Embed Size (px)
Citation preview

Experimental Gerontology 47 (2012) 170–178
Contents lists available at SciVerse ScienceDirect
Experimental Gerontology
j ourna l homepage: www.e lsev ie r .com/ locate /expgero
Blueberry extract prolongs lifespan of Drosophila melanogaster
Cheng Peng a,b, Yuanyuan Zuo a,b, Kin Ming Kwan a,b, Yintong Liang a,b, Ka Ying Ma a,b,Ho Yin Edwin Chan a,b, Yu Huang c, Hongjian Yu d, Zhen-Yu Chen a,b,⁎a School of Life Sciences, Chinese University of Hong Kong, Hong Kong, Chinab CUHK Shenzhen Research Institute, Shenzhen, Chinac School of Biomedical Science, Chinese University of Hong Kong, Hong Kong, Chinad Tianjin Jianfeng Natural Product Co., Ltd, Tianjin, China
Abbreviations: CAT, catalase; CcO, cytochrome c oxtaining superoxide dismutase; MTH, Methuselah; SOD2oxide dismutase; SOD, superoxide dismutase, ROS, rreactive oxygen species.⁎ Corresponding author at: School of Life Sciences, T
Kong, Shatin, NT, Hong Kong, China. Tel.: +852 2609 6E-mail address: [email protected] (Z.-Y. Che
0531-5565/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.exger.2011.12.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 26 July 2011Received in revised form 8 November 2011Accepted 8 December 2011Available online 17 December 2011
Section Editor: Andrzej Bartke
Keywords:CatalaseBlueberryFruit flyLifespanSuperoxide dismutase
Blueberry possesses greater antioxidant capacity than most other fruits and vegetables. The present study in-vestigated the lifespan-prolonging activity of blueberry extracts in fruit flies and explored its underlyingmechanism. Results revealed that blueberry extracts at 5 mg/ml in diet could significantly extend the meanlifespan of fruit flies by 10%, accompanied by up-regulating gene expression of superoxide dismutase(SOD), catalase (CAT) and Rpn11 and down-regulating Methuselah (MTH) gene. Intensive H2O2 and Paraquatchallenge tests showed that lifespan was only extended in Oregon-R wild type flies but not in SODn108 orCatn1 mutant strains. Chronic Paraquat exposure shortened the maximum survival time from 73 to 35 daysand decreased the climbing ability by 60% while blueberry extracts at 5 mg/ml in diet could significantlyincrease the survival rate and partially restore the climbing ability with up-regulating SOD, CAT, andRpn11. Furthermore, gustatory assay demonstrated that those changes were not due to the variation offood intake between the control and the experimental diet containing 5 mg/ml blueberry extracts. It wastherefore concluded that the lifespan-prolonging activity of blueberry extracts was at least partially associatedwith its interactions with MTH, Rpn11, and endogenous antioxidant enzymes SOD and CAT.
© 2011 Elsevier Inc. All rights reserved.
1. Introduction
Aging is a complex biological process. Oxidative stress has beenshown to increase with aging. It is believed that accumulation ofoxidative damages caused by reactive oxygen species (ROS) is oneof the major contributors responsible for aging (Harman, 1956). Toscavenge excess amount of ROS, antioxidant defenses includingendogenous antioxidant system and exogenous antioxidant intakeare indispensable. In this regard, superoxide dismutase (SOD), cata-lase (CAT), and glutathione peroxidase (GPx) serve as primary endog-enous antioxidants to deactivate ROS in vivo, while exogenousantioxidants, such as vitamin C and E, polyphenols, help to eliminatefree radicals by terminating the propagation of ROS reaction (Williset al., 2009; Matés and Sánchez-Jiménez, 1999).
Mitochondria and longevity-determined genes may play a vitalrole in aging process. Mitochondrial respiratory capacity has beenshown to decline with aging. As the terminal oxidoreductase of
idase; SOD1, copper–zinc con-, manganese containing super-eactive oxygen species; ROS,
he Chinese University of Hong382; fax: +852 2603 7246.n).
rights reserved.
mitochondrial electron transport chain (ETC), cytochrome c oxidase(CcO) shows an age-related decline in both invertebrates and verte-brates (Schwarze et al., 1998). CcO deficiency would lead to the re-duction of total ETC activity, resulting in increased amount of eithersuperoxide anion radicals or hydrogen peroxide in mitochondria(Sohal et al., 2008). In addition, Rpn11 is one lid component of themultiple subunits making up the 19S regulatory subunit, which is es-sential for 26S proteasome intact structure and activity. It has beenreported that knocking down of Rpn11 results in accumulated ubiqui-tinated proteins, reduces 26S proteasome activity, and shortenslifespan, whereas overexpressing Rpn11 can reduce age-related accu-mulation of ubiquitinated proteins, thus extending survival time(Tonoki et al., 2009). Moreover, one of the longevity-determinedgenes namely methuselah (MTH) is of great interest in Drosophila asMTH mutant flies can live 35% longer than the wild type strain aswell as they exhibit stronger resistance to exogenous oxidative stress(Lin et al., 1998). However, the specific function of MTH and itspotential connection with antioxidant defense systems remainunknown.
Fruit fly is one of the models to study aging and age-related dis-eases (Jafari, 2010). Humans actually share a huge amount of con-served biological pathways and diseases-causing genes with thistiny insect (Reiter et al., 2001; Bauer et al., 2004). Compared withother models, fruit fly is relatively easier to maintain in a largequantity due to their tiny body size and short lifespan. Previous

171C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
reports have revealed that dietary modification, including calorierestriction and dietary supplementation, can extend lifespan andameliorate certain age-related diseases (McCay et al., 1935; Linet al., 2002; Partridge et al., 2005; Lee et al., 2006; Piper andBartke, 2008).
