Embed Size (px)
Citation preview

Brain-Specific Interaction of a 91-kDa Membrane-Bound Protein with theCytoplasmic Tail of the 300-kDa Mannose 6-Phosphate Receptor
Olaf Rosorius,*,1 Olaf-Georg Issinger,† and Thomas Braulke*,2
*Institute for Biochemistry II, University of Göttingen, Gosslerstr. 12 d, D 37073 Göttingen, Germany; and†Biochemisk Institut, Odense Universitet, Campus Vej 55, DK 5230 Odense M, Denmark
Received March 5, 1996
The cytoplasmic tail of the 300 kDa mannose 6-phosphate receptor (MPR 300-CT) is thought to play animportant role in sorting and targeting of lysosomal enzymes and the insulin-like growth factor II along thebiosynthetic and endocytic pathway. In this study a brain specific 91 kDa protein and a 35 kDa proteinsalt-washed from membranes (referred as TIP 91-M and TIP 35-M) were found to interact with the cytoplasmicreceptor tail as assayed by cross-linkage with recombinant [32P] labeled MPR 300-CT. Subcellular fractionationrevealed a distinct pattern of distribution of TIP 35-M and TIP 91-M in microsomal and synaptosomal fractions.Furthermore, the formation of cross-link complexes with membrane proteins appeared to be developmentally andregionally regulated in the brain and inhibited upon ATP hydrolysis. The data suggest the requirement of specificprotein interactions for MPR 300 functions in neuronal cells.© 1996 Academic Press, Inc.
Newly synthesized soluble lysosomal hydrolases acquire mannose 6-phosphate residues in theGolgi, which function as high affinity recognition signals for mannose 6-phosphate receptors(MPRs), mediating segregation of the enzymes from the secretory route and the targeting tolysosomes. The sorting of lysosomal enzymes is believed to occur in thetrans-Golgi network,(TGN) followed by budding of clathrin-coated vesicles containing receptor-ligand complexes. Thetransport vesicles fuse with an endosomal/prelysosomal compartment. The low pH in the endo-somes induces dissociation of lysosomal enzymes which are then sorted and packed into lysosomesby a process that is as of yet unknown. The MPRs recycle to the Golgi complex to mediate furtherrounds of transport (1).Two distinct MPRs with molecular masses of 46 kDa (MPR 46) and 300 kDa (MPR 300)
function in the targeting of lysosomal enzymes differing in ligand binding properties, transportfunctions and efficiency. MPR 300 has the major function of sorting newly synthesized lysosomalenzymes in the TGN and mediates the endocytosis of exogenous mannose 6-phosphate-containingligands and insulin-like growth factor II (2, 3). The directed intracellular transport of the two MPRsalong the biosynthetic and endocytic pathway is thought to be mediated by signaling elementswhich are proposed as being localized mainly in the cytoplasmic domains. Thus, tyrosine-containing internalization signals, carboxyterminal di-leucine motifs involved in efficient sorting oflysosomal enzymes, and an endosomal retention determinant preventing the MPR transport to thelysosome have been identified (4–8). The interaction of these signal structures with components ofthe vesicular transport machinery, such as AP-1 adaptins in the Golgi or AP-2 in the plasmamembrane (9, 10), may be required to guarantee the distinct MPR trafficking routes.We have previously reported that both MPRs are phosphorylated in their cytoplasmic tail by a
1 Present address: Departement de Biologie Cellulaire, Universite de Geneve, Sciences III, CH-1211 Geneve 4, Swit-zerland.
2 Corresponding author. Fax: (49)-551-395979.Abbreviations: MPR 300, 300 kDa mannose 6-phosphate receptor; MPR 300-CT, cytoplasmic tail of the 300 kDa
mannose 6-phosphate receptor; TIP 35, tail interacting protein with a molecular mass of 35 kDa; CK-2, casein kinase 2;BS3, bis(sulfosuccinimidyl) suberate; GTPgS, guanosine 59-3-O-(thio) trisphosphate; AMPPNP, adenyl-imidodiphosphate;SDS–PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS221,525–530 (1996)ARTICLE NO. 0630
5250006-291X/96 $18.00Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

casein kinase 2 (CK-2)-like kinase (11, 12), and that this specific phosphorylation is required forthe interaction of the cytoplasmic tail of the MPR 300 (MPR 300-CT) with a 35 kDa cytosolicprotein (referred as TIP 35-C). In addition, cross-linkage analysis revealed the interaction of a 35-and 91 kDa protein, salt-washed from bovine brain membranes, with the cytoplasmic tail of theMPR 300 in a phosphorylation-independent manner (13). In the present study we have character-ized the conditions and the proteins forming complexes with the MPR 300 domain. We found thatTIP 35-C is present in various rat tissues, whereas the interaction with the 91 kDa salt-washedmembrane protein was observed only in the brain tissue with regional, temporal and subcellulardifferences in its localization.
