Embed Size (px)
Citation preview

Drought resistance and recovery mechanisms of tedera
(Bituminaria bituminosa var. albomarginata)
Kevin Foster
This thesis is presented for the degree of
Doctor of Philosophy
The University of Western Australia
School of Plant Biology
Faculty of Science
March 2015


PAGE | i
STATEMENT OF ORIGINAL CONTRIBUTION
The research presented in this thesis is an original contribution to the field of plant
ecophysiology. The hypotheses and results are my original ideas and writing. However, this
work could not have been achieved without the contribution of my supervisors who share
authorship on the manuscripts. Their contributions are acknowledged in each preface of the
experimental chapters.
This thesis has been completed during the course of my enrolment for a PhD degree at The
University of Western Australia, and has not been used previously for a degree or a diploma at
any other institution.
Primary authored manuscripts:
Foster K, Ryan M, Real D, Ramankutty P, Lambers H (2012) Drought resistance at the seedling
stage in the promising fodder plant tedera (Bituminaria bituminosa var. albomarginata). Crop
and Pasture Science 63, 1034–1042.
Foster K, Ryan M, Real D, Ramankutty P, Lambers H (2013) Seasonal and diurnal variation in
stomatal conductance and paraheliotropism of tedera (Bituminaria bituminosa var.
albomarginata L.) in the field. Functional Plant Biology 40, 719–729.
Foster K, Lambers H, Real D, Ramankutty P, Cawthray GR, Ryan M (2014) Drought resistance
and recovery in mature Bituminaria bituminosa (L.) C.H. Stirton. var. albomarginata. Accepted
Annals of Applied Biology. Published online October 30th 2014.
Kevin Foster
March 2015

PAGE | ii

PAGE | iii
ACKNOWLEDGEMENTS
My deepest gratitude to my primary supervisors: Associate Professor Megan Ryan for her
support and expert advice while working towards this thesis and for understanding the
challenges that can come with doing a PhD later in life. I would like to thank Professor Hans
Lambers for his critical appraisal of my work, his patience and the humorous emails at all hours
of the night, often across international time zones. I don’t know how you find the time Hans to
achieve so much and so well. Thanks must also go to Drs Mike Ewing and Kevin Goss from the
Future Farm Industries CRC for their help and financial support, and also to Scott Glyde, a
talented educator. I wish to thank Dr. Daniel Real for introducing me to a truly remarkable
plant in tedera and for having the confidence in me to unravel some of its secrets. I would like
to acknowledge Dr. Mark Sweetingham from DAFWA for supporting my study leave application
and for his encouragement and Professor John Kuo from the Centre for Microscopy,
Characterisation and Analysis (CMCA) for his generosity on the instruction of sample
preparation, a true gentleman and a scholar. Special acknowledgment must also go to Ben
Biddulph from DAFWA, one of our best and brightest young research leaders, who loaned rain-
exclusion shelters for my field experiments and was always generous with his time and advice.
Special thanks go to my fellow PhD students in the School of Plant Biology, Lalith Suriyagoda
and Padmaja Ramankutty, and to Professors Neil Turner and Erik Veneklaas for their
professional interest and encouragement during all aspects of this project. To Dion Nicol - few
students I have met could ever hope to undertake the PhD you embarked on; I hope you stay
in agricultural research, because we need your practical insights. I would like to especially
thank Dr. Clinton Revell from DAFWA, my manager, for all his valuable support, advice, and
understanding during my PhD. To Drs Phil Nichols, Ron Yates, Bradley Nutt, Angelo Loi, and to
Darryl McClements, Geoff Moore and Mike Davies from the Pasture Science group at DAFWA,
thanks for all your great support regarding various aspects of this project. It is no exaggeration
to say you are the best legume researchers in dryland agriculture on the planet. To David Budd
from transport section at DAFWA, thank you for locating “spare and cheap” vehicles, so that I
could travel to my field sites, over 45,000 kms in two years. I would like to thank my colleague
Daniel Kidd, one of the most organised and positive people I have had the pleasure to work
with. To the overworked Brad Wintle, thanks for your great sense of humour that you bring to
work every day. To Peter Skinner, Darryl McClements and Mark Holland, thank you for your
professional support and friendship over many, many years.

PAGE | iv
Sincere thanks are due to Rob Creasy and Bill Piasini from UWA and Ross Ramm from DAFWA
for their technical, professional and personal help that they provided while I was a student at
UWA. I would like to especially acknowledge the terrific support of the staff in the School of
Plant Biology for their help with administrative tasks, which are always done with a smile.
I would like to especially thank the late Professor Clive Francis (1938–2012 RIP), for without
him planting that small seed that one day—“You should actually do a PhD”—I would not have
contemplated this journey when the opportunity arose. I only wish you were here to see it
completed Clive.
I would like to thank my two sons, Liam and Patrick Foster who have been very understanding
and supportive over several years. We have some serious game time to catch up on, boys.
Also, I must thank my parents (Michael Foster 1927-1985 RIP), especially my amazing mother,
Mary Agnes Foster, who always believed in and stressed the importance of higher education to
all her 10 children, and went without much over many years so we could all have one. Finally,
there is my wife, Donna Chalwell. I have written many thousands of words in this thesis, and
read millions more of others. But for once, I am lost for words. This thesis simply would not
have been possible without your unwavering support, encouragement and your absolute
belief in me.

PAGE | v
ABSTRACT
Lucerne (Medicago sativa L.) often underperforms agronomically in Western Australia (WA),
due to its sensitivity to acidic soils and the prolonged period of water stress during summer,
which is typical of the WA Mediterranean climate. The development of new perennials
adapted to acid soils and low rainfall is a high priority for future research in the Australian
wheatbelt. Tedera (Bituminaria bituminosa var. albomarginata) is a recently-introduced
herbaceous perennial legume that has shown potential as a drought-resistant forage species
for Mediterranean environments across southern Australia and is currently being evaluated for
domestication. Tedera is native to the Canary Islands of Spain and, on the island of Lanzarote,
tedera can survive on as little as 150 mm per year, with a 5–6 month dry season.
There have been no detailed investigations into drought adaptation and physiological traits of
tedera in the field that have allowed this species to persist and be productive over summer.
The mechanisms of drought resistance in legumes have not been characterised as well as in
cereals. The objective of this thesis was to examine the drought resistance mechanisms of
tedera under both field and controlled-environment conditions by comparing the response of
tedera with lucerne. It is proposed that the drought resistance mechanisms of tedera are a
combination of several morphological and physiological processes such as osmotic adjustment,
an extensive root system, efficient stomatal control and paraheliotropism (leaf folding).
Drought stress was initiated at different growth stages (seedling and mature plants) by
withholding water in the controlled-environment study and by erecting a rain-exclusion shelter
over summer in the field. Parameters relating to aboveground dry matter production, root
growth, water-use efficiency, leaf-water relations, gas exchange, leaf folding, and individual
leaf size were measured. The concentration of compatible solutes was determined for mature
tedera plants in a controlled-environment study during both drought stress and recovery.
The ability of seedlings to survive a drought following sowing or seedling germination in a
regenerating pasture is important for successful pasture establishment/persistence. Thus, the
first experiment (Chapter 2) was conducted under controlled-environment conditions (using
soil reconstructed from the field) to investigate the physiological and morphological responses
to water stress of tedera seedlings compared with lucerne, the annual pasture legume
biserrula (Biserrula pelecinus L.) and the common summer-active weed Afghan melon (Citrullus
lanatus Thunb). This study revealed significant differences in rooting depth and stomatal
conductance among the three legume species when under water stress, with tedera being the
most drought resistant. Key drought resistance traits in tedera included vigorous seedling

PAGE | vi
growth, early taproot elongation, effective stomatal control and paraheliotropism. These traits
may allow tedera seedlings to survive a dry period following opening autumn or winter rains.
The abiotic stresses of the WA wheatbelt (low pH, aluminum toxicity, nutrient deficiencies)
and the low water-holding capacity of many soils could potentially impact the drought
resistance and survival of tedera. The second experiment (Chapter 3), conducted in the field in
the southern wheatbelt of WA, explored diurnal and seasonal variation (May, August and
February) in stomatal conductance and paraheliotropism in mature plants (from 11–18 months
old) of three tedera accessions and one lucerne cultivar. Tedera exhibited greater stomatal
conductance early in the morning in May and December, but lower conductance at midday
and during the afternoon. The reduction in stomatal conductance over the day in tedera shows
the capacity of this species to reduce water loss quickly when conditions for CO2 fixation
relative to water loss are highly unfavourable. Leaf water-use efficiency, relative leaf water
content and, importantly, green leaf retention were higher for tedera than for lucerne under
severe water deficit. Both tedera and lucerne displayed increased paraheliotropism over
autumn and summer, suggesting that this is also an adaptive trait to cope with water stress.
With the fluctuation in rainfall patterns predicted from climate change, the occurrence of
episodic drought may increase where plants will be repeatedly exposed to drought in the field.
However, the combined responses to water stress and re-watering among perennial legumes
are relatively unknown. The third experiment (Chapter 4) investigated the response of two
tedera accessions and one lucerne cultivar to drought stress and their ability, following re-
watering, to regrow and restore photosynthetic activity. Plants were grown outdoors and
watered for eight months (winter to mid-summer), and then moved to a controlled-
environment glasshouse where either watering was maintained or drought was imposed for
up to 47 days, before re-watering for 28 days. After re-watering, the previously drought-
stressed plants had higher photosynthetic rates and stomatal conductance than the well-
watered controls, indicating an overcompensation of gas exchange. However, the decline in
aboveground dry matter due to long-term severe drought stress was not completely replaced
by this enhanced growth following re-watering. Proline and pinitol accumulated in tedera
leaves during drought stress and their concentration declined after re-watering, suggesting
that they act as compatible solutes during drought stress and as a carbon source for regrowth
during recovery. Root systems in tedera also had greater plasticity (higher root:shoot ratio) in
response to drought stress and re-watering than lucerne.
Considering the results of the three experiments together, it is apparent that tedera and
lucerne have different physiological and morphological strategies to survive and recover from

PAGE | vii
drought stress. Significantly, the retention of green leaves in tedera over summer in the field
would be a highly attractive trait to farmers, as this could reduce the need for supplementary
feeding to meet animal nutritional requirements. This study has contributed to the
understanding of stress biology of plants by investigating the morphological and physiological
traits that confer drought resistance in tedera and identifying the role of proline and pinitol in
both drought tolerance and recovery. The presence of multiple drought resistance traits in
tedera highlights the value of examining whole-plant physiology in any new perennial legume
species.
“Education is the kindling of a flame, not the filling of a vessel.” Socrates.

PAGE | viii

PAGE | ix
TABLE OF CONTENTS
STATEMENT OF ORIGINAL CONTRIBUTION I
ACKNOWLEDGEMENTS III
ABSTRACT V
TABLE OF CONTENTS IX
LIST OF FIGURES XIII
LIST OF TABLES XV
LIST OF PLATES XVI
CHAPTER 1. GENERAL INTRODUCTION 1
THE NEED FOR NEW PERENNIAL LEGUMES 1
THE DEVELOPMENT OF NEW DEEP-ROOTED PERENNIALS ADAPTED TO LOW-TO-MEDIUM RAINFALL 2
NATIVE ENVIRONMENT OF TEDERA 4
DROUGHT RESISTANCE MECHANISMS 5
TEDERA AND ITS POTENTIAL RELEVANCE TO SOUTHERN AUSTRALIAN FARMING SYSTEMS 6
FOCUS OF PHD RESEARCH ON DROUGHT RESISTANCE MECHANISMS IN TEDERA 7
THESIS METHODOLOGICAL APPROACH 9
THESIS AIMS 11
THESIS STRUCTURE 12
CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN THE PROMISING FODDER PLANT TEDERA (BITUMINARIA BITUMINOSA VAR. ALBOMARGINATA) 13
PREFACE 14
ABSTRACT 15
INTRODUCTION 15
MATERIALS AND METHODS 16
Measurements 17
Statistical analysis 17
RESULTS 17
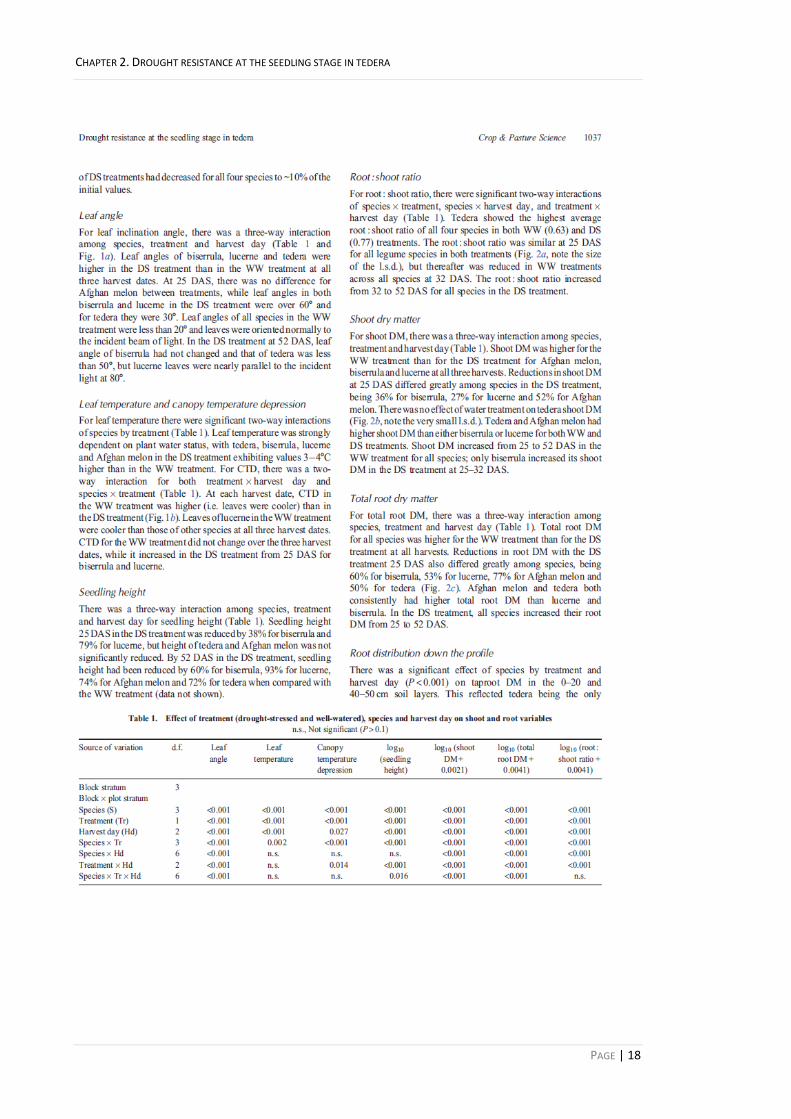
Plant water use 17
Leaf angle 18
Leaf temperature and canopy temperature depression 18
Seedling height 18
Root:shoot ratio 18
Shoot dry matter 18
Total root dry matter 18
Root distribution down the profile 18
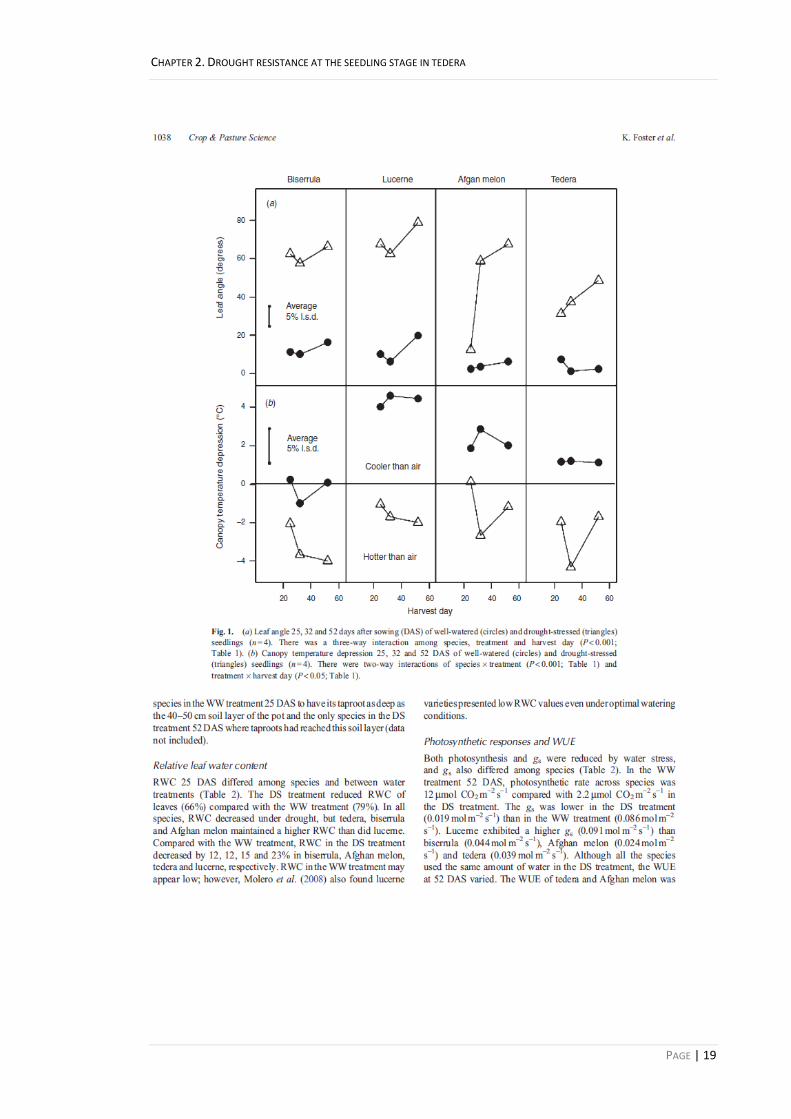
Relative leaf water content 19
Photosynthetic resposnes and WUE 19
DISCUSSION 20
Growth 20
Leaf movements 21
Dry matter allocation 21
Physiological adaptations to water stress 21
Agronomic implications 21
CONCLUDING REMARKS 22

