Embed Size (px)
Citation preview

Ingham et al.: Brain Correlates of Stuttering 321Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004 • ©American Speech-Language-Hearing Association1092-4388/04/4702-0321
Roger J. InghamUniversity of California,
Santa Barbaraand
University of Texas HealthScience Center at San Antonio
Peter T. FoxUniversity of Texas Health
Science Center at San Antonio
Janis C. InghamUniversity of California,
Santa Barbaraand
University of Texas HealthScience Center at San Antonio
Jinhu XiongFrank Zamarripa
L. Jean HardiesJack L. Lancaster
University of Texas HealthScience Center at San Antonio
Brain Correlates of Stutteringand Syllable Production: GenderComparison and Replication
This article reports a gender replication study of the P. T. Fox et al. (2000)performance correlation analysis of neural systems that distinguish betweennormal and stuttered speech in adult males. Positron-emission tomographic (PET)images of cerebral blood flow (CBF) were correlated with speech behavior scoresobtained during PET imaging for 10 dextral female stuttering speakers and 10dextral, age- and sex-matched normally fluent controls. Gender comparisonswere made between the total number of voxels per region significantly correlatedwith speech performance (as in P. T. Fox et al., 2000) plus total voxels per regionthat were significantly correlated with stutter rate and not with syllable rate.
Stutter-rate regional correlates were generally right-sided in males, butbilateral in the females. For both sexes the positive regional correlates forstuttering were in right (R) anterior insula and the negative correlates were in RBrodmann area 21/22 and an area within left (L) inferior frontal gyrus. Thefemale stuttering speakers displayed additional positive correlates in L anteriorinsula and in basal ganglia (L globus pallidus, R caudate), plus extensive righthemisphere negative correlates in the prefrontal area and the limbic and parietallobes. The male stuttering speakers were distinguished by positive correlates in Lmedial occipital lobe and R medial cerebellum. Regions that positively correlatedwith syllable rate (essentially stutter-free speech) in stuttering speakers andcontrols were very similar for both sexes.
The findings strengthen claims that chronic developmental stuttering isfunctionally related to abnormal speech-motor and auditory region interactions.The gender differences may be related to differences between the genders withrespect to susceptibility (males predominate) and recovery from chronic stuttering(females show higher recovery rates during childhood).
KEY WORDS: stuttering, brain imaging, gender differences, PET
There is a growing consensus that developmental stuttering is aproduct of central nervous system dysfunction with genetic origins(see Ingham, 2001; Yairi, Ambrose, & Cox, 1996). Incomplete left
lateralization of speech and other motor skills is a much-theorized neu-rologic source of the disorder (Moore, 1993; Travis, 1978). This theoreticposition, originally derived from inferential studies of the neural sys-tem (see Ingham, 1998), has been fortified by recent functional brainimaging studies using positron emission tomography (PET) and mag-netic resonance imaging (MRI). Conducted mainly with dextral adultmale stuttering speakers, these studies have detected incomplete leftlateralization of speech motor systems and overactive premotor areasduring stuttering (Braun et al., 1997; De Nil, Kroll, Kapur, & Houle, 2000;

322 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
Fox et al., 1996, 2000; Wood, Stump, McKeehan,Sheldon, & Proctor, 1980; Wu et al., 1995). In addition,less-than-normal activations, even inhibitions (decreasesin brain blood flow relative to control conditions), in ex-tra-primary auditory areas have been observed (Braunet al., 1997; Fox et al., 1996). These latter findings areconsistent with stuttering theories that have emphasizeddysfunctional auditory processing (e.g., Stromsta, 1986;Yates, 1963). Both theoretic positions gained supportfrom a recent performance correlation study (Fox et al.,2000) that showed stuttering frequency in adult maledextral stuttering speakers was correlated with cere-bral blood flow (CBF) increases in speech-motor regionsand decreases in auditory regions. These neurophysiol-ogy studies have now been complemented by reports ofneuroanatomic abnormalities in left hemisphere speech-motor regions of mixed-gender groups of adult stutteringspeakers (Foundas, Bollich, Corey, Hurley, & Heilman,2001; Sommer, Koch, Paulus, Weiller, & Büchel, 2002),highlighting a possible source(s) for the abnormal CBFfindings.
Many studies point to the importance of studyingwomen who stutter separately from men in brain imag-ing research. The neurophysiologic evidence that men’sand women’s brains generally differ in processingspeech-related tasks is beginning to emerge as a task-specific phenomenon, although these differences mayalso have structural foundations. Initial functional MRI(fMRI) studies showing that women display greater bi-laterality (vs. left lateralization for men) and less acti-vation in inferior frontal gyrus during phonologic pro-cessing (Pugh et al., 1996; Shaywitz et al., 1995) havenot been completely replicated (see Buckner, Raichle, &Petersen, 1995; Xiong, Rao, Jerabek, Woldorff, & Fox,2000). Similar mixed findings have been obtained fromfMRI studies during listening tasks: Frost et al. (1999)found no gender differences during a semantic monitor-ing task, but others (Kansaku, Yamaura, & Kitazawa,2000; Phillips, Lowe, Lurito, Dzemidzic, & Mathews 2001)have reported that women show greater bilateral tempo-ral lobe activations while listening to oral reading. Thefocus on possible gender differences in neurophysiologicstudies is also justified by morphometric studies (e.g.,McGlone, 1980; Witelson & Kigar, 1992). An autopsy study(Harasty, Double, Halliday, Kril, & McRitchie, 1997) foundthat the language-important planum temporale andBroca’s regions were, respectively, 30% and 20% largerin women than in men. And, in an extensive morpho-logical study of 465 adult brains, Good et al. (2001) con-cluded that there were substantial gender-based differ-ences in gray matter volume and concentration.
Gender differences are an important characteris-tic of developmental stuttering. Current theories ofstuttering must ultimately account for the fact that thedisorder occurs more frequently among males, typically
2:1–3:1 (Bloodstein, 1995). There is also a much higherprobability that the progeny of females who stutter, bycomparison with male stuttering speakers’ progeny, willhave the disorder (Kidd, Heimbuch, & Records, 1981).However, there is a remarkable paucity of research onstuttering in females, and no neurophysiological re-search on gender differences. Neural processing studieshave not found gender differences on bimanual hand-writing (Webster, 1988), central auditory processing(Nuck, Blood, & Blood, 1987), or dichotic listening (Blood& Blood, 1989) tasks. But these studies neither ad-dressed speech or language processing nor involved neu-rophysiologic analyses. Functional imaging studies ofstuttering in women are important in order to deter-mine if the neural regions previously found to be re-lated to stuttering in men are gender specific or are rep-licated across genders (and, hence, independent of speechprocessing differences between females and males).
Within-subject conditional contrasts, despite beingwidely used in neuroimaging studies—including mostPET studies on stuttering—do not always constitute astrong experimental strategy. Conditional contrasts relyon the assumption that behavioral/cognitive task com-ponents present in different conditions will be subservedby activation of the same brain areas and to the samedegree in both conditions. Thus, it is assumed that sub-tracting one condition from the other will remove thecommon activations, leaving those functionally relatedto the test task to be the product of the subtraction in theconditional contrast. The assumption that a single or“pure insertion” (Boring, 1950) test task will separate theconditions has been challenged on theoretic (Petersen,van Mier, Fiez, & Raichle, 1998) and empirical (Sidtis,Strother, Anderson, & Rottenberg, 1999) grounds. Fur-ther, conditional contrasts assume the phenomenon ofinterest (e.g., stuttering) can be isolated to one condi-tion. Although the symptoms of many neurologic andpsychiatric disorders can be iatrogenically modulated,they are rarely isolated. Thus, an experimental strat-egy applicable when the behavior of interest is not en-tirely under the experimenter’s control is desirable.
An alternative strategy, first introduced by Silber-sweig et al. (1995), is known as performance correlationanalysis. This strategy utilizes the principle that theintensity of brain activations is correlated with the fre-quency with which the neural elements are used duringimaging tasks (e.g., Fox & Raichle, 1984, 1985). It hasbeen used to study hallucination rates in schizophrenicpatients (Silbersweig et al., 1995), speech perceptionafter a cochlear implant (Fujiki et al., 1999), and tic fre-quency in patients with Tourette’s syndrome (Stern etal., 2000).
Braun et al. (1997) reported the first performancecorrelation strategy in a stuttering CBF study. Using a

