Embed Size (px)
Citation preview

THY JOUBNAL OF BIOLOGICAL CHEMISTBY
Vol. 260, No. 7, Issue of April 10, pp. 2487-2494,197s Printed in U.S.A.
Bovine Serum Albumin
STUDY OF THE FATTY ACID AND STEROID BINDING SITES USING SPIN-LABELED LIPIDS*
(Received for publication, August 22, 1974)
JOEL D. MORRISETT,~ HENRY J. POWNALL, AND ANTONIO M. GOTTO, JR.
From the Division of Atherosclerosis and Lipoprotein Research, Department of Medicine, The Methodist Hospital. Baylor College of Medicine, Houston, Texas 770.26
SUMMARY
Three spin-labeled derivatives of stearic acid and two derivatives of palmitic acid have been used to study the structure of the strong fatty acid binding site of bovine se- rum albumin. The steroid and indole binding sites have been studied using spin-labeled derivatives of androstol and in- dole, respectively. Paramagnetic resonance and fluorescence quenching data suggest that the fatty acid, steroid, and in- dole binding sites may be identical.
The mobility of the nitroxyl group at C-8 of pahnitic acid bound to albumin at a 1: 1 molar ratio is unaffected when the carboxyl group is esterified. When the nitroxyl group is lo- cated at C-5 on this acid, its motion is detectably increased by esterification of the carboxyl group but the magnitude of this change is small. This result suggests that the carboxyl group may play a minor role in the binding of fatty acids to the strongest fatty acid binding site of albumin.
When stearic acid derivatives bearing the nitroxide at C-5, C-12, and C-16 are bound to albumin at a ligand to albumin ratio of 1, the order of mobility at O-30” is C-16 > C-12 N_ C-5. Although motion at the methyl terminus is al- ways greater than at the COOH terminus in the range O-60”, a simple monotonic increase in chain motion between the two termini is not observed. Arrhenius plots of the motion parameters for these bound fatty acids show two abrupt changes in slope. The temperature ranges for these changes are 15-23” and 3845”. These results suggest that when one mole of spin-labeled fatty acid is bound to albumin, the pro- tein undergoes a conformational change in each of these temperature ranges.
Albumin is the most abundant protein in mammalian plasma. This molecule has a molecular weight of about 67,000 (1, 2), and exists as a single polypeptide chain (2). Of the approximately 588 amino acids present in bovine albumin, there are only 2 tryptophanyl, 1 cysteinyl, and 4 methionyl residues (1, 2). It
* This research was supported in part by grants from the De- nartment of Health. Education. and Welfare CHL-14194 and HL- ‘i5648) and from the’ John A. Hartford Foundation, Inc.
$ Established Investigator of the American Heart Association.
has been estimated that the protein possesses about 50% (Y- helical structure which is stabilized by 17 to 18 disulfide bridges (3).
The affinity of the protein for ionic ligands bearing alkyl side chains has been shown to depend 011 the hydrophobic character of the chain and the sign of the charged group. ;\lolecules with long chains and negative ionic groups such as stearic acid are very tightly bound, whereas molecules containing shorter side chains or positively charged ionic groups such as hexyltrimethyl- ammonium bromide are less tightly bound (4). Many of the anionic ligands have at least two classes of binding sites on albumin which differ by 1 to 2 orders of magnitude in relative binding affinity. Recent studies suggest that arginine and tryptophan are at or near these sites (5). Studies directed to- ward the identification of those amino acid residues in albumin which are involved in fatty acid binding are currently underway (6).
Albumin also binds a variety of other ligands, such as acetyl- choline, ascorbic acid, barbiturates, digitonin, penicillin, sulfona- mides, and thyroxine (3) and is the principal vehicle for the transport of several lipid classes, including free fatty acids (7), lysolecithin (8), and certain steroids (9). Because of our interest in the mechanisms of lipid binding and transport (lo-12), we have undertaken a study of the interaction between albumin and a variety of spin-labeled lipids. In this report, we describe the interaction of bovine albumin’ with nitroxide-containing derivatives of fatty acids, fatty acid methyl esters, androstol, and indole.
EXPERIMENTAL PROCEDURES
Spin Labels-Spin-labeled fatty acids (12,3-, 10,3-, 7,6-, 5,10-, and 1,14-SLFA* in Fig. 1) were obtained by hydrolysis of the corresponding ester in dioxane-aqueous sodium hydroxide (15) or from Syva (Palo Alto). Spin-labeled derivatives of fatty acid methyl esters (II) were prepared by m-chloroperbenzoic acid oxidation of the corresponding oxazolidine (16). Starting keto- esters were synthesized by standard methods (15). Spin-labeled
1 Albumin has also been studied by the use of covalentlg at- tached spin labels (13, 14).
2 The abbreviations used are: SLFA. snin-labeled fattv acid: SLFAME, spin-labeled fatty acid methyl ester; SLA, spin-labeled androstol; SLC, spin-labeled carboline; doxyl, 2,2-dimethyl- oxazolidinyl-1-oxyl; HDL, high density lipoprotein; apoLP-Gln-I, apolipoprotein-glutamine I is the most abundant apoprotein in human HDL; apoLP-Gln-II, apolipoprotein-glutamine II is the second most abundant apoprotein in human HDL.
2487
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from
2488
0.~0 I CH; ICH& c ICH21”.C02H m, n SLiA
FIG. 1. Chemical structures of spin-labeled lipid molecules used in this study. I, spin-labeled fatty acid; II, spin-labeled fatty acid methyl ester; III, spin-labeled androstol; IV, spin- labeled carboline.
androstol (SLA, III) was prepared as described by Hubbell and McConnell (18). Spin-labeled carboline (SLC, IV) was prepared by coupling phenylhydrazine with l-oxyl-2,2,6,6-tetramethyl- piperidone-4 followed by cyclization (19) then oxidation (20).
VOLUME (ml1
Instrumentation-Electron paramagnetic resonance (EPR) spectra were recorded on a Varian E-12 spectrometer equipped with a variable temperature controller. Acetone and Dry Ice were used to cool the nitrogen. The temperature of the cavity was measured before and after recording the spectrum with a Tri-It electronic thermometer to an accuracy of f0.5’. Samples were contained in flat quartz microcells (Varian). The splitting between the two extrema of the EPR spectra could be measured with an accuracy of about ~0.3 G. pH was measured with a Radiometer TTT2 pH meter equipped with a combination micro- electrode, type CK2321C. Protein absorption at 278 nm was measured with a Beckman Acta V spectrophotometer. Fluores- ence spectra were recorded at 22” with an Aminco-Bowman spectrofluorimeter equipped with an RCA lP28 phototube. Excita- tion wavelength was 280 nm and slit widths for the excitation and analyzing monochrometers were 2.0 mm.