PAGE | x
ACKNOWLEDGEMENTS 22
REFERENCES 22
SUPPORTING INFORMATION 24
CHAPTER 3. SEASONAL AND DIURNAL VARIATION IN THE STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA (BITUMINARIA BITUMINOSA VAR. ALBOMARGINATA) IN THE FIELD 27
PREFACE 28
ABSTRACT 29
INTRODUCTION 29
MATERIALS AND METHODS 30
Experimental site 30
Temperature and rainfall during the experiment 30
Photosynthesis and water-use efficieny 31
Stomatal conductance and leaf temperature 31
Leaf angle and individual leaf area 32
Dry matter production and leaf mass ratio 32
Statistical analysis 32
RESULTS 32
Photosynthesis and water-use efficiency 32
Dry matter production and LMR 32
RWC and leaf water potential 33
Stomatal conductance measure with leaf porometer 33
Leaf angle, individual leaf area and temperature 35
DISCUSSION 35
Physiological adaptations to water stress 35
Stomatal conductance (gs), relative leaf water content (RWC) and leaf water potential (LWP) 36
Morphological adaptation to water stress 36
Adaptations to native environment and concluding comments 37
ACKNOWLEDGEMENTS 37
REFERENCES 37
SUPPORTING INFORMATION 40
CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE BITUMINARIA BITUMINOSA (L.) C.H. STIRTON. VAR. ALBOMARGINATA. 43
PREFACE 44
ABSTRACT 45
INTRODUCTION 45
MATERIALS AND METHODS 46
Measurements 48
Plant water use 48
Photosynthesis and instantaneous leaf water-use efficieny 48
Relative leaf water content, pre-dawn and midday leaf water potential 48
Photosynthetically active radiation 48
Osmotic adjustment 48
Compatible solutes and sugars 49
Ethanol extraction of compatible solutes, soluble sugars and sugar ethanols from plant tissues 49

PAGE | xi
Shoot and root growth characters 49
Individual leaf angle and plant height 50
Statistical analysis 50
RESULTS 50
Soil water 50
Photosynthesis and stomatal conductance 50
Intrinsic leaf water-use efficiency 51
Relative leaf water content 51
Pre-dawn and midday leaf water potential 51
Other plant growth responses 52
Leaf angle and photosynthetically active radiation 52
Plant height and pubescence 53
Plant growth 54
Total shoot dry matter and leaf to total mass ratio 54
Root dry matter production 54
Root to shoot ratio 54
Osmotic adjustment 54
Compatible solutes and sugars 54
DISCUSSION 55
Physiological and morphological responses to drought stress 56
Plant growth responses to drought stress 56
Total shoot and root dry matter production 56
Other aboveground plant growth responses 56
Root to shoot ratio 56
Response of compatible solutes and sugars to drought stress 56
Physiological traits associated with post-drought recovery 57
Root growth responses associated with post-drought recovery 57
Changes in compatible solutes and sugars associated with post-drought recovery 58
CONCLUDING COMMENTS 58
ACKNOWLEDGEMENTS 58
REFERENCES 58
SUPPORTING INFORMATION 61
CHAPTER 5. GENERAL DISCUSSION 65
INTRODUCTION 66
KEY FINDINGS FROM THIS RESEARCH 67
ADAPTIVE RESPONSES OF TEDERA SEEDLINGS TO DROUGHT STRESS 68
ADAPTIVE RESPONSES OF MATURE TEDERA PLANTS TO DROUGHT STRESS 69
Physiological responses to water stress 69
Changes in organic solutes in response to water deficit and re-watering 70
MORPHOLOGICAL RESPONSES TO WATER STRESS 71
Paraheliotropism 71
Individual leaf area and pubescence under drought stress 72
Root characteristics and growth responses of mature plants to drought stress 73
POST-DROUGHT RECOVERY ABILITY OF TEDERA AND LUCERNE 77
SPECIES-SPECIFIC MECHANISMS FOR TEDERA TO COPE WITH ITS NATIVE ENVIRONMENT 78
INTEGRATION OF PERENNIALS INTO SOUTHERN AUSTRALIAN FARMING SYSTEMS 81
The challenge of establishment 81

PAGE | xii
Shift in sowing dates and depth 82
COMPARING WATER STRESS IN THE FIELD AND CONTROLLED-ENVIRONMENT EXPERIMENTS 84
LIMITATIONS OF THE RESEARCH PRESENTED IN THIS THESIS 85
FUTURE RESEARCH DIRECTIONS 85
CONCLUDING COMMENTS 88
REFERENCES 90

PAGE | xiii
LIST OF FIGURES
Figure 2.1. (a) Leaf angle 25, 32 and 52 days after sowing (DAS) of well-watered (circles) and drought-stressed (triangles) seedlings (n = 4). There was a three-way interaction among species, treatment and harvest day (P < 0.001; Table 2.1). (b) canopy temperature depression 25, 32 and 52 DAS of well-watered (circles) and drought-stressed (triangles) seedlings (n = 4). There were two-way interactions of species × treatment (P < 0.001; Table 2.1) and treatment × harvest day (P < 0.05; Table 2.1). 19
Figure 2.2. (a) Root:shoot ratio, (b) total shoot dry matter, and (c) total root dry matter of well-watered (circles) and drought-stressed (triangles) seedlings (n = 3). There were two-way interactions of species × treatment, species × harvest date and treatment × harvest day (P < 0.001; Table 2.1) for root:shoot ratio. There were three-way interactions of species × treatment × harvest date (P < 0.001; Table 2.1) for both total root dry matter and total shoot dry matter. 20
Figure 3.1. Monthly weather data for Newdegate, Western Australia from February 2009 to February 2010: (a) solar radiation; (b) temperature; (c) rainfall. A rainfall shelter was in place from 3 December 2009 (early summer). 31
Figure 3.2. Box plot of field data to compare (a) dry matter production and (b) leaf mass ratio for lucerne and three accessions of tedera in February 2010 (late summer, 63 days without water). 33
Figure 3.3. Comparison of stomatal conductance for lucerne and three accessions of tedera in May 2009 (late autumn), August 2009 (late winter, well watered; see Figure 3.1) and February 2010 (late summer, 63 days without water). 34
Figure 3.4. Comparison of leaf angle (degrees) for lucerne and three accessions of tedera in May 2009 (late autumn), August 2009 (late winter, well watered; see Figure 3.1) and February 2010 (late summer, 63 days without water); 0o is horizontal. 35
Figure 3.5. Paraheliotropism in tedera in February 2010 (later summer 63 days without water). Also note the strong pubescence on leaf margins and petioles, and the folding of leaf lamina along the midrib. 35
Figure 4.1. Experimental design (one of four replicates shown). Plants were either well watered throughout the 75 days of the experiment or experienced drought for up to 47 days, before being rewatered for 28 days. Plants in harvest 1 (H1) were harvested after 33 days of drought; plants in harvest 2 (H2) were harvested after 47 days of drought followed by a recovery phase of 28 days of weekly rewatering. 47
Figure 4.2. Photosynthesis for two genotypes of tedera and one genotype of lucerne (n = 4, SED = 0.9455). There was a three-way interaction among treatment, rewatering and day (P < 0.001), but no significant effect of genotype. Lucerne plants at days 31 and 47 of the drought phase were at permanent wilting point and hence photosynthesis was zero. 51
Figure 4.3. (A) Stomatal conductance (n = 4, SED = 0.0390) and (B) intrinsic leaf water-use efficiency (n = 4, SED = 7.46) for two genotypes of tedera and one genotype of lucerne. For (A) and (B) there was a four-way interaction among genotype, treatment, rewatering and day (P < 0.001). Lucerne plants at days 31 and 47 of the drought phase were at permanent wilting point and hence gs and WUELwere zero. 52
Figure 4.4. Relative leaf water content for two genotypes of tedera and one genotype of lucerne (n = 4, SED = 0.033). There was a three-way interaction among treatment, rewatering and day (P < 0.001), but no significant effect of genotype. 53
Figure 4.5. Mean leaf angle for two genotypes of tedera and one genotype of lucerne (n = 4, SED = 4.19). There was a four-way interaction (P < 0.001) of genotype, treatment, rewatering and day. Note that 0o is horizontal. 53

PAGE | xiv
Figure 4.6. Root:shoot ratio for two tedera genotypes and one genotype of lucerne at day 33 and day 47 of the drought phase and day 28 of the recovery phase (n = 4, SED = 0.12). There was a three-way interaction of genotype, treatment and harvest (P < 0.001). 55
Figure 4.7. Pinitol and fructose concentrations in leaf tissue of tedera 4 at days 1 and 47 of the
drought phase and day 28 of the recovery phase (mean SE, n = 4). 55

PAGE | xv
LIST OF TABLES
Table 2.1. Effect of treatment (drought-stressed and well-watered), species and harvest day on shoot and root variables. 18
Table 2.2. The effect of species and treatment (drought-stressed and well-watered) on relative water content, photosynthesis response and stomatal conductance. 20
Table 3.1. Predicted means for the effects of time for photosynthesis measurements on 3 February 2010 (late summer, 63 days without water) for all three tedera accessions and lucerne. 33
Table 3.2. Predicted means by accession for the water-use efficiency of the leaf and relative leaf water content in tedera and lucerne. 33
Table 3.3. Predicted means by accession by month individual leaf area in tedera and lucerne. 36
Table 4.1. Shoot DM and root DM for two genotypes of tedera and one genotype of lucerne. 54
Table 5.1. Climatic data from the airport of Arrecife, Lanzarote and Newdegate, Western Australia based on measurements from 1972 to 2000. Mean air temperature is T, Tmin and Tmax are mean minimum and maximum air temperatures, respectively, rH is relative humidity. Differences between sum of all months and years are the result of rounding. Adapted from Graf et al. (2004) and Bureau of Meteorology. 80

PAGE | xvi
LIST OF PLATES
Plate 1.1. Lanzarote is 115 km from the West coast of Africa. In summer there is high irradiance and high temperatures, combined with cloudless skies and strong winds, resulting in a very high evaporation rate. 3
Plate 1.2. Traditional cut and carry of tedera in the Canary islands (photos courtesy of P Mendez). 4
Plate 1.3. Soil removed in three layers from the field site at Newdegate in the southern wheatbelt of WA for controlled-environment studies. 10
Plate 2.1. Seedling drought experiment in controlled environment. 24
Plate 2.2. Upper root system of five-week-old tedera seedlings from well-watered controls showing tap and lateral roots with nodules. 24
Plate 2.3. Successful establishment of tedera seedlings after direct sowing of pods at a depth of 50–70 mm with an experimental seeder. 25
Plate 3.1. Newdegate field experiment January 2010 with rain exclusion shelter (Cover remains on from early December to late April the following year). 40
Plate 3.2. Tedera leaves at the Newdegate field site in mid-summer showing heliotropism from 08.00 h to 18.00 h. 41
Plate 4.1. Drought and re-watering experiment before imposing drought stress (Note pot crane for weekly weighing of pots). 61
Plate 4.2. Tedera plant 14 days after re-watering (top) and after 28 days (bottom). 62
Plate 5.1. Examining the root structure of two-year-old tedera plants at the Newdegate site. 65
Plate 5.2. Aerial view of tedera plants growing outside, showing moderate folding of upper leaves. Note the way the leaves are typically arranged around the crown which should enhance the interception of light on lower leaves. 72
Plate 5.3. Strong pubescence on leaves and petioles of tedera after 30 days of drought stress (right) compared to well-watered control plants (left). Source: chapter 4. 73
Plate 5.4. Tedera (accession 4) plant excavated at the end of summer (late March 2010) at the Newdegate field site (Chapter 3). Note the fine fibrous root mass just below the soil surface (a), and the extensive lateral root system (b and c). 56
Plate 5.5. Cross sections of stems from well-watered control (above) and drought-stressed tedera plants (below) after withholding water for 47 days. Stems were embedded in glycol methacrylate and tissue sections viewed under a light microscope. Structures that resemble tyloses in the xylem vessels of drought-stressed tedera are circled in yellow. Plants were 10 months old. 76
Plate 5.6. Leaves of well-watered tedera plants restrained from moving while outside in mid-summer. Note the bleaching of the leaf edges after 24 hours. 79
Plate 5.7. Two-year old tedera accessions at the Newdegate field site. Tedera plants are often a hemispheric shape at low density, and this shape is possibly more efficient from a light-harvesting point of view than an erect lucerne plant. 81
Plate 5.8. Tedera seedlings in late autumn (a) and four weeks later in early winter (b) competing with annual background species. 82
Plate 5.9. Tedera pods were directly sown in the field to a depth of 50–70 mm (similar to cereals) and seedlings successfully emerged. Note some lateral roots were lost from the roots during removal from the soil. 83
Plate 5.10. Drought-resistant tedera accession under a rain-exclusion shelter showing high leaf retention and folding but also high levels of flower abortion. 87

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 1
CHAPTER 1. GENERAL INTRODUCTION
THE NEED FOR NEW PERENNIAL LEGUMES
Legumes play a critical role in sustainable agricultural systems because of their nitrogen-fixing
symbiosis with rhizobia (Subbarao et al., 1995). However, the lack of profitable and productive
perennial legumes is limiting the productive base for Australian farmers, as they currently rely
on narrowly-based annual farming systems susceptible to economic and climatic challenges.
There is a need to develop more sustainable and profitable animal production systems, in
particular, more resilient pasture and feeding systems adapted to the predicted increase in
seasonal variability as climate change impacts are realised. Consequently, an understanding of
drought stress and water use in relation to plant growth is vital for sustainable farming
systems. However, the current range of perennial pasture legumes and grasses cultivated in
Australia is limited. In eastern Australia, perennial pasture species are still largely confined to
regions where annual rainfall exceeds 500–600 mm (400 mm for lucerne (Medicago sativa L.)
or, in the case of Western Australia (WA), where the growing season is 8 months or longer
(Biddiscombe et al., 1982). Even within those medium-rainfall regions, the productivity of
perennial-based pastures has not often been superior to that of annual-based pastures (Kemp
et al., 1989).
In the Mediterranean environment of WA, annual pasture feed quality often deteriorates in
late summer and autumn, particularly after summer rainfall events (Nicol et al., 2013), and
producers inherently face critical feed shortages across these months. Most crop residues and
other grazing resources are also low in digestibility and nitrogen content during these months.
Producers are required to either reduce the number of stock or provide costly feed
supplements. This is one of the main factors limiting the value of the stock enterprise in the
agricultural regions of southern Australia (Bathgate and Pannell, 2002) and a factor that could
be addressed through inclusion of summer-active perennial pastures. However, to achieve
adoption, perennial pasture systems must be as profitable and practical as the agricultural
enterprises they displace.
At present, lucerne is often the only perennial legume option. Where lucerne is well suited, it
demonstrates good production, drought tolerance, ability to respond to summer/autumn
rainfall and persistence (Moore et al., 2006). However, there are several major limitations to
the broader use of lucerne in WA, including its sensitivity to acid soils and high soil aluminium,
and its susceptibility to waterlogging (Humphries and Auricht, 2001). Hill (1996) suggested that
lucerne is limited to the higher-rainfall zones in WA, which comprise only 42% of agriculture

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 2
land. Yet, the area planted to lucerne in WA falls far short of its estimated potential (7.8 Mha),
with its production in pure and mixed stands being approximately 170,000 ha (Robertson,
2006). It is also risky to rely predominantly on one perennial legume pasture species due to the
risk of serious yield loss from disease and insect outbreaks (Cocks, 2001).
Thus, there is clearly a need for alternative perennial legumes to lucerne in the low-to-medium
rainfall cropping zone of southern Australia. The demand for new perennial legumes is also
likely to increase, given predictions of more intensive and out-of-season rainfall events, which
will be more suitable for deep-rooted, summer-active perennials than for winter annuals
(Marshall et al., 2011). New species will need to be well adapted to soils, climate and farming
systems, and be productive and persistent. However, improving the drought resistance of
crops first requires knowledge of the key physiological mechanisms and the genetic control of
the contributing drought resistance traits (Farooq et al., 2009).
New perennial species will need to have drought-adaptive traits, both as seedlings (in the
establishment phase) and as mature plants, if they are to sustain the productivity of our
pasture systems under predicted climate change. Even though the seedling stage is only a
short period in the life of a plant, it is critical for plant survival (Sanderson et al., 1997). The
primary reason why seedlings die in many environments is drought and soil desiccation (Moles
and Westoby, 2004). Thus, in a Mediterranean climate, any reduction in rainfall at the
beginning of the autumn growing season would have a large negative effect on seedling
survival and growth (Tadross et al., 2007). While screening for differences in seedling survival
during drought would be an easy and effective solution for future climate change scenarios
(Vadez et al., 2011), there has been little evaluation of seedling drought resistance and survival
in the major crops and pastures grown in south-west Australia (Turner et al., 2011).
THE DEVELOPMENT OF NEW DEEP-ROOTED PERENNIALS ADAPTED TO LOW-TO-
MEDIUM RAINFALL
Over the past decade, a large collaborative research effort has focused on developing new
perennial legumes as alternatives to lucerne for southern Australia. A large number of species,
both native and exotic, have been evaluated (Bell et al., 2006; Dear and Ewing, 2008). After
several years of trials by the Future Farm Industries Cooperative Research Centre (FFI CRC) and
the Department of Agriculture and Food Western Australia (DAFWA), the most promising
species was the exotic Bituminaria bituminosa var. albomarginata (common name tedera),
which showed good summer feed production and drought resistance (Suriyagoda et al., 2013).
There are a number of botanical varieties of B. bituminosa and their responses to drought

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 3
differ considerably. Martinez-Fernandez et al. (2010) found var. albomarginata from a low
rainfall area in Lanzarote, Canary Islands (Plate 1.1) to be the most drought-tolerant variety.
Notably, it had the most biomass under water stress relative to other varieties.
Plate 1.1. Lanzarote is 115 km from the West coast of Africa. In summer there is high
irradiance and high temperatures, combined with cloudless skies and strong winds, resulting in
a very high evaporation rates.

