Embed Size (px)
Citation preview

PDtr
HAFra N
37b In
1.
Sp
Veterinary Microbiology 171 (2014) 448–459
A
Ke
ES
E.
Re
W
*
htt
03
revalence and characteristics of ESBL-producing E. coli inutch recreational waters influenced by wastewatereatment plants
etty Blaak a,*, Patrick de Kruijf a, Raditijo A. Hamidjaja a,ngela H.A.M. van Hoek a, Ana Maria de Roda Husman a,b,anciska M. Schets a
ational Institute for Public Health and the Environment (RIVM), Centre for Zoonoses and Environmental Microbiology, PO Box 1,
20 BA Bilthoven, The Netherlands
stitute for Risk Assessment Sciences, Utrecht University, PO Box 80178, 3508 TD Utrecht, The Netherlands
Introduction
During the last two decennia, the prevalence of Extendedectrum Beta-Lactamase (ESBL)-producing bacteria has
increased worldwide (Canton et al., 2008; Castanheira et al.,2008). ESBL-producing bacteria are resistant to most beta-lactam antibiotics, including 3rd and 4th generationcephalosporins, and are often additionally resistant tomultiple other classes of antibiotics. This severely limitstreatment options for infections caused by these bacteria,which has led to an increased use of last-resort antibioticssuch as carbapenems (Canton et al., 2012). Although initially
R T I C L E I N F O
ywords:
BL
coli
creational water
astewater treatment plants
A B S T R A C T
Outside health care settings, people may acquire ESBL-producing bacteria through
different exposure routes, including contact with human or animal carriers or
consumption of contaminated food. However, contact with faecally contaminated surface
water may also represent a possible exposure route. The current study investigated the
prevalence and characteristics of ESBL-producing Escherichia coli in four Dutch
recreational waters and the possible role of nearby waste water treatment plants
(WWTP) as contamination source. Isolates from recreational waters were compared with
isolates from WWTP effluents, from surface water upstream of the WWTPs, at WWTP
discharge points, and in connecting water bodies not influenced by the studied WWTPs.
ESBL-producing E. coli were detected in all four recreational waters, with an average
concentration of 1.3 colony forming units/100 ml, and in 62% of all samples. In surface
waters not influenced by the studied WWTPs, ESBL-producing E. coli were detected in
similar concentrations, indicating the existence of additional ESBL-E. coli contamination
sources. Isolates with identical ESBL-genes, phylogenetic background, antibiotic resistance
profiles, and sequence type, were obtained from effluent and different surface water sites
in the same watershed, on the same day; occasionally this included isolates from
recreational waters.
Recreational waters were identified as a potential exposure source of ESBL-producing E.
coli. WWTPs were shown to contribute to the presence of these bacteria in surface waters,
but other (yet unidentified) sources likely co-contribute.
� 2014 Elsevier B.V. All rights reserved.
Corresponding author. Tel.: +31 30 274 7005; fax: +31 30 274 4434.
E-mail address: [email protected] (H. Blaak).
Contents lists available at ScienceDirect
Veterinary Microbiology
jou r nal h o mep ag e: w ww .e ls evier . co m/lo c ate /vetm i c
p://dx.doi.org/10.1016/j.vetmic.2014.03.007
78-1135/� 2014 Elsevier B.V. All rights reserved.

Eanm(LMcfoMnvthp(ewinpofr
hopscthmfetrrthch
F
b
W
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 449
SBL-production was typically associated with hospital-cquired infections caused by Klebsiella pneumoniae, it isow also associated with community-acquired infections,ainly urinary tract infections caused by Escherichia coli
ivermore et al., 2007; Paterson and Bonomo, 2005).oreover, ESBL-producing E. coli are present among the
ommensal E. coli population in healthy individuals andod-producing animals (Huijbers et al., 2013; Nethmap_-ARAN, 2013; Trott, 2013). Commensal E. coli generally do
ot cause disease, however, spread of ESBL-producingariants through human and animal populations is never-eless worrisome, and may lead to increased exposure of
opulations more susceptible to opportunistic infections.g. the elderly or hospitalized individuals). Additionally,ith an increasing number of ESBL-producing E. coli carriers
the human population the risk increases that gutathogens efficiently acquire resistance by gene transferf ESBL-genes as well as other antibiotic resistance genesom ESBL-producing E. coli in the intestinal tract.
Dissemination of ESBL-producing E. coli outside theealth care setting may be facilitated by contact with humanr animal carriers, or consumption of contaminated animalroducts. Additionally, a possible role for the environmenthould be considered in this regard. Since E. coli areommensal bacteria, they are abundantly excreted intoe environment, amongst others through application ofanure as fertilizer or droppings of pasture animals, withces of wild animals, and with discharge of (partially)eated wastewater, or with sewage overflows during heavy
ainfall. Some of the commensal as well as pathogenic E. coli
at are excreted into the environment may have theapacity to produce ESBL. Indeed, ESBL-producing E. coli
ave been detected in surface water worldwide, including
the Netherlands (Blaak et al., 2011; Chen et al., 2010; Dhanjiet al., 2011; Hong et al., 2004). Human exposure to thesebacteria may occur, for instance during recreation incontaminated surface water, or indirectly, when contami-nated surface water is used for irrigation of (raw consumed)crops, therewith contributing to community-associateddissemination of ESBL-producing E. coli. Additionally,contaminated surface water might contribute to exposureof animals (wild life as well as livestock) that drink from it.
In order to limit dissemination of ESBL-producing E. coli
through the environment, insight in the contribution ofdifferent possible environmental contamination sourcesand exposure routes is essential. The current studydetermined the presence of ESBL-producing E. coli in fourDutch recreational water regions, and the possiblecontribution of nearby wastewater treatment plants.
2. Materials and methods
2.1. Sampling and sampling locations
Three recreational waters appointed under EuropeanBathing Water Directive 2006/7/EC (‘official’) (Anony-mous, 2006) and one not appointed (‘unofficial’) recrea-tional water were sampled during the bathing seasons of2011 and/or 2012 which, in the Netherlands, lasts fromMay 1st until September 30th. The recreational waterswere situated in different regions in the Netherlands, andeach was located 1–2 km (as the crow flies) from awastewater treatment plant (WWTP) that did not disinfecttreated effluents before discharge (Fig. 1). Two of theofficial recreational waters (regions A and B) were situatedin freshwater lakes; the third (region C) was located in the
ig. 1. Sampling locations in regions A, B, C, and D. *For regions A and B, the direction of currents is variable and ‘upstream’ and ‘downstream’ annotations are
ased on the main current direction. ** The entire river is used for recreation, the indicated site is chosen to represent recreational water downstream of an
WTP.

No(recasu(‘dindewmrereofresmdowre
saDuthsaWsuwreWWof
Ta
Co
R
A
B
C
D
a
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459450
rth Sea. The fourth and unofficial recreational watergion D) was a small river that is frequently used for
noeing. All recreational waters were connected to therface water where WWTP effluents were dischargedischarge points’). At both lake-side recreational waters
regions A and B, the direction of the current is variable,pending on wind direction and operational schedules of
ater pumping engines and/or sluices. In both cases theain current direction is from WWTP discharge points tocreational waters, and for descriptional simplicity, thesecreational waters are referred to as located ‘downstream’
the WWTPs. The currents at WWTP discharge points ingion C and D are always directed from east to west. Theall river in region D was sampled both upstream andwnstream of the WWTP situated at the river bank, of
hich the downstream sample was taken to representcreational water under influence of the WWTP.
