Embed Size (px)
Citation preview

BMB 170 Lecture 16Lipids and Membranes, Nov 16, 2017
‘Architecture of the mammalian mechanosensitive Piezo1 channel.’ 3jac (Mouse, ~2500aa, 10-38 TMs predicted, 14 seen )Ge, Li, Zhao, Li, Chen, Zhi, Li Gao, Xiao, & Yang. Nature (2015) doi: 10.1038/nature15247
30 by TMHMM2.0

• Final problem set– Short (only three lectures)– Due Tuesday, Dec. 1st by 5pm (to Jingzhou or my office)
• Final exam– Due Friday, Dec. 8th by 5pm at my office or to Claudia– Timed – 3hrs– Short answer – taken directly from problem sets and
lecture notes– Roughly 5-10 minutes per question– Open book (no internet)– Familiarize yourself with what we covered – should be
straightforward
The end of the class
“…the hydrophobic force is the energetically dominant force for containment, adhesion, etc., in all life processes. This means that the entire nature of life as we know it is a slave to the hydrogen-bonded structure of liquid water.”
proteinwater
Charles Tanford Protein Science (1997) 6:1358-66
What happens when a protein leaves water (membrane proteins)?

-1 0 +1
Asp Ile
hydrophobicity
buriedsurfacewater-soluble proteins
buried / surfaceTM region, 16 α-helical mps
Spencer & Rees Annu Rev Biophys Biomol Struct (2002) 31:207-33Rees et al Science (1989) 245:510-3
Hydrophobicity scale: Eisenberg et al Faraday Symp Chem Soc (1982) 17:109-20
Hydrophobicities of buried/surface residues
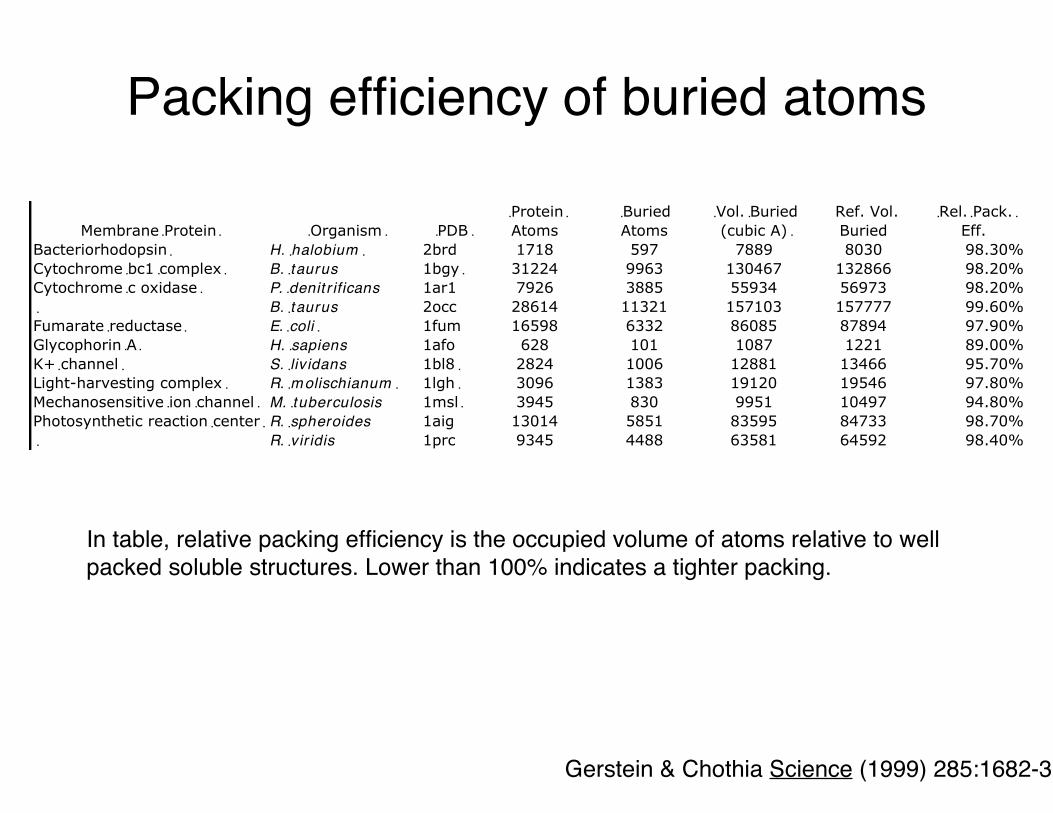
In table, relative packing efficiency is the occupied volume of atoms relative to well packed soluble structures. Lower than 100% indicates a tighter packing.
Gerstein & Chothia Science (1999) 285:1682-3
Packing efficiency of buried atoms
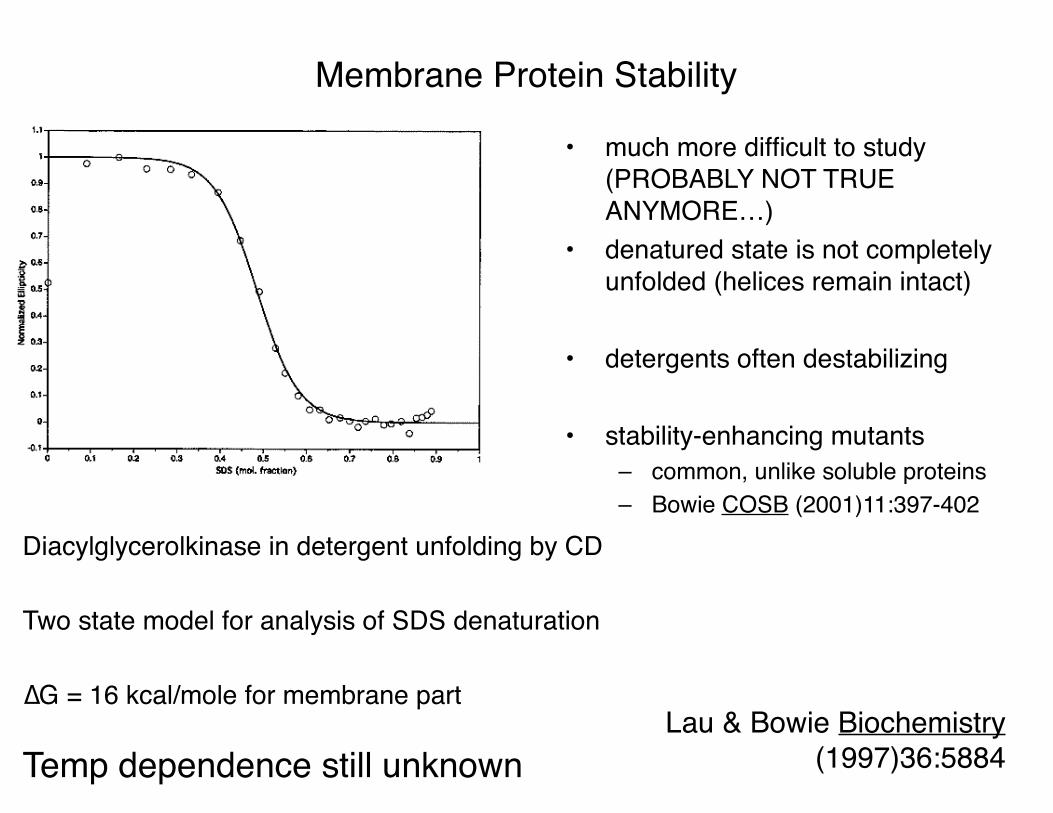
Diacylglycerolkinase in detergent unfolding by CD
Two state model for analysis of SDS denaturation
∆G = 16 kcal/mole for membrane part
Temp dependence still unknown
• much more difficult to study (PROBABLY NOT TRUE ANYMORE…)
• denatured state is not completely unfolded (helices remain intact)
• detergents often destabilizing
• stability-enhancing mutants– common, unlike soluble proteins– Bowie COSB (2001)11:397-402
Membrane Protein Stability
Lau & Bowie Biochemistry (1997)36:5884
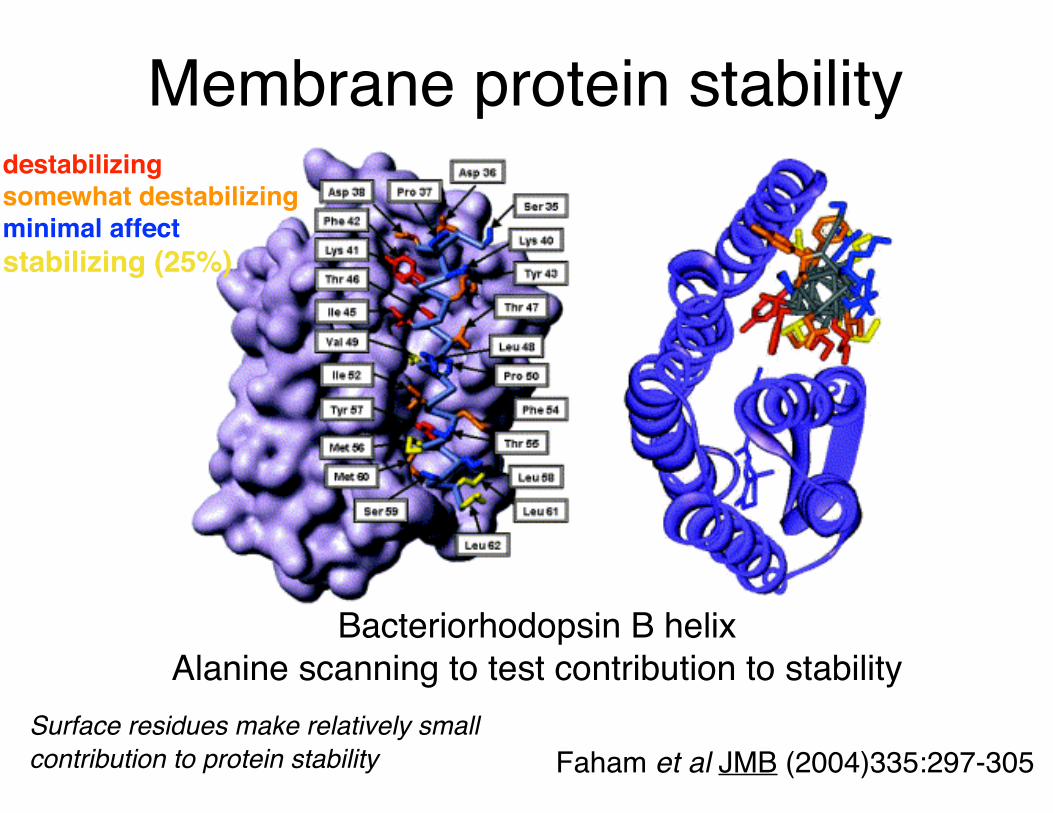
Faham et al JMB (2004)335:297-305
Bacteriorhodopsin B helixAlanine scanning to test contribution to stability
Surface residues make relatively small contribution to protein stability
destabilizingsomewhat destabilizingminimal affectstabilizing (25%)
Membrane protein stability