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 4
NATIVE ENVIRONMENT OF TEDERA
Tedera is native to the island of Lanzarote, which belongs to the Canaries archipelago in the
eastern north Atlantic, 100 km off the coast of Morocco (Plate 1.1). The Canary Islands are
described as a 'hotspot' of plant diversity within the Mediterranean global diversity hotspot
(Quezel and Medial, 1995). Tedera is found at low altitude and in areas with an annual rainfall
of 50–200 mm (depending on location) with a Mediterranean rainfall pattern and a 5–6 month
dry season. In the Canary Islands, tedera is a valuable source of summer-green feed for sheep
and goats after cutting and drying (Plate 1.2). The rainfall is seasonal, from November to
March (Table 5.1), but with high inter-annual variability. For most of the year, these islands are
subject to north-north-easterly trade winds which suppress the development of any rainfall
(Graf et al., 2004), but also bring high relative humidity (>75%) and high evapotranspiration
(>980 mm). Tedera is well adapted to this adverse environment of this Mediterranean summer
as most of its reproductive development (flowering and seed set) in the Canary Islands takes
place in the absence of any precipitation. Most annual plants in such a water-limited
environment would escape drought by finishing their reproductive phase before the onset of
drought.
Plate 1.2. Traditional cut and carry of tedera in the Canary Islands (Photos courtesy of P
Mendez).
The harsh environmental conditions of the Canary Islands have presumably resulted in the
evolution of drought resistance traits to allow plants to survive and reproduce. The scarce
rainfall, low altitude and high sun exposure on Lanzarote have all been reported to contribute
to adversity being a constant, thus favouring the most resistant species (Reyes-Betancort et al.,

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 5
2001). The distinctly seasonal precipitation is also likely to have shaped the evolution of water
relations and root system characteristics of plants in this ecosystem. Given the complexity of
limiting factors in the Canary Islands, it seems likely that combinations of morphological and
physiological features, not single traits, would be responsible for the adaptive capacity of
tedera to its native environment.
DROUGHT RESISTANCE MECHANISMS
The term 'drought' has different meanings for hydrologists and meteorologists. Within an
agricultural context it has been defined as “a climatic excursion involving a shortage of
precipitation sufficient to adversely affect crop production” (Rosenberg, 1980). In the
physiological literature, drought can be described as a water deficit, which impairs plant
growth and yield compared with the amount needed for optimum growth (Lawlor, 2013). The
majority of a plant’s physiological and metabolic processes are affected by water deficits. Once
a drop in water potential develops, a wide range of physiological responses are induced
(Cattivelli et al., 2008).
The ability of a crop to grow adequately in environments that experience water deficits has
been referred to as 'drought resistance', and the modification in plant structure and function
that increases the chances that these plants will survive is termed 'adaptation' (Kramer, 1990).
Plants express various mechanisms to cope with drought stress. Typically, plant adaptation to
drought has been separated into escape, avoidance or tolerance strategies (Turner, 1986).
However, these are not necessarily mutually exclusive and plants may combine a range of
these responses (Ludlow, 1989). Some of these responses are triggered directly by the
changing water status of the tissues, while others are brought about by plant hormones that
signal changes in water status (Chaves et al., 2003). A brief overview of drought resistance
mechanisms at various levels in crop plants is provided below.
Plants can escape drought by finishing their life cycle while sufficient moisture is still available,
as occurs for many flowering annuals, which allows plants to reproduce before the
environment becomes too dry (Smith et al., 1998; Chandler and Bartels, 2003). This can be
illustrated by early flowering (<100 days) in annual pastures and in crops such as wheat
(Triticum aestivum L.) or barley (Hordeum vulgare L.), or subclover (Trifolium subterraneum L.)
where stored water is used before the onset of terminal drought. However, early flowering to
escape drought may involve a cost in terms of reduced yield (Turner et al., 2001), as a longer
growing period is often associated with higher yields (Smith et al., 1998).

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 6
Drought avoidance in plants depends on maintenance of cell turgor through increased water
uptake and/or reduced water loss (Tuberosa et al., 2003). Efficient stomatal regulation, a
reduction in light absorbance, smaller leaf area, and vertical leaf orientation are among the
major drought-avoidance traits which minimise transpiration in order to conserve water under
stress conditions (Wang and Yamauchi, 2006). For instance, light-avoiding leaf movements
(paraheliotropism) have previously been reported in members of the Fabaceae family, such as
within species of Phaseolus (Bielenberg et al., 2003) and Macroptilium (Ludlow and Björkman,
1984). In addition, many desert plants have a thick covering of light-reflecting hairs on their
leaves which increases reflectance and reduces absorbance of solar radiation reaching the leaf
surface (Fahn, 1989).
Drought tolerance involves processes at the cellular level, particularly osmotic adjustment
(OA), and is one of the crucial components of acclimation to drought stress. Osmotic
adjustment is the accumulation of organic and inorganic solutes under drought which helps to
lower the plant water potential without decreasing actual water content (Serraj and Sinclair,
2002). Osmotic adjustment is normally a slow process in the shoots and is initiated above a
certain level of cell water deficit (Chaves et al., 2003). Osmotic adjustment is considered to
enhance growth and yield because it aids in the maintenance of leaf turgor, and has previously
been correlated with grain yield in some crop species (Gebre and Tschaplinski, 2000). Some of
the osmotic compounds produced include sugar alcohols, soluble sugars, glycinebetaine and
proline (Kiani et al., 2007; Lambers et al., 2008). The accumulation of proline under drought
stress has been reported in other legume crops such as pea (Pisum sativum) (Sánchez et al.,
1998). However, some studies have reported OA in legumes and not found any associated
benefits (Turner et al., 2007).
TEDERA AND ITS POTENTIAL RELEVANCE TO SOUTHERN AUSTRALIAN FARMING
SYSTEMS
Tedera has significant potential to contribute to southern Australian farming systems, to
persist and be productive in the low-medium rainfall cropping zones, and to provide critical
out-of-season forage production, which is one of the major constraints to increasing wool and
meat production and profitability. This is especially so in WA, where some soils limit cropping,
and where a longer-term more stable pasture phase is required. A phase-farming system is
where successive years of perennial-based pasture species are grown, followed by several
years of grain crop species (Cocks, 2001). If tedera can be a successful component of such a
phase system, it could extend the range of perennial species in WA.

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 7
Green leaf retention is a highly-desirable trait in a perennial pasture species over summer and
autumn, as leaf biomass is likely to make up most of the nutritional intake for livestock. Tedera
plants retain most of their green leaves during summer and autumn which contrasts with
lucerne, which often drops its leaves as a drought-avoidance mechanism (Irigoyen et al., 1992;
Moore et al., 2006). The whole-farm bio-economic model MIDAS (Model of an Integrated
Dryland Agricultural System) indicates that the provision of green forage in summer and
autumn can generate a large increase in profit by reducing the requirement for purchased
supplementary feed (Finlayson et al., 2012). A recent pilot grazing trial showed tedera plants
could be grazed during both summer and autumn without causing problems to livestock, and
the recovery of the plants after grazing in this same period has been excellent (D Real
personal. communication). Suriyagoda et al. (2013) also found excellent regrowth following
cutting in early summer.
The extended summer/autumn growth of perennials such as tedera should also confer an
ability to dry the soil to a greater depth than can be achieved by annual crops and pastures
(Bell et al., 2006), thereby reducing the potential for groundwater recharge and associated
secondary salinity (Dear et al., 2003). Additional benefits will likely arise from more permanent
soil cover and reduced soil erosion during summer and autumn.
FOCUS OF PHD RESEARCH ON DROUGHT RESISTANCE MECHANISMS IN TEDERA
Research into plant responses to water availability is increasingly important and the need to
improve plant drought tolerance is particularly urgent as most climate change models predict
an increase in aridity in many parts of the world (Petit et al., 1999; IPCC, 2007). The south-west
of Australia urgently needs new drought-tolerant perennial pasture species to be introduced
as part of the overall climate change management plan (Turner et al., 2011). With the
predicted fluctuations in rainfall patterns due to climate change, there may also be changes in
the occurrence of episodic drought (IPCC, 2007), where plants will be repeatedly exposed to
drought in the field. Repeated exposures to drought can increase the sensitivity of plants to
changes in the environment that induce stomatal closure, while also reducing the sensitivity to
changes that induce stomatal opening (Aasamaa and Sober, 2011). Understanding how
perennial legume species like tedera respond to rainfall after prolonged summer drought will
be important for farmers to implement pasture-management practices. This understanding is
essential for forages that are used for livestock feeding schedules aimed at high production
systems.
Many plant adaptations for survival in harsh environments also tend to reduce economic yield
(Richards, 1997) and persistence does not equal productivity. It is often difficult to tell how, or

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 8
even whether, a particular phenotypic character contributes to a plant’s survival. As an
example, Medicago asiatica (Sinskaya) is highly resistant to leaf drop in Afghanistan, but
undergoes severe leaf shedding when grown in Europe (Bennett, 1970). Strategies that are
good in an arid environment such as increased water-use efficiency and slower growth rates
may be a disadvantage under more favourable conditions as plants may be outcompeted by
less drought-tolerant plants (Arve et al., 2011). Tedera plants have also evolved under a humid
environment in the Canary Islands (Table 5.1); how will the plants respond to the lower-
humidity environment of the southern wheatbelt? Plants grown under constant high relative
humidity tend to contain less abscisic acid (ABA) and their stomata can be larger than those of
plants grown under lower-humidity environments (Nejad and van Meeteren, 2007).
There are also often significant abiotic stresses in many Australian wheatbelt soils, such as low
pH, aluminium toxicity, nutrient deficiencies and low water-holding capacity; all of these could
impact drought resistance, survival and productivity of plants. For instance, the roots of many
crops will not grow into an acid soil layer with a high aluminium concentration (Lambers et al.,
2008). This severely limits the ability of the plant to obtain nutrients and also restricts the
overall rooting depth and can lead to severe drought symptoms in the crop, even if moisture is
available at depth. The soils on Lanzarote were also high in alkalinity (pH up to 8.6 in saturated
paste (Arve et al., 2011); how will tedera plants respond to lower pH soils?
The drought resistance mechanisms of tedera may allow it to survive, but, critically, will it
remain productive over the summer and autumn drought in southern Australian
environments? How do we manage tedera to maximise yield and livestock production over
summer and yet maintain persistence? To answer these questions, understanding the
physiological and ecological significance of the various drought resistance mechanisms in
tedera is required. This will allow breeders to target new sites for germplasm collection,
undertake plant selection and breeding programs, produce new cultivars that will not only
survive over summer, but, critically, be productive and profitable, in our southern Australian
farming systems.
While drought resistance and recuperative potential are known to vary within several
perennial grass species (Su et al., 2008), the combined responses to water stress and re-
watering among perennial legumes is relatively unknown (Filippou et al., 2011). More
generally, the relationship between plant growth and ecophysiological functioning in response
to water stress and re-watering is not yet fully understood and needs to be further
investigated (Xu and Zhou, 2008). Understanding physiological factors that enable perennial

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 9
legumes to rapidly recover from drought damage after summer or autumn rainfall events is
also important in legume management, particularly in water-limiting environments.
THESIS METHODOLOGICAL APPROACH
Growing plants in pots in controlled environments enables tight control of many variables such
as humidity, light and temperature and are therefore more repeatable than field experiments
(Passioura, 2006). Extrapolating experimental pot results to the field is considered possible if
some precautions are taken, including ensuring that pots are large enough (Poorter et al.,
2012), the growing media is from the field, and soil hypoxia is avoided (Passioura, 2006).
However, many studies investigating stress effects on plants are short-term, which are often
not directly applicable to the field (Cai et al., 2010). Important considerations should also
include: the method used to impose dehydration, the maturity of the plants, the severity and
duration of dehydration, and the physiological status to be assessed.
In some studies, inert polymers of high molecular weight, such as polyethylene glycol (PEG),
have been applied to simulate a programmed level of drought stress by reducing the water
potential of the nutrient solution (Kaufmann and Eckard, 1971). The use of PEG can avoid
many of the problems associated with using soil for pot-grown plants for drought experiments
(Lawlor, 2013). As an example, it is possible to change the water status readily to particular
values and apply at specific times (Lawlor, 1970). However, these polymers have also entered
the plant and exerted a toxic effect, regardless of the water uptake limitation (Munns et al.,
1979). Moreover, the physiological responses to drought and osmotically-applied stress are
largely different in terms of shoot:root carbon allocation and the metabolic pathways involved
(Frechilla et al., 1993), and while the use of PEG in experiments may increase our knowledge of
stress physiology, it is unlikely to represent physiological conditions in the field (Lawlor, 2013).
Nonetheless, imposing drought stress in controlled environments by simply ceasing to water is
also problematic. For instance, depending on the media, plants in large pots may even develop
stress soon after watering has ceased (Pang et al., 2011), while development of water deficit in
the field is often a gradual process reflecting the greater volume of soil explored by the root
system. The plant’s response to the velocity of stress imposed will affect the metabolism in
distinct ways when acclimating to slowly-imposed drought stress or with a fast response to a
dramatic change in tissue water status (Waseem et al., 2011). A better plant growth system to
produce agronomically-relevant results is to create a physiological water stress in which the
drought stress is a gradual depletion of water using soil from the field, where water deficit
occurs over weeks, not days, and the timing (in terms of season) of the drought stress matches
the stress in the field.