In 2011, recreational waters in regions A–C werempled four or five times during the bathing season.ring the 2012 bathing season, waters were sampled
ree (A, C, D) or four (B) times (Table 1), and at eachmpling time-point additional samples were taken fromWTP effluents, surface water at WWTP discharge points,rface water located upstream of the WWTPs, and surfaceaters from connecting water bodies upstream of thecreational waters but not under influence of the studiedWTPs (Fig. 1). In region B, surface water ‘upstream of theWTP’ was not sampled. Instead, because of the presence
two dams between WWTP and the recreational water
(possibly interfering with passage of water between thetwo sites), an additional surface water sample ‘down-stream’ of the WWTP, but ‘upstream’ of the recreationalwater was taken. Due to logistic problems, dischargepoints were only sampled twice in regions B and D.
Prior to the day of sampling of surface water, effluentswere collected by WWTP staff using automated systemsthat continuously sample effluent, rendering 24 h flow-proportional, homogenous samples that are collected inmixing vessels. One liter of effluent sample was taken fromthese mixing vessels. On the third sampling date for regionC, no 24 h flow-proportional effluent sample was available,and a 1 l grab sample was taken from the effluent duct onthe morning of surface water sampling. Surface watersamples (1 l) were taken according to NEN-EN-ISO 19458(Anonymous, 2007). Samples were transported and storedat 5 � 3 8C, and analyzed within 24 h after sampling.
2.2. Isolation and enumeration of E. coli and ESBL-producing
E. coli
From each sample, multiple volumes were filteredthrough 0.45 mm pore size membrane filters (Millipore,Amsterdam, the Netherlands), and E. coli was isolatedaccording to ISO 9308-1 ‘Rapid test’ (Anonymous, 2000). Inshort, filters were incubated on TSA for 4–5 h at 36 � 2 8C,and then transferred to TBA and incubated for 19–20 h at44 � 0.5 8C. Presumptive E. coli colonies were stained withJames reagent to test for indole production (Biomerieux,
ble 1
ncentrations ESBL-E. coli and E. coli in Dutch recreational waters.
egion Sampling date Concentrations (cfu/100 ml) (95%-Confidence Interval) % ESBL
ESBL-E. coli E. coli
4-7-2011 0.29 (0.05–0.88) 2.9 � 102 (2.2 � 102–3.6 � 102) 0.10
25-7-2011 15 (12–18) 1.8 � 103 (1.6 � 103–1.9 � 103) 0.83
8-8-2011 1.9 (1.1–3.1) 1.2 � 103 (7.4 � 102–1.7 � 103) 0.17
6-9-2011 0.15 (0.0084–0.65) 67 (52–84) 0.22
18-6-2012 1.0 (0.44–2.0) 4.8 � 102 (4.0 � 102–5.8 � 102) 0.21
2-7-2012 1.2 (0.54–2.2) 1.3 � 102 (1.1 � 102–1.5 � 102) 0.93
13-8-2012 0 (0–0.33)a 83 (69–99) <0.4
7-6-2011 4.8 (3.7–6.1) 5.1 � 103 (4.2 � 103–6.2 � 103) 0.093
4-7-2011 0 (0.00–0.27) 1 1.4 � 102 (1.1 � 102–1.6 � 102) < 0.2
25-7-2011 0.15 (0.0084–0.65) 2.7 � 102 (2.1 � 102–3.4 � 102) 0.055
8-8-2011 0 (0.00–0.28) 1 2.3 � 102 (2.0 � 102–2.6 � 102) <0.1
6-9-2011 0.15 (0.0084–0.65) 2.1 � 102 (1.6 � 102–2.8 � 102) 0.069
21-5-2012 0 (0–0.28)a 23 (15.0–33.1) <1.2
2-7-2012 0 (0–0.28)a 32 (24.7–40.2) <0.9
13-8-2012 0 (0–0.28)a 48 (39.6–57.0) <0.6
24-9-2012 0.29 (0.049–0.91) 92 (75.5–111.6) 0.32
21-6-2011 0 (0.00–0.64)a 31 (21.3–43.2) <2
4-7-2011 0 (0.00–0.27)a 29 (19.7–40.9) <0.9
25-7-2011 3.4 (2.2–5.0) 6.2 � 102 (5.2 � 102–7.4 � 102) 0.54
8-8-2011 0.17 (0.01–0.76) 34 (23.8–46.7) 0.51
6-9-2011 1.0 (0.44–2.0) 1.3 � 102 (1.1 � 102–1.6 � 102) 0.77
4-6-2012 0 (0–0.29)a 23 (14.5–33.1) <1.3
16-7-2012 6.2 (4.5–8.2) 7.5 � 102 (6.1 � 102–9.2 � 102) 0.82
27-8-2012 0.29 (0.049–0.91) 45 (22.8–79.7) 0.65
18-6-2012 0.29 (0.049–0.91) 3.1 � 102 (2.8 � 102–3.4 � 102) 0.10
30-7-2012 8.1 (6.1–10) 8.3 � 102 (7.2 � 102–9.6 � 102) 0.97
10-9-2012 0.15 (0.0084–0.65) 2.6 � 102 (2.3 � 102–2.8 � 102) 0.057
No ESBL-producing E. coli were detected. cfu = colony forming units.

BwbcBcdncth
c
aSgcBwC2aw(3mcaainc2aclo3Ein9
2
psccscpmuIn
2
kpsAsrs
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 451
oxtel, the Netherlands), and only indole-positive coloniesere counted. Indole positive isolates were further confirmed
y testing for ß-glucuronidase-activity on Brilliance E. coli/oliform agar (BECSA; Oxoid, Badhoevedorp, the Netherlands).eta-glucuronidase-positive, indole-positive isolates wereonsidered to be E. coli. API20E (Biomerieux) was used toetermine whether or not indole-positive, ß-glucuronidase-egative isolates were E. coli. E. coli concentrations werealculated based on the number of indol-positive colonies ande fraction of these colonies that was confirmed to be E. coli.
For the isolation and enumeration of ESBL-producing E.
oli, membrane filters were placed on ChromIDTM ESBLgar (Biomerieux) and incubated for 18–24 h at 36 � 2 8C.pecies identity of suspected ESBL-E. coli isolates (i.e. ß-lucuronidase-positive on ChromIDTM ESBL agar) wereonfirmed by indole-testing using BBL Dry SlideTM (BD,reda, The Netherlands). Suspected ESBL-producing isolatesere tested for ESBL-production by disk diffusion following
LSI guidelines (Clinical and Laboratory Standards Institute,010), using Sensi-DiscTM (BD, Breda, the Netherlands)ccording to the manufacturer’s instructions. Zone diametersere determined for cefotaxime (30 mg/ml), cefotaxime0 mg/ml) + clavulanic acid (10 mg/ml), ceftazidime (30ug/l), ceftazidime (30ug/ml) + clavulanic acid (10ug/ml) and
efoxitin (30 mg/ml). ESBL-producing isolates were defineds strains resistant to cefotaxime (zone diameter �22 mm)nd/or ceftazidime (zone diameter �17 mm), and a reduction
zone diameter of �5 mm with the disks containinglavulanic acid (Clinical and Laboratory Standards Institute,010). Isolates without a significant effect of clavulanic acidnd resistant to cefoxitin (zone diameter �14 mm) wereonsidered AmpC-producing (Jacoby, 2009). Because of thew isolation frequency of AmpC-producing isolates (8 of
94), these were excluded from further analyses. E. coli andSBL-producing E. coli concentrations and 95%-confidencetervals (CI) were calculated using Mathematica software
.0.1 (WolframResearch, Champaign, IL, USA).
