Embed Size (px)
Citation preview

Cur J Biochem 118, 61 -70 (1981) ( FEBS 1981
Biosynthesis of the Light-Harvesting Chlorophyll a/b Protein Polypeptide Turnover in Darkness
John BENNETT
Department of Biological Sciences, University of Warwick, Coventry
(Received February 17, 1981)
1. When etiolated pea seedlings were exposed to continuous light for 24 h and then returned to darkness, 38 ”/, of the chlorophyll a, 74 of the light-harvesting chlorophyll a/b protein that had accumulated under illumination proved to be unstable in darkness. The unstable chlorophyll displayed a half-life of about 90 min. In contrast, the CI and a subunits of the chloroplast coupling factor and the large and small subunits of ribulose 1,5-bisphosphate carboxylase continued to accumulate in darkness, although at a slower rate than in plants maintained under light.
2. Short-term labelling in vivo with ~-[~’S]methionine showed that leaves continued to synthesize the light- harvesting protein and the small subunit of ribulose 1,5-bisphosphate carboxylase for up to 48 h after transfer of plants from light to darkness. However, after long-term labelling (16 h), the light-harvesting chlorophyll a/h protein was found to be labelled to high specific activity only in illuminated leaves.
3. I conclude that the light-harvesting chlorophyll a/h protein is subject to turnover after transfer of plants from light to darkness. The site of breakdown appears to be the photosynthetic membrane. I suggest that turn- over of the protein is part of the normal physiological mechanism for co-ordinating the accumulation of the pigment and protein components of the light-harvesting chlorophyll a/b complex.
of the chlorophyll h and 84
The chlorophyll molecules of the chloroplast photosyn- thetic membrane are organized within the lipid bilayer as complexes with specific polypeptides [l]. In green algae and higher plants, the most abundant chlorophyll-binding poly- peptide is termed the light-harvesting chlorophyll a/h protein [l, 21. There is considerable current interest in the structure, function and biosynthesis of this protein. In the present paper I examine the relationship between the biosynthesis of the pro- tein and the biosynthesis of chlorophyll.
In many species of plant there are at least two related forms of the light-harvesting chlorophyll a/b protein [3- 51. The forms differ in molecular weight (within the range 21 500 - 26 000) and in relative abundance. The function of the protein is to organize about one-third of the chlorophyll u and possibly all of the chlorophyll b molecules of the mem- brane into complexes that absorb light and transfer the result- ing excitation energy to the two photosystems [6,7]. The direction of excitation energy transfer is controlled by rever- sible phosphorylation of the light-harvesting protein [8]. In the phosphorylated state, the complex transfers excitation energy preferentially to photosystem I, while, in the dephos- phorylated state, the complex transfers excitation energy preferentially to photosystem 11.
The light-harvesting chlorophyll ajb protein is encoded in the nucleus and synthesized in the cytoplasm as a precursor molecule [9 - 131. The major light-harvesting protein of barley _______.
Abbreviation. CF1, chloroplast coupling factor. Eniyme.7. ~-Ribulose 1,5-bisphosphate carboxylase (EC 4.1.1.39);
Staphylococcus aureus V8 protease (EC 3.4.21.19).
has a molecular weight of about 25000 while the molecular weight of its precursor is 29500 [lo]. The precursor is trans- ported into chloroplasts by a post-translational mechanism and cleaved to the mature molecular weight [ l l , 121. Although transport, cleavage, membrane-attachment and the binding of chlorophyll are known to occur, the order in which these events take place is not clear. For example, it is not known whether cleavage takes place during or after transport of the precursor across the double envelope of the chloroplast.
Regulation of light-harvesting chlorophyll a/b protein synthesis is believed to involve light at two stages. Firstly, the level of the corresponding mRNA, as judged by translation in vitro, is increased by illumination and decreased by a sub- sequent dark period [13-151. It is hot known whether the increase in the level of the mRNA reflects increased tran- scription or processing, or decreased breakdown. However, it is clear that the photoreceptor mediating this response to light is phytochrome [13]. A single flash of red light is sufficient to initiate the appearance of the mRNA for the light-harvest- ing protein in barley.