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 10
For the reasons outlined above, the glasshouse experiments presented in this thesis (Chapters
2 and 4) used soils from a field site in a region targeted for tedera adoption. The soil (texture-
contrast soil, grey coarse sand over an alkaline clay loam; Plate 1.3) also represents a major
soil type of high agricultural importance in south-western Australia (Moore et al., 1998). This
approach was deemed the most ecologically relevant for water relations and ensured the
presence of many physical, chemical and biological characteristics of a natural soil. Commercial
potting mixes often behave differently from soil (more aeration and drainage) in their water-
holding capacity and water release characteristics and this is especially problematic for
drought stress experiments (http://plantstress.com/methods/PotExp.htm).
Plate 1.3. Soil removed in three layers from the field site at Newdegate in the southern
wheatbelt of WA for controlled-environment studies.
To gain a better understanding of the physiological and morphological responses of tedera
over summer, field studies were also undertaken (Chapter 3). Field-grown plants are likely to
be subjected to one or more cycles of lower soil water potential and consequently may
become pre-conditioned, or 'hardened' to subsequent drought stress (Ashley, 1993). Although
designing and setting up field experiments is a difficult task and can be complex with high risk
of failure, testing newly-developed stress-tolerant genotypes to multiple stresses under

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 11
controlled-environment conditions must be confirmed with studies in the field (Richards et al.,
2010). However, screening plants in the field under natural conditions for drought resistance
can be difficult due to the possibility of summer-rainfall events; this becomes more
manageable by using rain-exclusion shelters (Farooq et al., 2009).
THESIS AIMS
The specific aims of the research reported in this thesis were to:
a) Compare the drought resistance of tedera (one accession) with other pasture legume
species at the seedling stage in a controlled environment (Chapter 2).
b) Examine seasonal and diurnal variation in stomatal conductance and paraheliotropism
of tedera (three accessions) in the field (Chapter 3).
c) Investigate in detail in a controlled-environment the key physiological and
morphological traits related to the drought resistance mechanisms of tedera (two
accessions) identified in the field (Chapter 4).
d) Investigate in detail in a controlled-environment the key physiological and
morphological traits related to the mechanisms for recovery from drought stress of
tedera (two accessions) (Chapter 4).
Drought stress in tedera is likely to trigger a wide variety of leaf and root physiological and
morphological responses. This study is among the first to investigate the mechanisms that
confer drought resistance in tedera in the field and examine the level of variation within seven
accessions for this highly-desirable trait. Moreover, new detailed knowledge on the
acclimation of tedera (and lucerne) to severe drought conditions will increase the relatively
poor knowledge of drought resistance mechanisms in perennial legumes.
The results from this study will increase understanding of the ecophysiology and ecology of
tedera, and the agronomic potential for tedera to increase the sustainability of Australian
farming systems threatened by predicted climate change. A better understanding of the
mechanisms of drought resistance and recovery will allow specific traits to be targeted in
selection/breeding programs prior to the release of the first tedera cultivar and enhance the
potential for the adoption of this species into low-medium rainfall pasture systems.

CHAPTER 1. GENERAL INTRODUCTION
PAGE | 12
THESIS STRUCTURE
This thesis is in agreement with the Postgraduate and Research Scholarship Regulation
1.3.1.33(1) of The University of Western Australia and is presented as a series of three
scientific papers. Two manuscripts are published and included as PDFs with the other is
accepted and published online. All work presented has resulted from work done towards this
thesis. There are five main chapters in this thesis: General Introduction, three Experimental
Chapters and General Discussion. The General Introduction covers the broad background for
the work presented in the thesis in order to justify the research objectives presented. A more
focused review of literature is presented in the Introduction of each Experimental Chapter. The
three Experimental Chapters are presented in the format of scientific papers that can be read
individually or as part of the whole thesis. Each Experimental Chapter includes the following
sections: Summary, Introduction, Materials and Methods, Results, Discussion and References.
Each Experimental Chapter is preceded by a short Preface, which serves to link the chapter to
the broader hypotheses addressed by the thesis and clarifies the role of each listed author of
the paper. This 'thesis-as-a-series-of-papers' format results in some unavoidable repetition,
especially in the Materials and Methods sections of Chapters 2 with 4. The references for
Chapters 1 and 5 are combined at the end of the thesis.

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 13
CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN THE PROMISING FODDER PLANT TEDERA (BITUMINARIA BITUMINOSA VAR.
ALBOMARGINATA)
Kevin FosterABCD, Megan H. RyanAB, Daniel RealABC, Padmaja RamankuttyAB, Hans LambersA
B Future Farm Industries Cooperative Research Centre, The University of Western Australia,
Crawley, WA 6009, Australia
C Department of Agriculture and Food Western Australia, South Perth, WA 6157, Australia
D Corresponding author
Published: Crop & Pasture Science 63, 1034–1042
18 December 2012
Keywords: canopy temperature depression, novel pasture species, paraheliotropism, perennial
legume
A School of Plant Biology and Institute of Agriculture, The University of Western Australia,
Crawley, WA 6009, Australia

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 14
PREFACE
While many studies have investigated the response of crop and pasture species to drought
stress, there has been little evaluation of seedling drought resistance, especially in legumes.
This study was conducted to compare, in a controlled environment, the drought resistance of
tedera (one accession) with other pasture legume species at the seedling stage. I undertook
most of the work involved in preparing this chapter including all tasks related to setting up the
experiment, germplasm selection, planting, irrigation, physiological and morphological
measurements, harvesting, and interpretation of the results. Associate Professor Ryan, Adjunct
Professor Real and Winthrop Professor Lambers contributed meaningful comments on the
ideas presented. Dr Ramankutty provided statistical assistance and Associate Professor Renton
contributed towards the initial experimental design. Dr Christine Davies of Tweak Editing
contributed editorial assistance on the manuscript.

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 15

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 16

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 17
CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 18
CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 19

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 20

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 21

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 22

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 23

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 24
SUPPORTING INFORMATION
Plate 2.1. Seedling drought experiment in controlled environment.
Plate 2.2. Upper root system of five-week-old tedera seedlings from well-watered controls
showing tap and lateral roots with nodules.

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 25
Plate 2.3. Successful establishment of tedera seedlings after direct sowing of pods at a depth
of 50–70 mm with an experimental seeder.

CHAPTER 2. DROUGHT RESISTANCE AT THE SEEDLING STAGE IN TEDERA
PAGE | 26

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 27
CHAPTER 3. SEASONAL AND DIURNAL VARIATION IN THE STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA (BITUMINARIA
BITUMINOSA VAR. ALBOMARGINATA) IN THE FIELD
Kevin FosterABCD, Megan H. RyanAB, Daniel RealABC, Padmaja RamankuttyAB, Hans LambersA
B Future Farm Industries Cooperative Research Centre, The University of Western Australia,
Crawley, WA 6009, Australia
C Department of Agriculture and Food Western Australia, South Perth, WA 6157, Australia
D Corresponding author
Published: Functional Plant Biology 403, 719–729
10 July 2013
Keywords: adaptation, alfalfa, lucerne, paraheliotropism, perennial legume, Psoralea
bituminosa
A School of Plant Biology and Institute of Agriculture, The University of Western Australia,
Crawley, WA 6009, Australia

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 28
PREFACE
This chapter has been published as follows: “Foster, K., Ryan, M., Real, D., Ramankutty P.,
Lambers, H. (2013). Seasonal and diurnal variation in stomatal conductance and
paraheliotropism of tedera (Bituminaria bituminosa var. albomarginata L.) in the field,
Functional Plant Biology 40, 719–729”. The framework for this study is that light, relative
humidity and temperature are often quite contrasting in controlled-environment studies
compared with field conditions and as such, stomatal behaviour in response to water stress
may be considerably different. This study used two field sites with contrasting soil types within
the medium-low rainfall zone of the WA wheatbelt. The first site at Merredin has a naturally
highly-acidic soil (pH in CaCl2 3.8 at 100 cm); however, the site was destroyed by rabbits
following damage to a perimeter fence before many of the measurements could be completed
and no results are reported. The second site at Newdegate is the focus of this chapter. I
undertook most of the work related to this chapter, including all tasks related to setting up the
field experiments, germplasm selection, site preparation, planting, nutrition, irrigation design,
scheduling of watering, biomass sampling, plant physiological and morphological
measurements, and interpretation of the results. Associate Professor Ryan, Adjunct Professor
Real and Winthrop Professor Lambers contributed meaningful comments on the ideas
presented. Dr Ramankutty provided statistical assistance and Dr Jane Speijers was responsible
for the initial experimental design. Dr Christine Davies from Tweak Editing contributed
editorial assistance with the manuscript.

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 29

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 30

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 31

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 32

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 33

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 34

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 35

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 36

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 37

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 38

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 39

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 40
SUPPORTING INFORMATION
Plate 3.1. Newdegate field experiment January 2010 with rain exclusion shelter (cover remains
on from early December to late April the following year).

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 41
Plate 3.2. Tedera leaves at the Newdegate field site in mid-summer showing heliotropism from
08.00 h to 18.00 h.

CHAPTER 3. STOMATAL CONDUCTANCE AND PARAHELIOTROPISM OF TEDERA
PAGE | 42

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 43
CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE BITUMINARIA BITUMINOSA (L.) C.H. STIRTON. VAR. ALBOMARGINATA.
Kevin FosterABCD, Hans LambersA, Daniel RealABC, Padmaja RamankuttyAB, Gregory R.
CawthrayA, Megan H. RyanAB
B Future Farm Industries Cooperative Research Centre, The University of Western Australia,
Crawley, WA 6009, Australia
C Department of Agriculture and Food Western Australia, South Perth, WA 6157, Australia
D Corresponding author
Annals of Applied Biology. Published online October 30th 2014
Keywords: alfalfa, climate change, osmotic adjustment, paraheliotropism, plant adaptation,
Psoralea bituminosa, rehydration, water-use efficiency
A School of Plant Biology and Institute of Agriculture, The University of Western Australia,
Crawley, WA 6009, Australia

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 44
PREFACE
This chapter has been formatted for and submitted to Annals of Applied Biology. Despite its
importance, studies on the capacity of photosynthetic recovery from drought stress have only
been conducted on a few species. The ability of tedera to recover from drought stress is as
essential for plant growth and survival as drought resistance during water stress. For
consistency, this controlled-environment study used the same accessions and same soil as the
field experiment (Chapter 3). Growing plants in pots provided the opportunity to examine
changes in root systems that may occur during drought stress and recovery. I undertook most
of the work related to this chapter, including all tasks related to setting up the experiment,
germination, planting, nutrition, irrigation scheduling, plant physiological and morphological
measurements, sampling and preparation for chemical analysis, data entry, and preparation
and interpretation of the results. Associate Professor Ryan, Adjunct Professor Real and
Winthrop Professor Lambers contributed meaningful comments on the ideas presented. Dr
Ramankutty provided statistical assistance and Associate Professor Renton contributed
towards the initial experimental design. Dr Christine Davies from Tweak Editing also
contributed with editorial assistance on the manuscript.
Please note: There are several typeset errors in the paragraph starting at line 37 on page 6 (i.e.
no spaces between sentences) and on page 8 (second column line 39, should read “that is,”)
that were not amended on the online publication but have been corrected for the print version.

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 45

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 46

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 47

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 48

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 49

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 50

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 51

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 52

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 53

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 54

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 55

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 56

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 57

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 58

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 59

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 60 Available from Wiley Online Library via http://dx.doi.org/10.1111/aab.12171

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 61
SUPPORTING INFORMATION
Plate 4.1. Drought and re-watering experiment before imposing drought stress (Note pot
crane for weekly weighing of pots).

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 62
Plate 4.2. Tedera plant 14 days after re-watering (top) and after 28 days (bottom).
CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 63
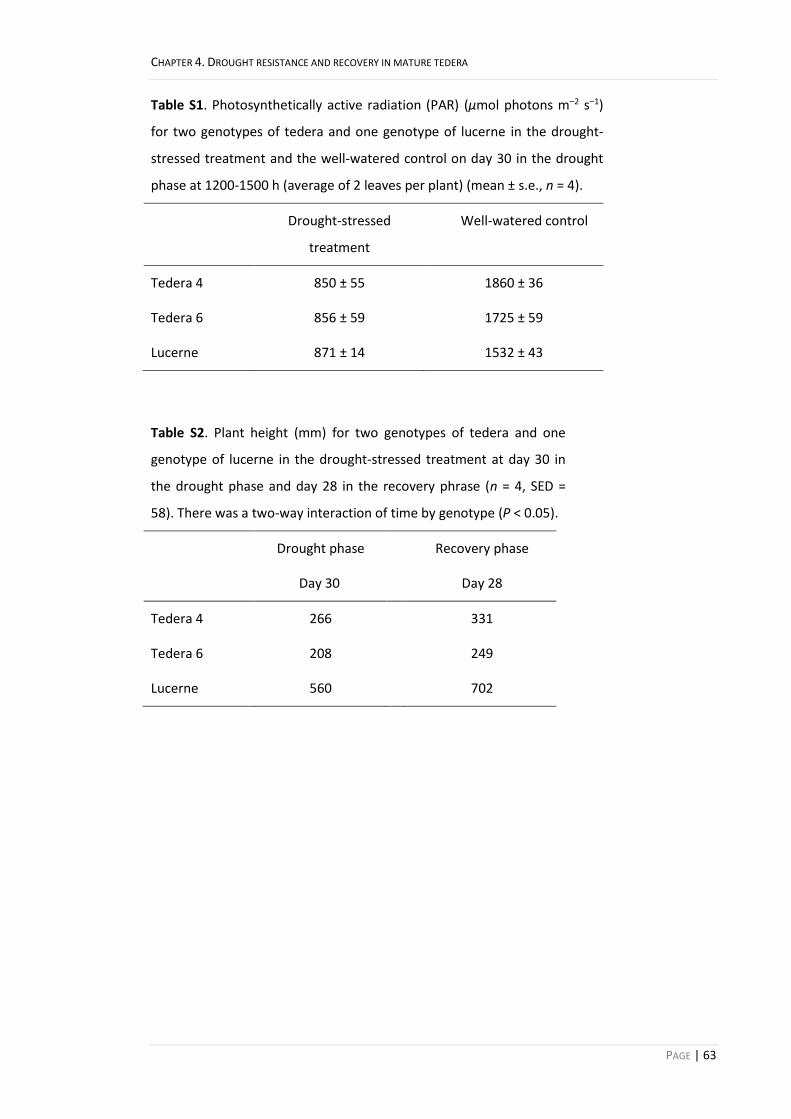
Table S1. Photosynthetically active radiation (PAR) (μmol photons m–2 s–1)
for two genotypes of tedera and one genotype of lucerne in the drought-
stressed treatment and the well-watered control on day 30 in the drought
phase at 1200-1500 h (average of 2 leaves per plant) (mean ± s.e., n = 4).
Drought-stressed
treatment
Well-watered control
Tedera 4 850 ± 55 1860 ± 36
Tedera 6 856 ± 59 1725 ± 59
Lucerne 871 ± 14 1532 ± 43
Table S2. Plant height (mm) for two genotypes of tedera and one
genotype of lucerne in the drought-stressed treatment at day 30 in
the drought phase and day 28 in the recovery phrase (n = 4, SED =
58). There was a two-way interaction of time by genotype (P < 0.05).
Drought phase Recovery phase
Day 30 Day 28
Tedera 4 266 331
Tedera 6 208 249
Lucerne 560 702

CHAPTER 4. DROUGHT RESISTANCE AND RECOVERY IN MATURE TEDERA
PAGE | 64
Table S3. Proline, proline betaine, fructose/pinitol and glucose concentrations in leaf tissue for two genotypes of tedera and one genotype of
lucerne at day 1 and day 47 of the drought phase, and day 28 of the recovery phase in the drought-stressed treatment (DS) and the well-watered
control (WW) (mean ± s.e., n=4). PWP = permanent wilting point, * = not detected, i.e. failure to detect any quantity above the detection limit:
proline (10 µmol g–1 DW), proline betaine (10 µmol g–1 DW), fructose (11 mg g–1 DW), glucose (16 mg g–1 DW) and pinitol (25 mg g–1 DW).
Phase Day Genotype Treatment Proline
(µmol g–1 DW)
Proline betaine
(µmol g–1 DW)
Fructose + Pinitol
(mg g–1 DW)
Glucose
(mg g–1 DW)
Drought Day 1 Lucerne WW * 35.5 ± 8.9 37.4 ± 9.3 17.1 ± 4.3
DS * 27.8 ± 2.1 41.6 ± 6.2 23.9 ± 1.4
Tedera 4 WW * * 20.5 ± 2.4 10.5 ± 1.6
DS * * 25.9 ± 1.2 18.4 ± 6.0
Tedera 6 WW * * 26.3 ± 2.2 16.1 ± 3.7
DS * * 22.0 ± 3.1 11.2 ± 2.7
Drought Day 47 Lucerne WW * 87.8 ± 23.5 71.7 ± 9.8 15.4 ± 3.8
DS PWP PWP PWP PWP
Tedera 4 WW * * 50.4 ± 4.8 15.1 ± 1.9
DS 205 ± 1 * 95.8 ± 3.4 36.9 ± 6.4
Tedera 6 WW * * 59.7 ± 5.6 23.5 ± 3.5
DS 191 ± 3 * 80.0 ± 4.2 40.4 ± 7.3
Recovery Day 28 Lucerne WW * 69.4 ± 12.1 49.9 ± 6.6 9.3 ± 1.6
DS * 58.0 ± 12.1 45.3 ± 1.2 4.7 ± 1.3
Tedera 4 WW * * 57.8 ± 2.0 16.3 ± 3.5
DS * * 48.4 ± 3.2 16.9 ± 7.0
Tedera 6 WW * * 51.1 ± 4.5 21.8 ± 3.0
DS * * 51.3 ± 4.1 25.7 ± 6.1