Blueberry, containing large amounts of polyphenols, possesses agreater antioxidant capacity than most other fruits and vegetables(Prior et al., 1998). It has been reported that consumption of blueber-ry can retard age-related physiological and functional deficits (Josephet al., 2005). Krikorian et al. (2010) have recently finished theirhuman trial study evaluating the health benefits of blueberry supple-mentation, revealing that daily consumption of wild blueberry juicefor 12 weeks could improve memory function in older adults withearly memory decline. However, larger sample size and more consis-tent clinical data are lacking to draw a conclusion. As to the experi-ments conducted in animal models, Galli et al. (2006) claimed thatblueberry-supplemented diet could reverse age-related decline inhippocampal heat shock protein (HSP) neuroprotection in rats.Similarly, blueberry is also effective in enhancing cognitive andmotor behavior as well as attenuating cognitive declines in object rec-ognition memory in aged rats (Goyarzu et al., 2004). Furthermore,age-related deficits in N-methyl-D-aspartate receptor-dependentlong-term potentiation, a cellular substrate for learning and memory,are also reported to be ameliorated by blueberry-enriched diet(Coultrap et al., 2008).
No report to date has studied the effect of blueberry on the lifespanofDrosophila melanogaster. The present study was therefore to investigate(i) anti-aging activity of blueberry extracts in fruit flies; (ii) interactionbetween supplementation of blueberry extracts and gene expressionsof endogenous antioxidant enzymes, CcO subunits III and VIb, Rpn11,and MTH in D. melanogaster.
2. Materials and methods
2.1. Fly strains
Fly strains chosen in this study included Oregon-R-C (OR), OE−/SM5;Catn1/TM3 (Catn1), and SODn108/TM3 (SODn108) (BloomingtonDrosophila Stock Center, Department of Biology, Indiana University,Bloomington, IN, USA). OR is a wild type fly strain that was used inall experiments unless specified otherwise. SODn108 is a mutant strainwith one pair of single SOD gene on 3L chromosome being knockedout while Catn1 is a mutant strain with CAT gene on chromosome 3Lbeing knocked out by a point mutation.
Minu
2 4 6 8 10 12 14 1
mV
olts
0
5
10
15
20
25UV
1
2
Fig. 1. HPLC chromatogram of blueberry extracts. Peaks: 1
2.2. Diet
The basal diet was prepared according to the standard formulationdescribed previously (Li et al., 2007; Peng et al., 2009). In brief,1000 ml diet contained 105 g cornmeal, 105 g glucose, 21 g yeast,and 13 g agar. Ethyl-4-hydroxybenzoate (0.4%) was added into dietas an anti-mold agent. Two experimental diets were prepared by add-ing blueberry extracts powder at 2 mg (Bbe) or 5 mg (BBE) in thecontrol diet per milliliter. For rearing the stocks, 15 ml of the basaldiet was poured and set into a vial. For the experimental flies, 5 mlof the basal or experimental diets were prepared per vial.
2.3. HPLC analysis of blueberry extracts
Blueberry extract was obtained from Tianjin Jianfeng NaturalProduct Co., Ltd, Tianjin, China. The extraction process is shown inthe Supplementary Information. In brief, individual components inblueberry extract were separated on a C-18 (250×4.6 mm) columnand quantified on a HPLC system with a UV detector at 520 nm. Thecolumn temperature and flow rate were set at 30 °C and 0.8 ml/minrespectively. The gradient mobile phase consisted of 0.5% H3PO4
(solvent A) and H2O:acetonitrile:acetic acid:phosphoric acid(50:48:5:1:0.5, solvent B). The ratio of A to B was programmed 4:1to 2:3 in 26 min and then back to 4:1 in 4 min, and then was heldfor another 5 min. The peaks were identified according to the reten-tion times of standards. The blueberry extracts in the present studymainly contained cyanidin-3-O-glu (49.2%) and petunidin-3-O-glu(20.1%) (Fig. 1).
2.4. Effect of blueberry extracts on longevity of OR flies fed the basal diet
Two independent trials were conducted. For each trial, newlyeclosed male flies were divided into 3 groups (n=200 each), andhoused in 10 vials (20 flies per vial). The first group was maintainedon the basal diet, while the two experimental groups were fed oneof the Bbe and BBE diets. Dead flies were counted every 2–3 daysand the remaining alive flies were transferred to a new vial contain-ing the same diet. The feeding lasted 76 days (Fig. 2 A1 and A2). Themaximum life spans in this study were calculated as the averagelife span of the 5% longest surviving flies. The same experimentsdescribed above were similarly repeated and the fruit flies were sacri-ficed at selected time points (days 0, 15, 25, 35, 45, 55) in order toquantify the expression of SOD, CAT, MTH, and Rpn11.
tes
6 18 20 22 24 26 28
mV
olts
0
5
10
15
20
25
3
, cyanidin-3-O-glu; 2, petunidin-3-O-glu; 3, unknown.

A1
0%
50%
100%
DAY
Su
rviv
al R
ate
CTL
Bbe
BBE
P<0.01, BBE Vs CTL
P=0.07, Bbe Vs CTL
0%
50%
100%
DAY
Su
rviv
al R
ate
CTL
Bbe
BBE
P<0.01, BBE Vs CTL
P=0.30, Bbe Vs CTL
A2
B
0
1
2
3
4
5
20 30 40 50 60 70
20 30 40 50 60 70
CTL Bbe BBE
Red
nes
s In
dex
Fig. 2. (A1 and A2) Lifespan curve of wild type flies (OR) fed diets containing 0 mg/ml(control, CTL), 2 mg (Bbe) and 5 mg blueberry extracts (BBE)/ml diet in two separatedtrials. Data were expressed as the maximum lifespan of last fly, 50% survival time andmean lifespan (n=200 flies) for each group (Table 1). The Kaplan–Meier test foundBBE could significantly extend the mean lifespan of fruit flies (Pb0.01). (B) Gustatoryassay compared the food intake on the basis of the differences in the degree of abdomenredness among CTL, Bbe and BBE groups. Data are expressed as mean±SD.
172 C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
2.5. Gustatory assay
To exclude the possibility that lifespan extension in survival assaymight be induced by dietary restriction, gustatory assay was carriedout as described by Bahadorani and Hilliker (2008) with slight mod-ification. In brief, 60 male flies were collected (20 flies per vial) andreared on a standard diet for 5 days and then starved for 24 h on Kim-wipes paper soaked with distilled water. Afterward, flies were main-tained on the control or the BBE diet containing 0.2% sulforhodamineB sodium salt (Acid-Red) for 2 h. Then the degree of fly abdomenredness was blind-scored using a grading scale ranging from grade0 (colorless abdomen) to grade 5 (fully red abdomen). Food intakewas compared on the basis of the difference in the degree of abdomenredness between the control and BBE group (Fig. 2B).