.2.1. Antibiotic resistance profiles
Using Sensi-DiscsTM, phenotypically confirmed ESBL-roducing E. coli isolates (n = 386) were tested forusceptibility to 12 antibiotics of human and veterinarylinical relevance: tetracycline, ampicillin, amoxicillin/lavulanic acid (co-amoxiclav), gentamicin, streptomycin,ulfisoxazole, trimethoprim, chloramphenicol, ciprofloxa-in, nalidixic acid, imipenem, and meropenem. Tests wereerformed according to CLSI guidelines following theanufacturer’s instructions, and resistance determined
sing CLSI breakpoints (Clinical and Laboratory Standardsstitute, 2010).
.3. Phylogenetic typing
Of 386 confirmed ESBL-producing E. coli isolates withnown antibiotic resistance profiles, 214 were selected forhylogenetic group analysis. Of these, 123 were randomlyelected across samples and from isolates with differentBR phenotypes (defined as ‘random subset’), and 91 wereelected because of an observed match in phenotype withespect to ABR profile between isolates from differentampling sites within the same region at the same
sampling time-point (defined as ‘matches subset). Isolateswere allotted to phylogenetic groups A, B1, B2 or D, using aPCR targeted to the chuA, yjaA genes and TspE4.C2 DNAfragment, using primers described by Clermont et al.(2000). Material from one single colony was suspended inTris EDTA buffer (pH 8.0, Sigma-Aldrich, Zwijndrecht, theNetherlands) and cells were lysed at 70 8C for 5 min. DNA-extracts were stored at �20 8C. Targets were amplifiedusing QIAGEN Multiplex PCR kit (Qiagen Benelux BV,Venlo, the Netherlands), in 1.5 ml of 10� diluted DNAextract and using 5 pmol of each primer. Amplificationconditions were adapted from Clermont et al. (2000):15 min 95 8C, followed by 35 cycles of 10 s 95 8C, 20 s 60 8C,30 s 72 8C, and a final elongation step of 5 min 72 8C. Strainswere sub-grouped according to (Escobar-Paramo et al.,2006): subgroup A0: chuA�, yjaA�, and TspE4.C2�;subgroup A1: chuA�, yjaA+, and TspE4.C2�; group B1:chuA�, yjaA�, and TspE4.C2+; subgroup B22: chuA+, yjaA+,and TspE4.C2�; subgroup B23: chuA+, yjaA+, and TspE4.C2+;subgroup D1: chuA+, yjaA�, and TspE4.C2�; subgroup D2:chuA+, yjaA�, and TspE4.C2+.
2.4. ESBL genotyping
ESBL-genotypes were determined for a subset of 154 of214 isolates with determined phylogroup, including 70from the ‘random subset’ and 56 from the ‘matches subset’(which were identified as matches with respect tophylogroup as well as ABR profiles). Additionally, ESBL-genotypes were determined for 28 isolates with uniqueABR profiles that were not included in phylogenetic groupanalysis. Isolates were analyzed for the presence of genesencoding CTX-M-group 1, CTX-M-group 2, and CTX-M-group 9, and OXA-, SHV- and TEM-genes, by multiplexPCRs using primers described by Dallenne et al. (2010). Foramplification, 3 ml of DNA extract (the same as was usedfor phylogenetic typing) was mixed with 10 pmol of eachprimer and 12.5 ml Qiagen Multiplex PCR mix (QiagenBenelux BV, Venlo, the Netherlands) in a final volume of25 ml. Amplification conditions were as described byDallenne et al. (2010). PCR-products were analyzed onagarose gel. To identify individual alleles, PCR-products ofthe expected size (TEM: 800 bp, SHV: 713, CTX-M group1:688 bp, CTX-M group 2:404 bp, CTX-M-group 9: 561 bp)were treated with ExoSAP-IT (GE Healthcare, Hoevelaken,the Netherlands) and sequenced using the same primersused to generate the PCR-products, and BigDye Terminatorv3.1 Cycle Sequencing kit (Applied Biosystems, Bleiswijk,the Netherlands). Thus obtained partial ESBL genesequences were compared with ESBL gene sequences inthe GenBank database and on the Lahey website (www.la-hey.org/Studies). Partial sequence analysis allows identifi-cation of particular clusters of homologous alleles, towhich, for sake of annotation, the lowest allele numberbelonging to a cluster was assigned, e.g. blaCTX-M-1 includesblaCTX-M-1 and blaCTX-M-61, see also Fig. 5).
2.4.1. Exclusion of within sample copy strains
Of all isolates for which ABR profiles, ESBL genotypesand phylogenetic groups were determined, 10 were ‘twin’isolates, i.e. isolates identical with respect to these

chwexwde
2.4
anidcochsawgeam(2MFom(In(InTaAmfopu
eloansizlaksaTeBlthdb
3.
3.1
su
reofco
tofrorepoco
efthupstuwre
3.2
ES
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459452
aracteristics, originating from the same sample. Since itas likely that these isolates were copy strains, they werecluded from prevalence analyses, leaving 204 isolatesith assigned phylogenetic groups and 144 isolates withtermined ESBL-genotypes.
.2. Multilocus sequence typing (MLST)
Among isolates for which ABR profiles, ESBL genotypesd phylogenetic groups were determined, 37 wereentified that could be grouped into 15 sets of isolatesnsisting of variants identical with respect to thesearacteristics, obtained at different sites in a region on theme sampling date. Of these 37 isolates, sequence typesere determined. For this purpose, seven house-keepingnes, adk, fumC, gyrB, icd, mdh, purA and recA, wereplified and sequenced as described by Wirth et al.
006). Primer sequences were obtained from the E. coli
LST database website http://mlst.ucc.i.e./mlst/dbs/Ecoli.r amplification, 2 ml of 10� diluted DNA extract wasixed with 200 pmol of each primer, 1� PCR buffervitrogen, Bleiswijk, the Netherlands), 2.5 mM MgCl2
vitrogen), 200 mM dNTP mix (Invitrogen), and 1.25Uq polymerase (Invitrogen) in a final volume of 50 ml.
plification conditions were as follows: 5 min 95 8C,llowed by 35 cycles of 30 s 95 8C, 30 s 60 8C (adk, icd, mdh,rA, recA) or 30 s 64 8C (fumC, gyrB), 45 s 72 8C, and a finalngation step of 10 min 72 8C. PCR-products were
alyzed on agarose gel and PCR-products of the expectede were treated with ExoSAP-IT (GE Healthcare, Hoeve-en, the Netherlands) followed by sequencing using the
me primers used to generate PCR-products, using BigDyerminator v3.1 cycle sequencing kit (Applied Biosystems,eiswijk, the Netherlands). Sequences were imported ine E. coli MLST database website (http://mlst.ucc.i.e./mlst/s/Ecoli) to determine MLST types.
Results
. ESBL-producing E. coli in recreational waters and other
rface waters in the vicinity
ESBL-producing E. coli were detected in all fourcreational waters, in 44% (region B) to 100% (region D)
samples (Table 1). Concentrations of ESBL-producing E.
li ranged from 0.15 to 15 cfu/100 ml. For comparison,tal E. coli concentrations in the same samples rangedm 23 to 5.2 � 103 cfu/100 ml. ESBL-producing E. coli
presented 0.05–1% of the total E. coli population insitive samples. In all regions, concentrations of ESBL-E.
li at WWTP discharge points were comparable to that influents, and on average 2- to 3-log 10 units higher thanat in the recreational waters (Fig. 2). In surface watersstream or not under the influence of the WWTPsdied, average concentrations of ESBL-producing E. coli
ere in the same range or slightly higher than those in thecreational waters in the same area (Fig. 2).
