Embed Size (px)
Citation preview

Hydrobiologia 275/276: 1-14, 1994.E. Mortensen et al. (eds), Nutrient Dynamics and Biological Structure in Shallow Freshwater and Brackish Lakes.© 1994 Kluwer Academic Publishers. Printed in Belgium. 1
Brackish and freshwater shallow lakes - different systems or variationson the same theme?
Brian MossDepartment of Environmental & Evolutionary Biology, University of Liverpool, Liverpool, U.K.
Abstract
Saline lakes may be divided into two categories - those primarily saline because they are endorheic andthose secondarily saline (brackish) because of natural or anthropomorphic inputs of sea water. Endorheiclakes have greatly varying composition and salinity, occur in arid regions and are not as abundant forthat reason as exorheic lakes. The area of land surface over which they are the characteristic types, is,however, very considerable. Many endorheic lakes have communities of low diversity but the reason forthis is not necessarily high salinity - the world's largest endorheic lake, the ocean, has a very high di-versity - but the temporariness of many of these habitats which has led to wide and unpredictablefluctuations in salinity and to periodic drying out.
Secondarily saline lakes are rather different. They generally have an ionic composition which is thatof diluted sea water, and although they do not have the high diversity of marine communities, they oftenhave comparable, if slightly lesser, diversities with those of freshwaters. There are no firm salinityboundaries between fresh and brackish waters, only ones drawn (e.g. at 5 per mille total salts) byconvention.
Brackish waters have the particular interest that estuarine organisms including mysid shrimps andessentially marine members of the algal division, the Haptophyta, may colonize and establish themselvesin food webs otherwise characteristic of freshwaters. A comparison is made between the systems ofbrackish and freshwater lakes in the Norfolk Broadland of eastern England, most of which have un-dergone eutrophication. Although at first sight appearing very different, both fall into a similar patternwhen processes of change are considered. Increases in salinity serve as indirect switch mechanismswhich, like other but different mechanisms in freshwaters, may cause the lake community to change fromsubmerged plant dominance to phytoplankton dominance at intermediate states of nutrient loading. Thecurrent drought in eastern England is inducing small salinity changes that support this assertion.
Introduction them with gold to replace the female companion-ship they had been led to anticipate and in doing
Fasolt and Fafner were the two giants who, in so germinated the spore that was to immolate theDas Rhinegold, the first music drama of Richard gods and all with them in the final G6tterdam-Wagner's cycle, Der Ring des Nibelungen, built a merung, the twilight of the gods.great fortress, Valhalla, for Wotan and his fellow It would be invidious to compare Augustegods. For the gods it was a refuge and a mani- Thienemann and Einar Naumann with Fafnerfestation of immense power. The giants built it and Fasolt, who had, between them, a number ofwell but Wotan failed to honour the bargain he serious character defects, but the models of olig-had made for payment to the giants. He corrupted otrophic and eutrophic lakes that they created

2
(Thienemann, 1915; Naumann, 1917) have beensolid Valhallas of immense usefulness. With them,knowledge of only a few variables allows predic-tion of much else about many lakes. The summeroxygen profile is a key which opens many doors.The models are part of a holism which is theimportant but under-recognised gift of limnologyto ecological science. But the models are so pow-erful that they have had corrupting consequences.They have become prisons, maintained despiteevidence that many lakes simply do not fit easilyinto the standard patterns. There have long beendifficulties with tropical lakes, for instance (Thien-emann, 1931; Kilham & Kilham, 1989). The con-cepts are products of north temperate limnologyand, through their dominance, one importantgroup of lakes, those that are saline, has beenrelegated to a peculiar outsider status.
Such lakes are not peculiar, however. The areaof land surface over which saline lakes are thenormal kinds is very large (Williams, 1986) butthese areas are often arid, remote and little popu-lated. The perhaps unwarranted emphasis of lim-nology on non-saline systems simply reflects thehome-distribution of limnologists, predominantlyin Europe and the better watered parts of NorthAmerica.
Saline lakes have much to contribute to limno-logical theory. They fall into two groups - thosethat are primarily saline (or endorheic) becausethey lie in closed basins where evaporative con-centration of salts occurs (the world ocean can beregarded as the largest and most constant ofthese) and the second group which is secondarilysaline (or brackish) because of introduction anddilution of sea water into otherwise freshwatersystems. There is also a small group of second-arily saline lakes in which the salinity is derivedfrom dissolution of ancient salt deposits, laiddown in basins that were formerly endorheic, butwhich are now, as a result of continental drift andclimatic change, in exorheic areas. The 'flashes' inNorth Cheshire, UK, are of this kind.
Most endorheic basins are small and shallowbecause of their tendency to dry up. One excep-tion is Lake Turkana, a large and deep East Af-rican rift valley lake (Kolding, 1992). Their chem-
istry varies considerably depending on the localgeochemistry and the degree to which their solu-tions become saturated as evaporation has pro-ceeded. Because they are highly susceptible tovariations in weather between years, they are en-vironmentally unpredictable and often extreme.Biological diversity is usually low, though thoseorganisms that can cope may become very abun-dant. The Great Salt Lake in Utah is either pinkor green depending on the dominant two speciesof green algae, (Morgan, 1973; Stokes, 1984),which inhabit the slightly different parts of thelake to either side of the railway embankmentwhich bisects it. Other endorheic lakes in Africaand South America are dominated by densegrowths of the cyanophyte, Spirulinaplatensis, andanimal communities are similarly sparse in vari-ety, with Crustacea usually predominant. Theocean is the exception, for it is deep, permanentand perhaps the most constant in chemistry of allnatural waters; furthermore it harbours commu-nities of immense diversity. These characteristicsdepend on its vastness, covering two-thirds of theplanet and the fact that it has had an uninter-rupted history as a water body (of varying shape)for the past three or more billion years.
Brackish lakes, which contain diluted seawater, are rather different. Ocean water may enterthem through tidal rivers or by percolationthrough permeable deposits lying between thebasin and the sea coast. They do not necessarilylie in the arid areas which spawn endorheic lakesand hence may have a relatively steady ionic com-position and concentration. Estuaries representthe intermediate stage of greatly varying brack-ishness and themselves illustrate that no sensibleabsolute borderline separates fresh and brackishwaters, though conventionally a value of 5 permille 5000 mg 1- total salts) (Remane &Schlieper, 1971) is often taken as a convenientlower limit to brackishness.
In principle, therefore, a complete continuumof concentration must exist between waters ofnegligible ionic content and full strength sea water.Brackish waters, if of steady concentration, maydevelop much greater diversity than endorheiclakes and have communities which comprise rep-

3
resentatives from both freshwaters and the coastalsea. Estuaries likewise have a mixed communitycomposition but diversity is low compared withlakes of steady brackishness on a daily basis. Thelatter lakes are often regarded as inevitably dif-ferent from freshwater lakes simply because oftheir brackishness. This issue will be examined bycomparisons among lakes in the Norfolk Broad-land.