Ingham et al.: Brain Correlates of Stuttering 323
“weighted dysfluency score” to compute voxel-wise cor-relations, they found positive correlations in several lefthemisphere motor regions and negative correlationswithin bilateral auditory association (A2) areas. Fox etal. (2000) separately measured stuttering and speechrate in order to identify the extent of regional dissocia-tion between stuttering and syllable rate among adultmale stuttering speakers and in relation to the controls’syllable rate during the Fox et al. (1996) study. In gen-eral, both groups’ syllable rates showed positive corre-lations in similar regions, though more extensively incerebellum for the stuttering speakers. The main dif-ference between stutter- and syllable-rate correlationswas that the regions where activations were positivelycorrelated with stutter rate were more right-lateralized,particularly in the speech-motor regions, and only stut-ter rate produced negatively correlated areas withinright (R) A2. Another feature of the Fox et al. (2000)study was the proposed classification of the voxel corre-lates into state, trait, and compensatory effects that arereflected in syllable and/or stutter rates during the im-aging conditions.
The present study replicates the Fox et al. (2000)procedure by comparing adult female stuttering speak-ers and normally fluent controls with previously stud-ied male counterparts so as to (a) identify regions func-tionally related to stuttering across genders, and (b)determine the extent to which the regional effects asso-ciated with stuttering are gender specific. Comparisonsare also made with a state-trait-compensatory effect clas-sification proposed by Fox et al.
MethodParticipants
Ten right-handed females with a history of develop-mental stuttering (mean age = 44 years; stuttering se-verity ratings [Lewis & Sherman, 1951]: mild = 3, mod-erate = 3, severe = 4]) formed the stuttering group. Allwere pretested to ascertain responsivity to chorus read-ing (Ingham & Packman, 1979). Neither the female normale participants who stutter had received therapy inthe past 5 years. Only persons who produced zero stut-tering during pretesting with chorus reading were par-ticipants. Ten right-handed normally fluent womenformed the control group (mean age = 42 years). Forcomparison, the data from 10 right-handed males whostutter (mean age = 32 years) and their matched con-trols (mean age = 32 years) were obtained from thosereported in Fox et al. (1996, 2000). The female and malegroups participated in identical experiments. Informedconsent was obtained from all participants in accordancewith the Declaration of Helsinki and under the auspicesof the Institutional Review Boards of the University of
Texas Health Science Center at San Antonio and theUniversity of California, Santa Barbara.
PET Imaging TasksThe tasks were identical to those described in Fox
et al. (1996, 2000). Each participant received nine PETscans: three trials in each of three conditions in a coun-terbalanced order. The conditions were as follows: oralparagraph reading of a text passage (Solo), oral para-graph reading while accompanied by an audio record-ing of the same passage being read by a fluent speaker(Chorus), and eyes-closed rest (Rest). The passage forreading (Abbey, 1975) was presented on a video monitorsuspended above the participant, approximately 14 in.from the eyes. For Chorus, the recorded passage was pre-sented via an earphone inserted in the participant’s leftear. To counter adaptation effects (Van Riper & Hull,1955), the 10-min intervals between scans were occupiedwith casual conversation. For each task, reading wasstarted at the moment of tracer injection, continued whilethe tracer circulated to the brain (~10 s), and was stopped40 s after image acquisition was triggered by the ar-rival of the tracer bolus in the brain (see below).
Speech MeasurementsAs in Fox et al. (1996, 2000), speech performance
data were derived from audiovisual recordings obtainedduring the 40-s PET scanning periods. Stuttering ratewas computed as the number of 4-s intervals judged tocontain stuttering (Ingham, Cordes, & Gow, 1993), witha maximum possible score of 10 in a 40-s scanning ep-och. Syllable production rate was computed as the totalnumber of syllables spoken in the 40-s scanning period,counting each repeated syllable in a stutter so as to fullyreflect speech-motor behavior. Speech naturalness wasrated on a 9-point scale (Martin, Haroldson, & Triden,1984). Data were collected using the Stuttering Mea-surement System program (Ingham, Bakker, Kilgo, &Moglia, 1999). To establish measurement reliability, arandomly selected sample of recordings (6 stutteringspeakers and 4 controls) was scored independently byan experimenter (the third author) and a trained judgewho was blind to task conditions and group. Measure-ment agreement was satisfactory. A Pearson r correla-tion between syllable counts for the two judges was .99(p < .001). The judges’ mean syllable counts were notsignificantly different (118.0 vs. 117.0; p > .1, paired ttest). A .99 (p < .001) correlation was also obtained be-tween the judges’ stuttered interval (SI) counts on thestuttering speakers’ samples. The judges’ mean SI countswere also not significantly different on a paired t test(3.74 vs. 3.60; p > .1). The judges each provided 60 speechnaturalness scores, and 59 were within ±1 scale unit.

324 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
Both judges agreed that no stuttering occurred on anyof the controls’ recordings.
Measurement reliability data for the male groups’SIs, syllable counts, and speech naturalness ratings werereported in Fox et al. (2000). They were similar to thosereported for the females.
Image Acquisition and AnalysisThe H2
15O PET imaging and analysis procedureswere identical to those reported in Fox et al. (2000). Ananatomical MRI was acquired for each participant andused to optimize spatial normalization. PET and MRimages for each participant were spatially normalizedrelative to the Talairach and Tournoux (1988) brain at-las using software devised by Lancaster et al. (1995).Brain dimensions obtained from the MR image wereapplied to both the MR and PET images, which werethen transformed into 3-D, spatially normalized imagesusing 2 × 2 × 2 mm voxels.
Three r-value, statistical parametric images (SPI{r})were computed as voxel-wise correlations with each ofthree speech performance measures: (a) correlation withstuttering rate in the stuttering groups, (b) correlationwith syllable production rate in the stuttering groups,and (c) correlation with syllable production rate in thenonstuttering groups. Thus, correlations were calculatedbetween CBF (in every voxel per condition) and each ofthe three speech measures. The calculation of these threeperformance correlations permits the identification ofneural correlates that are specifically associated withstuttering or with syllable production in both gendergroups. Syllable rate measures the rate of articulatorymovement as distinguished from movement associatedwith stuttering and essentially measures nonstutteredspeech. For each SPI{r}(syllable-rate correlates and stut-ter-rate correlates), PET images from all three test con-ditions (Rest, Solo, Chorus) were included. The inclu-sion of conditions in which stuttering is absent (Rest andChorus) increased the overall power of the correlationsby ensuring that the stutter-related correlations wouldbe independent of those that might occur in the absenceof stuttering. This correlation analysis was identical tothat used in Fox et al. (2000), as were all procedures usedfor normalizing images and identifying locations. P val-ues were assigned from the image z distributions. Onlypositive and negative r values at or greater than .30 (p< .001) and forming contiguous clusters of 15 or morevoxels are reported (Xiong, Gao, Lancaster, & Fox, 1995).
It should be noted that for these analyses, voxel clus-ters that were correlated with stutter rate could also becorrelated with syllable rate. Therefore, an additionallogical contrast analysis (not performed in Fox et al.,2000) was conducted using images derived from the
stutter-rate correlates and from the syllable-rate corre-lates for both sexes so as to identify voxel clusters sig-nificantly correlated with stutter rate and not with syl-lable rate. As an additional insurance that these clusterswere unique to stutter rate, the selected clusters of voxelsidentified as exclusively related to stutter rate werethose that were outside a 53 voxel area surrounding anyvoxel that was significantly correlated with syllable rate.
Anatomical labels and Brodmann area (BA) desig-nations were applied automatically, using a 3-D, elec-tronic brain atlas (the Talairach Daemon™; Lancasteret al., 1997) and subsequently checked by hand. Fifty-four regions or structures (27 in each hemisphere) ac-counted for all performance-correlated voxel clusters.These regions are listed in Table 3 (below).1 These re-gions were preselected by using those activated inspeech task studies summarized by Fiez and Peterson(1998) and Indefrey and Levelt (2000). A coarser levelof categorization was used for the parietal and occipitallobes and cerebellum according to criteria used byIndefrey and Levelt. Their criteria identified subregionswithin these lobes that they found, via a meta-analysis,were associated with speech and language production.
Data analyses within this study were conductedin two independent ways in order to determine the ro-bustness of the findings: (a) For each group, the totalvoxels per region (occurring in clusters of 15 or more)that correlated with stutter and/or syllable rate was cal-culated. This permitted comparisons within and betweenthe groups for the number of correlated voxels per re-gion. (b) For the stuttering speakers groups, the totalvoxels that correlated with stutter rate and not withsyllable rate was determined within the regions iden-tified in (a). The latter permitted comparisons betweengenders in regard to brain areas linked exclusively tostuttering.
ResultsSpeech Performance
Chorus reading was a very effective fluency-induc-ing procedure for the female stuttering speakers, al-though, as Table 1 shows (see the Stutters column), itwas slightly less effective for the females than the males.(The male groups’ data, previously reported in Fox etal., 2000, are also included in Table 1.) Stuttering waseliminated during all chorus-reading trials (three perparticipant) for 7 of the 10 females who stuttered—theother 3 produced at least 80% reduction in stuttering.
1 The regions listed in Table 3 do not obviously total 54. Some regions arelisted twice because they contain positively and negatively correlatedvoxels, and because basal ganglia is not divided into its separate caudateand globus pallidus sections (as shown in Table 4).

Ingham et al.: Brain Correlates of Stuttering 325
In addition, chorus reading produced equivalent meansyllable production rates in the stuttering and controlgroups. As expected, stuttered speech was judged rela-tively unnatural sounding, but the mean speech natu-ralness ratings during chorus reading were similar forthe female and male stuttering speakers and controls,and within the range expected for normally fluent speak-ers (Ingham, 1988).
Table 1 shows the behavioral data for this study.The female stuttering group averaged 7.3 stuttered in-tervals (SD = 3.1) and 78 syllables (SD = 28.8) per 40-sscan in the Solo condition. In the Chorus condition, anaverage of 0.5 stuttered intervals occurred in the femalestuttering group (SD = 0.8); the mean number of syl-lables spoken was 128 (SD = 11.6). For the female stut-tering group, the difference in mean stuttering rate be-tween the Solo and Chorus conditions was highlysignificant, t(28) = 6.21, p < .0001; the difference in meansyllable rate was also highly significant, t(28) = 5.18, p< .0001. Stuttering rate and syllable rate were inverselycorrelated only in the Solo condition (r = –.84, p < .005).
Female control participants did not stutter in ei-ther condition and spoke an average of 149 syllables (SD= 18.7) in the Solo condition and 128 syllables (SD =12.5) in the Chorus condition. The difference in meansyllable rate between the Solo and Chorus conditionswas also significantly different for the control group, t(28)= 4.16, p < .01. No difference in mean syllable rate wasexpected between stuttering speakers and controls dur-ing the Chorus condition because stuttering speakers’
self-selected chorus reading stimulus tapes were eachmatched for speech rate across the two groups.
CBF CorrelationsThe voxel totals from both genders that were posi-
tively and negatively correlated with stutter and syllablerates for the stuttering speakers, and with syllable ratefor the controls, are shown in bar graph form in Figure 1.
The total voxels correlating positively and negativelywith syllable rate and for stutter rate (stuttering speak-ers only) for each gender group were tested for genderdifferences using separate 2 × 2 chi-square tests. Sig-nificant gender differences were obtained for both thestuttering speakers’ stutter rates, !2(1, N = 8866) = 73.76,p < .001, and syllable rates, !2(1, N = 30082) = 228.41, p< .001, plus the controls’ syllable rates, !2(1, N = 30656)= 3.99, p < .05. In total, the females produced 29.8%fewer positively correlated voxels and 15.7% fewer nega-tively correlated voxels than their male counterparts.This finding is consistent with previous findings regard-ing overall CBF activations in males and females dur-ing speaking tasks (e.g., Buckner et al., 1995), althoughthe differences favoring the males could be related tothe age differences between the gender groups—a dif-ference known to influence the number of neocorticalneurons in the brain (Pakkenberg & Gundersen, 1997).
Because the total number of significantly correlatedvoxels differed between the sexes, at least for the stut-tering speakers, this difference confounded any direct
Table 1. Speech performance scores and ages for the female participants in the present study are shown(mean and range) along with parallel data for the male participants from Fox et al. (2000).
Stutters Syllables NaturalnessAge
Group Age Range Condition M Range M Range M Range
Female stutterers 44 25–73 Solo 7.3 1–10 78 35–130 6.5 3–9Chorus 0.5 0–3 128 109–147 2.9 2–4
Female controls 42 26–75 Solo 0.0 0 149 89–183 2.4 1–3Chorus 0.0 0 128 103–141 2.7 2–4
Male stutterers 32 21–55 Solo 6.2 1–10 113 82–154 5.5 3–9Chorus 0.0 0 144 121–171 2.5 1–4
Male controls 32 21–53 Solo 0.0 0 147 124–188 2.4 1–4Chorus 0.0 0 146 121–181 2.5 1–4
Note. “Stutters” indicates the mean number of 4-s epochs during the 40-s scanning period judged to containstuttering (i.e., stutter rate). “Syllables” indicates the mean number of syllables uttered during the 40-s scanningperiod (i.e., syllable production rate). “Naturalness” indicates the mean speech naturalness scores, with a rangeof 1 (highly natural) to 9 (highly unnatural). For syllable and stutter scores, n = 30 for each condition (3 scanepochs per participant × 10 participants), giving a total of 90 scores (Solo, Chorus, and Rest) for syllables and forstutters for stutterers, and 90 scores for syllables for the controls. These 90-score sets were used as correlates todetermine the PET blood flow for regional correlations with each speech variable (stutter rate and syllableproduction rate). Naturalness scores were not used as potential correlates for image analysis.