CHAPTER 5. GENERAL DISCUSSION
PAGE | 65
CHAPTER 5. GENERAL DISCUSSION
Plate 5.1. Examining the root structure of two-year-old tedera plants at the Newdegate site.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 66
INTRODUCTION
The predicted impacts of human-induced climate change are the highest in Mediterranean
ecosystems (Hulme et al., 1999), with changes predicted in temperature regimes and rainfall
patterns, as well as an increase in heat waves and droughts (IPCC, 2007). From 2000–2008,
Australia experienced two of its worst droughts on record. The years 2002–03 and 2006–07
had the most widespread adversity, with record high temperatures and dry conditions (ABS,
2008). Hennessy et al. (2007) concluded that without further adaptation to climate change,
negative impacts on agricultural production in Australia would likely be substantial and
projected that by 2030, production from agriculture could decline over much of southern
Australia. The recent Australian Bureau of Agricultural and Resource Economics report (ABARE,
2007) on climate change predicted Australia to be one of the most adversely-affected regions
from future changes in climate in terms of reductions in agricultural production and exports.
Australian farmers who rely on narrow-based annual farming systems will be susceptible to
both economic and climatic challenges. Based on the above, it can be concluded that our
current farming practices, based on predominantly shallow-rooted annual crops and pastures
with relatively low drought resistance, are not sustainable in the medium to long term.
To decrease the effects of climate change on agricultural land, farming systems in southern
Australia will likely need to be redesigned in this century to include crops and pastures with
improved maximum water-use efficiency. The resilience of our legume crops to current
weather extremes, such as drought, is also considered to predict their adaptation to future
climate change (Cutforth et al., 2007). Therefore, drought-resistant perennial species can be
part of this redesign, but they must be as profitable as the agricultural enterprises they
displace. Perennial pastures can be profitable by providing high-quality feed at strategic times
(i.e. autumn) and, preferably, being grown where they are economically competitive against
established cash crop systems (i.e. cereals and oilseeds). However, there appear to be few
herbaceous perennial legumes from elsewhere in the world that meet these criteria and are
suited for the southern Australian wheatbelt, particularly the acid soils (Dear et. al., 2003)
which current available species such as lucerne are poorly adapted. To maximise productivity
from new perennial legumes, we therefore need to consider a broad number of previously
unexplored genera and, likely, focus on those with different physiology to current options. We
also need to understand drought resistance mechanisms of any new perennial species
introduced.
There are new drought-resistant perennial herbaceous legumes from overseas currently being
evaluated that could play a key role in this redesigned agriculture. A list of priority species for

CHAPTER 5. GENERAL DISCUSSION
PAGE | 67
further evaluation was developed by the Future Farm CRC, following assessment of material
from collection missions and genetic resource centres. Tedera was regarded as a promising
species to provide out-of-season forage across all rainfall zones in southern Mediterranean
Australia. Tedera is from Lanzarote, the Canary Islands, has excellent fodder value and grazing
tolerance, is adapted to summer aridity, and can be found in soils with pH from 4.7 to 8.5 (Real
et al., 2009). However, the exact adaptations and/or drought resistance mechanisms that
enable tedera to grow in the harsh habitat of the Canary Islands as either a seedling or as a
mature plant are not known. It is likely not a single trait, but a suite of morphological and
physiological attributes are responsible for the adaptation of tedera to its native environment.
This thesis has provided an overview of morpho-physiological and biochemical responses of
tedera plants to drought stress by comparing it with lucerne. Lucerne (SARDI 10) was used,
where suited, as it is highly productive, relatively drought resistant and persistent, well
adapted to a Mediterranean climate (Bouizgaren et al., 2010) and is the standard by which
other perennial legumes are compared (Dear et. al., 2003; Pang et al., 2011; Suriyagoda et al.,
2013). In this chapter, several areas that have been highlighted by the research presented in
this thesis are discussed: a) the drought resistance mechanisms of tedera as a seedling and
mature plant; b) the species-specific mechanisms in tedera which allow it to survive in its
native environment; and c) the agronomic implications of the research findings. The results
from each experiment and their agronomic implications are considered, and future research
priorities are considered.
KEY FINDINGS FROM THIS RESEARCH
This study has shown that drought stress triggers a wide variety of physiological, biochemical,
and leaf and root morphological responses in tedera, which allow it to survive and recover
from drought stress. Comparing the water relations of tedera and lucerne, both as seedlings
and as mature plants, confirmed the superior drought resistance of tedera, which continues to
function during periods when water stress restricts lucerne growth. Tedera has physiological
traits such as osmotic adjustment (OA) that permit continued water transport, gas exchange,
and cell survival at very low relative leaf water content (RWC) and leaf water potential (LWP).
Water uptake is maximised in tedera by early taproot elongation in the seedling, by changes in
the allocation pattern of roots, namely an increase in the root:shoot ratio and in lateral root
growth under drought stress. Water loss is minimised in tedera by effective stomatal control,
increased water-use efficiency (WUEL) and reduced light absorbance through
paraheliotropism, leaf rolling and increased pubescence of stems and leaves (Chapter 4).

CHAPTER 5. GENERAL DISCUSSION
PAGE | 68
A significant finding was the confirmation that tedera can sustain shoot biomass under
extreme drought stress in the field over summer (Chapter 3). A novel finding from this
research was the significant accumulation of proline and pinitol in leaf tissue in tedera in
response to drought stress; both are likely central in maintaining water balance in tedera and
also in recovery from drought stress (Chapter 4). The ability of tedera to maintain
photosynthetic activity during a slowly-developing drought may be directly related to its
survival and recovery following a drought. Tedera demonstrated enhanced physiological
plasticity in changing environments relative to lucerne. Thus, it seems that tedera is a drought-
resistant plant, which not only maximises photosynthetic water-use efficiency, but,
importantly, maximises the use of available soil water.
ADAPTIVE RESPONSES OF TEDERA SEEDLINGS TO DROUGHT STRESS
Lack of intense rainfall in the Canary Islands (Chapter 1) combined with high evaporation rates
means seedlings need to implement short to medium-term physiological responses to rapidly
acclimate and survive in that environment. Chapter 2 examined the drought resistance
mechanisms of tedera at the seedling stage, compared with two other pasture legumes,
biserrula and lucerne, and the drought-resistant summer-active Afghan melon. All four species
decreased photosynthetic rate (A) and stomatal conductance (gs) in the drought treatment.
The photosynthetic rate of leaves of all plants decreased as RWC and LWP decreased. Tedera
and Afghan melon had tighter stomatal control and used water more efficiently; consequently
achieving a water-use efficiency (WUEL defined as the ratio of A/gs) up to three times greater
than that of biserrula and lucerne. The reduced photosynthesis in drought-stressed tedera
seedlings can be attributed to stomatal (stomatal closure) and possibly non-stomatal factors.
The early taproot elongation in tedera seedlings (Chapter 2) should allow for high water
extraction from the soil and would be a key factor in this early drought resistance. If tedera
were grown in the field under severe water-deficit conditions, the rapid elongation of the tap
root should keep up with the receding moisture front down the soil profile. Tedera and Afghan
melon also had more lateral roots in the 0–20 cm soil layer than either biserrula or lucerne in
both treatments (K Foster unpublished data). Observations in the field and the controlled-
environment study also showed that some of the early roots of tedera grew horizontally (Plate
5.1) in the upper soil surface (K Foster personal observations). Although water from upper soil
layers is often lost due to soil evaporation (Johansen et al., 1992), these lateral roots of tedera
seedlings may still provide a competitive advantage in early water extraction.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 69
ADAPTIVE RESPONSES OF MATURE TEDERA PLANTS TO DROUGHT STRESS
PHYSIOLOGICAL RESPONSES TO WATER STRESS
The field experiment at Newdegate (Chapter 3) is the first physiological study of tedera in the
wheatbelt of southern Australia. The morphological and physiological data collected over the
entire year provided valuable insight into the productivity and survival of tedera over summer
under extreme drought.
Daily rhythms have been recognised for a long time in different plant processes, namely
photosynthesis and stomatal aperture (Waseem et al., 2011). The depression in gs in tedera
during peak evaporative demand in the middle of the day in May (late autumn) and February
(late summer) may have resulted from an evolutionary pressure in the Canary Islands to avoid
physiological responses that may be disadvantageous during these parts of the day (when
temperature and radiation are excessive). This stomatal behaviour in tedera would be of major
adaptive significance to plants growing in arid areas, and this water conservation at the
expense of carbon gain is likely a critical adaptation of this species to water-limiting
environments. The stomatal control mechanisms in tedera may not only result in water being
used more efficiently, but may minimise the risk of xylem embolism by reducing the
probability of cavitation through stomatal closure during periods of high transpirational
demand (Meinzer et al., 2009).
Stomatal conductance is determined by a combination of stomatal size, density and aperture.
This is not controlled by soil water availability alone, but by a complex interaction of factors
internal and external to the leaf (Medrano et al., 2002). One possible reason for tedera’s more
effective control of stomata is the accumulation of abscisic acid (ABA) in leaves; ABA is
commonly associated with stomatal closure (Zhang and Davies, 1990). Previous studies also
suggest that stomates respond to xylem ABA under water-stressed conditions when gs has
decreased to a low value (Trejo et al., 1995). Initial soil drying may be sensed by part of the
shallow lateral root system observed in the field (see below) and root-sourced ABA in the
xylem may regulate gs such that water loss in the shoots may be reduced and leaf water deficit
can be avoided. It is possible that some tedera accessions are also hypersensitive to ABA, and
more resistant to drought stress (i.e. tedera 4), whereas those insensitive to ABA are more
drought sensitive (i.e. tedera 22).
Usually, abaxial surfaces from amphistomatous leaves (stomata on both surfaces) have higher
stomatal density compared with adaxial surfaces, so higher stomatal conductances are often
measured on abaxial leaf surfaces (Lu et al., 1993). However, the differences in gs of adaxial

CHAPTER 5. GENERAL DISCUSSION
PAGE | 70
and abaxial leaf surfaces found both species (Chapter 3) are probably directly influenced by
environmental variables and mediated by soil water deficits. The abaxial surface of leaves of
both species would be more exposed to wind, light and heat reflected from the ground, and
consequently experience a higher evaporative demand. Stomatal conductance decreased
more on the abaxial than adaxial leaf surface in rice cultivars, and this was due to a change in
leaf form and folding of the upper leaf surface (O’Toole and Cruz, 1980).
CHANGES IN ORGANIC SOLUTES IN RESPONSE TO WATER DEFICIT AND RE-WATERING
Chapter 4 reported for the first time the substantial concentration of proline in tedera in
response to water deficits and the presence of pinitol (Chapter 4). The accumulation of proline
and pinitol in tedera at the same time as significant changes in RWC and midday LWP suggests
it has a role in OA in leaf tissue, and contributes to stress tolerance during drought. Blum
(1989) suggested there was a positive linear relationship between growth and OA only when
OA equalled or exceeded 0.2 MPa. However, we found evidence of OA up to 0.5 MPa for both
tedera accessions during drought stress (Chapter 4). Osmotic adjustment may allow stomata
to remain open, at least partially, and CO2 assimilation to continue at very low LWPs (<–6.5
MPa) that could otherwise be inhibitory. High leaf proline concentrations have also been
reported to protect the photosynthetic system from permanent damage during drought
(Lawlor, 2001) and have been reported previously to not interfere with normal biochemical
processes (Stewart, 1981).
Pinitol, a sugar alcohol has been found in legumes especially under water stress conditions
(McManus et al., 2000) and may also serve as a scavenger of reactive oxygen species
(Sengupta et al., 2008). The accumulation of pinitol in accession tedera 4 may also contribute
to the drought tolerance of this accession and its ability to tolerate low LWPs. Proline was not
detected in leaves of tedera at the end of the recovery phase: the catabolism of proline has
been suggested to supply energy for plants to resume growth after stress (Szabados et al.,
2011). This strongly suggests a role for proline in plant drought recovery in tedera. Pinitol
concentrations decreased substantially in leaves of tedera 4 following re-watering, but were
still higher than pre-drought values, reflecting a slower turnover that likely contributed to
metabolism for growth of new shoots. Water uptake is likely enhanced in tedera by the
accumulation of these solutes to both lower tissue water potential and improve root growth.
The pool of these solutes, and possibly others, which built up in tedera during drought stress
probably helped to produce the new and rapid growth exhibited upon re-watering.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 71
MORPHOLOGICAL RESPONSES TO WATER STRESS
PARAHELIOTROPISM
Water availability plays a major role in modulating heliotropism in tedera, its deficit resulting
in steeper midday leaf angles (Chapters 3 and 4). These leaf movements appear to be an
important adaptation to reduce leaf temperature—not only from direct radiation from above,
but probably also from reflection and radiation from the soil surface—and thus the vapour
pressure gradient between leaf and atmosphere which, in turn, would reduce transpirational
water loss. Rapid leaf movements could also offer tedera seedlings a fast-acting stress-
avoidance mechanism in the case of a sudden rise in temperature or hot dry winds, and may
be critical when seedlings are highly exposed to the physical environment (due to a lack of self-
shading). Secondary pulvini on the base of the petioles adjoining the stem further increase the
upwards angle of the leaf (K Foster personal observations; Plate 5.9).
Although in this field study I did not report on leaf azimuthal direction—the compass direction
of leaves—sunlit leaves of tedera accessions in summer were oriented east in the early
morning (Plate 3.2) and faced west in the late afternoon, i.e. diaheliotropism (Koller, 1986).
Such diaheliotropic leaf angles may have evolved to improve day-long carbon gain by
increasing light interception from low angles and minimising water use with respect to daily
carbon gain (Cowan, 1982). This would allow tedera leaves to operate at maximum
photosynthetic capacity in the morning and the afternoon, increasing carbon gain (relative to
leaves in a fixed orientation) when evaporative demand is low and WUE is high. Any potential
loss of carbon assimilation due to this lower PAR when leaves are folded is likely to be offset by
favourable effects of reduced leaf temperature and transpiration as the heat load on these
upper leaves would be reduced compared with a horizontal leaf. Some of the lost PAR from
the upper folded leaves may be intercepted by leaves lower in the canopy (Plate 5.2), thereby
providing most of the required incident radiant energy for tedera to maintain photosynthetic
activity.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 72
Plate 5.2. Aerial view of tedera plants growing outside, showing moderate folding of upper
leaves. Note the way the leaves are typically arranged around the crown which should
enhance the interception of light on lower leaves.
INDIVIDUAL LEAF AREA AND PUBESCENCE UNDER DROUGHT STRESS
Tedera has a high degree of plasticity, which includes the ability to adjust its leaf canopy
according to soil moisture availability, and involves individual leaf area adjustment over the
seasons (Chapter 3). Smaller leaves were produced by tedera and lucerne over summer, and
this may not only reduce water loss but also allow more photoassimilates to be directed
towards root growth. These two mechanisms in tedera may help balance water conservation
and extraction in its native environment during summer. Smaller leaves are also well adapted
to high temperature and high light environments, and provide more efficient control of water
loss by stomatal closure (Jarvis and McNaughton, 1986). The new leaves of drought-stressed
tedera plants were increasingly pubescent on stems and leaves (Plate 5.3) which would partly
protect plants from excessive light injury and decrease transpirational water loss (Sandquist
and Ehleringer, 2003). The leaves produced in summer by tedera may not only differ
morphologically (e.g. smaller size, more leaf hairs), but also physiologically from the autumn
and winter-produced leaves.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 73
Plate 5.3. Strong pubescence on leaves and petioles of tedera after 30 days of drought stress
(right) compared to well-watered control plants (left). Source: Chapter 4.
ROOT CHARACTERISTICS AND GROWTH RESPONSES OF MATURE PLANTS TO DROUGHT STRESS
As an adaptive trait to drought resistance, plants in arid conditions often decrease
aboveground biomass production and allocate more biomass to roots, maintaining a higher
root:shoot ratio (Erice et al., 2007). We also observed higher root:shoot ratios in tedera
relative to lucerne under water-deficit conditions in the controlled-environment experiment
(Chapter 4). Root systems in tedera in this study had greater plasticity in response to drought
stress and re-watering than those of lucerne. Lateral root mass actually increased under
drought stress for tedera 4, with a large increase in root:shoot ratio at day 47 of drought
stress. This suggests stronger acclimation in tedera 4 to prolonged drought stress than in
tedera 6 or lucerne.
When tedera plants were removed from the soil at Newdegate at the end of summer, they had
an extensive lateral root system that spread over a large area with a mat of finer roots just
below the surface (Plate 5.4). These shallow and finer lateral roots may provide an opportunity
to access early-season precipitation before it is lost through evaporation, and may have
different responses to soil water than the primary tap root system. Pang et al. (2011) also

CHAPTER 5. GENERAL DISCUSSION
PAGE | 74
found that several perennial legume species, including tedera, increased allocation to surface
roots in response to drought stress. Other species from dry environments also have similar
root systems, with shallow lateral roots taking advantage of light rainfall events that may not
infiltrate far below the soil surface (Canadell and Zedler, 1995). Large summer-rainfall events
may increase the productivity of deep-rooted species like lucerne; however, with smaller
rainfall events predicted from climate change, these should benefit and increase productivity
in shallow-rooted species like tedera. These shallow lateral roots also appear to dry the upper
soil reducing competition from weeds during summer (K Foster personal observations).
Plate 5.4. Tedera (accession 4) plant excavated at the end of summer (late March 2010) at the
Newdegate field site (Chapter 3). Note the fine fibrous root mass just below the soil surface
(a), and the extensive lateral root system (b and c).
During the summer months, how do tedera's shallow lateral roots remain alive? The surface
soil dries out quickly during hot, dry periods, so these lateral roots likely release whatever
water they contain to the surrounding dry soil; thus the roots would likely die unless this water
was replaced. However, at the end of the drought phase (Chapter 4), upper lateral root
diameter (50 mm below the crown) declined by 22% in tedera when compared to the well-

CHAPTER 5. GENERAL DISCUSSION
PAGE | 75
watered controls (K Foster unpublished data). This may cause a root–soil air gap as the roots
shrink radially. Similar results have been reported by Nobel and Huang (1992) for drought-
resistant desert plants, who proposed that the air gap restricts water loss from the roots to the
drying soil in desert succulents. Drought stress may also cause embolism of the xylem vessels
(Chapter 4), and could reduce axial conductance for lateral roots as well as main roots.
Drought-induced embolisms form when xylem sap tension reaches a critical threshold and air
is aspirated through pit membranes. North and Nobel (1991) found that embolism reduced
axial conductance by up to 98% in lateral roots (hydraulically isolating the plants from soil),
thereby limiting water loss to dry soil. However, plants have evolved mechanisms to restore
xylem transport capacity once embolisms occur (Brodersen and McElrone, 2013). By re-
watering, axial conductance would recover by eliminating the embolism from xylem vessels
(North and Nobel, 1991), and remobilised root constituents from the taproot may supply
energy for the resumption of new shoot growth in tedera.
The long-term responses to water stress may include changes in xylem anatomy and a
reduction in xylem hydraulic conductivity to water flow along the soil–plant–atmosphere
continuum. The formation of tyloses in the vascular system may restrict water uptake in tedera
over summer. Tyloses are outgrowths of xylem parenchyma cells into vessel-parenchyma pit
pairs into the lumen of tracheary elements (Esau, 1948). Canny (1997) proposed that
formation of tyloses is initiated by the frequent embolisation of vulnerable vessels. Structures
that resemble tyloses were found in the xylem vessels of stems in drought-stressed tedera
plant (M J Canny personal communication) which were not observed in the well-watered
controls (Plate 5.5).