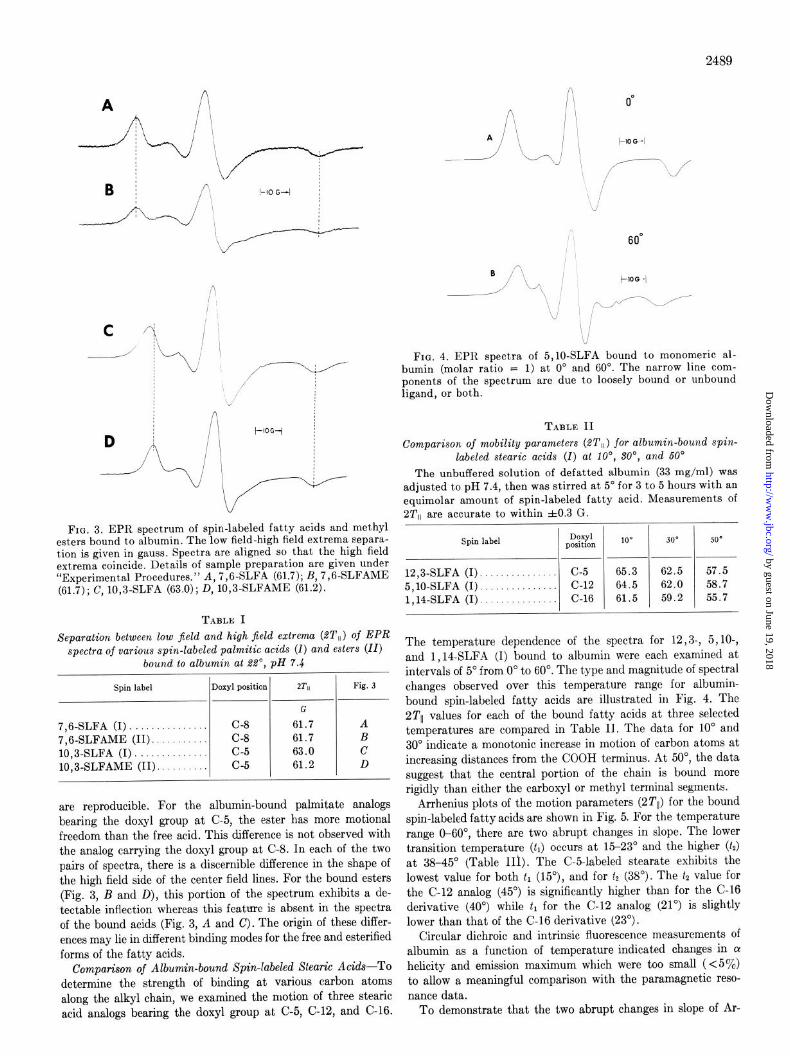
FIG. 2. Gel filtration of charcoal-treated albumin. Chroma- tography conditions: column (2.5 X 100 cm) of Sephadex G-150 eluted with 0.2 M NaCl-0.05 M Tris, pH 8.1, 5”. Fractions of 9 ml were collected at a flow rate of 14 ml/hour. The mixture was composed of 4.7% polymer, 19.4% dimer, and 75.8% monomer. The EPR spectrum of 5,10-SLFA when bound to monomeric albumin was not detectably different from that of the label bound to the unfractionated mixture.
Analysis of EPR Spectra-For spectra which indicated strong impedence of spin label motion, the splitting between the low and high field extrema (2T,,) was used as an index fbr immobiliaa- tion. For an isotropic ensemble of nitroxides whose correlation times (TV) fall within a certain range, 2T,, increases* as the mo- tional freedom of the nitroxide decreases. In the range of motion where 2T,, is a measurable quantity, this parameter is useful for comparing various degrees of immobilization. Since 2Tl values were not always quantifiable, calculation of order parameters (S) was not feasible (15).
The column was developed at 14 ml/hour with the same buffer and g-ml fractions were collected. The protein content of each fraction was determined by its absorbance (Ef% = 6.67 at 278 nm) (23). Under these conditions, defatted albumin contained about 76yo monomers, 19% dimers, and 57’ polymers (Fig. 2). When monomeric albumin from this experiment was adjusted to pH 7.4 and labeled with 5,10-SLFA as described above, the reso- nance spectrum was identical to that obtained with the unre- solved mixture prior to chromatography. Chromatography of labeled monomeric albumin gave a single peak coincident with unlabeled albumin monomer.
Apolipoproteins-High density lipoproteins were isolated by ultracentrifugation of human plasma as described previously (24). then were delipidated by extraction with chloroform-meth- andi (3:l). The apohrotein components were solubilized in 0.2 M Tris-0.01% EDTA. DH 8.5. AnoLP-Gln-I was prepared by a --,., previously described-method (i4). Each of these proteins was dialyzed against distilled water and adjusted to pH 7.4 before
Albumin-Crystalline bovine serum albumin (fatty acid-poor) was obtained from Miles Laboratories (Lot 21) and used as re- ceived, except where indicated otherwise. In a typical experi- ment, 100 mg of albumin (-1.5 pmol) were dissolved in 3 ml of distilled, deionized water and the pH was adjusted to 7.4. Ap- proximately 1.5 pm01 of spin label in ethanol were transferred to a test tube and evaporated to dryness with a stream of nitrogen. The solution of albumin was then added and stirred gently for 3 to 6 hours at 5“.
labeling bythe method described above for albumin. Fluorescence Quenching-In a typical quenching experiment,
the fluorescence spectrum of a 2-ml solution of albumin (1.57 mg/ml, pH = 7.4j contained in a quartz fluorescence cell (1 X 1 X 3 cm) was recorded in the absence of quencher. Then 25+1 al&ots df the quenching ligand (0.5 to 0.8 mg/ml) in ethanol were added with a Hamilton syringe to the albumin solution and the fluorescence spectrum recorded after each addition. The intensities at the eI$ission maxima were used in the Stern-Volmer treatment of the quenching data (Table IV, Figs. 7 and 10).