The second photocontrol is mediated by protochlorophyl- lide, a late intermediate in the biosynthesis of chlorophyll [16]. The reduction of protochlorophyllide to chlorophyllide by the NADPH-dependent enzyme protochlorophyllide reduc- tase is dependent in angiosperms on the excitation of the protochlorophyllide molecule itself [I 71. Continuous synthesis of chlorophyll therefore requires continuous illumination. In barley leaves that have received a single flash of red light before being returned to darkness, the mRNA for the light- harvesting protein accumulates over a period of several hours

62
but the protein cannot be detected [13]. The protein accumu- lates only under continuous illumination. In mutants of barley that contain chlorophyll a but lack chlorophyll b, the major light-harvesting polypeptide cannot be detected even in leaves exposed to continuous illumination [lo, 18,191, a fact that has led to the notion that accumulation of the protein is governed by the synthesis of chlorophyll b rather than of chlorophyllide or chlorophyll a. Support for this notion comes from the observation that pea and bean plants grown under intermittent illumination (such as 2 min of white light every 2 min for 2 days) synthesize low levels of chlorophyll a but seem to be devoid of chlorophyll b and the light-harvesting polypeptides [20,21]. In addition, in the leaves of lettuce hearts, a similar situation is found: the dim light received by the leaves is sufficient to permit formation of low levels of chlorophyll a but not chlorophyll b or the light-harvesting apo-proteins [22].
It would be very reasonable to conclude from these results that the synthesis of the light-harvesting chlorophyll ajb pro- tein is controlled by the synthesis of chlorophyll b. However, the results in the present paper offer a different explanation for these observations : light, through the synthesis of chloro- phyll a and chlorophyll b, prevents turnover of the light- harvesting apo-protein in the photosynthetic membrane. Most studies on the regulation of the formation of the light- harvesting complex have dealt with the on-set of synthesis in response to illumination or with mutants unable to synthesize the complex. In this study I have examined the response of greening pea leaves actively synthesizing the light-harvesting apparatus to the sudden inhibition of chlorophyll synthesis by darkness. The results point very clearly to the existence of turnover of the apo-protein in darkness.
MATERIALS AND METHODS
Plants
Seeds of pea (Pisum sativum L. var. Feltham First) were sown in damp potting fibre and grown for seven days in dark- ness at 20-22°C. They were then illuminated for up to six days under continuous white light (cool fluorescent tubes, photosynthetically active radiation = 100 pmol quanta m-' s-l) and then returned to darkness.
Chlorophyll Determination
The chlorophyll content of the third leaf pair of pea seed- lings was determined in triplicate on groups of five plants. Leaves were ground in a mortar with 5 ml of 90% (v/v) aqueous acetone and the homogenate was made up to 10 ml with the same solvent. After 30 min at room temperature insoluble material was removed by centrifugation at 2000 x g for 5 min. The supernatant was diluted with 80% aqueous acetone and the visible absorption spectrum was recorded with a SP800 spectrophotometer (Pye Unicam Ltd, Cam- bridge, UK). The chlorophyll a and b contents were deter- mined from the equations given in [23].
Assay of Polypeptide Content by Staining of Polyacrylamide Gels
The chlorophyll content of the third leaf pair was deter- mined by acetone extraction of a small sample of leaves as described above. The remaining third leaves were homog- enized in a cold mortar with 10 ml of SO mM Tricine/KOH
pH 8.0, 5 mM 2-mercaptoethanol, filtered through one layer of Miracloth (Calbiochem, San Diego, CA, USA) and centri- fuged at 30000 x g for 10 min. The pellet (representing a crude membrane fraction) was taken up in a volume of homogenis- ing buffer equal to the volume of the supernatant (crude soluble protein fraction). Proteins of each fraction were pre- cipitated by the addition of four volumes of acetone. After overnight precipitation at 4 "C, proteins were collected by centrifugation, dispersed in 200-400 p1 of 0.1 M NazC03, 0.1 M dithiothreitol (solutionA) and solubilised in an equal volume of 4% (w/v) sodium dodecyl sulphate, 15% (v/v) glycerol and 0.1 % bromophenol blue (solution B) by heating at 70 "C for 4 min, prior to gel electrophoresis. The acetone supernatant above the membrane protein pellet was assayed for chlorophyll. By comparison with the chlorophyll content per leaf pair, the yield of material in the filtrate was calculated as deriving from a certain number of leaf pairs. From the volumes of solutions A and B used to solubilize the membrane- bound and soluble proteins, it was possible to calculate the volume of each protein sample that had to be loaded onto the 10 - 30 % (w/v) polyacrylamide gel so that each track should carry the protein equivalent to one-tenth of a leaf pair (Fig. 2 and 3 ) . After electrophoresis and staining, the gel was scanned with a model 3CS microdensitometer (Joyce-Loebl Ltd, Gateshead, Tyne and Wear, UK).