MATERIALS AND METHODS
The cytoplasmic tail of the human 300 kDa mannose 6-phosphate receptor (MPR 300-CT) and thea subunit of humanCK-2 were expressed inEscherichia coliand purified as described (11, 14).g[32P] ATP was from Amersham, and thebifunctional crosslinker bis(sulfosuccinimidyl) suberate (BS3) was purchased from Pierce Chemical Co. Nucleotides andanalogs were obtained from Boehringer Mannheim.Subcellular fractionation.Liver, heart, lung and brain tissue were isolated from fed Wistar rats (Thomae GmbH,
Biberach). Cytosolic and membrane fractions were prepared as follows: the sliced tissues were homogenized in a Douncehomogenizer (Wheaton) in buffer A (20 mM Tris/HCl pH 7.4 containing 50 mM NaCl, 1 mM PMSF, 1 mM EDTA, 1 mMDTT), yielding a 30% (wt/vol) homogenate. Total membranes were prepared by centrifugation of the homogenate at100.000×g for 60 min in a TL100 centrifuge (Beckman). Alternatively, membrane fractions were prepared by differentialcentrifugation. After centrifugation at 1000×g for 10 min, the post nuclear supernatant was successively centrifuged at10.000×g for 10 min, 60.000×g for 60 min and 100.000×g for 60 min in a TL100 centrifuge to yield membrane pellets P2,P3 and P4, respectively. A purified synaptosomal fraction was prepared from the cerebral cortex of adult rats by sucrosedensity gradient centrifugation (15). Proteins were extracted by incubation of membranes in 6 volumes of buffer A,containing 1 M NaCl for 60 min on ice. The salt-washed fraction was clarified by centrifugation at 100.000×g for 60 minand the supernatants were dialyzed against 20 mM HEPES pH7.2 and 150 mM NaCl over night.Other methods.Cross-linkage analysis of [32P] labeled MPR 300-CT with cytosolic or salt-washed membrane proteins
were carried out as described previously (13). Protein was determined according to Bradford (16).
RESULTS
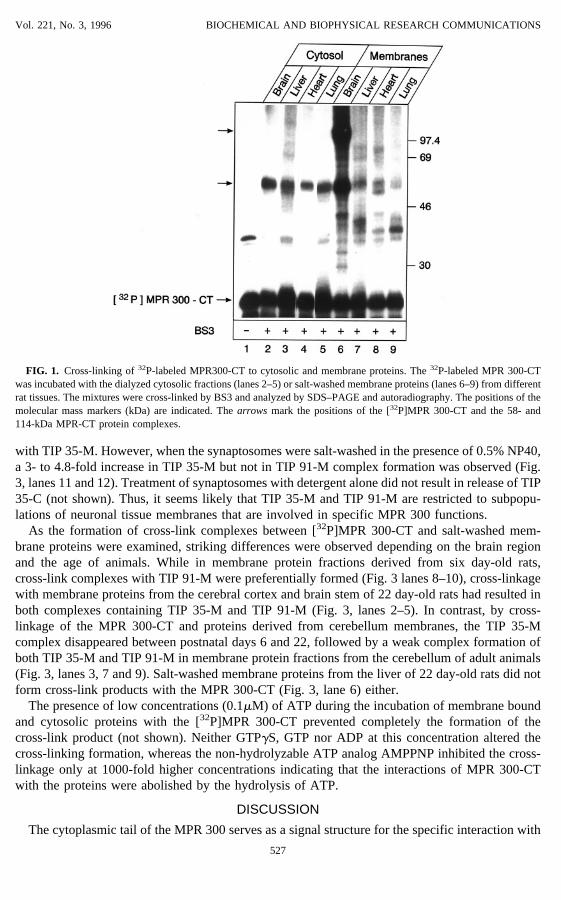
By cross-linkage, a MPR 300-cytoplasmic tail interacting protein of 35 kDa in bovine braincytosol (TIP 35-C) and a 35- and 91 kDa protein, salt-washed from bovine brain membranes, (TIP35-M and TIP 91-M) were detected (13). When cytosolic fractions from rat brain, liver, heart andlung were incubated with the 23 kDa [32P] MPR 300-CT, followed by cross-linking with BS3, the58 kDa cross-link product with TIP 35-C was detected in each tissue preparation (Fig. 1). Theincubation of [32P] MPR 300-CT with BS3 alone in the absence of cellular proteins did not resultin the appearance of multimeric cross-link forms (not shown). The formation of cross-link com-plexes of32P-labeled MPR 300-CT with TIP 91-M was only observed with salt-washed brainmembrane proteins and not with proteins derived from liver, heart or lung membranes (Fig. 1). Inaddition, a strong interaction of the MPR 300-CT with TIP 35-M from brain membranes wasdetected, whereas cross-linkage with membrane proteins from other tissues resulted in weaksignals. In all further experiments, rat brain tissue was used as source of cytosol and/or membraneproteins.To characterize the membranes from which TIP 35-M and TIP 91-M could be salt-washed, a