. Antibiotic resistance profiles of ESBL-producing E. coli
Next to cephalosporins and ampicillin (explained by
from surface waters, including recreational waters, andWWTP effluents were resistant to sulfoxisole (74%),trimethoprim (67%), tetracycline (60%), and nalidixic acid(60%). Resistance to ciprofloxacin (42%) and streptomycin(49%) was also common, while resistance to gentamicin(29%), chloramphenicol (19%), coamoxiclav (10%), andcefoxitin (3%) was less frequently observed. None of theisolates were resistant to the carbapenem antibioticsimipenem and meropenem. Among ESBL-producing iso-lates from recreational waters, resistance to (fluoro)qui-nolones, aminoglycosides and tetracycline appeared lowercompared to isolates from wastewater and other surfacewaters (Fig. 3a). Overall, 71% of isolates from WWTPeffluents, 63% of isolates from discharge points, 62% ofisolates from surface waters upstream and not underinfluence of the investigated WWTPs, and 41% of isolatesfrom recreational waters were resistant to at least threeclasses of antibiotics additional to beta-lactams, andtherewith defined as multidrug-resistant (MDR)(Fig. 3b). In total, 14% of all ESBL-producing E. coli wereresistant to five (12%), six (1.8%) or even all seven (0.7%)additional classes of antibiotics.
For each sampling region, the resistance phenotypesof isolates resistant to one to four classes of antibiotics inaddition to beta-lactams were compared betweensampling sites (Fig. 4). Resistance phenotypes detectedin recreational waters were frequently, but not always,detected in WWTP effluents and/or in surface waterslocated upstream of recreational waters, includingsurface water at WWTP discharge points and surfacewater upstream of WWTPs. Isolates resistant to five or sixclasses of antibiotics were not included in this analysis,because there was hardly any variation in phenotypesamong such isolates (e.g. almost two-third of all isolatesresistant to five classes of antibiotics next to beta-lactams were resistant to sulfanomides/trimethoprim,(fluor)quinolones, tetracycline, aminoclycosides and
Fig. 2. Concentrations ESBL-producing E. coli in surface water and
wastewater. Error bars represent the standard error of the means.
Numbers on top of the bars represent the number of samples on which
average concentrations were based. For region A, results from two sites
upstream of the WWTP have been pooled, for region C results from two
surface waters not influenced by studied WWTP have been pooled;
ns = not sampled; SW = surface water; n.u.i. = not under the influence of
studied WWTP.
loramphenicol).
BL production), the majority of all ESBL-producing E. coli ch
3
ow(FfotithspindogdwW
F
a
in
o
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 453
.3. ESBL-genotypes and phylogenetic groups
Almost two-third of 144 ESBL-producing E. coli
btained from Dutch surface waters, including recreationalater, and WWTP effluents carried blaCTX-M-15 or blaCTX-M-1
ig. 5a). These two ESBL-genes were predominant in allur regions, although blaCTX-M-1 was detected about twomes less frequently in region C compared to the otherree regions (Fig. 5b). For some ESBL-genotypes region-
pecific patterns were observed, e.g. blaCTX-M-14 was morerevalent in regions A and B, blaSHV-12 was more prevalent
regions B and C, blaCTX-M-3 and blaCTX-M-9 were onlyetected in region C, and blaCTX-M-32 and blaTEM-52 werenly detected in regions A and C. The least frequentenotypes were blaCTX-M-2 and blaCTX-M-24 which were bothetected once (0.7% of all isolates). All ESBL-genotypes thatere detected in recreational waters were also detected inWTP effluents and/or surface waters located upstream of
the recreational waters (including discharge points andwaters upstream of WWTPs). This was also the case forrelatively infrequently observed and region-specific geno-types, such as blaCTX-M-9, blaCTX-M-14, blaCTX-M-32, blaSHV-12,and blaTEM-52 (Fig. 5b).
When ESBL-genotypes were compared within thecontext of E. coli phylogenetic groups, it appeared that eventhough the same ESBL-genes were detected at different siteswithin a region, they were often present in diversephylogenetic backgrounds (Fig. 6). In a minority of cases(n = 13) in regions A, C and D, E. coli of the same phylogeneticgroup and carrying the same ESBL-genes were observed inrecreational waters as well as WWTP effluents or upstreamlocated surface waters. Of note, even though the majority ofESBL-producing E. coli from surface water belonged tophylogenetic groups generally associated with commensalE. coli, also phylogenetic (sub)groups associated withvirulent strains, i.e. B2 and D2 (Anastasi et al., 2012; Duriez
ig. 3. Resistance phenotypes of ESBL-producing E. coli from surface waters and wastewaters. Shown are the percentages of isolates resistant to each
ntibiotic tested (A) and percentages of isolates resistant to different numbers of antibiotic classes besides beta-lactam antibiotics (B). N.U.I = not under the
fluence of the studied WWTP (i.e. upstream of the WWTP and other connecting water bodies not under influence), D.P. = discharge point. The percentages
n top of the bars in (B) represent the percentages of multidrug resistant isolates.
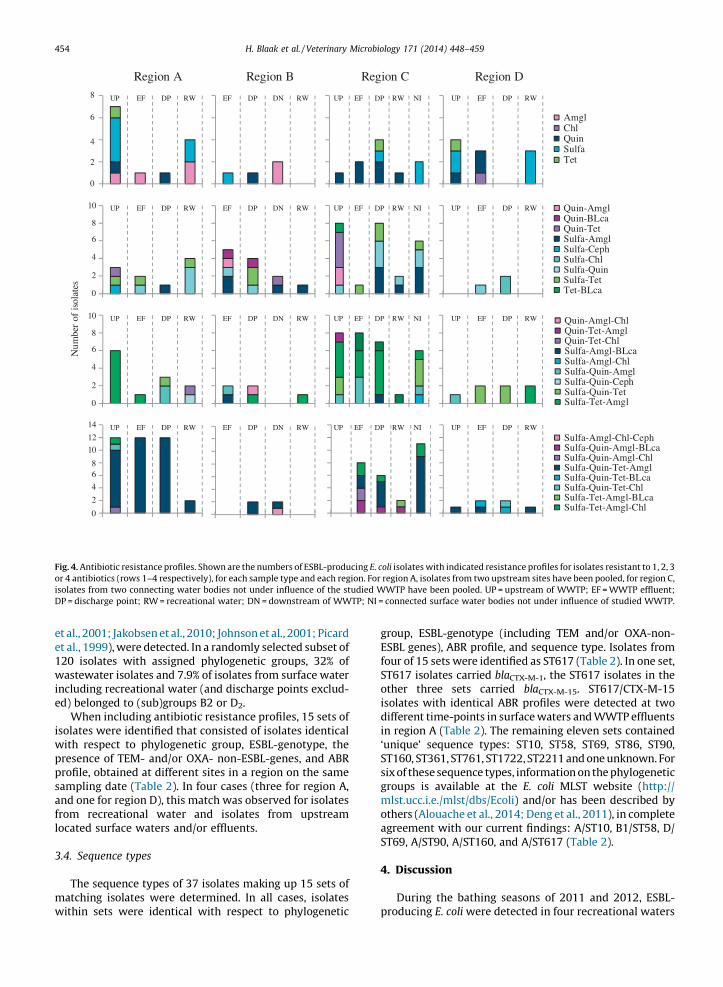
etet12wined
isowprprsaanfrolo
3.4
mw
Fig
or
iso
DP
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459454
al., 2001; Jakobsen et al., 2010; Johnson et al., 2001; Picard al., 1999), were detected. In a randomly selected subset of0 isolates with assigned phylogenetic groups, 32% of
astewater isolates and 7.9% of isolates from surface watercluding recreational water (and discharge points exclud-) belonged to (sub)groups B2 or D2.When including antibiotic resistance profiles, 15 sets oflates were identified that consisted of isolates identical
ith respect to phylogenetic group, ESBL-genotype, theesence of TEM- and/or OXA- non-ESBL-genes, and ABRofile, obtained at different sites in a region on the samempling date (Table 2). In four cases (three for region A,d one for region D), this match was observed for isolatesm recreational water and isolates from upstream
cated surface waters and/or effluents.