Labelling of Proteins with [35S]Methionine in vivo
The third leaf pair was detached under water, the cut end of the petiole was immersed in 100 pl of water containing 100-200 pCi [35S]methionine (1150 Ci/mmol, The Radio- chemical Centre, Amersham, Bucks, UK) and the leaf pair was allowed to take up isotope for 6 h in the light (100 pmol quanta m-2sp') or darkness. Three, four or five leaf pairs were labelled and homogenised in a cold mortar with 5 - 10 ml of SO mM Tricine/KOH pH 8.0, 5 mM 2-mercaptoethanol. After filtration and differential centrifugation as described above, the membrane-bound and soluble proteins were pre- cipitated with acetone and prepared for electrophoresis on 10 - 30 % polyacrylamide gels by solubilization with solu- tions A and B.
Staphylococcal V8 Protease Digestion
Membrane proteins, labelled in vivo with [35S]methionine in either light or darkness, were electrophoresed on a 20% (w/v) polyacrylamide gel slab, the gel was dried without staining or fixation, and the 26000-Mr polypeptides were located by autoradiography. The appropriate regions of the gel were excised and fragments were placed in the wells of a second gel slab (10 - 30 % linear gradient of polyacrylamide with a 5 % polyacrylamide stacking gel). The fragments in the wells were covered with a little buffer (62.5 mM Tris/HCl pH 6.8, 10% glycerol, 2.3% sodium dodecyl sulphate, 5 % 2-mercaptoethanol) and allowed to swell. Each well was then overlaid with a solution of Staphylococcus aureus V8 protease (Miles Research Products, Elkhardt, IN, USA). The concen- tration of the protease was 5 pg ml-' and the buffer was an equal mixture of the buffer used to swell the gel fragments and 50 pM Naz EDTA pH 6.8. The S. aureus V8 protease and the radioactive proteins were electrophoresed quickly into the stacking gel, the current was switched off for 1 h to permit digestion to proceed to the extent of about SO % hydrolysis of the 26 000-M, proteins, and then electrophoresis was continued overnight. The gel was fixed and fluorographed

[24]. The fluorogram was developed and scanned with the microdensitometer.
Preparation of Antigen
Isolated intact chloroplasts were incubated in the light with [32P]orthophosphate to label thylakoid polypeptides such as the 26000-M, light-harvesting chlorophyll a/b protein doublet [25,26]. Thylakoids were prepared from the chloro- plasts and extracted with chloroform/methanol [26]. The chloroform/methanol-soluble polypeptides were precipitated with an equal volume of ether, solubilised by heating in 500 pl of solution A and 500 pl of solution B and fractionated by sodium dodecyl sulphate/polyacrylamide gel electrophoresis using a 20% polyacrylamide gel slab. The surface of the stacking gel ( 5 % polyacrylamide) was flat so that the sample could be applied across the entire width of the slab. After electrophoresis, the wet, unfixed gel was autoradiographed to locate the 26000-M, light-harvesting protein doublet. A narrow band containing the doublet was excised from the gel and the polypeptides were eluted from macerated frag- ments by stirring overnight at room temperature with 1 ”/, sodium dodecyl sulphate, 1 % (v/v) 2-mercaptoethanol. The proteins were precipitated by the addition of four volumes of acetone and dissolved in phosphate-buffered saline (0.8 ”/, NaCl, 0.02% KC1, 0.115% Na2HP04, 0.02% KH2P04, pH 7.4, anhydrous salts) to a final light-harvesting protein concentration of about 500 pg ml-’.
Preparation of Antiseru against Purified Light-Harvesting Polypeptides
An immune response was elicited in female New Zealand White rabbits by injection of 200 pg of protein emulsified with Freund’s complete adjuvant. Booster injections of 200 pg of protein emulsified in Freund’s incomplete adjuvant were followed by bleeding and preparation of serum. The immuno- globulin fraction was precipitated from serum at 40% (NH4)2S04, dissolved to a concentration of about 20 mg ml-’ in phosphate-buffered saline, dialysed against the same buffer and stored at - 20 “C.
