Embed Size (px)
Citation preview

Journal of Ecology
2001
89
, 1–13
© 2001 British Ecological Society
1
Blackwell Science, Ltd
Crown development in tropical rain forest trees: patterns with tree height and light availability
FRANK J. STERCK and FRANS BONGERS
Silviculture and Forest Ecology Group, Department of Environmental Sciences, Wageningen University, PO Box 342, 6700 AH Wageningen, The Netherlands, tel.: 31 317478008, fax: 31 317478078
Summary
1
Monitoring of two canopy species
Dicorynia guianensis
and
Vouacapoua americana
(Caesalpiniaceae) in a tropical rain forest in French Guiana was used to investigatevegetative crown development at five organizational levels: leaf, metamer, extensionunit, sympodial unit and whole crown. The effects of light availability and tree height ondifferent traits were evaluated in trees < 25 m in height and compared with tallerindividuals (25–37 m). Path-analysis is used to illustrate the consequences of traitchanges at multiple levels of organization for the whole crown level.
2
Tree height and canopy openness influenced crown development at each organizationallevel. Crowns in higher light levels had lower specific leaf area, greater leaf spacing,greater extension of all branches, and greater extension of the leader shoot. Withincreasing tree height, crowns had a lower specific leaf area, greater leaf area index andgreater relative crown depth.
3
Vouacapoua
showed some responses to light not seen in
Dicorynia
. In particular,
Vouacapoua
increased meristem activity with light, but the lack of response in
Dicorynia
may be due to moderate light levels rather than inability to respond.
4
Low leaf-display costs at low light availability may enable trees to survive light suppression.
5
Light availability cannot explain trait changes with tree height. Alternative explana-tions for trait changes with tree height are discussed.
6
Several of the relationships between plant traits and tree height or canopy opennessbecame non-linear when taller trees (25–37 m) were included. In these taller trees,vegetative growth was reduced at all organizational levels, particularly in
Vouacapoua
,which does not grow as tall as
Dicorynia
.
7
Qualitatively, plant responses to light did not differ between trees of different height,and were similar to seedling and sapling data in the literature. Responses were, however,quantitatively different, suggesting that small saplings cannot serve as model organismsfor crown development in taller trees.
Key-words
: canopy, crown structure,
Dicorynia guianensis
, French Guiana, ontogeny,rain forest trees, tree architecture,
Vouacapoua americana
.
Journal of Ecology
(2001)
89
, 1–13
Introduction
Trees growing through a tropical rain forest understoreyencounter different light environments (e.g. Chazdon& Fetcher 1984; Terborgh 1985; Clark
et al
. 1996).Generally, they will occupy sites with more light as theyget taller (e.g. Yoda 1974; Clark & Clark 1992; Bongers
& Sterck 1998), until they reach the canopy and nolonger compete with neighbours for light. If pioneertrees are to survive, they must expand their crownsquickly, shading their neighbours, and grow to maturityin the high light conditions associated with canopy gaps(e.g. Alvarez-Buylla & Martinez-Ramos 1992; Ackerly1996). Slower growing, shade-tolerant trees cannotmonopolize canopy gaps and need to survive periodsof light suppression, expanding during periods whensufficient light is available, until they reach maturity(e.g. Hartshorn 1978). Vegetative crown developmentplays an important role in the shade-tolerant strategy
Correspondence: Frank J. Sterck, Utrecht University, SectionPlant Ecology, Postbox 80084, 3508 TB Utrecht, The Nether-lands (e-mail [email protected]).
JEC525.fm Page 1 Monday, March 12, 2001 4:08 PM

2
F. J. Sterck & F. Bongers
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1–13
(e.g. Canham 1988; Küppers 1989; Kohyama & Hotta1990; King 1994; Aiba & Kohyama 1997).
Our approach to the process in structurally complexrain forest trees defines vegetative crown developmentas the production, positioning and morphologicalproperties of the various components listed in Table 1(see also White 1979; Barthélémy 1991; Bell 1991; Room
et al
. 1994). These plant components characterize thedifferent levels of organization in the crown hierarchy,and are produced and spaced according to an inheritedpattern (Hallé & Oldeman 1970; Hallé
et al
. 1978). Thishierarchy remains the same as trees get taller or respondto environmental conditions, and can be used to integratedevelopment at different organizational levels and tocompare crown development among trees in differentlight environments and in different ontogenetic phases.
Components at each of the various organizationallevels are well known to respond to light, i.e. they arelocated at different positions and produced at differentrates (King 1994), and have different sizes or archi-tectures (Fisher 1986; Shukla & Ramakrishnan 1986).Theoretical predictions that crown development atlow light availability will favour survival if it is asso-ciated with slow growth rates (Veneklaas & Poorter1998), if bifurcation ratios are low and crowns areflatter (Leopold 1971), if there is reduced self-shadingof the leaves (Horn 1971) and if there is higher allocationof mass to leaves relative to wood (Givnish 1988) aregenerally supported by empirical studies of seedlingsand small saplings (e.g. Canham 1988; King 1994).
The influence of ontogeny on vegetative developmentis less well known and, although it has been describedfor some rain forest trees (Hallé
et al
. 1978; Edelin1990), information on the effects on bifurcation ratios,crown allometry, leaf self-shading and leaf allocationremains anecdotal and ambiguous. Trees often becomemore exposed as they grow taller and crown develop-ment may therefore change with tree height, just asit does with increasing light availability. This wouldparallel physiological leaf traits where assimilation and
respiration rates are reported to increase as either treeheight or light increased (Kitajima 1996; Rijkers
et al
.2000). Yet some trees become less exposed with increas-ing tree height (e.g. Clark & Clark 1992). Moreover,trees become increasingly woody and have to be main-tained by relatively low light interception (or leaf) areas.We would therefore expect any change in vegetativecrown development with tree height to parallel thatwith declining light levels. Most of the available datarelates to the pioneer species that monopolize light gaps(e.g. Alvarez-Buylla & Martinez-Ramos 1992; Ackerly1996), rather than the shade-tolerant trees that growin variable light conditions (Strauss-Debenedetti &Bazzaz 1996).
Ontogenetic stage and light environment are likely toinfluence crown development simultaneously. Althoughthe effects of both tree size and light level have beenconsidered for leaf morphology (Niinemets & Kull1995; Niinemets 1997; Rijkers
et al
. 2000), such jointinvestigations on crown development are rare andusually focus on smaller individuals and few organiza-tional levels (e.g. Canham 1988). The combined effectsof development at the various organizational levels forthe whole crown are also poorly understood.
We explicitly distinguished between the effects oftree height and light availability on each of the planttraits in Table 2 (i.e. at the various organizationallevels) to evaluate their influence on crown growth, leafdisplay and leaf display costs. We studied individuals oftwo shade-tolerant species in French Guiana, rangingin size from small saplings to canopy trees, to test twohypotheses: (i) plant traits will vary with light avail-ability such that trees will have slower crown growth,wider crowns, less self-shading, smaller leaf displaycost, smaller leaf spacing and larger specific leaf area atlower light levels; and (ii) because an increase in treeheight is accompanied by a gradual increase in lightavailability, trait changes with increasing tree height(up to 25 m) will parallel those seen under conditionsof increasing light.