For some experiments, defatted (21) monomeric (22) albumin was used. It was prepared in the following manner. One gram of albumin was dissolved in 10 ml of water and to this was added 0.5 g of powdered charcoal (Norit A, MCB). The mixture was adjusted to pH 3.0 with 1 M HCl and was stirred for 2 hours at room temnerature. The charcoal was removed by centrifugation (18,000 rpm, 5”, 20 min). The supernatant was adiusted to pH 8.15 with 1.0 M NaQH. A 2-ml aliquot (200 mg) of this defatted albumin solution was applied to a column (2.5 X 100 cm) of Sephadex G-150 equilibrated with 0.2 M NaCl-0.05 M Tris, pH 8.0, at 5”.
RESULTS
Comparison of Albumin-bound Spin-labeled Palmitic Acids and Methyl Esters-The resonance spectra of albumin-bound 7,6- and 10,3-spin-labeled fatty acids (A, c) and their corres- ponding esters (B, D) are shown in Fig. 3. All four spectra indi- cate strong association of the nitroxide-bearing alkyl chain with albumin. While alI these spectra appear similar, there are subtle, but significant spectral differences which distinguish any single spectrum from the others; viz. the total separation be- tween the low and high field extrema (2Tll) and the shape of the high field side of the center field line (MO). Values of 2Tll for albumin-bound spin-labeled fatty acids and esters are collected in Table I. While the differences in values of 2Tll are small, they
* For nitroxides which are part of an ordered ensemble of molecules such as a crystal or a phospholipid multibilayer, 2T,, also increases as the ensemble is experimentally disposed in space so as to increase the parallelism between the long axis of the 2p ?r orbital of the nitroxide and the direction of the applied laboratory magnetic field (Ho).
I.2 - MONOMER ( 75 8%)
1.0 - 0 0
os-
A280 -
06- I * /
0.4
.~jLzJf :.._
90 IS0 270 360 450
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from
2489
C
D
i
;\\:..’
FIG. 3. EPR spectrum of spin-labeled fatty acids and methyl esters bound to albumin. The low field-high field extrema sepsra- tion is given in gauss. Spectra are aligned so that the high field extrema coincide. Details of sample preparation are given under “Experimental Procedures.” A, 7,6-SLFA (61.7); B, 7,6-SLFAME (61.7); C, 10,3-SLFA (63.0); D, 10,3-SLFAME (61.2).
TABLE I Separation between low jield and high field extreme (ST,,) of EPR
spectra of various spin-labeled palmitic acids (I) and esters (II) bound to albumin at RI’, pH Y.4
Spin label Doxyl position ZTll Fig. 3
G
7,6-SLFA (I). C-8 61.7 A 7,6-SLFAME (II). C-8 61.7 B 10,3-SLFA (I) c-5 63.0 c 10,3-SLFAME (II). c-5 61.2 D
are reproducible. For the albumin-bound palmitate analogs bearing the doxyl group at C-5, the ester has more motional freedom than the free acid. This difference is not observed with the analog carrying the doxyl group at C-8. In each of the two pairs of spectra, there is a discernible difference in the shape of the high field side of the center field lines. For the bound esters (Fig. 3, B and D), this portion of the spectrum exhibits a de- tectable inflection whereas this feature is absent in the spectra of the bound acids (Fig. 3, A and c). The origin of these differ- ences may lie in different binding modes for the free and esterilied forms of the fatty acids.
Comparison of Albumin-bound Spin-labeled Stearic Acids-To determine the strength of binding at various carbon atoms along the alkyl chain, we examined the motion of three stearic acid analogs bearing the doxyl group at C-5, C-12, and C-16.
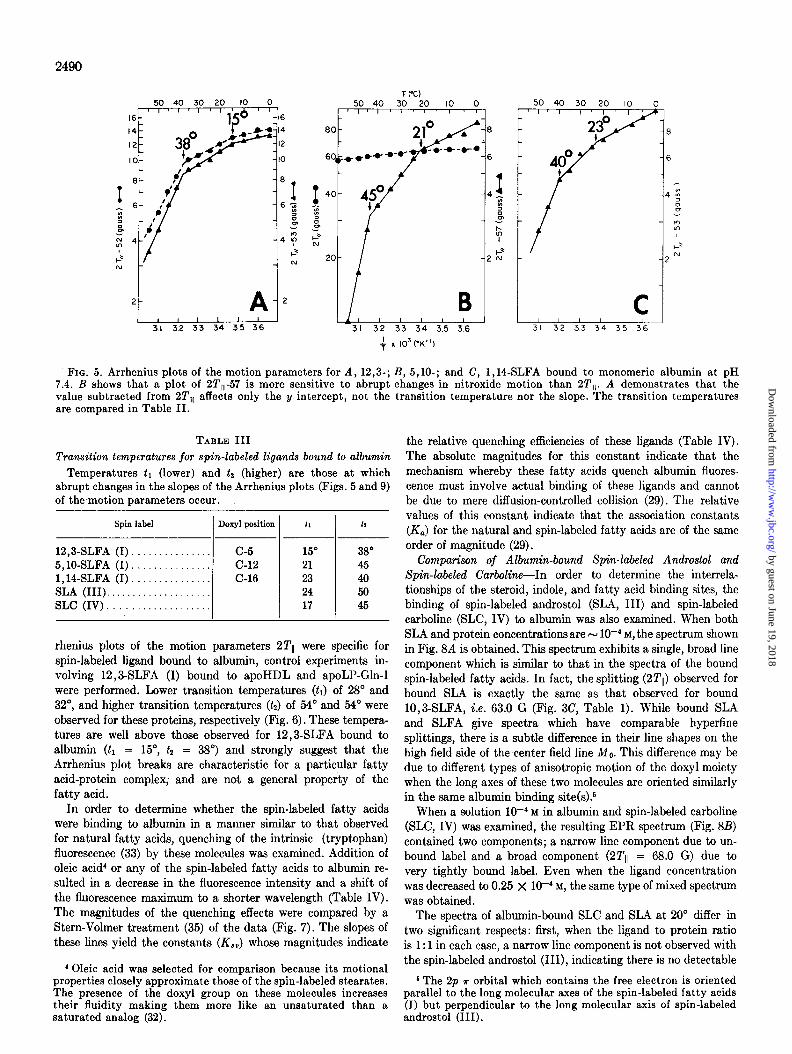
FIG. 4. EPR spectra of 5,10-SLFA bound to monomeric al- bumin (molar ratio F 1) at 0” and 60”. The narrow line com- ponents of the spectrum are due to loosely bound or unbound ligand, or both.
