect the presence of the respiratory horn. These methods could be developed also foruse in fruit disinfestation, e.g., addition of a surfactant to a fruit bath may allow waterto enter the chorion interior and limit egg respiration.
ACKNOWLEDGMENTS
The authors thank Dr. Mario Vargas and Dr. Francisco Hernández who collabo-rated in this study, and Dr. William Ramírez for his suggestions in early manuscripts.Dr. Allen Norrbom gave valuable suggestions and manuscript reviewing. We thankUnidad de Microscopia Electrónica (UME) of the Universidad de Costa Rica for all fa-cilities they gave for this study, and for the training course we received in that Center;also, Organismo Internacional Regional de Sanidad Agropecuaria (OIRSA) for speci-mens, and Jorge Ramírez for his comments on the final manuscript draft. Finally, theauthors thank the Consejo Nacional de Investigaciones Científicas (CONICIT) for eco-nomic support for the publication of this paper.
REFERENCES CITED
BENJAMIN, F. H. 1934. Descriptions of some native trypetid flies with notes on theirhabits. United States Dept. Agric. Technical Bulletin 401.
CARROLL, L. E., AND R. A. WHARTON. 1989. Morphology of the immature stages ofAnastrepha ludens (Diptera: Tephritidae). Ann. Entomol. Soc. America 82:201-214.
CAVENDER, G. L., AND R. D. GOEDEN. 1984. The life history of Paracantha cultaris(Coquillet) on wild sunflower, Helianthus annus L. ssp. lenticularis (Douglas)

348 Florida Entomologist 77(3) September, 1994
Cockerell, in southern California (Diptera: Tephritidae). Pan-Pacific. Entomol.60:213-218.
DREW, R. A. I., AND A. C. LLOYD. 1989. Bacteria associated with fruit flies and theirhost plants, pp. 131-140 in A.S. Robinson and G. Hooper, [eds], World CropPests. Fruit Flies, their biology, natural enemies and control. Amsterdam,Elsevier Science. v. 3A.
EMMART, E. W. 1933. The eggs of four species of the fruit flies of the genus Anas-trepha. Proc. Entomol. Soc. Washington 35: 184-191.
HEADRICK, D., AND R. D. GOEDEN. 1990. Description of the immature stages of Para-cantha gentilis (Diptera: Tephritidae). Ann. Entomol. Soc. America 83: 220-229.
HINTON, H. E. 1981. Biology of insect eggs. Pergamon Press, Oxford. v. I, 316 p.HOWARD, D. J., G. L. BUSH, AND J. A. BREZNAK. 1985. The evolutionary significance
of bacteria associated with Rhagoletis. Evolution 39:405-417.LAWRENCE, P. O. 1979. Immature stages of the Caribbean fruit fly Anastrepha sus-
pensa. Florida Entomol. 62: 214-219.MOUZAKI, D. G., AND L. H. MARGARITIS. 1987. Comparative structural study of the
egg-shell (chorion) in Dacus oleae, Rhagoletis cerasi, Ceratitis capitata, and Eu-rytoma amygdali. Fruit Flies Procs. II International Symposium, Crete, Sept.1986. p. 79-87.
MURILLO, T., P. RIVERA, F. HERNANDEZ, AND L. F. JIRON. 1990. Indigenous microfloraof the West Indies fruit fly, Anastrepha obliqua (Diptera Tephritidae). Fruits(France) 45: 629-631.
NORRBOM, A. L. 1985. Phylogenetic analysis and taxonomy of the cryptostrepha, dac-iformis, robusta and schausi species groups of Anastrepha Schiner (Diptera: Te-phritidae). Ph.D. Thesis. Pennsylvania Sta. Univ. 359 p.
NORRBOM, A. L., AND R. H. FOOTE. 1989. The taxonomy and zoogeography of the ge-nus Anastrepha (Diptera: Tephritidae), pp. 15-26 in A.S. Robinson and G.Hooper, [eds.], World Crop Pests. Fruit Flies, their biology, natural enemiesand control. Amsterdam, Elsevier Science. v. 3A.
PROKOPY, R. J., AND B. ROITBERG. 1984. Foraging behavior of true fruit flies. Ameri-can Scientist 72: 41-49.
SEIN, F. 1933. Anastrepha (Trypetidae, Diptera) fruit flies in Puerto Rico. J. Dept. Ag-ric. Puerto Rico 17:183-196.
STECK, G. J., AND A. MALAVASI. 1988. Description of immature stages of Anastrephabistrigata (Diptera Tephritidae). Ann. Entomol. Soc. America 81: 1004-1009.
STECK, G. J., AND R. A. WHARTON. 1988. Description of immature stages of Anas-trepha interrupta, A. limae, A. grandis (Diptera: Tephritidae). Ann. Entomol.Soc. America 81: 994-1003.
TAKASU, K., AND Y. HIROSE. 1988. Host discrimination in the parasitoid Ooencyrtusnezarae: the role of the egg stalk as an external marker. Entom. Exper. & Appl.47:45-48.

Childers: Copper Formulations and Rust Mite Control
349
EFFECT OF DIFFERENT COPPER FORMULATIONS TANK-MIXED WITH FENBUTATIN-OXIDE
FOR CONTROL OF CITRUS RUST MITES (ACARI: ERIOPHYIDAE) ON FLORIDA CITRUS
C
ARL
C. C
HILDERS
University of Florida, IFASCitrus Research and Education Center
700 Experiment Station RoadLake Alfred, FL 33850
A
BSTRACT
One or more rates of fenbutatin-oxide (= Vendex
) 4 L applied alone were comparedwith the same rates of Vendex tank-mixed with one or more formulations of copperand an untreated check for control of citrus rust mite,
Phyllocoptruta oleivora
(Ash-mead) (Acari: Eriophyidae), in 4 citrus grove sites between 1988 and 1991. The copperformulations corresponded to recommended rates in the Florida Citrus Spray Guideand included: 99% GC basic copper sulfate (pentahydrate) with 53% metallic copper,a 77% WP copper hydroxide formulation containing 50% metallic copper and a 61.4%copper hydroxide DF formulation containing 40% metallic copper. Reduced effective-ness of Vendex occurred when combined with all 3 copper formulations. Both copperhydroxide formulations were more disruptive in reducing the effectiveness of Vendexin controlling citrus rust mite compared with the basic copper sulfate formulation. In-creases in citrus rust mite numbers occurred in the copper-only treatments in 2 of the3 field experiments. In both instances, population increases in the copper-only treat-ments occurred at the same time as those in the untreated check trees, suggesting astimulatory effect by the copper compounds.
Key Words: Chemical control,
Phyllocoptruta oleivora
, miticides, spray tank-mixes
R
ESUMEN
Varias dosis de oxido de fenbutatina (Vendex) 4 L aplicadas solas fueron compara-das con mezclas en tanque de diferentes formulaciones de cobre mas fenbutatina a lasmismas dosis y con un testigo no tratado, para el control del acaro tostador,
Phyllo-coptruta oleivora
(Ashmead) (Acari: Eriophyidae), en cuatro localidades con huertasde citricos entre 1988 y 1991. Las formulaciones de cobre correspondieron a las reco-mendadas en la Guía de Aspersiones de los Cítricos de la Florida e incluyeron: 99%GC de sulfato de basico de cobre (pentahidratado) con 53% de cobre metalico, una for-mulación de hidroxido de cobre al 77% WP conteniendo 50% de cobre metalico y unaformulación del 6l.4% de hidroxido de cobre DF con un 40% de cobre metalico. La efec-tividad del Vendex se redujo cuando este fue combinado con las tres formulaciones decobre. Las formulaciones de hidroxido de cobre redujeron mas fuertemente la efecti-vidad del Vendex que el sulfato basico de cobre en el control del acaro tostador. En dosde los tres campos experimentales se observaron aumentos en la mortalidad de losacaros tostadores. En ambos casos, la población en los tratamientos de cobre solo au-mentó al mismo tiempo que en los arboles no tratados, sugiriendo un efecto estimula-
dor por parte de los compuestos de cobre.
Copper compounds are recommended for use on Florida citrus to control severalfungal diseases including greasy spot (
Mycosphaerella citri
Whiteside), melanose (
Di-
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

350
Florida Entomologist
77(3) September, 1994
aporthe citri
Wolf) and citrus scab (
Elsinoe fawcettii
Bitancourt & Jenkins) (Timmer& McGovern 1993, Whiteside 1988). Several formulations of copper, that contain me-tallic copper, including copper sulfate and copper hydroxide, have been used by Flor-ida citrus growers for many years. The materials are relatively inexpensive whileproviding fungicidal activity. Many citrus growers apply copper in the postbloom andsummer sprays, in combination with a miticide or insecticide, for broad spectrum pestcontrol and to minimize application costs.
The citrus rust mite,
Phyllocoptruta oleivora
(Ashmead), and the spider mites,
Eu-tetranychus banksi
(McGregor) and
Panonychus citri
(McGregor), can be serious pestsduring the postbloom and summer periods and thus may require a miticide applica-tion during one or both times, depending upon citrus variety, mite presence and riskof population increase (Childers 1987).
Thompson (1939) found that
P. citri
increased following copper sprays on citrusand Holloway et al. (1942) reported that this spider mite was more abundant follow-ing sprays containing compounds of copper, zinc and lime compared with untreatedtrees. Griffith & Fisher (1949) observed that
Phyllocoptruta oleivora
and
Panonychuscitri
increased abnormally on citrus in Florida following the use of sprays containingcopper, zinc, or lime or exposure to road dust or fertilizer residues on fruit and leaves.Johnson (1960a) evaluated 5 copper formulations, including basic copper sulfate, cop-per oxide, copper hydroxysulfate, copper sulfate, and tetra copper calcium oxychlo-ride, in combination with zineb for control of
Phyllocoptruta oleivora
. All of the coppercompounds and rates tested reduced the effectiveness of zineb in the summer spraybut not in the postbloom spray. This supported earlier work by Johnson et al. (1957)that showed zineb provided less control of
P. oleivora
when combined with fixed cop-per compared with zineb alone.
Residual control by chlorobenzilate or dicofol against the citrus rust mite was re-duced when copper or methidathion was tank-mixed for use on citrus in Texas (Dean1979). Densities of
Brevipalpus phoenicis
(Geijskes) (Acari: Tenuipalpidae) increasedafter frequent copper oxychloride applications on tea in Indonesia (Oomen 1982).Eger et al. (1985) evaluated tank mixtures of copper with chlorpyrifos or ethion ap-plied in 2.338 or 4.676 kiloliters of water per hectare using an airblast sprayer. Theyfound that residual control of the citrus rust mite was reduced with the copper + or-ganophosphate combinations compared with the organophosphate compounds ap-plied alone.
Hirsutella thompsonii
Fisher, a fungus pathogenic to citrus rust mite, wassuspected of being adversely affected by the copper.
Vendex is not affected by differences in water pH because the organotin compoundis extremely stable. The formulation forms a suspension, not a solution, in water (Sea-man & Riedl 1986). This would suggest that a coating of the miticide by petroleum oilwould reduce the efficacy of the contact miticide (Childers & Selhime 1983).
Grower complaints of poor citrus rust mite control with fenbutatin-oxide + coppertank mixes prompted the field evaluations reported in this paper. The residual activ-ity of one or more copper formulations in combination with one or more rates of Ven-dex was evaluated between 1987 and 1991 and compared with the miticide appliedalone at the same rates. Results of these studies are reported here.
M
ATERIALS
AND
M
ETHODS
One or more rates of Vendex, 4 pounds active ingredient per gallon liquid formu-lation (4 L) (479 g AI per liter) (E. I. DuPont de Nemours, Wilmington, DE), appliedalone were compared with the same rates of Vendex tank-mixed with one or more for-
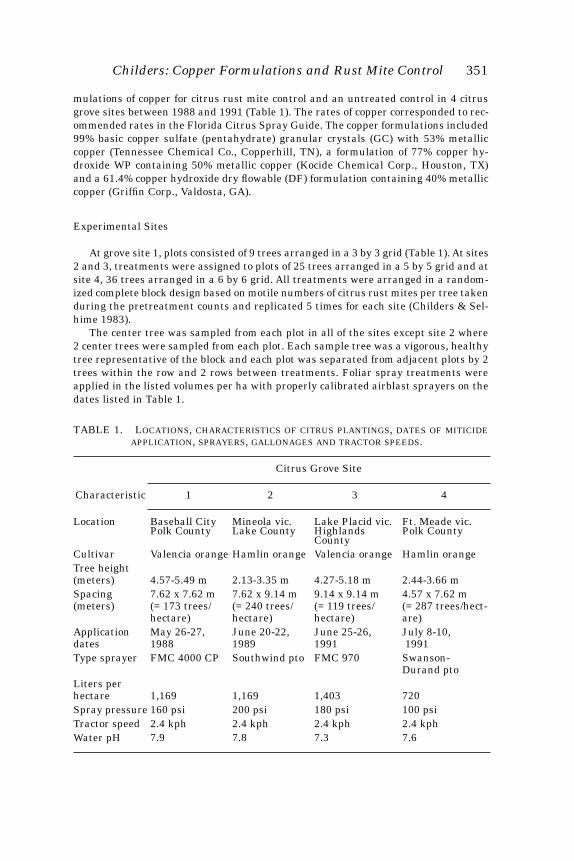
Childers: Copper Formulations and Rust Mite Control
351
mulations of copper for citrus rust mite control and an untreated control in 4 citrusgrove sites between 1988 and 1991 (Table 1). The rates of copper corresponded to rec-ommended rates in the Florida Citrus Spray Guide. The copper formulations included99% basic copper sulfate (pentahydrate) granular crystals (GC) with 53% metalliccopper (Tennessee Chemical Co., Copperhill, TN), a formulation of 77% copper hy-droxide WP containing 50% metallic copper (Kocide Chemical Corp., Houston, TX)and a 61.4% copper hydroxide dry flowable (DF) formulation containing 40% metalliccopper (Griffin Corp., Valdosta, GA).
Experimental Sites
At grove site 1, plots consisted of 9 trees arranged in a 3 by 3 grid (Table 1). At sites2 and 3, treatments were assigned to plots of 25 trees arranged in a 5 by 5 grid and atsite 4, 36 trees arranged in a 6 by 6 grid. All treatments were arranged in a random-ized complete block design based on motile numbers of citrus rust mites per tree takenduring the pretreatment counts and replicated 5 times for each site (Childers & Sel-hime 1983).
The center tree was sampled from each plot in all of the sites except site 2 where2 center trees were sampled from each plot. Each sample tree was a vigorous, healthytree representative of the block and each plot was separated from adjacent plots by 2trees within the row and 2 rows between treatments. Foliar spray treatments wereapplied in the listed volumes per ha with properly calibrated airblast sprayers on thedates listed in Table 1.
TABLE 1. L
OCATIONS
,
CHARACTERISTICS
OF
CITRUS
PLANTINGS
,
DATES
OF
MITICIDEAPPLICATION
,
SPRAYERS
,
GALLONAGES
AND
TRACTOR
SPEEDS
.
Characteristic
Citrus Grove Site
1 2 3 4
Location Baseball CityPolk County
Mineola vic.Lake County
Lake Placid vic.Highlands County
Ft. Meade vic.Polk County
Cultivar Valencia orange Hamlin orange Valencia orange Hamlin orangeTree height (meters) 4.57-5.49 m 2.13-3.35 m 4.27-5.18 m 2.44-3.66 mSpacing (meters)
7.62 x 7.62 m(= 173 trees/hectare)
7.62 x 9.14 m(= 240 trees/hectare)
9.14 x 9.14 m(= 119 trees/hectare)
4.57 x 7.62 m(= 287 trees/hect-are)
Application dates
May 26-27, 1988
June 20-22, 1989
June 25-26, 1991
July 8-10, 1991
Type sprayer FMC 4000 CP Southwind pto FMC 970 Swanson-Durand pto
Liters per hectare 1,169 1,169 1,403 720Spray pressure 160 psi 200 psi 180 psi 100 psiTractor speed 2.4 kph 2.4 kph 2.4 kph 2.4 kphWater pH 7.9 7.8 7.3 7.6

352
Florida Entomologist
77(3) September, 1994
Mite Sampling
Twenty fruit were examined at random around the canopy perimeter of each sam-ple tree at recorded time intervals before and after spraying of the treatments. Livemotile citrus rust mites were counted using a 10x hand lens equipped with a 1 cm
2
grid subdivided into 4 mm
2
subunits (Childers & Selhime 1983). Two places on oppo-site sides of each fruit outside of the exposed sunspot were examined. Population den-sity estimates of citrus rust mite were obtained by counting the number of livingmotile mites present within the combined 2 cm
2
area per fruit and recorded as one ob-servation. In test 2, 10 fruit were examined at random around the canopy perimeterfrom each of 2 sample trees with a total of 20 fruit per plot.
Twenty leaves from the spring flush were picked at random around the canopy pe-rimeter of each sample tree at designated time intervals before and after spraying.Each leaf sample was placed in separately labeled paper bags and returned to the lab-oratory. A stereomicroscope was used to examine a 1 cm
2
area on both the upper andlower leaf surfaces in test 1, or a 2 cm
2
area on both leaf surfaces in tests 4 and 5. Livemotile citrus rust mites were counted within each area on both the upper and lowerleaf surfaces. The 2 counts were combined as 1 observation and equalled either 2 or 4cm
2
area of leaf surface. In test 3, 10 leaves were collected from each of 2 trees in thecenter of each plot.
Cumulative citrus rust mite-days were determined for each treatment within agrove each year by calculating the area under the population growth curve over time(Allen 1976). Mite-days were calculated using the formula presented by Childers et al.(1987).
Fruit Damage Ratings
One hundred fruit were examined randomly around the perimeter of each sampletree following completion of a field experiment. The cumulative percentage of surfacearea with russeting from citrus rust mite feeding injury was recorded for each fruit.This was determined by estimating the percentage area of russeting on one side ofeach fruit and then turning the fruit 180
°
and repeating the estimate of russeted areaagain. The combined estimated russeted area of sides one and two for each fruit wasthen divided by two. A series of plates with line drawings of mature sized fruit withcalculated proportional areas of surface injury were prepared to provide a consistentreference for the person determining rind blemish injury. A fruit was classified as re-jected for the fresh market when rind blemish from citrus rust mite feeding exceeded5% of the total surface area.
Statistical Analysis
In all experiments, data were subjected to analysis of variance; Duncan’s (1955)multiple range test was used to separate treatment means when the ANOVA provideda significant F value (P
≤
0.05) (SAS Institute 1991). Motile citrus rust mite countswere subjected to Log
10
(X + 1) transformations for statistical analysis. Untrans-formed means are shown in all tables.
R
ESULTS
Site 1, ‘Valencia’ Orange
Sustained moderate numbers of citrus rust mites were present in the untreatedcheck trees through August 2 following treatment applications on May 26 and 27. Cit-