326 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
comparisons between the sexes with respect to total cor-related voxels in particular regions. Therefore, compari-sons between the sexes’ regional activations were madeby estimating the similarity (or otherwise) of their dis-tributions of positively and/or negatively correlatedvoxels across the 54 regions studied. This was accom-plished by deriving correlation coefficients between thevoxel totals across regions from both gender groups (stut-tering speakers and controls) for syllable and stutterrate (for the stuttering speakers) (see Table 2). A sig-nificant and strong positive correlation between the to-tal number of positively correlated voxels per region forsyllable rate for both sexes would mean that the rate ofsyllable production by male and female speakers in-volved similar distributions of across-region neural ac-tivity. Conversely, nonsignificant correlations betweenthe sexes for numbers of performance-correlated voxelsacross the regions would point to substantial differencesbetween the distributions of regional activity by the maleand female speakers.
Table 2 confirms that there were strong positivecorrelations between the females and males in regionswhere voxel clusters occurred that were positively cor-related with the rate of syllable production—for stut-tering speakers, r(22) = .870, p < .01; for controls, r(19)= .805, p < .01—and negatively correlated with syllablerate—for stuttering speakers, r(36) = .665, p < .01; forcontrols, r(34) = .661, p < .01. The corresponding corre-lation coefficients for stutter rate were significant forthe positively correlated voxels, r(16) = .558, p < .05,but not for the negatively correlated voxels, r(26) = .052,ns. In other words, the genders were not consistently
similar with respect to the proportion of voxels per re-gion that were negatively correlated with the rate of stut-tering, but there were consistent similarities betweenthe genders with respect to syllable rate. Table 2 also
Figure 1. The volume (in total voxels) of blood flow changes positively or negatively correlated with stutterand syllable rate in female and male stuttering speakers and controls. All positive voxel cluster correlateshave a peak-voxel r " +0.3 and size " 15; negative correlates have a peak-voxel r # –0.3 and size " 15.
Table 2. Correlations between total voxels per region that werepositively or negatively correlated with stutter or syllable rate.
Voxels ××××× Rate correlations Females ××××× Males r
(a)Positively correlated voxels
Stutter rate .558*Stutterers’ syllable rate .870**Controls’ syllable rate .805**
Negatively correlated voxelsStutter rate .052Stutterers’ syllable rate .665**Controls’ syllable rate .661**
Females r Males r
(b)Positively correlated voxels
Stutterers’ × Controls’ Syllable Rate .885** .915**Stutter Rate × Stutterers’ Syllable Rate .646** .825**
Negatively correlated voxelsStutterers’ × Controls’ Syllable Rate .746** .678**Stutter Rate × Stutterers’ Syllable Rate .150 .009
Note. Table 2A shows between-gender correlations derived from thetotal number of significantly activated voxel clusters within each of 27regions in the left and right hemisphere (*p < .05; **p < .01). Table 2Bshows the within-gender correlations.

Ingham et al.: Brain Correlates of Stuttering 327
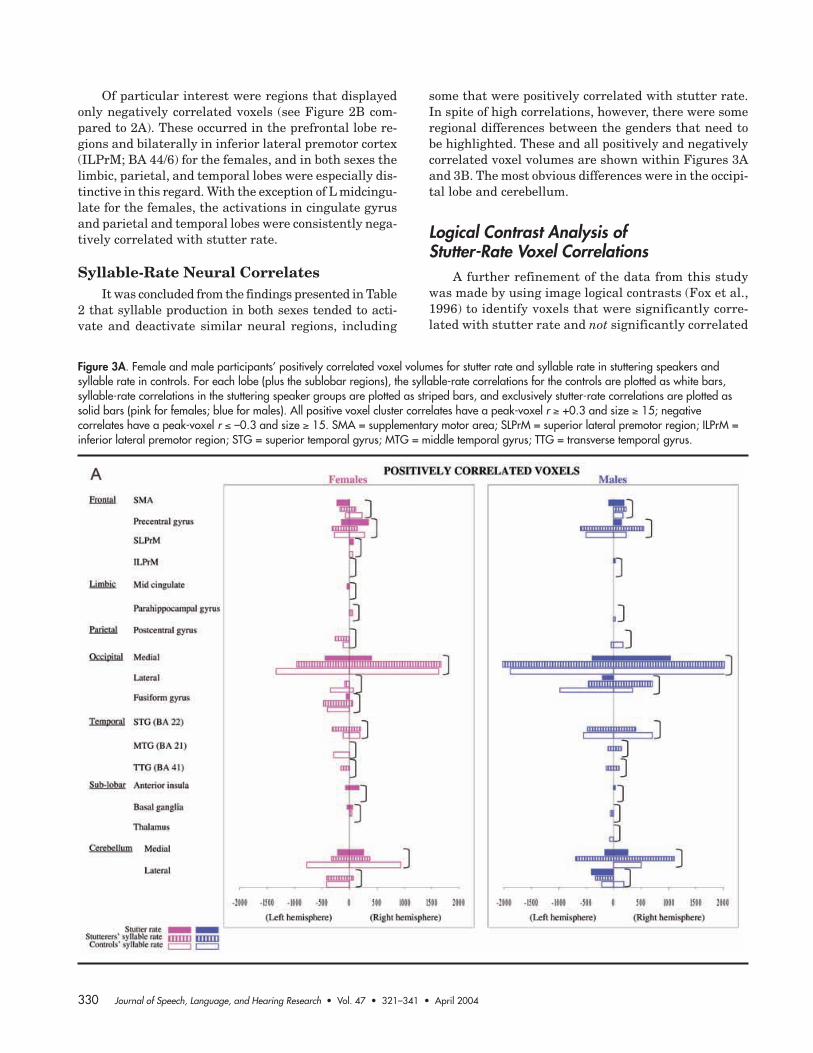
shows that the female stuttering speakers’ and controls’syllable rates were significantly correlated—for posi-tively correlated voxels, r(20) = .885, p < .01; for nega-tively correlated voxels, r(36) = .746, p < .01—across the54 regions, and this was also true for the male stutter-ing speakers and controls. By contrast, although therewas a strong relationship between regions that were posi-tively related to stutter and syllable rate—for females,r(22) = .646, p < .01; for males, r(18) = .825, p < .01—thiswas not the case for regions that were negatively corre-lated with stutter and syllable rate—for females, r(39) =.150, ns; for males, r(28) = .009, ns. The total negativelycorrelated voxels were also significantly different for stut-ter and syllable rate for both genders: for females, t(40) =4.150, p < .001; for males, t(29) = 4.221, p < .001. Conse-quently, the principal focus in the analysis of the find-ings in this article is directed mainly toward the stutterrate, rather than syllable rate results. However, syllablerate findings for the stuttering and control populationsare shown graphically (see Figures 3A and 3B, below).
Analysis of Total Performance-Correlated Voxels per Region
Because each region contains a single count of thecorrelated voxels for each group, it was not possible touse within-region parametric comparisons to determinethe statistical significance of any differences betweenthe sexes for the magnitude of within-region correlatedvoxels. However, unambiguous differences are obviouswhen significantly correlated voxels occur within a re-gion for one group but not for the other group. In thisinitial comparison, no attempt was made to identifyvoxels that were correlated with stutter rate and notwith syllable rate. Indeed, we hypothesize (see the Dis-cussion section) that when specific regions are correlatedwith both kinds of behaviors, then those particular re-gions may still be necessary for stuttering to occur. Whenspecific regions were later identified as correlated withstutter rate but not with syllable rate (see the LogicalContrast Analysis of Stutter-Rate Voxel Correlations sec-tion), then we hypothesize that these regions are notonly necessary but also sufficient for stuttering to oc-cur—at least during oral reading.
Stutter-Rate Neural CorrelatesThe within-region local maxima coordinates for
voxels that correlate with stutter rate are shown inTable 3. In most regions the coordinates relate to onecluster, but in some larger regions, such as cerebellum,multiple clusters were located. Table 3 shows only thelocal maxima coordinates for the largest and most highlycorrelated voxel cluster.
Table 3 is complemented by Figures 2A and 2B,which show, respectively, the total number of positively
and negatively correlated voxels (including multiple foci)within each region for the female and male stutteringspeakers.
Positive Correlations WithStutter Rate
Women and men both showed bilateral correlationsin the medial occipital lobes and in medial cerebellum,plus the men had correlations in L lateral cerebellumas well. Neither showed any correlations with the pari-etal lobe or the temporal lobe. The sexes were partiallyalike with respect to correlations with supplementarymotor area (SMA), primary motor cortex (M1) moutharea, and anterior insula, except the women showed bi-lateral correlations with M1 and anterior insula whilethe men were right-sided; the men only showed R BA 44(ILPrM) correlations and bilateral correlations with SMA(the women showed only L SMA). An obvious source ofthe low between-gender correlations for stutter rate re-ported in Table 2 was the four areas in which only thefemales showed significant correlations: R SLPrM, Lmidcingulate, L occipital fusiform gyrus, and bilateralbasal ganglia.
Negative Correlations WithStutter Rate
Figure 2B shows that there were more areas thatproduced negative correlations than positive correlationswith stuttering. Females and males were alike in thatthey both showed right-sided negative correlations withposterior cingulate and posterior parietal lobe and bi-lateral negative correlations with the superior tempo-ral (BA 22) and middle temporal gyrus (BA 39). Neithergender produced any negative correlations in cerebel-lum. The genders were partially alike in that the fe-males produced bilateral correlations and the men pro-duced unilateral correlations in the inferior frontal lobe(men, left-sided), anterior cingulate (men, right-sided),and the middle temporal lobe BA 21 (men, right-sided).The reverse was true for prefrontal areas where the menshowed bilateral negative correlations and the women’swere right-sided. Three other areas produced negativecorrelations in both genders, although in the right hemi-sphere for the females and the left for males: parahippo-campus, anterior parietal lobe, and the fusiform gyrus ofthe temporal lobe. In contrast, clearly differentiating be-tween females and males were six regions of negativecorrelation displayed by females only (L superior lateralpremotor cortex [SLPrM] and mostly left lateral occipitallobe, R midcingulate and R postcentral gyrus, and L basalganglia and bilateral thalamus). The only area with nega-tive correlations exclusive to males was BA 41, part ofthe group of negative correlations in the auditory com-plex that was common to both genders (see above).