Table 1 Plant components in order of increasing size (partly adapted from Bell 1991; Room et al. 1994) and representing thevarious organizational levels in the crown
Name Definition
Meristem Region of active cell division and enlargement. There is typically a meristematic zone at the apex (apical meristem) and each node (axillary meristem) along every axis. Axillary meristems become apical meristems once they become active and produce a new shoot.
Leaf Photosynthetic structure.
Metamer An internode with axillary bud(s) at its proximal end and one or more leaves at its distal end, but without shoots resulting from growth of axillary buds.
Extension unit The vegetative axis, displayed by one period of activity of a meristem (‘flush’), either of apical or axillary origin. It consists of a sequence of metamers.
Sympodial unit The total sequence of metamers and/or extension units produced by one meristem (also referred to as module, Prévost 1967; Hallé et al. 1978).
Crown The assemblage of leaf-supporting branches, including portions of the stem that support leaves or leaf-supporting branches.
JEC525.fm Page 2 Monday, March 12, 2001 4:08 PM

3
Crown development in tropical trees
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1–13
Materials and methods
Field work was carried out in a pristine lowlandtropical rain forest at the biological field station ‘lesNouragues’ (4
°
05
′
N, 52
°
40
′
W), French Guiana. Annualrainfall is approximately 3000 mm, with dry seasons(< 100 mm month
–1
) in September and October, andsometimes in March. The forest is dominated by speciesof Lecythidaceae, Leguminosae, Sapotaceae, Chryso-balanaceae and Burseraceae (Sabatier & Prévost 1989).The area is covered with well-drained, clayey to sandy-clayey soils on weathered granite parent material.
Dicorynia guianensis
Amshoff. and
Vouacapouaamericana
Aubl., hereafter referred to by their genericnames alone, are abundant in the canopy. SuchCaesalpiniaceae or other leguminous species are(co-)dominant in many neo-tropical forests and mostare characterized as shade-tolerant (e.g. Schulz 1960)and as conforming either to Troll’s model of canopyarchitecture or to other mixed plagiotropic models(Oldeman 1974; Hallé
et al
. 1978; Oldeman 1989).Thus, the stem leans over to support a horizontal sprayof distichously arranged leaves and the process isrepeated at different positions in the crown as a result ofrelays sprouting from the bends. Stem and branches can-not be distinguished from one another at early stagesof crown development, but one branch subsequentlystraightens towards the vertical and continues secondarythickening growth and thus becomes the stem, whilethe other branches fall or develop in a more horizontaldirection. Both study species are known for their com-mercial timber value in French Guiana and Surinam.
Figure 1 shows how different components (leaves,metamers, extension units and sympodial units, Table 1)contribute to crown development in the study species.For
Vouacapoua
, metamers were not considered becausethey are structurally variable: those at the bottom of anextension unit support scale leaves, and the ones at thetop support compound leaves. For
Dicorynia
, however,metamers are structurally equivalent, with each producingone compound leaf, and therefore extension unitscannot always be unambiguously distinguished (butsee Drénou 1994).
Populations of both species were inventoried inOctober 1992 as part of a long-term study of tree growth(Sterck 1997). A 12-ha plot was mapped for trees with astem diameter
≥
10 cm at 1.30 m above ground level (d.b.h.),and a central 1.5-ha plot was mapped for trees with a stemdiameter < 10 cm, but with a height greater than 0.50 m.
For each species, we selected 20 of the inventoriedindividuals that were less than 4 m tall, ensuring thatthe full range of light levels within the site was repres-ented. Their crown development was measured fromthe ground, either by bending the stem over or usingnatural elevations within the forest. The crowns of tallerindividuals could only be studied from neighbouringtrees (climbed using spikes or alpinist ropes). Becauseonly a few of the inventoried individuals were accessiblein this way, we included a selection of the accessible4–25 m trees (13
Dicorynia
and 20
Vouacapoua
) and allfive accessible > 25 m trees (
Vouacapoua
, 25, 35 and37 m, and
Dicorynia
, 26 and 37 m) from the adjacentarea outside the plots. Measurements were made in
Table 2 Plant traits, ordered by organizational level. Metamer is equivalent to extension unit in Vouacapoua
Crown levelRelative crown depth = Crown depth/mean crown width (m m–1)Total branch extension = Total branch length produced per year (m year–1)Leader extension = Total leader length produced per year (m year–1)Crown area = Ground projection of the crown area (m2)Total leaf area = Mean leaf area × total number of leaves (m2)Leaf area index = Total leaf area/crown area (m2 m–2)Number of meristems = Number of apical meristems
Sympodial unit levelMetamer production = Number of metamers produced per sympodial unit per year (year–1)Leader metamer production = Number of leader metamers produced per year (year–1)Sympodial unit survival = Percentage of sympodial units surviving per year (% year–1)Sympodial unit production = Percentage of sympodial units that is produced in preceding year (% year–1)
Metamer levelMetamer length = Mean length of metamer (cm)Leader metamer length = Mean length of leader metamer (cm)Leaf production = Number of leaves produced per metamer per year (year–1)Leaf fall = Number of fallen leaves per metamer per year (year–1)Leaf spacing = Mean length between leaves (cm)Relative leaf spacing = Leaf spacing/√leaf area (cm cm–1)
Leaf levelLeaf area = Mean surface area of individual leaf (cm2)Specific leaf area = Leaf area/leaf mass (cm2 g–1)
JEC525.fm Page 3 Monday, March 12, 2001 4:08 PM

4
F. J. Sterck & F. Bongers
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1–13
November 1992, November 1993, and November 1994for all individuals.
The light environment was recorded using hemi-spherical photographs, taken in November 1993. ACanon Ti 70 body and Canon 7.5 mm/5.6 lens wasmounted in a leveller and telescopic poles (up to 6 m long)were used to position the camera just above the top ofeach individual. The light environment of taller individualswas also measured by climbing neighbouring trees withropes or spikes. Black-and-white prints were scannedusing a Hewlett Packard Desk Scan II and canopy open-ness (the percentage of open sky) was calculated usingthe programme Winphot 5 (Ter Steege 1996). Canopyopenness is used as an estimate of light availability.
Trees were marked and drawn to scale in 1992. Allbranches and apical meristems of individuals < 4 m weretagged and coded, as was a selection of branches plustheir apical meristems (
n
= 10–45) within the top of thecrown of taller individuals. In later censuses (1993 and1994), we measured size, structure and production rateof each plant component (for details see below).
Production was assessed by counting leaves, leaf scars,metamers (
Dicorynia
), extension units (
Vouacapoua
)and apical meristems (or active sympodial units) thathad been produced by 1994 on top of apical meristems
tagged in 1992. For
Vouacapoua
, the number of leavesand scars included photosynthetic but not scale leaves.Although for trees taller than 4 m detailed counts weremade only for the selected apical meristems, weassessed the total number of leaves and apical meristemsby climbing carefully along the length of these individuals.In each individual, the axis (linear sequence of metamersor extension units) that supported the uppermost apicalmeristem of the crown in 1994 was defined as the leader.
Leaf area was determined (Delta Image AnalysisSystem, Eijkelkamp 1991) as the mean value for 20 leavestaken from random positions in the crown. Lengthswere measured for all metamers (
Dicorynia
) or exten-sion units (
Vouacapoua
) produced distal to monitoredmeristems. Mean metamer length and extension unitlength were calculated for both the whole crown andthe leader separately. Lengths of sympodial units arenot reported because they may have continued lengthgrowth after the last census in November 1994. We alsomeasured the greatest crown width, the crown widthperpendicular to this, and crown depth, i.e. the verticaldistance between the uppermost and the bottom leaves.