TABLE II Comparison of mobility parameters (ST,,) for albumin-bound spin-
labeled steak acids (I) at lo”, ?30”, and 60’ The unbuffered solution of defatted albumin (33 mg/ml) was
adjusted to pH 7.4, then was stirred at 5” for 3 to 5 hours with an equimolar amount of spin-labeled fatty acid. Measurements of 2T,, are accurate to within f0.3 G.
Spin label Doxyl position 100 300 SO0
p--P 12,3-SLFA (I). C-5 65.3 62.5 57.5 5,10-SLFA (I). C-12 64.5 62.0 58.7 1,14-SLFA (I). . C-16 61.5 59.2 55.7
The temperature dependence of the spectra for 12,3-, 5, lo-, and 1,14-SLFA (I) bound to albumin were each examined at intervals of 5” from 0” to 60”. The type and magnitude of spectral changes observed over this temperature range for albumin- bound spin-labeled fatty acids are illustrated in Fig. 4. The 2Tll values for each of the bound fatty acids at three selected temperatures are compared in Table II. The data for 10” and 30” indicate a monotonic increase in motion of carbon atoms at increasing distances from the COOH terminus. At 50”, the data suggest that the central portion of the chain is bound more rigidly than either the carboxyl or methyl terminal segments.
Arrhenius plots of the motion parameters (2Tll) for the bound spin-labeled fatty acids are shown in Fig. 5. For the temperature range O-60”, there are two abrupt changes in slope. The lower transition temperature (tl) occurs at 15-23” and the higher (tz) at 38-45“ (Table III). The C-5-labeled stearate exhibits the lowest value for both tl (15”), and for t2 (38”). The tz value for the C-12 analog (45”) is significantly higher than for the C-16 derivative (40”) while tl for the C-12 analog (21”) is slightly lower than that of the C-16 derivative (23”).
Circular dichroic and intrinsic fluorescence measurements of albumin as a function of temperature indicated changes in LY helicity and emission maximum which were too small ( <5%) to allow a meaningful comparison with the paramagnetic reso- nance data.
To demonstrate that the two abrupt changes in slope of Ar-
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from
2490
50 40 30 20 IO c 1)1111l~ I
16-
*I[, ( , , ,+ 3.1 32 33 34 35 36
) -16
I-1’ -1;
-I(
-E
-6
-‘
T 1°C) 50 40 30 20 IO 0
lIJ!I 71 ‘1’ 1 ’ I-
B I I I I I I 31 32 33 34 3.5 3.6
f x IO3 (“K-‘I
-6
-4
-2
C I I I I I 31 32 33 34 35 36
FIG. 5. Arrhenius plots of the motion parameters for A, 12,3-; B, 5,10-; and C, 1,14-SLFA bound to monomeric albumin at pH 7.4. B shows that a plot of 2T,,-57 is more sensitive to abrupt changes in nitroxide motion than 2T,,. A demonstrates that the value subtracted from 2T,, affects only the y intercept, not the transition temperature nor the slope. The transition temperatures are compared in Table II.
TABLE III
Transition temperatures for spin-labeled ligands bound to albumin Temperatures tl (lower) and ta (higher) are those at which
abrupt changes in the slopes of the Arrhenius plots (Figs. 5 and 9) of the motion parameters occur.
Spin label Doxyl position 11 la
12,3-SLFA (I) c-5 15” 38” 5,lOSLFA (I). . c-12 21 45 1,14-SLFA (I). . . C-16 23 40 SLA (III). . 24 50 SLC (IV). . . . 17 45
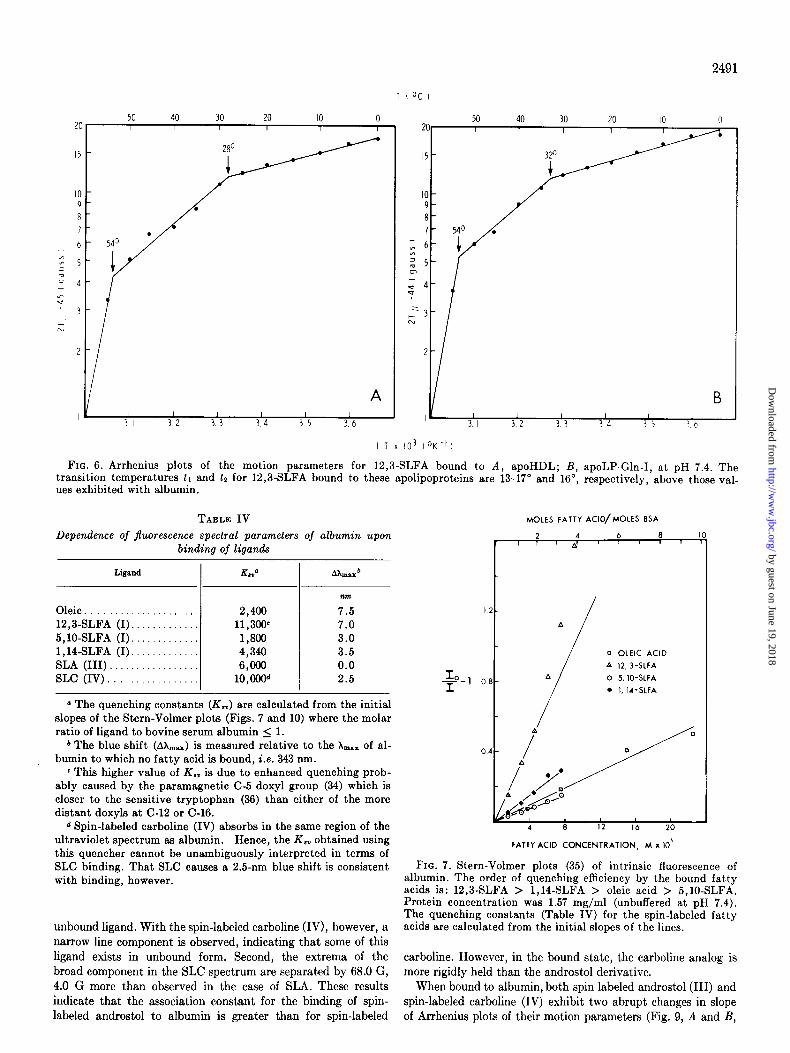
rhenius plots of the motion parameters 2Tll were specific for spin-labeled ligand bound to albumin, control experiments in- volving 12,8SLFA (I) bound to apoHDL and apoLl’-Gln-I were performed. Lower transition temperatures (tr) of 28” and 32”, and higher transition temperatures (tz) of 54” and 54’ were observed for these proteins, respectively (Fig. 6). These tempera- tures are well above those observed for 12,8SLFA bound to albumin (tr = 15”, t2 = 38”) and strongly suggest that the Arrhenius plot breaks are characteristic for a particular fatty acid-protein complex, and are not a general property of the fatty acid.