2.6. Intensive paraquat challenge experiment
Dietary paraquat (1,1′-dimethyl-4,4′-bi-pyridinium dichloride,Sigma, St. Louis, MO, USA) is able to generate superoxide anion
radicals (Michaelis and Hill, 1933). To examine the resistance offlies against superoxide-induced oxidative stress, both OR flies(n=400 in 20 vials) and SODn108 mutant flies (n=400 in 20 vials)were maintained on their corresponding control and BBE diet, and in-cubated at 25 °C. At day 25, the fruit flies in two groups were firststarved for 2 h, and then transferred into new vials containing a filterpaper saturated with 1 ml of 20 mM paraquat in a 6% glucose solu-tion. Every 4–6 h, dead flies were counted until all flies died.
2.7. Hydrogen peroxide (H2O2) challenge test
H2O2 is able to generate a hydroxyl radical (OH). It was chosen toexamine the resistance of flies against OH-induced oxidative stress inthis study. OR flies (n=400) and Catn1 mutant flies (n=400) weremaintained on their corresponding control diet or BBE diet at 25 °Cfor 25 days. Then, the fruit flies in the two groups were first starvedfor 2 h, and then were transferred into new vials containing a filterpaper saturated with 1 ml of 30% H2O2 in a 6% glucose solution.Every 4–6 h, dead flies were counted until all flies died.
2.8. Chronic paraquat challenge
A long-term exposure to paraquat is one of the potential risk fac-tors to induce neurodegenerative diseases, such as Parkinson's dis-ease. To examine the resistance of flies against paraquat-inducedmortality and locomotor deficiency, 800 newly eclosed male OR flieswere randomly divided into three groups, namely blank controlgroup (BCTL), control group (CTL), and the BBE diet. Both BCTL andCTL flies were maintained on the control diet, while BBE group wasfed the diet containing 5 mg/ml blueberry extract. Every 3 days, fliesin CTL and BBE groups, after 2-hour starvation, were transferredinto vials containing a filter paper had saturated with 1 ml of20 mM paraquat in a 6% glucose solution. After 24 h, flies weremoved to new vials containing only water-saturated filter paper for2 h before they were transferred back into vials containing respectivediets. Flies in BCTL group also went through similar transfer cycles.However, they were exposed to 1 ml of 6% glucose solution withoutany paraquat. The experiment lasted 45 days until all the fruit fliesin BBE group died. Another set of the experiment described abovewas similarly repeated and the fruit flies were sacrificed at days 0,10, 20, and 30 to quantify the expression of SOD, CAT, MTH, Cco sub-units, ubiquitinated proteins and Rpn11.
2.9. Climbing assay
Locomotor function of fruit flies was assessed using the climbingassay as described previously (Peng et al., 2011). Briefly, 10 maleflies were placed in a plastic vial, given 10 s to climb up. At the endof each trial, the number of flies that climbed up to a vertical distanceof 8 cm or above was recorded. Each trial was performed three times.Flies were tested at days 0, 10, 20, and 30 during the chronic paraquatchallenge survival assay.
2.10. SOD activity
An assay kit was used to quantify SOD activity in fruit flies(Cayman Chemical, Michigan, USA). The principle is that a tetrazoli-um salt can detect superoxide anion radicals generated by xanthineoxidase and hypoxanthine while SOD is able to remove the superox-ide anion. In general, one unit of SOD was defined as the amount ofenzyme needed to exhibit 50% dismutation of the superoxide radical.The fruit flies (10 homogenates of 20 flies each) were homogenized in1 ml of cold 20 mM HEPES buffer (pH 7.2, with 1 mM EGTA, 210 mMmannitol and 70 mM sucrose) followed by centrifugation at a speedof 1500 g for 5 min at 4 °C. The supernatant was transferred into anew tube on ice and then subjected to centrifugation at 10,000 g for

Table 1Lifespan of OR wild type flies fed the control diet and the two experimental diets con-taining blueberry extracts at 2 mg (Bbe) and 5 mg (BBE)/ml in two separated trials.
Maximum lifespan (average of thelast 5% of surviving flies) (day)
50% Survival(day)
Mean lifespan(Mean±SD, day)
Trial 1Control 66 47 48±1a
Bbe 70 47 50±1a
BBE 71 53 53±1b
Trial 2Control 65 48 48±1a
Bbe 70 49 49±1a
BBE 72 54 54±1b
a,bMeans with different letters differ significantly at Pb0.01.
173C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
15 min at 4 °C. The supernatant contained the cytosolic copper zinccontaining SOD (CuZnSOD or SOD1), and the pellet contained mito-chondrial manganese containing SOD (MnSOD or SOD2). The super-natant was removed into a new tube and the mitochondrial pelletwas suspended in 0.5 ml cold HEPES buffer. The sample (10 μl) intriplicates was used for each test. The diluted radical detector con-taining tetrazolium salt (200 μl) was added onto 96 well plates to-gether with 10 μl sample. The reaction was initiated by adding 20 μlof diluted xanthine oxidase followed by shaking the plate for20 min at room temperature. After incubation, the absorbance wasrecorded at 450 nm in a micro-plate reader.
2.11. CAT activity
CAT was measured using a catalase assay kit (Sigma, St. Louis, MO,USA). The principle is based on the measurement of the hydrogenperoxide substrate remaining after the action of catalase present inthe sample. In brief, flies (10 homogenates of 20 flies each) were ho-mogenized in 1 ml enzyme dilution buffer followed by centrifugationat a speed of 1500 g for 5 min at 4 °C. The supernatant was movedinto a new tube and diluted 15 times by 1× assay buffer (5 mM potas-sium phosphate buffer, pH 7.0) in triplicates. The resultant sample(10 μl) was diluted again with 65 μl of 1× assay buffer. Then, 25 μlof 200 mM hydrogen peroxide solution was added to initiate thereaction. At exactly 1 min, 900μl of stop solution (15 mM sodium
SOD1 Group: P<0.05 Age: P<0.01
Group x Age: P=0.55
0
0.2
0.4
0.6
0.8
1
1.2
1.4
DAY
SO
D1
/ Rp
49
CTL
BBE
SOD2
0
0.2
0.4
0.6
0.8
1
1.2
DAY
SO
D2
/ Rp
49
CAT Group: P<0.01 Age: P<0.05
Group x Age: P<0.05
0
0.2
0.4
0.6
0.8
1
1.2
1.4
DAY
CA
T /
Rp
49
CTL
BBE
0 15 25 35 45 55 0 15 25
0 15 25 35 45 55
Fig. 3. Effect of 5 mg blueberry extracts (BBE)/ml diet on mRNA of copper–zinc containingRpn11, catalase (CAT) and Methuselah (MTH) compared with those in the control diet (C0, 15, 25, 35, 45 and 55 days. Data are expressed as mean±S.D.
azide) was added. The reaction mixture (10 μl) was mixed with1 ml color reagent containing 0.25 mM 4-aminoantipyrine, 2 mM3,5-dichloro-2-hydroxybenzenesulfonic acid, and freshly added per-oxidase (0.8–1.2 U/mg). After incubation at room temperature for15 min, the absorbance of each samplewasmeasured in a spectrometerat 520 nm.