Immunoreaction of [3sSJMethionine- Labelled Light-Harvesting Protein
Acetone-precipitated membrane proteins were dispersed in 300 pl of phosphate-buffered saline and the solution was made 2 % (w/v) in sodium dodecyl sulphate. After heating at 70°C for 4 min, a portion of the solution (50 pl) was diluted to 500 p1 with phosphate-buffered saline and 50 pl of anti- serum was added. The immunoreaction was allowed to proceed at 37 “C for 2 h. Protein-A - Sepharose (50 mg, Pharmacia Fine Chemicals, Uppsala, Sweden) was washed once with phosphate-buffered saline and resuspended in 400 p1 of the same buffer. At the end of the immunoreaction, the 400 pl suspension of protein-A - Sepharose was added and left overnight at 4°C to adsorb the antibody-antigen complexes. The gel was poured into a small chromatography column containing a glass wool plug, was washed with 10 ml of phosphate-buffered saline containing 1 % (v/v) Nonidet P-40 and was finally washed with 1.5 ml of 1 M acetic acid to elute the antibody-antigen complex. The eluted protein was precipitated at 0 “C by the addition of trichloroacetic acid to a final concentration of 10 (w/v) and prepared for electro- phoresis by solubilization with solutions A and B.
Sodium Dodecyl Sulphate/Polyacrylamide Gel Electrophoresis
The buffers used were those of Laemmli [27] and slab gels were formed as described [28]. The slabs consisted of either 10-30% gradients of acrylamide or a uniform 20% acryl- amide gel. In the 10 %, 20 % and 30 ”/, acrylamide solutions from which the gels were made, methylene-bisacrylamide constituted 2.7%, 1.5% and 0.5%, respectively, of the total acrylamide content. Gels were stained and destained as described [25,26]. Where gels were fixed without staining, they were immersed for at least 2 h in a solution containing 7 ”/, (v/v) acetic acid and 30 (v/v) methanol.
RESULTS
Breakdown of Light-Harvesting Chlorophylls in Darkness
Seven-day-old etiolated peas were exposed to continuous white light to initiate leaf expansion and chloroplast devel- opment. The third leaf pair developed more rapidly than the other leaves and synthesized chlorophyll for about 6 days (Fig. 1 A). When seedlings were returned to darkness after just one day of illumination, almost half of the chlorophyll in the third leaves proved to be unstable, with a half-life of about 90 min (Fig. 1 B).
The loss of chlorophyll in darkness was accompanied by a rise in the chlorophyll alchlorophyll h ratio (Table 1). In plants exposed to one day of illumination the ratio increased in darkness from 2.46 to 5.76, but in plants exposed to a longer period of illumination the rise in the ratio was less pronounced. After one day of illumination 38 % of the chlorophyll a and 74 % of the chlorophyll b were unstable and the chlorophyll a/ chlorophyll b ratio of the unstable molecules was 1.27. This low value for the ratio closely resembles the values obtained for isolated light-harvesting chlorophyll a/b protein com- plexes [29,30] and indicates that the unstable chlorophylls were derived from the light-harvesting apparatus of the im- mature thylakoid membrane.
Breakdown of the Light-Harvesting Chlorophyll a/b Protein
The accumulation of the light-harvesting chlorophyll a/ b protein and of certain other chloroplast polypeptides was assayed in plants transferred from light to darkness. Mem- brane-bound proteins and soluble proteins were separated by differential centrifugation of leaf homogenates. The two fractions were analyzed by sodium dodecyl sulphate/poly- acrylamide gel electrophoresis followed by staining and scanning of the gel. Scan profiles of membrane proteins are presented in Fig. 2 and of soluble proteins in Fig. 3. In each case, tracks were loaded with the membrane or soluble pro- teins corresponding to one-tenth of a leaf pair.
After one day of illumination the 26000-MI light-harvest- ing chlorophyll a/b protein and the 59000-Mr(a) and 56000- M,(p) subunits of the coupling factor CF1 were detectable among the membrane proteins of the leaf (Fig.2C). With the exponential 10 - 30 % polyacrylamide gradient gel used here, the a and p subunits of CF1 were not sufficiently separated to appear as two distinct peaks on the scan. Among the soluble proteins were the 54000-M1 and 14000-MI subunits of ribu- lose 1,5-bisphosphate carboxylase (Fig. 3 C). After 3 days of illumination (Fig. 2A and 3A), these five chloroplast poly- peptides had become very prominent.