water membrane
• membrane proteins and water soluble proteins have similar– interior apolarities– packing densities– surface areas– patterns of residue conservation– stabilities
• and differ in – surface polarities– helix-helix packing distributions– tertiary folds
Membrane protein - water-soluble protein comparison

Slovic et al. PNAS (2004) 101:1828
Water-solubilization of KcsA
• Designed computationally then made it – confirmed binding to known inhibitors
• Different surface hydrophobicity for different solvents - same internal packing
• Water soluble proteins are membrane proteins with built-in detergent
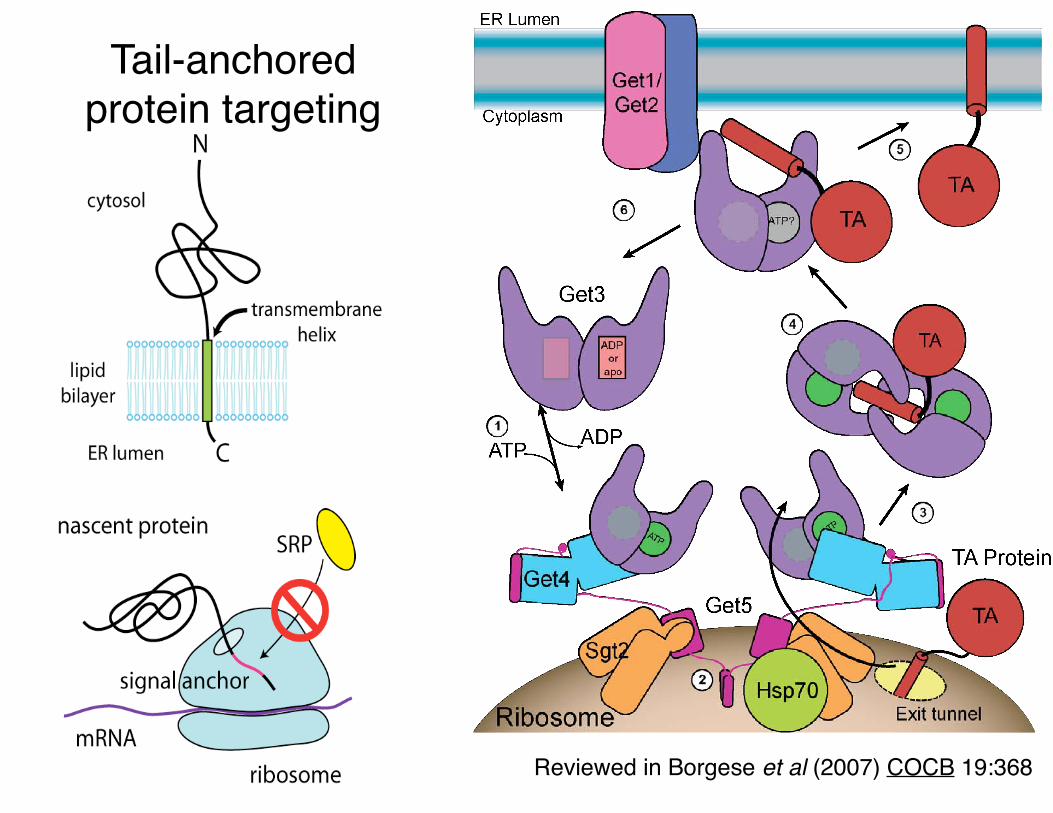
Reviewed in Borgese et al (2007) COCB 19:368
Tail-anchored protein targeting

Cyanobacterial Get3Nostoc. PCC 7120 All4481 (3IGF)
S. cerevisiae Get3 (2WOJ)
Synechocystis PCC 6803,Hohmann-Marriott et al. 2009