2.12. Real-time PCR
Gene expression of SOD1, SOD2, CAT, MTH and Rpn11 was mea-sured as previously described (Peng et al., 2011). In brief, total RNAwas extracted using the commercial extraction agent TRIzol (Invitro-gen, Carslbad, CA, USA). Fruit flies (6 homogenates of 15 flies each)were homogenized in 800 μl of TRIzol solution, and then centrifugedat 12,000 g at 4 °C for 10 min. The supernatant was transferred to an-other new tube containing 160 μl chloroform. The mixture was thensubjected to centrifugation at 12,000 g at 4 °C for 15 min. The upperlayer was mixed with 400 μl isopropanol. After 10 min of incubationat room temperature, the samples were centrifuged at 12,000 g at4 °C for 10 min, and the pellet was saved and washed in 1 ml of75% ethanol followed by re-centrifugation. Finally, 25 μl DEPCwater was employed to resuspend the RNA pellet. The concentrationand purity of RNA obtained were determined by measuring theirabsorbance at 260 nm and 280 nm. High Capacity cDNA ReverseTranscription Kit (Applied Biosystems, Foster City, CA, USA) wasused to construct cDNA. RNA (2 μg) was used for each reaction to-gether with MgCl2, 10× RT buffer, dNTP, random hexamers, RNaseinhibitor, and MultiScribe Transcriptase. The final volume was ad-justed to 10 μl. cDNA was synthesized in the thermocycler GeneAmpPCR system 9700 (Applied Biosystems, Foster City, CA, USA) andstored at −20 °C.
Real-time PCR amplification was carried out on a Fast Real-timePCR System 7500 (Applied Biosystems, Foster City, CA, USA). Fivetarget genes included SOD1 (NCBI Reference Sequence NM_057387.3), SOD2 (NCBI Reference Sequence NM_057577.2), CAT (NCBIReference Sequence NM_080483.2), MTH (NCBI Reference SequenceNM_079147.2), and Rpn11 (NCBI Reference Sequence NM_135061.2). The expressions of target genes were normalized with thatof Rp49 (NCBI Reference Sequence NM_079843.2), a housekeepinggene used as the internal control. Gene expressions were calculated
Group: P<0.01 Age: P<0.01
Group x Age: P=0.72
CTL
BBE
MTH Group: P<0.05 Age: P<0.01
Group x Age: P=0.28
0
0.2
0.4
0.6
0.8
1
1.2
1.4
DAY
MT
H /
Rp
49
CTL
BBE
Rpn11 Group: P<0.05 Age:P<0.01
Group x Age: P=0.49
00.20.40.60.8
11.21.41.6
35 45 55 0 15 25 35 45 55
0 15 25 35 45 55
DAY
Rp
n11
Rp
49
CTL
BBE
superoxide dismutase (SOD1), manganese containing superoxide dismutase (SOD2),TL). The wild type (OR) flies (n=300/group, n=20/vial) were incubated at 25 °C for

174 C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
based on the comparative Threshold cycle (CT) value. Levels of geneexpressions in all groups were shown as a ratio of the day 0 controlgroup.
SOD1
0.8
1
1.2
1.4 *SOD1
0.8
1
1.2
1.4
0 15 25 35 45 55
0 15 25 35 45 55
0 15 25 35 45 55
*SOD1
0.8
1
1.2
1.4
DAY
SO
D1
/β-
acti
n
SO
D2
/β-
acti
n
CA
T /β
- ac
tin
CTL
BBE
*
C0 B0 C15 B15 C25 B25 C35 B35 C45 B45 C55 B55
SOD1
β - actin
Group: P<0.05Age: P<0.01
Group x Age: P=0.22
SOD2
0.85
0.9
0.95
1
1.05
1.1 SOD2
0.85
0.9
0.95
1
1.05
1.1
DAY
CTL
BBE
C0 B0 C15 B15 C25 B25 C35 B35 C45 B45 C55 B55
SOD2
β - actin
Group: P=0.71Age: P<0.01
Group x Age: P=0.86
CAT
0.8
0.85
0.9
0.95
1
1.05
1.1 CAT
0.8
0.85
0.9
0.95
1
1.05
1.1
C0 B0 C15 B15 C25 B25 C35 B35 C45 B45 C55 B55
CAT
β - actin
CAT
0.8
0.85
0.9
0.95
1
1.05
1.1
DAY
CTL
BBE
Group: P=0.88Age: P<0.01
Group x Age: P=0.97
Fig. 4. Effect of 5 mg blueberry extracts (BBE)/ml diet on the relative proteinmass of SOD1,SOD2 and CAT compared with those in the control diet (CTL). The wild type (OR) flies(n=300/group, n=20/vial) were incubated at 25 °C for 0, 15, 25, 35, 45 and 55 days.C0–C55 represented the protein bands for the control at days 0–55 days while B0–55represented the protein bands for BBE at days 0–55. Data are expressed as mean±SD.