Childers: Copper Formulations and Rust Mite Control
353
rus rust mite densities on treated trees were significantly higher between August 2and 25 in the Vendex + copper sulfate or hydroxide combinations compared with theVendex only treatments at the same rate (Table 2). Cumulative mite-days were sig-nificantly higher with the 2.10 kg rate of Vendex + copper hydroxide and the 1.40 kgrate of Vendex + copper sulfate compared with the same rates of Vendex applied alone.The other combinations of copper + Vendex did not result in significantly higher cu-mulative mite-days than the corresponding rates of Vendex alone. Damage rating dif-ferences were not significantly different between the miticide treatments in thisexperiment (Table 2). Based on these data, all Vendex treatments performed well.
Site 2, ‘Hamlin’ Orange
Moderate to high densities of citrus rust mites were present on the untreated treesbetween July 27 and October 12 following treatment applications (Table 3). Vendex at1.68 kg + copper hydroxide had a significantly higher citrus rust mite density on thefruit by September 20 compared with Vendex at 1.68 kg applied alone. Vendex at 1.68kg + copper sulfate had a significantly higher citrus rust mite density by September27 compared with Vendex at the same rate applied alone. These differences continuedthrough October 18. Vendex at 2.24 kg + copper hydroxide had significantly higher cit-rus rust mite densities by September 27 compared with Vendex at 2.24 kg appliedalone. Again, these differences continued through October 18. The Vendex at 2.24 kg+ copper sulfate combination was not significantly different from Vendex at 2.24 kguntil October 3. These differences in citrus rust mite densities continued to be signif-icantly higher through October 12. Vendex at 2.80 kg combined with either copper hy-droxide or copper sulfate had significantly higher citrus rust mite densities on thefruit by September 20 and 27 and October 18 compared with Vendex at 2.80 kg ap-plied alone. These high rust mite densities occurred while the population was increas-ing in the untreated check trees.
Consistently higher citrus rust mite densities ranging from 1 to 4 times higherwere present in both copper hydroxide and copper sulfate treated trees between Au-gust 17 and September 27 compared with the untreated trees.
Cumulative mite-day comparisons were significantly different between the Ven-dex at 1.68 kg + copper hydroxide treatment compared with Vendex at 1.68 kg appliedalone (Table 4). Cumulative mite-day differences between the remaining miticidetreatments were not significantly different although numerically higher densities ofcitrus rust mites were consistently obtained in the Vendex + copper treatments com-pared with the Vendex only treatments.
Significantly higher percentages of russeted and rejected fruit were obtained inthe low rate of Vendex combined with either copper hydroxide or copper sulfate com-pared with the low rate of Vendex applied alone (Table 4). The highest percentages ofrusseted and rejected fruit were obtained in the copper-only treatments comparedwith all other treatments including the untreated check trees.
Site 3, ‘Valencia’ Orange
Low to moderate citrus rust mite densities were present on both fruit and leavesbetween July 11 and August 19 on the untreated check trees (Tables 5, 6). Low but sig-nificantly higher citrus rust mite densities were present on the Vendex + copper DFtreatment by August 12 compared with Vendex applied alone. Significantly higher cit-rus rust mite densities on the fruit were consistently obtained on the Vendex + copperDF treatment between August 12 and September 17 compared with Vendex applied
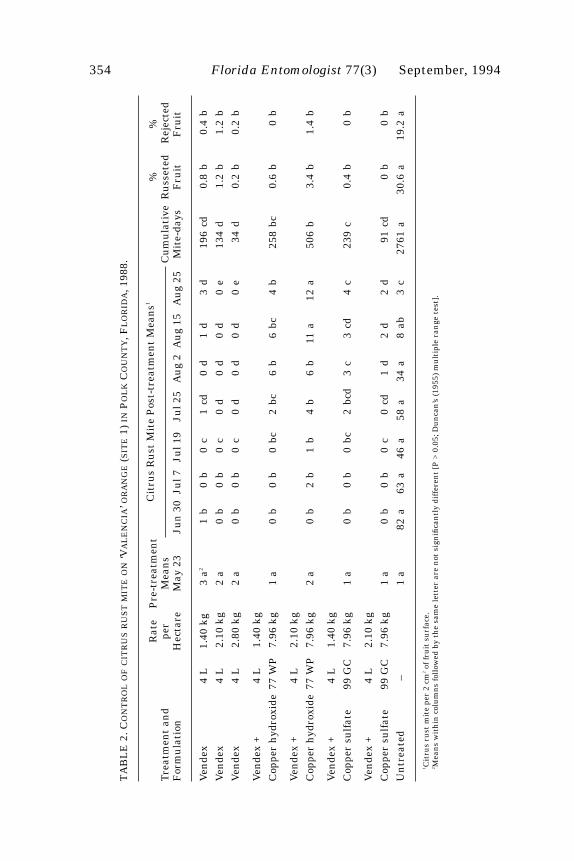
354
Florida Entomologist
77(3) September, 1994
1
Cit
rus
rust
mit
e pe
r 2
cm
2
of
fru
it s
urf
ace.
2
Mea
ns
wit
hin
col
um
ns
foll
owed
by
the
sam
e le
tter
are
not
sig
nifi
can
tly
diff
eren
t [P
> 0
.05;
Du
nca
n’s
(19
55)
mu
ltip
le r
ange
tes
t].
TA
BL
E 2
. C
ON
TR
OL
OF
CIT
RU
S
RU
ST
MIT
E
ON
‘V
AL
EN
CIA
’
OR
AN
GE
(
SIT
E
1)
IN
P
OL
K
C
OU
NT
Y
, F
LO
RID
A
, 198
8.
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tre
atm
ent
Mea
ns
May
23
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns
1
Cu
mu
lati
veM
ite-
days
%R
uss
eted
F
ruit
% R
ejec
ted
Fru
itJu
n 3
0Ju
l 7Ju
l 19
Jul 2
5A
ug
2A
ug
15A
ug
25
Ven
dex
4 L
1.40
kg
3 a
2
1b
0b
0c
1cd
0d
1d
3d
196
cd0.
8b
0.4
bV
ende
x 4
L2.
10 k
g2
a0
b0
b0
c0
d0
d0
d0
e13
4d
1.2
b 1
.2b
Ven
dex
4 L
2.80
kg
2 a
0b
0b
0c
0d
0d
0d
0e
34d
0.2
b 0
.2b
Ven
dex
+ 4
L1.
40 k
gC
oppe
r h
ydro
xide
77 W
P7.
96 k
g1
a0
b0
b0
bc2
bc6
b6
bc4
b25
8bc
0.6
b 0
b
Ven
dex
+ 4
L2.
10 k
gC
oppe
r h
ydro
xide
77 W
P7.
96 k
g2
a0
b2
b1
b4
b6
b11
a12
a50
6b
3.4
b 1
.4b
Ven
dex
+ 4
L1.
40 k
gC
oppe
r su
lfat
e99
GC
7.96
kg
1 a
0b
0b
0bc
2bc
d3
c3
cd4
c23
9c
0.4
b 0
b
Ven
dex
+ 4
L2.
10 k
gC
oppe
r su
lfat
e99
GC
7.96
kg
1 a
0b
0b
0c
0cd
1d
2d
2d
91cd
0b
0b
Un
trea
ted
–
1 a
82a
63a
46a
58a
34a
8ab
3c
2761
a30
.6a
19.2
a
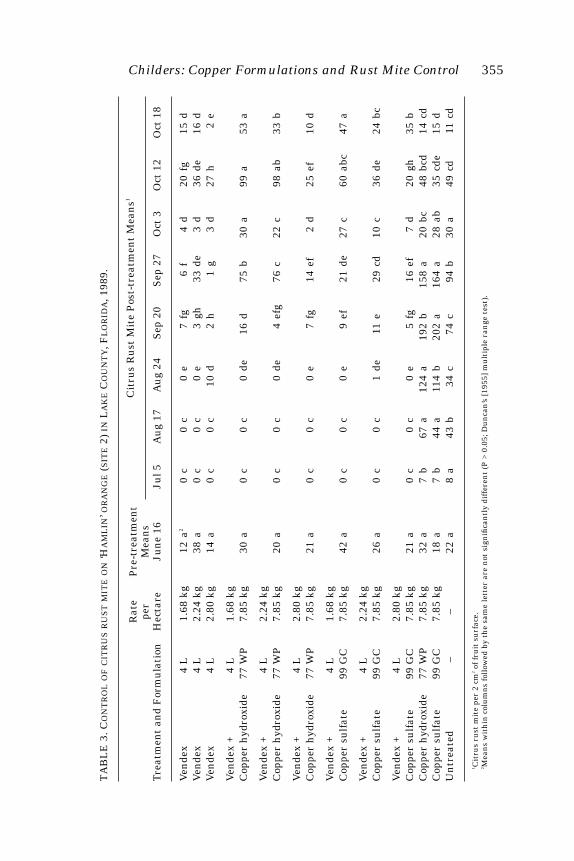
Childers: Copper Formulations and Rust Mite Control
355
1
Cit
rus
rust
mit
e pe
r 2
cm
2
of
fru
it s
urf
ace.
2
Mea
ns
wit
hin
col
um
ns
foll
owed
by
the
sam
e le
tter
are
not
sig
nifi
can
tly
diff
eren
t (P
> 0
.05;
Du
nca
n’s
[19
55]
mu
ltip
le r
ange
tes
t).
TA
BL
E 3
. C
ON
TR
OL
OF
CIT
RU
S
RU
ST
MIT
E
ON
‘H
AM
LIN
’
OR
AN
GE
(
SIT
E
2)
IN
L
AK
E
C
OU
NT
Y
, F
LO
RID
A
, 198
9.
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tre
atm
ent
Mea
ns
Jun
e 16
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns
1
Jul 5
Au
g 17
Au
g 24
Sep
20
Sep
27
Oct
3O
ct 1
2O
ct 1
8
Ven
dex
4 L
1.68
kg
12a
2
0c
0c
0e
7fg
6f
4d
20fg
15d
Ven
dex
4 L
2.24
kg
38a
0c
0c
0e
3gh
33de
3d
36de
16d
Ven
dex
4 L
2.80
kg
14a
0c
0c
10d
2h
1g
3d
27h
2e
Ven
dex
+ 4
L1.
68 k
gC
oppe
r h
ydro
xide
77 W
P7.
85 k
g30
a0
c0
c0
de16
d75
b30
a99
a53
a
Ven
dex
+ 4
L2.
24 k
gC
oppe
r h
ydro
xide
77 W
P7.
85 k
g20
a0
c0
c0
de4
efg
76c
22c
98ab
33b
Ven
dex
+ 4
L2.
80 k
gC
oppe
r h
ydro
xide
77 W
P7.
85 k
g21
a0
c0
c0
e7
fg14
ef2
d25
ef10
d
Ven
dex
+ 4
L1.
68 k
gC
oppe
r su
lfat
e99
GC
7.85
kg
42a
0c
0c
0e
9ef
21de
27c
60ab
c47
a
Ven
dex
+ 4
L2.
24 k
gC
oppe
r su
lfat
e99
GC
7.85
kg
26a
0c
0c
1de
11e
29cd
10c
36de
24bc
Ven
dex
+ 4
L2.
80 k
gC
oppe
r su
lfat
e99
GC
7.85
kg
21a
0c
0c
0e
5fg
16ef
7d
20gh
35b
Cop
per
hyd
roxi
de77
WP
7.85
kg
32a
7b
67a
124
a19
2b
158
a20
bc48
bcd
14cd
Cop
per
sulf
ate
99 G
C7.
85 k
g18
a7
b44
a11
4b
202
a16
4a
28ab
35cd
e15
dU
ntr
eate
d–
–22
a8
a43
b34
c74
c94
b30
a49
cd11
cd
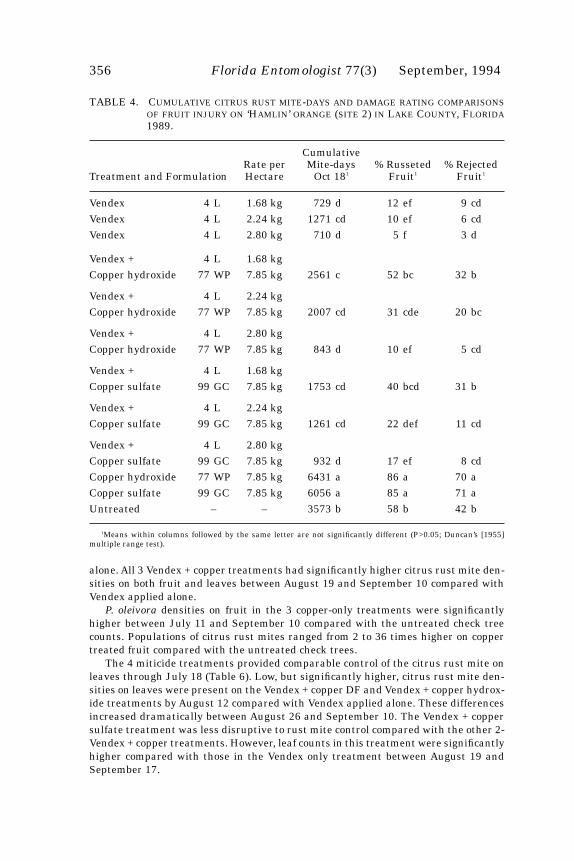
356
Florida Entomologist
77(3) September, 1994
alone. All 3 Vendex + copper treatments had significantly higher citrus rust mite den-sities on both fruit and leaves between August 19 and September 10 compared withVendex applied alone.
P. oleivora
densities on fruit in the 3 copper-only treatments were significantlyhigher between July 11 and September 10 compared with the untreated check treecounts. Populations of citrus rust mites ranged from 2 to 36 times higher on coppertreated fruit compared with the untreated check trees.
The 4 miticide treatments provided comparable control of the citrus rust mite onleaves through July 18 (Table 6). Low, but significantly higher, citrus rust mite den-sities on leaves were present on the Vendex + copper DF and Vendex + copper hydrox-ide treatments by August 12 compared with Vendex applied alone. These differencesincreased dramatically between August 26 and September 10. The Vendex + coppersulfate treatment was less disruptive to rust mite control compared with the other 2-Vendex + copper treatments. However, leaf counts in this treatment were significantlyhigher compared with those in the Vendex only treatment between August 19 andSeptember 17.
1
Means within columns followed by the same letter are not significantly different (P>0.05; Duncan’s [1955]multiple range test).
TABLE 4. C
UMULATIVE
CITRUS
RUST
MITE
-DAYS AND DAMAGE RATING COMPARISONSOF FRUIT INJURY ON ‘HAMLIN’ ORANGE (SITE 2) IN LAKE COUNTY, FLORIDA1989.
Treatment and FormulationRate per Hectare
Cumulative Mite-days
Oct 181% Russeted
Fruit1% Rejected
Fruit1
Vendex 4 L 1.68 kg 729 d 12 ef 9 cdVendex 4 L 2.24 kg 1271 cd 10 ef 6 cdVendex 4 L 2.80 kg 710 d 5 f 3 d
Vendex + 4 L 1.68 kgCopper hydroxide 77 WP 7.85 kg 2561 c 52 bc 32 b
Vendex + 4 L 2.24 kgCopper hydroxide 77 WP 7.85 kg 2007 cd 31 cde 20 bc
Vendex + 4 L 2.80 kgCopper hydroxide 77 WP 7.85 kg 843 d 10 ef 5 cd
Vendex + 4 L 1.68 kgCopper sulfate 99 GC 7.85 kg 1753 cd 40 bcd 31 b
Vendex + 4 L 2.24 kgCopper sulfate 99 GC 7.85 kg 1261 cd 22 def 11 cd
Vendex + 4 L 2.80 kgCopper sulfate 99 GC 7.85 kg 932 d 17 ef 8 cdCopper hydroxide 77 WP 7.85 kg 6431 a 86 a 70 aCopper sulfate 99 GC 7.85 kg 6056 a 85 a 71 aUntreated – – 3573 b 58 b 42 b
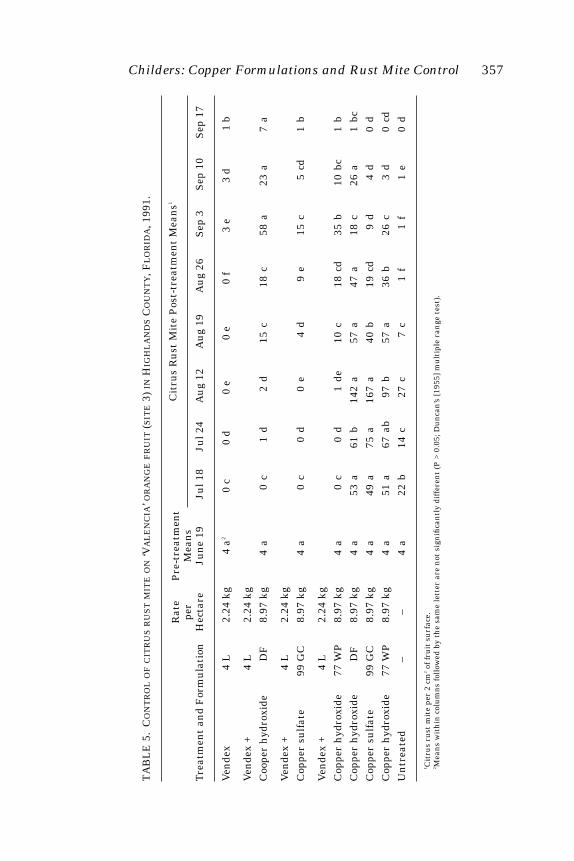
Childers: Copper Formulations and Rust Mite Control 357
1 Cit
rus
rust
mit
e pe
r 2
cm2 o
f fr
uit
su
rfac
e.2 M
ean
s w
ith
in c
olu
mn
s fo
llow
ed b
y th
e sa
me
lett
er a
re n
ot s
ign
ifica
ntl
y di
ffer
ent
(P >
0.0
5; D
un
can
’s [
1955
] m
ult
iple
ran
ge t
est)
.
TA
BL
E 5
.C
ON
TR
OL O
F C
ITR
US R
US
T M
ITE
ON
‘VA
LE
NC
IA’ O
RA
NG
E F
RU
IT (
SIT
E 3
) IN
HIG
HL
AN
DS C
OU
NT
Y, F
LO
RID
A, 1
991.
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tre
atm
ent
Mea
ns
Jun
e 19
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns1
Jul 1
8Ju
l 24
Au
g 12
Au
g 19
Au
g 26
Sep
3S
ep 1
0S
ep 1
7
Ven
dex
4
L2.
24 k
g4
a20
c0
d0
e0
e0
f3
e3
d1
b
Ven
dex
+
4 L
2.24
kg
Coo
per
hyd
roxi
de
D
F8.
97 k
g4
a0
c1
d2
d15
c18
c58
a23
a7
a
Ven
dex
+
4 L
2.24
kg
Cop
per
sulf
ate
99
GC
8.97
kg
4a
0c
0d
0e
4d
9e
15c
5cd
1b
Ven
dex
+
4 L
2.24
kg
Cop
per
hyd
roxi
de 7
7 W
P8.
97 k
g4
a0
c0
d1
de10
c18
cd35
b10
bc1
bC
oppe
r h
ydro
xide
DF
8.97
kg
4a
53a
61b
142
a57
a47
a18
c26
a1
bcC
oppe
r su
lfat
e 9
9 G
C8.
97 k
g4
a49
a75
a16
7a
40b
19cd
9d
4d
0d
Cop
per
hyd
roxi
de 7
7 W
P8.
97 k
g4
a51
a67
ab97
b57
a36
b26
c3
d0
cdU
ntr
eate
d–
–4
a22
b14
c27
c7
c1
f1
f1
e0
d
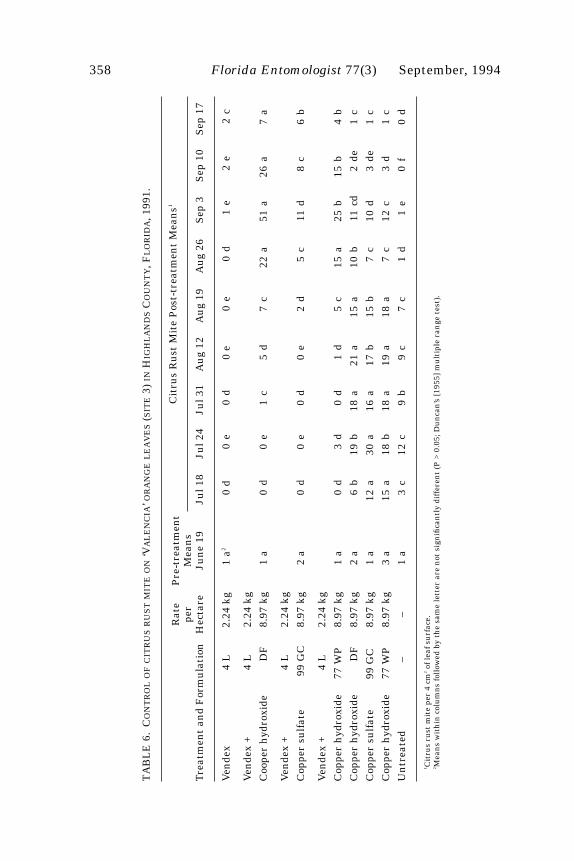
358 Florida Entomologist 77(3) September, 1994
1 Cit
rus
rust
mit
e pe
r 4
cm2 o
f le
af s
urf
ace.
2 Mea
ns
wit
hin
col
um
ns
foll
owed
by
the
sam
e le
tter
are
not
sig
nifi
can
tly
diff
eren
t (P
> 0
.05;
Du
nca
n’s
[19
55]
mu
ltip
le r
ange
tes
t).
TA
BL
E 6
.C
ON
TR
OL O
F C
ITR
US R
US
T M
ITE
ON
‘VA
LE
NC
IA’ O
RA
NG
E L
EA
VE
S (
SIT
E 3
) IN
HIG
HL
AN
DS C
OU
NT
Y, F
LO
RID
A, 1
991.
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tre
atm
ent
Mea
ns
Jun
e 19
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns1
Jul 1
8Ju
l 24
Jul 3
1A
ug
12A
ug
19A
ug
26S
ep 3
Sep
10
Sep
17
Ven
dex
4
L
2.2
4 kg
1 a
2
0
d
0
e
0
d
0
e
0
e
0
d
1
e
2
e
2
c
Ven
dex
+
4 L
2.24
kg
Coo
per
hyd
roxi
de
D
F8.
97 k
g1
a0
d0
e1
c5
d7
c22
a51
a26
a7
a
Ven
dex
+
4 L
2.24
kg
Cop
per
sulf
ate
99
GC
8.97
kg
2 a
0d
0e
0d
0e
2d
5c
11d
8c
6b
Ven
dex
+
4 L
2.24
kg
Cop
per
hyd
roxi
de77
WP
8.97
kg
1 a
0d
3d
0d
1d
5c
15a
25b
15b
4b
Cop
per
hyd
roxi
de
D
F8.
97 k
g2
a6
b19
b18
a21
a15
a10
b11
cd2
de1
cC
oppe
r su
lfat
e99
GC
8.97
kg
1 a
12a
30a
16a
17b
15b
7c
10d
3de
1c
Cop
per
hyd
roxi
de77
WP
8.97
kg
3 a
15a
18b
18a
19a
18a
7c
12c
3d
1c
Un
trea
ted
––
1 a
3c
12c
9b
9c
7c
1d
1e
0f
0d
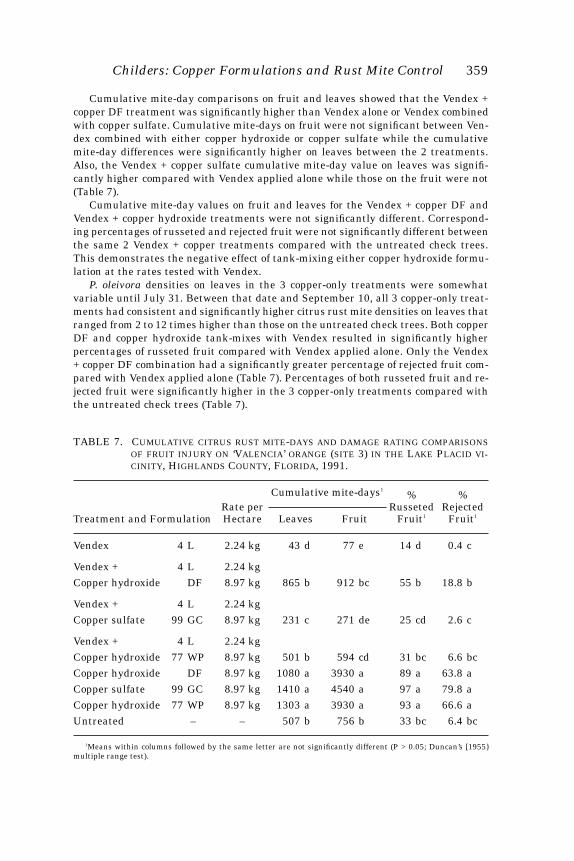
Childers: Copper Formulations and Rust Mite Control 359
Cumulative mite-day comparisons on fruit and leaves showed that the Vendex +copper DF treatment was significantly higher than Vendex alone or Vendex combinedwith copper sulfate. Cumulative mite-days on fruit were not significant between Ven-dex combined with either copper hydroxide or copper sulfate while the cumulativemite-day differences were significantly higher on leaves between the 2 treatments.Also, the Vendex + copper sulfate cumulative mite-day value on leaves was signifi-cantly higher compared with Vendex applied alone while those on the fruit were not(Table 7).
Cumulative mite-day values on fruit and leaves for the Vendex + copper DF andVendex + copper hydroxide treatments were not significantly different. Correspond-ing percentages of russeted and rejected fruit were not significantly different betweenthe same 2 Vendex + copper treatments compared with the untreated check trees.This demonstrates the negative effect of tank-mixing either copper hydroxide formu-lation at the rates tested with Vendex.
P. oleivora densities on leaves in the 3 copper-only treatments were somewhatvariable until July 31. Between that date and September 10, all 3 copper-only treat-ments had consistent and significantly higher citrus rust mite densities on leaves thatranged from 2 to 12 times higher than those on the untreated check trees. Both copperDF and copper hydroxide tank-mixes with Vendex resulted in significantly higherpercentages of russeted fruit compared with Vendex applied alone. Only the Vendex+ copper DF combination had a significantly greater percentage of rejected fruit com-pared with Vendex applied alone (Table 7). Percentages of both russeted fruit and re-jected fruit were significantly higher in the 3 copper-only treatments compared withthe untreated check trees (Table 7).
1Means within columns followed by the same letter are not significantly different (P > 0.05; Duncan’s [1955}multiple range test).
TABLE 7. CUMULATIVE CITRUS RUST MITE-DAYS AND DAMAGE RATING COMPARISONSOF FRUIT INJURY ON ‘VALENCIA’ ORANGE (SITE 3) IN THE LAKE PLACID VI-CINITY, HIGHLANDS COUNTY, FLORIDA, 1991.
Treatment and FormulationRate per Hectare
Cumulative mite-days1%
Russeted Fruit1
% Rejected
Fruit1Leaves Fruit
Vendex 4 L 2.24 kg 43 d 77 e 14 d 0.4 c
Vendex + 4 L 2.24 kgCopper hydroxide DF 8.97 kg 865 b 912 bc 55 b 18.8 b
Vendex + 4 L 2.24 kgCopper sulfate 99 GC 8.97 kg 231 c 271 de 25 cd 2.6 c
Vendex + 4 L 2.24 kgCopper hydroxide 77 WP 8.97 kg 501 b 594 cd 31 bc 6.6 bcCopper hydroxide DF 8.97 kg 1080 a 3930 a 89 a 63.8 aCopper sulfate 99 GC 8.97 kg 1410 a 4540 a 97 a 79.8 aCopper hydroxide 77 WP 8.97 kg 1303 a 3930 a 93 a 66.6 aUntreated – – 507 b 756 b 33 bc 6.4 bc