In order to determine whether the spin-labeled fatty acids were binding to albumin in a manner similar to that observed for natural fatty acids, quenching of the intrinsic (tryptophan) fluorescence (33) by these molecules was examined. Addition of oleic acid* or any of the spin-labeled fatty acids to albumin re- sulted in a decrease in the fluorescence intensity and a shift of the flnorescence maximum to a shorter wavelength (Table IV). The magnitudes of the quenching effects were compared by a Stern-Volmer treatment (35) of the data (Fig. 7). The slopes of these lines yield the constants (Z&J whose magnitudes indicate
* Oleic acid was selected for comparison because its motional properties closely approximate those of the spin-labeled stearates. The presence of the doxyl group on these molecules increases their fluidity making them more like an unsaturated than a saturated analog (32).
the relative quenching efficiencies of these ligands (Table IV). The absolute magnitudes for this constant indicate that the mechanism whereby these fatty acids quench albumin fluores- cence must involve actual binding of these ligands and cannot be due to mere diffusion-controlled collision (29). The relative values of this constant indicate that the association constants (K,) for the natural and spin-labeled fatty acids are of the same order of magnitude (29).
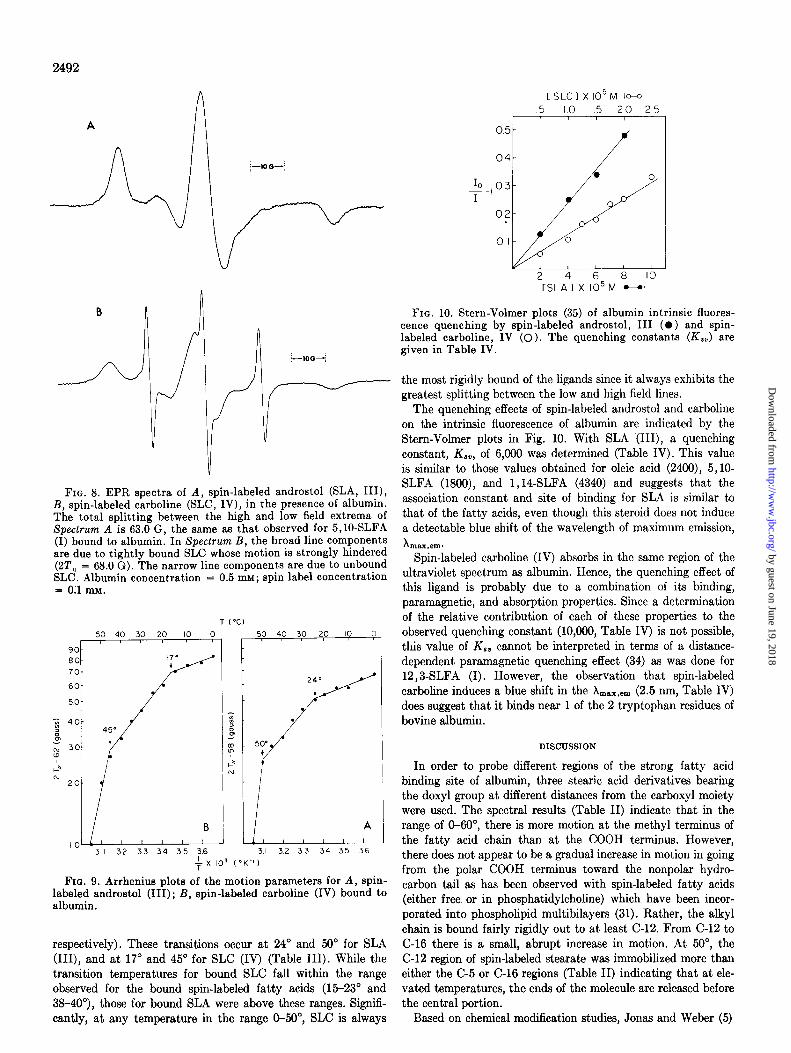
Comparison of Albumin-bound Spin-labeled Androstol and Spin-labeled Carboline-In order to determine the interrela- tionships of the steroid, indole, and fatty acid binding sites, the binding of spin-labeled androstol (SLA, III) and spin-labeled carboline (SLC, IV) to albumin was also examined. When both SLA and protein concentrations are N lo-* M, the spectrum shown in Fig. 8A is obtained. This spectrum exhibits a single, broad line component which is similar to that in the spectra of the bound spin-labeled fatty acids. In fact, the splitting (2Tll) observed for bound SLA is exactly the same as that observed for bound lO,BSLFA, i.e. 63.0 G (Fig. 3C, Table 1). While bound SLA and SLFA give spectra which have comparable hyperfine splittings, there is a subtle difference in their line shapes on the high field side of the center field line Me. This difference may be due to different types of anisotropic motion of the doxyl moiety when the long axes of these two molecules are oriented similarly in the same albumin binding site(s).”
When a solution lo-* M in albumin and spin-labeled carboline (SLC, IV) was examined, the resulting ERR spectrum (Fig. 8B) contained two components; a narrow line component due to un- bound label and a broad component (22’11 = 68.0 G) due to very tightly bound label. Even when the ligand concentration was decreased to 0.25 x lo-* M, the same type of mixed spectrum was obtained.
The spectra of albumin-bound SLC and SL4 at 20” differ in two significant respects: first, when the ligand to protein ratio is 1: 1 in each case, a narrow line component is not observed with the spin-labeled androstol (III), indicating there is no detectable
6 The 2p 7 orbital which contains the free electron is oriented parallel to the long molecular axes of the spin-labeled fatty acids (I) but perpendicular to the long molecular axis of spin-labeled androstol (III).