CHAPTER 5. GENERAL DISCUSSION
PAGE | 76
Plate 5.5. Cross sections of stems from well-watered control (above) and drought-stressed
tedera plants (below) after withholding water for 47 days. Stems were embedded in glycol
methacrylate and tissue sections viewed under a light microscope. Structures that resemble
tyloses in the xylem vessels of drought-stressed tedera are circled in yellow. Plants were 10
months old.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 77
POST-DROUGHT RECOVERY ABILITY OF TEDERA AND LUCERNE
For a perennial plant species, recovery from drought stress is one of the most relevant survival
strategies (Hu et al., 2010). Tedera and lucerne had different physiological and morphological
strategies to recover from drought stress (Chapter 4). The leaves of tedera accessions which
dehydrate up to a minimum 40–45% RWC were still pliable and green. Maintenance of plant
functions in tedera at low LWP and RWC and the more rapid recovery after re-watering than
lucerne in the field should contribute to more aboveground dry matter yield under cycles of
stress and recovery from stress. Despite more than 16 days of almost full stomatal closure in
both tedera accessions in the drought treatment, severe drought stress did not appear to
impair the vascular capacity for water transport as photosynthesis and gs completely
recovered after re-watering. With this type of effective stomatal control mechanism in tedera,
reversibility is vital for lessening negative impacts on plant function. Since stomatal closure
limits gas exchange, plants would have to open stomata as soon as rainfall occurs or a slow
rate of CO2 entry and photosynthesis would restrict growth. On re-watering, tedera also
rapidly unfolded and unrolled its green leaves, which played an important role in the more
rapid resumption of plant growth. In contrast to tedera, lucerne plants sacrificed all their green
leaves under drought stress, with new shoots and leaves initiated from new basal buds on the
crown of the plant following re-watering.
Tedera evolved in an environment where rainfall events tend to be light (<10 mm d–1) during
the year (Diaz, 2004). As a result, even a small rainfall event in such an ecosystem may induce
a rapid response in tedera to activate plant growth. Importantly, a shallow lateral root system
in tedera should allow small summer-rainfall events to efficiently affect plant water status and
influence processes that determine productivity. In contrast, lucerne does not appear to
respond to small rainfall events (20 mm d–1) during summer months in southern Australia
(Lolicato, 2000) and under severe drought conditions, does not respond to rainfall unless it is
also defoliated (P Dolling personal communication).
Lucerne is likely more deeply rooted than tedera in the field which may also account for its lack
of response to small summer-rainfall events. Also, rather than tolerating severe drought stress
in the field by protecting foliage from excessive dehydration and turgor loss, drought-stressed
plants typically exploit the water when it is available, drop their leaves, then re-foliate when
re-watered. For lucerne then to recover from drought stress, new shoots and leaves must be
initiated following re-watering. Consequently, the growth rate of lucerne was slower than that
of tedera over the four weeks of the recovery phase in the controlled-environment study
(Chapter 4). However, both lucerne and tedera had high biological activity following re-

CHAPTER 5. GENERAL DISCUSSION
PAGE | 78
watering, including increased photosynthetic capacity and new leaf growth, although this did
not compensate for the limitation to plant growth and metabolic activity due to previous
drought. Root dry matter for tedera accessions, but not for lucerne, also declined following re-
watering of drought-stressed plants; remobilised root constituents may supply energy and
nutrients for the resumption of new shoot growth in tedera following re-watering.
SPECIES-SPECIFIC MECHANISMS FOR TEDERA TO COPE WITH ITS NATIVE
ENVIRONMENT
Tedera presumably evolved various drought resistance mechanisms to cope with the drought-
stress patterns under which this species naturally occurs. Species like tedera that are adapted
to semi-arid systems may be expected to display traits associated with water conservation and
light avoidance, such as leaf folding, effective stomatal control and high water-use efficiency.
The greater degree of effective stomatal control in tedera when compared to lucerne
(Chapters 3 and 4) reflects the environment of their native habitats. In the Canary Islands, with
persistent water limitation over summer (Table 5.1), tedera likely maintains a state of minimal
biological activity, such as maintaining a low gas exchange level and effective stomatal closure
so plants can conserve available soil water over summer. Tedera plants likely open their
stomata only to the extent that is required to survive.
The folding and rolling of tedera leaves in the field during summer, while reducing gs on the
abaxial leaf surface, demonstrates a strong interaction between the two functions to decrease
exposure of the upper leaf surface to transpirational water loss. This would be an adaptive
mechanism in this species to water stress. Kao and Tsai (1998) also proposed that
paraheliotropic leaf movements and photosynthetic characteristics evolved in a mutual way in
species adapted to arid environments, so that WUE is enhanced under drought stress. Under
the Mediterranean conditions in the Canary Islands, where drought and high-light stresses
occur simultaneously over the summer period, photoprotection would also be a key
adaptation for evergreen species like tedera. When the leaves of tedera plants were restrained
from moving freely outside during mid-summer, thermal damage was apparent even on well-
watered plants (Plate 5.6).

CHAPTER 5. GENERAL DISCUSSION
PAGE | 79
Plate 5.6. Leaves of well-watered tedera plants restrained from moving while outside in mid-
summer. Note the bleaching of the leaf edges after 24 hours.
While increased root depth is an adaptation to survival in drought conditions for lucerne, for
tedera in its native environment with shallow soils and light rainfall events, an increase in
rooting depth alone in response to drought is unlikely to aid plant survival if water is
unavailable at depth. Growth of a large tap root system in tedera would also consume
photosynthetic end-products and indirectly negate lateral root and shoot growth during water
deficit. It is also doubtful if lucerne would survive with no water in the root zone (Kemp et al.,
1989). That lucerne is not adapted to the native environment of tedera is demonstrated by the
fact that on Lanzarote all lucerne stock feed is imported (Arevalo et al., 2012). This also
highlights that a particular mechanism of drought resistance in a plant will not work for all
drought environments.
These results may appear to contradict the widely-held view that rooting depth increases in
drier environments. However, Blum (1996) questioned the role of increased rooting depth
when there is no water at depth to extract. Kemp and Culvenor (1994) also proposed that
survival in drying soils does not always require deep roots. The disadvantage is, with such an
expansive lateral root system, tedera plants could compete for soil water if growing at high

CHAPTER 5. GENERAL DISCUSSION
PAGE | 80
density. However, tedera is reported to occur at low density on Lanzarote (P Mendez personal
communication) and the shallow root structure in tedera may be more ecologically suited to
low density stands (Plate 5.7). Also, the soil horizons could be repeatedly explored for water
and nutrients by the shallow lateral roots in tedera. In support of this, at several experimental
field sites in WA over several years, no tedera plantings developed into swards (K Foster
personal observations) despite some seedling recruitment being observed.
Table 5.1. Climatic data from the airport of Arrecife, Lanzarote and Newdegate, Western
Australia based on measurements from 1972 to 2000. Mean air temperature is T, Tmin and
Tmax are mean minimum and maximum air temperatures, respectively, rH is relative humidity.
Differences between sum of all months and years are the result of rounding. Adapted from
Graf et al. (2004) and Bureau of Meteorology.
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Year
Lanzarote
T (°C) 17.0 17.5 18.5 19.0 20.2 21.9 23.8 24.7 24.4 22.5 20.3 18.1 20.7
Tmin (°C) 13.7 13.9 14.6 15.0 16.3 18.1 19.9 20.7 20.4 18.7 16.8 14.8 16.9
Tmax (°C) 20.4 21.2 22.5 23.0 24.2 25.7 27.8 28.8 28.4 26.3 23.9 21.4 24.5
Rainfall (mm) 24.0 14.0 15.0 6.0 2.0 0.0 0.0 0.0 2.0 7.0 12.0 27.0 110.0
rH (%) 71.0 71.0 69.0 69.0 69.0 69.0 70.0 71.0 73.0 73.0 72.0 73.0 71.0
VPD (kPa) 0.7 0.7 0.9 0.9 0.9 1.0 1.1 1.2 1.0 0.9 0.8 0.7 0.9
Newdegate
T (°C) 23.1 23.1 21.3. 18. 14.5 11.6 10.4 11.0 12.7 16 19.4 21.4 16.9
Tmin (°C) 15.3 15.5 14.3 11.7 8.8 6.3 5.2 5.4 6.2 8.9 11.8 13.6 10.2
Tmax (°C) 30.8 30.7 28.0 24.0 20.3 17.0 15.6 16.7 19.2 23.1 26.9 29.3 23.5
Rainfall (mm) 24.1 18.7 27.2 14.5 39.3 31.9 41.1 38.1 32.9 30.3 25.7 29.5 342.6
rH (%) 58.0 60.4 63.2 69.4 76.1 81.8 84.2 82.9 76.4 66.5 59.8 58.3 70.0
VPD (kPa) 1.9 1.8 1.3 0.9 0.6 0.3 0.3 0.3 0.5 0.9 1.2 1.7 1.0

CHAPTER 5. GENERAL DISCUSSION
PAGE | 81
Plate 5.7. Two-year old tedera accessions at the Newdegate field site. Tedera plants are often
a hemispheric shape at low density, and this shape is possibly more efficient from a light-
harvesting point of view than an erect lucerne plant.
INTEGRATION OF PERENNIALS INTO SOUTHERN AUSTRALIAN FARMING SYSTEMS
THE CHALLENGE OF ESTABLISHMENT
Perennial seedlings are particularly vulnerable to competition in the first year of
establishment, typically having less vigorous growth than many annual weeds and annual
pasture species (Humphries and Auricht, 2001; Bell et al., 2006). Lucerne seedlings are
reportedly poor competitors, and their slow growth at the seedling stage may be due to their
small seed size (Humphries and Auricht, 2001). Perennials are also expensive to establish;
estimated between $250 and $400/ha (Virgona and Hildebrand, 2006). A major limitation of
phase-farming system adoption has been the high failure rate establishing lucerne pastures
after a cropping phase compared with annual pasture species (Ransom and Barr, 1994).
However, tedera has large seeds (over 30 mg) and seedlings have strong early vigour in the
glasshouse compared to other legumes (Chapter 2). The high seedling vigour of tedera should
allow for early light capture and for seedlings (depending on seedling density) to compete with
annual weeds during establishment (Plate 5.8). This would be a major advantage to the
establishment and integration of a new perennial species such as tedera into phase-farming

CHAPTER 5. GENERAL DISCUSSION
PAGE | 82
systems. The successful and early seedling establishment would reduce both the cost and the
time until first grazing.
Plate 5.8. Tedera seedlings in late autumn (a) and four weeks later in early winter (b)
competing with annual background species.
SHIFT IN SOWING DATES AND DEPTH
The effect of drought stress on tedera can be managed at the seedling stage in conjunction
with farm-level adaptations such as: a) shifting sowing dates earlier to make best use of early
available moisture and warmer soil temperatures; and b) using deeper sowing depths to avoid
rapid soil drying of upper soil layers (Plate 5.9). As discussed in Chapter 2, the superior drought
resistance of tedera seedlings would enable seed planting on limited rainfall early in the
season to achieve early biomass production when the pasture feed base is traditionally low.
This would provide significant economic benefits to farmers by reducing purchased feed
supplements. Economic modelling data have also stressed the importance of early-season

CHAPTER 5. GENERAL DISCUSSION
PAGE | 83
feed; even an extra megajoule of feed from pastures in autumn is worth about 10 times its
value in spring when green feed is usually abundant (J Young personal communication).
Plate 5.9. Tedera pods were directly sown in the field to a depth of 50–70 mm (similar to
cereals) and seedlings successfully emerged. Note some lateral roots were lost from the roots
during removal from the soil.
Alternatively, spring sowing of tedera may have significant advantages over winter sowing for
tedera. The establishment of a new pasture after an extended cropping phase often occurs in
the presence of a large weed seed-bank. Spring sowing would provide an opportunity to
control previously emerged or herbicide-resistant weeds in the cropping phase with a non-
selective herbicide (glyphosate) or for the paddock to be used for heavy winter grazing.
Importantly, this practice of later sowing would also conserve soil moisture by removing more

CHAPTER 5. GENERAL DISCUSSION
PAGE | 84
established plants and allowing a short fallow during seedling establishment where soil
moisture can build. However, the disadvantage is that there is a higher risk that soil water will
run out before perennial plants can establish roots sufficiently to persist over summer (Bell et
al., 2006).
COMPARING WATER STRESS IN THE FIELD AND CONTROLLED-ENVIRONMENT
EXPERIMENTS
In Chapters 2 and 4, controlled-environment experiments were used to examine traits of
drought resistance in tedera to remove the variability present in a field environment.
Encouragingly, the results of the controlled-environment experiments on drought stress for
tedera generally agreed with the findings from the field, i.e. leaf folding and rolling, effective
stomatal control and increased pubescence were key mechanisms of drought resistance.
Grzesiak (1996) also found that drought resistance ratings of legume plants in the laboratory
were overall consistent with the ratings based on estimating the direct effects of soil drought
on yield in field experiments. However, I did find greater resistance to drought in field-grown
mature plants (Chapter 3) than in plants maintained in the controlled-environment experiment
(Chapter 4). The leaves of tedera plants had a lower leaf-folding angle (70°) and higher midday
LWP (–2 MPa) and RWC in the field (77%) in February after severe drought stress (63 days of
rainfall excluded) when compared with the controlled-environment study (47 days of no
watering): leaf angle 85°, LWP –6.5 MPa and RWC 42%. There are several possible hypotheses
to explain this difference. Field-grown plants are likely to have been subjected to one or more
cycles of lower soil water in spring or early summer, and consequently become pre-
conditioned to subsequent drought stress. Alternatively, exposure to moisture stress at the
seedling stage may confer some degree of hardening against later drought periods. It is likely
that plant roots in the field could explore a larger soil volume for stored soil water. This finding
highlights the importance of the velocity of the imposition of drought stress (i.e. faster in the
controlled environment and slower in the field), as both may generate different physiological
results or acclimations.
The implications of relying primarily on deep root systems alone for drought resistance were
clearly observed in the lucerne plants in Chapter 4. In this controlled-environment experiment,
severe leaf desiccation occurred in lucerne three weeks after withholding water, while tedera
leaves were still green and pliable after seven weeks of drought stress. Evans (1977) identified
that lucerne resists drought by extracting water from deeper in the soil in the field, rather than
by efficient control of water use by the plant, and in the controlled-environment study the
roots of lucerne plants were likely restricted by the pot. The restriction of water loss (i.e.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 85
stomatal closure) is not the primary mechanism of drought avoidance for lucerne because this
failed to prevent rapid water loss in the pots in the drought treatment.
LIMITATIONS OF THE RESEARCH PRESENTED IN THIS THESIS
A number of limitations to the experiments reported in this thesis warrant mentioning.
Climatic conditions in the controlled environments (Chapters 2 and 4) are likely to have
differed from those in the field where complex interactions may occur as a result of different
stresses, especially in terms of lower humidity, wind and heat. Root growth may also have
been artificially restricted by pots. Pot experiments on water relations can be difficult to relate
to plants growing in the field, even if field soils are reconstructed and used, because of the loss
of the structural features that can dominate such interactions in the field, such as pre-existing
biopores, which may have different physical or chemical properties from those of the bulk soil
(Pankhurst et al., 2002).
A larger-diameter pot and more seedlings per pot (Chapter 2) would allow for additional
sampling to monitor RWC over time and identify any potential compatible osmolytes at the
seedling stage. For the drought resistance and recovery experiment (Chapter 4), re-watering
occurred while air temperatures were constant and without wetting the upper foliage.
However, in the field, showers often fall when air temperatures are relatively low which also
wet plant foliage. The role of proline in drought stress is still unclear; it reportedly does not
accumulate when plants are exposed to both heat and drought stress simultaneously, and may
be phytotoxic at high temperatures (Rizhsky et al., 2004). Therefore, although proline may play
a direct role in the drought resistance in tedera in its native environment (and in our
controlled-environment study), this compound may not accumulate under both abiotic
stresses of heat (up to 46°C) and drought in the field environment of WA over summer.
FUTURE RESEARCH DIRECTIONS
There are still several gaps in our understanding with regard to drought resistance mechanisms
of tedera in the field. Suggestions for further research are to: a) further characterise stomatal
responses of tedera and lucerne by examining gs between dawn and 08:00 h and after 17:00 h
in summer; b) examine changes to stomatal density on the leaf surfaces of tedera over the
season; c) investigate accumulation of abscisic acid in leaves and roots under moisture deficit;
d) examine if proline accumulates in the leaves of tedera when plants are under both heat and
drought stress in the field (i.e. over summer); e) examine the extent of xylem cavitation at low
water potentials; f) examine changes to photosynthetic pigments such as Chl a and Chl b under
drought stress; g) examine the effects of drought stress on the PSII photochemistry of tedera;