360 Florida Entomologist 77(3) September, 1994
Site 4, ‘Hamlin’ Orange
Low to moderate citrus rust mite densities on fruit in the untreated check treeswere recorded between July 15 and September 10 following treatment applications onJuly 8, 9 and 10. Population development of P. oleivora was evident on the back sidesof fruit by August 8 with no citrus rust mites present on the front sides of the samefruit in the Vendex treatments. Poor spray coverage resulted from inadequate pene-tration of pesticide spray through the canopy since only 720 liters of finished spraywere applied per hectare.
All Vendex treatments provided comparable control of citrus rust mite on the fruitthrough August 13 (Table 8). The Vendex + copper hydroxide and Vendex + copper DFcombinations generally had significantly higher citrus rust mite densities on the fruitbetween August 21 and September 10 compared with Vendex applied alone and theVendex + copper sulfate treatments. As in the previous field experiment (site 3), theamount of metallic copper applied per hectare was highest in the copper sulfate for-mulation followed by copper hydroxide WP and copper hydroxide DF. The 3 copper-only treatments did not have higher rust mite densities on fruit during this experi-ment compared with the untreated check trees (Table 8).
All 4 Vendex treatments had significantly lower cumulative mite-day values onfruit compared with the untreated check trees (Table 9). Only the Vendex + copper hy-droxide 77% WP treatment had significantly more russeted fruit compared with theother Vendex treatments in this experiment. Numerically higher percentages of re-jected fruit were found in the Vendex + copper DF and Vendex + copper hydroxidetreatments. However, they were not significantly different compared with the other 2Vendex treatments.
P. oleivora densities on leaves were low in all treatments throughout this experi-ment (Table 10). However, significantly higher citrus rust mite densities were re-corded on the Vendex + copper DF and Vendex + copper hydroxide treatments onAugust 27 and September 5 compared with Vendex applied alone or Vendex + coppersulfate. The 3 copper-only treatments did not have consistently higher citrus rustmite densities on the leaves compared with the untreated check trees. Cumulativemite-day differences between the copper-only and untreated check treatments werenot significantly different in this experiment (Table 9).
DISCUSSION
Results from this study show that the different copper formulations tested are in-compatible with Vendex when applied at the rates tested. The copper hydroxide DFformulation demonstrated the greatest degree of incompatibility followed by copperhydroxide WP. The copper sulfate formulation in combination with Vendex showedthe least disruption in residual rust mite control of the 3 formulations tested. Theamount of available metallic copper applied per hectare in tests 3 and 4 differed be-tween the 3 formulations as follows: copper DF = 1.45 kg, copper hydroxide = 1.81 kgand copper sulfate = 1.93 kg. This would suggest that one or more other componentsof the copper DF formulation were involved in creating increased antagonism in com-bination with the organo-tin miticide compared with the metallic copper contentalone. This phenomenon is not unique and has been reported with other tank mix-tures. For example, Venkata Ram (1963) found that nickel chloride hexahydrate con-trolled the fungal pathogen, Exobasidium vexans, on tea as effectively as copperoxychloride without increasing purple mite, Calacarus carinatus (Green) (Acari: Erio-phyidae).
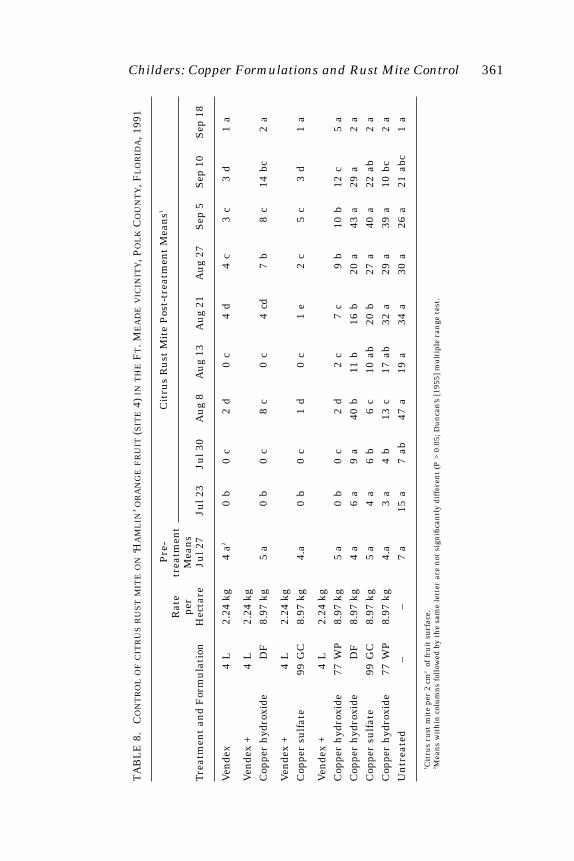
Childers: Copper Formulations and Rust Mite Control 361
1 Cit
rus
rust
mit
e pe
r 2
cm2 o
f fr
uit
su
rfac
e.2 M
ean
s w
ith
in c
olu
mn
s fo
llow
ed b
y th
e sa
me
lett
er a
re n
ot s
ign
ifica
ntl
y di
ffer
ent
(P >
0.0
5; D
un
can
’s [
1955
] m
ult
iple
ran
ge t
est.
TA
BL
E 8
. C
ON
TR
OL O
F C
ITR
US R
US
T M
ITE
ON
‘HA
ML
IN’ O
RA
NG
E F
RU
IT (
SIT
E 4
) IN
TH
E F
T. M
EA
DE
VIC
INIT
Y, P
OL
K C
OU
NT
Y, F
LO
RID
A, 1
991
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tr
eatm
ent
Mea
ns
Jul 2
7
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns1
Jul 2
3Ju
l 30
Au
g 8
Au
g 13
Au
g 21
Au
g 27
Sep
5S
ep 1
0S
ep 1
8
Ven
dex
4L
2.24
kg
4 a2
0b
0c
2d
0c
4d
4c
3c
3d
1a
Ven
dex
+4
L2.
24 k
gC
oppe
r h
ydro
xide
DF
8.97
kg
5 a
0b
0c
8c
0c
4cd
7b
8c
14bc
2a
Ven
dex
+4
L2.
24 k
gC
oppe
r su
lfat
e99
GC
8.97
kg
4.a
0b
0c
1d
0c
1e
2c
5c
3d
1a
Ven
dex
+4
L2.
24 k
gC
oppe
r h
ydro
xide
77W
P8.
97 k
g5
a0
b0
c2
d2
c7
c9
b10
b12
c5
aC
oppe
r h
ydro
xide
DF
8.97
kg
4 a
6a
9a
40b
11b
16b
20a
43a
29a
2a
Cop
per
sulf
ate
99G
C8.
97 k
g5
a4
a6
b6
c10
ab20
b27
a40
a22
ab2
aC
oppe
r h
ydro
xide
77W
P8.
97 k
g4.
a3
a4
b13
c17
ab32
a29
a39
a10
bc2
aU
ntr
eate
d–
–7
a15
a7
ab47
a19
a34
a30
a26
a21
abc
1a
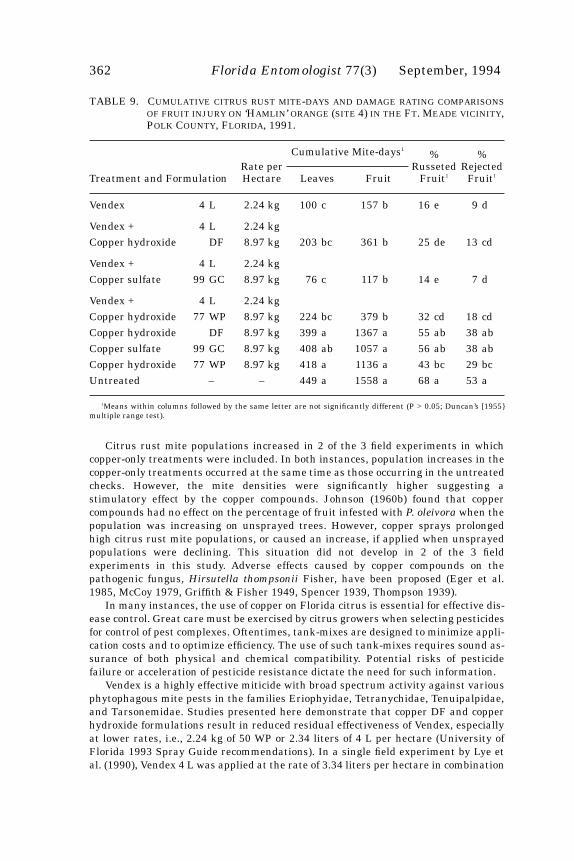
362 Florida Entomologist 77(3) September, 1994
Citrus rust mite populations increased in 2 of the 3 field experiments in whichcopper-only treatments were included. In both instances, population increases in thecopper-only treatments occurred at the same time as those occurring in the untreatedchecks. However, the mite densities were significantly higher suggesting astimulatory effect by the copper compounds. Johnson (1960b) found that coppercompounds had no effect on the percentage of fruit infested with P. oleivora when thepopulation was increasing on unsprayed trees. However, copper sprays prolongedhigh citrus rust mite populations, or caused an increase, if applied when unsprayedpopulations were declining. This situation did not develop in 2 of the 3 fieldexperiments in this study. Adverse effects caused by copper compounds on thepathogenic fungus, Hirsutella thompsonii Fisher, have been proposed (Eger et al.1985, McCoy 1979, Griffith & Fisher 1949, Spencer 1939, Thompson 1939).
In many instances, the use of copper on Florida citrus is essential for effective dis-ease control. Great care must be exercised by citrus growers when selecting pesticidesfor control of pest complexes. Oftentimes, tank-mixes are designed to minimize appli-cation costs and to optimize efficiency. The use of such tank-mixes requires sound as-surance of both physical and chemical compatibility. Potential risks of pesticidefailure or acceleration of pesticide resistance dictate the need for such information.
Vendex is a highly effective miticide with broad spectrum activity against variousphytophagous mite pests in the families Eriophyidae, Tetranychidae, Tenuipalpidae,and Tarsonemidae. Studies presented here demonstrate that copper DF and copperhydroxide formulations result in reduced residual effectiveness of Vendex, especiallyat lower rates, i.e., 2.24 kg of 50 WP or 2.34 liters of 4 L per hectare (University ofFlorida 1993 Spray Guide recommendations). In a single field experiment by Lye etal. (1990), Vendex 4 L was applied at the rate of 3.34 liters per hectare in combination
1Means within columns followed by the same letter are not significantly different (P > 0.05; Duncan’s [1955}multiple range test).
TABLE 9. CUMULATIVE CITRUS RUST MITE-DAYS AND DAMAGE RATING COMPARISONSOF FRUIT INJURY ON ‘HAMLIN’ ORANGE (SITE 4) IN THE FT. MEADE VICINITY,POLK COUNTY, FLORIDA, 1991.
Treatment and FormulationRate per Hectare
Cumulative Mite-days1%
Russeted Fruit1
% Rejected
Fruit1Leaves Fruit
Vendex 4 L 2.24 kg 100 c 157 b 16 e 9 d
Vendex + 4 L 2.24 kgCopper hydroxide DF 8.97 kg 203 bc 361 b 25 de 13 cd
Vendex + 4 L 2.24 kgCopper sulfate 99 GC 8.97 kg 76 c 117 b 14 e 7 d
Vendex + 4 L 2.24 kgCopper hydroxide 77 WP 8.97 kg 224 bc 379 b 32 cd 18 cdCopper hydroxide DF 8.97 kg 399 a 1367 a 55 ab 38 abCopper sulfate 99 GC 8.97 kg 408 ab 1057 a 56 ab 38 abCopper hydroxide 77 WP 8.97 kg 418 a 1136 a 43 bc 29 bcUntreated – – 449 a 1558 a 68 a 53 a
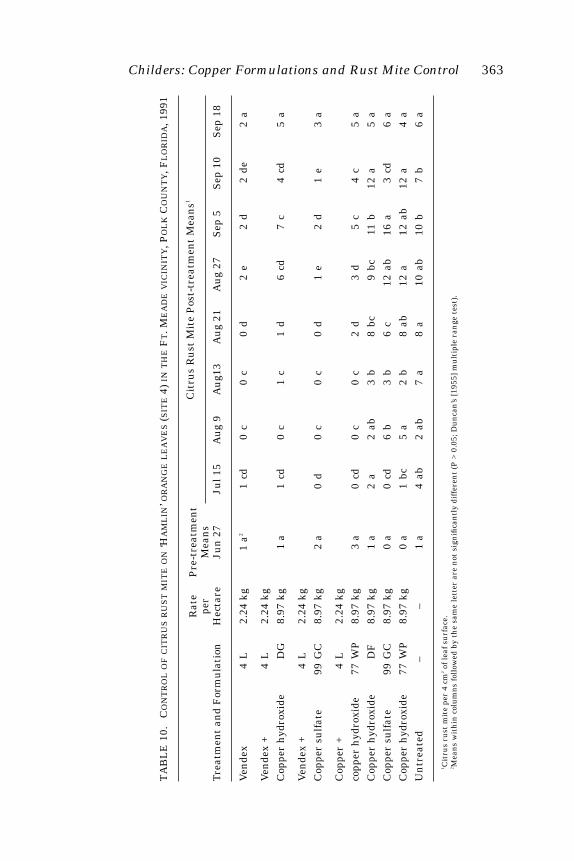
Childers: Copper Formulations and Rust Mite Control 363
1 Cit
rus
rust
mit
e pe
r 4
cm2 o
f le
af s
urf
ace.
2 Mea
ns
wit
hin
col
um
ns
foll
owed
by
the
sam
e le
tter
are
not
sig
nifi
can
tly
diff
eren
t (P
> 0
.05;
Du
nca
n’s
[19
55]
mu
ltip
le r
ange
tes
t).
TA
BL
E 1
0.
CO
NT
RO
L O
F C
ITR
US R
US
T M
ITE
ON
‘HA
ML
IN’ O
RA
NG
E L
EA
VE
S (
SIT
E 4
) IN
TH
E F
T. M
EA
DE
VIC
INIT
Y, P
OL
K C
OU
NT
Y, F
LO
RID
A, 1
991
Trea
tmen
t an
d F
orm
ula
tion
Rat
epe
rH
ecta
re
Pre
-tre
atm
ent
Mea
ns
Jun
27
Cit
rus
Ru
st M
ite
Pos
t-tr
eatm
ent
Mea
ns1
Jul 1
5A
ug
9A
ug1
3A
ug
21A
ug
27S
ep 5
Sep
10
Sep
18
Ven
dex
4L
2.24
kg
1 a2
1cd
0c
0c
0d
2e
2d
2de
2a
Ven
dex
+4
L2.
24 k
gC
oppe
r h
ydro
xide
DG
8.97
kg
1 a
1cd
0c
1c
1d
6cd
7c
4cd
5a
Ven
dex
+4
L2.
24 k
gC
oppe
r su
lfat
e99
GC
8.97
kg
2 a
0d
0c
0c
0d
1e
2d
1e
3a
Cop
per
+4
L2.
24 k
gco
pper
hyd
roxi
de77
WP
8.97
kg
3 a
0cd
0c
0c
2d
3d
5c
4c
5a
Cop
per
hyd
roxi
deD
F8.
97 k
g1
a2
a2
ab3
b8
bc9
bc11
b12
a5
aC
oppe
r su
lfat
e99
GC
8.97
kg
0 a
0cd
6b
3b
6c
12ab
16a
3cd
6a
Cop
per
hyd
roxi
de77
WP
8.97
kg
0 a
1bc
5a
2b
8ab
12a
12ab
12a
4a
Un
trea
ted
––
1 a
4ab
2ab
7a
8a
10ab
10b
7b
6a