The Norfolk Broadland
The Norfolk Broadland in eastern England, con-tains examples of lakes of different but relativelyconstant brackishness on a daily and seasonalbasis (Moss, 1983). It is a system (Fig. 1) of riv-ers, close to the sea, in whose valleys are aboutfifty small, shallow lakes or Broads. The Broads
Fig. 1. Map of the Norfolk Broadland showing locationsmentioned in the text, and current mean salinities as parts permille (in parentheses).
were dug out as peat pits prior to the fourteenthcentury (Lambert et al., 1961), but then becameflooded to form a more or less completely inter-connected system extending inland from themouth of the estuary, Breydon Water, to its dis-tal extremities in the Broads and in the sources ofthe freshwater rivers.
Formerly there were probably three estuaries,but only the central one, Breydon Water, stillfunctions. The southernmost, formerly serving theR. Waveney, is now occluded by a lock which isnever opened and the river water is divertedthrough an artificial channel, the New Cut, toBreydon Water. The northernmost, which serveda small river, represented now by parts of theR. Thurne, either silted up or, more likely, wasdeliberately dammed, diverting water from thisarea indirectly to the central estuary and the coastvia the R. Bure. These changes occurred in the18th and 19th Centuries and were linked with thepolitics of navigation and commerce (George,1992). They leave us now, however, with a systemwith three groups of lakes.
The first is unaffected by sea water beyond thecontribution sea spray makes to rainfall near thecoast and includes only two, completely land-locked Broads, Upton Broad and CalthorpeBroad (Fig. 1). The second group is of Broadsconnected with the main rivers and is essentiallyfreshwater. But because the river gradients arevery small, physical tides are normally experi-enced well inland. On the highest tides, pulses ofsea water may extend far upriver (Fig. 2), inject-ing salts into all of the broads on occasion. Insevere cases there may be fish and invertebratedeaths but the events are short lived. Fluvial flowsrapidly dilute the salt out and the system rapidlyrecovers its normal freshwater state. The com-munities of these Broads. which are the majority,are essentially freshwater in nature in conse-quence.
The third group of Broads is associated withthe R. Thurne in the north of the area and in-cludes Hickling Broad, Heigham Sound, HorseyMere and Martham Broad. Heigham Sound andthe south eastern part of Hickling Broad may beremnants of the original northern estuary, for
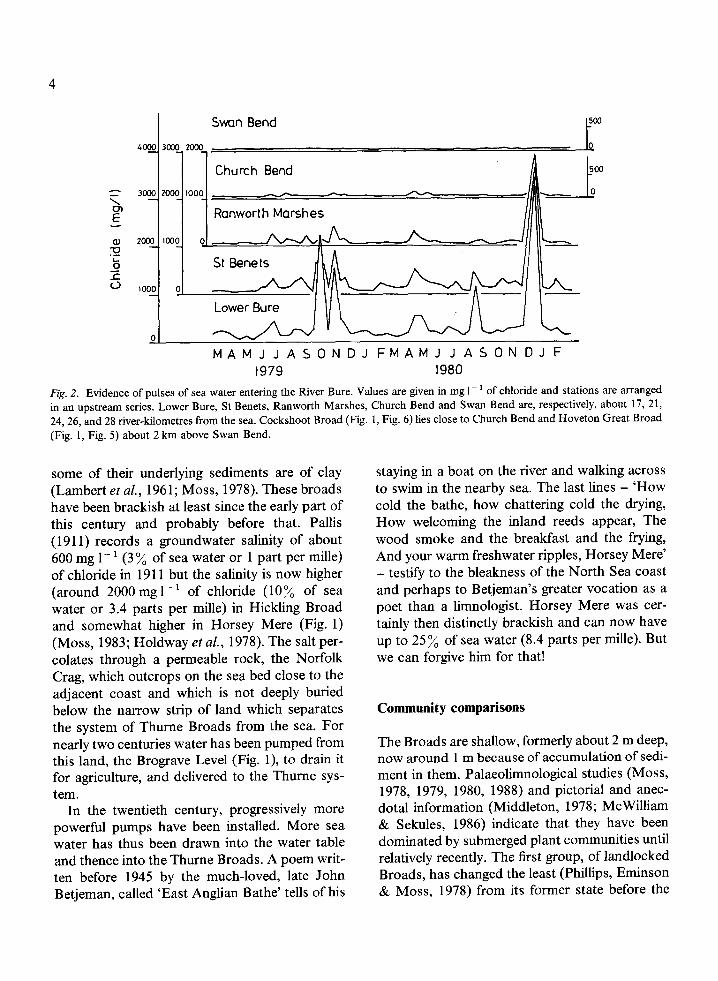
4
E
-o0
MAM J J ASON DJ FMAMJ J AS ON DJ F1979 1980
Fig. 2. Evidence of pulses of sea water entering the River Bure. Values are given in mg 1-' of chloride and stations are arranged
in an upstream series. Lower Bure, St Benets, Ranworth Marshes, Church Bend and Swan Bend are, respectively, about 17, 21,
24, 26, and 28 river-kilometres from the sea. Cockshoot Broad (Fig. 1, Fig. 6) lies close to Church Bend and Hoveton Great Broad
(Fig. 1, Fig. 5) about 2 km above Swan Bend.
some of their underlying sediments are of clay(Lambert et al., 1961; Moss, 1978). These broadshave been brackish at least since the early part ofthis century and probably before that. Pallis(1911) records a groundwater salinity of about600 mg 1- ' (3 % of sea water or 1 part per mille)of chloride in 1911 but the salinity is now higher(around 2000 mg 1' of chloride (10% of seawater or 3.4 parts per mille) in Hickling Broadand somewhat higher in Horsey Mere (Fig. 1)(Moss, 1983; Holdway et al., 1978). The salt per-colates through a permeable rock, the NorfolkCrag, which outcrops on the sea bed close to theadjacent coast and which is not deeply buriedbelow the narrow strip of land which separatesthe system of Thurne Broads from the sea. Fornearly two centuries water has been pumped fromthis land, the Brograve Level (Fig. 1), to drain itfor agriculture, and delivered to the Thurne sys-tem.
In the twentieth century, progressively morepowerful pumps have been installed. More seawater has thus been drawn into the water tableand thence into the Thurne Broads. A poem writ-ten before 1945 by the much-loved, late JohnBetjeman, called 'East Anglian Bathe' tells of his
staying in a boat on the river and walking acrossto swim in the nearby sea. The last lines - 'Howcold the bathe, how chattering cold the drying,How welcoming the inland reeds appear, Thewood smoke and the breakfast and the frying,And your warm freshwater ripples, Horsey Mere'- testify to the bleakness of the North Sea coastand perhaps to Betjeman's greater vocation as apoet than a limnologist. Horsey Mere was cer-tainly then distinctly brackish and can now haveup to 25% of sea water (8.4 parts per mille). Butwe can forgive him for that!