postnuclear supernatant from a rat brain homogenate was subfractionated by differential centrifu-gation. The various membrane fractions were washed with 1 M NaCl, centrifuged, dialyzed andtested for cross-linkage with the [32P]MPR 300-CT. Fig. 2 shows that only weak cross-linkproducts with TIP 35-M but not with TIP 91-M could be detected in the 10.000×g membrane pellet(lysosomes, mitochondria, synaptosomes). Even though TIP 35-M was equally distributed betweenthe 60.000×g and 100.000×g membrane fractions (“heavy” and “light” microsomes, respectively),a cross-link product with TIP 91-M was observed only in the “heavy” microsome fraction. Cross-linkage analysis with salt-washed synaptosomal proteins resulted in a weak complex formation
Vol. 221, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
526
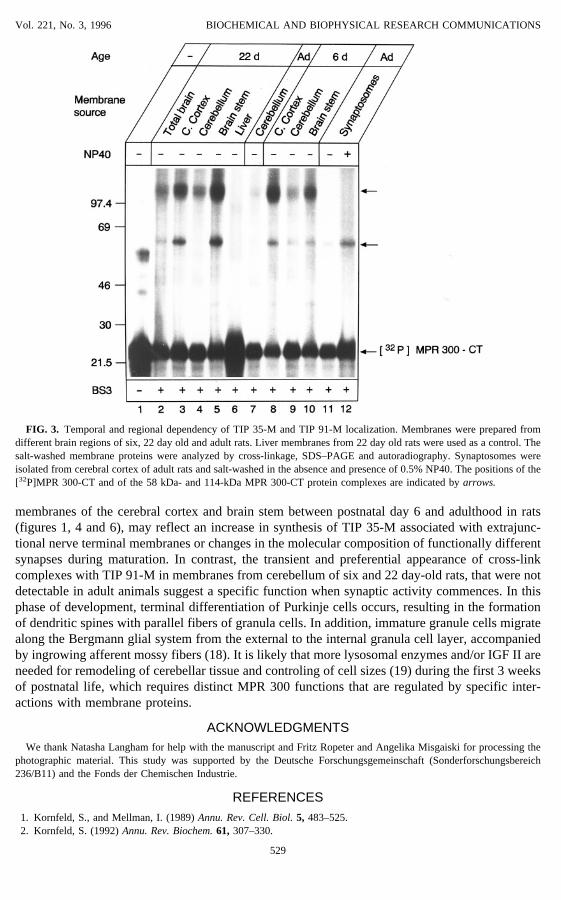
with TIP 35-M. However, when the synaptosomes were salt-washed in the presence of 0.5% NP40,a 3- to 4.8-fold increase in TIP 35-M but not in TIP 91-M complex formation was observed (Fig.3, lanes 11 and 12). Treatment of synaptosomes with detergent alone did not result in release of TIP35-C (not shown). Thus, it seems likely that TIP 35-M and TIP 91-M are restricted to subpopu-lations of neuronal tissue membranes that are involved in specific MPR 300 functions.As the formation of cross-link complexes between [32P]MPR 300-CT and salt-washed mem-
brane proteins were examined, striking differences were observed depending on the brain regionand the age of animals. While in membrane protein fractions derived from six day-old rats,cross-link complexes with TIP 91-M were preferentially formed (Fig. 3 lanes 8–10), cross-linkagewith membrane proteins from the cerebral cortex and brain stem of 22 day-old rats had resulted inboth complexes containing TIP 35-M and TIP 91-M (Fig. 3, lanes 2–5). In contrast, by cross-linkage of the MPR 300-CT and proteins derived from cerebellum membranes, the TIP 35-Mcomplex disappeared between postnatal days 6 and 22, followed by a weak complex formation ofboth TIP 35-M and TIP 91-M in membrane protein fractions from the cerebellum of adult animals(Fig. 3, lanes 3, 7 and 9). Salt-washed membrane proteins from the liver of 22 day-old rats did notform cross-link products with the MPR 300-CT (Fig. 3, lane 6) either.The presence of low concentrations (0.1mM) of ATP during the incubation of membrane bound
and cytosolic proteins with the [32P]MPR 300-CT prevented completely the formation of thecross-link product (not shown). Neither GTPgS, GTP nor ADP at this concentration altered thecross-linking formation, whereas the non-hydrolyzable ATP analog AMPPNP inhibited the cross-linkage only at 1000-fold higher concentrations indicating that the interactions of MPR 300-CTwith the proteins were abolished by the hydrolysis of ATP.