364 Florida Entomologist 77(3) September, 1994
with copper hydroxide. No reduction in miticidal activity was recorded. However, thisrate exceeds that which is recommended in the Florida Citrus Spray Guide.
Additional studies are needed to identify those miticides currently recommendedfor use during the postbloom and summer sprays that have minimal compatibilityproblems with specific available copper formulations.
ENDNOTE
The technical assistance of Paul M. Keen, Jr., Michael G. Warmington, andDeanna K. Threlkeld is gratefully acknowledged. Florida Agricultural ExperimentStation Journal No. R-03528.
REFERENCES CITED
ALLEN, J. C. 1976. A model for predicting citrus rust mite damage on Valencia orangefruit. Environ. Entomol. 5:1083-1088.
BRUSSEL, E. W. VAN. 1975. Interrelations between citrus rust mite, Hirsutella thomp-sonii and greasy spot on citrus in Surinam. Agric. Exp. Sta. Surinam (Parama-ribo). Bull. 98.
BULLOCK, R. C., C. C. CHILDERS, J. L. KNAPP, C. W. MCCOY, AND P. A. STANSLY. 1993.Citrus rust mite and spider mites, in J. L. Knapp [ed.], Florida Citrus SprayGuide. Florida Coop. Ext. Serv., IFAS, Univ. Florida, Gainesville, SP-43.
CHILDERS, C. C. 1987. Chemical control of phytophagous mite pests on Florida citrus.Proc. Int. Conf. on Pests in Agric. Paris III, pp. 119-126.
CHILDERS, C. C., L. W. DUNCAN, T. A. WHEATON, AND L. W. TIMMER. 1987. Arthropodand nematode control with aldicarb on Florida citrus. J. Econ. Entomol.80:1064-1071.
CHILDERS, C. C., AND A. G. SELHIME. 1983. Reduced efficacy of fenbutatin-oxide incombination with petroleum oil in controlling the citrus rust mite Phyllocop-truta oleivora. Florida Entomol. 66:310-319.
DEAN, H. A. 1979. Citrus rust mite control affected by certain pesticides. J. RioGrande Valley Hort. Soc. 33:55-61.
DUNCAN, D. B. 1955. Multiple range and multiple F tests. Biometrics 11:1-42.EGER, J. E., JR., V. M. FERGUSON, AND K. G. TOWNSEND. 1985. Efficacy of selected
miticides and spray tank mixtures used to control rust mite in Florida citrus.Proc. Florida State Hort. Soc. 98:11-14.
GRIFFITH, J. T., JR., AND F. E. FISHER. 1949. Residues on Citrus trees in Florida. J.Econ. Entomol. 42:829-833.
HOLLOWAY, J. K., C. F. HENDERSON, AND H. V. MCBURNIE. 1942. Population increasesof citrus red mite associated with the use of sprays containing inert granularresidues. J. Econ. Entomol. 35:348-350.
JOHNSON, R. B. 1960a. The effect of copper compounds on control of citrus rust mitewith zineb. J. Econ. Entomol. 53:395-397.
JOHNSON, R. B. 1960b. The effect of copper on rust mite control with four rust mitemiticides. Florida State Hort. Soc. 73:84-89.
JOHNSON, R. B., J. R. KING, AND J. J. MCBRIDE, JR. 1957. Zineb controls citrus rustmite. Proc. Florida State Hort. Soc. 70:38-48.
LYE, B. H., C. W. MCCOY, AND J. FOJTIK. 1990. Effect of copper on residual efficacy ofacaricides and population dynamics of citrus rust mite (Acari: Eriophyidae).Florida Entomol. 73:230-237.
MCCOY, C. W. 1977. Horticultural practices affecting phytophagous mite populationson citrus. Proc. Int. Soc. Citriculture 2:459-462.
OOMEN, P. A. 1982. Population dynamics of the scarlet mite, Brevipalpus phoenicis, apest of tea in Indonesia. Meded. Landbouwhogeschool Wageningen. 82-1, 88 p.

Childers: Copper Formulations and Rust Mite Control 365
SAS INSTITUTE. 1991. SAS language and procedures: Usage 2, version 6, First edition.SAS Institute. Cary, North Carolina.
SEAMAN, A. J., AND H. RIEDL. 1986. Preventing decomposition of agricultural chemi-cals by alkaline hydrolysis in the spray tank. Cornell Univ., New York’s Foodand Life Science Bull. 118. 7 p.
SPENCER, H. 1939. Increases in citrus scale insect infestations from heavy residue andfrom copper spray mixtures. J. Econ. Entomol. 32:686-688.
THOMPSON, W. L. 1939. Cultural practices and their influence upon citrus pests. J.Econ. Entomol. 32:782-789.
TIMMER, L. W., AND R. MCGOVERN. 1993. Fungus diseases, fruit and/or foliage, pp 11-13 in J. L. Knapp [ed.], Florida Citrus Spray Guide. Florida Coop. Ext. Serv.,IFAS, Univ. Florida, Gainesville. SP-43.
VENKATA RAM, C. S. 1963. Application of copper and nickel fungicides with motorizedmist blowers for blister blight control. 3. Effect of yield and buildup of purplemites. Rep. Unit Plant Assoc. South India Sci. Dept. (Tea Sect.) 1962-63(Coonoor) pp. XVII-XXIV.
WHITESIDE, J. O. 1988. Greasy spot, melanose and scab diseases, pp. 15-17, 20-21, 26-27 in J. O. Whiteside, S. M. Garnsey, and L. W. Timmer [eds.], Compendium of
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
citrus diseases. APS Press, St. Paul, Minnesota.