When plants were illuminated for one day and then returned to darkness for two days, four out of the five poly-

64
peptides continued to accumulate, although at a lower rate than in the light (Fig.2B and 3B). The exception was the light-harvesting chlorophyll a/b protein : this polypeptide not only failed to accumulate in darkness but appeared actually to have declined in concentration (compare peak heights in Fig. 2B and C).
In view of the decline in the concentration of chlorophyll a and chlorophyll h after transfer of plants from light to dark- ness (Table I), it seemed desirable co place these estimates of the concentration of the light-harvesting protein on a more quantitative basis. The scans in Fig. 2 and 3 refer to a stained polyacrylamide gel slab where each track was loaded with proteins derived from the same fraction of a leaf pair. This type of loading permits the relative levels of proteins in different samples to be compared at a glance but is not the most appropriate type of loading for quantification of protein levels. For example, although it is quite easy to determine the relative amounts of the sinall subunit of ribulose 1,s-bisphos- phate carboxylase in Fig. 3 A and B, the peak in Fig. 3 C is too small and too poorly resolved from neighbouring peaks to be quantified. For quantification, two changes in procedure are required. Firstly, the gel should be cast from a single concen- tration of polyacrylamide chosen to give optimal separation of the peak of interest from neighbouring peaks of similar molecular weights. In the case of both the small subunit of the carboxylase and the light-harvesting protein, a 20 poly- acrylamide gel was most appropriate. Secondly, it is best to load the gel slab with several different volumes of each sample
Fig. 1. Insiabiliiy of chlorophyll in tliwloping pea seedlings. After seven days of etiolated growth, plants were exposed to continuous white light for six days (A) or for one day followed by return to darkness for a further 24 h (B). At intervals the chlorophyll content of the third leaf pair was determined
(decreasing volumes of concentrated samples and increasing volumes of dilute samples) to obtain a more accurate measure- ment of relative abundances.
The area under a peak on the scan is very nearly propor- tional to the volume of sample loaded. This is illustrated in Fig. 4A by reference to the small subunit of the carboxylase. Linearity can sometimes be improved if corrections are made for variations that can occur in the width of tracks. Width can vary not only from track to track but also down the length of a single track.
Table 2 presents the results of an experiment in which the levels of the small subunit of ribulose 1,5-bisphosphate car- boxylase and of the light-harvesting chlorophyll a/b protein were determined from scans of stained gels. The assays were
Migration
Fig. 2. Instability qf the light-harvesting chlorophyll a/b protein in devel- opingpea seed/iing.r. Etiolated plants were exposed to white light for one day (C), three days (A) or one day followed by two days of darkness (B). The third leaf pairs were homogenized and cellular membranes, including the thylakoids, were sedimented at 30000 xg. Membrane proteins were analyzed by sodium dodecyl sulphatelpolyacrylamide gel electrophoresis. The gel was stained and scanned with a microdensitometer. Each track was loaded with the membrane proteins from one-tenth of a leaf pair. The position of the 26000-M, light-harvesting chlorophyll alh protein is indicated (26), as is the position of the doublet of coupling factor 56000-M, and 59000-Mr subunits (59 + 56)
Table 1 Degradation of thloiophqll a and chlorophyll b after tranyfer of immature qreenrng pea wcdlrngr from hnht to daikneys Etiolntcd pea seedlings were exposed to continuous illurninntion for 1, 2 or 4 days and then trninfei red to darkness for 2 days The chlorophyll content o f the third leaf pairs w n ~ determined before and afler the 2-day dark period The diflerence between the two figures was taken to rcpre\cnt unstable chlorophyll molecules that weie degraded during darkness
Chlorophyll type
Chlorophyll Loiiteiit per third lent pnir nfter
1 day of light
total stable unstable total stable unstable total stable unstable