DISCUSSION
The cytoplasmic tail of the MPR 300 serves as a signal structure for the specific interaction with
FIG. 1. Cross-linking of32P-labeled MPR300-CT to cytosolic and membrane proteins. The32P-labeled MPR 300-CTwas incubated with the dialyzed cytosolic fractions (lanes 2–5) or salt-washed membrane proteins (lanes 6–9) from differentrat tissues. The mixtures were cross-linked by BS3 and analyzed by SDS–PAGE and autoradiography. The positions of themolecular mass markers (kDa) are indicated. Thearrowsmark the positions of the [32P]MPR 300-CT and the 58- and114-kDa MPR-CT protein complexes.
Vol. 221, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
527

at least five distinct proteins, i.e. AP-1, AP-2, TIP 35-C, TIP 35-M and TIP 91-M.In vitroexperiments have shown that the recruitment of AP-1 adaptor proteins to Golgi membranes, animportant step in the formation of coated buds, and the interaction with the cytosolic TIP 35 requirethe phosphorylation of the MPR 300-CT at two serine residues catalyzed by CK-2 (10, 13).Analysis of wild type and mutant MPR 300-CT suggested that a tyrosine residue present in the tightb turn internalization sequence is part of the recognition motif of the AP-2 adaptin, a constituentof clathrin-coated vesicles derived from the plasma membrane. Both adaptor complexes recognizedistinct and non-overlapping receptor sites. The determinants in the MPR 300-CT, necessary forthe interaction with the membrane-bound TIP 35-M and TIP 91-M, have not yet been identified.Due to i) the ubiquitous occurrence of TIP 35-C in various tissues, in comparison with thepreferential localization of TIP 35-M in brain membranes, ii) the independence of TIP 35-Minteraction on receptor tail phosphorylation, and iii) the differences in dependency on divalentcations (O. Rosorius, unpublished results), it is rather unlikely that TIP 35-M represents a modifiedor activated form of TIP 35-C.The different subcellular localization and the developmentally related changes in the cross-
linkage pattern may indicate distinct functions mediated by TIP 35-M and TIP 91-M. The cellularlocalization of membrane-bound TIPs in brain tissue cannot be resolved from this study. However,in the synaptosomal preparation the majority of TIP 35-M is localized in intrasynaptosomal mem-branes which become accessible in the presence of detergents. Thus, the faint signal of TIP 35-Min the salt-washed 10.000×g membrane fraction most likely arised from “leaky” synaptosomes thatare present. Whether TIP 35-M is a constituent of the synaptic membrane, endosomes, cytoskeletonor of the synaptic vesicles has to be determined by refined fractionation procedures of lysedsynaptosomes and co-immunoprecipitation experiments. However, the presence of TIP 35-M insynaptosomes may not reflect the necessity of interaction with the MPR 300 but could be importantfor MPR 300 function in endocytosis at nerve terminals. In addition, the fact that the IGF IIimmunoreactivity in synaptosomes is 8- to 10-fold higher than in the crude extract from motorcortex (17), suggests that synaptosomal MPR 300 may be required to control local IGF II effects.The relative increase in the appearance of cross-linked products with TIP 35-M, derived from
FIG. 2. Subcellular distribution of TIP 35-M and TIP 91-M. Rat brain membrane fractions obtained either afterdifferential centrifugation of a post nuclear supernatant at 10.000×g (1), 60.000×g (2), 100.000×g (3) or after one step100.000×g centrifugation (4) were washed with 1 M NaCl. The dialyzed extracts were cross-linked with the [32P]MPR300-CT and analyzed by SDS–PAGE and autoradiography. The positions of the [32P]MPR 300-CT, TIP 35-M and TIP91-M are indicated byarrows.