Plant traits were categorized into various organ-izational levels (as detailed in Table 2). At crown levelwe used the measurements to calculate relative crown
Fig. 1 Plant component architecture of Dicorynia guianensis and Vouacapoua americana, two shade-tolerant canopy tree species oftropical rain forest in French Guiana. For definitions of plant components see Table 1. Figure adapted from Sterck (1999).
JEC525.fm Page 4 Monday, March 12, 2001 4:08 PM

5
Crown development in tropical trees
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1–13
depth, total branch extension, leader extension, crownarea, total leaf area, leaf area index and number of mer-istems. At sympodial unit level we calculated metamer(or extension unit) production, leader metamer produc-tion, sympodial unit survival and sympodial unit pro-duction. At metamer (or extension unit, in the case of
Vouacapoua
) level we calculated metamer length, leadermetamer length, leaf production, leaf fall, leaf spacing (ameasure for the average distance between leaves) andrelative leaf spacing (a measure for the leaf spacingper unit area). At leaf level we calculated leaf area andspecific leaf area.
Multiple linear regression was used to assess the rel-ative importance of tree height and canopy openness indetermining plant traits. Trees taller than 25 m wereexcluded because their inclusion caused relationshipsbetween plant traits and tree height to become non-linear in 11 out of 36 cases. In some cases, plant traitvalues were transformed if this improved the linearityamong variables and/or the homogeneity of variances.The following regression model was used:
Plant trait = c + (
b
1
×
height) + (
b
2
×
openness) + (
b
3
×
height
×
openness) +
e
eqn 1
Here, c is the constant (intercept), the
b
1
,
b
2
,
b
3
are theregression coefficients and e is the error term.
We used path-analysis (e.g. Wright 1934; Kingsolver& Schemske 1991) to integrate the effects of tree heightand canopy openness on plant traits and to show theconsequences of these plant traits for the whole crown(Fig. 2). Path-analysis shows direct (or causal) effectsas path-coefficients, and relations without causal effectas Pearson product moment correlation coefficients.The analysis distinguishes direct, indirect and total effects(e.g. Sokal & Rohlf 1981). In general, path-analysisindicates the change in a criterion variable (expressedin SD units) as a result of a change of 1 SD in each ofits predictors, both direct and indirect. Correlationamong predictors (collinearity) inflates both the path-coefficients and their confidence intervals. Consequently,the effect of a predictor on a criterion variable is eitheroverestimated with respect to its magnitude, or neglected
because it is harder to get a coefficient that differssignificantly from 0 (Petraitis
et al
. 1996). In our analysis,we had to deal with collinearity between tree heightand canopy openness, as well as among plant traits.We did not evaluate the magnitudes of significant effectssince the coefficient values cannot be accurately estimated.We realise that by only distinguishing between effects thatare significant and those that are not, we may erroneouslyhave rejected actual effects (increased type II errors).
Results
.
Canopy openness of individuals < 25 m tall rangedfrom 0.8% to 30% (Fig. 3). For the five taller individuals itranged from 12% to 80%. Tree height and canopyopenness were correlated (Pearson’s product momentcoefficient of 0.54 in
Dicorynia
and 0.74 in
Vouacapoua
,
P
< 0.001). Although canopy openness tended to behigher above taller trees, there was a wide range ofcanopy openness values for any particular tree height.In our multiple regression analyses and path-analyseswe concentrated on how crown development is affectedby canopy openness independent of tree height, and bytree height independent of canopy openness.
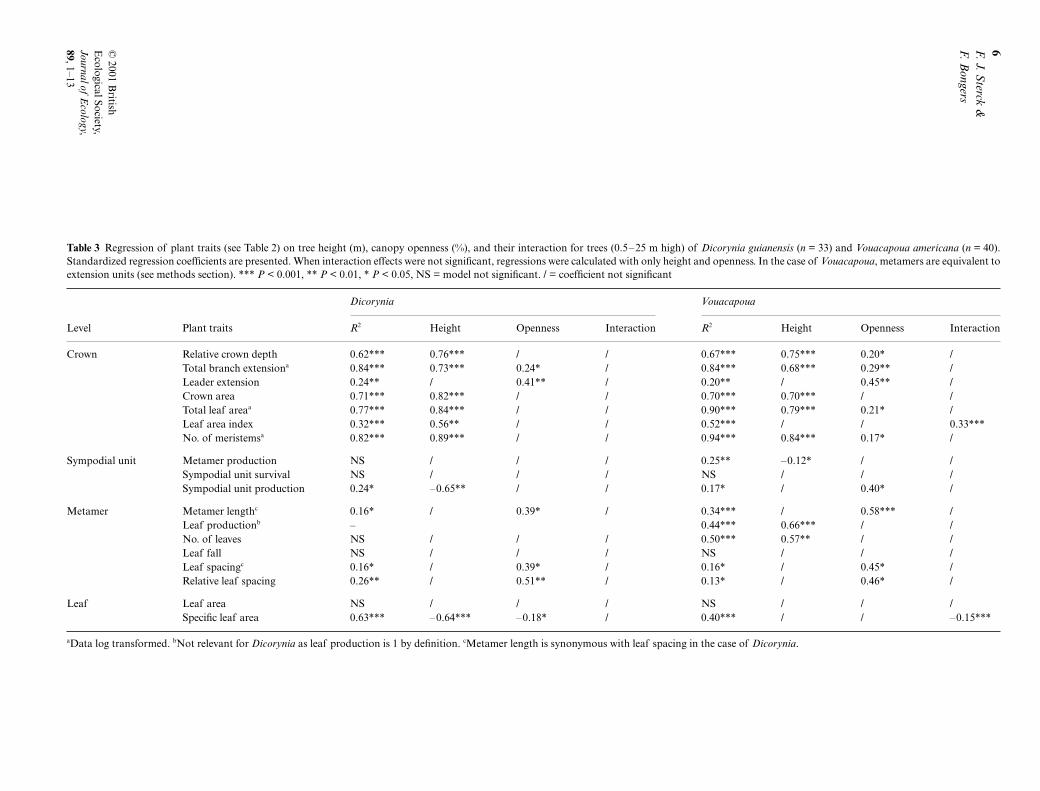
Multiple regression analysis was used to evaluate changesin plant traits with tree height and canopy openness(Table 3) for trees < 25 m high. For clarity, the plant traitchanges are described for each organizational level.
Crown level
: In both species, total branch extensionincreased with both tree height and canopy openness,and leader extension increased with canopy opennessindependent of tree height. All other traits in
Dicorynia
increased with height alone, whereas in the case of
Vouacapoua
they increased with both tree height andcanopy openness, except for crown area (height only)and leaf area index (interaction only).
Fig. 2 Model presenting the expected influences of treeheight and light availability on plant traits at the crown levelafter two successive steps. Initially, light availability and treeheight act on different plant traits, which then influence aplant trait at the specific (usually the whole crown) level.Single-headed arrows indicate causal relationships; double-headed arrows indicate correlations.
Fig. 3 Canopy openness vs. tree height for sampled trees. s,Dicorynia guianensis; d, Vouacapoua americana.