2.13. Western blot analysis
Total proteins or mitochondria proteins were extracted and sub-jected to western blot analysis. For total proteins, 50 flies were homog-enized in a 1.5 ml tube containing 500 μl homogenizing buffer (20 mMTris–HCl, 2 mM MgCl2, 0.2 M sucrose and protease inhibitor cocktail(Roche, Mannheim, Germany). The extracts were centrifuged at13,000 g for 5 min at 4 °C and the supernatant was collected; As forthe mitochondria extraction, flies (8 homogenates of 50 flies each)were homogenized in mitochondrial isolation medium (MIM;250 mM sucrose, 10 mM Tris pH 7.4, 0.15 mM MgCl2). The sampleswere centrifuged twice at 1000g for 5 min to remove debris. The super-natant was centrifuged at 13,000g to obtain a mitochondria-enrichedpellet which was washed with 1 ml of MIM. The pellet was re-suspended in 50 μl MIM and the membranes were disrupted by twofreeze thaw cycles. Protein concentration was determined using a pro-tein concentration assay kit according to the manufacturer's instruc-tions (Bio-Rad, Hercules, CA, USA). After adding 6× loading dye andhomogenizing buffer to adjust the volume, the protein was boiled at95 °C for 5 min, and then stored at −80 °C. Mitochondria protein wassubjected to western blot analysis for Cco subunit III, VIb, and porin.
SOD1
0
25
50
75
100
0
25
50
75
100
0 15 25 35 45 55
0 15 25 35 45 55
0 15 25 35 45 55
DAY
DAY
SO
D1
Act
ivit
y (U
/µg
Pro
tein
)S
OD
2 A
ctiv
ity
(U/µ
g P
rote
in)
CA
T A
ctiv
ity
(U/m
g P
rote
in)
CTL
BBE
SOD2
CTL
BBE
CAT
0
100
200
300
400
DAY
CTL
BBE
Group: P<0.05Age: P<0.01
Group x Age: P=0.24
Group: P<0.01Age: P<0.01
Group x Age: P=0.13
Group: P<0.05Age: P<0.01
Group x Age: P=0.26
Fig. 5. Effect of 5 mg blueberry extracts (BBE)/ml diet on enzymatic activity of SOD1,SOD2 and CAT compared with those in the control diet (CTL). The wild type (OR)flies (n=300/group, n=20/vial) were incubated at 25 °C for 0, 15, 25, 35, 45, and55 days. Data are expressed as mean±SD.

A
0%
50%
100%
0%
50%
100%
0 10 20 30 40 50
DAY
0 10 20 30
DAY
Su
rviv
al R
ate
BCTL
CTL
BBE
B
Clim
bin
g a
bili
ty
BCTL
CTL
BBE
Fig. 7. (A) Lifespan of OR fruit flies fed either a basal diet without exposure to paraquat
175C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
For the measurement of CAT and β-actin, 20 μg total protein weresize-fractionated on 7% SDS-PAGE gel at 130 V for 70 min, while thesame amount of total protein was loaded to measure SOD1 andSOD2 on 15% SDS-PAGE gel at 130 V for 180 min. To measure Cco sub-unit III, VIb, and porin, 10 μg mitochondria protein was size-fractionated on 10% SDS-PAGE gel at 110 V for 100 min. The proteinwas then transferred to a Hybond-P PVDF membrane (Millipore, Bil-lerica, MA, USA). The membranes with total proteins were incubatedfor 1 h in blocking solution (5% non-fat milk) at room temperatureand then in the same solution containing diluted anti-catalase(Abcam, Cambridge, USA) / anti-actin (Santa Cruz Biotechnology,Inc., California, USA) / anti-CuZnSOD (Abcam, Cambridge, USA) /anti-MnSOD (Abcam, Cambridge, USA) / anti-Ub (Mito Sciences,USA) antibodies respectively at 4 °C overnight. For immunodetectionof mitochondria proteins, membranes were incubated with the anti-CcO subunits III, VIb, and porin (Mito Sciences, USA) at 37 °C for 1 h.The membrane was washed in 1× TBST and was then incubated for1 h at 4 °C in diluted horseradish peroxidase-linked goat anti-rabbitIgG (Santa Cruz Biotechnology, Inc., California, USA) or anti-mouseIgG (Santa Cruz Biotechnology, Inc., California, USA). The washeswere repeated before the membranes were developed with ECLenhanced chemiluminescence agent (Santa Cruz Biotechnology, Inc.,California, USA) and subjected to autoradiography for 1 s to 5 minon SuperRX medical X-ray film (Fuji, Tokyo, Japan). Densitometrywas quantified using the computer software Quantity one (Bio-Rad,CA, USA).
(BCTL) or a basal diet with the paraquat chronic exposure (CTL) with addition of 5 mgblueberry extracts (BBE)/ml diet at 25 °C. The Kaplan–Meier test found BBE group sur-vived better than the CTL (Pb0.01). (B) Effect of BBE on the climbing ability in BCTL,CTL, and BBE groups. BBE group could significantly alleviate locomotor deficiency inCTL group.
2.14. Statistics
Data were expressed as mean±standard deviation. All survival as-says, gustatory assay were conducted twice. Real Time PCR, Westernblot, and Enzyme Activity assays were also conducted at least twicewith three parallel samples in each group. The Kaplan–Meier test wasemployed to compare the difference between the survival curvesusing SPSS 15.0 (Statistical Package for the Social Sciences software,SPSS Inc, Chicago, USA). The significance of difference between meanswas assessed using two-way ANOVA (age and diet as two variables).Differences were considered significant when Pb0.05.
Wild Type Flies
0%
50%
100%
0%
50%
100%
0 20 40 60
Hour
Su
rviv
al R
ate
Su
rviv
al R
ate
CTL
BBE
Wild Type Flies
0 50 100
Hour
CTL
BBE
H2O2 Treatm
Paraquat Trea
P<0.01
P<0.01
Fig. 6. Effect of paraquat treatment or hydrogen peroxide treatment on the survival time of tor 5 mg blueberry extracts (BBE)/ml compared with that of the wild type (OR) flies. The KaplOR control (Pb0.05) while the survival rate of BBE-fed SODn108 and Catn1 groups was not d
3. Results
3.1. Effect of BBE on longevity of OR flies fed the basal diet and BBE diets
Two separated survival trials showed similar results. Fruit flies inBBE (5 mg/ml) group had the longest survival time among the threegroups of OR wild type male flies (Fig. 2 A1 and A2). In the first
Cat n1 Flies
CTL
BBE
0%
50%
100%
0%
50%
100%
0 20 40 60
Hour
Su
rviv
al R
ate
Su
rviv
al R
ate
SOD n108 Flies
0 20 40 60
Hour
CTL
BBE
ent
tment
P=0.79
P=0.92
he mutant flies (SODn108) or mutant flies (Catn1) fed the diets containing 0 mg/ml (CTL)an–Meier test found both that BBE-fed OR group survived better than its correspondingifferent from their corresponding control groups.