328 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
Table 3. Local maxima Talairach coordinates per regions (Brodmann area number shown in parentheses) for females’ and males’ voxels thatare correlated positively or negatively with stutter rate.
Females Males
Left hemisphere Right hemisphere Left hemisphere Right hemisphere
Region r x y z r x y z r x y z r x y z
Positive correlationsFrontal lobe
SMA (6) .39 –2 –6 54 — — — — * .36 –6 –8 56 .42 12 0 58M1-mouth (4) .41 –44 –16 30 .35 52 –16 34 — — — — * .35 50 –16 28SLPrM (6) — — — — .32 44 –8 35 — — — — — — — —ILPrM (44/6) — — — — — — — — — — — — .35 44 22 1
Limbic lobeMidcingulate (24) .32 –4 10 36 — — — — — — — — — — — —
Occipital lobeMedial (18) * .41 –6 –89 –8 * .38 4 –86 10 * .40 –14 –93 14 * .43 2 –88 2Lateral (18) — — — — — — — — * .32 –33 –83 –14 — — — —Fusiform gyrus (18) .32 –20 –84 –21 — — — — — — — — — — — —
SublobarAnterior insula (13) .31 –31 18 4 * .35 44 20 1 — — — — .37 36 8 –2Basal ganglia .32 –14 –5 –1 .34 16 19 10 — — — — — — — —
CerebellumMedial * .35 –10 –78 –16 * .33 0 –72 –22 .34 –8 –72 –8 * .51 10 –70 –16Lateral — — — — — — — — * .42 –36 –66 –28 — — — —
Negative correlationsFrontal lobe
SLPrM (6) –.33 –30 –4 31 — — — — — — — — — — — —Prefrontal (8,9) — — — — * –.36 20 32 33 * –.34 –25 16 47 –.31 39 10 40ILPrM (44/6) * –.36 –39 13 21 * –.41 43 14 25 — — — — — — — —Inferior frontal (GFi) — — — — — — — — –.33 –29 2 28 — — — —
Limbic LobeAnterior cingulate (32) –.33 –18 38 9 * –.32 18 36 20 — — — — * –.37 12 39 –3Midcingulate (24) — — — — –.33 20 –4 33 — — — — — — — —Posterior cingulate (31) — — — — –.32 6 –54 26 — — — — * –.31 5 –60 24Parahippocampal (36) — — — — * –.36 37 –19 –9 –.31 –16 –16 –20 — — — —
Parietal lobeAnterior (40) — — — — –.31 61 –34 30 –.34 –49 –41 38 — — — —Posterior (7) — — — — –.34 4 –62 44 — — — — –.39 6 –54 36Postcentral gyrus (5) — — — — * –.31 36 –28 38 — — — — — — — —
Occipital lobeLateral (19) * –.38 –37 –69 13 –.32 32 –75 30 — — — — — — — —
Temporal lobeSuperior temporal gyrus (22) * –.31 –53 –41 –1 –.31 64 –32 8 –.31 –48 –56 26 –.35 54 –54 20Middle temporal gyrus (21) * –.34 –44 –30 –2 * –.30 60 –34 –8 — — — — * –.39 55 0 –13Middle (39) * –.35 –34 –70 28 * –.31 52 –56 22 –.31 –43 –67 22 –.33 51 –64 14Fusiform gyrus (37, 20) — — — — –.32 36 –38 –10 –.33 –46 –26 –16 — — — —Transverse gyrus (41) — — — — — — — — — — — — * –.35 53 –18 11
SublobarBasal ganglia –.33 –20 –6 22 — — — — — — — — — — — —Thalamus –.31 –26 –28 10 –.31 26 –28 9 — — — — — — — —
Note. Coordinates that correlate significantly with stutter rate and not also with syllable rate are in bold type. The asterisk indicates that multiple foci(with different voxel clusters) were identified in that particular area, but only the local maxima coordinate with the highest correlation is shown. Thedashes indicate that there is no significantly correlated voxel cluster in the homologous region within a gender. SMA = supplementary motor area;SLPrM = superior lateral premotor region; ILPrM = inferior lateral premotor region (left side = Broca’s area).

Ingham et al.: Brain Correlates of Stuttering 329
Figure 2. Female and male stuttering speakers’ total voxels per region that are correlated with stutter rate positively (Figure 2A) or negatively(Figure 2B). Voxels were located within 26 regions in the left hemisphere and 26 in the right hemisphere, with some regions containing bothpositively and negatively correlated voxels. All positive voxel cluster correlates have a peak-voxel r " +0.3 and size " 15; negative correlateshave a peak-voxel r # –0.3 and size " 15. SMA = supplementary motor area; SLPrM = superior lateral premotor region; ILPrM = inferiorlateral premotor region.
330 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
Of particular interest were regions that displayedonly negatively correlated voxels (see Figure 2B com-pared to 2A). These occurred in the prefrontal lobe re-gions and bilaterally in inferior lateral premotor cortex(ILPrM; BA 44/6) for the females, and in both sexes thelimbic, parietal, and temporal lobes were especially dis-tinctive in this regard. With the exception of L midcingu-late for the females, the activations in cingulate gyrusand parietal and temporal lobes were consistently nega-tively correlated with stutter rate.
Syllable-Rate Neural CorrelatesIt was concluded from the findings presented in Table
2 that syllable production in both sexes tended to acti-vate and deactivate similar neural regions, including
some that were positively correlated with stutter rate.In spite of high correlations, however, there were someregional differences between the genders that need tobe highlighted. These and all positively and negativelycorrelated voxel volumes are shown within Figures 3Aand 3B. The most obvious differences were in the occipi-tal lobe and cerebellum.
Logical Contrast Analysis ofStutter-Rate Voxel Correlations
A further refinement of the data from this studywas made by using image logical contrasts (Fox et al.,1996) to identify voxels that were significantly corre-lated with stutter rate and not significantly correlated
Figure 3A. Female and male participants’ positively correlated voxel volumes for stutter rate and syllable rate in stuttering speakers andsyllable rate in controls. For each lobe (plus the sublobar regions), the syllable-rate correlations for the controls are plotted as white bars,syllable-rate correlations in the stuttering speaker groups are plotted as striped bars, and exclusively stutter-rate correlations are plotted assolid bars (pink for females; blue for males). All positive voxel cluster correlates have a peak-voxel r " +0.3 and size " 15; negativecorrelates have a peak-voxel r # –0.3 and size " 15. SMA = supplementary motor area; SLPrM = superior lateral premotor region; ILPrM =inferior lateral premotor region; STG = superior temporal gyrus; MTG = middle temporal gyrus; TTG = transverse temporal gyrus.

Ingham et al.: Brain Correlates of Stuttering 331
266). The negatively correlated stutter rate voxels inthe cerebrum for females and males both showed rightlaterality (for females, L = 722, R = 1171; for males, L =350, R = 1136). The cerebellum contained no voxels thatwere negatively correlated with stutter-rate for eithergender. These broad trends were reflected in the regionalfindings.
Table 3 shows (in bold) the local maxima coordi-nates of the identified stutter-rate only voxel clusters—these are isolated in Table 4. All Table 4 coordinatesare also plotted within the BrainMap© database (Fox& Lancaster, 1996) and displayed in Figures 4A and 4B.
Table 4 shows that stuttering, for both sexes, wasassociated with positive correlations in anterior insulaand negative correlations in inferior frontal gyrus (GFi)and the auditory association area of the temporal lobe(BA 21 or 22). Anterior insula and BA 21 or 22 were bilat-eral in the females and right-lateralized in the males—only the GFi negative correlates were left-lateralized in
with syllable rate. In order to avoid identifying singlestutter-rate-only voxels embedded within syllable-rateclusters, the identified voxels were those that were iso-lated from syllable-rate voxels by at least five cubicvoxels. The clusters for stutter-rate-only voxels weremuch smaller than was the case for those that corre-lated with syllable-rate, and for that reason we used acluster criterion of three or more voxels.
The findings from the logical contrast analysis wereconditioned by some striking gender differences at abroad hemispheric level. The total voxels that were posi-tively correlated for stutter rate in the cerebrum showedbilaterality in the female stuttering speakers (L = 1040;R = 1039), but strong right-sidedness in the male stut-tering speakers (L = 681; R = 1428). That trend wasreproduced appropriately in reverse within the cerebel-lum, where the female stuttering speakers remainedessentially bilateral (L = 221; R = 260) and the malestuttering speakers showed left laterality (L = 552; R =
Figure 3B. Female and male participants’ negatively correlated voxel volumes for stutter rate and syllable rate in stuttering speakers andsyllable rate in controls. See Figure 3A caption for details.