Fusion

Organelle fusion
Review: Wickner & Schekman (2008) NSMB 15:658

• low pH of endosomes induces conformational change
• Eventual coiled-coil leads to fusion
Wiley & Skehel labs: Bullough et al Nature (1994) 371:37-43
Fusion peptide Membrane
fusion
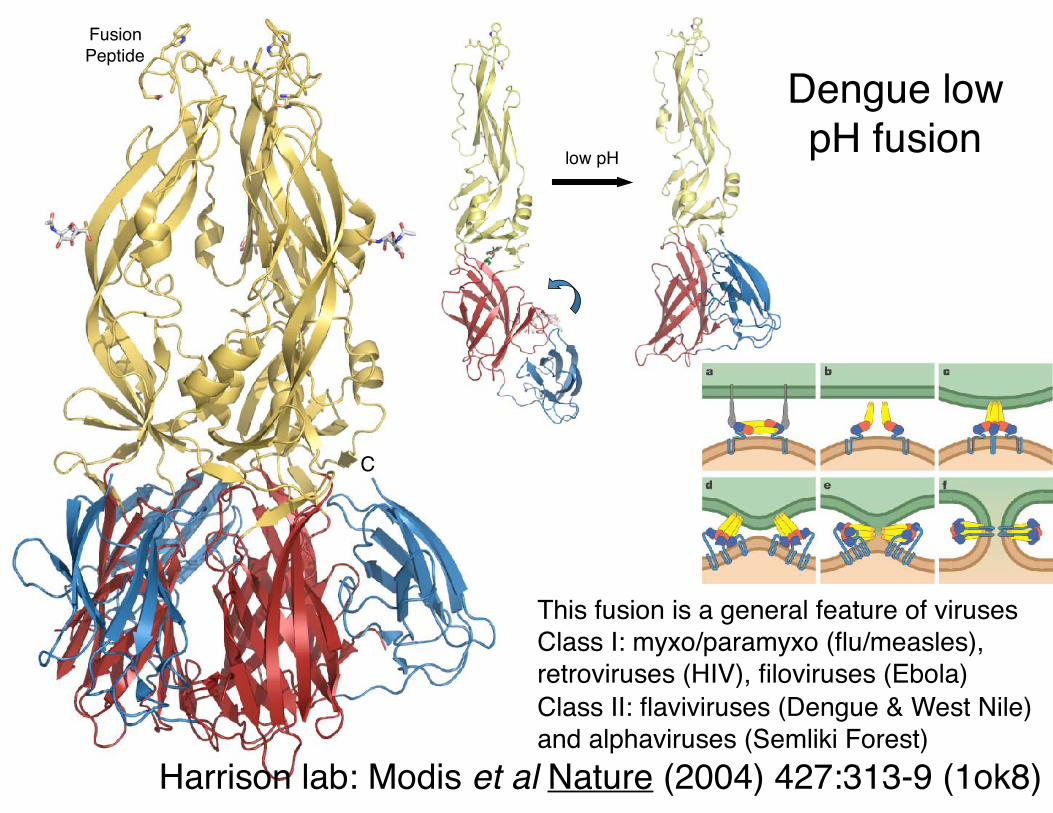
Dengue low pH fusion
Harrison lab: Modis et al Nature (2004) 427:313-9 (1ok8)
This fusion is a general feature of viruses Class I: myxo/paramyxo (flu/measles), retroviruses (HIV), filoviruses (Ebola)Class II: flaviviruses (Dengue & West Nile) and alphaviruses (Semliki Forest)
low pH
C
Fusion Peptide
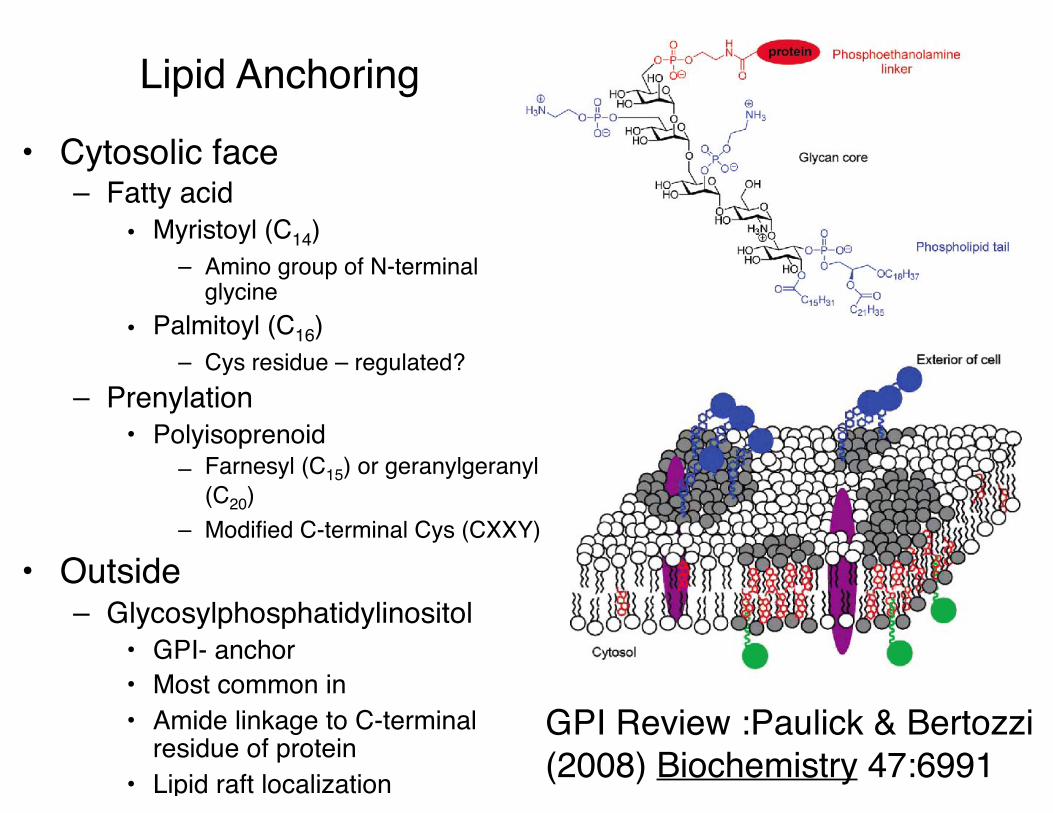
Lipid Anchoring
• Cytosolic face– Fatty acid
• Myristoyl (C14)– Amino group of N-terminal
glycine• Palmitoyl (C16)
– Cys residue – regulated?– Prenylation
• Polyisoprenoid– Farnesyl (C15) or geranylgeranyl
(C20)– Modified C-terminal Cys (CXXY)
• Outside– Glycosylphosphatidylinositol
• GPI- anchor• Most common in • Amide linkage to C-terminal
residue of protein• Lipid raft localization
GPI Review :Paulick & Bertozzi (2008) Biochemistry 47:6991

2D Crystal structure of Aqp0
Walz & Harrison lab, Gonen et al Nature (2005) 438:633-8

Weiner (2000) Structure 8:R171-5
R
a
b
α
1 n
2� R
S
n12• • •
• α
123
ab
• • •
R=radius n=number of strands
α=tilt angle S=shear numbera=Cα
β-barrel membrane proteins
Schulz (2003) Adv Prot Chem 63:47-70
OmpA 8 (1bxw)
OMPLA 12 (1qd5)
OmpF 16 (2omf) LamB 18
(1mal)
FhuA 22 (1by5 & 2fcp)
α-hemolysin 14 (7) (7ahl)
TolC 12 (3) (1ek9)
Outer membrane proteins (OMPs)
Bacteria, mitochondria and chloroplasts

Porins
Cowan et al (1992) Nature 358:727-33 PDB 1pho
bold residues face the outside of the barrel
n=16, S=20,R=15.5 Å, α=43˚

TolCPart of the multidrug efflux and
protein export pathway
Spans the periplasm
Trimer; n=12, S=20,R=13.6 Å, α=51˚Koronakis et al.(2000) Nature 405:914-9
(1ek9)
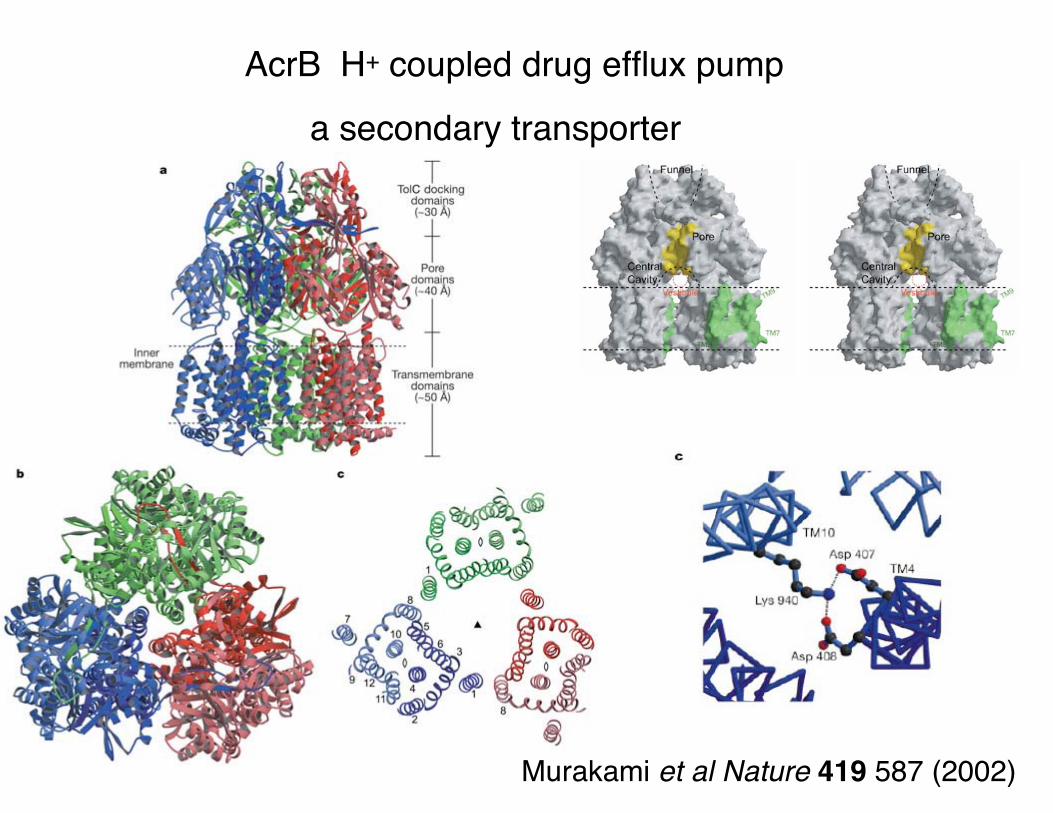
AcrB H+ coupled drug efflux pump
a secondary transporter
Murakami et al Nature 419 587 (2002)