Community comparisons
The Broads are shallow, formerly about 2 m deep,now around 1 m because of accumulation of sedi-ment in them. Palaeolimnological studies (Moss,1978, 1979, 1980, 1988) and pictorial and anec-dotal information (Middleton, 1978; McWilliam& Sekules, 1986) indicate that they have beendominated by submerged plant communities untilrelatively recently. The first group, of landlockedBroads, has changed the least (Phillips, Eminson& Moss, 1978) from its former state before the
V

5
accelerated use of the area for agriculture and theincreases in population which have taken place inthe last two centuries (Moss, 1983). The twoBroads in this group have rich aquatic plant com-munities and clear water although they have un-dergone some agricultural eutrophication re-flected in changes in the community composition.The second (riverine freshwater) group has be-come eutrophicated to a greater degree because ofdischarge of sewage effluent into the rivers whichserve the lakes (Osborne, 1980, 1981; Moss et al.,1988). And the larger members of the third,brackish group have suffered guanotrophicationby black-headed gulls (Larus ridibundus) (Leahet al., 1978; Moss & Leah, 1982) which have mi-grated in winter to eastern England and whichhave roosted at night on Hickling Broad. Only thesmall Martham Broad, lying at the very end of thesystem and distant from the other Broads (Fig. 1)has escaped severe eutrophication and retains di-verse plant communities (Moss, 1983).
It has been conventional to regard the fresh-water and brackish groups of severely eutrophi-cated broads as fundamentally different. Chemi-cally they are distinctive, of course, withchlorinities from 50 to 90 mg -1 in the freshwatergroup except immediately after tidal surges whenconcentrations may rise to a few hundred mg 1 - '.In the brackish group, Hickling Broad has 1500-2000 mg Cl - ' and Horsey Mere around 6000 mg- . The least saline of the latter group, Martham
Broad has about 1000 mg - '. There is thus nor-mally no overlap in salinities between the twogroups.
The phytoplankton communities of the brack-ish group appear distinctive in that certain generalike Chaetoceros are present (Leah etal., 1978)and the marine Prymnesium parvum has had animportant role in causing fish kills (Holdway et al.,1978). The characteristic cyanophyte is a colonialAphanothece sp. Anabaenopsis sp. is also distinc-tively present. This contrasts with the freshwatergroup where Oscillatoria limnetica and Oscillatoriaagardhii are often predominant with Anabaenaspecies in summer (Osborne, 1981; Moss & Balls,1989). Aulacosira (Melosira) spp are common insummer in the freshwater group but absent from
the brackish whilst centric diatoms predominatein spring in the former (Timms & Moss, 1984;Moss & Balls, 1989) and pennate species (Di-atoma, Synedra) (Leah et al., 1978; Eminson &Moss, 1980; Moss, 1981) in the latter, though thismay be related to differences in flushing rates(Moss, 1977, 1983; Moss etal., 1989). Overall,the algae are smaller in the brackish system thanin the freshwater one (Moss, 1977) though thismay partly reflect a lower nutrient loading. It ap-plies however to species like Scenedesmus quad-ricauda that are shared between the two groups.
The submerged aquatic plant communities ofthe two groups do not differentiate them. Charo-phytes formerly dominated both systems and al-though Najas marina has been common in thebrackish group, it is, despite its name, equallyabundant in Upton Broad, one of the two isolatedfreshwater Broads. The floras of both groups arenow much diminished but the species character-istic of the more saline of the brackish group,Myriophyllum spicatum and Potamogeton pectina-tus, are essentially tolerant freshwater plantsrather than distinctive brackish indicators.
Certain invertebrates such as Eurytemoralacinulata and Neomysis integer were recordedfrom the brackish group earlier this century whensalinities were lower, but not from the freshwatergroup. However, many species of zooplanktonwere common to both including many Cladoceraand Copepoda (Gurney, 1905-1919). Other es-sentially brackish animals like E. affinis andPalaemonetes varians, together with Neomysis in-teger, now characterise the brackish group. Sedi-ment cores and contemporary studies have shownfewer taxa of Cladocera in the brackish groupthan in the freshwater (Stansfield et al., 1989).The fish communities of both groups are fresh-water in nature and dominated by roach (Rutilusrutilus) and bream (Abramis brama) with a varietyof other cyprinid fish plus pike (Esox lucius) anda very few perch (Percafluviatilis). Apart from thecatadromous eel (Anguilla anguilla) only flounder(Platichthys flesus) gives some indication of ma-rine connection and it is present in both groups(Wortley, 1974).
From a survey of community composition it is

6
thus clear that there are differences between thetwo groups but also many similarities. There aresome hints of reduced diversity in the brackishgroup, at least in the invertebrate communities.The sense is, however, of variations on a themerather than of two completely separate themes.
Patterns of change
Freshwater Broads
There appear to be some differences in the waysthat the two groups have changed as increasednutrient loading has affected them. The rates, de-grees and sources of eutrophication have differedbetween the two groups but the differences con-cern specific organisms and their roles. In thefreshwater group, as revealed in studies of sedi-ment cores, there was an initial switch fromcharophyte communities to aquatic plant com-munities dominated by vascular plants, often ofvigorous growth form, towards the end of thenineteenth century (Broads of the R. Bure) (Moss,1988) or in the early decades of the twentieth(Barton Broad on the R. Ant) (Moss, 1980). TheR. Yare Broads (Moss, 1979) may also haveswitched quite early. Palaeolimnological studies(Moss, 1983) suggest that these changes were inresponse largely to the disposal of sewage or itstreated effluent and also to increasing intensifica-tion of agriculture.
Although the nutrient loading increased furtherthereafter, there is evidence that the next majorchange, to phytoplankton dominance, which oc-curred rather abruptly in the 1950s and 1960s,required impacts additional to nutrient loading(Irvine et al., 1989; Balls et al., 1989; Moss, 1989,1991; Bales etal., 1993). Attempts to displacesubmerged plant communities in experimentalponds by high loading failed because it was foundthat the system buffered the nutrient additions(Irvine etal., 1989; Bales etal., 1989). Plant-associated Cladocera grazed any phytoplanktonthat developed and the water remained clear.Other observations and experiments (Timms &Moss, 1984) have shown that lily-dominated or
submerged plant communities in shallow lakesare not necessarily precluded by high nutrientloading so long as the buffering mechanisms areable to operate.