JEC525.fm Page 5 Monday, March 12, 2001 4:08 PM
6
F. J. Sterck &
F. B
ongers
© 2001 B
ritish E
cological Society,
Journal of Ecology
,
89
, 1–13
Table 3
Regression of plant traits (see Table 2) on tree height (m), canopy openness (%), and their interaction for trees (0.5–25 m high) of
Dicorynia
guianensis
(
n
= 33) and
Vouacapoua
americana
(
n
= 40).Standardized regression coefficients are presented. When interaction effects were not significant, regressions were calculated with only height and openness. In the case of
Vouacapoua
, metamers are equivalent toextension units (see methods section). ***
P
< 0.001, **
P
< 0.01, *
P
< 0.05, NS = model not significant. / = coefficient not significant
Dicorynia Vouacapoua
Level Plant traits
R
2
Height Openness Interaction
R
2
Height Openness Interaction
Crown Relative crown depth 0.62*** 0.76*** / / 0.67*** 0.75*** 0.20* /Total branch extension
a
0.84*** 0.73*** 0.24* / 0.84*** 0.68*** 0.29** /Leader extension 0.24** / 0.41** / 0.20** / 0.45** /Crown area 0.71*** 0.82*** / / 0.70*** 0.70*** / /Total leaf area
a
0.77*** 0.84*** / / 0.90*** 0.79*** 0.21* /Leaf area index 0.32*** 0.56** / / 0.52*** / / 0.33***No. of meristems
a
0.82*** 0.89*** / / 0.94*** 0.84*** 0.17* /
Sympodial unit Metamer production NS / / / 0.25** –0.12* / /Sympodial unit survival NS / / / NS / / /Sympodial unit production 0.24* –0.65** / / 0.17* / 0.40* /
Metamer Metamer length
c
0.16* / 0.39* / 0.34*** / 0.58*** /Leaf production
b
– 0.44*** 0.66*** / /No. of leaves NS / / / 0.50*** 0.57** / /Leaf fall NS / / / NS / / /Leaf spacing
c
0.16* / 0.39* / 0.16* / 0.45* /Relative leaf spacing 0.26** / 0.51** / 0.13* / 0.46* /
Leaf Leaf area NS / / / NS / / /Specific leaf area 0.63*** –0.64*** –0.18* / 0.40*** / / –0.15***
a
Data log transformed.
b
Not relevant for
Dicorynia
as leaf production is 1 by definition.
c
Metamer length is synonymous with leaf spacing in the case of
Dicorynia
.
JEC
525.fm P
age 6 Monday, M
arch 12, 2001 4:08 PM

7
Crown development in tropical trees
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1–13
Sympodial unit level
: Extension unit productiondecreased with tree height in
Vouacapoua
. Sympodialunit production decreased with tree height in Dicoryniaand increased with canopy openness in Vouacapoua.
Metamer level (only for Dicorynia): Metamer length(and thus also leaf spacing) and relative leaf spacing in-creased with canopy openness independent of tree height.
Extension unit level (only for Vouacapoua): Extensionunit length, leaf spacing and relative leaf spacingincreased with canopy openness independent of treeheight. Leaf production and the number of leavesincreased with tree height.
Leaf level: In Dicorynia, specific leaf area decreasedwith tree height and canopy openness. In Vouacapoua,the decrease in specific leaf area required increases withboth tree height and canopy openness, as illustrated bythe highly significant interaction term.
The path-analysis shows how tree height and canopyopenness influence plant traits at a higher level throughtheir effects on plant traits at lower organizationallevels (Figs 4–7). Path-analysis does not account forinteractions between tree height and canopy openness,nor for non-linear relationships, and is therefore restrictedto trees shorter than 25 m.
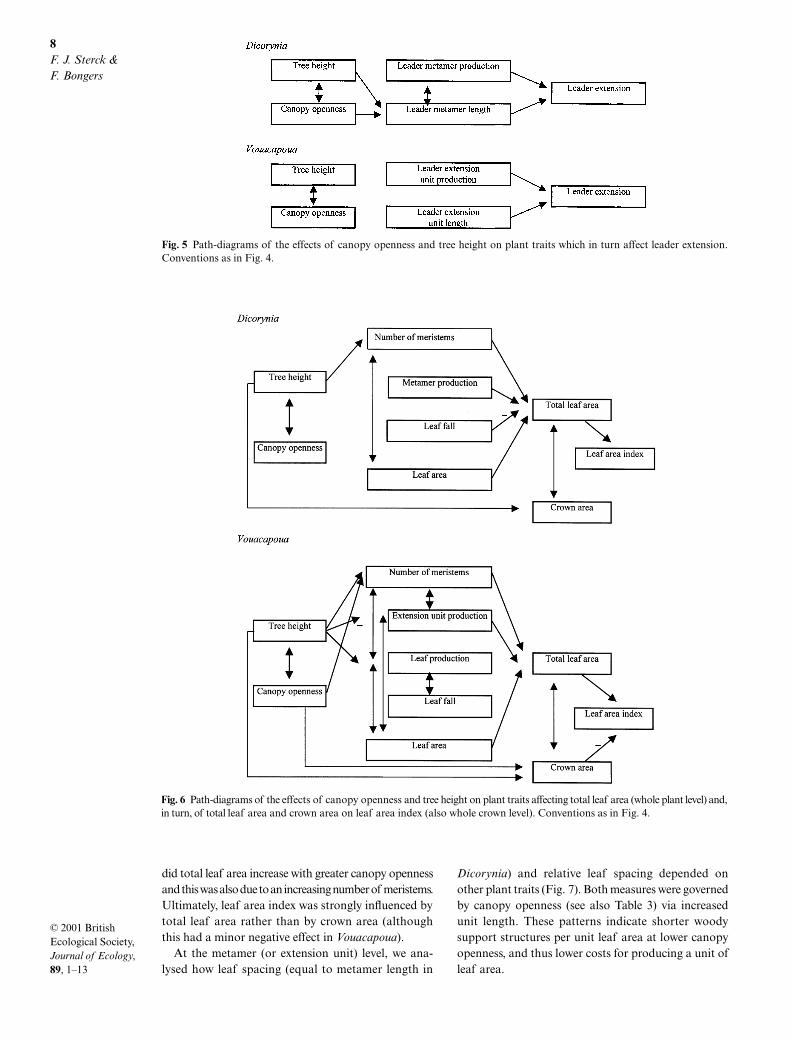
Total branch extension depends on the number ofmeristems, metamer (or extension unit) productionper meristem, and metamer (or extension unit) length(Fig. 4). In both species tree height contributed to theincrease in total branch extension because of the greater
number of meristems. In Vouacapoua, extension unitproduction was also affected (in this case negatively) bytree height, but the effect on total branch extension wasminor, compared to the strong positive effect of thenumber of meristems (values of effects not shown).Canopy openness increased total branch extension inVouacapoua due to both increased numbers of meristemsand greater extension unit length. In turn, the greaternumber of meristems resulted from a greater sympo-dial unit production (Table 3). In Dicorynia, however,the increase in total branch extension with canopyopenness resulted only from the positive effect onmetamer length. Thus, only one organizational levelcontributed to the more rapid crown growth of Dicoryniawith both greater tree height and with greater canopyopenness, whereas two levels were involved in Vouacapoua.