332 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
Table 4. Regional coordinates in Talairach space for females’ and males’ positive and negative correlations with stutter rate that are not alsocorrelated significantly with syllable rate.
Females Males
Left hemisphere Right hemisphere Left hemisphere Right hemisphere
Region r x y z r x y z r x y z r x y z
Positive correlationsOccipital lobe
Lateral (18) .32 –33 –83 14 — — — —
SublobarAnterior insula (13) .31 –31 18 4 .35 44 20 1 — — — — .37 36 8 –2
.32 44 11 2
Basal ganglia (caudate nucleus) — — — — .34 16 19 10Basal ganglia (globus pallidus) .32 –14 –5 –1 — — — —
CerebellumMedial — — — — .51 10 –70 –16
.32 2 –51 –15
Negative correlationsFrontal lobe
Prefrontal (8,9) — — — — –.36 20 32 33
ILPrM (44/6) –.36 –39 13 21 –.41 43 14 25–.34 –34 34 5 — — — —–.32 –36 24 17 — — — —
Inferior frontal (GFi) –.33 –26 10 21 — — — — –.33 –29 2 28 — — — —
Limbic lobeMidcingulate (24) — — — — –.33 20 –4 33
Parahippocampal (36) — — — — –.36 37 –19 –9–.31 28 –38 –2
Parietal lobePosterior (7) — — — — –.34 4 –62 44
Occipital lobe Lateral (19) –.38 –37 –69 13 — — — —
Temporal lobeSuperior temporal gyrus (22) –.31 –53 –41 –1 –.31 64 –32 8Middle temporal gyrus (21) — — — — –.39 55 0 –13
–.35 61 –9 –12–.35 45 –13 –2–.34 66 –39 –1
Fusiform gyrus (37, 20) — — — — –.32 36 –38 –10Transverse gyrus (41) — — — — –.35 53 –18 11
–.35 49 –27 16
Note. The local maxima coordinates are shown for voxel clusters of three or more that are isolated from syllable rate voxels by a 53 voxel kernel.These include the coordinates shown in bold type in Table 3 plus other significantly correlated voxel clusters within a region (inferior frontal gyrus[GFi] is added here because smaller voxel cluster criteria were used for Table 4). Arrows in Figure 4 also identify coordinates shown in bold for bothsexes within an area. ILPrM = inferior lateral premotor region (left side = Broca’s area). The dashes indicate that there is no significantly correlatedvoxel cluster in the homologous region within a gender.

Ingham et al.: Brain Correlates of Stuttering 333
Figure 4. Brain images showing locations of the local maxima coordinates for female and male stuttering speakers’ voxel clusters that arecorrelated with stutter rate positively (Figure 4A) or negatively (Figure 4B). The data tabulated in Table 4 are plotted using the BrainMapdatabase (Fox & Lancaster, 1996). The arrows in the axial views of the figures point to local maxima coordinates for activations in the sameregion for both sexes. The Figure 4A arrows show coordinates from both sexes in right (R) anterior insula. The Figure 4B arrows showcoordinates for both sexes in left inferior frontal gyrus (GFi) and in R Brodmann’s area (BA) 21 or 22 (auditory association area).

334 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
both sexes. It is noteworthy that the three pairs of coor-dinates common to both genders (see arrows in Figure4) occurred within areas that are functionally related tospeech planning/production (anterior insula, auditoryassociation area, and GFi). Table 4 and Figure 4 alsoshow that there were within-area differences betweenthe genders in the location of the paired local maximacoordinates and the number of stutter-related coordi-nates within these areas.
There were also distinct differences between the re-gional positive and negative correlates produced by thefemale and male stuttering speakers. The females dis-played positive correlates in basal ganglia (R caudate,globus pallidus) and extensive negative correlatesthroughout the right hemisphere—in the prefrontal re-gion (BA 9) and in the limbic (BA 24 and 36), parietal(BA 7), and temporal (fusiform gyrus) lobes. Intriguingly,the female stuttering speakers showed strong negativecorrelations in the L occipital lobe (BA 19), whereas themale stuttering speakers displayed positive correlationsin the L occipital lobe (BA 18). Particular to the malestuttering speakers were positive correlations in R me-dial cerebellum and additional negative correlations inthe primary auditory area (R transverse gyrus or BA 41).
In the present study, cerebellum discriminated be-tween female and male stuttering speakers but was notclearly distinctive for stuttering (see Figures 3A and 3B).The original data on male stuttering speakers and con-trols (see Figures 3A and 3B and Fox et al., 2000) sug-gested that hyperactivity within cerebellum correlatedwith syllable production and stutter rate and was, there-fore, an essential feature of stuttering. However, thefemale stuttering speakers’ data showed much less cer-ebellum activity when compared to controls for syllablerate and less than the males for stutter rate. It may bethat high levels of cerebellum activity are gender re-lated rather than stuttering related.
Fox et al. (2000) interpreted their performance cor-relation findings in terms of state, trait, and compensa-tory effects of stuttering. The findings from that studywere derived from performance correlated, voxel-basedblood flow that was positively or negatively correlatedwith syllable rate and/or stutter rate. The same simplerule that Fox et al. used to categorize their findings intonormal, state, trait, and compensatory effects of stut-tering was used with the current data. In the controls,all effects were considered normal effects, by definition.The stutter-rate correlates that differed substantiallyfrom the syllable correlates in the controls were consid-ered state effects of stuttering. In the stuttering group,regional syllable-rate effects that resembled in voxelmagnitude the controls’ correlates were considered nor-mal. Other syllable-rate effects in the stuttering groupwere classified as trait or compensatory. Trait effects
were those that were intermediate in magnitude, thatis, between state effects and normal effects. Compensa-tory effects were those whose pattern was specific to theproduction of essentially nonstuttered syllables by thestuttering group, that is, responses present only insofaras the stuttering group spoke syllables fluently. The re-sults from both genders, shown in Table 5, highlight thestate effect gender commonalities in the frontal lobe (M1-mouth and SMA), in L anterior insula, and in R A2 (BA21/22). Compensatory effects showed possible gendercommonalities in L M1-mouth and, more decisively, inthe primary auditory area (A1). The additional analysisof the stutter-specific regions (see Table 4) appears tohave refined the state effect commonalities to three re-gions: R anterior insula, R BA 21/22, and an area withinL GFi. Arguably, L GFi, which is negatively correlatedwith stutter rate, is a state effect that was not found byusing the preceding system for identifying state-trait-compensatory effects.
DiscussionThe present study and its companion study (Fox et
al., 2000) offer added support to the suggestion thatdevelopmental stuttering is functionally related to ab-normalities within the speech planning/production andauditory systems during speech. The principal findingshave now been replicated in a different population us-ing an identical methodology. The specific region thatemerged as aberrantly activated during stuttering—andcommon to both genders—was R anterior insula (speechplanning), while R BA 21/22 (auditory processing and/or monitoring of speech) and L GFi (near to regions as-sociated with articulatory movement planning) wereaberrantly deactivated, the latter evolving in this studyfrom the more refined logical contrast analysis. Andbecause the logical contrast showed that these regionswere associated with stuttering, and not with syllableproduction, there is an additional reason why theymight be fundamentally related to the occurrence ofstuttering. Interestingly, two of these areas (R ante-rior insula, R BA 21/22) were significantly correlatedwith the rate of stuttering unilaterally in males andbilaterally in females. It is also noteworthy that essen-tially these same regional abnormalities appear to bepresent when stuttering speakers imagine stuttering.Ingham, Fox, Ingham, and Zamarripa (2000) showedthat most of these same regions were significantly acti-vated or deactivated even when stuttering speakersimagined they were reading aloud and stuttering—andtended to normalize when these individuals imaginedreading fluently. Thus, it would seem possible that acircuit involving these regions is essentially “primed”for stuttering to occur prior to the execution of the move-

Ingham et al.: Brain Correlates of Stuttering 335
ments of speech production. The overall findings con-tinue to support a blending of two broad theories of stut-tering: theories that emphasize abnormalities in speech-motor regions (e.g., Webster, 1993), especially in thenondominant (right) cerebral hemisphere (Travis, 1978),and theories that focus on the auditory system (e.g.,Stromsta, 1986; Yates, 1963).
Some issues that might compromise the reportedcomparison between the male and female stutteringspeakers are some potentially important differences be-tween them (see Table 1). The most obvious differenceswere in average age (females = 44 years; males = 32years) and chorus reading effects (only 7 of 10 femaleswere completely stutter-free during chorus reading). Themean oral reading stuttering frequency difference be-tween the genders (females = 7.3 stuttered intervals;males = 6.2 stuttered intervals) was not significant, t(28)= 1.313, ns, although differences in speech rate duringstuttered readings (males = 113 syllables; females = 78syllables) were significant, t(28) = 2.863, p < .01. Con-ceivably, some or all these variables could have accountedfor the obtained neurophysiologic gender differences.However, if persistent stuttering in adults implicatesspecific regions (or, more likely, regional interactions),then it seems unlikely that their functionality wouldchange markedly between the ages of 32 and 44 years.Indeed, the differences between these populations (sex,age, speaking rate, and chorus reading response) actu-ally strengthen the importance of the regions that wereassociated only with stutter rate and were common toboth sexes.
Gender and Neural RegionDifferences
The present findings for females and males all in-volve regions that have been generally reported to beassociated with oral reading tasks during PET studies(Fiez & Petersen, 1998)—albeit in studies using single-word utterances. Not a great deal is known about re-gions specifically engaged during continuous oral read-ing, although some information can be gleaned fromstudies that have investigated sentence generation dur-ing PET. Müller et al. (1997), for example, found that LBA 46 displayed strong activations during sentence gen-eration. However, in the present study neither Broca’sarea nor R BA 46 showed positive syllable correlationsin stuttering speakers or controls. This is consistent withtwo recent studies (Murphy et al., 1997; Wise, Greene,Buchel, & Scott, 1999) that also failed to obtain signifi-cant activation in BA 44/45 during speaking tasks withnormal speakers. It is noteworthy that there is consid-erable agreement between the areas identified in otherPET studies involving speaking tasks and the areas thatconsistently correlated with syllable production amongthe male and female controls and stuttering speakers(see Indefrey & Levelt, 2000). By contrast, much lessagreement exists between the areas in the male and fe-male stuttering speakers that were consistently corre-lated with stuttering and areas that are normally asso-ciated with speech production.
Anterior insula, so prominently associated withstuttering in the present study, has been implicated in
Table 5. Stuttering-specific responses and their physiological interpretations.
Females Males
Area Side Sign State Trait Comp. Side Sign State Trait Comp.
M1-mouth L + — — ? L + — — !
M1-mouth R + !* — — R + !* — !
SMA L + !* ! — L + !* — —SMA — — — R + ! — !
ILPrM — — — R + ! — —Postcentral L + — — ! — — —Anterior insula B + !* — — R + !* — —A1 L + — — !* B + — — !*A2 B – !* — — R – !* — —Basal ganglia B + ! ? — L + — — !
CBM L — — — L + ! — !
CBM R — — — R + — — !
Note. “Side” indicates hemisphere: R = right; L = left; B = bilateral. “Sign” indicates positive or negative correlates of stuttering. “State”, “Trait”, and“Compensatory” (“Comp.”) are physiological classifications, as explained in the text. ! = an area’s response was interpreted as fitting the physiologi-cal category; — = an area’s response was not so interpreted; ? = an area’s response categorization was equivocal; !* = an area’s response wascommon to both genders. The areas have been slightly expanded on those previously reported by Fox et al. (2000). SMA = supplementary motorarea; ILPrM = inferior lateral premotor region; A1 = primary auditory area (BA 41); A2 = auditory association area (BA 21/22); CBM = cerebellum.