. Sequence types
The sequence types of 37 isolates making up 15 sets ofatching isolates were determined. In all cases, isolatesithin sets were identical with respect to phylogenetic
group, ESBL-genotype (including TEM and/or OXA-non-ESBL genes), ABR profile, and sequence type. Isolates fromfour of 15 sets were identified as ST617 (Table 2). In one set,ST617 isolates carried blaCTX-M-1, the ST617 isolates in theother three sets carried blaCTX-M-15. ST617/CTX-M-15isolates with identical ABR profiles were detected at twodifferent time-points in surface waters and WWTP effluentsin region A (Table 2). The remaining eleven sets contained‘unique’ sequence types: ST10, ST58, ST69, ST86, ST90,ST160, ST361, ST761, ST1722, ST2211 and one unknown. Forsix of these sequence types, information on the phylogeneticgroups is available at the E. coli MLST website (http://mlst.ucc.i.e./mlst/dbs/Ecoli) and/or has been described byothers (Alouache et al., 2014; Deng et al., 2011), in completeagreement with our current findings: A/ST10, B1/ST58, D/ST69, A/ST90, A/ST160, and A/ST617 (Table 2).
4. Discussion
During the bathing seasons of 2011 and 2012, ESBL-producing E. coli were detected in four recreational waters
Quin- Amg l Quin-B Lca Qu in- Tet Sulf a-Amgl Sulfa-Cep h Sulfa-C hl Sulf a-Quin Sulfa -Tet Tet-BLca
Amgl Chl Quin Sulf a Tet
Quin- Amg l-Chl Qui n-Te t-Amg l Quin- Tet- Chl Sulfa-A mgl-BLca Sulfa-A mgl-Chl Sulfa-Quin-Amgl Sulfa-Q uin-Ceph Sulfa -Qu in-Tet Sulf a-Tet-Am gl
Sulfa-A mgl-Chl-C eph Sulfa-Q uin-Amg l-BLca Sulfa-Q uin-Amg l-Chl Su lfa-Quin-Tet-Amg l Sulfa-Quin-Tet-B Lca Sulfa-Q uin-Tet-Chl Sulf a-Tet-Am gl- BLca Sulfa-Tet-A mgl-Chl
Region A Region B Region C Region D8
6
2
0
4
Num
ber
of is
olat
es
UP EF DP RW EF DP DN RW
10
8
4
0
6
2
UP EF DP NIRW
UP EF DP RW
UP EF DP RW
UP EF DP RW UP EF DP RW
EF DP DN RW
UP EF DP RW
UP EF DP RW
EF DP DN RW
EF DP DN RW
UP EF DP RW
UP EF DP NIRW
UP EF DP NIRW
UP EF DP NIRW
10
8
4
0
6
2
10
8
4
0
6
2
12
14
. 4. Antibiotic resistance profiles. Shown are the numbers of ESBL-producing E. coli isolates with indicated resistance profiles for isolates resistant to 1, 2, 3
4 antibiotics (rows 1–4 respectively), for each sample type and each region. For region A, isolates from two upstream sites have been pooled, for region C,
lates from two connecting water bodies not under influence of the studied WWTP have been pooled. UP = upstream of WWTP; EF = WWTP effluent;
= discharge point; RW = recreational water; DN = downstream of WWTP; NI = connected surface water bodies not under influence of studied WWTP.

ssmatipas
F
is
u
v
3
‘T
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 455
ituated nearby and under the influence of WWTPs. Aubstantial proportion of these ESBL-producing E. coli wasultidrug-resistant. Detection of ESBL-producing E. coli
ppeared to be associated with relatively high concentra-ons of total E. coli. Nevertheless, the proportion of ESBL-roducing E. coli relative to total E. coli numbers variedmong sites, as well as between time-points at the sameite, ranging from 0.05% to 1%. Overall, concentrations of
ESBL-producing E. coli were similar in both bathingseasons, although in region B the frequency of detectionappeared lower in 2012 as compared to 2011. At this site,the concentration of total E. coli was also lower in 2012than in the previous year. A possible explanation may bethat in 2012 a small part of the lake adjacent to the WWTPwas dammed to retain sewage overflows (but noteffluents) which were therewith no longer discharged
A.
B.
24,31%
42,36%1,39%
3,47%
0,69%
2,78%
9,72%
2,08%
0,69%
8,33%
4,17%CTX-M-1
CTX-M-15
CTX-M-3
CTX-M-32
CTX-M-2
CTX-M-9
CTX-M-14
CTX-M-27
CTX-M-24
SHV-12
TEM-52
0%
20%
40%
60%
80%
100%
Total (n=45 ) UpstreamWWTP (n=17)
WWTPeffluent(n=11)
Dischargepoint (n =6)
Recrea�onalwater (n=11 )
%Iso
lates
Region A
CTX-M-1 CTX-M-15 CTX-M-32 CTX-M-14 SHV-12 TEM-52
0%
20%
40%
60%
80%
100%
Total (n=15 ) WW TPeffluent (n=3)
Dischargepoint (n =5)
DownstreamWWTP (n=5)
Recrea�onalwater (n=2)
%Iso
lates
Region B
CTX-M-1 CTX-M-15 CTX-M-14 CTX-M-24 SHV-12
0%
20%
40%
60%
80%
100%
Total (n=60 ) UpstreamWWTP (n=4)
WWTPeffluent(n=17 )
Dischargepoint (n=17)
Recrea�onalwater (n=9)
Surfacewater N.U.I(n=13 )
%Iso
lates
Region C
CTX-M-1 CTX-M-3 CTX-M-15 CTX-M-32 CTX-M-9
CTX-M-14 CTX-M-27 SHV-12 TEM-52 CTX-M-2
0%
20%
40%
60%
80%
100%
Total (n=24 ) UpstreamWWTP (n =7)
WWTPeffluent (n=3)
Dischargepoint (n=4)
Recrea�onalwater (n=10 )
%Iso
lates
Region D
CTX-M-1 CTX-M-15 CTX-M-14 CTX-M-27 SHV-12
n=144
ig. 5. ESBL-genes detected in ESBL-producing E. coli from Dutch wastewater and surface water. Shown are the overall distribution of ESBL-genotypes in 144
olates from wastewaters and different surface waters (A) and the distribution of ESBL-genotypes per sampling region and sample type (B). N.U.I. = not
nder influence of the WWTP studied. Each indicated genotype represents a cluster of closely related, on the basis of partial sequence indistinguishable,
ariants: ‘CTX-M-1’ = CTX-M-1/61, ‘CTX-M-2’ = CTX-M-2/20/44/56/59/97, ‘CTX-M-3’ = CTX-M-3/22/66, ‘CTX-M-15’ = CTX-M-15/28, ‘CTX-M-32’ = CTX-M-
2, ‘CTX-M-9’ = CTX-M-9/51, ‘CTX-M-14’ = CTX-M-14/21/83/90/113, ‘CTX-M-27’ = CTX-M-27/98, ‘CTX-M-24’ = CTX-M-24/65, ‘SHV-12’ = SHV-12/129,
EM-52’ = TEM-52/92.