Proposed docking model of the complex between the multidrug efflux transporter AcrB and TolC
Murakami et al (2002) Nature 419:587-93 PDB 1iwg
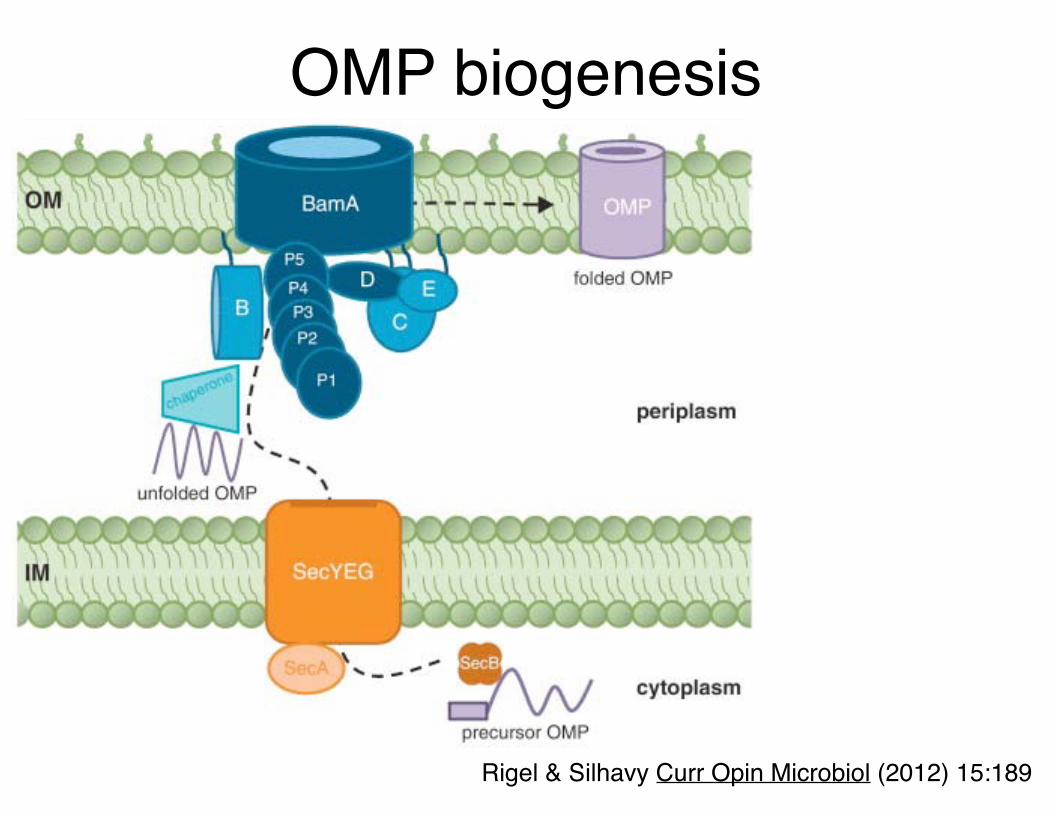
OMP biogenesis
Rigel & Silhavy Curr Opin Microbiol (2012) 15:189

BamA structure
Noinaj..Lithgow, Buchanan Nature (2013) 501:385–390 (4k3c, 4k3b)
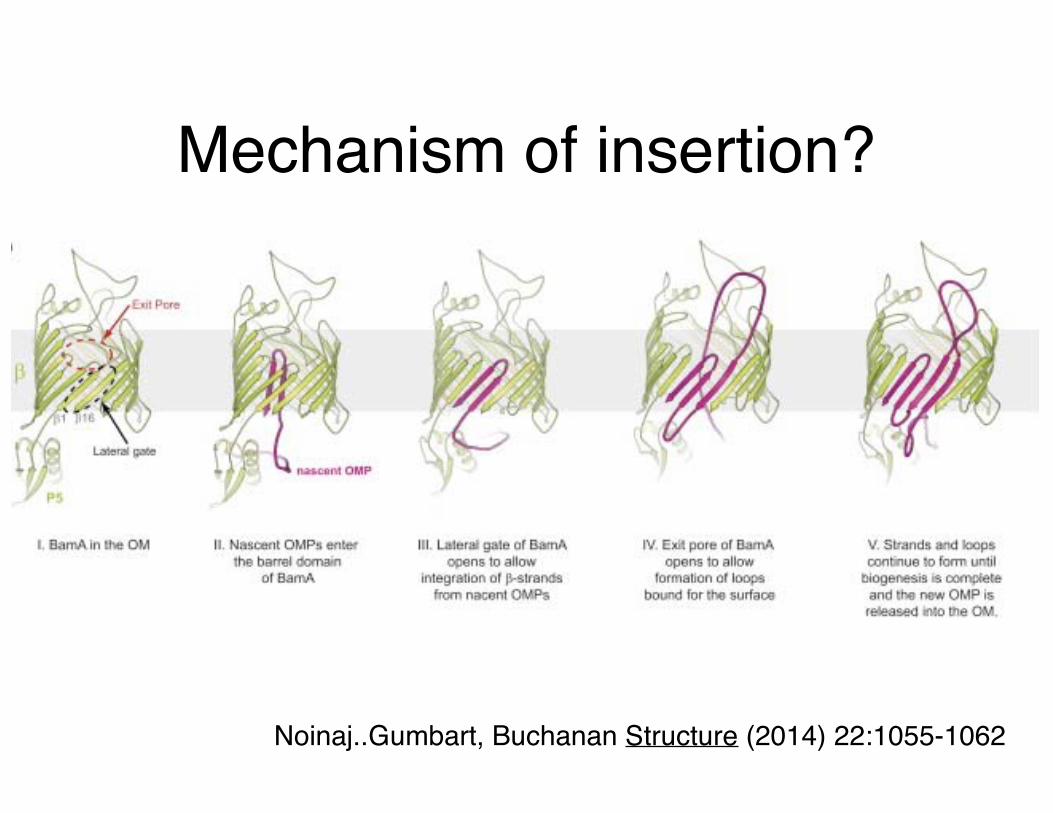
Mechanism of insertion?
Noinaj..Gumbart, Buchanan Structure (2014) 22:1055-1062

MP Expression• One of the biggest hurdles for structure
determination• Proteins must be synthesized into host membrane• Typical yield is ~1 mg/L
– in E. coli in the best cases the protein is ~0.05% of cell mass (0.5-2% for soluble proteins)
– Expression system is key• Observations
– Functional expression but no protein when overexpressed
– Strong species variability tending to match along Class divisions
– Not all E. coli MPs can be overexpressed– Often no protein by Western
• Expression seems to be working (sfGFP)• Degradation?