CHAPTER 5. GENERAL DISCUSSION
PAGE | 86
and, h) examine the potential for improving drought resistance by modifying potassium
fertiliser inputs. A better understanding of the mechanisms of drought resistance will allow
specific traits to be targeted in selection/breeding programs and enhance the potential for the
adoption of this species into low-rainfall pasture systems.
The soils on Lanzarote often have low organic matter (0–3%) and low plant available
phosphorus (67 mg kg–1 Olsen P test), but very high available potassium levels (450–3000 mg
kg–1) (Arevalo et. al., 2012). Potassium improves the resistance of plants to environmental
stress, and is involved in physiological processes related to drought resistance, such as leaf
movements, transmission of signals and the maintenance of turgor pressure, and plays a key
role in photosynthesis (Wang et al., 2013). However, potassium levels in many WA wheatbelt
duplex soils are inherently low (Wong, 2001), and current farming practices are rapidly
depleting them. It is possible to measure 1000 mg kg–1 of potassium on some heavy WA
wheatbelt soils, but certainly not on light-textured soils where 100 mg kg–1 is considered good
and 200 mg kg–1 is considered high (B Bowden personal communication). The inherently-low
soil potassium levels in the WA wheatbelt (i.e. <100 mg kg–1 dry soil at Newdegate) is likely to
result in reduced productivity and persistence of many perennial legumes and grass species.
Reproductive failure under extreme drought stress in tedera has been observed in the field
(Plate 5.10). Some tedera accessions, ironically, may be too drought resistant at the expense of
seed production, and this may be related to a whole-plant signalling system involving
hormones (particularly ABA). The plentiful lateral roots of tedera in the field combined with
higher summer daytime temperatures in WA could mean that drying of the upper soil during
commercial seed production (on light-textured soils) is detrimental (i.e. possible abortion of
seed florets from ABA signalling). Also, the higher temperature and lower humidity during
summer (high VPD) in the WA wheatbelt environment may affect tedera’s seed production. A
pollen-based screening of tedera germplasm for drought or high temperatures or ABA
sensitivity may need to be included as part of the breeding program. Also, mechanical root
pruning of lateral roots in the field may increase seed yield in commercial seed crops by
reducing the ABA signalling from shallow lateral roots.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 87
Plate 5.10. A drought-resistant tedera accession under a rain-exclusion shelter showing high
leaf retention and folding but also high levels of flower abortion.
The prospect of slightly higher summer rainfall under climate change may create new
opportunities for growing summer-active perennial crops in some areas in WA. However, it is
critical that the cyclic pattern of defoliation and the effects of assimilate depletion and re-
accumulation in new perennial legumes during exposure to drought stress and re-watering are
investigated. The longevity of perennial legumes is highly dependent on their response to
grazing (Cocks, 2001). Tedera demonstrates good recovery from heavy grazing, although it is
likely that some form of rotational grazing may be required to sustain plant persistence and
production over summer and autumn. Comparative physiological research is necessary in
Lanzarote over summer to fully understand how tedera is adapted to the WA environment,
with possible changes in root morphology (i.e. extent that root growth is redistributed to
greater soil depths as soil water is depleted) on our lighter textured and deeper soils.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 88
CONCLUDING COMMENTS
The primary role of the leaf movements (heliotropism) in tedera seedlings and mature plants is
to regulate the amount of incident light on the leaf surface, maximising it in the morning and
late afternoon on sunlit leaves. Consequently, these leaf movements have a direct effect on
leaf temperature, transpirational water loss, carbon gain, WUE and the leaf energy budget,
while likely also reducing photoinhibition of the photosynthetic apparatus. Drought resistance
at the seedling stage in tedera accessions may also correlate with the performance of mature
plants in the field. If so, screening at the seedling stage for tedera could be a simple and
efficient method to select for drought-resistant accessions from large germplasm collections.
Effective stomatal control in tedera is a promising physiological trait to screen large numbers
of new accessions for drought resistance using a hand-held poromoter. This method is non-
destructive, has relatively quick-throughput, and precise instruments (Decagon™ poromoter)
for the measurement of conductance are available. Infrared thermometry is rapid enough to
also be a potential selection tool, and can serve as an effective surrogate for stomatal
conductance measurements.
Lucerne displays a substantially different gs response to drought than tedera both in the field
and in controlled-environment studies which raises the question as to how conclusions derived
from studies on the response of lucerne alone to drought can be widely generalised to other
plant species, especially undomesticated perennial legume species like tedera. The stomatal
response patterns of tedera accessions to drought stress during the day may also have
implications, from a modelling perspective, in determining the plant survival response to
naturally-fluctuating environmental conditions (where control of gs is a function of the model)
or in future climate-change scenarios.
For current southern Australian farming systems, this research has shown that the drought
resistance mechanisms of tedera allow it to not only survive, but remain productive, over the
summer drought, hence offering substantial gains in sustainability and productivity over
lucerne. Tedera could be used as a permanent pasture in low-productive areas of the WA
wheatbelt where intensive cropping is unsuitable (now or in the future) or as pasture in an
extended phase system.
The highly variable climatic conditions typical of the WA Mediterranean climate have always
posed severe limitations on agriculture and to livestock production. However, the
sustainability of southern Australia’s dryland farming systems is likely under threat, due to the
predicted increases in seasonal variability as climate change impacts are realised. If the drying