The switch to dense phytoplankton communi-ties and complete loss of aquatic plants that oc-curred in the freshwater broads thus requiredsome additional mechanism. There is a variety ofpossibilities including herbicide use, grazing by anintroduced rodent, the coypu, Myocaster coypus,and mechanical damage to the plants by boatingactivity. All of these would remove the refugeswhich the grazer Cladocera require to coexist withzooplanktivorous fish (Timms & Moss, 1984; Ir-vine et al., 1989). In the event, none of these ex-planations has proved satisfactory for the Broads.There is evidence, however, for a mechanismwhich directly removes the Cladocera, minimiz-ing algal grazing for a time and allowing the sys-tem to switch from a stable plant-dominated stateto an equally stable phytoplankton-dominatedone. In the freshwater Broads this may have beenthe use of organochlorine pesticides, to whichCladocera are particularly susceptible, on the sur-rounding land in the 1950s and 1960s (Stansfieldet al., 1989).
Different switch mechanisms (e.g. changes inthe fish community arising from differential lossof piscivorous fish following winter kills (Bron-mark & Weisner, 1992) may pertain elsewhere(Moss, 1991). I have, for example, noticed that inthe connected narrow canals which intersectDelft, as in many other Dutch cities, stretcheswhich are dominated by phytoplankton andwhich have turbid water, correspond with themotor boat route which transports visitors to seethe attractions of the town. Adjacent stretches,not on the boat route, have attractive, clear water,lily- and submerged plant-dominated communi-ties, occasionally supplemented, if not enhanced,by the electric fountains installed by day by en-terprising restaurant owners!
Beyond the stage of nutrient loading where al-ternative phytoplankton and aquatic plant domi-nated communities can potentially exist, there isprobably a unique state of plankton dominance(Fig. 3), where, at very high loading, large algal

Nutrient concentration-MedFum High
PHASE IClear water. Lowaquatic plant biomassbut high diversity.Low phytoplankton(nutrient- limited)
Increaseload -
Decrease load
IExtreme decrease innutrient load
I PHASE 21Clear water. Highaquatic plant biomass
c butlow to moderate Increase nutrient oad· diversity. Low (grazer- Increase organicX controlled] phytoplankton load?
I load?PHASE 3 l'. IO PHASE4 PHASE 5
Turbid water.Low As " Turbid water. No Turbid water Noaquatic plant biomass aquatic plants aquatic plants. Highand diversity(often High, cyanophyte phytoplankton ofabsent). High( NorP rich,grazer-res- light limited, pot-limited but potentially istent phytopl - entially grazablegrazable)phytoplankton ankton (?CO, chlorophyte algae
limited ) ,? Biomanipulatlon
|' Decrease external & internal nutrient loads
Fig. 3. Relationship between states of the ecosystem in shallow lakes, nutrient loadings, and approaches necessary for restora-tion.
species, including certain cyanophytes, dominatethe summer phytoplankton and which are poorlygrazed, if at all, by Cladocera (Moss, Stansfield& Irvine, 1991). There is also a fifth phase (Jeppe-sen et al., 1991) at extremely high nutrient con-centrations, where the phytoplankton becomesdominated by green algae rather than cyanophytesand in which zooplankton grazing may be a us-able restoration technique provided fish stockscan be kept low on a continuous basis. Equally,at very low nutrient loading, there is probably aunique state of plant dominance (Fig. 3) to whichthe system might return spontaneously with asuitably severe degree of reduction in nutrientloading. This is often practically impossible underthe present conditions in most countries thoughnot necessarily precluded (Simons et al., 1994).
It is possible to switch back from phytoplank-ton dominance to plant dominance, a processgenerally needing three steps. First the nutrientloads must be reduced to appropriate levels or thesystem changed in other ways (e.g. by increased
flushing) so that it supports phytoplankton com-munities that are grazable. Secondly zooplank-tivorous fish must be removed to allow Cladocerato increase, even in an unstructured environmentlacking in refuges for them, and to clear the water.And thirdly plants must be introduced and oftengiven protection from grazing birds to allow themto establish and create the refuges necessary forthe system to become stable when a normal fishcommunity is allowed to redevelop.
Brackish Broads
The pattern of change with eutrophication in thebrackish Broads is similar in some ways butshows what may be important differences. Thebrackish Broads do not receive sewage effluent.They lie in the remotest part of the region at thedistal end of the main river system. Although therewere small signs of eutrophication in the earlydecades of this century, reflected in increases in
Low
7
Very high

8
Cladophora, they appear to have retained theircharophyte communities and very clear water intothe 1960s. The nutrient loads they received mustnonetheless have been steadily increasing, for theflock of black headed gulls which roosted onHickling Broad was increasing (Moss & Leah,1982) and tidal movements allow mixing of thewaters of Hickling Broad, Heigham Sound andHorsey Mere. The War years of the 1940s and thepost-war period also saw some increase in theintensification of agriculture in the low lying Lev-els which form the catchment to the system.
Then, in 1969, there began a series of changes.There was a major fish kill that year, caused byan increase in the ichthyotoxic, haptophytan alga,Prymnesium parvum. The water became turbidwith phytoplankton, and the aquatic plant biom-ass declined markedly (Leah et al., 1978; Moss &Leah, 1982). The remaining plant community wasdominated by Myriophyllum spicatum and Pota-mogeton pectinatus with patches of Hippuris vul-garis. The zooplankton community was found tobe devoid of Cladocera though Cladocera hadbeen abundant formerly (Gurney, 1905-1919,1965).
Although the changes followed closely on themajor reduction in fish stock and were clearlysymptomatic of eutrophication, there was no ob-vious single event that could readily explain them.Nutrient loading from gulls had, from records ofgull numbers (Moss & Leah, 1982), been steadilyincreasing for many years but had not undergonea quantum leap around 1969. Moss & Leah (1982)proposed that some threshold of loading musthave been passed. It was realised that salinitylevels had also been increasing but less impor-tance was attributed to this in interpreting thechanges. More powerful pumps had been installedin the area during the 1960s to drain the landsufficiently for arable agriculture and these haddrawn water from deeper saline layers of theground water. Following further work in the late1980s (Moss et al., 1991; Bales et al., 1993; Irvineet al., 1993) it was realised that the increased sa-linity may have acted as a switch mechanism inconverting the system from plant to phytoplank-ton dominance.
Buffer mechanisms which had stabilised theformer, clear-water, plant-dominated system in-cluded grazing by Cladocera on the phytoplank-ton and also grazing of periphyton on the plantsby Neomysis integer, a brackish water mysid,which is also a predator on zooplankton (Irvineet al., 1990, 1993). Even at its greatest the nutri-ent loading was less than in the effluent-pollutedBroads and this may have served to delay theonset of the changes. The fish kills which her-alded the changes might have been expected toincrease the numbers of Cladocera and to en-hance the clarity of the water. This is the effectfish removal has in the freshwater Broads. How-ever, exactly the opposite effect was experienced.Cladocera disappeared and the zooplankton be-came dominated by a calanoid brackish watercopepod, Eurytemora affinis. It seems that the in-creased salinity was an extremely powerful switchmechanism leading to more complex conse-quences of eutrophication than in the freshwatergroup of Broads. Many Cladocera, particularlydaphnids, do not survive at salinity values above3-4 per mille (e.g. see Dwyer et al., 1992; Jeppe-sen et al., 1994) which is what is presently expe-rienced in Hickling Broad in summer, other Cla-docera may be more tolerant and there are somewhich survive at quite high salinities (see Alonso,1990, 1991; Aladin, 1991). There also appears tobe a great deal of genetically based ecotypic vari-ation within zooplankton species with differentialadaptation to local conditions (Weider & Hebert,1987; Mort, 1991).