176 C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
trial, the maximum lifespan increased more than 5% in BBE groupwith the 50% survival time being increased from 47 days to 53 dayscompared with that of the control flies. The mean lifespan for the con-trol, Bbe and BBE groups were 48, 50, and 53 days, respectively (Fig. 2A1; Table 1). In the second trial, the maximum lifespan also increasedmore than 5% in BBE group with the 50% survival time being in-creased from 48 days to 54 days compared with that of the controlflies. The mean lifespan for the control, Bbe and BBE groups were
Fig. 8. The relative immunoreactive mass of SOD1, SOD2, CAT, CcO Subunit III, CcO Sub-unit VIb, and ubiquitinated proteins in OR fruit flies fed either a basal diet withoutgoing through the paraquat challenge cycle (BCTL; BC10–BC30) or a basal diet withthe paraquat chronic exposure (CTL; CL0–CL30) with addition of 5 mg blueberry ex-tracts (BBE/ml (BB10–BB30). The results were normalized with the values of day0 being 1.0.
Fig. 8 (continued).
48, 49, and 54 days, respectively (Fig. 2 A2; Table 1).Therein, a signif-icant difference was found between BBE and the control in both trials(Pb0.01).
As to the gene expression, both SOD1 and SOD2 were significantlyup-regulated in BBE group compared with that in the control (Pb0.05for SOD1; Pb0.01 for SOD2, Fig. 3). It was also observed that the geneexpression of SOD1 and SOD2 was decreasing with the age in both theBBE and control flies (Pb0.01). Similarly, expression of CAT andRpn11 was increased in BBE group compared with that in the control.In contrast, MTH gene decreased with the age and was significantlydown-regulated in BBE group compared with that in the control(Fig. 3). The detail ANOVA results for groups, ages and groups×ageshereafter in each figure were presented as in Supplemental Informa-tion (attached file).
177C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
Western blot data showed that BBE group had protein mass ofSOD1 greater than that in the control (Pb0.05, Fig. 4). This studyalso investigated effect of BBE on the activity of SOD1, SOD2 and CATin OR wild type male flies at days 0, 15, 25, 35, 45, and 55. Comparedwith the control group, BBE could significantly increase the activity ofSOD1, SOD2 and CAT (Fig. 5).
3.2. Effect of BBE on Paraquat and H2O2-challenged OR, Catn1 andSODn108 flies
Intensive paraquat challenge test revealed that BBE OR group hada longer survival time than the control flies (Pb0.01). The maximumsurvival time was increased from 74 h in the control to 88 h in BBEgroup, while the mean survival time was prolonged by 16%(Pb0.01). However, no difference was observed in SODn108
flies fedthe control and BBE diet. Similarly, supplementation of BBE prolongedthe maximum survival time, 50% survival time and mean survivaltime only in OR wild type but not in Catn1 fly strain (Fig. 6).
3.3. Effect of BBE on chronic paraquat challenge in OR flies
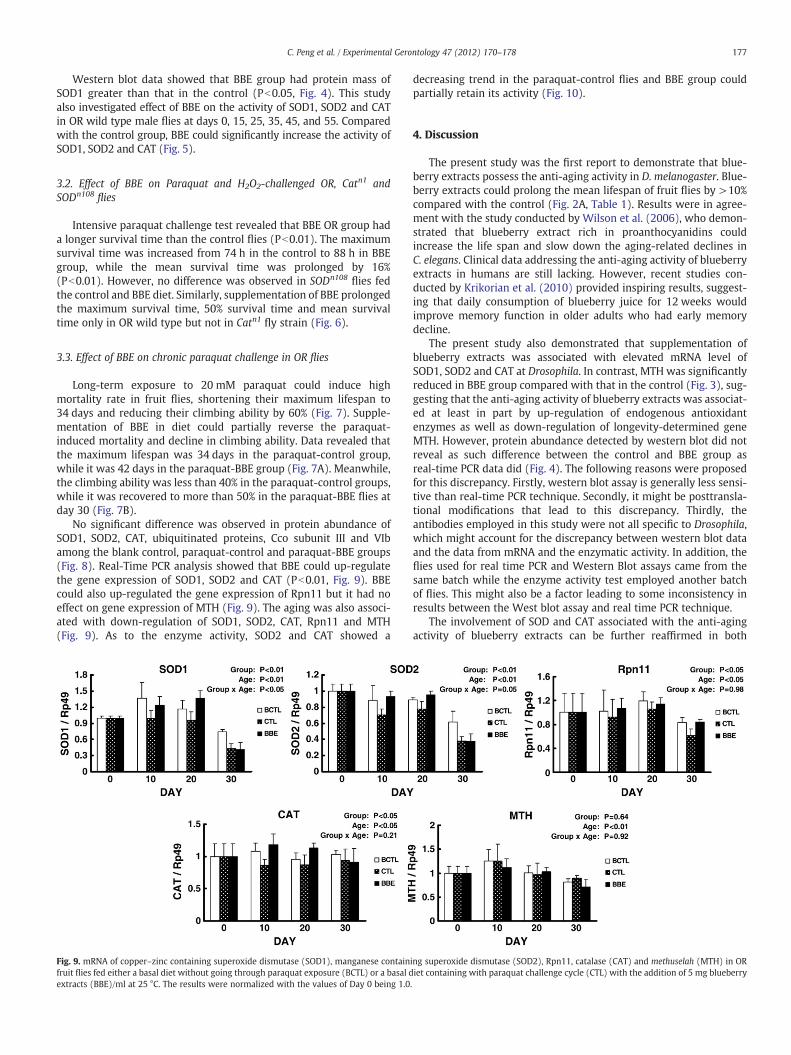
Long-term exposure to 20 mM paraquat could induce highmortality rate in fruit flies, shortening their maximum lifespan to34 days and reducing their climbing ability by 60% (Fig. 7). Supple-mentation of BBE in diet could partially reverse the paraquat-induced mortality and decline in climbing ability. Data revealed thatthe maximum lifespan was 34 days in the paraquat-control group,while it was 42 days in the paraquat-BBE group (Fig. 7A). Meanwhile,the climbing ability was less than 40% in the paraquat-control groups,while it was recovered to more than 50% in the paraquat-BBE flies atday 30 (Fig. 7B).