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from
T C"CI
2C
IO 9 8 7
6 z * 5 ; = 4
0 3
r;
2
I
50 40 30 20 IO 0
1 I 1 I I 1 3 I 3. 2 3. 3 3. 4 3. 5 3. 6
I3
I T x IO3 1°K.‘I
FIG. 6. Arrhenius plots of the motion parameters for 12,3-SLFA bound to A, apoHDL; B, apoLP-Gln-I, at pH 7.4. The transition temperatures ti and 12 for 12,3-SLFA bound to these apolipoproteins are 13-17” and 16”, respectively, above those val- ues exhibited with albumin.
TABLE IV MOLES FATTY ACID/MOLES 8SA
Dependence of jluorescence spectral parameters of albumin upon binding of ligands
2 4 6 8
I ”
’ a’ ’ ’ r ’ ’
Liiand
Oleic 12,3-SLFA (I) 5,10-SLFA (I). 1,14-SLFA (I). SLA (III). SLC (IV)
L”
2,400 11,300c 1,806 4,340 ‘3,‘3oC’
10,OoOd
nm
7.5 7.0 3.0 3.5 0.0 2.5
o The quenching constants (K,,) are calculated from the initial slopes of the Stern-Volmer plots (Figs. 7 and 10) where the molar ratio of ligand to bovine serum albumin 2 1.
b The blue shift (A&,.,) is measured relative to the A,., of al- bumin to which no fatty acid is bound, i.e. 343 nm.
E This higher value of K., is due to enhanced quenching prob- ably caused by the paramagnetic C-5 doxyl group (34) which is closer to the sensitive tryptophan (36) than either of the more distant doxyls at C-12 or C-16.
d Spin-labeled carboline (IV) absorbs in the same region of the ultraviolet spectrum as albumin. Hence, the K,, obtained using this quencher cannot be unambiguously interpreted in terms of SLC binding. That SLC causes a 2.5-nm blue shift is consistent with binding, however.
unbound ligand. With the spin-labeled carboline (IV), however, a narrow line component is observed, indicating that some of this ligand exists in unbound form. Second, the extrema of the broad component in the SLC spectrum are separated by 68.0 G, 4.0 G more than observed in the case of SLA. These results indicate that the association constant for the binding of spin- labeled androstol to albumin is greater than for spin-labeled
0 OLEIC ACID A 12, 3-SLFA 0 5, IO-SLFA
l 1, II-SPA
10
3
-
FATTY ACID CONCENTRATION, M x 10’
FIG. 7. Stern-Volmer plots (35) of intrinsic fluorescence of albumin. The order of quenching efficiency by the bound fatty acids is: 12,3-SLFA > 1,14-SLFA > oleic acid > 5,10-SLFA. Protein concentration was 1.57 mg/ml (unbuffered at pH 7.4). The quenching constants (Table IV) for the spin-labeled fatty acids are calculated from the initial slopes of the lines.
carboline. However, in the bound state, the carboline analog is more rigidly held than the androstol derivative.
When bound to albumin, both spin-labeled androstol (III) and spin-labeled carboline (IV) exhibit two abrupt changes in slope of Arrhenius plots of their motion parameters (Fig. 9, A and B,
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from
2492
A A
j--100- J--&f-v 1
B 1
i-100+
I
Y
FIG. 8. EPR spectra of A, spin-labeled androstol @LA, III), B, spin-labeled carboline (SLC, IV), in the presence of albumin. The total splitting between the high and low field extrema of Speclrum A is 63.0 G, the same as that observed for 5,10-SLFA (I) bound to albumin. In Spectrum B, the broad line components are due to tightly bound SLC whose motion is strongly hindered (2T,, = 68.0 G). The narrow line components are due to unbound SLC. Albumin concentration = 0.5 mM; spin label concentration = Oil mM.
T (‘C)
90. SO- 70.
60.
IO 3 I 32 33 34 35 36
i L 31 32 33 34 35 36
+X IO’ (OK-‘)
Fxa. 9. Arrhenius plots of the motion parameters for A, spin- labeled androstol (III); B, spin-labeled carboline (IV) bound to albumin.
respectively). These transitions occur at 24” and 50” for SLA (III), and at 17” and 45” for SLC (IV) (Table III). While the transition temperatures for bound SLC fall within the range observed for the bound spin-labeled fatty acids (15-23’ and 38409, those for bound SLA were above these ranges. Signifi- cantly, at any temperature in the range O-50”, SLC is always
C SLC 1 X IO5 M 1~
5 IO 15 20 25 I I
IO I ’
FIG. 10. Stern-Volmer plots (35) of albumin intrinsic fluores- cence quenching by spin-labeled androstol, III (0) and spin- labeled carboline, IV (0). The quenching constants (K.,) are given in Table IV.
the most rigidly bound of the ligands since it always exhibits the greatest splitting between the low and high field lines.
The quenching effects of spin-labeled androstol and carboline on the intrinsic fluorescence of albumin are indicated by the Stern-Volmer plots in Fig. 10. With SLA (III), a quenching constant, K,,, of 6,000 was determined (Table IV). This value is similar to those values obtained for oleic acid (2400), 5, lo- SLFA (1800), and 1,14-SLFA (4340) and suggests that the association constant and site of binding for SLA is similar to that of the fatty acids, even though this steroid does not induce a detectable blue shift of the wavelength of maximum emission, x rnax.l?rn~
Spin-labeled carboline (IV) absorbs in the same region of the ultraviolet spectrum as albumin. Hence, the quenching effect of this ligand is probably due to a combination of its binding, paramagnetic, and absorption properties. Since a determination of the relative contribution of each of these properties to the observed quenching constant (10,000, Table 1V) is not possible, this value of K,, cannot be interpreted in terms of a distance- dependent paramagnetic quenching effect (34) as was done for 12,3-SLFA (I). However, the observation that spin-labeled carboline induces a blue shift in the X,,,,,, (2.5 nm, Table IV) does suggest that it binds near 1 of the 2 tryptophan residues of bovine albumin.
DISCUSSION
In order to probe different regions of the strong fatty acid binding site of albumin, three stearic acid derivatives bearing the doxyl group at different distances from the carboxyl moiety were used. The spectral results (Table II) indicate that in the range of O-60”, there is more motion at the methyl terminus of the fatty acid chain than at the COOH terminus. However, there does not appear to be a gradual increase in motion in going from the polar COOH terminus toward the nonpolar hydro- carbon tail as has been observed with spin-labeled fatty acids (either free or in phosphatidylcholine) which have been incor- porated into phospholipid multibilayers (31). Rather, the alkyl chain is bound fairly rigidly out to at least C-12. From C-12 to C-16 there is a small, abrupt increase in motion. At 50”, the C-12 region of spin-labeled stearate was immobilized more than either the C-5 or C-16 regions (Table II) indicating that at ele- vated temperatures, the ends of the molecule are released before the central portion.