In the 1980s, following the almost completeloss of aquatic plants during the 1970s, there hasbeen a recovery of the plant biomass though it isstill dominated by vascular plants rather thancharophytes (Bales et al., 1993). There has beenan apparent increase also in Neomysis integer. Fishstocks remain low perhaps because fish avoidmoving into the Broad because of low levels ofPrymnesium toxin in the water. There have beenno significant fish kills for some time and Prym-nesium is now scarce in the phytoplankton (Baleset al., 1993). The gull flock has declined (owing toclosure of a nearby municipal waste tip) andphosphorus levels have declined proportionately.

9
However the water remains turbid, salinities re-main high and Cladocera are still absent. All ofthis provides clues to the mechanisms operatingwhen the major changes were occurring in the1970s. Stabilisation of the former plant commu-nity probably depended not only on grazing ofphytoplankton by Cladocera but also on grazingof periphyton by Neomysis. Grazing of both kindsprobably declined. Large invertebrates are sus-ceptible to Prymnesium toxin just as are fish.Neomysis probably declined for this reason in the1970s and the plants became doubly vulnerable.Now that Prymnesium numbers have decreased,Neomysis has recovered and has probably rein-stated effective periphyton grazing. The plant bio-mass is increasing and this may be because thelight climate at its surface has improved despiteno change in the turbidity of the water column.The decline in Prymnesium might be attributed todecline in nutrient loading either of inorganic nu-trients or of the several vitamins it requires andof which decaying guano was undoubtedly a po-tent source. Moss (1992), Irvine et al. (1993) andBales et al. (1993) provide a detailed treatment ofthese issues.
In explaining the changes on eutrophication inboth the freshwater group and in the brackishgroup therefore, it seems that increased nutrientloading alone is insufficient. There must be addi-tional switch mechanisms. The apparent differ-ence between the two groups is that the switchesin the freshwater group, and affecting other fresh-water systems, have needed to be only tempo-rarily present whilst that in the saline group (in-creased salinity) has been effectively permanent.One of the conditions for restoring shallow lakesto plant dominance is that the forward switchmust be removed so that the reverse switch canbe operated (usually by biomanipulation). If in-creased salinity is truly permanent then full res-toration of the brackish Broads will not be pos-sible. However in the Broads' area, reduction ofsalinity is, in principle, easily possible. It simplyinvolves reduction in the intensity of pumpeddrainage in the area. In turn this means changesin agricultural land use back to stock farming andpasture rather than arable. The area has been
designated an Environmentally Sensitive Area(ESA) which means that grants are available tosupport this. Little has yet happened but, in therapidly changing state of agriculture in the Euro-pean Community, the future is optimistic. In sucha case, salinity will have become a temporaryswitch mechanism and the changes in both thefreshwater and brackish groups of broads appearin principle to be rather similar and not funda-mentally different from one another.
Drought and its effects on the freshwater broads
Since 1989, rainfall in eastern and southern En-gland has been well below the long term averageand some of its effects emphasise the rather smalldifferences that exist between the freshwater andbrackish groups of Broads despite their obvioussalinity differences. The system of Broadland riv-ers is a tidal one and salt water normally pen-etrates upriver on very high tides especially whenthese occur under meteorological conditionswhich give rise to surges in the North Sea. Thesouthern part of the North Sea geological basin,of which Broadland is a part, is also sinking at therate of several mm per year (Funnell, 1979). Thismeans that sea water salinity is affecting the riv-ers further and further upstream. Long termrecords from a drinking water intake on the riverBure show a slight but significant increase inbackground chlorinity and in the incidence oflarge peaks associated with surge tides (Fig. 4).Shots of quite high salinity water enter the Broadseach autumn and winter but the effects decreaseupstream (Fig. 2) and the salinity is rapidlywashed out by continued river flow. The recentdrought, with reduced fluvial flows. has meantthat winter salinity has been removed less rapidlyin some areas and less effectively in others.
A case of the former is Hoveton Great Broad,lying in the upper part of the middle R. Bure(Fig. 1). Phosphate is removed from the effluentof the larger of the upstream sewage treatmentworks which potentially affect the Broad, and a2 ha enclosure has been established in the Broad.The aim is to build up areas of plant bed sequen-

11I _/ X .6
A **A/~j ~ --
@140
XI \, *
0, 11 1. "' _ .,*
1952 1953 1954 1955 \. 1956 1957 1958 , 1959
50 * *
~~0 ~~ ~04 2
1.960 1961 1962' 1963 1964 )1965,' 1966 144\ 1968
50 * i-. ** , , S
1969 1970 1971 19720, , . ..
· *.--- ' \,/ *'- - -. ,'* '.. 1973 ' 197/ ' 1975 ' 1976 ' 1977
1978 1979 1980 1981 Fig. 4. Chloride concentrations at a water supply intake on the River Bure from 1951 until 1981. The intake lay close to ChurchBend (Fig. 2) and was abandoned for routine use in the early 1980s because of the increasing risk of salt contamination.
tially thus avoiding the need to remove fish fromthe entire Broad to effect a switch back to plantdominance (Moss, 1990). Fish have been re-moved from the enclosure, which is made of solidwalls fixed deep into the sediment but with 1 mmmesh stainless steel panels to allow free inter-change of water between the enclosure and themain 20 ha Broad. In 1991, before the wall wasundermined and fish re-entered, the water clearedas daphnid populations developed. After repair ofthe wall in early 1992 and removal of virtually allfish, Daphnia failed to increase. However Neomy-sis integer was common in the enclosure until Maywhen it disappeared and Daphnia hyalina in-creased, clearing the water and allowing growthof aquatic plants. Figure 5 shows details of chlo-ride changes in the Broad and at a river sitedownstream of it in a non-drought year (precipi-tation 610 mm) and in the drought years of 1990-1992. A much larger injection of sea water oc-curred in 1990 than is usual, taking theconcentration of chloride in the Broad up to atleast 358 mg 1- . However this was washed outin the ensuing month and values returned to
around 60 mg 1- . A similar shot of saline waterentered in autumn 1991 and a further one in Janu-ary 1992 which was only slowly diluted as rain-fall and river flow continued to be low. The Broadentered spring with more than 100 mg 1- 1 chlo-ride and only after several weeks had chlorideconcentrations returned to the background 55-60 mg 1- 1. Neomysis disappeared subsequently inearly June. [In the higher salinity Hickling Broadit is, of course, present for the whole year]. Somefish remained in the enclosure and may have con-tributed to the demise of the mysid but since itcoexists with somewhat larger fish stocks inHickling Broad, it is likely that its disappearancewas related to the return to freshwater conditions.