Leader extension depends on leader metamer (orextension unit) production, and on the length of theseplant components (Fig. 5). The increase in leaderextension in Dicorynia with greater canopy opennesswas mainly due to increases in metamer length. Vouaca-poua showed the same tendency, although the effectof canopy openness on leader extension unit lengthwas not significant (P = 0.07).
Total leaf area depends on the number of meristems,metamer (or extension unit) production per meristem,leaf production and leaf fall per metamer (or extensionunit), and leaf area (Fig. 6). The increase in totalleaf area with tree height resulted primarily from an in-creasing number of meristems. Although extension unitproduction in Vouacapoua decreased with tree height,its effect was minor compared to the strong positiveeffect of the number of meristems. Only in Vouacapoua
Fig. 4 Path-diagrams presenting the effects of tree height and canopy openness on total branch extension for the canopy tree speciesDicorynia guianensis and Vouacapoua americana in a tropical rain forest in French Guiana. The effects are split into the effects ofcanopy openness and tree height on plant traits at various organizational levels and the subsequent effects of these traits on totalbranch extension (whole crown level). Significant (P < 0.05) correlations and direct effects are indicated by double- and single-headedarrows, respectively. Relationships were usually positive; instances of negative relationship are shown by a negative (–) sign.
JEC525.fm Page 7 Monday, March 12, 2001 4:08 PM
8F. J. Sterck & F. Bongers
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
did total leaf area increase with greater canopy opennessand this was also due to an increasing number of meristems.Ultimately, leaf area index was strongly influenced bytotal leaf area rather than by crown area (althoughthis had a minor negative effect in Vouacapoua).
At the metamer (or extension unit) level, we ana-lysed how leaf spacing (equal to metamer length in
Dicorynia) and relative leaf spacing depended onother plant traits (Fig. 7). Both measures were governedby canopy openness (see also Table 3) via increasedunit length. These patterns indicate shorter woodysupport structures per unit leaf area at lower canopyopenness, and thus lower costs for producing a unit ofleaf area.
Fig. 5 Path-diagrams of the effects of canopy openness and tree height on plant traits which in turn affect leader extension.Conventions as in Fig. 4.
Fig. 6 Path-diagrams of the effects of canopy openness and tree height on plant traits affecting total leaf area (whole plant level) and,in turn, of total leaf area and crown area on leaf area index (also whole crown level). Conventions as in Fig. 4.
JEC525.fm Page 8 Monday, March 12, 2001 4:08 PM
9Crown development in tropical trees
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
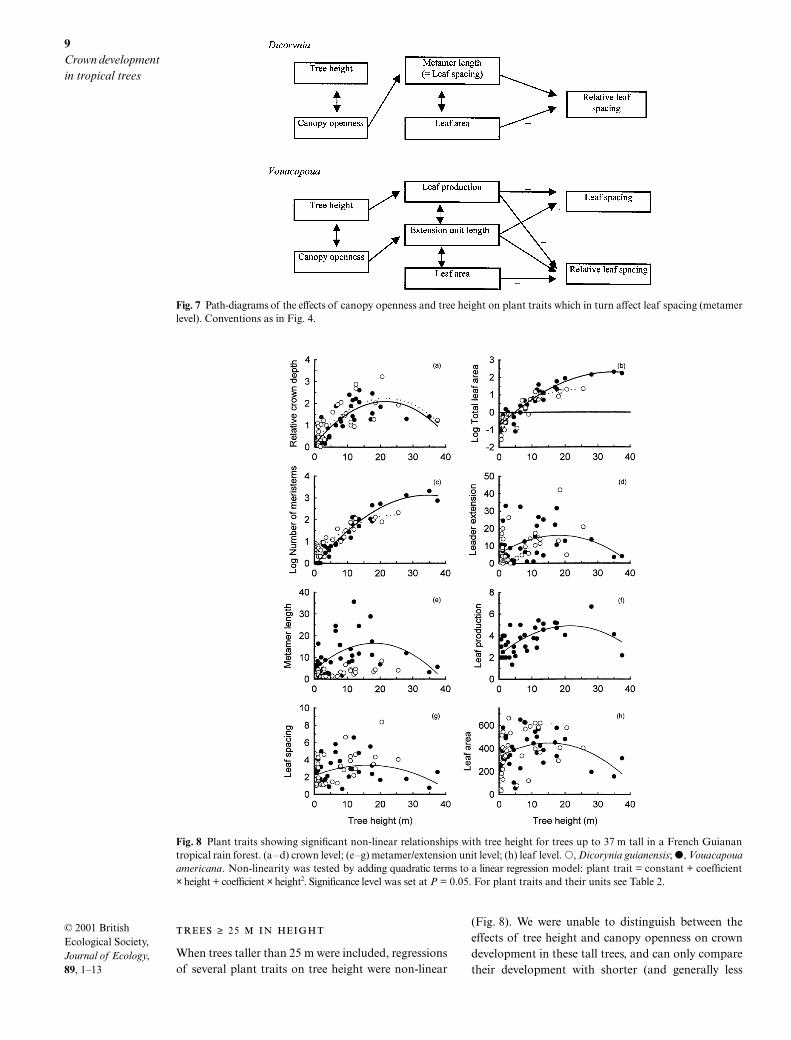
≥ 25
When trees taller than 25 m were included, regressionsof several plant traits on tree height were non-linear
(Fig. 8). We were unable to distinguish between theeffects of tree height and canopy openness on crowndevelopment in these tall trees, and can only comparetheir development with shorter (and generally less
Fig. 7 Path-diagrams of the effects of canopy openness and tree height on plant traits which in turn affect leaf spacing (metamerlevel). Conventions as in Fig. 4.
Fig. 8 Plant traits showing significant non-linear relationships with tree height for trees up to 37 m tall in a French Guianantropical rain forest. (a–d) crown level; (e–g) metamer/extension unit level; (h) leaf level. s, Dicorynia guianensis; d, Vouacapouaamericana. Non-linearity was tested by adding quadratic terms to a linear regression model: plant trait = constant + coefficient× height + coefficient × height2. Significance level was set at P = 0.05. For plant traits and their units see Table 2.
JEC525.fm Page 9 Monday, March 12, 2001 4:08 PM

10F. J. Sterck & F. Bongers
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
exposed) trees. At the crown level, non-linear relation-ships became apparent generally for both speciesbetween tree height and relative crown depth, totalleaf area, number of meristems, and leader extension(Fig. 8a–d). There were no non-linear relationships atthe sympodial unit level, neither were there for Dicory-nia at the metamer level, whereas extension unit length,leaf production and leaf spacing in Vouacapouashowed maximal values around 25 m (Fig. 8e–g). Atthe leaf level, leaf area decreased above 25 m, particu-larly in Vouacapoua (Fig. 8h).
Discussion
Total crown growth and leader growth (which is corre-lated with height growth) increased with canopy open-ness, independently of height in the latter. Only one ortwo organizational levels contributed to these crowngrowth responses, in contrast with another study ofthese species (Sterck 1999) where both measures ofcrown growth of juveniles (4–18 m) were higher in gapsthan in the understorey because of changes at all pos-sible levels. It may be that some traits respond only tothe high light levels characteristic of gaps and these arerarely experienced by trees shorter than 25 m. Alterna-tively, gradual changes may become significant onlywhen well-separated light levels are compared.