No significant difference was observed in protein abundance ofSOD1, SOD2, CAT, ubiquitinated proteins, Cco subunit III and VIbamong the blank control, paraquat-control and paraquat-BBE groups(Fig. 8). Real-Time PCR analysis showed that BBE could up-regulatethe gene expression of SOD1, SOD2 and CAT (Pb0.01, Fig. 9). BBEcould also up-regulated the gene expression of Rpn11 but it had noeffect on gene expression of MTH (Fig. 9). The aging was also associ-ated with down-regulation of SOD1, SOD2, CAT, Rpn11 and MTH(Fig. 9). As to the enzyme activity, SOD2 and CAT showed a
Fig. 9. mRNA of copper–zinc containing superoxide dismutase (SOD1), manganese containfruit flies fed either a basal diet without going through paraquat exposure (BCTL) or a basal dextracts (BBE)/ml at 25 °C. The results were normalized with the values of Day 0 being 1.0
decreasing trend in the paraquat-control flies and BBE group couldpartially retain its activity (Fig. 10).
4. Discussion
The present study was the first report to demonstrate that blue-berry extracts possess the anti-aging activity in D. melanogaster. Blue-berry extracts could prolong the mean lifespan of fruit flies by >10%compared with the control (Fig. 2A, Table 1). Results were in agree-ment with the study conducted by Wilson et al. (2006), who demon-strated that blueberry extract rich in proanthocyanidins couldincrease the life span and slow down the aging-related declines inC. elegans. Clinical data addressing the anti-aging activity of blueberryextracts in humans are still lacking. However, recent studies con-ducted by Krikorian et al. (2010) provided inspiring results, suggest-ing that daily consumption of blueberry juice for 12 weeks wouldimprove memory function in older adults who had early memorydecline.
The present study also demonstrated that supplementation ofblueberry extracts was associated with elevated mRNA level ofSOD1, SOD2 and CAT at Drosophila. In contrast, MTH was significantlyreduced in BBE group compared with that in the control (Fig. 3), sug-gesting that the anti-aging activity of blueberry extracts was associat-ed at least in part by up-regulation of endogenous antioxidantenzymes as well as down-regulation of longevity-determined geneMTH. However, protein abundance detected by western blot did notreveal as such difference between the control and BBE group asreal-time PCR data did (Fig. 4). The following reasons were proposedfor this discrepancy. Firstly, western blot assay is generally less sensi-tive than real-time PCR technique. Secondly, it might be posttransla-tional modifications that lead to this discrepancy. Thirdly, theantibodies employed in this study were not all specific to Drosophila,which might account for the discrepancy between western blot dataand the data from mRNA and the enzymatic activity. In addition, theflies used for real time PCR and Western Blot assays came from thesame batch while the enzyme activity test employed another batchof flies. This might also be a factor leading to some inconsistency inresults between the West blot assay and real time PCR technique.
The involvement of SOD and CAT associated with the anti-agingactivity of blueberry extracts can be further reaffirmed in both
ing superoxide dismutase (SOD2), Rpn11, catalase (CAT) and methuselah (MTH) in ORiet containing with paraquat challenge cycle (CTL) with the addition of 5 mg blueberry.

Group: P=0.77Age: P<0.01
Group x Age: P=0.99
0
25
50
75S
OD
1 A
ctiv
ity
(U/µ
g P
rote
in)
BCTL
CTL
BBE
SOD2
SOD1
Group: P<0.05Age: P<0.01
Group x Age: P=0.74
0
25
50
75
SO
D2
Act
ivit
y (U
/µg
Pro
tein
)
BCTL
CTL
BBE
CAT Group: P<0.05Age: P<0.01
Group x Age: P=0.33
0
50
100
150
200
250
300
350
0 10 20 30
DAY
CA
T A
ctiv
ity
(U/m
g P
rote
in)
BCTL
CTL
BBE
Fig. 10. Enzymatic activity of SOD1, SOD2 and CAT in OR fruit flies fed either a basal dietwithout going through paraquat exposure (BCTL) or a basal diet containing with para-quat challenge cycle (CTL) with addition of 5 mg blueberry extracts (BBE)/ml at 25 °C.
178 C. Peng et al. / Experimental Gerontology 47 (2012) 170–178
paraquat and H2O2 challenge tests. Results in intensive Paraquat /H2O2 challenge assays showed that BBE could only prolong thesurvival time in OR wild type flies but did not affect that of SODn108
and Catn1 mutant strains, in which gene of either SOD or CAT waspartially knocked out (Fig. 6), supporting the hypothesis that theanti-aging activity of blueberry was possibly associated in part withits interaction with genes of SOD and CAT.
Blueberry extracts in diet could partially and significantly reversethe chronic Paraquat exposure-induced high mortality rate and de-cline in climbing ability (Fig. 7), demonstrating that superoxideanion radicals generated by Paraquat could accelerate while dietaryBBE could delay the aging process. Chronic paraquat exposure hasbeen regarded as one of the factors contributing to neurodegenera-tive disorder such as Parkinsonian syndrome. Data collected from sur-vival and locomotor activity tests demonstrated that blueberryextracts could help ameliorate the neurodegenerative disorder.With chronic paraquat exposure, gene expression of SOD1, SOD2,CAT, and Rpn11 were down-regulated. Blueberry extracts couldpartially restore the decreasing trend.
In conclusion, blueberry extracts in diet could prolong the meanlifespan, alleviate paraquat-induced mortality rate and partiallyreverse the decline of locomotor deficiency in fruit flies. The anti-aging activity and anti-neurodegenerative disorder of blueberrywere at least partially associated with its interaction with genesSOD, CAT, Rpn11, and MTH.
Appendix A. Supplementary data
Supplementary data to this article can be found online at doi:10.1016/j.exger.2011.12.001.
References
Bahadorani, S., Hilliker, A.J., 2008. Cocoa confers life span extension in Drosophila mel-anogaster. Nutr. Res. 28, 377–382.
Bauer, J.H., Goupil, S., Garber, G.B., Helfand, S.L., 2004. An accelerated assay for the iden-tification of lifespan-extending interventions in Drosophila melanogaster. Proc.Natl. Acad. Sci. U. S. A. 101, 12980–12985.