Isolating membrane
proteins

Detergent solubilization• Detergents form
micelles instead of bilayers
• Decision is important as micelle size affects crystallization
• Detergents have different effects on MPs so many have to be tested

Property of micelles
• Cfree = [detergent not in micelle]• Cmicelle = [detergent micelle]• N = aggregation number (molecules in
micelle)• Ctotal=NCmicelle + Cfree
• Critical micelle concentration (CMC)– Point at which NCmicelle = Cfree
– Below CMC micelles cannot form

• DHPC - diheptanoylphosphatidylcholine
• β glucosides (βOG:octyl- βNG:nonyl-)
• Maltosides (DDM:dodecyl- UDM:undecyl-DM:decyl-)
• Cn-Em – polyoxyethylene (n) dodecyl ether (10-13,6-12)
• Cymals (4-7)– cyclohexyl β maltoside
• LDAO – lauryl dimethylamineoxide
• FosCholine (7-15)
Extraction (vary detergent)

Detergent descriptions• CMC
– critical micelle concentration– Minimal concentration for
micelle formation– mM and %
• Aggregation number• Molecular weight of
micelle• These are all salt
dependent - generally increasing salt lowers CMC
• CMC mm(%), Agg#,MW• Maltosides
– DM 2.2(0.07),69,22.1– UDM 0.59(0.029),71,31.5– DDM 0.17(0.009),
114,58.2• Glucosides
– βOG 19(0.556),18,5.3– βNG 6.5(0.199)
• FosCholines– 12 1.5(0.058),55,21.4– 14 0.12(0.005),108,41
• Cymals– 5 2.4(0.119),66,32.6– 6 0.56(0.028),63,32
• LDAO 2(0.046),76,17.4
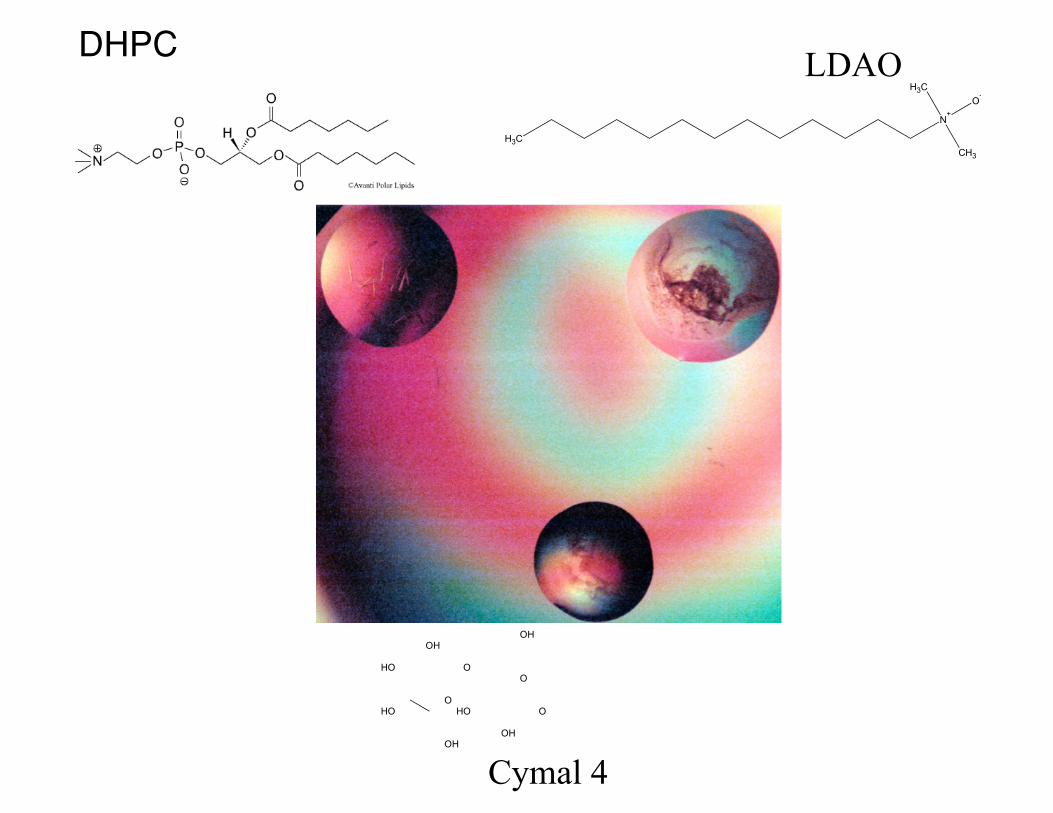
DHPC LDAO
Cymal 4

2D Crystallization• Goal is to form
crystals in bilayers• Protein is solubilized
in lipid/detergent mix• Slowly remove
detergent - leads to crystals
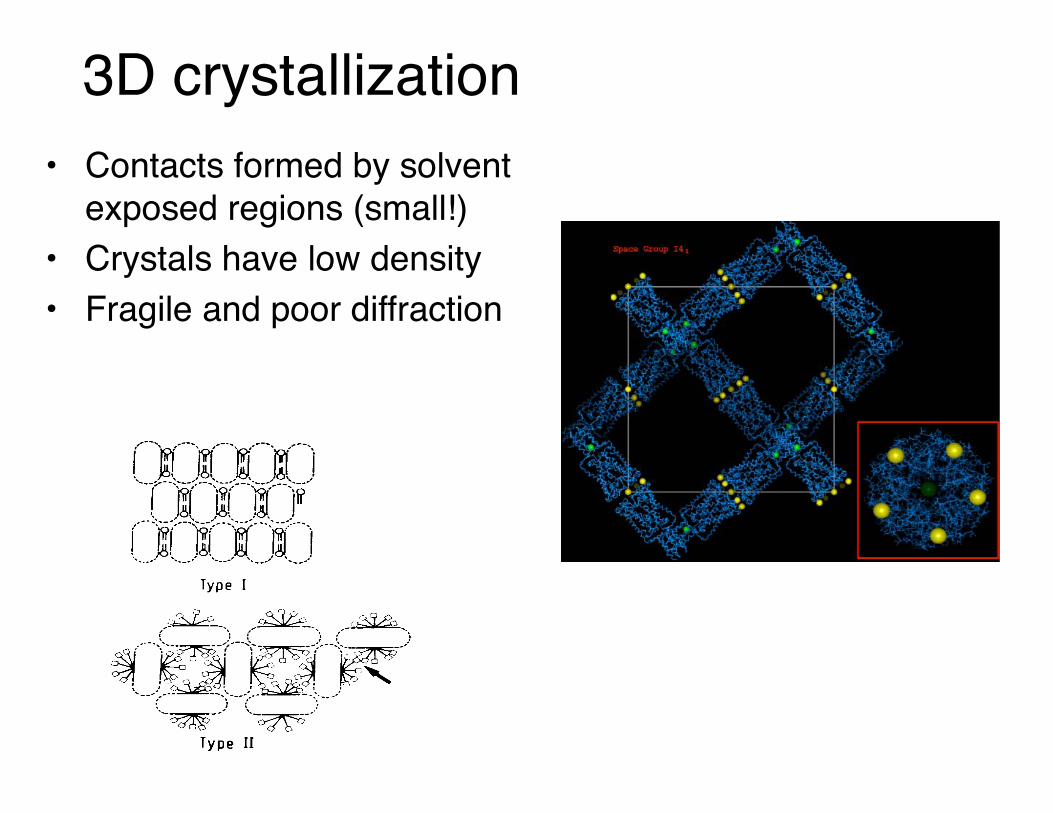
3D crystallization• Contacts formed by solvent
exposed regions (small!)• Crystals have low density• Fragile and poor diffraction

Other Methods• Antibody Fragments
– Use of Fv or Fab fragments in complexes– Majority of crystal contacts made through ab– ELISA-positive, Western-negative– Fv
• Cytochrome c oxidase (bacterial) [Michel]• bc1 complex (yeast) [Michel, Hunte]
– Fab• KcsA, ClC, KvAP [MacKinnon]
• Inserting domains– GPCRs (lysozyme)