336 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
motor programming by several studies, including stud-ies of speech planning/production (Dronkers, 1996; Fiez& Petersen, 1998; Fox et al., 1996; Paulesu, Frith, &Frackowiak, 1993; Raichle et al., 1994). Dronkers re-ported that lesions in L anterior insula (identified byCT or MRI) were consistent across a group that had beendiagnosed as displaying apraxia of speech, a speech dis-order characterized by deficient speech-motor planning(Wertz, Lapointe, & Rosenbek, 1991). R anterior insulahas been shown to be activated during difficult speakingtasks, such as translating (Price, Green, & von Studnitz,1999), and also during swallowing (Zald & Pardo, 1999).Equally interesting is evidence that R and L anteriorinsula activations have been found to correlate with ticfrequency in patients with Tourette’s syndrome (Sternet al., 2000), a disorder that shares many features incommon with stuttering (Abwender et al., 1998), includ-ing genetic associations (Comings & Comings, 1994).
The pattern of negatively correlated voxels withinthe temporal lobe was clearly different according to thegroup (stuttering speakers or controls) and speech in-dex (syllable rate or stutter rate). This finding suggeststhat stuttering speakers have a diminished capacity forauditory monitoring during speech—a conclusion con-sistent with reports of diminished auditory processingin stuttering speakers (Braun et al., 1997; Rosenfield &Jerger, 1984; Salmelin et al., 1998; Stromsta, 1972). Itis of interest that Zatorre, Meyer, Gjedde, and Evans(1996) review of PET studies on speech processing con-cluded that BA 21/22 shows CBF decreases during pas-sive listening tasks and increases during speech tasks.This is consistent with the present findings showingpositive correlations between BA 21/22 and syllable ratesfor both genders in both speaker groups, and it also un-derscores the abnormality of the negative correlates ofstuttering with R BA 21/22.2 Finally, a recent fMRI studyby Belin, Zatorre, Lafaille, Ahad, and Pike (2000) pro-vided compelling information on the importance of thisfinding. They identified areas on the R superior tempo-ral gyrus (STG) that were sensitive to the human voice;the coordinates for one of the most consistently activatedregions were very similar to the coordinates for the re-gion that was found to be negatively correlated with stut-tering in both genders in the present study.
Given that females may differ from males in thepattern of neural regions used during speech (Jaeger etal., 1998; Shaywitz et al., 1995), the gender differences
in the present study are not surprising. Syllable-rateproduction in the male stuttering speakers appeared tobe more right-sided, whereas in females who stutter itinvolved more bilateral positive correlations. Perhaps themost substantial difference was basal ganglia. Positivestutter-rate correlations with basal ganglia that occurredonly among female stuttering speakers may relate totheir relatively large area of stutter-rate frontal lobecorrelations (SMA and M1). Strong interactions have beenreported between basal ganglia and SMA/M1 during dif-ferent motor activities, including hand movement (Boekeret al., 1998). Because the female stuttering speakers andnot the controls showed positive syllable-rate correla-tions with basal ganglia and with SMA and M1, per-haps an interrelationship between these regions providesa compensatory system that aids the production of flu-ent speech during chorus reading—or any other fluency-inducing condition (Wildgruber, Ackermann, & Grodd,2001). The very prominent cerebellum syllable-produc-tion correlates that were identified in male stutteringspeakers (Fox et al., 2000) may also be gender-relatedbecause they were not found among the female stutter-ing speakers.
Findings in Relation to OtherBrain Imaging Studies
Other brain imaging studies of stuttering haveyielded findings that are relevant to those reported inthis article (Braun et al., 1997; De Nil et al., 2000; Sal-melin et al., 1998; Salmelin, Schnitzler, Schmitz, &Freund, 2000; Wu et al., 1995), although the task andmethodological differences among the studies compli-cate direct comparisons (Ingham, 2001). The most per-tinent of these studies was reported by Braun et al. Inthis mixed-gender PET study, right-handed adult stut-tering speakers (n = 18; 8 females) and controls (n = 20;8 females) completed a hierarchy of speaking tasks; twowere designed to reduce or eliminate stuttering andthe others were nonlinguistic oral-motor tasks. In gen-eral, the stuttering speakers showed either bilateralor right-lateralized activations. Comparisons betweentheir fluent and dysfluent tasks suggested that duringstuttering the speech-motor regions were disproportion-ately active in the right hemisphere, whereas the post-rolandic regions (including the auditory areas) wererelatively inactive. This study also involved a perfor-mance correlation analysis using weighted measures of“dysfluency,” but there were few similarities betweenregions that were strongly correlated with stutteringand those reported in the present study. The main ex-ception was that BA 22 was negatively correlated, as itwas in the present study. Braun et al. also obtainedreduced activations (relative to rest) when their stut-tering speakers made laryngeal and oral movements
2 The findings reported in the present study and in Fox et al. (2000)illustrate the differences that may occur between those obtained throughthe use of condition contrast and performance correlation methods. In theFox et al. (1996) study, which used condition contrasts, the deactivationsin BA 21/22 were found in the left hemisphere. In the current study andin the study reported by Fox et al. (2000), both of which used performancecorrelation, the BA 21/22 negatively correlated voxels principally occurredin the right hemisphere.

Ingham et al.: Brain Correlates of Stuttering 337
that were related to speech, which points to an unusualinteraction between stuttering speakers’ speech-motorbehavior in general and the A2 region. The differencesbetween the Braun et al. study and the present studymight help to explain the limited overlap in findings.There was no separation of CBF correlations with non-stuttered syllables spoken and with stuttering, whichmay partly explain why there were few resemblancesbetween Braun et al.’s correlation data and the corre-lation data obtained in the present study. Braun et al.also used a complex mixture of speech tasks to evokeand reduce stuttering. The task requirements in thefluency-inducing tasks were considerably different fromthe stuttering tasks (rate differences, rehearsed vs. un-rehearsed speech), but no behavioral data were re-ported, so it is not possible to learn the effect of thetasks on stuttering or speech rate. And finally, themixed-gender population might have created an addi-tional confound.
De Nil et al. (2000) included 10 right-handed malestuttering speakers and 10 age-matched controls whocompleted silent and oral readings from a series of sepa-rately presented single words (fixation on an X servedas a rest control). Only conditional contrasts and acti-vation data were reported. Some important similaritiesand differences with the findings of the present studywere reported. In contrast to the controls, during oralreading the stuttering speakers displayed large activa-tions in right insula that extended into the right pre-central and inferior lateral premotor cortex. Despite theabsence of deactivation data, a direct comparison be-tween the controls and stuttering speakers did revealthat L STG was significantly less activated in the lattergroup. De Nil et al. also obtained significant activationsin anterior cingulate cortex (ACC), whereas the presentstudy found extensive negative correlations with stut-tering in ACC. It is not clear why, although the use ofrelatively unfamiliar word lists may have some connec-tion with a novel task condition that Raichle et al. (1994)found prompted ACC activations. Nevertheless, if, as inthe present study, ACC is not activated during stutter-ing, then it is probably not necessary for stuttering tooccur.
The only other PET study on stuttering was reportedby Wu et al. (1995) on a mixed-gender population of 4right-handed stuttering speakers (1 female) and 4 con-trols (1 female). In this F-18 fluoro-deoxyglucose (FDG)PET study, chorus reading and solo reading conditionswere used, but no rest condition was used and scanningwas not extended to include all premotor regions. Strongcaudate activations were identified as consistently com-mon to the stuttering speakers only, but the study’s de-sign limitations severely restrict comparisons with find-ings from other PET studies on stuttering.
Two magnetoencephalography (MEG) studies bySalmelin and colleagues (1998, 2000) have yielded find-ings that partially complement those derived from thePET studies. Salmelin et al. (1998) first located unusualtemporal lobe processing among stuttering speakers thatis reasonably consistent with the deactivations reportedin the San Antonio studies (Fox et al., 1996, 2000;Ingham et al., 2000) and Braun et al. (1997), plus thepresent study. More recently, Salmelin et al. (2000) re-ported finding an unusual neural processing sequenceamong stuttering speakers during (nonstuttered) wordproduction: among controls the utterance was first pro-cessed in Broca’s area (where articulatory planning ispresumed to occur) and then in the lateral central sul-cus and premotor area (where motor preparation occurs);among stuttering speakers (n = 9; 2 females) this se-quence was reversed with premotor and motor process-ing occurring before articulatory planning. Salmelin etal. used only nonstuttered words, but the findings maybe consistent with PET findings of negatively correlatedvoxels in the stuttering speakers’ Broca’s area or GFiduring stuttered speech. Unfortunately, MEG onlyrecords near to the surface of the brain (12 mm belowthe skull in Salmelin et al.), so any involvement of deepercortical regions in this process (e.g., anterior insula andACC) cannot be identified.
An important issue that arises from the female andmale studies reported here is whether the apparentreciprocal relationships among the regions associatedwith stuttering, the seemingly “key regions” of R ante-rior insula, L GFi, and R BA 21/22, is unique to devel-opmental stuttering and, if so, what implications thishas for future research. These are considered key re-gions because, as mentioned above, they were the onlyareas that correlated with stutter rate alone, in bothgenders. It is not possible at present to fully resolvethe uniqueness issue because so few imaging studieshave used connected speech tasks. There are knowninterconnections among these regions (Mesulam &Mufson, 1985), which does make this interaction fea-sible. That is more manifest in Griffiths, Bench, andFrackowiak’s (1994) finding that when participantswere attending to a moving sound stimulus, there wasincreased CBF in right insula. Conceivably, therefore,anterior insula and the temporal lobe may work in con-cert to achieve induced fluency, but that is still onlypartially relevant to the interaction pattern that oc-curred during stuttering. In fact, an extensive searchof imaging reports reveals no study to date on patientor nonpatient populations that has recorded such areciprocal relationship between these two regions dur-ing speech.
An additional important consideration is the rela-tionship between neurophysiologic findings and those