The injections of sea water have been less ef-fectively removed from Cockshoot Broad whichlies about 6 km downstream of Hoveton GreatBroad. Cockshoot Broad has been isolated fromthe River Bure by dams and a sluice since 1982(Moss et al., 1986). Its water supply comes froma small stream draining the land to the south ofthe broad but, in winter, surge tides occasionallyovertop the dams bringing saline water into the
10
50
0
- , .'6· - - p -- 0-00-9 --*3-
39P
oE
_)
0
50
0
. .I I , I _ I I

11
( 150
O 100
r 50
0
150
100
50
.l
Hoveton Gt Broad & R. Bure
1979Rainfall 610 mm /\ /\Re / a '
Broad
1990Rainfall 472 mm
I I I I I2574-' '969
(2537
Cockshoot Broad
Q10
0
I I I I I I I I I '. . I I361- ' 7 a. "2789
1991 /284, \ .Rainfall 467mm | Neomysisnot detect. /\ /
o ,n 0 000 0
NE
:9r-
zU
I I I I I I I I I
1992 Drought continueso Neomysis present in fish-removal exclosure
\'- o Neomysis absente"o~ a; !o~-
J I- M A M J J A b U N U
Fig. 5. Changes in the chloride concentrations of the RiverBure at Church Bend, and in Hoveton Great Broad, some5 km upriver, in a normal year (1979) and in recent droughtyears (1990-1992). A large enclosure is in place in the Broad,from which fish have been exhaustively removed but throughwhich main broad water and hence river water, freely circu-lates.
Broad. This has normally been washed out by thestream flow which flushes the Broad about fourtimes a year in non-drought periods and a back-ground chloride concentration of about 60-70 mg1- is restored. Figure 6 gives details of chlorideconcentrations in the inflow stream, which re-mained steady and only slightly increased abovethe background value in the drought years of 1991and 1992, and in the Broad. The Broad receivedinjections of saline river water (see Fig. 5 for riverdata) in the winter periods of 1990/91 and 1991/92which took the chloride concentrations to over400 mg 1- in early 1991 and over 500 mg 1- 1 in1992. In 1991, reduced summer dilution still leftconcentrations around 150 mg 1- and this valuestill pertains in 1992. Neomysis integer was not
100
0
30Q
200
100
0
300
200
100
o4F16Pre-isolation 1979Rainfall 610 mm
Broad I/ Stream)Broad /Stream')
I I I I I I I I I I IPost-isolation 1987Rainfall 616 mmNeomysis absent
! W 9° 8 d 8Q 86 8'. g 8
I I I I I I I I I I I 0
\ Fish removed 1991 I -j°" Rainfall 467mm / u/
_ioa . Neomysis absent
··. ....... -. . ........ .0 . .®®*·iee ~ e® · iii i·® · ® · e·
\ Fish removed Drought continues 1992-f aNeormysis abundant lup to800/ m2 )
\ until mid August
I I 1..I. I..I I , I . I-~l 11J I M A M J J A S U N'U'
Fig. 6. Changes in the chloride concentrations of CockshootBroad in meteorologically normal years before the Broad hadbeen isolated from the river by dams (1979) and after suchisolation (1987) and in the drought years of 1991 and 1992.Fish have been exhaustively removed from the Broad since1998/89. The water supply to the Broad comes from a smallstream, data for which are shown. This stream is not influ-enced by events in the main river from which the Broad hasbeen isolated. In extreme tidal events, water overtops theisolating dams and brings saline water into the Broad.
detected in the Broad in 1991 and the water, fromwhich the fish had been removed, and which sup-ported large populations of Daphnia hyalina andother cladocerans, remained very clear. In 1991Neomysis was very abundant, Daphnia was virtu-ally absent and chlorophyll a values (around50 pg 1-') were much higher than in previousyears (< 10 ,g 1- ) for the early to mid summerperiod. The water was turbid with algae. Neomy-sis disappeared in mid August when chloride con-centrations remained at about 140 mg 1- . I donot know why Neomysis was not present in 1991but its simultaneous appearance in both Hoveton
150
50
0
150
100
50
0-
E
I . . . . . . .... . .
I . . . . . . . . . . .. : . .. .. ....
I
l .l. . I . l. l.I I I I I I I
I I I I . I . I I I . I I I I -
I·· · ·

12
Great and Cockshoot Broads in 1992 suggeststhat it came in with the sea water injection up theRiver Bure in January 1992. The mysid is nor-mally present in the Thurne system, which has itsconfluence with the Bure a few km downstreamof Cockshoot Broad (Fig. 1). Thurne water maypass directly downstream to the estuary or beturned up into the Bure depending on the riverflows and tidal state. There is thus a great deal ofstochasticity concerning the chances of Neomysisbeing brought upriver in sufficient numbers to es-tablish a population.
What does seem clear, however, is that salini-ties equivalent to only 50-150 mg 1- of chloride(0.26-0.78 % of sea water), and hence well belowconventional definitions of brackishness (Remane& Schlieper, 1971), seem able to convert a fresh-water food web into something that has key fea-tures of the supposedly very different brackishBroads in the importance of an essentially estua-rine mysid. The conversion appears to have beenonly temporary but it suggests that only a mod-est further increase in salinity might lead to per-manent breeding populations of the mysid. Thecauses of the drought which is affecting southernand eastern England may or may not be linkedwith changes in global climate but the drought ismore severe than that in 1975/76, which was cal-culated to have a return time of several hundredyears. It is thus not inconceivable that permanentsignificant increases in salinity in the freshwaterbroads could be a feature of the future.
If drier weather is to become a long term fea-ture of the area, restoration strategies for thefreshwater Broads may need to become even morecomplex. Following nutrient control, simple tem-porary removal of all fish, from either wholeBroads or enclosures within them, may allowNeomysis, in the absence of refuges for Cladocera,to flourish to the detriment both of Daphnia andwater clarity. We will need information on theoptimal fish communities which will reduceNeomysis whilst not making severe inroads onDaphnia. This information is not available andwill be difficult to establish.
It might seem that in contemplating closelymanaged fish communities, we are planning a
parallel biological fix to the technological fix ofresponding to environmental problems caused bytechnology with further technology. In such a casewe might face further and further complications,and the already difficult goal of full restoration ofshallow lakes will more distantly recede. Ulti-mately control of cause is preferable to control ofsymptoms, however sophisticatedly this is done.It may be, then, that we will have to control notonly our nutrient inputs but also the weather -most preferably not by any arrogant schemes tomanipulate it actively but by changing our soci-eties in ways which reduce our present pollutionof the atmosphere. In his negotiations with Fafnerand Fasolt, Wotan wanted to retain a ring fromthe hoard of gold with which he proposed to paythem. The ring was supposed to give great pow-ers to control events. Erda, the primeval earthgoddess emerged from the rocks to counsel himotherwise: 'Flieh' des Ringes Fluch! Rettungslosdunklem Verderben weiht dich sein Gewinn' [Flyfrom the Ring's curse! Utter ruin past salvation,its gain will bring you.] On that occasion, Wotan,ever responsive to the charms of women, waswise enough to listen.