Coultrap, S.J., Bickford, P.C., Browning, M.D., 2008. Blueberry-enriched diet amelioratesage-related declines in NMDA receptor-dependent LTP. Age (Dordr.) 30, 263–272.
Galli, R.L., Bielinski, D.F., Szprengiel, A., Shukitt-Hale, B., Joseph, J.A., 2006. Blueberrysupplemented diet reverses age-related decline in hippocampal HSP70 neuropro-tection. Neurobiol. Aging 27, 344–350.
Goyarzu, P., Malin, D.H., Lau, F.C., Taglialatela, G., Moon, W.D., Jennings, R., Moy, E.,Moy, D., Lippold, S., Shukitt-Hale, B., Joseph, J.A., 2004. Blueberry supplementeddiet: effects on object recognition memory and nuclear factor-kappa B levels inaged rats. Nutr. Neurosci. 7, 75–83.
Harman,D., 1956. Aging: a theorybased on free radical and radiation chemistry. J. Gerontol.11, 298–300.
Jafari, M., 2010. Drosophila melanogaster as a model system for the evaluation of anti-aging compounds. Fly (Austin) 4 [Epub ahead of print].
Joseph, J.A., Shukitt-Hale, B., Casadesus, G., 2005. Reversing the deleterious effects ofaging on neuronal communication and behavior: beneficial properties of fruitpolyphenolic compounds. Am. J. Clin. Nutr. 81, 313S–316S.
Krikorian, R., Shidler, M.D., Nash, T.A., Kalt, W., Vinqvist-Tymchuk, M.R., Shukitt-Hale,B., Joseph, J.A., 2010. Blueberry supplementation improves memory in older adults.J. Agric. Food Chem. 58, 3996–4000.
Lee, G.D., Wilson, M.A., Zhu, M., Wolkow, C.A., de Cabo, R., Ingram, D.K., Zou, S., 2006.Dietary deprivation extends lifespan in Caenorhabditis elegans. Aging Cell 5,515–524.
Li, Y.M., Chan, H.Y., Huang, Y., Chen, Z.Y., 2007. Green tea catechins upregulate superoxidedismutase and catalase in fruit flies. Mol. Nutr. Food Res. 51, 546–554.
Lin, Y.J., Seroude, L., Benzer, S., 1998. Extended life-span and stress resistance in theDrosophila mutant methuselah. Science 282, 943–946.
Lin, S.J., Kaeberlein, M., Andalis, A.A., Sturtz, L.A., Defossez, P.A., Culotta, V.C., Fink, G.R.,Guarente, L., 2002. Calorie restriction extends Saccharomyces cerevisiae lifespan byincreasing respiration. Nature 418, 344–348.
Matés, J.M., Sánchez-Jiménez, F., 1999. Antioxidant enzymes and their implications inpathophysiologic processes. Front. Biosci. 4, 339–345.
McCay, C.M., Crowell, M.F., Maynard, L.A., 1935. The effect of retarded growth upon thelength of life and upon the ultimate body size. J. Nutr. 10, 63–79.
Michaelis, L., Hill, E.S., 1933. Potentiometric studies of semi-quinones. J. Am. Chem. Soc.55, 1481–1494.
Partridge, L., Piper, M.D., Mair, W., 2005. Dietary restriction in Drosophila. Mech. AgingDev. 126, 938–950.
Peng, C., Chan, H.Y., Li, Y.M., Huang, Y., Chen, Z.Y., 2009. Black tea theaflavins extend thelifespan of fruit flies. Exp. Gerontol. 44, 773–783.
Peng, C., Chan, H.Y., Huang, Y., Yu, H., Chen, Z.Y., 2011. Apple polyphenols extend themean lifespan of Drosophila melanogaster. J. Agric. Food Chem. 59, 2097–2106.
Piper, M.D., Bartke, A., 2008. Diet and aging. Cell Metab. 8, 99–104.Prior, R.L., Cao, G., Martin, A., Sofic, E., McEwen, J., O'Brien, C., Lischner, N., Ehlenfeldt,
M., Kalt, W., Krewer, G., Mainland, M., 1998. Antioxidant capacity as influencedby total phenolic and anthocyanin content, maturity and variety of Vaccinium species.J. Agric. Food Chem. 46, 2586–2593.
Reiter, L.T., Potocki, L., Chien, S., Gribskov, M., Bier, E., 2001. A systematic analysis ofhuman disease-associated gene sequences in Drosophila melanogaster. GenomeRes. 11, 1114–1125.
Schwarze, S.R., Weindruch, R., Aiken, J.M., 1998. Oxidative stress and aging reduce COXI RNA and cytochrome oxidase activity in Drosophila. Free Radic. Biol. Med. 25,740–747.
Sohal, R.S., Toroser, D., Brégère, C., Mockett, R.J., Orr, W.C., 2008. Age-related decreasein expression of mitochondrial DNA encoded subunits of cytochrome c oxidase inDrosophila melanogaster. Mech. Aging Dev. 129, 558–561.
Tonoki, A., Kuranaga, E., Tomioka, T., Hamazaki, J., Murata, S., Tanaka, K., Miura, M.,2009. Genetic evidence linking age-dependent attenuation of the 26S proteasomewith the aging process. Mol. Cell. Biol. 29, 1095–1106.
Willis, L.M., Shukitt-Hale, B., Joseph, J.A., 2009. Modulation of cognition and behavior inaged animals: role for antioxidant- and essential fatty acid-rich plant foods. Am. J.Clin. Nutr. 89, 1602S–1606S.
Wilson, M.A., Shukitt-Hale, B., Kalt, W., Ingram, D.K., Joseph, J.A., Wolkow, C.A., 2006.Blueberry polyphenols increase lifespan and thermotolerance in Caenorhabditiselegans. Aging Cell 5, 59–68.


![Blueberry Hill - Kiama Blowhole Buskers€¦ · Blueberry Hill [C] /// /// [F] /// /// [G7] stop [Tacet] I found my [F]thrill ..... on Blueberry [C]Hill ..... On Blueberry [G] Hill](https://img.pdfslide.us/doc/110x75/5f334258c6a82b4839519c93/blueberry-hill-kiama-blowhole-buskers-blueberry-hill-c-f-g7.jpg)


