Additional protein forms part of crystal contacts
Yeast cytochrome bc1 Complex with Fv
Hunte et al Structure (2000) 8:669-84 (1ezv)
β2-adrenergic GPCRLysozyme insert
Cherezov et al Science (2007) 318:1258 (2rh1)
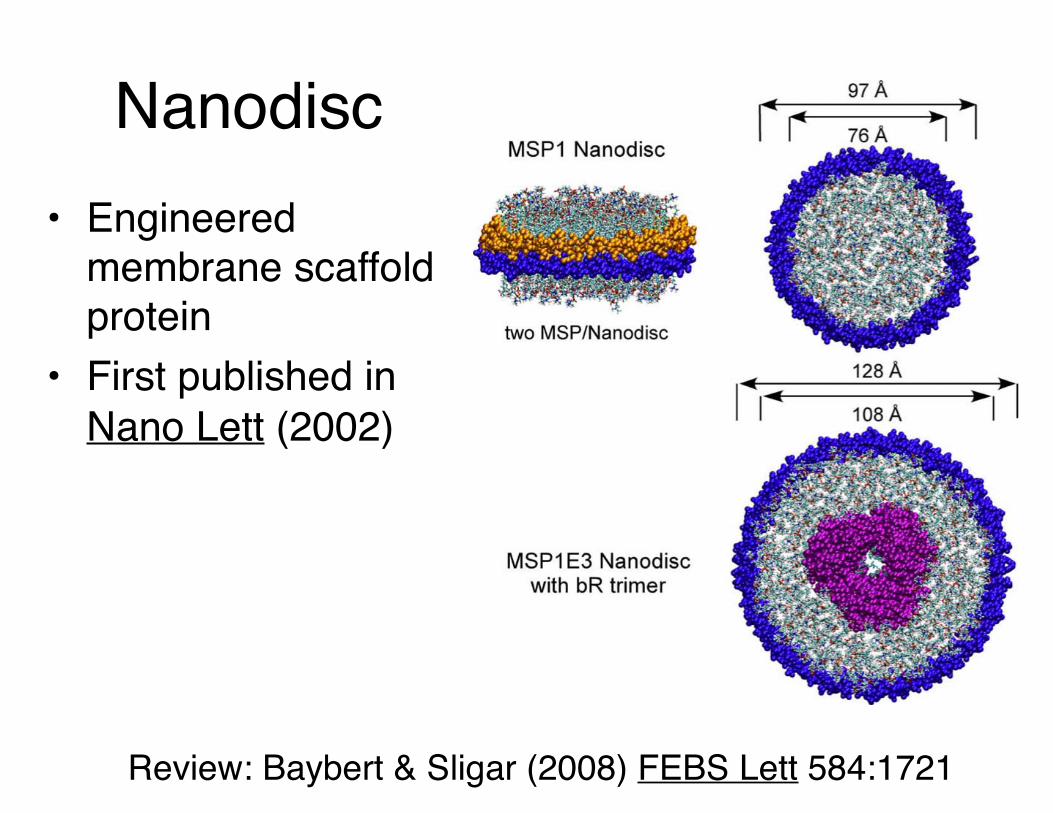
Nanodisc• Engineered
membrane scaffold protein
• First published in Nano Lett (2002)
Review: Baybert & Sligar (2008) FEBS Lett 584:1721

Nanodiscs continuedRyanodine receptor (EM 6.1Å)
KRas4b (NMR)
Review: Denisov & Sligar (2016) NSMB 23 (6) 481-6

Cherezov COSB (2011) 21:559 LCP methods review
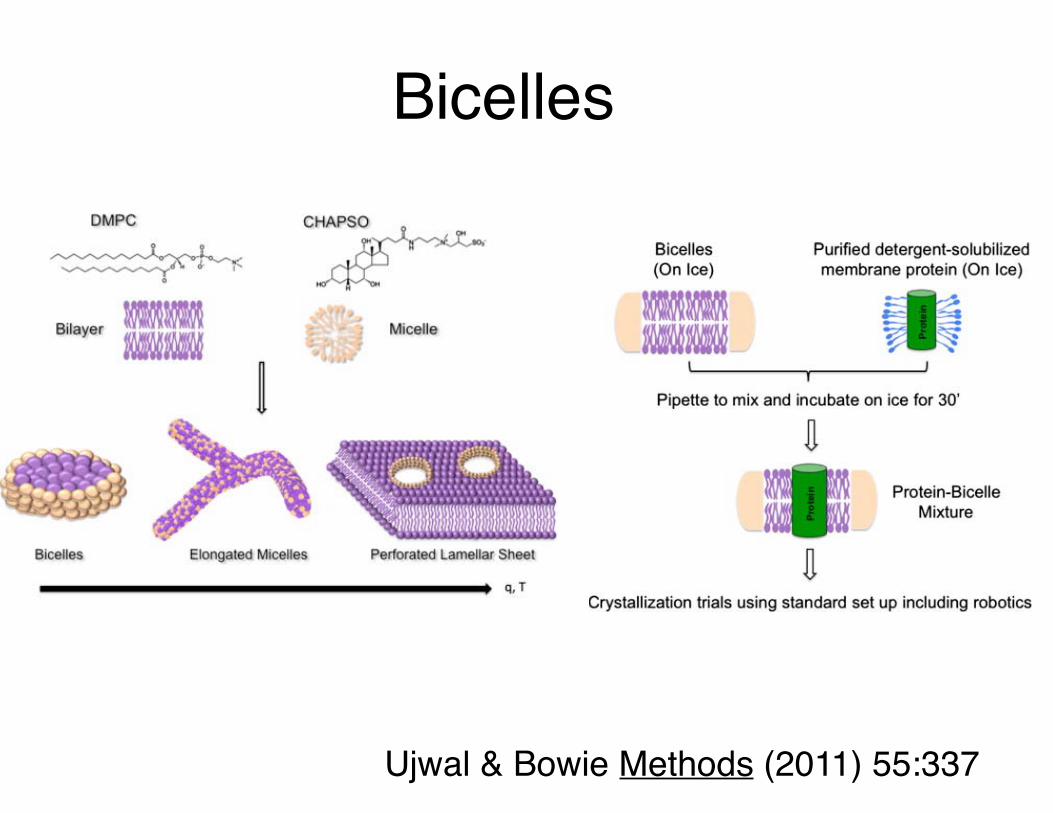
Bicelles
Ujwal & Bowie Methods (2011) 55:337

The first high resolution membrane protein structureRps. viridis photosynthetic reaction center
Deisenhofer, Michel, Huber, et al. JMB 180, 385 (1984)Nature 318, 618 (1985)
JMB 246, 429 (1995)
L, M, and H membrane-spanning subunits + cytochrome

Henderson & Unwin Nature (1975) 257:28
The beginning of membrane protein structural biology
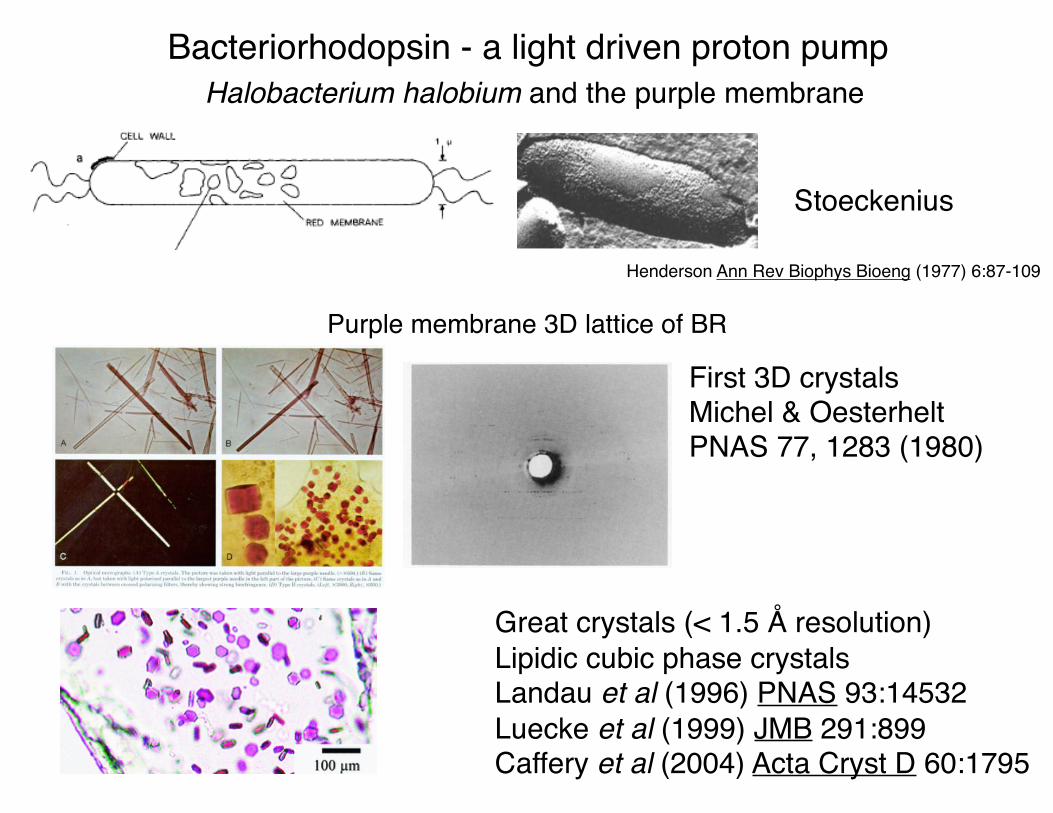
Bacteriorhodopsin - a light driven proton pump
Purple membrane 3D lattice of BR
Halobacterium halobium and the purple membrane
Stoeckenius
First 3D crystalsMichel & OesterheltPNAS 77, 1283 (1980)
Great crystals (< 1.5 Å resolution)Lipidic cubic phase crystalsLandau et al (1996) PNAS 93:14532Luecke et al (1999) JMB 291:899Caffery et al (2004) Acta Cryst D 60:1795
Henderson Ann Rev Biophys Bioeng (1977) 6:87-109