The influence of light availability on leader extensionhas been shown for seedlings (e.g. Popma & Bongers1988), saplings (e.g. King 1991) and taller juveniles(Sterck 1999). The increase in height growth with sizein eight Costa Rican tree species (Clark & Clark 1992)might also be due to light availability rather than sizeitself. Leader growth therefore generally seems tobe controlled by light in trees up to at least 25 m,whereas total branch extension (which has rarely beenstudied explicitly) is more affected by tree height(Table 3).
Total leaf area and leaf area index increased with treeheight in Dicorynia. In Vouacapoua, total leaf areaincreased with both tree height and canopy openness,but leaf area index was affected only by their interaction.An increasing number of meristems explained most ofeach effect.
Small individuals (< 10 m) of the pioneer tree Cecropiaobtusifolia (Alvarez-Buylla & Martinez-Ramos 1992)show a similar increase in total leaf area with heightbut as a result of a more than 100-fold increase in leafarea, rather than meristem activation. Dicorynia andVouacapoua typically have c. 20 apical meristems bythe time they reach 10 m in height, and larger individ-uals continued to increase total leaf area by producingmore apical meristems, as is the case with Cecropiaindividuals > 10 m in height. Similar changes may
therefore be the result of different plant traits in eachspecies.
The early branching of Dicorynia and Vouacapouafollows Troll’s model, while Cecropia persists for longeras a single pole axis and its subsequent branching pat-tern is described by Rauh’s model (Alvarez-Buylla &Martinez-Ramos 1992). Below 10 m, Cecropia (still inpole phase) may have a higher leaf area index than thebranched Dicorynia and Vouacapoua, but self-shadingis reduced by the positioning (Ashton 1978; Orians1982) and by wider spacing of its leaves (King 1994).
The higher sympodial unit production in Vouacapouawith greater canopy openness indicates both an increasedbranching rate and a higher bifurcation ratio (an indexof the degree of branching from one branching order tothe next) with greater canopy openness (Canham 1988)and was associated with narrower crowns. Leopold(1971) and Whitney (1976) argued that such high bifurca-tion ratios are more efficient (in terms of stem tissueused for leaf display) for narrower crowns in higherlight environments, while low bifurcation ratios aremore efficient for planar crowns (which have less leafself-shading) at lower light levels. Similar increases inbifurcation ratios at higher light availability have alsobeen found in other studies (Steingraeber et al. 1979;Shukla & Ramakrishnan 1986).
Although juveniles of both species had larger total leafarea, higher leaf area index, and longer and relativelynarrower crowns in gaps than in the darker understorey(Sterck 1999), the lack of such effects in Dicorynia inthe present study may (as with crown growth) reflectthe moderate light levels. Similar differences in resultswere found between studies of small saplings of thetemperate species Acer saccharum, which showed aresponse only when fully exposed sites (highest lightlevels) were included (Steingraeber et al. 1979; Canham1988; Bonser & Aarssen 1994).
Leaf display costs can be defined as the amount ofcarbon invested to produce one unit of leaf area, anddepend on the relative amounts, densities and con-struction costs of leaf and wood material. Such costsmust be reduced at low light levels if light interceptionis to increase, but only the size components show sig-nificant plasticity (e.g. King 1991, 1994). Leaf spacing,relative leaf spacing and specific leaf area reflect the(relative) amounts of wood and leaves along newshoots, although the impacts of secondary thickeningand root growth are not considered.
Specific leaf area (a leaf area trait) is negativelyrelated to both tree height and light availability inDicorynia, but only to their interaction in Vouacapoua,similar to patterns observed for other species (Jurik1986; Oberbauer & Strain 1986; Poorter et al. 1995;Rijkers et al. 2000). Everything else being equal, a higherspecific leaf area at lower light levels and/or tree heightwill result in a greater light interception area per leaf
JEC525.fm Page 10 Monday, March 12, 2001 4:08 PM

11Crown development in tropical trees
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
mass, and thus contribute to reduced leaf displaycosts. However, morphological information alone maybe misleading because many other leaf-level factors,including content of expensive metabolites such aschlorophyll, may vary (Boardman 1977; Pearcy 1987;Ellsworth & Reich 1993) and contribute to reducingcosts in higher, more open canopies.
Leaf area was unaffected by either factor, but leafspacing and relative leaf spacing both decreased whencanopy openness is less; this decrease was due to shortermetamers (Dicorynia) or extension units (Vouacapoua).Leaf spacing also decreased at lower light in othertropical trees (King 1991, 1993, 1994), but there is as faras we know no comparable data on relative leaf spacing.All three traits considered appear to contribute to areduction in the leaf display costs at lower light availab-ility, which may enable trees to lower their whole-plantcompensation point and thus survive light suppression.
Regressions of plant traits against tree height showedlinear relationships in trees under 25 m in height, whileseveral regressions became non-linear if larger treeswere also included. This apparent switch in crowndevelopment might be triggered at a certain height,greater exposure or some other environmental factor.On attaining the canopy, branching patterns may changedrastically (King & Maindonald 1999) and even mul-tiple tree-like structures (trunk-like branches, each withits own subcrown) may develop in a process referred toas reiteration or metamorphosis (Hallé et al. 1978;Edelin 1990); however, no data are available on associ-ated quantitative changes in crown development traits.
Many crown-level traits in both species lost theirlinear increase with tree heights above 25 m. Thechanges in relative crown depth and leader extensionsuggest that crowns which have been narrowing withincreasing height then widen above 25 m, as seen fortaller trees in other tropical forests (Bongers & Sterck1998; Bongers et al. 1988; King 1996). This is probablydue to the higher light levels in the upper open canopybecause the change occurs at the height where lightlevels increased sharply (Koop & Sterck 1994) andjuvenile plantation-grown trees of Dicorynia andVouacapoua had wider crowns than forest-grown treesof the same height (F.J. Sterck, unpublished data). Inthe open upper canopy, where there is little chance ofbeing over-topped, increasing crown width may enabletrees to shade and thus out-compete their smallerneighbours.
In Vouacapoua, changes with tree height and canopyopenness also became non-linear above 25 m for sometraits at the extension unit level, indicating a reductionin vegetative crown growth; this observation is expectedbecause maximum height is about 35 m for this species.No such effects were seen in Dicorynia where the largesttrees (25 and 37 m) were still below the 45–50 m heightof full-grown adults.
At the leaf level, leaf area decreased above 25 mfor Vouacapoua and this may reflect adaptations tophysiological drought in the open upper canopy, butthere was no effect in Dicorynia. In general, adults ofcanopy tree species have smaller leaves than juvenilesof the same species (Thomas & Ickes, 1995).
General conclusions
Traits generally responded to light in the same ways aspreviously reported for smaller saplings. Returning toour first hypothesis, under conditions of decreasinglight availability, trees were (as expected) found to becharacterized by slower crown growth rates, relativelywide crowns, fewer leaf layers, smaller leaf display costs,lower sympodial unit production, smaller leaf spacingand larger specific leaf areas. The absence of someresponses in Dicorynia probably reflected the generalabsence of light levels able to induce an effect, ratherthan an inherent inability to respond.