Fig. 6. Phylogenetic groups and ESBL-genes of ESBL-producing E. coli. Shown are the numbers of isolates with indicated ESBL genotypes in phylogenetic
backgrounds for different samples and for each region. Isolates from wastewater and from surface water at the discharge points have been pooled. *For
region C, surface water upstream of recreational waters encompasses surface waters upstream of WWTP as well as surfacewaters not under influence of the
studied WWTP.
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459456

H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 457
onto the lake (unless in situations of extreme rainfallcausing spill-over of the dam).
All WWTP effluents contained ESBL-producing E. coli, asdid receiving surface waters at the points of discharge. Atthese sites, ESBL-producing E. coli isolates can be assumedto directly reflect the bacterial population present indischarged effluents at that moment of sampling, althougha small proportion of isolates may have been derived fromupstream locations. In line with this assumption, concen-trations of ESBL-producing E. coli at discharge points weresimilar to that in effluents, and on average 1- to 2-log 10
units higher than concentrations upstream of the WWTPs,or in connecting water bodies not influenced by theinvestigated WWTPs. Considering an average concentra-tion of 2 � 102–5 � 102 ESBL-producing E. coli in 100 ml ofeffluent, and an average daily discharge of 10,000–35,000 m3 of effluent by the investigated WWTPs, approx-imately 1010–1011 ESBL-producing E. coli are dischargeddaily per WWTP. Even though these findings demonstratethat WWTPs contribute to the presence of ESBL-producingE. coli in surface water, ESBL-producing E. coli were alsofrequently detected upstream of the WWTPs, and inconnecting water bodies not influenced by the studiedWWTPs, implicating the existence of additional sources ofESBL-producing E. coli. Moreover, since ESBL-producing E.
coli were detected in multiple water bodies and in differentregions in the Netherlands, these data suggest thatcontamination sources are abundant. However, in arecreational water located within approximately 200 mof, but not connected to, recreational water D, ESBL-producing E. coli were not detected at three time-pointsduring bathing season 2012 (data not shown), demon-strating that at least some surface waters are onlyinfrequently, if at all, exposed to ESBL-producing E. coli
– containing fecal contamination sources.The current study focused on discharged effluents of
nearby located WWTPs as possible source of ESBL-producing E. coli in recreational waters, but did notinvestigate the contribution of sewage overflows or moreremote WWTPs. Since overflows contain untreated sew-age, they may be an important source of ESBL-producing E.
coli in surface water during heavy rainfall. Locations ofoverflow exhausts in the investigated regions were notmapped, however, both overflows and more remoteWWTPs may have contributed to the fecal contaminationin the investigated surface waters, in recreational waters aswell as at the other sampling sites. Additionally, animalmanure and droppings from pasture animals may havecontributed, since ESBL-producing E. coli are abundant inDutch food animals, in particular in broilers, but also inpigs and veal calves (Nethmap_MARAN, 2013). Finally, alsofeces of wild animals such as birds, may contribute toESBL-producing E. coli in surface water (Guenther et al.,2011). With the exception of region C, all investigatedregions are located in rural areas, making it probable thatthe ESBL-producing E. coli in the investigated surfacewaters are of mixed human and animal origin.
ESBL-producing E. coli recovered from recreationalwaters carried similar ESBL-genes, partially in the samephylogenetic background, as ESBL-producing E. coli ineffluents and/or upstream located surface waters. MostT
ab
le2
Ch
ara
cte
rist
ics
of
sets
of
ide
nti
cal
iso
late
so
bta
ine
dfr
om
dif
fere
nt
sam
ple
sin
the
sam
ere
gio
n.
Re
gio
nT
ime
-po
int
Ph
ylo
ge
ne
tic
(su
b)g
rou
p
ES
BL
ge
ne
No
n-E
SB
LT
EM
/OX
A
ge
ne
a
MLS
TA
BR
Pro
file
Up
stre
am
1b
Up
stre
am
2b
WW
TP
Dis
cha
rge
po
int
Re
cre
ati
on
wa
ter
SW
n.u
.i
A1
A1
CT
X-M
-1T
EM
ST
61
7A
m,
Cx
,G
e,
St,
Te
,C
i,N
a,
Su
,T
r+
++
+�
n.s
.
A1
A1
CT
X-M
-15
OX
AS
T6
17
Am
,C
x,
Cz,
Ge
,T
e,
Ci,
Na
,S
u,
Tr
�+
++
+n
.s.
A1
B1
CT
X-M
-14
�S
T8
6A
m,
Cx
,S
t,T
e,
Ci,
Na
,S
u,
Tr,
Ch
+�
+�
�n
.s.
A1
D1
CT
X-M
-15
TE
M,
OX
AS
T6
9A
m,
Cx
,C
z,S
t,T
e,
Su
,T
r+
+�
��
n.s
.
A1
B1
CT
X-M
-1T
EM
ST
58
Am
,C
x,
St,
Te
,S
u,
Tr
+�
+�
�n
.s.
A2
A1
CT
X-M
-15
�S
T9
0A
m,
Cx
,C
z,C
i,N
a�
+�
+�
n.s
.
A2
D1
CT
X-M
-15
TE
MS
T1
72
2A
m,
Cx
,C
z,G
e+
�+
�+
n.s
.
A2
A1
CT
X-M
-15
OX
AS
T6
17
Am
,C
x,
Cz,
Ge
,T
e,
Ci,
Na
,S
u,
Tr
++
++
�n
.s.
A2
A0
CT
X-M
-1�
ST
16
0A
m,
Cx
,S
u,
Tr
�+
��
+n
.s.
C1
A1
CT
X-M
-15
OX
AS
T3
61
Am
,C
x,
Cz,
Ax
,G
e,
Ci,
Na
,S
u,
Tr�
n.s
.+
+�
�C
1A
1C
TX
-M-1
5�
ST
10
Am
,C
x,
Na
�n
.s.
++
��
C3
A1
CT
X-M
-15
TE
MS
T2
21
1A
m,
Cx
,C
z,S
t,T
e,
Su
,T
r�
n.s
.+
+�
�C
3B
1C
TX
-M-1
5�
Un
kn
ow
nc
Am
,C
x,
Cz,
Te
,S
u,
Tr
�n
.s.
++
��
D2
A1
CT
X-M
-15
OX
AS
T6
17
Am
,C
x,
Cz,
Te
,C
i,N
a,
Su
,T
r�
n.s
.+
+�
n.s
.
D2
A1
CT
X-M
-1�
ST
76
1A
m,
Cx
+n
.s.
��
+n
.s.
aId
en
tifi
ed
usi
ng
PC
R,
typ
es
we
ren
ot
de
term
ine
d.
bFo
rre
gio
nA
two
site
su
pst
rea
mW
WT
Pw
ere
sam
ple
d(s
ee
Fig
.1
).c
All
ele
nu
mb
ers
for
ho
use
ke
ep
ing
ge
ne
sa
dk
,fu
mC
,g
yrB
,ic
d,
md
h,
pu
rA,
recA
we
re6
,1
33
,1
2,
1,
9,
5,
an
d7
resp
ect
ive
ly.
n.s
.=n
ot
sam
ple
d;
SW
n.u
.i=
wa
ter
no
tu
nd
er
the
infl
ue
nce
of
stu
die
dW
WT
P,
Am
=a
mp
icil
lin
,C
x=
cefo
tax
ime
,C
z=
ceft
azi
dim
e,
Ge
=g
en
tam
icin
,S
t=
stre
pto
my
cin
,T
e=
tetr
acy
clin
e,
Ci=
cip
rofl
ox
aci
n,
Na
=n
ali
dix
ica
cid
,S
u=
sulf
ox
izo
le,
Tr
=tr
ime
tho
pri
m.