Vol. 221, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
528
membranes of the cerebral cortex and brain stem between postnatal day 6 and adulthood in rats(figures 1, 4 and 6), may reflect an increase in synthesis of TIP 35-M associated with extrajunc-tional nerve terminal membranes or changes in the molecular composition of functionally differentsynapses during maturation. In contrast, the transient and preferential appearance of cross-linkcomplexes with TIP 91-M in membranes from cerebellum of six and 22 day-old rats, that were notdetectable in adult animals suggest a specific function when synaptic activity commences. In thisphase of development, terminal differentiation of Purkinje cells occurs, resulting in the formationof dendritic spines with parallel fibers of granula cells. In addition, immature granule cells migratealong the Bergmann glial system from the external to the internal granula cell layer, accompaniedby ingrowing afferent mossy fibers (18). It is likely that more lysosomal enzymes and/or IGF II areneeded for remodeling of cerebellar tissue and controling of cell sizes (19) during the first 3 weeksof postnatal life, which requires distinct MPR 300 functions that are regulated by specific inter-actions with membrane proteins.
ACKNOWLEDGMENTSWe thank Natasha Langham for help with the manuscript and Fritz Ropeter and Angelika Misgaiski for processing the
photographic material. This study was supported by the Deutsche Forschungsgemeinschaft (Sonderforschungsbereich236/B11) and the Fonds der Chemischen Industrie.
REFERENCES
1. Kornfeld, S., and Mellman, I. (1989)Annu. Rev. Cell. Biol.5, 483–525.2. Kornfeld, S. (1992)Annu. Rev. Biochem.61, 307–330.
FIG. 3. Temporal and regional dependency of TIP 35-M and TIP 91-M localization. Membranes were prepared fromdifferent brain regions of six, 22 day old and adult rats. Liver membranes from 22 day old rats were used as a control. Thesalt-washed membrane proteins were analyzed by cross-linkage, SDS–PAGE and autoradiography. Synaptosomes wereisolated from cerebral cortex of adult rats and salt-washed in the absence and presence of 0.5% NP40. The positions of the[32P]MPR 300-CT and of the 58 kDa- and 114-kDa MPR 300-CT protein complexes are indicated byarrows.
Vol. 221, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
529

3. Hille-Rehfeld, A. (1995)Biochim. Biophys. Acta1241,177–194.4. Johnson, K. F., Chan, W., and Kornfeld, S. (1990)Proc. Natl. Acad. Sci. USA87, 10010–10014.5. Canfield, W. M., Johnson, K. F., Ye, R. D., Gregory, W., and Kornfeld, S. (1991)J. Biol. Chem.266,5682–5688.6. Johnson, K. F., and Kornfeld, S. (1992)J. Cell Biol.119,249–257.7. Johnson, K. F., and Kornfeld, S. (1992)J. Biol. Chem.267,17110–17115.8. Rohrer, J., Schweizer, A., Johnson, K. F., and Kornfeld, S. (1995)J. Cell Biol.130,1297–1306.9. Glickman, J. N., Conibear, E., and Pearse, B. M. F. (1989)EMBO J8, 1041–1047.10. LeBorgne, R., Schmidt, A., Mauxion, F., Griffiths, G., and Hoflack, B. (1993)J. Biol. Chem.268,22552–22556.11. Rosorius, O., Mieskes, G., Issinger, O.-G., Körner, C., Schmidt, B., von Figura, K., and Braulke, T. (1993)Biochem.
J. 292,833–838.12. Körner, C., Herzog, A., Weber, B., Rosorius, O., Hemer, F., Schmidt, B., and Braulke, T. (1994)J. Biol. Chem.269,
16529–16532.13. Rosorius, O., Issinger, O.-G., and Braulke, T. (1993)J. Biol. Chem.268,21470–21473.14. Grankowski, N., Boldyreff, B., and Issinger, O.-G. (1991)Eur. J. Biochem.198.15. Bradford, M. M. (1976)Anal. Biochem.72, 248–254.16. Marchbanks, R. M. (1968)Biochem. J.110,533–541.17. Haselbacher, G. K., Schwab, M. E., Pasi, A., and Humbel, R. E. (1985)Proc. Natl. Acad. Sci. USA82, 2153–2157.18. Hatten, M. E., and Heintz, N. (1995)Annu. Rev. Neurosci.18, 385–408.19. Nixon, R. A., and Cataldo, A. M. (1995)Trends Neurosci.18, 489–496. 25–30.
Vol. 221, No. 3, 1996 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
530







![Prognostic significance of cytoplasmic S100A2 ... · expression of S100A2 in OSCC in comparison with histo-logically normal oral mucosa [5,6]. S100A2, an 11.4 kDa protein, is a member](https://img.pdfslide.us/doc/110x75/602b77c31cce2f3d31452980/prognostic-significance-of-cytoplasmic-s100a2-expression-of-s100a2-in-oscc-in.jpg)




