Bacteriorhodopsins and the BR photocycle
Oesterhelt (1998) Curr Opin Struc Biol 8:489-500
Cartailler & Luecke ARBBS 32, 285 (2003)

Helical packing and lipid interactions in bacteriorhodopsin
A
BC
D
E
FG
From periplasm - up/down topo
Cartailler & Luecke ARBBS 32, 285 (2003)
BR - lipid interactions
7TM - 1 retinal (1brx)Luecke et al Science (1998)
280:1934-7

1.55Å bacteriorhodopsin structure
1c3w: Luecke et al (1999) JMB 291:899

Proton pumping
• Protons are shuttled coupled to the cis-trans isomerization of retinal
Fig. 10.17 & 11.19
Excited state

How we see
cGMP phosphodiesterase

Rhodopsin• GPCR
• Bovine light receptor
• Couples light to signalling pathway
Palczewski et al (2000) Science 289:739-45 PDB 1f88

Changes with activation
• Most rhodopsin crystals lost diffraction upon exposure to light
• Red – conformational changes in activated state
• Blue – disordered when activated
Salom..Palczewski (2006) PNAS 103:16123

Transducin• Gα has two domains
– Blue – G domain– Red – helical domain
• Release goes to hydrolytically competent conformation
Gβγ
Gα
Fig. 11.24
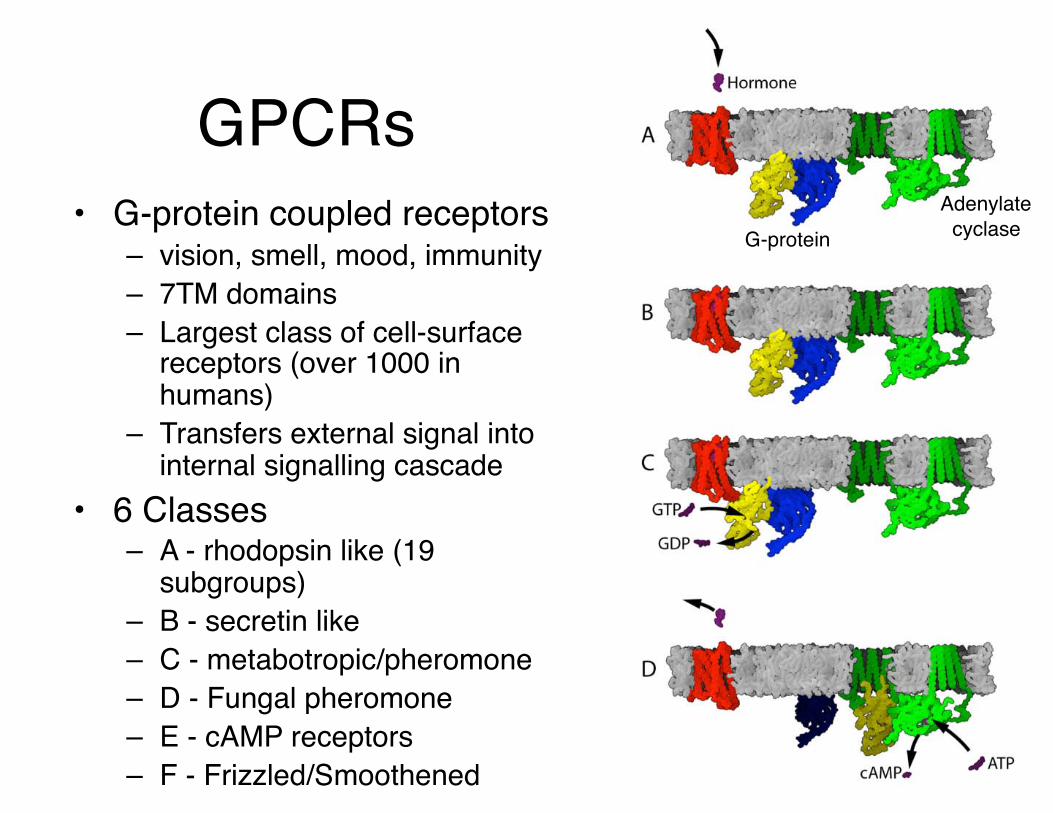
GPCRs• G-protein coupled receptors
– vision, smell, mood, immunity– 7TM domains– Largest class of cell-surface
receptors (over 1000 in humans)
– Transfers external signal into internal signalling cascade
• 6 Classes– A - rhodopsin like (19
subgroups)– B - secretin like– C - metabotropic/pheromone– D - Fungal pheromone– E - cAMP receptors– F - Frizzled/Smoothened
Adenylate cyclaseG-protein

Combinatorial coupling
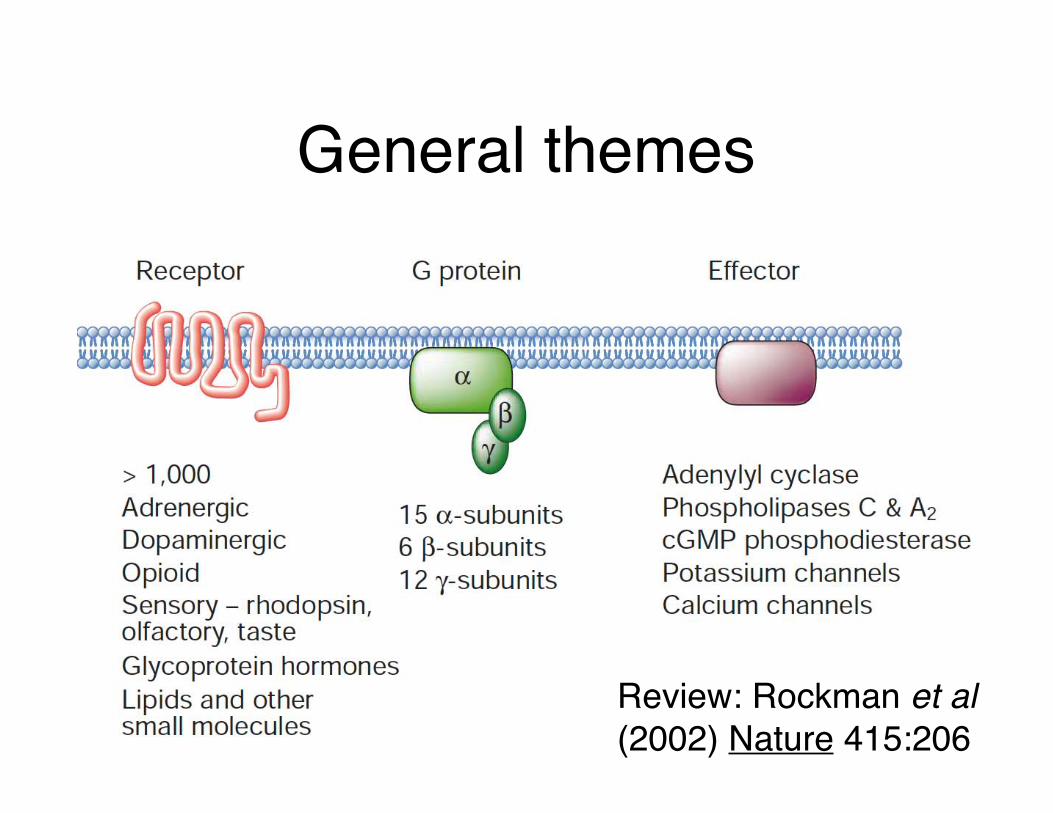
General themes
Review: Rockman et al (2002) Nature 415:206