Based on chemical modification studies, Jonas and Weber (5)
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from

2493
have suggested that there are arginine residues at or near the strong hydrophobic anion binding sites of bovine albumin. The implication of their study is that the cationic guanidino groups of these arginine residues form electrostatic or hydrogen bonds with the carboxyl groups of fatty acids. Our results indicate this or a similar interaction is operative but is not the dominant mechanism whereby albumin binds fatty acids. Rather, our re- sults suggest that the entire length of the fatty acid chain is bound by hydrophobic interactions which are somewhat variable along the length of the binding site. This view is supported by the spectral data obtained from the uncharged spin-labeled fatty acid esters (Table 1, Fig. 3). These data show that esterifi- cation of the carboxyl group causes no change in the mobility of the C-8 doxyl function, and only a small (albeit reproducible) increase in the mobility of the C-5 doxyl group (1.8 G). That the mobility increase for the doxyl group is greatest for the fatty acyl chain which bears this functionality nearest the carboxyl group is consistent with the view that an arginine side chain interacts with this carboxyl group.
Further information about the preferential interaction of specific regions of fatty acids with albumin was obtained from a study of the temperature dependence of the resonance spectra of the bound doxyl stearates. Results from these studies (Fig. 5, Tables II, III) indicate that the doxyl groups of the bound spin-labeled fatty acids undergo two abrupt changes in motion, the lower of these occurring between 15” and 23” and the higher between 38” and 45’. The temperature required to bring about the lower transition (ti) is lowest (15”) for the fatty acid with the doxyl group closest to the COOH terminus (12 ,&SLFA). It is the same fatty acid which exhibits the lowest temperature (38’) for the higher transition (tz). This does not mean that the carboxyl end of the fatty acid is bound more loosely than seg- ments of the molecule closer to the methyl terminus. On the contrary, the experiments involving doxyl palmitic acids (Table I) demonstrate that the carboxyl end is the most rigidly bound portion of the molecule. Furthermore, the mobility parameter of albumin-bound doxyl stearates (Table II) is greatest (i.e. the motion is slowest) for the derivative carrying the nitroxide nearest the carboxyl group, up to a temperature of 50”. What this result does mean is that temperature-dependent changes in mobility of the bound fatty acid are first observed in the car- boxy1 portion of the molecule. The transitions ti and t2 may represent small conformational changes in the protein at or near the fatty acid binding site. Such changes could result in decreased interaction between the fatty acid and the protein, thereby al- lowing new degrees of motional freedom (e.g. trans-gauche isomerization) for the carbon atoms at or near the doxyl moieties. That these transitions are observed first with 12,3-SLFA sug- gests that these conformational changes in the strong fatty acid binding site begin in the local region of this site which interacts with the carboxyl segment of the molecule. Although the spectra of the bound fatty acids do change with temperature, the net change between 0” and 60” is not particularly great, probably due to stabilization of the tertiary structure of the protein by its 17 to 18 disulfide bonds or by the bound fatty acid itself, or both. That these transitions were not observed by circular dichroism or tryptophan fluorescence suggests the local nature of these structural changes.
steroids similar to SLA have been shown previously to bind to bovine albumin with an affinity somewhat lower than that of long chain fatty acids (36). These relative affinities are not surprising when the surface contact areas of these molecules are compared. However, our experiments indicate that once SLA is bound, it is as rigidly held (as strongly immobilized) as the spin- labeled fatty acids. A similar result has been obtained by Cham- baa et al. (26) with a different nitroxyl steroid. The quenching constant, K,,, for SLA (III) (Table IV) is comparable to that of oleic acid and the spin-labeled fatty acids (except 12,3SLFA which exhibits distance dependent paramagnetic quenching in addition to the normal quenching). This result suggests that SLA is (a) binding at the strong fatty acid binding site, or (b) binding at a different site but in a manner which produces the perturbation of tryptophan emission. A determination of whether this perturbation is due to ligand binding near the affected tryptophan so as to change directly its environment, or is the result of a conformational change induced indirectly by ligand binding at some site more remote from this tryptophan, will require further experiments. The first alternative is very attractive in light of the recent isolation of an albumin tryptic peptide with the sequence Lys-Trp-Ala-Val-Ala-Arg (37). Efforts to locate the steroid binding site(s), have recently re- sulted in the isolation from a peptic digest of albumin, a tyrosine- rich peptide of molecular weight 10,050 which retains the ca- pacity to bind progesterone, testosterone, and 17P-estradiol (27).
The carboline derivative (SLC, IV), exhibits several properties which make it an attractive probe molecule. First, the nitroxide ring is coplanar with the adjacent ring system so that the com- plete molecule is flat, closely resembling the structure of indole. Fatty acids bearing the doxyl function do not so closely mimic the parent molecules since the plane of the nitroxide ring lies perpendicular rather than parallel to the long axis of t’he mole- cule. Second, in the unoxidized form, the molecule is fluorescent. In favorable cases where the system of interest does not contain a fluorophore which absorbs (emits) in the region of 280 (360) nm (e.g. tryptophan), the paramagnetic and diamagnetic forms of this molecule may be used to probe the same binding site by both paramagnetic resonance and fluorescence methods.
In the presence of albumin, SLC (IV) gives a mixed spectrum containing a broad component with a splitting of 68.0 G, indi- cating that the bound label has almost no motion independent of the albumin molecule. Kuznetsov et al. (17) have obtained similar results using a similar molecule which also contains the indole nucleus. It is likely that SLC binds at the single site where indole, skatole, acetyl-n-tryptophan, tryptophan, and indole- propionate bind (38). This site is thought to be flexible and adaptable since several different small affinity labels are bound. Our results suggest that once this label is bound, it is rigidly held in the binding site. Due to absorption of SLC in the same region as albumin, the observed quenching constant is of no value in estimating the distance from this bound ligand to the perturbed tryptophan. However, the 2.5-nm blue shift which SLC induces demonstrates the ability of this molecule to de- crease the polarity near the sensitive tryptophan. Further experi- ments to determine whether the site(s) at which SLFA (I), SLA (III), and SLC (IV) bind are identical, equivalent, or different are currently in progress.