365
INCISITERMES FURVUS
, A NEW DRYWOOD TERMITE (ISOPTERA: KALOTERMITIDAE) FROM PUERTO RICO
R
UDOLF
H. S
CHEFFRAHN
Ft. Lauderdale Research and Education CenterUniversity of Florida, Institute of Food & Agric. Sciences
3205 College Avenue, Ft. Lauderdale, FL 33314
A
BSTRACT
The imago and soldier of
Incisitermes furvus
n. sp. from western Puerto Rico aredescribed for the first time. This is the seventh species of
Incisitermes
reported fromthe West Indies.
R
ESUMEN
Se describen por vez primera el imago y el soldado de
Incisitermes furvus
sp. n. dePuerto Rico. Esta es la septima especie de
Incisitermes
registrada de las Indias Occi-dentales.
Key Words: New species description, soldier, winged imago, West Indies.
The genus
Incisitermes
was established by Krishna (1961) to accommodate the re-classification of a large subgroup of kalotermitid species previously placed together inthe genus
Kalotermes
Hagen. The known distribution of
Incisitermes
is limited to both
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

366
Florida Entomologist
77(3) September, 1994
the Nearctic and Neotropical Americas (Snyder 1949, Araujo 1977), Australia (Gay1976), India (Roonwal & Verma 1973), and islands of the Pacific Ocean (Krishna1961).
A new dark and rather small species of
Incisitermes
was collected during a recentsurvey of the termites of Puerto Rico. The imago and soldier castes of
Incisitermes fur-vus
n. sp. are described herein.
M
ATERIALS
AND
M
ETHODS
Fourteen colonies of
I. furvus
n. sp. were sampled in four locations in Puerto Ricoincluding three colonies from Reserva Forestal Maricao (18
°
09’N, 66
°
59’W) on19-V-1992 by S.C. Jones and one on 2-VI-1993 by J. R. Mangold; eight from BosqueEstatal de Guajataca (18
°
25’N, 66
°
58’W) on 3-VI-1993; one from Area Recreativa RioAbajo (18
°2
0’N, 66
°
43’W) on 4-VI-1993; and one sample from Bosque Estatal de Cam-balache (18
°
26’N, 66
°
36’W) on 4-VI-1993; the latter ten collected by J. A. Chase, J. dela Rosa Guzman, J. R. Mangold and R. H. Scheffrahn. Colonies were dwelling insidestanding and fallen native dead tree trunks and branches of sound condition. The ter-mites were collected by aspirator and field-preserved in 85% ethanol.
Measurements of specimens, made with a calibrated ocular micrometer to a max-imum accuracy of
±
0.01 mm, follow the conventions of Krishna & Emerson (1962).Thirty-four imagos (14 from Maricao, 18 from Guajataca and one each from Rio Abajoand Cambalache) and 52 soldiers (14 from Maricao, 31 from Guajataca, four from RioAbajo, and three from Cambalache) were used for measurements. Scanning electronmicrographs were made with a Hitachi S-4000 field emission microscope (10kV) ofspecimens dehydrated in absolute ethanol and 1,1,1,3,3,3-hexamethyldisilazane (Na-tion 1983) and then sputter-coated with gold.
The holotype imago from Guajataca and morphotype soldier from the Maricao site(2-VI-1993) will be deposited in the collection of the National Museum of Natural His-tory, Washington, D.C. Paratype soldiers and alates will be deposited in the AmericanMuseum of Natural History, New York, New York, in the Florida State Collection ofArthropods, Florida Department of Agriculture and Consumer Services., Division ofPlant Industry, Gainesville, Florida, and in the author’s collection.
Incisitermes furvus
, new species
IMAGO
(Figs. 1-5, Table 1). Head capsule, antennae, and notum very dark brown;wing scales and membranes blackish, costal veins including costal margin, radius,and radial sector very dark brown, in all, giving the winged imago a strikingly uni-form black appearance when viewed dorsally. Clypeus concolorous at base with headgrading to pale yellow anteriorly; labrum light brown. Head with about 16 fine setaeprojecting from vertex and frons in lateral profile. Compound eye subtriangular, com-posed of very small facets; ocellus small, oblong, and hyaline, margin sharply defined-surrounding head pigmentation, less than one half diam above eye. Antennaecomposed of 15-17 segments, usually 16; third or fourth segment shortest. Mandibledentition as in Fig. 4 and as described for
Incisitermes
by Krishna (1961); Pronotumwider than long, about as wide as head; anterior margin squarely incised to formabout a 160
°
angle, anterior corners rounded; posterior margin shallowly concave,posterior corners reduced to two rounded, about 45
°
curves; all margins with scat-tered long and short setae. Wing membrane tuberculate; median and cubitus veinsand their branches delineated by a single row of papillae smaller than those on mem-brane, costal veins densly covered with even smaller papillae. In forewing, radial sec-
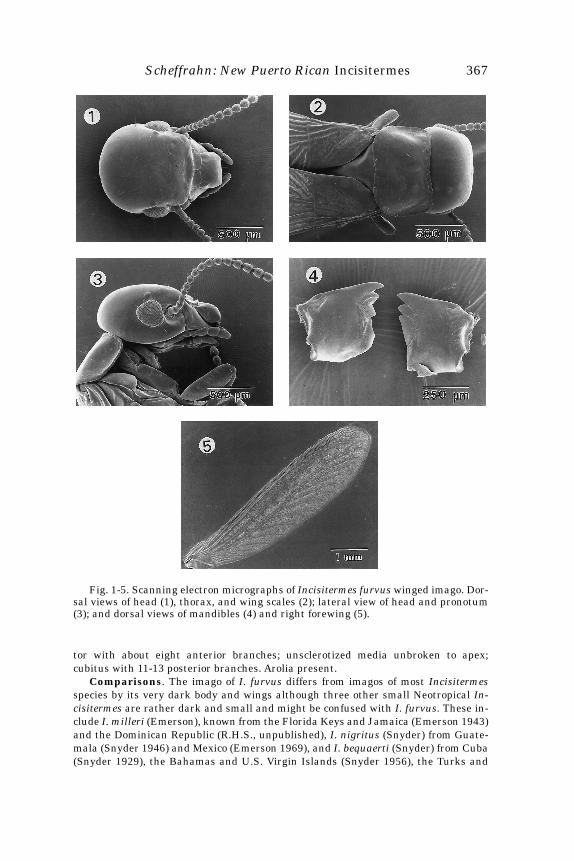
Scheffrahn: New Puerto Rican
Incisitermes 367
tor with about eight anterior branches; unsclerotized media unbroken to apex;cubitus with 11-13 posterior branches. Arolia present.
Comparisons
. The imago of
I. furvus
differs from imagos of most
Incisitermes
species by its very dark body and wings although three other small Neotropical
In-cisitermes
are rather dark and small and might be confused with
I. furvus
. These in-clude
I. milleri
(Emerson), known from the Florida Keys and Jamaica (Emerson 1943)and the Dominican Republic (R.H.S., unpublished),
I. nigritus
(Snyder) from Guate-mala (Snyder 1946) and Mexico (Emerson 1969), and
I. bequaerti
(Snyder) from Cuba(Snyder 1929), the Bahamas and U.S. Virgin Islands (Snyder 1956), the Turks and
Fig. 1-5. Scanning electron micrographs of Incisitermes furvus winged imago. Dor-sal views of head (1), thorax, and wing scales (2); lateral view of head and pronotum(3); and dorsal views of mandibles (4) and right forewing (5).

368
Florida Entomologist
77(3) September, 1994
Caicos (Scheffrahn et al. 1990), the Dominican Republic (R.H.S., unpublished), andPuerto Rico [S.C. Jones (USDA-FS, Tucson, AZ) & R.H. S. unpublished]. Table 2 givesrelevant characters to separate these species from
I. furvus
and each other. The ima-gos of
I. milleri
are smaller and have 1-4 fewer antennal segments than
I. furvus
.Wings of
I. bequaerti
are hyaline and larger and wider than
I. furvus
.
Incisitermes nig-ritus
imagos are closest to
I. furvus
but can be separated by the larger and more ro-bust bodies, larger eyes and ocelli, and more tuberiferous wing membranes of
I.furvus
.
SOLDIER
(Figs. 6-9, Table 3). Head capsule, when viewed dorsally, red-brown infront grading to pale yellow posteriorly; from above, sides parallel, anterior cornersnearly square at mandibular articulations, posterior margin evenly rounded; ventralsurface pale yellow except for reddish anterior margin. Head capsule, when viewedlaterally, with 40-50 dorsal setae of variable length to 0.13 mm; setae shorter but ofsame density on ventral surface. Frons slopes about 30
°
from vertex, weakly andbroadly concave, surface very weakly rugose. Anteclypeus a narrow, white trapezoid.Labrum light yellow-brown, spatulate. In most specimens, eyespot composed of apoorly defined hyaline ellipse having diffuse border with surrounding pigmentation;eyespot regions about same diam and standing about one-half diam behind, and atsame height, as antennal fossae. In a few specimens, a small more well-defined hya-line or gray eye rudiment visible within center of unpigmented region. Mandiblesrather slender with outer margins smooth, angled dorsally about 30
°
from plane ofhead, dentition as in Fig. 9; left mandible with long, narrow distal blade and threemarginal teeth, first two pointed forward, prominently arising in distal third of man-dible, the third tooth less prominent, arising from a shelf on the basal half; right man-dible with serrate blade in distal half and two triangulate teeth in basal half.Antennae with 10-14 segments, usually 12, rarely 10 as in Fig. 6; third longest, nar-rower than first; third as long as fourth and fifth together; in some specimens, fourthfused to third. Postmentum as in Fig. 8. Pronotum with median incision in anteriormargin forming square angle of about 135
°
; anterior corners sharply rounded; poste-rior margin with a slight or no concavity; posterior corners broadly rounded; setae onmargins of same length and density as head. Femora weakly inflated.
TABLE 1. M
EASUREMENTS
OF
I
NCISITERMES
FURVUS
IMAGO
Measurement in mm (n=34) Range Mean
±
SD Holotype
Number antennal segments 15-17 15.72
±
0.53 16Head length to labrum tip 1.25–1.48 1.37
±
0.045 1.44Head length to lateral base of mandibles 0.96–1.10 1.02
±
0.030 1.10Head maximum width at eyes 1.04–1.16 1.09
±
0.029 1.12Eye maximum diameter 0.26–0.31 0.29
±
0.015 0.30Ocellus maximum diameter 0.10–0.14 0.11
±
0.009 0.11Distance from eye to lower margin of head 0.14–0.20 0.17
±
0.016 0.16Pronotum median length 0.58–0.72 0.65
±
0.029 0.64Pronotum maximum length 0.68–0.84 0.76
±
0.036 0.76Pronotum maximum width 0.98–1.16 1.09
±
0.037 1.08Hind tibia length 0.83–0.93 0.87
±
0.030 0.93Forewing length at suture 5.12–6.42 5.83
±
0.36 5.83Forewing maximum width 1.46–1.88 1.68
±
0.12 1.69Body length without wings 4.73–6.09 5.45
±
0.33 4.92Total length with wings 7.45–8.94 8.23
±
0.49 8.23
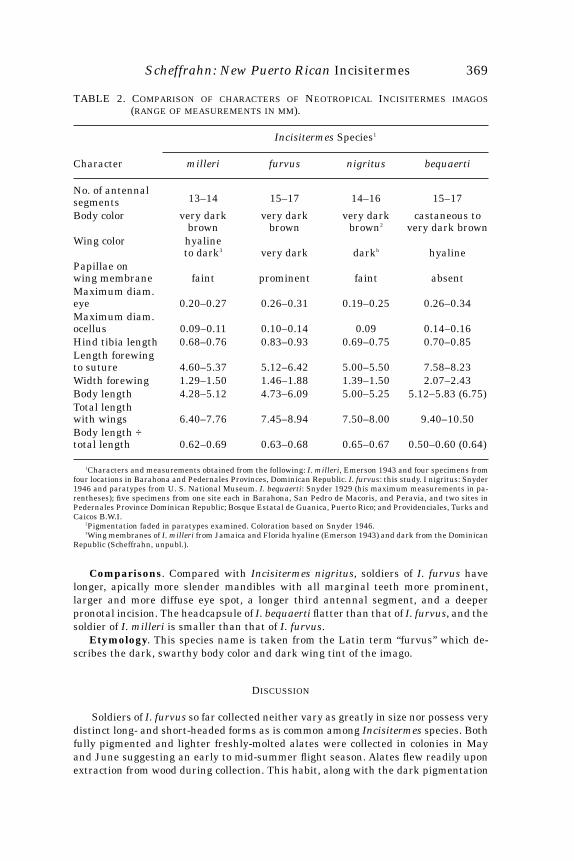
Scheffrahn: New Puerto Rican
Incisitermes 369
Comparisons
. Compared with
Incisitermes nigritus
, soldiers of
I. furvus
havelonger, apically more slender mandibles with all marginal teeth more prominent,larger and more diffuse eye spot, a longer third antennal segment, and a deeperpronotal incision. The headcapsule of
I. bequaerti
flatter than that of
I. furvus
, and thesoldier of
I. milleri
is smaller than that of
I. furvus
.
Etymology
. This species name is taken from the Latin term “furvus” which de-scribes the dark, swarthy body color and dark wing tint of the imago.
D
ISCUSSION
Soldiers of
I. furvus
so far collected neither vary as greatly in size nor possess verydistinct long- and short-headed forms as is common among
Incisitermes
species. Bothfully pigmented and lighter freshly-molted alates were collected in colonies in Mayand June suggesting an early to mid-summer flight season. Alates flew readily uponextraction from wood during collection. This habit, along with the dark pigmentation
1
Characters and measurements obtained from the following:
I. milleri
, Emerson 1943 and four specimens fromfour locations in Barahona and Pedernales Provinces, Dominican Republic.
I. furvus: this study. I nigritus: Snyder1946 and paratypes from U. S. National Museum. I. bequaerti: Snyder 1929 (his maximum measurements in pa-rentheses); five specimens from one site each in Barahona, San Pedro de Macoris, and Peravia, and two sites inPedernales Province Dominican Republic; Bosque Estatal de Guanica, Puerto Rico; and Providenciales, Turks andCaicos B.W.I.
2Pigmentation faded in paratypes examined. Coloration based on Snyder 1946.3Wing membranes of I. milleri from Jamaica and Florida hyaline (Emerson 1943) and dark from the Dominican
Republic (Scheffrahn, unpubl.).
TABLE 2. COMPARISON OF CHARACTERS OF NEOTROPICAL INCISITERMES IMAGOS(RANGE OF MEASUREMENTS IN MM).
Incisitermes Species1
Character milleri furvus nigritus bequaerti
No. of antennalsegments 13–14 15–17 14–16 15–17
Body color very darkbrown
very darkbrown
very dark brown2
castaneous to very dark brown
Wing color hyalineto dark3 very dark darkb hyaline
Papillae onwing membrane faint prominent faint absentMaximum diam. eye 0.20–0.27 0.26–0.31 0.19–0.25 0.26–0.34Maximum diam. ocellus 0.09–0.11 0.10–0.14 0.09 0.14–0.16Hind tibia length 0.68–0.76 0.83–0.93 0.69–0.75 0.70–0.85Length forewing to suture 4.60–5.37 5.12–6.42 5.00–5.50 7.58–8.23Width forewing 1.29–1.50 1.46–1.88 1.39–1.50 2.07–2.43Body length 4.28–5.12 4.73–6.09 5.00–5.25 5.12–5.83 (6.75)Total lengthwith wings 6.40–7.76 7.45–8.94 7.50–8.00 9.40–10.50Body length ÷total length 0.62–0.69 0.63–0.68 0.65–0.67 0.50–0.60 (0.64)
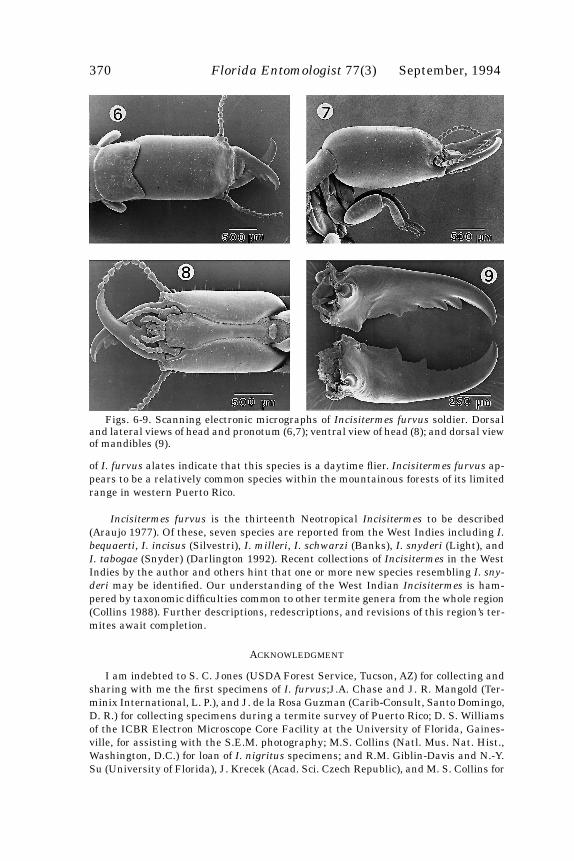
370 Florida Entomologist 77(3) September, 1994
of I. furvus alates indicate that this species is a daytime flier. Incisitermes furvus ap-pears to be a relatively common species within the mountainous forests of its limitedrange in western Puerto Rico.
Incisitermes furvus is the thirteenth Neotropical Incisitermes to be described(Araujo 1977). Of these, seven species are reported from the West Indies including I.bequaerti, I. incisus (Silvestri), I. milleri, I. schwarzi (Banks), I. snyderi (Light), andI. tabogae (Snyder) (Darlington 1992). Recent collections of Incisitermes in the WestIndies by the author and others hint that one or more new species resembling I. sny-deri may be identified. Our understanding of the West Indian Incisitermes is ham-pered by taxonomic difficulties common to other termite genera from the whole region(Collins 1988). Further descriptions, redescriptions, and revisions of this region’s ter-mites await completion.
ACKNOWLEDGMENT
I am indebted to S. C. Jones (USDA Forest Service, Tucson, AZ) for collecting andsharing with me the first specimens of I. furvus;J.A. Chase and J. R. Mangold (Ter-minix International, L. P.), and J. de la Rosa Guzman (Carib-Consult, Santo Domingo,D. R.) for collecting specimens during a termite survey of Puerto Rico; D. S. Williamsof the ICBR Electron Microscope Core Facility at the University of Florida, Gaines-ville, for assisting with the S.E.M. photography; M.S. Collins (Natl. Mus. Nat. Hist.,Washington, D.C.) for loan of I. nigritus specimens; and R.M. Giblin-Davis and N.-Y.Su (University of Florida), J. Krecek (Acad. Sci. Czech Republic), and M. S. Collins for
Figs. 6-9. Scanning electronic micrographs of Incisitermes furvus soldier. Dorsaland lateral views of head and pronotum (6,7); ventral view of head (8); and dorsal viewof mandibles (9).