Acknowledgements
I am grateful to a great many present and formercolleagues, discussion with whom has contrib-uted greatly to the understanding, though still veryimperfect, that we now have about the aspects ofthe Broadland system discussed above. Particu-lar among these are Drs G. L. Phillips, K. Irvineand M. Bales and Ms Julia Stansfield. I am alsograteful for the perspective supplied by interac-tion with colleagues from Denmark, The Nether-lands, and Norway in the Northern EuropeanShallow Lakes Database Group. I am also grate-ful for logistic support from the National RiversAuthority and the Broads Authority. Data areused in this paper from projects funded by theBroads Authority, the Natural Environment Re-search Council (UK), the European Commissionand the National Rivers Authority.

13
References
Aladin, N. V., 1991. Salinity tolerance and morphology of theosmoregulation organs in Cladocera with special referenceto Cladocera from the Aral sea. Hydrobiologia 225/Dev.Hydrobiol. 71: 291-300.
Alonso, M., 1990. Anostraca, Cladocera and Copepoda ofSpanish saline lakes. Hydrobiologia 197/Dev. Hydrobiol.59: 221-231.
Alonso, M., 1991. Review ot Iberian Cladocera with remarkson ecology and biogeography. Hydrobiologia 225/Dev.Hydrobiol. 71: 37-44.
Balls, H. R., B. Moss & K. A. Irvine, 1989. The loss of sub-merged plants with eutrophication I Experimental design,water chemistry, aquatic plant and phytoplankton biomassin experiments carried out in ponds in the Norfolk Broad-land. Freshwat. Biol. 22: 71-87.
Bales, M., B. Moss, G. L. Phillips, K. Irvine & H. Stansfield,1993. The changing ecosystem of a shallow, brackish lake,Hickling Broad,Norfolk II Long term trends in waterchemistry and ecology and their implications for restorationof the lake. Freshwat. Biol. 29: 141-165.
Bronmark, C. & S. Weisner, 1992. Indirect effects of fishcommunity structure on submerged vegetation in shallow,eutrophic lakes: an alternative mechanism. Hydrobiologia243-244/Dev. Hydrobiol. 79: 293-302.
Dwyer, F. J., S. A. Burch, C. G. Ingersoll & J. B. Hunn, 1992.Toxicity of trace element and salinity mixtures to stripedbass (Morone saxatilis) and Daphnia magna. Envir. Toxicol.Chem. 11: 513-520.
Eminson, D. F. & B. Moss, 1980. The composition and ecol-ogy of periphyton communities in freshwaters 1 The influ-ence of host type and external environment on communitycomposition. Br. phycol. J. 15: 429-456.
Funnell, B. M., 1979. History and prognosis of subsidenceand sea level changes in the lower Yare valley, Norfolk.Bull. Geol. Soc. Norfolk 31: 35-44.
George, M., 1992. The land use, ecology and conservation ofBroadland. Packard Publishing ltd, Chichester, 558 pp.
Gurney, R., 1905-1919. Unpublished manuscript notebooks.Held at the castle Museum, Norwich, U.K.
Gurney, R., 1965. Crustacea in Ellis, E. A. The Broads, Col-lins, London, 325 pp.
Holdway, P. A., R. A. Watson & B. Moss, 1978. Aspects ofthe ecology of Prymnesium parvum (Haptophyta) and waterchemistry in the Norfolk broads, England. Freshwat. Biol.8: 295-311.
Irvine, K., B. Moss, M. Bales & D. Snook, 1993. The chang-ing ecosystem of a shallow, brackish lake, Hickling Broad,Norfolk, U.K. I Trophic relationships with special refer-ence to the role of Neomysis integer Leach. Freshwat. Biol.29: 119-139.
Irvine, K., B. Moss & H. Balls, 1989. The loss of submergedplants with eutrophication II Relationships between fishand zooplankton in a set of experimental ponds, and con-clusions. Freshwat. Biol. 22: 89-107.
Irvine, K., B. Moss, H. Stansfield & D. Snook, 1990. Trophicrelations in Hickling Broad - a shallow and brackisheutrophic lake. Verh. int. Ver. theor. angew. Limnol. 24:576-579.
Jeppesen, E., P. Kristensen, J. P. Jensen, M. Sondergaard,E. Mortensen & T. Lauridsen, 1991. Recovery resiliencefollowing a reduction in external phosphorus loading ofshallow, eutrophic, danish lakes: duration, regulating fac-tors and methods for overcoming resilience. Mem. Ist. ital.Idrobiol. 48: 127-148.
Jeppesen, E., M. Sondergaard, E. Kanstrup, B. Petersen,R. B. Eriksen, M. Hammersh0j, E. Mortensen, J. P. Jensen& A. Have, 1994. Does the impact of nutrients on the bio-logical structure and function of brackish and freshwaterlakes differ? Hydrobiologia 275-276/Dev. Hydrobiol. 94:15-30.
Kilham, P. & S. S. Kilham, 1989. Endless summer: internalloading processes dominate nutrient cycling in tropicallakes. Freshwat. Biol 23: 379-389.
Kolding, J., 1992. A summary of Lake Turkana: an everchanging mixed environment. Mitt. int. Ver. Limnol. 23:25-35.
Lambert, J. M., J. N. Jennings, C. T. Smith, C. Green &J. N. Hutchinson, 1960. The making of the Broads: a re-construction of their origin in the light of new evidence.Royal Geographical Society, London.
Leah, R. T., B. Moss & D. E. Forrest, 1978. Experiments withlarge enclosures in a fertile, shallow, brackish lake, HicklingBroad, United Kingdom. Int. Revue ges. Hydrobiol. 63:291-310.
McWilliam, N. & V. Sekules (eds), 1986. Life and Landscape:P. H. Emerson. Art and Photography in East Anglia 1885-1890. Sainsbury Centre for the Visual Arts, University ofEast Anglia, Norwich, 200 pp.
Middleton, C. S., 1978. The Broadland Photographers. Wen-sum Publishing, Norwich, 189 pp.
Morgan, D. L., 1973. The Great Salt Lake. University ofMexico, Albuquerque, 432 pp.
Mort, M. A., 1991. Ecological genetics of freshwater zoo-plankton: current research and future perspectives. Arch.Hydrobiol. 123: 129-141.
Moss, B., 1977. Conservation problems in the Norfolk Broadsand rivers of East Anglia - phytoplankton, boats and thecauses of turbidity. Biol. Conserv. 12: 95-114.