Abrupt changes in the slopes for the Arrhenius plots of the motion parameter for albumin-bound spin-labeled fatty acids are also observed for the spin-labeled steroid derivative (SLA, III) and the spin-labeled indole derivative (SLC, IV). Androstyl
Acknourlegments-We thank Mr. Richard Plumlee for superior technical assistance and Ms. Debbie Mason for typing the manuscript (TBTGA).
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from

2494
REFERENCES
SPAHR, P. F., AND EDSALL, J. T. (1964) J. Biol. Chem. 239, 850-854
GOOSSENS. W.. PREAUX. G., AND LONTIE. R. (1973) Biochimie
20. ROZANTSEV, E. G., AND SHAPRIO, A. B. (1964) Izu. Akad. Nauk. SSSR. Ser. Khim. 1123-1125
21. CHEN, R. F. (1967) J. Bicl. Chem. 242, 173-181 22. ROSSENEU-MOTREFF, M., BLATON, V., DECLERCQ, B., VAN-
DAMME, D., AND PEETERS, H. (1970) J. Biochem. 68, 369-377 23. LEONARD, W. J., JR., VIJAI, K. K., AND FOSTER, J. F. (1963)
J. Biol. Chem. 238, 1984-1988 24. JACKSON, R. L., AND GOTTO, A. M. (1972) Biochim. Biophys.
Acta 286, 36-47 25. GLOMSET, J. A. (1972) in Blood Lipids and Lipoproteins: Quan-
titation, Composition and Metabolism (NELSON, G. J., ed) pp. 745-787, Wiley-Interscience, New York
26. CHAMBAZ, E., DEFAYE, G., HADJIAN, A., MARTIN, P., RA- MASSEUL, R., AND RASSAT, A. (1971) Fed. Eur. Biochem. Sot. Lett. 19, 55-59
27. PEARLMAN, W. H., AND FONG, I. F. F. (1972) J. Biol. Chem. 247, 8078-8084
28. GAMBHIR, K. K., AND MCMENAMY, R. H. (1973) J. Biol. Chem. 248, 1966-1960
29. PARKER, C. A. (1968) Photoluminescence of Solutions, pp. 72- 74, Elsevier, Amsterdam
30. FEW, A. V., OTTEWILL, R. H., AND PARREIRA, H. C. (1955) Biochim. Biophys. Acta 18, 136-137
31. MCCONNELL, H. M., AND MCFARLAND, B. G. (1972) Ann. N. Y. Acad. Sci. 196, 207-217
32. CADENHEAD, D. A., AND MUELLER-LANDAU, F. (1973) Biochim. Biophys. Acta 307, 279-286
33. SPECTOR, A. A., AND JOHN, K. M. (1968) Arch. Biochem. Bio- phys. 127, 65-71
34. GREEN, J. A., SINGER, L. A., AND PARKS, J. H. (1973) J. Chem. Phys. 68, 2690-2695
35. BIRKS, J. B. (1970) Photophysics of Aromatic Molecules, Chap. 10, Wiley-Interscience, London
36. ATTALLAH, N. A., AND LATA, G. F. (1968) Biochem. Biophys. Acta 168, 321-333
37. SWANEY, J. B., AND KLOTZ, I. M. (1970) Biochemistry 9, 2570- 2574
38. PANDE, C. S., AND MCMENAMY, R. H. (1974) Biochim. Biophys. Acta 342, 60-68
1.
2.
3.
4.
5. 6.
7.
8. 9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
66,119411207 ’ ’ .
PUTNAM, F. W. (1965) in The Proteins (NEURATH, H., ed) Vol. 3. DD. 187-199. Academic Press. New York
SPECTOR, A. A., FLETCHER, J. E., AND’ASHBROOK, J. D. (1971) Biochemistry 10, 3229-3232
JONAS, A., AND WEBER, G. (1971) Biochemistry 10, 1335-1339 PETERS, T., JR., TANIUCHI, H., AND ANFINSEN, C. B., JR.
(1973) J. Biol. Chem. 248, 2447-2451 SKIPSKI, V. P. (1972) in Blood Lipids and Lipoproteins: Quanti-
tation, Composition and Metabolism (NELSON, G. J., ed) pp. 539-545, Wiley-Interscience, New York
SWITZER, S., AND EDER, H. A. (1965) J. Lipid Res. 6,506-511 WESTPHAL, U. (1971) Steroid-Protein Interactions, Springer-
Verlag, New York MORRISETT, J. D., DAVID, J. S. K., POWNALL, H. J., AND
GOTTO, A. M., JR. (1973) Biochemistry 12, 1290-1299 SPARROW, J. T., GOTTO, A. M., AND MORRISETT, J. D. (1973)
Proc. hfatl. Acad. Sci..U. S. A. 70, 2124-2128 GOTTO. A. M.. Lux. S. E.. AND GOODMAN. D. S. (1972) Bio-
chim: Biophhs. Acia 271,‘429-435 ’ STONE, T. i., BUCKMAN, T., NORDIO, P. L., AND MCCONNELL,
H. M. (1965) Proc. Natl. Acad. Sci. U. S. A. 64, lOl(t1017 GRIGORI~N,~~. L., KALMANSON, A. E., ROZANT~EV, E. G.,
AND SUSKINA, V. S. (1967) Nature 216, 927-928 HUBBELL, W. L., AND MCCONNELL, H. M. (1971) J. Am.
Chem. Sot. 93, 314-326 KEANA, J. F. W., KEANA, S. B., AND BEETHAM, D. (1967)
J. Am. Chem. Sot. 89, 3055-3056 KUZNETSOV, A. N., EBERT, B., AND LASSMANN, G. (1973)
Stud. Biophys. 40, 173-180 HUBBELL, W. L., AND MCCONNELL, H. M. (1969) Proc. Null.
Acad. Sci. U. S. A. 63, 16-22 ROSNATI, V., AND PALAZZO, G. (1945) Gazz. Chim. Ital. 84,
644-648 by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from

J D Morrisett, H J Pownall and A M Gotto, Jrspin-labeled lipids.
Bovine serum albumin. Study of the fatty acid and steroid binding sites using
1975, 250:2487-2494.J. Biol. Chem.
http://www.jbc.org/content/250/7/2487Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/250/7/2487.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 19, 2018http://w
ww
.jbc.org/D
ownloaded from





