Scheffrahn: New Puerto Rican Incisitermes 371
critically reviewing and improving this contribution no. R-03458 of the University ofFlorida Experiment Stations Series.
REFERENCES CITED
ARAUJO, R. L. 1977. Catálogo dos Isoptera do novo mundo. Acad. Brasileira de Ciên-cias, Rio de Janeiro, RJ. 92 pp.
COLLINS, M. S. 1988. Taxonomic problems with termites of North America, Canadathrough Panama. Sociobiology 14: 207-210.
DARLINGTON, J. P. E. C. 1992. Survey of termites in Guadeloupe, Lesser Antilles(Isoptera: Kalotermitidae, Rhinotermitidae, Termitidae). Florida Entomol. 75:104-109.
EMERSON, A. E. 1943. Kalotermes milleri, a new species of termite from the FloridaKeys and Jamaica (Isoptera, Kalotermitidae). Psyche 50: 18-22.
EMERSON, A. E. 1969. A revision of the Tertiary fossil species of the Kalotermitidae(Isoptera). American Mus. Nov. 2359: 1-57.
GAY, F. J. 1976. An Australian species of Incisitermes Krishna (Isoptera: Kalotermiti-dae). J. Australian Entomol. Soc. 14: 395-398.
KRISHNA, K. 1961. Generic revision and phylogenetic study of the family Kalotermiti-dae (Isoptera). Bull. American Mus. Nat. Hist. 122: 303-408
KRISHNA, K., AND A. E. EMERSON. 1962. New species of the Genus Glyptotermes Frog-gatt from the Papuan, Oriental, Ethiopian, and Neotropical Regions (Isoptera,Kalotermitidae). American Mus. Novitates No. 2089: 1-65.
NATION, J. A. 1983. A new method using hexamethyldisilazane for the preparation ofsoft insect tissue for scanning electron microscopy. Stain Technol. 55: 347-352.
ROONWAL, M. L., AND S. C. VERMA. 1973. First record of termite genus IncisitermesKrishna (Kalotermitidae) from Indian Region, with description of a new speciesfrom Rajasthan. Zool. Anzeiger 191: 390-397.
TABLE 3. MEASUREMENTS OF INCISITERMES FURVUS SOLDIERS.
Measurement in mm (n=52) Range Mean ± SD Morphotype
Number antennal segments 11–14 12.4 ± 0.78 13Third antennal seg. length 0.15–0.24 0.19 ± 0.021 0.19Third antennal segmentmaximum width 0.088–0.11 0.10 ± 0.006 0.10Third antennal segmentminimum width 0.050–0.069 0.060 ± 0.005 0.056Head length with mandibles 2.53–3.33 2.90 ± 0.20 2.86Head length to lateral base of mandibles 1.50–2.20 1.86 ± 0.16 1.87Left mandible length todorsal base 1.13–1.40 1.26 ± 0.068 1.33Pronotum median length 0.53–0.80 0.66 ± 0.057 0.67Pronotum maximum length 0.68–1.00 0.83 ± 0.072 0.80Head maximum width 1.03–1.33 1.20 ± 0.064 1.21Pronotum maximum width 1.00–1.35 1.20 ± 0.071 1.20Head maximum height 0.85–1.07 0.96 ± 0055 0.95Postmentum maximum width 0.44–0.58 0.51 ± 0.030 0.51Postmentum minimum width 0.16–0.27 0.21 ± 0.021 0.22Postmentum length 1.12–1.78 1.44 ± 0.15 1.44Hind tibia length 0.70–0.96 0.85 ± 0.063 0.88

372 Florida Entomologist 77(3) September, 1994
SCHEFFRAHN, R. H., N.-Y. SU, AND B. DIEHL. 1990. Native, introduced, andstructure-infesting termites of the Turks and Caicos Islands, B.W.I. (Isoptera:Kalotermitidae, Rhinotermitidae, Termitidae). Florida Entomol. 73: 622-627.
SNYDER, T. E. 1929. New termites from the Antilles and Middle America. Proc. Ento-mol. Soc. Washington 31: 79-87.
SNYDER, T. E. 1946. A small, dark-colored new Kalotermes from Guatemala. Proc. En-tomol. Soc. Washington 48: 158-160.
SNYDER, T. E. 1949. Catalog of the termites (Isoptera) of the world. Smithsonian Misc.Coll. 122: 1-490.
SNYDER, T. E. 1956. Termites of the West Indies, the Bahamas, and Bermuda. J. Ag-ric. Univ. Puerto Rico 40: 189-202.

Scientific Notes
373
FECUNDITY AND FERTILITY OF
RHYNCHOPHORUS CRUENTATUS
(COLEOPTERA: CURCULIONIDAE)
T
HOMAS
J. W
EISSLING
1
AND
R
OBIN
M. G
IBLIN-
D
AVIS
University of Florida, Institute of Food andAgricultural Sciences, Ft. Lauderdale Research and Education
Center, 3205 College Ave., Ft. Lauderdale, FL 33314
The palmetto weevil (
R
.
cruentatus
F.) breeds in a variety of stressed or dyingpalms (Giblin-Davis & Howard 1988, 1989). These large (1.9 - 3.0 cm long) weevils areassociated with the native cabbage palmetto,
Sabal
palmetto
(Walter), in Florida(Woodruff 1967). Semiochemicals emanating from stressed or dying palms and maleconspecifics (Weissling et al. 1992, 1993, 1994, Giblin-Davis et al. 1994) are attractiveto
R
.
cruentatus
adults. Although not fully understood, mating apparently takes placeon dying palms and females lay their eggs in the leaf bases or directly into the woundsof the host. Larvae develop primarily in the crown region but can occasionally befound in the stem tissue. Last instar larvae migrate to the fibrous stem periphery orpetiolar bases and construct cocoons from fiber (Giblin-Davis & Howard 1988).
Research on an improved method to culture
R
.
cruentatus
has required the collec-tion of a large number of eggs to produce neonate larvae for evaluation of diets. Usingpineapple,
Anana comosus
(L.), as an ovipositional substrate, Giblin-Davis et al.(1989) reported the mean lifetime fecundity of field-collected females as 26
±
15 eggsper female. However, pineapple proved to be a difficult media to dissect for removal ofeggs. More suitable ovipositional substrates were investigated and we found that ap-ple (
Pyrus malus
L.) slices were easily dissected and were readily accepted by
R
.
cruentatus
females. Using apple slices, we reinvestigated the fecundity of
R
.
cruenta-tus
females and determined fertility.Cocoons were harvested in the field from infested palms or in the laboratory from
sugarcane (
Saccharum officinarum
L.) stem (Giblin-Davis et al. 1989). Cocoons wereplaced individually in covered 100-ml plastic cups with moistened tissue paper(Giblin-Davis et al. 1989) and were stored at 29
o
C until adult emergence. One maleand one female were placed in a 500-ml covered plastic container with moistened tis-sue. After 48-72 h, males were removed and a slice of apple (‘Red Delicious’) wasadded. Slices were thin (5-15 mm; 10-18 g wet weight), convex segments covered bypeel. Females oviposit through the apple pulp and most eggs are found along the peel.All containers were placed in an environmental chamber at 29
o
C with a 13:11 (L:D)photoperiod. Apple slices were usually replaced every 1-3 days until female death.Slices removed from containers were carefully dissected and eggs were removed. Thistest was repeated five times with four or five females per test (22 females total). Datawere converted to the number of eggs laid per female per week. In addition, the totalnumber of eggs laid per female was determined. During two of the tests, eggs removedfrom apple slices were placed in 15 x 100 mm plastic petri dishes lined with water-moistened filter paper, sealed with parafilm, and placed in the environmental cham-ber. Neonate larvae were removed from each dish at daily intervals and the dish re-sealed until all eggs hatched or decomposed. For the first test, fertility wasdetermined every one to three days for 45 d. During the second test, eggs were col-
1
Current address: Yakima Agricultural Research Laboratory, USDA-ARS, 3706 W. Nob Hill Blvd., Yakima, WA98902
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.
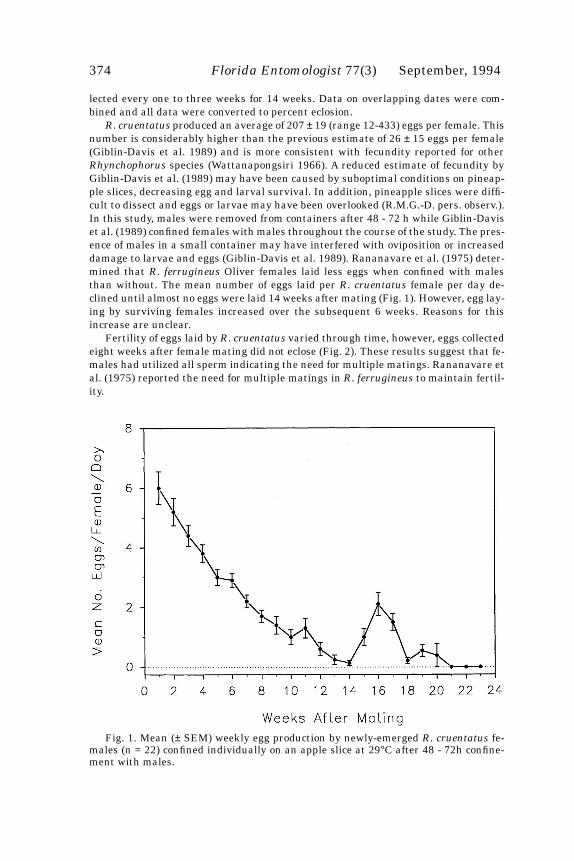
374
Florida Entomologist
77(3) September, 1994
lected every one to three weeks for 14 weeks. Data on overlapping dates were com-bined and all data were converted to percent eclosion.
R
.
cruentatus
produced an average of 207
±
19 (range 12-433) eggs per female. Thisnumber is considerably higher than the previous estimate of 26
±
15 eggs per female(Giblin-Davis et al. 1989) and is more consistent with fecundity reported for other
Rhynchophorus
species (Wattanapongsiri 1966). A reduced estimate of fecundity byGiblin-Davis et al. (1989) may have been caused by suboptimal conditions on pineap-ple slices, decreasing egg and larval survival. In addition, pineapple slices were diffi-cult to dissect and eggs or larvae may have been overlooked (R.M.G.-D. pers. observ.).In this study, males were removed from containers after 48 - 72 h while Giblin-Daviset al. (1989) confined females with males throughout the course of the study. The pres-ence of males in a small container may have interfered with oviposition or increaseddamage to larvae and eggs (Giblin-Davis et al. 1989). Rananavare et al. (1975) deter-mined that
R
.
ferrugineus
Oliver females laid less eggs when confined with malesthan without. The mean number of eggs laid per
R
.
cruentatus
female per day de-clined until almost no eggs were laid 14 weeks after mating (Fig. 1). However, egg lay-ing by surviving females increased over the subsequent 6 weeks. Reasons for thisincrease are unclear.
Fertility of eggs laid by
R
.
cruentatus
varied through time, however, eggs collectedeight weeks after female mating did not eclose (Fig. 2). These results suggest that fe-males had utilized all sperm indicating the need for multiple matings. Rananavare etal. (1975) reported the need for multiple matings in
R
.
ferrugineus
to maintain fertil-ity.
Fig. 1. Mean (± SEM) weekly egg production by newly-emerged R. cruentatus fe-males (n = 22) confined individually on an apple slice at 29°C after 48 - 72h confine-ment with males.
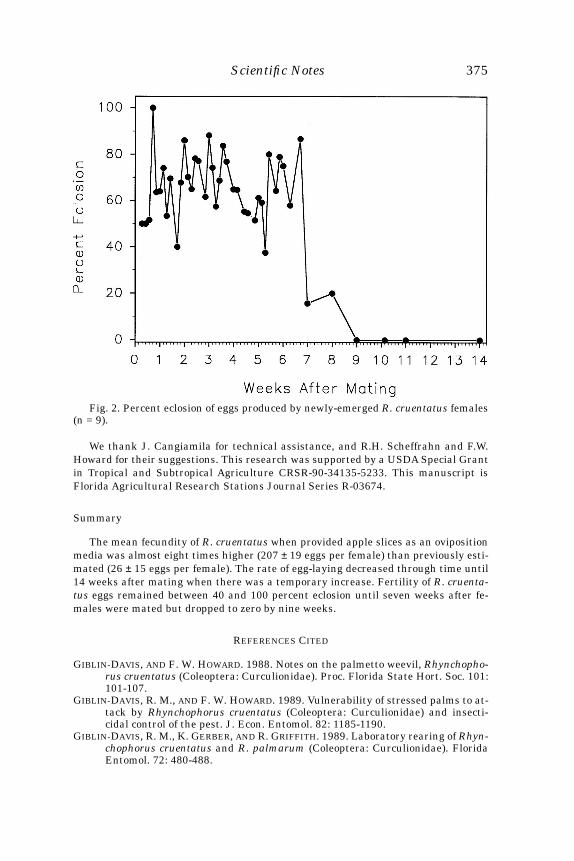
Scientific Notes
375
We thank J. Cangiamila for technical assistance, and R.H. Scheffrahn and F.W.Howard for their suggestions. This research was supported by a USDA Special Grantin Tropical and Subtropical Agriculture CRSR-90-34135-5233. This manuscript isFlorida Agricultural Research Stations Journal Series R-03674.
Summary
The mean fecundity of
R
.
cruentatus
when provided apple slices as an ovipositionmedia was almost eight times higher (207
±
19 eggs per female) than previously esti-mated (26
±
15 eggs per female). The rate of egg-laying decreased through time until14 weeks after mating when there was a temporary increase. Fertility of
R
.
cruenta-tus
eggs remained between 40 and 100 percent eclosion until seven weeks after fe-males were mated but dropped to zero by nine weeks.
R
EFERENCES
C
ITED
G
IBLIN-
D
AVIS
,
AND
F. W. H
OWARD
. 1988. Notes on the palmetto weevil,
Rhynchopho-rus
cruentatus
(Coleoptera: Curculionidae). Proc. Florida State Hort. Soc. 101:101-107.
G
IBLIN-
D
AVIS
, R. M.,
AND
F. W. H
OWARD
. 1989. Vulnerability of stressed palms to at-tack by
Rhynchophorus
cruentatus
(Coleoptera: Curculionidae) and insecti-cidal control of the pest. J. Econ. Entomol. 82: 1185-1190.
G
IBLIN-
D
AVIS
, R. M., K. G
ERBER
,
AND
R. G
RIFFITH
. 1989. Laboratory rearing of
Rhyn-chophorus
cruentatus
and
R
.
palmarum
(Coleoptera: Curculionidae). FloridaEntomol. 72: 480-488.
Fig. 2. Percent eclosion of eggs produced by newly-emerged R. cruentatus females(n = 9).

376
Florida Entomologist
77(3) September, 1994
G
IBLIN-
D
AVIS
, R. M., T. J. W
EISSLING
, A. C. O
EHLSCHLAGER
,
AND
L. M. G
ONZALEZ
.1994. Field response of
Rhynchophorus cruentatus
(F.) (Coleoptera: Curculion-idae) to its aggregation pheromone and fermenting plant volatiles. Florida En-tomol. 77: 164-177.
R
ANANAVARE
, H. D., K. S
HANTARUM
, M. R. H
ARWALKAR
,
AND
G. W. R
AHLKAR
. 1975.Method of laboratory rearing of red palm weevil,
Rhynchophorus ferrugineus
Oliv. J. Plantation Crops 3: 65-67.W
ATTANAPONGSIRI
, A. 1966. A revision of the genera
Rhynchophorus
and
Dynamis
(Coleoptera: Curculionidae). Dept. Agric. Science Bulletin, Bangkok 1: 1-328.W
EISSLING
, T. J., R. M. G
IBLIN-DAVIS, R. H. SCHEFFRAHN, AND N. M. MENDOZA. 1992.Trap for capturing and retaining Rhynchophorus cruentatus (Coleoptera: Cur-culionidae) adults using Sabal palmetto as bait. Florida Entomol. 75: 212-221.
WEISSLING, T. J., R. M. GIBLIN-DAVIS, AND R. H. SCHEFFRAHN. 1993. Laboratory andfield evidence for male-produced aggregation pheromone in Rhynchophoruscruentatus (F.) (Coleoptera: Curculionidae). J. Chem. Ecol. 19: 1195-1203.
WEISSLING, T. J., R. M. GIBLIN-DAVIS, G. GRIES, R. GRIES, A. L. PEREZ, H. D. PIERCE,JR., AND A. C. OEHLSCHLAGER. 1994. Aggregation pheromone of palmetto wee-vil Rhynchophorus cruentatus (Coleoptera: Curculionidae). J. Chem. Ecol. 20:505-515.
WOODRUFF, R. E. 1967. A giant palm weevil, Rhynchophorus cruentatus (Fab.), inFlorida (Coleoptera: Curculionidae). Florida Dept. Agr. Div. Plant Industry. En-
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
tomology Circ. No. 63.