notimdebecirco
thwphsevadastrWmAldicodisuSTDuor2020
buincorepridmhophdeconumsaphthanisocofloandiunWoffombeth(PPCNe
co
re
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459458
tably, infrequently detected ESBL-genotypes – some-es exclusively observed in certain regions – were also
tected in recreational waters, suggesting a relationtween variants in recreational water and variantsculating in the region. In four cases, ESBL-producing E.
li variants from recreational waters were identical toose in wastewater and/or upstream located surfaceater sites obtained on the same day, with respect toylogenetic group, ESBL-genotype ABR-profiles andquence types. In two of these four cases this concernedriants from recreational water and wastewater. Theseta may indicate that these four ESBL-producing E. coli
ains present in recreational waters originated fromWTP effluents or from a source further upstream, andigrated to the recreational waters with the current.ternatively, identical ESBL-producing variants present atfferent sites may originate from different sources, whichuld be the case when specific variants are widelyspersed among humans and/or animals. Some of therface water and effluent E. coli sequence types, i.e. ST10,58, ST69 and ST617, have previously been detected intch human and animal clinical isolates obtained, either
not associated with the same ESBL-genes (Dierikx et al.,12; Leverstein-van Hall et al., 2011; Overdevest et al.,11; Reuland et al., 2013).Even though the current study demonstrates a contri-tion of WWTPs to the presence of ESBL-producing E. coli
surface waters, their relative contribution to thentamination of the recreational waters under studymains undetermined. Only a small number of ESBL-oducing E. coli variants from recreational waters hadentical counterparts in effluents or at discharge points. Inost samples including recreational water samples,wever, a high diversity of ESBL-producing E. coli
enotypes was observed. As a consequence, the odds oftecting copy isolates from two different places innnecting water bodies is small and dependent on thember of isolates that are analyzed. In the current study
aximally 10 isolates were isolated and characterized permple, which considering the observed large variation inenotypes, may have resulted in an underestimation ofe level of accordance between isolates from wastewaterd recreational waters. Analysis of a larger number oflates per site, mapping and sampling of additional
ntamination sources in the investigated area (i.e. over-ws, other, more remote WWTPs, and animal manure),d taking into account current velocities betweenscharge points and recreational waters, are essential toequivocally determine the relative contributions ofWTPs and other contamination sources to the presence
ESBL-producing E. coli in recreational waters. Moreover,r the purpose of tracing variants to specific sources, aore thorough characterization of isolates would need to
considered, including an additional method for typing ofe E. coli strains, such as pulsed-field gel electrophoresisFGE), and analysis of the ESBL-carrying plasmids usingR based replicon typing (PBRT) (Carattoli et al., 2005;moy et al., 2005).Based on obligatory measurements of fecal indicators E.
li and intestinal enterococci of 2009–2011 in official
according to guidelines described in the European BathingWater Directive (Anonymous, 2006), the water quality ofthe three official recreational waters was classified as‘poor’ (region B), ‘acceptable’ (region A), or ‘good’ (regionC). As of 2013, ‘signal values’ are used to assess individualsamples for compliance; the signal value for E. coli is set at1800 cfu/100 ml. Based on this value and our 2011 and2012 measurements, non-compliance was observed onlyat one time-point in recreational waters A and B (both in2011). So, multidrug resistant ESBL-producing E. coli weredetected in recreational waters classified from ‘poor’ to‘good’, at time-points when recreational waters compliedwith bathing water legislation.
The rationale for investigation of ESBL-producing E. coli
in surface and recreational waters is twofold. Besidesdetermining the relative contribution of different contam-ination sources for the purpose of intervention strategies, itis necessary to determine the contribution of recreationalwater (or other environment-related exposure) to theoverall community-associated exposure to ESBL-produc-ing bacteria, relative to that of consumption of contami-nated food or direct contact with animal or human carriers.Considering that with the exception of people working atfarms, people may have a better opportunity to come intocontact with contaminated recreational water than withlivestock, the detection of ESBL-producing E. coli inapproved recreational waters suggests that recreationalwaters should be considered as a potential exposure routeof these bacteria.
Conflict of interest
None to declare.
Role of the funding source
The funding source has had no involvement in studydesign, the collection, analysis and interpretation of data,the writing of the manuscript, or the decision to submit forpublication.
Acknowledgements
The authors are thankful to Arieke Docters vanLeeuwen, Gretta Lynch, and Christiaan Veenman forexcellent technical assistance, to the water boards‘Wetterskip Fryslan’, ‘Hoogheemraadschap Rijnland’,‘Waterschap Rijn en IJssel’, and ‘Rijkswaterstaat IJssel-meergebied’, for providing essential information regardingWWTPs and recreational waters in the respective areas,and to the people working at the WWTPs for providingeffluent samples. This study was financed by the DutchMinistery of Infrastructure and the Environment.
References
Alouache, S., Estepa, V., Messai, Y., Ruiz, E., Torres, C., Bakour, R., 2014.Characterization of ESBLs and associated quinolone resistance inEscherichia coli and Klebsiella pneumoniae isolates from an urban
wastewater treatment plant in Algeria. Microb. Drug Resist. 20,30–38. creational waters performed by water boards, and
A
A
A
A
B
C
C
C
C
C
C
C
D
D
D
D
D
H. Blaak et al. / Veterinary Microbiology 171 (2014) 448–459 459
nastasi, E.M., Matthews, B., Stratton, H.M., Katouli, M., 2012. PathogenicEscherichia coli found in sewage treatment plants and environmentalwaters. Appl. Environ. Microbiol. 78, 5536–5541.
nonymous, 2000. NEN-EN-ISO 9308-1 Water – Detectie en enumeratievan Escherichia coli en bacterien van de coligroep – Deel 1: Methodemet membraanfiltratie.
nonymous, 2006. Directive 2006/7/EC of the European Parliament and ofthe Council of 15 February 2006 concerning the management ofbathing water quality and repealing directive 76/160/EEC OfficialJournal of the European Union L64, pp. 37–51.
nonymous, 2007. NEN-EN-ISO 19458. Water quality – Sampling formicrobiological analysis.
laak, H., van Rooyen, S.R., Schuijt, M.S., Docters van Leeuwen, A.E., van denBerg, H.H.J.L., Lodder-Verschoor, F., Italiaander, R., Schets, F.M., de RodaHusman, A.M., 2011. Prevalence of antibiotic resistant bacteria in therivers Meuse, Rhine and New Meuse. RIVM report 703719071.
anton, R., Novais, A., Valverde, A., Machado, E., Peixe, L., Baquero, F.,Coque, T.M., 2008. Prevalence and spread of extended-spectrum ß-lactamase-producing Enterobacteriaceae in Europe. Clin. Microbiol.Infect. 14 (Suppl. 1), 144–153.
anton, R., Akova, M., Carmeli, Y., Giske, C.G., Glupczynski, Y., Gniad-kowski, M., Livermore, D.M., Miriagou, V., Naas, T., Rossolini, G.M.,Samuelsen, O., Seifert, H., Woodford, N., Nordmann, P., 2012. Rapidevolution and spread of carbapenemases among Enterobacteriaceaein Europe. Clin. Microbiol. Infect. 18, 413–431.
arattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K.L., Threlfall, E.J., 2005.Identification of plasmids by PCR-based replicon typing. J. Microbiol.Methods 63, 219–228.
astanheira, M., Mendes, R.E., Rhomberg, P.R., Jones, R.N., 2008. Rapidemergence of blaCTX-M among Enterobacteriaceae in U.S. MedicalCenters: molecular evaluation from the MYSTIC Program (2007).Microb. Drug Resist. 14, 211–216.