Further components
Review: Rockman et al (2002) Nature 415:206

β-adrenergic receptor• Linked to diseases such
as asthma, hypertension and heart failure
• Found in smooth muscle • Targeted by a number of
drugs
Cherezov et al (2007) Science online 2rh1

Crystallization
Review: Rosenbaum, Rasmussen & Kobilka (2009) Nature 459:356
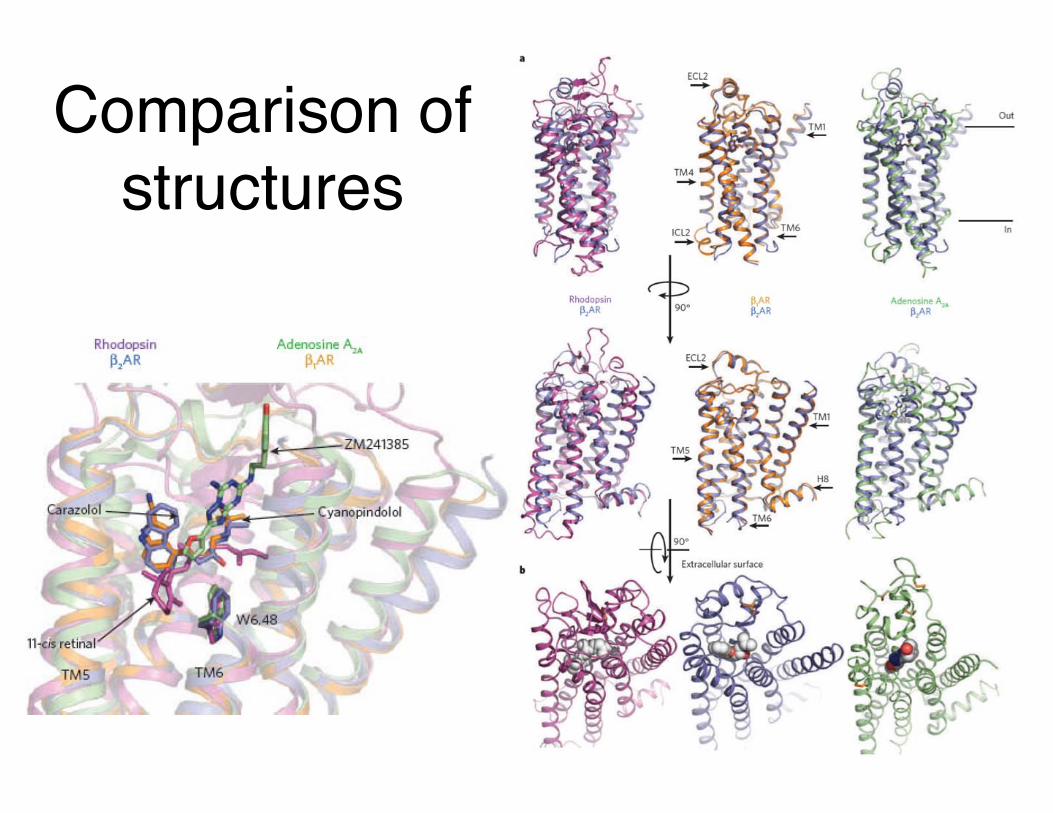
Comparison of structures

Full structure?
Rasmussen...Kobilka (2011) Nature 477:549 (3sn6)

Summary
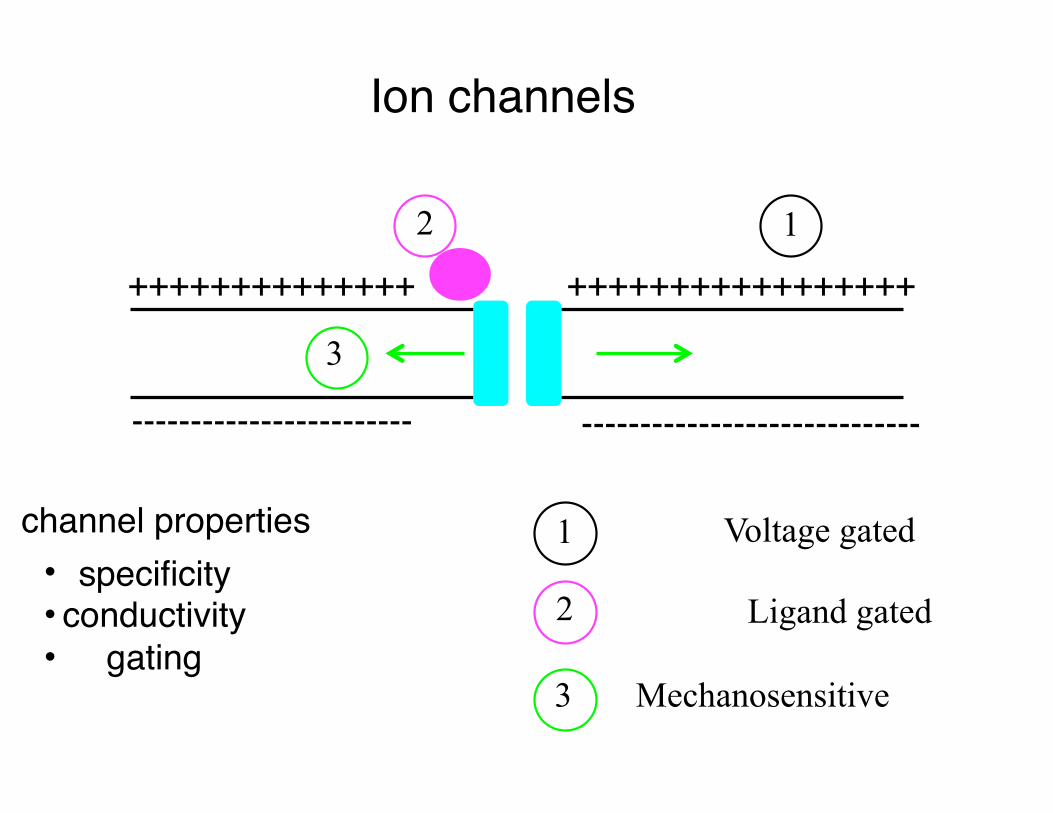
Ion channels
++++++++++++++ +++++++++++++++++
------------------------ -----------------------------
12
3
1 Voltage gated
2
3
Ligand gated
Mechanosensitive
• specificity• conductivity• gating
channel properties

Introduction to potassium channels
Hille, Ion Channels of Excitable Membranes 3rd ed (2001)
• Tetramers• 6 TM helices S1-S6• voltage sensor S4 (charges)• pore S5-S6 • selectivity filter GYG
Voltage sensor domainsPhosphoinositide phosphatase - Murata et al (2005) Nature
435:1239-43Miller (2006) Science 312:
534-5

MacKinnon lab: Doyle et al. (1998) Science 280:69 PDB 1BL8 Zhou et al. Nature 414, 43 (2001) PDB 1K4C
K+ permeation pathway

Morais-Cabral et al (2001) Nature 414:37-42
Mechanism of K+ permeation: Conduction state diagram
Zhou et al. Nature 414, 43 (2001) PDB 1K4C

KvAP structureMacKinnon lab: Opening and closing the channel in response
to changes in membrane potential - the charged S4 helix.
“voltage sensor paddles operate somewhat like hydrophobic cations
attached to levers, enabling the membrane electric field to open and
close the pore”
Jiang et al. (2003) Nature 423:33-41 1orq

Are charges in contact with membrane?
Charges fly in channel fieldControversy over how voltage-gated K+ channels sense
voltage continues to bubbleYarnell (2004) C&EN 82:35-6

Spin labeling experiments
Site directed spin labeling studies on KvAP in bilayers without FabPerozo lab: Cuello et al (2004) Science 306:491-5

buried arginines: fact or fiction?
von Heijne lab: Hessa et al (2005) Science 307:1427
“S4 is poised near the threshold of efficient
bilayer insertion, which makes sense for a voltage-dependent
switch”
“mAb fragments are not the source of non-native conformations of KvAP in crystal structures” and “a lipid membrane is required
to maintain their correct relative orientations”MacKinnon lab: Lee et al (2005) PNAS 102:15441-6

Kv1.2 crystal structure
MacKinnon Lab: Long et al (2005) Science 309:897-903 PDB 2a79
Long et al (2005) Science 309:903-8



