Scientific Notes
376
NEW SYNONYMY IN THE GENUS
POLYTES
STÅL(HETEROPTERA: SCUTELLERIDAE)
J. E. E
GER
, J
R
.DowElanco
Ste. 780, One MetroCenter4010 Boy Scout Boulevard
Tampa, FL 33607and
Florida State Collection of ArthropodsFlorida Department of Agriculture and Consumer Products
Gainesville, FL 32602
In a revision of the genus
Polytes
Stål (Eger 1990), I overlooked two previously de-scribed species. One of these species was originally described in the genus
Pachycoris
Burmeister, the type of which I examined recently while researching the latter genus.The other was an oversight of a species described in
Polytes
. The purpose of this paperis to rectify those errors. Measurements are given in mm.
Polytes discrepans
(Uhler 1875)
New Combination
Pachycoris discrepans
Uhler 1875:282-283.
DIAGNOSIS. Dorsum black with red markings as follows: 4 evenly spaced mac-ules on pronotum (lateral 2 oval, mesial 2 elongate, extending from near posterior
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Scientific Notes
377
pronotal margin to just anterad of cicatrices); cicatrices; two oval macules near lateralmargins on anterior half of scutellum. Dorsal punctation concolorous, dense, mostdense laterally. Venter entirely black, coxae and rostrum slightly lighter in color thanremainder of venter. Posterior margin of last abdominal sternite lacking colored lineor macule. Length 8.9.
Head very broadly rounded anteriorly; 2.1 long, 2.5 wide across eyes. Length of an-tennal segments 1-4 (5 missing): 0.6; 0.4; 0.5; 1.0, respectively.
Pronotum 2.7 long mesially, 5.4 wide across humeri. Anterolateral marginsslightly convex. Scutellum 5.4 long, 5.0 broad at widest point. Pleura unmarked,densely punctate. Venter of abdomen densely punctate, less so mesially and on stri-ated areas.
Posterior margin of genital cup shallowly concave mesially, convex laterally. Ven-trally exposed portion of cup densely punctate.
TYPE MATERIAL. Holotype: male, labeled: (a) Peru, Orton. (b) PR Uhler Collec-tion. (c)
Pachycoris discrepans
, Peru, Uhler. The type is deposited in the National Mu-seum of Natural History, Smithsonian Institution, Washington, D. C.
Polytes discrepans
resembles a number of other species in the
P. fenestra
Breddinspecies group (
P. bullocki
Eger,
P. bicolor
Distant,
P. lattini
Eger,
P. leopardinus
Dis-tant,
P. speculiger
Breddin, and
P. ruth
Breddin) in having a broadly rounded head. Itdiffers from all of these except
P. ruth
in lacking a large bilobed mesial macule on theposterior margin of the last abdominal sternite. There is a thin pale line on the pos-terior margin of this sternite in
P. ruth
, but
P
.
discrepans
has no pale markings on thissternite.
Polytes ruth
is also larger and is very differently colored than
P. discrepans
.The dorsal coloration of
P. discrepans
is most similar to that of
P. lattini
, but the headof
P. discrepans
is distinctly more broadly rounded than that of
P. lattini
, and thepronotal vittae are longer than those of
P
.
lattini
.Because
P. discrepans
appears to be distinct from all known congeners, I opted notto dissect the unique male holotype.
Polytes discrepans
will key to couplet 12 in the key by Eger (1990), but does not fiteither option given for this couplet. The following changes to the key will allow recog-nition of
P. discrepans
:
Polytes speculiger
Breddin 1914
12(11). Head broadly rounded anteriorly............................................................. 12’Head narrowly rounded anteriorly ...........................................................13
12’(12). Pronotum with two large yellow to red macules, each with distinct brownto black lanceolate intrusion from posterior margin; larger, 9.5-10.9 long
.............................................................................................P. ruth
(Breddin)Pronotum with four evenly spaced macules; smaller, 8.9 long....................
......................................................................................P. discrepans
(Uhler)
Polytes speculiger
Breddin 1914:53-54.
Polytes quechuus
Eger 1990:137-139, figs. 104-109.
New Synonymy.
I overlooked Breddin’s paper when researching the
Polytes
revision. His descrip-tion clearly indicates the presence of an impunctate area on the pronotum, a diagnos-tic character for this species, and leaves little doubt that my species is a juniorsynonym of his. I have not been able to locate Breddin’s types although one of theparatypes of
P. quechuus
was a syntype of
P. fenestra
Breddin, 1903, and was cer-

378
Florida Entomologist
77(3) September, 1994
tainly seen by him. It bore no labeling to indicate that it was a type of
P
.
speculiger
,though. Breddin also described two varieties (as ‘var. a’ and ‘var. b’) and suggestedthat these may be synonymous with
P. bicolor
and
P. leopardinus
. The color forms hedescribed were also seen by me and are apparently typical for
P. speculiger
.I thank Drs. R. C. Froeschner (Smithsonian Institution) and T. J. Henry (USDA-
ARS) for the loan of the type of
Pachycoris discrepans
and Drs. L. H. Rolston (Louisi-ana State University) and D. A. Rider (North Dakota State University) for assistancein locating literature for this study.
S
UMMARY
Corrections to a revision of the genus
Polytes
are presented.
Pachycoris discrepans
Uhler is transferred to
Polytes
with a diagnosis and revised key to allow recognitionof this species.
Polytes quechuus
Eger is placed in the synonymy of
P
.
speculiger
Bred-din.
L
ITERATURE
C
ITED
B
REDDIN
, G. 1903. Neue Rhynchoten aus den Anden. Societas Entomologica 18:122-124.
B
REDDIN
, G. 1914. Neue odor wenig gekannte neotropische Hemiptera. Abh. Senck-enb. Naturforsch. Gesellschaft 36:53-59.
E
GER
, J. E., J
R
. 1990. Revision of the genus
Polytes
Stål (Heteroptera: Scutelleridae).Ann. Entomol. Soc. America 83(2):115-141.
U
HLER
, P. R. 1874 [1875]. List of the species of Hemiptera and Neuroptera obtainedby Prof. James Orton, in Northern Peru. Proc. Boston Soc. Nat. Hist. 17:282-
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
286.

378
Florida Entomologist
77(3) September, 1994
FIRST RECORDS OF ENTOMOPATHOGENIC DISEASES IN THE PARAGUAY TEA AGROECOSYSTEM IN ARGENTINA
D
ANIEL
R. S
OSA-
G
ÓMEZ
1
, E
LLIOT
W. K
ITAJIMA
2
AND
M
ARCELO
E. R
OLON
3
1
Centro Nacional de Pesquisa da Soja, EMBRAPA - Caixa Postal 1061, 86001-970, Londrina, PR, Brasil
2
Dept. Biol. Cel., Univ. Brasilia, 70919-970, Brasilia, DF, Brasil
3
Establecimiento “Las Marías”, 3342 Gobernador Virasoro, Corrientes, Argentina.
The northeastern region of Argentina leads that country in the production of Par-aguay tea (
Ilex paraguariensis
Saint Hilarie) with 170 metric tons per year. The mostimportant pests of tea in this region are: a psyllid
Metaphalara spegazziniana
(Lizer)(Homoptera: Psyllidae), the “paraguay tea hornworm”
Pengonia lusca ilus
Bsd.(Lepidoptera: Sphingidae), and the gall bud mite
Dichopelnus
notus
Keifer (Acarina:Eriophyid)].
M. spegazziniana
is usually controlled by foliar sprays with dimethoate
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Scientific Notes
379
directed against adults to prevent damage by feeding and oviposition.
P. lusca ilus
iscontrolled mainly by pyrethroids.
The use of microbial control is a potentially valuable alternative to the high costs,possible pest resurgence, development of resistance, and environmental contamina-tion associated with chemical insecticides. Thus, as a first step toward the develop-ment of a biocontrol program, an investigation was begun to find natural enemies ofthese pests in field-collected individuals. This note reports the finding of a fungus inthe psyllid and a baculovirus in the “paraguay tea hornworm”. These species areknown by the common names “psílido de la yerba mate” and “marandová de la yerbamate”, respectively.
Sampling was done by randomly collecting the terminal shoot of paraguay teaplants from groves in Gobernador Virasoro, Corrientes, Argentina. Cadavers of thepsyllid were placed in humid chambers to induce the fungus to sporulate. After thisthey were mounted in Hoyer’s medium and observed under a stereomicroscope at 400magnification. Percent disease estimates were made by counting cadavers with myce-lium and sporulating structures. The conidial stage of the fungus was observed dur-ing March, April and early May of 1991 and 1992. A high prevalence (82% mortality,n=28) of the disease caused by the fungus was observed in the first week of May,1991.In 1992, the infection rate on April 24, May 11 and July 10 was, respectively, 30%(n=10), 93% (n=30) and 2% (n=88). We identified the fungus as
Zoophthora radicans
(Brefeld) Batko based on the descriptions of Ben-Zeév & Kenneth (1981), Balazy(1986) and by careful comparisons of morphology and cultural characters with a spec-imen from a culture collection (ARSEF 2282). This fungus has potential value as a bi-ological control agent, and basic knowledge about it’s production process is available(McCabe & Soper, 1985).
In the summer of 1988, some larvae (1% mortality, n=102) of
P. lusca ilus
displayedsymptoms typical of viral infection – the larvae changed color from typical green topale green, they fed less, and after death they were found hanging from the branchesof paraguay tea plants, attached by their anus and legs (Fig. 1). To confirm viral eti-ology, we examined hemolymph under the light microscope. This revealed large num-bers of polyhedra-like particles. The suspected viral polyhedra were concentrated byhomogenization of larval tissue, filtered through cheese cloth, and centrifuged at low(120 g for 2 min) and high speeds (6,000 g for 15 min). The pellet resulting from high-speed centrifugation was fixed in a modified Karnovsky fixative (2% glutaraldehyde,2% paraformaldehyde in 0.05 M cacodylate buffer, pH 7.2), postfixed in 1% osmiumtetroxide, dehydrated in acetone, and embedded in Spurr low viscosity medium.Blocks were sectioned in a LKB Ultratome III microtome equipped with a diamondknife, and the sections were stained with uranyl acetate and lead citrate before beingexamined in a JEOL JEM 100C electron microscope. Also, the suspension of sus-pected viral polyhedra was applied directly onto a specimen holder, air-dried, sputter-coated with gold in a Balzer’s sputter coater, and examined in a JEOL 840A scanningelectron microscope.
Electron microscopic examination demonstrated that the particles were typicalbaculovirus polyhedra containing large numbers of singly embedded rod-shaped viri-ons (Fig. 2). Scanning electron microscopy revealed large numbers of polyhedralstructures as the sole component of the nuclear polyhedra suspension (Fig. 3). Thesepolyhedra measured 1-3 micrometers in diam, matching in size the polyhedra seen bytransmission electron microscopy.
A crude preparation of this virus has been successfully used by farmers to control
P. lusca ilus
larvae, thus avoiding defoliation. This was achieved in a preliminarystudy conducted by farmers over approximately 900 ha in Gobernador Virasoro

380
Florida Entomologist
77(3) September, 1994
county, in the province of Corrientes, Argentina in February of 1992. A large amountof the virus was obtained by collecting larvae from artificially-infested fields andstored frozen for use in the subsequent season. The virus was applied by airplane us-ing the hemolymph from 15 infected last instar larvae per ha although, at that time,the etiology of the disease was not well understood. Cadavers were collected for usethe next year. In January of 1993, the treated area reached 2,362 ha. This baculovirusis currently used empirically without the benefit of prior research on dosage, timing,and population levels.
We thank Dr. James R. Fuxa of Louisiana State University for the critical manu-script review.
S
UMMARY
We report for the first time natural and artificial epizootics of pest populations onParaguay tea in Corrientes Province, Argentina. The natural epizootic was caused by
Zoophthora radicans
on
Metaphalara spegazziniana
and the artificial epizootic wascaused by a nuclear polyhedrosis virus on
Pengonia lusca ilus.
Fig. 1. Diseased larvae of Pengonia lusca ilus with symptoms typical of viral infec-tion.
Scientific Notes
381
Fig. 2. Transmission electron micrographs of thin sections of polyhedral inclusionbodies of the virus. Note that the m=nucleocapsids are wrapped individually by themembrane characterizing a SNPV (x 40,000)

382
Florida Entomologist
77(3) September, 1994
R
EFERENCES
C
ITED
B
ALAZY
, S. 1986. Taxonomic criteria for inter- and intraspecific differentiation in theentomophthoraceae, exemplified by the subgenus
Zoophthora
, pp. 201-205
in
R.A. Samson, J.M. Vlak and D. Peters [eds.] Fundamental and Applied aspectsof Invertebrate Pathology. Foundation 4th International Colloquium.
B
EN
-Z
EÉV
, I.,
AND
R. G. K
ENNETH
. 1981.
Zoophthora radicans
and
Zoophthora petchi
sp. nov. [Zygomycetes: Entomophthorales], two species of “Sphaerospermagroup” attacking leaf-hoppers and frog-hoppers [Hom.]. Entomophaga. 26: 131-142.
♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦♦
M
C
C
ABE
, D.,
AND
R. S. S
OPER
. 1985. US Patent 4530834 (July,23, 1985).
Fig. 3. Scanning electron micrographs of purified polyhedral inclusion bodies ofthe virus.

382
Florida Entomologist
77(3) September, 1994
BEE FLIES OF THE BRITISH VIRGIN ISLANDS(DIPTERA: BOMBYLIIDAE)
N
EAL
L. E
VENHUIS
AND
S
COTT
E. M
ILLER
Bishop Museum, Box 19000-A, Honolulu, HI 96817-0916
Heretofore no bee flies have been recorded from the British Virgin Islands (BVI)(Evenhuis, 1983, 1992). Survey work for other insects in the BVI by The ConservationAgency from 1984 to 1993 has recorded 6 species of bee flies from a total of 9 islands.The intensity of survey has varied considerably from island to island. Guana Island
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

Scientific Notes
383
has been extensively surveyed, while some islands were visited only briefly (e.g., Eus-tacia, Necker, Scrub).
Fieldwork was based on Guana Island and many specimens were collected in Mal-aise traps at North Bay. Guana is a small island on the north side of Tortola in theBritish Virgin Islands (18
°
28’N, 64
°
35’W). While it is small, only 297 ha, and the max-imum elevation is 266 m, it bears a relatively rich biota and has sustained less dam-age by feral animals and man than have many adjacent islands (Becker & Miller,1992). Scott E. Miller and collaborators were in the field VII-1984, VII-1985, VII-1986,VII-1987, VII-1988, and X-XI-1990. Roy R. Snelling was in the field X-1991, X-1992,IV-1993, and VI-VII-1993. Material is split between the Bishop Museum and Smith-sonian Institution, with synoptic vouchers in the Natural History Museum of Los An-geles County and The Natural History Museum (London). A few additional recordswere provided from the private collections of Michael Ivie (Norman Island) and Rich-ard Miller.
The material available is summarized in Table 1. All the BVI species are knownfrom Puerto Rico, and all but
Exoprosopa cubana
are known from the U.S. Virgin Is-lands (Curran, 1928, 1931). Most of these are also more widespread in the West In-dies. Despite recent collecting in the BVI and U.S. Virgin Islands,
Chrysanthax nero
Fabricius remains known only from the type described from “Americae meridionalisinsulis” (either St. Thomas or St. Croix). Species known from Puerto Rico and theSouth American continent, but not yet recorded from the BVI include:
Anthrax insu-lans
Marston,
Poecilanthrax lucifer
Fabricius, and
Ligyra cerberus
Fabricius. We be-lieve that with further collecting in the BVI, these species may also be found.
Field work by Miller and Snelling was supported by The Conservation Agency(Rhode Island), through a grant from the Falconwood Corporation (New York). Wethank Michael Ivie, Tina M. Kuklenski, James D. Lazell, Richard Miller, and Roy R.Snelling for assistance in obtaining specimens.
S
UMMARY
Six species of bee flies are known from the British Virgin Islands, including recordsfrom 9 islands. The BVI fauna is a subset of that of the Puerto Rican Bank.
TABLE 1. D
ISTRIBUTION
OF
BOMBYLIDAE
IN
THE
B
RITISH
V
IRGIN
I
SLANDS
.
An
egad
a
Eu
stac
ia
Gre
at C
aman
oe
Gu
ana
Nec
ker
Nor
man
Scr
ub
Tort
ola
Vir
gin
Gor
da
Heterostylum ferrugineum
(Fabr., 1805) x
Anthrax oedipus
Fabr., 1805 x x
Chrysanthrax gorgon
(Fabr., 1805) x x x x x
Neodiplocampta roederi
(Curran, 1931) x x x
Villa lateralis
(Say, 1825) x x x x x x x x
Exoprosopa cubana
Loew, 1869 x

384
Florida Entomologist
77(3) September, 1994
R
EFERENCES
C
ITED
B
ECKER
, V. O.,
AND
S. E. M
ILLER
. 1992. The butterflies of Guana Island, British Vir-gin Islands. Bull. Allyn Mus. 136: 1-9.
C
URRAN
, C. H. 1928. Diptera. Scientific Survey of Porto Rico and the Virgin Islands11: 1-118. New York Academy of Sciences.
C
URRAN
, C. H. 1931. First supplement to the ‘Diptera of Porto Rico and the Virgin Is-lands’. American Mus. Novitates 456: 1-23.
E
VENHUIS
, N. L. 1983. An indexed bibliography of Bombyliidae (Insecta, Diptera).Theses Zoologicae 4: 1-493.
E
VENHUIS
, N. L. 1992. An indexed bibliography of Bombyliidae (Insecta, Diptera):Supplement I. Bishop Mus. Tech. Rep. 2: 1-136.