Our second hypothesis, that increasing tree heightand increasing light availability influence tree charac-teristics in a similar way, was supported by parallelresponses in some traits at the crown and the leaf level.However, even in these cases changes with height maynot be due to the higher light levels: at the leaf level,effects on specific leaf area are more likely to reflectphysiological drought, and effects at the whole crownlevel were due to the influence of height on the number ofapices. At the three intermediate levels (metamer, exten-sion unit and sympodial unit), the changes in plant traitswith tree height never paralleled the patterns with lightavailability. The decrease in the proportion of product-ive (leaves) to unproductive (wood) biomass with increas-ing tree height may counterbalance the positive effectsof increasing light availability. Overall, light availabilitycannot be said to have been an important selection forceacting on architectural changes with tree height.
Our approach also suggests that small saplingscannot serve as model organisms for the study of crowndevelopment in trees of other sizes. While the directionsof responses of saplings and larger trees to light aresimilar, no quantitative predictions can be made. Aswell as changing with tree height per se, many traitresponses to light show changes in magnitude, or evenin direction (for trees above 25 m in height), as the treegrows. Furthermore, responses to light at the crownlevel are predominantly driven at intermediateorganizational levels (sympodial unit, extension unitand metamer), which rarely receive attention in smallsaplings. The modelling of crown development in treestherefore requires data from trees of a range of sizes.In addition, even closely related species such as thosestudied here (same plant family and similar generalbranching pattern) may differ in responses of sometraits. This may have far reaching consequences formodelling forest canopy dynamics on the basis ofcrown development of individual trees, particularly inspecies-rich habitats such as tropical rain forests.
JEC525.fm Page 11 Monday, March 12, 2001 4:08 PM

12F. J. Sterck & F. Bongers
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
Acknowledgements
We thank L. Poorter, T. Rijkers and M. Smeenge forgiving field support; D. Ackerly, C.D. Canham, P.A.Jansen, D. King, E. Leigh Jr., J. den Ouden, L. Poorter,H.H.T. Prins, S. Wijdeven and J. Wright for giving com-ments on earlier drafts of the manuscript; M.G. Bornfor drawing Fig. 1. This study was supported by grantW 85-239 of the Netherlands Foundation for theAdvancement of Tropical Research (WOTRO).
References
Ackerly, D. (1996) Canopy structure and dynamics: integra-tion of growth processes in tropical pioneer trees. TropicalForest Plant Ecophysiology (eds S.S. Mulkey, R.L. Chazdon& A.P. Smith), pp. 619–658. Chapman & Hall, New York.
Aiba, S. & Kohyama, T. (1997) Crown architecture andlife-history traits of 14 tree species in a warm-temperaterain forest: significance of spatial heterogeneity. Journal ofEcology, 85, 611–624.
Alvarez-Buylla, E.R. & Martinez-Ramos, M. (1992) Demo-graphy and allometry of Cecropia obtusifolia, a neotropicalpioneer tree – an evaluation of the climax – pioneer para-digm for tropical rain forests. Journal of Ecology, 80, 275–290.
Ashton, P.S. (1978) Crown characteristics of tropical trees.Tropical Trees as Living Systems (eds P.B. Tomlinson &M.H. Zimmermann), pp. 591–615. Cambridge UniversityPress, Cambridge.
Barthélémy, D. (1991) Levels of organisation and repeti-tion phenomena in seed plants. Acta Biotheoretica, 39,309–323.
Bell, A.D. (1991) Plant form. An Illustrated Guide to FloweringPlant Morphology. Oxford University Press, Oxford.
Boardman, N.K. (1977) Comparative photosynthesis of sunand shade plants. Annual Review of Plant Physiology, 28,355–377.
Bongers, F., Popma, J., Meave del Castillo, J. & Carabias, J.(1988) Structure and floristic composition of the lowlandrain forest of Los Tuxtlas, Mexico. Vegetatio, 74, 55–80.
Bongers, F. & Sterck, F.J. (1998) The architecture of tropicalrain forest trees: responses to light. Dynamics of TropicalCommunities (eds D.M. Newbery, H.H.T. Prins & N. Brown),pp. 125–162. Blackwell Scientific Publications, Oxford.
Bonser, S.P. & Aarssen, L.W. (1994) Plastic allometry inyoung sugar maple (Acer saccharum): adaptive responses tolight availability. American Journal of Botany, 81, 400–406.
Canham, C.D. (1988) Growth and canopy architecture ofshade-tolerant trees: response to canopy gaps. Ecology, 69,786–795.
Chazdon, R.L. & Fetcher, N. (1984) Light environments oftropical forests. Physiological Ecology of Plants of the WetTropics (eds E. Medina, H.A. Mooney & C. Vasquez- Yanes),pp. 27–36. Dr W. Junk, The Hague.
Clark, D.A. & Clark, D.B. (1992) Life history diversity ofcanopy and emergent trees in a Neotropical rain forest.Ecological Monographs, 62, 315–344.
Clark, D.B., Clark, D.A., Rich, P.M., Weiss, S. & Oberbauer, S.F.(1996) Landscape-evaluation of understorey light and canopystructure: methods and application to a neotropical lowlandrain forest. Canadian Journal of Forest Research, 26, 747–757.
Drénou, C. (1994) Approche architectural de la scenesencedes arbres: le cas de quelques angiospermes temperées ettropicales. PhD Thesis, Université de Montpellier II,Montpellier.
Edelin, C. (1990) The Monopodial Architecture: the Case ofsome Tree Species from Tropical Asia. Research Pamphletno. 105. Forest Research Institute Malaysia.
Eijkelkamp, B.V. (1991) Data Image Analysis System: a Manual.Enschede.
Ellsworth, D.S. & Reich, P.B. (1993) Canopy structure andvertical patterns of photosynthesis and related plant traitsin a deciduous forest. Oecologia, 96, 169–178.
Fisher, J.B. (1986) Branching patterns and angles in trees. Onthe Economy of Plant Form and Function. (ed. T.J. Givnish),pp. 493–523. Cambridge University Press, Cambridge.
Givnish, T.J. (1988) Adaptation to sun and shade: a wholeplant perspective. Australian Journal of Plant Physiology,15, 63–92.
Hallé, F. & Oldeman, R.A.A. (1970) Essai sur l’Architectureet Dynamique de Croissance des Arbres Tropicaux. Masson,Paris.
Hallé, F., Oldeman, R.A.A. & Tomlinson, P.B. (1978) Tropicaltrees and forests. An Architectural Analysis. SpringerVerlag, Berlin.
Hartshorn, G.S. (1978) Tree falls and forest dynamics.Tropical Trees as Living Systems (eds P.B. Tomlinson &M.H. Zimmermann), pp. 617–638. Cambridge UniversityPress, Cambridge.
Horn, H.S. (1971) The Adaptive Geometry of Trees. PrincetonUniversity Press, Princeton.
Jurik, T.W. (1986) Temporal and spatial patterns of specificleaf weight in successional northern hardwood species.American Journal of Botany, 73, 1083–1092.
King, D.A. (1991) Correlations between biomass allocation,relative growth rate and light environment in tropical forestsaplings. Functional Ecology, 5, 485–492.
King, D.A. (1993) Growth history of a Neotropical treeinferred from the spacing of leaf scars. Journal of TropicalEcology, 9, 525–532.
King, D.A. (1994) Influence of light level on the growth andmorphology of saplings in a Panamanian forest. AmericanJournal of Botany, 81, 948–957.