338 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
reported in two recent neuroanatomic studies of stut-tering. Foundas et al. (2001) reported finding differencesbetween adult stuttering speakers (n = 15; 5 females)and controls (matched for sex, age, and handedness)using 3-D MRI, particularly within the perisylvian fis-sure. The most prominent were “a second diagonal sul-cus and extra gyri along the superior bank of the sylvianfossa” (Foundas et al., 2001, p. 207) that were uniqueto the stuttering speakers. It was also reported that thestuttering speakers’ planum temporales were enlargedand showed reduced interhemispheric asymmetry.Sommer et al. (2002) used diffusion tensor imaging tocompare stuttering speakers and matched controls andfound that they “differed significantly immediately be-low the laryngeal and tongue representation in the leftsensorimotor cortex” (p. 380). This region, they deter-mined, was consistent with that reported by Foundaset al.
In the present study, abnormal CBF negatively cor-related voxels were identified in a region (L GFi) that isreasonably proximate to the anatomic areas identifiedby Foundas et al. (2001) and Sommer et al. (2002). Pre-sumably, the other abnormal regions identified in thecurrent study would constitute the extended effects ofthis structural problem on other speech-related regions.Of course it is yet to be established that such structuralabnormalities also occur in fully recovered stutteringspeakers. If they are not present in this population, thentheir absence needs to be explained. Such explanationsmay also differ according to the population’s age of re-covery. If they are present, then it will be important toidentify the compensatory neural changes that mustoccur in order to support recovery.
ImplicationsThe important issue that emerges from these find-
ings is that replicating the Fox et al. (2000) study withfemales has contributed converging information on theregions that functionally control stuttering, and thatthere may be functional regions that differ between thesexes. We postulate that abnormally activated R ante-rior insula and an abnormally deactivated area withinL GFi and R BA 21/22 constitute the common “state ef-fects” and an interactive system sufficient for stutteringto occur. There also appear to be some gender-specificstate effects, mainly within the frontal lobe and cerebel-lum for males and in basal ganglia for females. The ef-fects we describe as “compensatory” (excessive activa-tion or deactivation during syllable production) werecommon to both genders in L M1 mouth and A1 andshowed gender differences in frontal lobe and cerebel-lum for males and in the sensorimotor area for females.
The findings also suggest that for stuttering to oc-cur within each gender then it is necessary for certain
neural regions to be activated or deactivated duringspeech. Given that developmental stuttering is a geneti-cally based disorder, these gender-specific regional ab-normalities may emerge as the epigenetic features ofthe disorder. In other words, the genetic expression ofthe disorder may be modulated by a gene function thathas been selectively activated or inactivated accordingto the individual’s sex. Thus, the phenotypic differencesthat have been observed (sex ratio, different early ratesof recovery) may derive from epigenetic neurophysiologicor neuroanatomic features.
As was previously mentioned, this is the first imag-ing study of stuttering that has focused on females whostutter, and the findings point to the need for furthergender-oriented investigations. One interesting possi-bility is that these neurophysiologic differences have astructural basis. The previously mentioned neuroana-tomic investigations of developmental stuttering haveactually included a small number of female stutteringspeakers (3 of 16 in Foundas et al., 2001; 5 of 15 inSommers et al., 2002), but no attempt was made withineither study to parse out the findings for each sex. Itshould be obvious that a neuroanatomic investigationof the differences, if any, that occur between a popula-tion of males with persistent stuttering and a popula-tion of female stuttering speakers could make an im-portant contribution to the growing understanding ofthe neurologic foundations of this disorder. That contri-bution will likely be even greater when this research isextended to those who have recovered from this disorder.
AcknowledgmentsThis work was supported by Grants 1RO1MH60246-01,
1RO1DC036801-A1, and PO1MH/DA52176 from theNational Institutes of Health and by Grant LM06858 fromthe National Library of Medicine. Special thanks are alsodue to Shawn Mitiken for assistance in preparation of thefigures.
ReferencesAbbey, E. (1975). The monkey wrench gang. New York: Avon
Books.
Abwender, D. A., Trinidad, K. S., Jones, K. R., Como, P.G., Hymes, E., & Kurlan, R. (1998). Features resemblingTourette’s syndrome in developmental stutterers. Brainand Language, 62, 455–464.
Belin, P., Zatorre, R. J., Lafaille, P., Ahad, P., & Pike, B.(2000). Voice-selective areas in human auditory cortex.Nature, 403, 309–312.
Blood, G. W., & Blood, I. M. (1989). Laterality preferencesin female and male stutterers. Journal of FluencyDisorders, 14, 1–10.
Bloodstein, O. (1995). A handbook on stuttering. San Diego,CA: Singular.

Ingham et al.: Brain Correlates of Stuttering 339
Boecker, H., Dagher, A., Ceballos-Baumann, A. O.,Passingham, R. E., Samuel, M., Friston, K. J., et al.(1998). Role of the human rostral supplementary motorarea and the basal ganglia in motor sequence control:Investigations with H2
15O PET. Journal of Neurophysiol-ogy, 79, 1070–1080.
Boring, E. G. (1950). A history of experimental psychology.New York: Appleton.
Braun, A. R., Varga, M., Stager, S., Schulz, G., Selbie,S., Maisog, J. M., Carson, R. E., & Ludlow, C. L.(1997). Altered patterns of cerebral activity during speechand language production in developmental stuttering: AnH2
15O positron emission tomography study. Brain, 120,761–784.
Buckner, R. L., Raichle, M. E., & Petersen, S. E. (1995).Dissociation of human prefrontal cortical areas acrossdifferent speech production tasks and gender groups.Journal of Neurophysiology, 74, 2163–2173.
Comings, D. E., & Comings, B. G. (1994). TS, learning,and speech problems. Journal of the American Academy ofChild and Adolescent Psychiatry, 33, 429–430.
De Nil, L. F., Kroll, R. M., Kapur, S., & Houle, S. (2000).A positron emission tomography study of silent and oralsingle word reading in stuttering and nonstutteringadults. Journal of Speech, Language, and HearingResearch, 43, 1038–1053.
Dronkers, N. F. (1996). A new brain region for coordinatingspeech articulation. Nature, 384, 159–161.
Fiez, J. A., & Petersen, S. E. (1998). Neuroimaging studiesof word reading. Proceedings of the National Academy ofSciences, USA, 95, 914–921.
Foundas, A. L., Bollich, A. M., Corey, D. M., Hurley, M.,& Heilman, K. M. (2001). Anomalous anatomy of speech-language areas in adults with persistent developmentalstuttering. Neurology 57, 207–215.
Fox, P. T., Ingham, R. J., Ingham, J. C., Zamarripa, F.,Xiong, J.-H., & Lancaster, J. (2000). Brain correlates ofstuttering and syllable production: A PET performance-correlation analysis. Brain, 123, 1985–2004.
Fox, P. T., Ingham, R. J., Ingham, J. C., Hirsch, T.,Downs, J. H., Martin, C., et al. (1996). A PET study ofthe neural systems of stuttering. Nature, 382, 158–162.
Fox, P. T., & Lancaster, J. L. (1996). Un atlas du cerveausur Internet [A brain atlas on the Internet]. Recherche,289, 49–51.
Fox, P. T., & Raichle, M. E. (1984). Stimulus rate depen-dence of regional cerebral blood flow in human striatecortex, demonstrated by positron emission tomography.Journal of Neurophysiology, 51, 1109–1120.
Fox, P. T., & Raichle, M. E. (1985). Stimulus rate deter-mines regional brain blood flow in striate cortex. Annalsof Neurology, 17, 303–305.
Frost, J. A., Binder, J. R., Springer, J. A., Hammeke,T. A., Bellgowan, P. S., Rao, S. M., & Cox, R. W. (1999).Language processing is strongly left lateralized in bothsexes: Evidence from functional MRI. Brain, 122, 199–208.
Fujiki, N., Naito, Y., Hirano, S., Kojima, H., Shiomi,Y., Nishizawa, S., et al. (1999). Correlation betweenrCBF and speech perception in cochlear implant users.Auris Nasus Larynx, 26, 229–236.
Good, C. D., Johnsrude, I., Ashburner, J., Henson,R. N. A., Friston, K. J., & Frackowiak, R. S. J. (2001).Cerebral asymmetry and the effects of sex and handed-ness on brain structure: A voxel-based morphometricanalysis of 465 normal adult human brains. Neuroimage,14, 685–700.
Griffiths, T. D., Bench, C. J., & Frackowiak, R. S.(1994). Human cortical areas selectively activated byapparent sound movement. Current Biology, 4, 892–895.
Harasty, J., Double, K. L., Halliday, G. M., Kril, J. J., &McRitchie, D. A. (1997). Language-associated corticalregions are proportionally larger in the female brain.Archives of Neurology, 54, 171–176.
Indefrey, P., & Levelt, W. J. M. (2000). The neuralcorrelates of language production. In M. S. Gazzaniga(Ed.), The new cognitive neurosciences (2nd ed., pp. 845–866). Cambridge, MA: MIT Press.
Ingham, R. J. (1988). Speech naturalness and stutteringresearch: A review. In S. E. Gerber & G. T. Mencher (Eds.),International perspectives on communication disorders(pp. 168–180). Washington, DC: Gallaudet UniversityPress.
Ingham, R. J. (1998). On learning from speech-motorcontrol research on stuttering. In A. K. Cordes & R. J.Ingham (Eds.), Treatment efficacy for stuttering: Asearch for empirical bases (pp. 67–101). San Diego, CA:Singular.
Ingham, R. J. (2001). Brain imaging studies of developmen-tal stuttering. Journal of Communication Disorders, 34,493–516.
Ingham, R. J., Bakker, K., Kilgo, M., & Moglia, R.(1999). Stuttering Measurement System (SMS). SantaBarbara: University of California. Retrieved March 31,2004, from http://www.speech.ucsb.edu/
Ingham, R. J., Cordes, A. K., & Gow, M. L. (1993). Timeinterval measurement of stuttering: Modifying interjudgeagreement. Journal of Speech and Hearing Research, 36,503–515.
Ingham, R. J., Fox, P. T., Ingham, J. C., & Zamarripa,F. (2000). Is overt speech a prerequisite for the neuralactivations associated with chronic developmentalstuttering? Brain and Language, 75, 163–194.
Ingham, R. J., & Packman, A. (1979). A further evaluationof the speech of stutterers during chorus- and nonchorus-reading conditions. Journal of Speech and HearingResearch, 22, 784–793.
Jaeger, J. J., Lockwood, A. H., Van Valin, R. D., Jr.,Kemmerer, D. L., Murphy, B. W., & Wack, D. S. (1998).Sex differences in brain regions activated by grammaticaland reading tasks. Neuroreport, 9, 2803–2807.
Kansaku, K., Yamaura, A., & Kitazawa, S. (2000). Sexdifferences in lateralization revealed in the posteriorlanguage areas. Cerebral Cortex, 10, 866–872.
Kidd, K. K., Heimbuch, R. C., & Records, M. A. (1981).Vertical transmission of susceptibility to stuttering withsex-modified expression. Proceedings of the NationalAcademy of Sciences, USA, 78, 606–610.
Lancaster, J. L., Glass, T. G., Lankipalli, B. R., Downs,H., Mayberg, H., & Fox, P. T. (1995). A modality-indepen-dent approach to spatial normalization of tomographic