Book Review
385
BOOK REVIEW
B
EDDING
, R. A., R. J. A
KHURST
,
AND
H. K. K
AYA
. (eds.). 1993. Nematodes and thebiological control of insects. CSIRO; East Melbourne. vi + 178 p. Hardback. ISBN 0-643-05479-0. (Available from ISBS, 5602 NE Hassalo St., Portland, OR 97213-3640 atUS $60.00 plus shipping).
Following closely on the heels of the successful (and out-of-print) CRC Press texton entomopathogenic nematodes (Gaugler, R. and H. K. Kaya. 1990. Entomopatho-genic Nematodes in Biological Control. CRC Press, Boca Raton, Florida) is this con-tribution from Australia’s Commonwealth Scientific and Industrial ResearchOrganization (CSIRO). Unlike its predecessor, which dealt exclusively with ento-mopathogenic nematodes in the families Steinernematidae and Heterorhabditidae,this text also contains information on species in the families Phaenopsitylenchidae,Aphelenchoididae, and Mermithidae which are considered insect-parasitic. Somemembers of these families eventually kill their host and they have been used prima-rily as classical biological control agents. In contrast, entomopathogenic nematodeskill their hosts rapidly and are used as biological insecticides.
The book’s 25 contributors hail from seven nations and they offer a diversity ofsubject matter. Seven chapters provide case studies of pest insect control with nema-todes in Australia, Korea, the United States and China. One chapter each is devotedto Phaenopsitylenchidae, Aphelenchoididae and Mermithidae and the remaining fourdeal with steinernematids and heterorhabditids. The account of the phaenopsity-lenchid
Daledenus siricidicola
and its use as a control agent for woodwasps makes forthe most interesting reading of the entire book. This parasitic nematode was the firstemployed successfully against an insect pest. Its history as detailed here provides in-sights into the preparation for, and implementation of, a successful biological controlproject including the selection of correct strains, continued monitoring after release toassess effectiveness, and maintenance of lab colonies to ensure viable control agentstock. Other chapters of note describe the control efforts for the banana weevil borerand for scarab grubs in turf (the latter providing an excellent literature review). Un-fortunately, some chapters are superficial and provide very little more informationthan in a typical scientific journal paper.
The remaining ten chapters pertain to the study and use of steinernematids andheterorhabditids, and cover subjects of a more general nature including post-applica-tion biology, ecological genetics, integrated control, and bacterial symbionts. Threechapters are devoted to bacterial symbionts (the “power behind the throne” as one au-thor describes them) which provide the toxin for target hosts and essential nutrientsfor the nematodes. As with those of case studies, the form and information content ofthese chapters vary somewhat. Most are engaging and informative, but a few arequite dry, narrow in scope, and almost painful to read.
Due primarily to their increasing importance as commercially-available biologicalcontrol agents and their relative ease of rearing, research on steinernematids andheterorhabditids has proliferated in the past few years, which explains why the greatmajority of the book is devoted to them. Much of what is presented was not availableor known at the writing of the Gaugler and Kaya text.
The layout of the book is attractive and functional. Each chapter begins with anabstract (summary) and the top of each right-hand page contains the chapter title.The paper is of medium-heavy weight and is semi-glossy. The contributing authors’affiliations are listed as an addendum glued to the last page.
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

386
Florida Entomologist
77(3) September, 1994
Moderately priced, this text would be a valuable (and affordable) addition to the li-brary of anyone studying insect-parasitic or entomopathogenic nematodes or thosewishing to stay abreast of this increasingly-important group of biological controlagents.
Patrick ParkmanEntomology & Nematology Dept.University of FloridaGainesville, FL 32611-0620

I
N
M
EMORIAM
387
I
N
M
EMORIAM
H
ARRY
R. G
ROSS
, J
R
.
(1939-1994)
Harry R. Gross, Jr., was born in New Orleans, LA, on March 16, 1939, and passedaway on May 3, 1994 in Wesley Woods Hospital in Decatur, GA. He is survived by hiswife, Marlyne Adam Gross of Tifton, and a daughter, Lisa Gross of Atlanta, GA. Dr.Gross was a Supervisory Research Entomologist and Research Leader of the InsectBiology/Management Systems Research Unit, USDA-ARS, Insect Biology and Popu-lation Management Research Laboratory at Tifton, GA.
He graduated from Louisiana State University, obtaining his B.S. (1960), M.S.(1964), and Ph.D. (1967) degrees in Entomology. Throughout his career, Dr. Gross wasemployed by the U.S. Department of Agriculture, Agricultural Research Service. Hebegan his research career in 1967 at Gulfport, MS, working on the white-fringed bee-tle before transferring in 1971 to the Insect Biology and Population Management Re-search Laboratory (formerly Southern Grain Insects Research Laboratory), Tifton,GA. Dr. Gross was a member of Sigma Xi, the Entomological Society of America, theSoutheastern Branch of ESA, the Southeastern Biological Control Working Group(SBCWG), and the Georgia and Florida Entomological Societies. Dr. Gross was Pres-ident of the SBCWG in 1990. He was awarded the USDA Certificate of Merit in 1983for exemplary leadership and managerial accomplishments as Director and ResearchLeader for the Insect Suppression Research Unit. He served as a member of an advi-sory panel to provide expertise in the development of a National Biological ControlProgram for ARS. Dr. Gross published 74 articles and made over 70 scientific presen-tations at professional meetings (10 of which were invitational) during his career.
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

388
Florida Entomologist
77(3) September, 1994
Harry served on numerous committees within the Sigma Xi organization (ProgramChairman), the Georgia Entomological Society (Public Relations, Resolutions, and O.I. Snapp Awards committees), the Entomological Society of America (Fellows Commit-tee), and Southeastern Branch ESA (Committee on Constitutional Amendments, Pro-gram Chairman, Agricultural Awards, and Auditing Committees). Dr. Gross alsoserved as Chairman of the Southern Regional Project S-59, Fall Armyworm Confer-ence and the
Heliothis
Conference.Dr. Gross was a dedicated scientist who spent many long and hard hours at his
work. His invention (08/062,516) on “Beehive-mounted device for utilizing honeybees(Hymenoptera: Apidae) in the dissemination of biocontrol agents” was allowed in May1994. Dr. Gross was an expert on integrated biocontrol strategies for key insect pestsand highly knowledgeable of their behavior and population dynamics. He unraveledseveral mysteries of how parasites and predators use an array of chemical signals toidentify and locate pest insects, and how man might manipulate parasites and pred-ators to provide better control of insect pests. He also helped build a foundation for ba-sic knowledge that is, and will continue to be, used by entomologists throughout theworld. Dr. Gross was the first to demonstrate the potential for augmentative releasesof
Archytas marmoratus
, a primary parasitoid of the corn earworm and fall army-worm, and developed a system for its large scale rearing. He demonstrated that cornearworm male moths use visual cues to locate mates and showed that these visualcues could improve the efficiency of insect pheromone traps. Dr. Gross also demon-strated the presence of previously unknown indigenous pupal parasitoids for the fallarmyworm and the tobacco budworm. The frequent citations of his research in inter-national scientific journals offer but a glimpse of the significance with which Dr.Gross's contributions are viewed by the world scientific community. Harry will long beremembered by his friends, colleagues, coworkers, and especially by the technicalsupport personnel, because of his friendliness, fairness and leadership at the IBP-MRL. At the request of Mrs. Gross, a “Harry R. Gross, Jr. Memorial Fund” has beenestablished with the Tifton Sigma Xi Club. Contributions may be made in Harry'sname and sent to the Sigma Xi Club, c/o Dr. Larry Newton, CPES, P.O. Box 748, Tif-ton, GA. 31793-0748.
The Insect Biology and Population Management Research Laboratory:J. E. Carpenter, L. D. Chandler, J. J. Hamm, W. J. Lewis, R. E.Lynch, O. G. Marti, S. D. Pair, W. D. Perkins, C. E. Rogers, A. N.Sparks, H. R. Sumner, N. W. Widstrom, and B. R. Wiseman.USDA, ARS, IBPMRL, Tifton, GA 31793.

I
N
M
EMORIAM
389
I
N
M
EMORIAM
D
R
. Y
IN
-C
HI
H
SU
The standard reference on mayfly biology remains the 1935 classic
Biology of May-flies
by J. G. Needham, J. R Traver, and
Yin-Chi Hsu
, originally published by Com-stock Publishing of Ithaca, New York. Needham and Traver are familiar names, butthe third author is somewhat of an enigma to North American entomologists. Manyare surprised to learn that Hsu continued his research for more than 50 years and, inaddition to his role in North American entomology, Hsu was the founder ofEphemeroptera study in China, as well as being that country’s leading human para-sitologist. I was fortunate to have met him when he visited Florida A&M Universityin 1983, and I am happy to have the opportunity to submit this tribute, prepared bycolleagues in Nanjing, to one historic American mayfly scientist.
William L. Peters, Florida A&M University, Tallahassee
OUR TUTOR DR. XU YINQI (HSU YIN-CHI)(SEPT. 9, 1905–JAN. 29, 1986)
Prof. Xu Yinqi commanded the highest respect in China as a prominent entomolo-gist, parasitologist, and educator. A member of an educated family, he was a highly in-telligent and dedicated student, and earned a BS in Biology from Soochow Universityin 1926, with honorary titles of Beta Beta Beta and Phi Tau Phi. In 1927, he enteredYenching University, Peking, and graduated with an MS in Zoology in 1929; his MSdissertation was entitled “Gryllidae in China.” Because of his outstanding work atYenching, he was awarded a Rockefeller Foundation Fellowship to Cornell University,completing there his PhD degree, with honors, in 1932. He was also offered the Prize
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

390
Florida Entomologist
77(3) September, 1994
of Scientific Research under auspices of China’s Cultural Fund and later received theSino-American Service Professor Fellowship at the University of Minnesota. In 1932,he was elected a member of Sigma Xi; also in 1932, he attended the Vth InternationalCongress of Entomology in Paris, France.
Early in his professional career, Prof. Xu Yinqi was devotedly engaged in researchon Ephemeroptera taxonomy and morphology. After coauthoring (with J. G. Needhamand J. R. Traver) the
Biology of Mayflies
, published in 1935, a universally acknowl-edged classic in Ephemeroptera research, he (1936-1938) published a monograph on
The Mayflies of China
. This work presented a comprehensive survey and summary ofall previous Ephemeroptera in China and added many descriptions of new species ofChina. Prof. Xu Yinqi, considered the founder of Ephemeroptera research in China,enjoyed international renown.
In the early 1950s, he turned his studies to Parasitology and Acarology, investigat-ing serious problems of mite typhus in China. He and his assistants began a system-atic probe into the classification, ecotype, and fauna of chigger mites and their role intransmission of communicable diseases. His forty-odd pieces of published academictreatises on the subject have been frequently quoted, and provide a foundation for thisfield of medical acarology.
In the latter part of the 1970’s, he and his research team studied interrelation-ships between Demodicidae and demodicidosis-dermatopathy. He conducted clinicalcase analyses and pathological studies on the morphological structure, classification,and distribution of Demodicidae on folliculitus, rosacea, etc. as well as treatments forthese conditions, publishing extensively on the subject. Prof. Xu Yinqi constantly pro-moted Chinese science and published numerous treatises, monographs and text-books.
Prof. Xu Yinqi dedicated his life to teaching in the biological sciences. He served asa professor in, sequentially, Soochow University, Yenching University, St. John’s Uni-versity, Shanghai First Medical College (with a long term as Dean of the Biology De-partment), Soochow University (1938-1952), and Professor-in-Charge of the Board ofStudies of Parasitology, Shanghai First Medical College (1952-1984). He was on theCommittee of Experts, Shanghai Medical University from 1984 until 1986. For 59years, he taught courses on Invertebrate Zoology, Comparative Anatomy, Human Par-asitology, Entomology, Medical Entomology, Entomological Taxonomy, Medical Ac-arology, Histology, Embryology, etc. He was an erudite scholar, an animated lecturer,and an indefatigable teacher who was loved and esteemed by his students. His publi-cation,
How to Deliver Successfully a Lecture in Class
, elevated and elucidated the im-portance of instruction for younger faculty members. Numbered among his studentsare academicians of the Academy of Sciences of China, presidents of institutions ofhigher learning, professors, and doctors, all playing important roles in biological,medical, and educational circles in China.
Prof. Xu Yinqi, one of the sponsors of the Zoology Society of China and EntomologySociety of China, served long terms on the Boards of Directors of both societies, as wellas acting as Deputy Director-General of the Shanghai Parasitology Society and as ac-ademic advisor to the Shanghai Museum of Natural Science. Concurrently, he wasDeputy Editor-in-Chief of the
Journal of Parasitology and Parasitic Disease
and amember of the Editorial Board of
Acta Entomologica Sinica, Acta Zootaxonomica Sin-ica, Acta Academiae Medicinae
Primae
Shanghai, Lexicon
(section on medicine),
Eng-lish-Chinese Dictionary of Agricultural Entomology, Glossary and Nomenclature ofAcarology
, and others.Professor Xu Yinqi adopted a vigorous scientific approach in his academic work
and was always well respected by his colleagues. As a professor, he was direct, decent,

I
N
M
EMORIAM
391
enthusiastic and encouraging. He will be long remembered for his outstanding contri-butions to Chinese education and scientific research.
Publications on Ephemeroptera by Xu Yinqi [Hsu Yin-Chi]
1. Hsu, Yin-Chi. 1931. Two new species of mayflies from China. Peking Nat. Hist.Bull. 6(2):39-41.
2. — 1932. Morphology, anatomy, and ecology of genus
Heptagenia
. Ph.D. Diss.,Cornell Univ. Graduate School, Dep. Entomol.
3. — 1933. Some new morphological findings in Ephemeroptera. 5th Int. Congr.Entomol., Paris, 1932, 2:361-368. 2 pls.
4. — 1935. New Chinese mayflies from Kiangsi Province. Peking Nat. Hist. Bull.10(40):319-326.
5. Needham, J. G., J. R. Traver and Y.-C. Hsu. 1935.
The Biology of Mayflies witha Systematic Account of North American Species
. Comstock Publ., Ithaca, NewYork. xi + 759 pp., 168 figs., plates I-XL.
6. Hsu, Y.-C. 1936. Mayflies of Hong Kong with description of two new species(Ephemeroptera). Hong Kong Nat. 7:233-238.
7. — 1936-1937. The Mayflies of China I. Peking Nat. Hist. Bull. 11:129-148. 8. — 1936-1937. The Mayflies of China II. Peking Nat. Hist. Bull. 11:287-296. 9. — 1936-1937. The Mayflies of China III. Peking Nat. Hist. Bull. 11:433-440.10. — 1937-1938. The Mayflies of China IV. Peking Nat. Hist. Bull. 12:53-56.11. — 1937-1938. The Mayflies of China V. Peking Nat. Hist. Bull. 12:123-126.12. — 1937-1938. The Mayflies of China VI. Peking Nat. Hist. Bull. 12:221-224.13. You Da-Shou, Wu Tian, Gui Hong and Hsu Yin-Chi. 1980. A new species of the
genus
Choroterpes
from Nanjing (Ephemeroptera: Leptophlebiidae). Acta Zoot-axonomica Sinica 5(4):388-391.
14. Xu Jia-zhu, You Da-shou, Su Cui-rong and Xu Yin-qi. 1980. Two new species ofgenus
Ephemerella
(Ephemeroptera: Ephemerellidae). J. Nanjing Teacher’sCollege 1980(2):60-63.
15. You Da-shou, Wu Tian, Gui Hong and Hsu Yin-Chi. 1981. Two new species anddiagnostic characters of genus
Cinygmina
(Ephemeroptera; Ecdyoneuridae). J.Nanjing Teacher’s College 1981(3):26-32.
16. You Da-shou, Wu Tian, Gui Hong and Hsu Yin-Chi. 1982. Genus
Potamanth-odes
and two new species (Ephemeroptera: Potamanthidae). Acta Zootaxonom-ica Sinica, 7(4):410-415.
17. You Da-shou, Sui Cui-rong and Hsu Yin-chi. 1982. A new species of the genus
Afronurus
from Fujian Province (Ephemeroptera: Heptageniidae). J. NanjingTeacher’s College, 1982(4):61-65.
18. Xu Jiazhu, You Da-shou and Hsu Yin-Chi. 1984. A new species of
Ephemerella
(Ephemeroptera: Ephemerellidae). Acta Zootaxonomica Sinica, 9(4):413-415.
Gui Hong Zhang JunDepartment of Biology Nanjing NormalUniversity

392
Florida Entomologist
77(3) September, 1994
I
N
M
EMORIAM
H
ARRY
K
YDD
G
OUCK
Harry Kydd Gouck, a retired research entomologist from the USDA-ARS Medicaland Veterinary Entomology Research Laboratory, Gainesville, FL died 9 May, 1994 af-ter a long illness.
Harry Gouck was born and raised in Andover, MA. He attended the University ofNew Hampshire where he earned a B.S. degree in entomology in 1935 and played var-sity football. He continued his education at the University of Massachusetts where hereceived his M.S. degree in entomology in 1936. While a student, and after receivinghis M.S. degree, he worked for the USDA under the direction of Dr. C. N. Smith on theisland of Martha’s Vineyard, MA. Together, they studied the biology and ecology of theAmerican dog tick. When this laboratory was closed in 1942, the staff, includingHarry, moved with their families to Savannah, GA, to continue research on ticks.
The laboratory in Savannah was split in 1947, and some of the staff, includingHarry, joined other scientists at a newly organized facility expressly devoted to thecontrol of medically-important insects. Established at Orlando, FL in 1942, this labo-ratory was the Insects Affecting Man and Animals Research Laboratory which hadthe primary task of solving insect-borne disease problems facing U.S. combat troopsduring World War II. Ultimately, this laboratory and its scientists received global rec-ognition for their contributions to the field of medical entomology.
In 1950, Harry was transferred to the USDA Corn Research laboratory at Cham-paign-Urbana, IL, where he worked on the control of the corn earworm for two years.
In 1952, Harry and his family returned to the Orlando laboratory to find that hiscolleague, Dr. C. N. Smith, had become laboratory director. Harry’s new job was toevaluate and develop insect repellents. He tested numerous chemicals as repellents
This article is from
Florida Entomologist Online
, Vol. 77, No. 3 (1994).
FEO
is available from the Florida Center for Library Automation gopher (sally.fcla.ufl.edu) and is identical to
Florida Entomologist (An International Journal for the Americas).FEO
is prepared by E. O. Painter Printing Co., P.O. Box 877, DeLeon Springs, FL. 32130.

I
N
M
EMORIAM
393
against mosquitoes, biting flies, ticks, chiggers, and leaches. This research took himto field sites in Asia, Africa, Central America and the Bahama Islands. He collabo-rated in the discovery and development of the repellent deet which, to this day, is themost widely used insect repellent in the world.
When the laboratory in Orlando was moved to the campus of the University ofFlorida in 1963, Harry was named head of the mosquito repellents and attractants re-search project. In 1968, together with laboratory chemists, he participated in researchthat led to the identification of lactic acid from human skin as a mosquito attractant.
During his career Harry wrote and/or co-authored more than 60 scientific publica-tions and reported this research at major scientific meetings. He retired from theUSDA in 1973 and, together with his wife, started a second career as a flea marketeerand antique dealer. He continued this work until his health declined three years ago.
Harry is survived by his daughter, Polly Hall of Atlanta, GA, son Peter E. Gouckof North Attleboro, MA, and three grandsons.
Richard S. PattersonCarl E. SchreckMedical and Veterinary Entomology Research LaboratoryUSDA-ARS, SAA MAVERL1600 SW 23rd DriveGainesville, FL 32604





