Moss, B., 1978. The ecological history of a mediaeval man-made lake, Hickling Broad, Norfolk, United Kingdom. Hy-drobiologia 60: 23-32.
Moss, B., 1979. Algae and other fossil evidence for majorchanges in Strumpshaw broad, Norfolk, England in the lasttwo centuries. Br. phycol. J. 14: 263-283.
Moss, B., 1980. Further studies on the palaeolimnology andchanges in the phosphorus budget of Barton Broad, Nor-folk. Freshwat. Biol. 10: 261-279.
Moss, B., 1981. The composition and ecology of periphytoncommunities in freshwaters: 2. Inter-relationships betweenwater chemistry, phytoplankton populations and peri-

14
phyton populations in a shallow lake and associated ex-perimental reservoirs ('Lund Tubes'). Br phycol.J. 16: 59-76.
Moss, B., 1983. The Norfolk Broadland: Experiments in therestoration of a complex wetland. Biol. Rev. 58: 521-561.
Moss, B., 1988. The palaeolimnology of Hoveton GreatBroad, Norfolk: clues to the spoiling and restoration ofBroadland. Symposia of the Association for Environmen-tal Archaeology 7 (The Exploitation of Wetlands): 163-191.
Moss, B., 1989. Water pollution and the management of eco-systems: a case study of science and scientist. In Towarda More Exact Ecology. Thirtieth Symposium of the BritishEcological Society. Blackwell Scientific, Oxford: 401-422.
Moss, B., 1990. Engineering andBiological approaches to therestoration from eutrophication of shallow lakes in whichaquatic plant communities are important components. Hy-drobiologia 200-201/Dev. Hydrobiol. 61: 367-387.
Moss, B., 1991. The role of nutrients in determining the struc-ture of lake ecosystems and implications for the restoringof submerged plant communities to lakes which have lostthem. International Conference on N, P and Organic mat-ter. Contributions by invited international experts. NationalAgency for Environmental Protection, Copenhagen, Den-mark: 75-86.
Moss, B., M. Bales & K. Irvine, 1991. Food web and waterquality relationships in the saline broads of the RiverThurne, Norfolk. Mem. Ist. ital.Idrobiol 48: 342-362.
Moss, B. & H. R. Balls, 1989. Phytoplankton distribution ina floodplain lake and river system II Seasonal changes inthe phytoplankton communities and their control by hy-drology and nutrient availability. J. Plankton Res. 11: 839-867.
Moss, B., H. R. Balls, I. Booker, K. Manson & M. Timms,1988. Problems in the construction of a nutrient budget forthe River Bure and its broads (Norfolk) prior to its resto-ration from eutrophication. In Algae and the Aquatic en-vironment, F. E. Round (ed.), Biopress, Bristol: 326-353.
Moss, B., H. R. Balls, K. Irvine & J. H. Stansfield, 1986.Restoration of two lowland lakes by isolation from nutri-ent - rich water sources with and without removal of sedi-ment. J. appl. Ecol. 3: 391-414.
Moss, B. & I. Booker, H. R. Balls & K. Manson, 1989. Phy-toplankton distribution in a temperate floodplain lake andriver system I Hydrology, nutrient sources and phytoplank-ton biomass. J. Plankton Res. 11: 813-838.
Moss, B. & R. T. Leah, 1982. Changes in the ecosystem of aguanotrophic and brackish shallow lake in Eastern En-gland: potential problems in its restoration. Int. Revue ges.Hydrobiol. 67: 625-659.
Moss, B., J. H. Stansfield & K. Irvine, 1991. Development ofdaphnid communities in diatom- and cyanophyte-domi-nated lakes and their relevance to lake restoration bybiomanipulation. J. appl. Ecol. 2: 586-602.
Naumann, E., 1917. Undersokningar over fytoplankton ochunder den pelagiska regionen forsiggaende gyttje- och dy-bildningar inom vissa syd-och mellansvenska urbergsvat-ten. Kungl. Svenska Vetenskapsakademiens Handlingar56:1-165.
Osborne, P. L., 1980. Prediction of phosphorus and nitrogenconcentrations in lakes from both internal and externalloading rates. Hydrobiologia 69: 229-223.
Osborne, P. L., 1981. Phosphorus and nitrogen budgets ofBarton Broad and predicted effects of a reduction in nu-trient loading on phytoplankton biomass in Barton, Suttonand Stalham Broads, Norfolk, United Kingdom. Int. Revueges. Hydrobiol. 66: 171-202.
Pallis, M., 1911. Salinity in the Norfolk Broads I On the causeof salinity of the Broads of the River Thurne. Geogr. J. 37:214-291.
Phillips, G. L., D. Eminson & B. Moss, 1967. A mechanismto account for macrophyte decline in progressively eutrophi-cated freshwaters. Aquat. Bot. 4: 103-126.
Remane, A. & C. Schlieper, 1971. The biology of brackishwater. Die Binnengewasser 2nd (English) Edn. 25: 1-37.
Simons, J., M. Ohm, R. Daalder, P. Boers & W. Rip, 1994.Restoration of Botshol (The Netherlands) by reduction ofexternal nutrient load: recovery of a characean communityby Chara connivens. Hydrobiologia 275-276/Dev. Hydro-biol. 94: 243-253.
Stansfield, J. H., B. Moss & K. Irvine, 1989. The loss of sub-merged plants with eutrophication III Potential role of or-ganochlorine pesticides: a palaeoecological study. Fresh-wat. Biol. 2: 109-132.
Thienemann, A., 1915. Physikalische und chemische Unter-suchungen in den Maaren der Eifel II Verh. Naturhist. Ver.preuB. Rheinlande u. Westfalens 71: 273-389.
Thienemann, A., 1931. Tropische Seen und Seetypen. Arch.Hydrobiol. Suppl. 9: 205-231.
Timms, R. M. & B. Moss, 1984. Prevention of growth ofpotentially dense phytoplankton populations by zooplank-ton grazing in the presence of zooplanktivorous fish, in ashallow wetland ecosystem. Limnol. Oceanogr. 29: 472-486.
Stokes, W. L., 1984. The Great Salt lake. Starstore Publish-ing Co. Salt Lake City, 30 pp.
Weider, L. J. & P. D. N. Hebert, 1987. Ecological and physi-ological differentiation among low-Arctic clones of Daphniapulex. Ecology 68: 188-198.
Williams, W. D., 1986. Limnology, the study of inland waters:a comment on perceptions of studies of salt lakes. past andpresent. In De Deckker, P. & W. D. Williams (eds), Lim-nology in Australia. Dr W. Junk Publishers, Dordrecht:471-486.
Wortley, J. S., 1974. The river systems of Norfolk. In Naturein Norfolk. Ed by Norfolk Naturalists Trust, Jarrold. Nor-folk: 113-122.





