hen, H., Shu, W., Chang, X., Chen, J.A., Guo, Y., Tan, Y., 2010. The profileof antibiotics resistance and integrons of extended-spectrum beta-lactamase producing thermotolerant coliforms isolated from theYangtze River basin in Chongqing. Environ. Pollut. 158, 2459–2464.
lermont, O., Bonacorsi, S., Bingen, E., 2000. Rapid and simple determi-nation of the Escherichia coli phylogenetic group. Appl. Environ.Microbiol. 66, 4555–4558.
linical and Laboratory Standards Institute, 2010. Performance Standardsfor Antimicrobial Susceptibility Testing; Twentieth InformationalSupplement M100-S20. CLSI, Wayne, PA, USA.
allenne, C., Da Costa, A., Decre, D., Favier, C., Arlet, G., 2010. Developmentof a set of multiplex PCR assays for the detection of genes encodingimportant b-lactamases in Enterobacteriaceae. J Antimicrob Che-mother 65, 490–495.
eng, Y., Zeng, Z., Chen, S., He, L., Liu, Y., Wu, C., Chen, Z., Yao, Q., Hou, J.,Yang, T., Liu, J.H., 2011. Dissemination of IncFII plasmids carryingrmtB and qepA in Escherichia coli from pigs, farm workers and theenvironment. Clin. Microbiol. Infect. 17, 1740–1745.
hanji, H., Murphy, N.M., Akhigbe, C., Doumith, M., Hope, R., Livermore,D.M., Woodford, N., 2011. Isolation of fluoroquinolone-resistantO25b:H4-ST131 Escherichia coli with CTX-M-14 extended-spectrumß-lactamase from UK river water. J. Antimicrob. Chemother. 66, 512–516.
ierikx, C.M., van Duijkeren, E., Schoormans, A.H., van Essen-Zandbergen,A., Veldman, K., Kant, A., Huijsdens, X.W., van der Zwaluw, K.,Wagenaar, J.A., Mevius, D.J., 2012. Occurrence and characteristicsof extended-spectrum-beta-lactamase- and AmpC-producing clinicalisolates derived from companion animals and horses. J. Antimicrob.Chemother. 67, 1368–1374.
uriez, P., Clermont, O., Bonacorsi, S., Bingen, E., Chaventre, A., Elion, J.,Picard, B., Denamur, E., 2001. Commensal Escherichia coli isolates arephylogenetically distributed among geographically distinct humanpopulations. Microbiology 147, 1671–1676.
Escobar-Paramo, P., Le Menac’h, A., Le Gall, T., Amorin, C., Gouriou, S.,Picard, B., Skurnik, D., Denamur, E., 2006. Identification of forcesshaping the commensal Escherichia coli genetic structure by compar-ing animal and human isolates. Environ Microbiol 8, 1975–1984.
Guenther, S., Ewers, C., Wieler, L.H., 2011. Extended-spectrum beta-lactamases producing E. coli in wildlife, yet another form of environ-mental pollution? Front Microbiol 2, 246.
Hong, H., Chun, J., Lee, Y., 2004. Detection of extended-spectrum b-lacamase-producing, multidrug-resistant environmental isolated ofEscherichia coli that bind to human bladder cells. Microb. Drug Resist.10, 184–189.
Huijbers, P.M., de Kraker, M., Graat, E.A., van Hoek, A.H., van Santen, M.G.,de Jong, M.C., van Duijkeren, E., de Greeff, S.C., 2013. Prevalence ofextended-spectrum beta-lactamase-producing Enterobacteriaceae inhumans living in municipalities with high and low broiler density.Clin. Microbiol. Infect. 19, E256–E259.
Jacoby, G.A., 2009. AmpC ß-lactamases. Clin. Microbiol. Rev. 22, 161–182.Jakobsen, L., Spangholm, D.J., Pedersen, K., Jensen, L.B., Emborg, H.D.,
Agerso, Y., Aarestrup, F.M., Hammerum, A.M., Frimodt-Moller, N.,2010. Broiler chickens, broiler chicken meat, pigs and pork as sourcesof ExPEC related virulence genes and resistance in Escherichia coliisolates from community-dwelling humans and UTI patients. Int. J.Food Microbiol. 142, 264–272.
Johnson, J.R., Delavari, P., Kuskowski, M., Stell, A.L., 2001. Phylogeneticdistribution of extraintestinal virulence-associated traits in Escher-ichia coli. J. Infect. Dis. 183, 78–88.
Leverstein-van Hall, M.A., Dierikx, C.M., Cohen Stuart, J., Voets, G.M., vanden Munckhof, M.P., van Essen-Zandbergen, A., Platteel, T., Fluit, A.C.,van de Sande-Bruinsma, N., Scharinga, J., Bonten, M.J., Mevius, D.J.,2011. Dutch patients, retail chicken meat and poultry share the sameESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880.
Livermore, D.M., Canton, R., Gniadkowski, M., Nordmann, P., Rossolini,G.M., Arlet, G., Ayala, J., Coque, T.M., Kern-Zdanowicz, I., Luzzaro, F.,Poirel, L., Woodford, N., 2007. CTX-M: changing the face of ESBLs inEurope. J. Antimicrob. Chemother. 59, 165–174.
Nemoy, L.L., Kotetishvili, M., Tigno, J., Keefer-Norris, A., Harris, A.D.,Perencevich, E.N., Johnson, J.A., Torpey, D., Sulakvelidze, A., MorrisJr., J.G., Stine, O.C., 2005. Multilocus sequence typing versus pulsed-field gel electrophoresis for characterization of extended-spectrumbeta-lactamase-producing Escherichia coli isolates. J. Clin. Microbiol.43, 1776–1781.
Nethmap_MARAN 2013, 2013. Monitoring of antimicrobial resistanceand antibiotic usage in animals in the Netherlands in 2010/2011.www.swab.nl.
Overdevest, I., Willemsen, I., Rijnsburger, M., Eustace, A., Xu, L., Hawkey,P., Heck, M., Savelkoul, P., Vandenbroucke-Grauls, C., van der Zwaluw,K., Huijsdens, X., Kluytmans, J., 2011. Extended-spectrum beta-lacta-mase genes of Escherichia coli in chicken meat and humans, TheNetherlands. Emerg. Infect. Dis. 17, 1216–1222.
Paterson, D.L., Bonomo, R.A., 2005. Extended-spectrum beta-lactamases:a clinical update. Clin. Microbiol. Rev. 18, 657–686.
Picard, B., Garcia, J.S., Gouriou, S., Duriez, P., Brahimi, N., Bingen, E., Elion,J., Denamur, E., 1999. The link between phylogeny and virulence inEscherichia coli extraintestinal infection. Infec. Immun. 67, 546–553.
Reuland, E.A., Overdevest, I.T., Al Naiemi, N., Kalpoe, J.S., Rijnsburger, M.C.,Raadsen, S.A., Ligtenberg-Burgman, I., van der Zwaluw, K.W., Heck, M.,Savelkoul, P.H., Kluytmans, J.A., Vandenbroucke-Grauls, C.M., 2013.High prevalence of ESBL-producing Enterobacteriaceae carriage inDutch community patients with gastrointestinal complaints. Clin.Microbiol. Infect. 19, 542–549.
Trott, D., 2013. Beta-lactam resistance in gram-negative pathogens iso-lated from animals. Curr. Pharm. Des. 19, 239–249.
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L.H., Karch, H.,Reeves, P.R., Maiden, M.C., Ochman, H., Achtman, M., 2006. Sex andvirulence in Escherichia coli: an evolutionary perspective. Mol. Micro-biol. 60, 1136–1151.





