King, D.A. (1996) Allometry and life history of tropical trees.Journal of Tropical Ecology, 12, 25–44.
King, D.A. & Maindonald, J.H. (1999) Tree architecture inrelation to leaf dimensions and tree stature in temperate andtropical rain forests. Journal of Ecology, 87, 1012–1024.
Kingsolver, J.G. & Schemske, D.W. (1991) Path analyses ofselection. Trends in Ecology and Evolution, 6, 276–280.
Kitajima, K. (1996) Ecophysiology of tropical tree seedlings.Tropical Forest Plant Ecophysiology (eds S.S. Mulkey,R.L. Chazdon & A.P. Smith), pp. 559–596. Chapman &Hall, New York.
Kohyama, T. & Hotta, M. (1990) Significance of allometry intropical saplings. Functional Ecology, 4, 515–521.
Koop, H. & Sterck, F.J. (1994) Light penetration throughstructurally complex forest canopies: an example of a low-land tropical rainforest. Forest Ecology and Management,69, 111–122.
Küppers, M. (1989) Ecological significance of above groundarchitectural patterns in woody plants: a question of cost-benefit relationships. Trends in Ecology and Evolution, 4,375–379.
Leopold, L.B. (1971) Trees and streams: the efficiency ofbranching patterns. Journal of Theoretical Biology, 31,339–354.
Niinemets, U. & Kull, O. (1995) Effect of light availabilityand tree size on the architecture of assimilative surface inthe canopy of Picea abies: variation in needle morphology.Tree Physiology, 15, 307–315.
Niinemets, U. (1997) Acclimation to low irradiance inPicea abies: influence of past and present light climate onfoliage structure and function. Tree physiology, 17,723–732.
Oberbauer, S.F. & Strain, B.R. (1986) Effects of canopy posi-tion and irradiance on the leaf physiology and morphologyof Pentaclethra macroloba (Mimosaceae). American Journalof Botany, 73, 409–416.
JEC525.fm Page 12 Monday, March 12, 2001 4:08 PM

13Crown development in tropical trees
© 2001 British Ecological Society, Journal of Ecology, 89, 1–13
Oldeman, R.A.A. (1974) Architecture de la Forêt Guyanaise.Mémoire 73. ORSTOM, Paris.
Oldeman, R.A.A. (1989) Biological implications ofLeguminous tree architecture. Advances in Legumebiology. Monographs in Systematic Botany of MissouriBotanical Garden, no. 29 (eds C.H. Stirton & J.L. Zarucchi),pp. 17–34. Missouri Botanical Garden, Missouri.
Orians, G.H. (1982) The influence of treefalls in tropicalforests in tree species richness. Journal of Tropical Ecology,23, 255–279.
Pearcy, R.W. (1987) Photosynthetic gas exchange responsesof Australian tropical forest trees in canopy, gap and under-story micro-environments. Functional Ecology, 1, 169–178.
Petraitis, P.S., Dunham, A.E. & Niewiarowski, P.H. (1996)Inferring multiple causality: the limitations of path analysis.Functional Ecology, 10, 421–431.
Poorter, L., Oberbauer, S.F. & Clark, D.B. (1995) Leaf opticalproperties along a vertical gradient in a tropical rain forestcanopy in Costa Rica. American Journal of Botany, 82,1257–1263.
Popma, J. & Bongers, F. (1988) The effect of canopy gaps ongrowth and morphology of seedlings of rain forest species.Oecologia, 75, 625–632.
Rijkers, T., Pons, T.L. & Bongers, F. (2000) The effects of treeheight and light availability on photosynthetic leaf traitsof four neotropical species differing in shade tolerance.Functional Ecology, 14, 77–86.
Prévost, M.F. (1967) Architecture de quelques Apocynacéesligneuses. Mémoires de la Société Botanique de France.Colloque sur la physiology de l’arbre, 23–36.
Room, P.M., Maillette, L. & Hanan, J.S. (1994) Modular andmetamer dynamics and virtual plants. Advances in EcologicalResearch, 25, 105–157.
Sabatier, D. & Prévost, M.F. (1989) Quelques donneés surla composition floristique et la diversité des peuplementsforestiers de Guyana française. Revue Bois et Forêts desTropiques, 219, 31–55.
Schulz, J.P. (1960) Ecological Studies on Rain Forest in NorthernSuriname. Mededelingen Botanisch Museum en Herbarium,RUU, no. 163. Noord-Hollandsche Uitgevers Maatschappij,Amsterdam.
Shukla, R.P. & Ramakrishnan, P.S. (1986) Architecture andgrowth strategies of tropical trees in relation to successionalstatus. Journal of Ecology, 74, 33–46.
Sokal, R.R. & Rohlf, F. (1981) Biometry: the Principles and
Practice of Statistics in Biological Research. WH Freemanand company, San Francisco.
Steingraeber, D.A., Lawrence, J.K. & Franck, D.H. (1979)Variation of shoot morphology and bifurcation ratio insugar maple (Acer saccharum) saplings. American Journalof Botany, 66, 441–445.
Sterck, F.J. (1997) Trees and light: tree development andmorphology in relation to light availability in a tropical rainforest in French Guiana. PhD Thesis, Wageningen AgriculturalUniversity, Ponser & Looijen, Wageningen.
Sterck, F.J. (1999) Crown development in gaps and understorey.Plant Ecology, 143 (1), 89–98.
Strauss-Debenedetti, S. & Bazzaz, F.A. (1996) Photosyntheticcharacteristics of tropical trees along successional gradients.Tropical Forest Plant Ecophysiology (eds S.S. Mulkey,R.L. Chazdon & A.P. Smith), pp. 162–186. Chapman &Hall, New York.
Terborgh, J. (1985) The vertical component of plant speciesdiversity in temperate and tropical forests. AmericanNaturalist, 126, 760–776.
Ter Steege, H. (1996) Winphot 5: a programme to analyzevegetation indices, light and light quality from hemisphericalphotographs. Tropenbos Guyana Preports 95–2, TropenbosGuyana Programme, Georgetown, Guyana.
Thomas, S.C. & Ickes, K. (1995) Ontogenetic changes inleaf area in Malaysian rain forest trees. Biotropica, 27,427–434.
Veneklaas, E.J. & Poorter, L. (1998) Growth and carbon part-itioning of tropical tree seedlings growing in contrastinglight environments. Inherent variation in plant growth.Physiological Mechanisms and Ecological Consequences(eds H. Lambers, H. Poorter & M.I. van Vuuren), pp. 337–361. Backhuys Publishers, Leiden.
White, J. (1979) The plant as a metapopulation. Annual Reviewof Ecology and Systematics, 10, 109–145.
Whitney, G.G. (1976) The bifurcation ratio as an indicator ofadaptive strategy in woody plant species. Bulletin of theTorrey Botanical Club, 103, 67–72.
Wright, S. (1934) The method of path coefficients. Annals ofMathematics and Statistics, 5, 161–215.
Yoda, K. (1974) Three dimensional distribution of lightintensity in a tropical rain forest of West Malaysia. JapaneseJournal of Ecology, 24, 247–254.
Received 29 September 1999 received 25 February 2000
JEC525.fm Page 13 Monday, March 12, 2001 4:08 PM





