340 Journal of Speech, Language, and Hearing Research • Vol. 47 • 321–341 • April 2004
images of the human brain. Human Brain Mapping, 3,209–223.
Lancaster, J. L., Rainey, L. H., Summerlin, J. L.,Frietas, C. S., Fox, P. T., Evans, A. C., et al. (1997).Automated labeling of the human brain: A preliminaryreport on the development and evaluation of a forward-transform method. Human Brain Mapping, 4, 238–242.
Lewis, D., & Sherman, D. (1951). Measuring the severityof stuttering. Journal of Speech and Hearing Disorders,16, 320–326.
Martin, R. R., Haroldson, S. K., & Triden K. A. (1984).Stuttering and speech naturalness. Journal of Speech andHearing Disorders, 49, 53–58.
McGlone, J. (1980). Sex differences in human brainasymmetry: A critical survey. Behavioral and BrainSciences, 3, 216–263.
Mesulam, M. M., & Mufson, E. J. (1985). The insula ofReil in man and monkey. In A. Peters & E. Jones (Eds.),Cerbral cortex (pp. 179–226). New York: Plenum Press.
Moore, W. H., Jr. (1993). Hemisphere processing research.In E. Boberg (Ed.), Neuropsychology of stuttering (pp. 39–72). Edmonton, Alberta, Canada: University of AlbertaPress.
Müller, R. A., Rothermel, R. D., Behen, M. E., Muzik, O.,Mangner, T. J., & Chugani, H. T. (1997). Receptive andexpressive language activations for sentences: A PETstudy. Neuroreport, 8, 3767–3770.
Murphy, K., Corfield, D. R., Guz, A., Fink, G. R., Wise,R. J., Harrison, J., & Adams, L. (1997). Cerebral areasassociated with motor control of speech in humans.Journal of Applied Physiology, 83, 1438–1447.
Nuck, M. E., Blood, G. W., & Blood, I. M. (1987). Fluentand disfluent normal speakers’ responses on a syntheticsentence identification (SSI) task. Journal of Communica-tion Disorders, 20, 161–169.
Pakkenberg, B., & Gundersen, H. J. G. (1997). Neocorti-cal neuron number in humans: Effect of sex and age.Journal of Comparative Neurology, 384, 312–320.
Paulesu, E., Frith, C. D., & Frackowiak, R. S. (1993).The neural correlates of the verbal component of workingmemory. Nature, 362, 342–345.
Petersen, S. E., van Mier, H., Fiez, J. A., & Raichle,M. E. (1998). The effects of practice on the functionalanatomy of task performance. Proceedings of the NationalAcademy of Sciences, USA, 95, 853–860.
Phillips, M. D., Lowe, M. J., Lurito, J. T., Dzemidzic,M., & Mathews, V. P. (2001). Temporal lobe activationdemonstrates sex-based differences during passivelistening. Radiology, 220, 202–207.
Price, C. J., Green, D. W., & von Studnitz, R. (1999).A functional imaging study of translation and languageswitching. Brain, 122, 2207–2208.
Pugh, K. R., Shaywitz, B. A., Shaywitz, S. E., Con-stable, R. T., Skudlarski, P., Fulbright, R., et al.(1996). Cerebral organization of component processes inreading. Brain, 119, 1221–1238.
Raichle, M. E., Fiez, J. A., Videen, T. O., MacLeod, A.M., Pardo, J. V., Fox, P. T., & Petersen, S. E. (1994).Practice-related changes in human brain functional
anatomy during nonmotor learning. Cerebral Cortex, 4,8–26.
Rosenfield, D. B., & Jerger, J. (1984). Stuttering andauditory function. In R. F. Curlee & W. H. Perkins (Eds.),Nature and treatment of stuttering: New directions (pp.73–87). San Diego, CA: College-Hill Press.
Salmelin, R., Schnitzler, A., Schmitz, F., & Freund,H. J. (2000). Single word reading in developmentalstutterers and fluent speakers. Brain, 123, 1184–1202.
Salmelin, R., Schnitzler, A., Schmitz, F., Jäncke, L.,Witte, O. W., & Freund, H. J. (1998). Functionalorganization of the auditory cortex is different in stutter-ers and fluent speakers. Neuroreport, 9, 2225–2229.
Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Con-stable, R. T., Skudlarski, P., Fulbright, R., et al.(1995). Sex differences in the functional organization ofthe brain for language. Nature, 373, 607–609.
Sidtis, J. J., Strother, S. C., Anderson, J. R., &Rottenberg, D. A. (1999). Are brain functions reallyadditive? Neuroimage, 9, 490–496.
Silbersweig, D. A., Stern, E., Frith, C., Cahill, C.,Holmes, A., Grootoonk, S., et al. (1995). A functionalneuroanatomy of hallucinations in schizophrenia. Nature,378, 176–179.
Sommer, M., Koch, M. A., Paulus, W., Weiller, C., &Büchel, C. (2002). Disconnection of speech-relevant brainareas in persistent developmental stuttering. Lancet, 360,380–383.
Stern, E., Silbersweig, D. A., Chee, K. Y., Holmes, A.,Robertson, M. M., Trimble, M., et al. (2000). A func-tional neuroanatomy of tics in Tourette syndrome.Archives of General Psychiatry, 57, 741–748.
Stromsta, C. (1972). Interaural phase disparity of stutter-ers and nonstutterers. Journal of Speech and HearingResearch, 15, 771–780.
Stromsta, C. (1986). Elements of stuttering. Oshtemo, MI:Atsmorts.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxicatlas of the human brain. New York: Thieme.
Travis, L. E. (1978). The cerebral dominance theory ofstuttering: 1931–1978. Journal of Speech and HearingDisorders, 43, 278–281.
Van Riper, C., & Hull C. J. (1955). The quantitativemeasurement of the effect of certain situations onstuttering. In W. Johnson & R. R. Leutenegger (Eds.),Stuttering in children and adults (pp. 199–206) Minneapo-lis: University of Minnesota Press.
Webster, W. G. (1988). Neural mechanisms underlyingstuttering: Evidence from bimanual handwriting perfor-mance. Brain and Language, 33, 226–244.
Webster, W. G. (1993). Hurried hands and tangled tongues.In E. Boberg (Ed.), Neuropsychology of stuttering (pp. 73–127). Edmonton, Alberta, Canada: University of AlbertaPress.
Wertz, R. T., Lapointe, L. L., & Rosenbek, J. C. (1991).Apraxia of speech: The disorder and its management. SanDiego, CA: Singular.
Wildgruber, D., Ackermann, H., & Grodd, W. (2001).Differential contributions of motor cortex, basal ganglia,

Ingham et al.: Brain Correlates of Stuttering 341
and cerebellum to speech motor control: Effects ofsyllable repetition rate evaluated by fMRI. Neuroimage,13, 101–109.
Wise, R. J. S., Greene, J., Buchel, C., & Scott, S. K.(1999). Brain regions involved in articulation. Lancet, 353,1057–1061.
Witelson, S. F., & Kigar, D. L. (1992). Sylvian fissuremorphology and asymmetry in men and women: Bilateraldifferences in relation to handedness in men. Journal ofComparative Neurology, 323, 326–340.
Wood, F., Stump, D., McKeehan, A., Sheldon, S., &Proctor, J. (1980). Patterns of regional cerebral bloodflow during attempted reading aloud by stutterers both onand off haloperidol medication: Evidence for inadequateleft frontal activation during stuttering. Brain andLanguage, 9, 141–144.
Wu, J. C., Maguire, G., Riley, G., Fallon, J., LaCase, L.,Chin, S., et al. (1995). A positron emission tomography(F-18) deoxyglucose study of developmental stuttering.Neuroreport, 6, 501–505.
Xiong, J.-H., Gao, J., Lancaster, J. L., & Fox, P. T.(1995). Clustered pixels analysis for functional MRIactivation studies in the human brain. Human BrainMapping, 3, 287–301.
Xiong, J., Rao, S., Jerabek, P., Woldorff, M., & Fox, P. T.
(2000). Inter-subject variability in cortical activationsduring a complex language task. Neuroimage, 12, 326–339.
Yairi, E., Ambrose, N., & Cox, N. (1996). Genetics ofstuttering: A critical review. Journal of Speech andHearing Research, 39, 771–784.
Yates, A. J. (1963). Recent empirical and theoreticalapproaches to the experimental manipulation of speech innormal subjects and in stammerers. Behaviour Researchand Therapy, 1, 95–119.
Zald, D. H., & Pardo, J. V. (1999). The functional neu-roanatomy of voluntary swallowing. Annals of Neurology,46, 281–286.
Zatorre, R. J., Meyer, E., Gjedde, A., & Evans, A. C.(1996). PET studies of phonetic processing of speech:Review, replication, and reanalysis. Cerebral Cortex, 6,21–30.
Received May 16, 2003
Accepted August 8, 2003
DOI: 10.1044/1092-4388(2004/026)
Contact author: Roger J. Ingham, PhD, Department of Speechand Hearing Sciences, University of California, SantaBarbara, CA 93106. E-mail: [email protected]





