CHAPTER 5. GENERAL DISCUSSION
PAGE | 89
trends continue as projected, large areas of the WA wheatbelt currently used for cropping may
not be sustainable for this purpose.
This study of morphological and physiological response to drought stress and re-watering in
tedera has shown that many mechanisms coexist and contribute to whole-plant acclimation
and resistance to drought stress. This research has shown that tedera is well adapted to
drought, both as a seedling and a mature plant, and hence may help to sustain the productivity
of new pasture systems in the face of predicted climate change. However, drought is not one
simple stress. The agronomic practices that seek to adapt to the predicted climate change
must take into account multiple aspects of both the stress induced by water deficit and other
interacting stresses such as heat, low-nutrient status, and the acidity of many wheatbelt soils.
Nevertheless, in plant breeding now, breeders need to work more closely with plant
physiologists to develop more drought-resistant perennial legumes and to take a longer-term
view (up to 20–30 years) and not focus too strongly on the drought conditions that we
experience now, but prepare for the likely droughts in the future.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 90
REFERENCES
Aasamaa K, Sober A (2011) Stomatal sensitivities to changes in leaf water potential, air
humidity, CO2 concentration and light intensity, and the effect of abscisic acid on the
sensitivities in six temperate deciduous tree species. Environmental and Experimental
Botany 71, 72–78.
ABARE (Australian Bureau of Agricultural and Resource Economics Climate change). Impacts on
Australian Agriculture. December, 2007.
ABS (Australian Bureau of Statistics). Year Book Australia. No. 90, Catalogue No. 1301.0
Australian Bureau of Statistics, Canberra. 2008.
Arve LE, Torre S, Olsen JE, Tanino KK (2011) Stomatal responses to drought stress and air
humidity, abiotic stress in plants — mechanisms and adaptations, (Ed A Shanker),
Available from: http://www.intechopen.com/books/abiotic-stress-in-plants-
mechanismsandadaptations/stomatal-responses-to-drought-stress-and-air-humidity.
Arevalo JR, Mora JL, Chinea E (2012) Forage quality of native pastures on Lanzarote Island
(Canary Islands). Journal of Food, Agriculture and Environment 10(1), 696–701.
Ashley J (1993) Drought and crop adaptation. In ‘Dryland Farming in Africa’ (Ed JRJ Rowland) 3,
pp. 47–66. (Macmillan Press Ltd, UK).
Bathgate A, Pannell DJ (2002) Economics of deep-rooted perennials in Western Australia.
Agricultural Water Management 53(1–3), 117–132.
Bell LW, Ryan MH, Moore GA, Ewing MA (2006) Comparative water use by Dorycnium
hirsutum, lucerne, and annual-based pastures in the Western Australian wheatbelt.
Australian Journal of Agricultural Research 57, 857–865.
Bennett E (1970) Adaptation in Wild and Cultivated Plant Populations. In ‘Genetic Resources in
Plants—Their Exploration and Cultivation’. (Eds OH Frankel, E Bennett). IBP Handbook No.
11 pp. 115–129. (Blackwell Scientific Publications: Oxford, UK).
Biddiscombe EF, Paterson JG, Rogers AL, Nicholas DA, Glencross RN, Arkell PT, Anderson GW
(1982) Perennial pasture grasses in south western Australia. 4. Potential areas of use. WA
Department of Agriculture, Technical Bulletin No 60.
Bielenberg D, Miller J, Berg S (2003) Paraheliotropism in two Phaseolus species: combined
effects of photon flux density and pulvinus temperature, and consequences for leaf gas
exchange. Environmental and Experimental Botany 49, 95–105.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 91
Blum A (1989) Osmotic adjustment and growth of barley genotypes under drought stress. Crop
Science 29, 230–233.
Blum A (1996) Crop responses to drought and the interpretation of adaptation. Plant Growth
Regulation 20(2), 135–148.
Bouizgaren A, Kallida R, Al Faiz C (2010) Evaluation of drought tolerance variability in
Mediterranean alfalfa populations in the field under Moroccan conditions. In 'Sustainable
Use of Genetic Diversity in Forage and Turf Breeding' (Ed C Huyghe) pp. 283–288.
(Springer Science+Business Media B.V.).
Brodersen C, McElrone A (2013) Maintenance of xylem network transport capacity: a review of
embolism repair in vascular plants. Frontiers in Plant Science 4, 108.
Cai YF, Zhanf SB, Hu H, Li SY (2010) Photosynthetic performance and acclimation of Incarvillea
delavayi to water stress. Biologia Plantarum 54(1), 89–96.
Canadell J, Zedler PH, (1995) Underground structures of woody plants in Mediterranean
Ecosystems of Australia, California and Chile. In ‘Ecology and Biogeography of
Mediterranean Ecosystems in Chile, California and Australia’ (Eds MT Arroyo, PH Zedler,
MD Fox) pp. 177–210. (Springer Verlag, Berlin).
Canny MJ (1997) Tyloses and the maintenance of transpiration. Annals of Botany 80, 565–570.
Cattivelli L, Rizza F, Badeck F, Mazzucotelli E, Mastrangelo A, Francia E, Mare C, Tondelli A,
Michele S (2008) Drought tolerance improvement in crop plants: An integrated view from
breeding to genomics. Field Crops Research 105(1–2), 1–14.
Chandler J, Bartels D (2003) Drought avoidance and drought adaptation. In ‘Encyclopaedia
Water Science’ (Eds BA Stewart, TA Howell) pp. 163–165. (Marcel Dekker.
Chaves M, Maroco J, Pereora J (2003) Understanding plant response to drought from genes to
the whole plant. Functional Plant Biology 30, 239–264.
Cocks PS (2001) Ecology of herbaceous perennial legumes: a review of characteristics that may
provide management options for the control of salinity and waterlogging in dryland
cropping systems. Australian Journal of Agricultural Research 52, 137–151.
Cowan I (1982) Regulation of water use in relation to carbon gain in higher plants.
Encyclopaedia of Plant Physiology 12, 589–612.
Cutforth HW, McGinn SM, McPhee KE, Miller PR (2007) Adaptation of pulse crops to the
changing climate of the Northern Great Plains. Agronomy Journal 99, 1684–1699.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 92
Dear BS, Moore GA, Hughes SJ (2003) Adaptation and potential contribution of temperate
perennial legumes to the southern Australian wheatbelt: a review. Australian Journal of
Experimental Agriculture 43, 1–18.
Dear BS, Ewing MA (2008) The search for new pasture plants to achieve more sustainable
production systems in southern Australia. Australian Journal of Experimental Agriculture
48, 387–396.
Díaz F (2004) Sistemas agrícolas tradicionales de las zonas áridas de las Islas Canarias. Tesis
Doctoral. Universidad de La Laguna.
Erice G, Irigoyen JJ, Sanchez-Díaz M, Avice JC, Ourry A (2007) Effect of drought, elevated CO2
and temperature on accumulation of N and vegetative storage proteins (VSP) in taproot of
nodulated alfalfa before and after cutting. Plant Science 172, 903–912.
Esau K (1948) Anatomic effects of the viruses of Pierce's disease and phony peach. Hilgardia
18, 423–482.
Evans PS (1977) Root distribution and water-withdrawal patterns of some crop and pasture
species. Department of Scientific and Industrial Research Information Series. Proceedings
of the Soil and Plant Water Symposium, Palmerston North 126, 186–190.
Fahn A (1989) Plant Anatomy, 3rd Edition. Pergamon Press, Oxford.
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects,
mechanisms and management. Agronomy for Sustainable Development 29, 185–212.
Filippou P, Antoniou C, Fotopoulos V (2011) Effect of drought and rewatering on the cellular
status and antioxidant response of Medicago truncatula plants. Plant Signaling and
Behavior 6, 270–277.
Finlayson JD, Real D, Nordblom T, Revell C, Ewing MA, Kingwell R (2012) Farm level
assessments of a novel drought tolerant forage: Tedera (Bituminaria bituminosa C.H. Stirt
var. albomarginata). Agricultural Systems 112, 38–47.
Frechilla S, Royuela M, Arrese-Igor C, Aparicio-Tejo PM (1993) Effect of osmotically-applied
water stress or drought on photosynthesis in Pisum sativum. Plant Physiology 102, 177
(Suppl).
Gebre G, Tschaplinski T (2000) Environmental sciences role of osmotic adjustment in plant
productivity. Summary Report and Review of Current Literature. Colorado State
University. Publication No. 4961.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 93
Graf A, Kuttler W, Werner J (2004) Dewfall measurements on Lanzarote, Canary Islands.
Meteorologische Zeitschrift 13, 405–412.
Grzesiak S, Filek W, Skrudlik G, Nizioł B (1996) Screening for drought tolerance: Evaluation of
seed germination and seedling growth for drought resistance in legume plants. Journal of
Agronomy and Crop Science 177, 245–252.
Hennessy KB, Fitzharris BC, Bates N, Harvey SM, Howden L, Hughes J, Salinger R, Warrick
(2007) Australia and New Zealand. Climate Change 2007: Impacts, Adaptation and
Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the
Intergovernmental Panel on Climate Change. (Eds ML Parry, OF Canziani, JP Palutiko, J van
der Linden, CE Hanson) pp. 507–540 (Cambridge University Press, Cambridge, UK).
Hill MJ (1996) Potential adaptation zones for temperate pasture species as constrained by
climate: a knowledge-based logical modelling approach. Australian Journal of Agricultural
Research 47, 1095–1107.
Hu HH, Boisson-Dernier A, Israelsson-Nordstrom M, Bohmer M, Xue SW, Ries A, Godoski J,
Kuhn JM, Schroeder JI (2010) Carbonic anhydrases are upstream regulators of CO2-
controlled stomatal movements in guard cells. Nature Cell Biology 12, 87–93.
Hulme ME, Barrow M, Arnell NW, Harrison PA, Johns TC, Downing TE (1999) Relative impacts
of human-induced climate change and natural climate variability. Nature 397(6721), 688–
691.
Humphries A, Auricht G (2001) Breeding lucerne for Australia's southern dryland cropping
environments. Australian Journal of Agricultural Research 52, 153–169.
Irigoyen JJ, Emerich DW, Sánchez-Díaz M (1992) Alfalfa leaf senescence induced by drought
stress: photosynthesis, hydrogen peroxide metabolism, lipid peroxidation and ethylene
evolution. Physiologia Plantarum 84, 67–72.
IPCC (Intergovernmental Panel on Climate Change) (2007) Fourth Assessment Report of
Working Group I. Climate Change 2007: The Physical Science Basis, Summary for
Policymakers. (Eds S Solomon, D Qin, M Manning, Z Chen, M Marquis, KB Averyt, M
Tignor, HL Miller) (Cambridge University Press, Cambridge, United Kingdom and New
York, NY, USA).
Jarvis PG, McNaughton KG (1986) Stomatal control of transpiration: scaling up from leaf to
region. Advances in Ecological Research 15, 1–49.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 94
Johansen C, Baldev B, Brouwer JB, Erskine W, Jermyn WA, Li-Juan L, Malik BA, Ahad Miah, A,
Silim SN (1992) Biotic and abiotic stresses constraining productivity of cool season food
legumes in Asia, Africa and Oceania. In ‘Expanding the production and use of cool season
food legumes’ (Eds FJ Muehlbauer, WJ Kaiser) pp. 175–194 (Kluwer: Dordrecht).
Kao WY, Tsai TT (1998) Tropic leaf movements, photosynthetic gas exchange, leaf δ13C and
chlorophyll a fluorescence of three soybean species in response to water availability.
Plant, Cell & Environment 21, 1055–1062.
Kaufman MR, Eckard AN (1971) Evaluation of water stress control with PEG by analysis of
guttation. Plant Physiology 47, 453–458.
Kemp DR, Nicholas DA, Oldham CM (1989) New discoveries in pasture research: current status
& developments with temperate perennial pastures and shrubs in southern Australia.
Proceesings 5th Australian Agronomy Conference, 96–107.
Kemp DR, Culvenor RA (1994) Improving the grazing and drought tolerance of temperate
perennial grasses. New Zealand Journal of Agricultural Research 37, 365–378.
Kiani SP, Talia P, Maury P, Grieu P, Heinz R, Perrault A, Nishinakamasu V, Hopp E, Gentzbittel L,
Paniego N, Sarrafi A (2007) Genetic analysis of plant water status and osmotic adjustment
in recombinant inbred lines of sunflower under two water treatments. Plant Science 172,
773–787.
Koller D (1986) The control of leaf orientation by light. Journal of Photochemistry and
Photobiology 44, 819–826.
Kramer P (1980) Drought stress and the origin of adaptations. In ‘Adaptation of plants to water
and high temperature stress’. (Eds N Turner, P Kramer) pp. 7–20 (Wiley: New York).
Lambers H, Chapin III F, Pons T (2008) Plant Physiological Ecology. 2nd edition. Springer. New
York.
Lawlor DW (1970) Absorption of polyethylene glycols by plants and their effects on plant
growth. New Phytologist 69, 501–503.
Lawlor DW (2001) Photosynthesis, 3rd edition. Oxford. Bios Scientific Publishers.
Lawlor DW (2013) Genetic engineering to improve plant performance under drought:
physiological evaluation of achievements, limitations, and possibilities. Journal of
Experimental Botany 64(1), 83–108.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 95
Lolicato SJ (2000) Soil water dynamics and growth of perennial pasture species for dryland
salinity control. Australian Journal of Experimental Agriculture 40, 37–45.
Lu Z, Quiñones MA, Zeiger E (1993) Abaxial and adaxial stomata from Pima cotton (Gossypium
barbadense L.) differ in their pigment content and sensitivity to light quality. Plant, Cell &
Environment 16(7), 851–858.
Ludlow M (1989) Strategies of responses to water stress. In ‘Structural and functional
responses to environmental stresses’. (Eds K Krebb, H Richter, T Hickley) pp. 269–291 (SPB
Academic the Hague).
Ludlow M, Björkman O (1984) Paraheliotropic leaf movement in Siratro as a protective
mechanism against drought induced damage to primary photosynthetic reactions:
damage by excessive light and heat. Planta 161, 505–518.
Martínez-Fernández D, Walker DJ, Romero P, Correal E (2010) The physiology of drought
tolerance in tedera (Bituminaria bituminosa) In ‘The contributions of grasslands to the
conservation of Mediterranean biodiversity Zaragoza: (Eds C Porqueddu S Ríos)
CIHEAM/CIBIO/FAO/SEEP (Options Méditerranéennes: Série A. Séminaires
Méditerranéens; n. 92) pp.155–159.
Marshall EM, Humphries AW, Kobelt ET, Rowe TD, Auricht GC (2011) Drought response of
lucerne in south east & mallee environments in SA.
http://www.sardi.sa.gov.au/pastures/perennial_pastures/lucerne_breeding_projects/res
ponse_of_lucerne_to_drought. Data retrieved on 10 November 2011.
McManus MT, Bieleski RL, Caradus JR, Barker DJ (2000) Pinitol accumulation in mature leaves
of white clover in response to a water deficit. Environmental and Experimental Botany 43,
11–18.
Medrano H, Escalona JM, Bota J, Gulias J, Flexas J (2002) Regulation of photosynthesis of C3
plants in response to progressive drought: stomatal conductance as a reference
parameter. Annals of Botany 89, 895–905.
Meinzer FC, Johnson DM, Lachenbruch B, McCulloh KA, Woodruff DR (2009) Xylem hydraulic
safety margins in woody plants: coordination of stomatal control of xylem tension with
hydraulic capacitance. Functional Ecology 23, 922–930.
Moles AT, Westoby M (2004) What do seedlings die from, and what are the implications for
evolution of seed size? Oikos 106, 193–199.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 96
Moore G, Dolling P, Porter B, Leonard L (1998) Soil acidity. In ‘Soil guide. A handbook for
understanding and managing agricultural soils’. (Ed G Moore) pp. 127–140 (Department
of Agriculture and Food Western Australia).
Moore G, Sanford P, Wiley T (2006) Perennial pastures for Western Australia. Department of
Agriculture and Food Western Australia, Bulletin 4690, Perth.
Munns R, Brady CJ, Barlow EWR (1979) Solute accumulation in the apex and leaves of wheat
during water-stress. Australian Journal of Plant Physiology 6, 379–389.
Nejad AR, van Meeteren U (2007) The role of abscisic acid in disturbed stomatal response
characteristics of Tradescantia virginiana during growth at high relative air humidity.
Journal of Experimental Botany 58(3), 627–636.
Nicol DL, Finlayson J, Colmer TD, Ryan MH (2013) Opportunistic Mediterranean agriculture –
Using ephemeral pasture legumes to utilize summer rainfall. Agricultural Systems 120,
76–84.
Nobel PS, Huang B (1992) Hydraulic and structural changes for lateral roots of desert
succulents in response to soil drying and rewetting. International Journal of Plant Sciences
153, S163–S170.
North GB, Nobel PS (1991) Changes in hydraulic conductivity and anatomy caused by drying
and rewetting roots of Agave deserti (Agavaceae). American Journal of Botany 78, 906–
915.
O’Toole JS, Cruz RT (1980) Response of leaf water potential, stomatal resistance, and leaf
rolling to water stress. Plant Physiology 65(3), 428–432.
Pang J, Yang J, Ward P, Siddique KM, Lambers H, Tibbett M, Ryan M (2011) Contrasting
responses to drought stress in herbaceous perennial legumes. Plant and Soil 348(1–2),
299–314.
Pankhurst CE, Kirkby CA, Hawke BG (2002) Impact of a change in tillage and crop residue
management practice on soil chemical and microbiological properties in a cereal-
producing red duplex soil in NSW. Australia. Biology and Fertility of Soils 35, 189–196.
Passioura JB (2006) The perils of pot experiments. Functional Plant Biology 33, 1075–1079.
Petit J, Jouzel D, Raynaud N, Barkov J, Barnola I, Basile M, Bender J, Chappella M, Davis G,
Delaygue M, Delmotte V, Kotlyakov M, Legrand V, Lipenkov C, Lorius L, Pepin C, Ritz E,
Saltzman E, Stievenard M (1999) Climate and atmospheric history of the past 420,000
years from the Vostok ice core, Antarctica. Nature 399, 429–436.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 97
Poorter H, Bühler J, van Dusschoten D, Climent J, Postma JA (2012) Pot size matters: a meta-
analysis of the effects of rooting volume on plant growth. Functional Plant Biology 39,
839–850.
Quézel P, Medail F (1995) La région circuí-méditerranéene, centre mundial majeur de
biodiversité végétale. In 'Actes des 6èmes rencontres de l’Agence Régionale pour
l’Environment, Provence-Alpes-Côte d’Azur' pp. 152–160 (Gap).
Ransom K, Barr N (1994) The adoption of dryland lucerne in North-Central Victoria. Research
Report Series No. 151, Department of Agriculture, Melbourne.
Real D, Correal E, Méndez P, Ríos Ruiz S, Sternberg M, Dini-Papanastasi O, Pecetti L, Tava A
(2009) Grassland species profiles. Bituminaria bituminosa C.H. Stirton (synonym: Psoralea
bituminosa L.). (Food and Agriculture Organization: Rome) Available
at:http://www.fao.org/ag/AGP/agpc/doc/Gbase/new_species/tedera/bitbit.htm.
Reyes-Betancort JA, Wildpret W, León Arencibia MC (2001) The vegetation of Lanzarote
(Canary Islands). Phytocoenologia 31(2), 185–247.
Richards RA (1997) Defining selection criteria to improve yield under drought. In ‘Drought
Tolerance in Higher Plants: Genetical, Physiological and Molecular Biological Analysis’. (Ed
E Belhassen) pp.79–88 (Kluwer Academic Publishers, Dordrecht, The Netherlands).
Richards RA, Rebetzke GJ, Watt M, Condon A, Spielmeyer W, Dolferus R (2010) Breeding for
improved water productivity in temperate cereals: phenotyping, quantitative trait loci,
markers and the selection environment. Functional Plant Biology 37(2), 85–97.
Rizhsky L, Liang H, Shuman J, Shulaev V, Davletova S, Mittler R (2004) When defense pathways
collide. The response of Arabidopsis to a combination of drought and heat stress. Plant
Physiology 134(4), 1683–1696.
Robertson M (2006) Lucerne prospects: Drivers for widespread adoption of lucerne for profit
and salinity management. Cooperative Research Centre for Plant-based Management of
Dryland Salinity, Perth.
Rosenberg N (1980) Drought in the Great Plains: Research on Impacts and Strategies. Water
Resources Publications. Proceedings of the Workshop on Research in Great Plains Drought
Management Strategies held at The University of Nebraska-Lincoln March 26–28, 1979.
Sanchez FJ, Manzanares M, De-Andres EF, Tenorio J, Ayerbe L (1998) Turgor maintenance,
osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in
response to water stress. Field Crops Research 59(3), 225–235.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 98
Sanderson MA, Stair DW, Hussey MA (1997) Physiological and morphological responses of
perennial forages to stress. Advances in Agronomy 59, 171–224.
Sandquist DR, Ehleringer JR (2003) Population and family-level variation of brittlebush (Encelia
farinosa, Asteraceae) pubescence: its relation to drought and implications for selection in
variable environments. American Journal of Botany 90, 1481–1486.
Sengupta S, Patra B, Ray S, Majumder AL (2008) Inositol methyl tranferase from a halophytic
wild rice, Porteresia coarctata Roxb. (Tateoka): Regulation of pinitol synthesis under
abiotic stress. Plant, Cell & Environment 31(10), 1442–1459.
Serraj R, Sinclair TR (2002) Osmolyte accumulation: can it really help increase crop yield under
drought conditions? Plant, Cell Environment 25, 333–341.
Smith F, Cocks P, Ewing M (1998) Seed production in cluster clover (Trifolium glomeratum L.).
2. Effect of sowing time and sowing rate on flowering time, abortion, seed size, and
hardseededness. Australian Journal of Agricultural Research 49, 965–971.
Stewart CR (1981) Proline accumulation: Biochemical aspects. In ‘Physiology and Biochemistry
of drought resistance in plants’. (Eds LG Paleg, D Aspinall) pp. 243–251.
Su K, Bremer DJ, Keeley SJ, Fry JD (2008) Rooting characteristics and canopy responses to
drought of turfgrasses including hybrid bluegrasses. Agronomy Journal 100, 949–956.
Subbarao GV, Johansen C, Slinkard AE, Rao RC, Saxena NP, Chauhan YS (1995) Strategies for
improving drought resistance in grain legumes. Critical Reviews in Plant Sciences 14, 469–
523.
Suriyagoda LDB, Real D, Renton M, Lambers H, Ryan MH (2013) Establishment, survival, and
herbage production of novel, summer-active perennial pasture legumes in the low-rainfall
cropping zone of Western Australia as affected by plant density and cutting frequency.
Crop and Pasture Science 64, 71–85.
Szabados L, Kovács H, Zilberstein A, Bouchereau A (2011) Chapter 4. Plants in extreme
environments: Importance of protective compounds in stress tolerance. Advances in
Botanical Research 57, 105–150.
Tadross M, Suarez P, Lotsch A, Hachigonta S, Mdoka M, Unganai L, Lucio F, Kamdonyo
D,Muchinda M (2007) Changes in growing-season rainfall characteristics and downscaled
scenarios of change over southern Africa: Implications for growing maize. In ‘IPCC
Regional Expert Meeting on Regional Impacts, Adaptation, Vulnerability, and Mitigation’
pp. 193–204, Nadi, Fiji, June 20–22, 2007.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 99
Trejo CL, Clephan A, Davies WJ (1995) How do stomata read abscisic acid signal? Plant
Physiology 109, 803–811.
Tuberosa R, Grillo S, Ellis R (2003) Unravelling the genetic basis of drought tolerance in crops.
In ‘Abiotic stresses in plants’. (Eds L di Toppi, B Pawlik-Skowronska) pp. 71–122 (London:
Kluwer Academic Publishers).
Turner NC (1986) Crop water deficits: a decade of progress. Advances in Agronomy 39, 1–51.
Turner NC, Wright GC, Siddique KHM (2001) Adaptation of grain legumes (pulses) to water
limited environments. Advances in Agronomy 71, 193–231.
Turner NC, Abbo S, Berger JD, Chaturvedi SK, French RJ, Ludwig C, Mannur DM, Singh SJ,
Yadava HS (2007) Osmotic adjustment in chickpea (Cicer arietinum L.) results in no yield
benefit under terminal drought. Journal of Experimental Botany 58, 187–194.
Turner NC, Molyneux N, Yan S, Xiong YC, Siddique KHM (2011) Climate change in south-west
Australia and north-west China: challenges and opportunities for crop production. Crop &
Pasture Science 62, 445–456.
Vadez V, Kholova J, Choudhary S, Zindy P, Terrier M, Krishnamurtrty L, Kumar PR, Turner NC
(2011) Whole plant response to drought under climate change. In ‘Crop Adaptation to
Climate Change’. (Eds SS Yadav, R Redden, JL Hatfield, H Lotze-Campen, A Hall) pp. 186–
197 (Wiley-Blackwell).
Virgona J, Hilderbrand S (2006) Survey of perennial pasture establishment practices in
southern NSW. Proceedings of the 13th Australian Agronomy Conference, Western
Australia Groundbreaking Stuff. (Eds Neil Turner, Tina Acuna). 10–14 September 2006
Perth.
Wang H, Yamauchi A (2006) Growth and functions of roots under abiotic stress in soil. In
‘Plant–Environment Interactions, 3rd edition’. (Ed B Huang) pp. 271–320 (CRC Press, New
York).
Wang M, Zheng Q, Shenm Q, Guo S (2013) The critical role of potassium in plant stress
response. International Journal of Molecular Sciences 14, 7370–7390.
Waseem M, Asghar Ali, Tahir M, Nadeem MA, Ayub M, Tanveer Asif, Ahmad R, Hussain M
(2011) Mechanism of drought tolerance in plant and its management through different
methods. Continental Journal of Agricultural Science 5(1), 10–25.
Wong M (2001) Sustain potassium reserves to enhance yields. Farming Ahead No. 118.
Kondinin Group. 40–41.

CHAPTER 5. GENERAL DISCUSSION
PAGE | 100
Xu ZZ, Zhou GS (2008) Responses of leaf stomatal density to water status and its relationship
with photosynthesis in a grass. Journal Experimental Botany 59, 3317–3325.
Zhang J, Davies WJ (1990) Changes in the concentration of ABA in the xylem sap as a function
of changing soil water status can account for changes in leaf conductance and growth.
Plant, Cell and Environment 13, 277–285.





