~ - ~ __ __ -~ -~
2 days of light 4 days of light - - ~ ~ ~ ~~ ~~ ~ ~ ~
~
PLg ( 'U Cif ( " 3 P& (<'")
n 8 4 5 2 3 2 (38) 6 3 4 4 7 8 1 5 6 1248 8 7 7 37 1 (30)
~ ~ ~ ~ ~ - -~
a + h 1 1 8 6 1 5 7 (48) 9 5 1 6 9 5 2 5 6 (27) 1872 127 1 601 (32)
h 3 4 0 9 2 5 (74) 3 1 7 21 7 1 0 0 (25 ) (12) 6 2 4 3 9 4 23 0 (37 ) a h ratio 2 4 6 5 76 1 2 7 2 0 2 2 0 156 2 0 0 2 2 3 1 6 1

54
Migration
Fig. 3. Accumulution of .soluble proteins in developing pea sccdlings. Etio- lated plants were exposed to white light for one day (C), three days (A) or one day followed by two days of darkness (B). The third leaf pairs were homogenized and cellular membranes were removed by centrifugation at 30000 xg. The soluble proteins of the supernatant were analyzed by sodium dodecyl sulphatc/polyacrylamide gel electrophoresis. The gel was stained and scanned with a microdensitometer. Each track was loaded with the soluble proteins from one-tenth of a leaf pair. The posi- Lions of (lie 14000-Mr and 54000-12fr subunits of ribulose 1,5-biaphos- phate carboxylase are indicated (14 and 54)
Sample volume ( ~ 1 )
Fig. 4. Qucrntirurivr niic~rork~~rz,sitomc~ir~ of stained polj,uc~rylumide , ~ c ~ l s orid autor-adio~?rain.s. (A) Different volumes (19 - 78 pl) of the soluble protein sample frorn Fig. 3 A wcrc applied to it sodium dodecyl sulpliate,'poly- acrylamide gel. After electrophoresis, the gel was stained and scanned with a microdensitometer. The area under the peak corresponding to the small (14OOO-M,) subunit of ribulose 1,5-bisphosphate carboxylasc was determined for each track. (B) Different volumes (3-80 pl) of the ['sS]mett~ionine-labcllcd soluble protein sample from Fig. 8 D were applied to a sodium dodecyl sulphate,'polyacrylamide gel. After electro- phoresis. the gel was dried, and autoradiographed, and the autoradio- graph was scanned with ;L microdensitomcter. The area under the small subunit peak was determined for each track
performed on third leaf pairs that had been illuminated for one day, three days or one day followed by two days of dark- ness. The small subunit accumulated after transfcr of plants from light to darkness but at a slower rate than Lhat observed in plants kept in the light. In the dark the lcvel increased 2.4-fold whereas in the light the increase was 7.3-fold. In con-
Table 2. Effect of transfer of pea seedlings front l i ~ l i i to durkne.ss 011 thr concentration in the third leafpair of the IiRIit-harvrstiiig chloropliyll a / b protein mid tlir m a l l subunit qf ribuhr I,5-bi~pho,~plicrte cnrho.rylu.sr Etiolated pea seedlings were exposed to continuous illumination for 1 day, 3 days or 1 day followed by 2 days of darkness. The membrane-bound and soluble proteins were prepared from the third leaf pairs and analyzed by electrophoresis on a 20 :" polyacrylamide gel containing sodium dode- cylsulphate. The gel was stained and scanned, and the areas under the relevant peaks were used to calculate the absolute concentration of the light-harvesting protein and the small subunit per leaf pair. Thc figures in the table are the averages of duplicate assays and are exprcssed as a percentage of the polypeptide concentration in leaves that had received 3 days of illumination
Sainplc Time in Protcin concentration -~ per third ledf pdir
light daik light-harvesting ri bulo\e chlorophyll u/h 1,5-bi\phoiph,ite protein c'irboxyldse
mal l subunit
tldys /" ~~
1 1 - I8 14 b 3" - 100 100 C. 1 2 3 33 100 c/a 16 240
* After 3 days of' continuous illumination the concentration of the small subunit of the carboxylase was twice that of the light-harvesting protein.
trast the level of the light-harvesting protein increased only in the light (by 5.4-fold) and decreased in absolute amount in the dark (by 84%).
These results clearly confirm the impression created by the scans in Fig. 2 and 3. That is, the light-harvesting protein, unlike the other chloroplast polypeptides that have been studied, fails to accumulate in darkness and is subject to exten- sive breakdown. The figure of 84 y,; breakdown is comparable with the figure of 74 "/, breakdown in the case of chlorophyll h (Table 1).
Synthesis of thc Light- Harvesling C'l~loropl~j~ll a/b Prolciri in Dcrrkize.ss
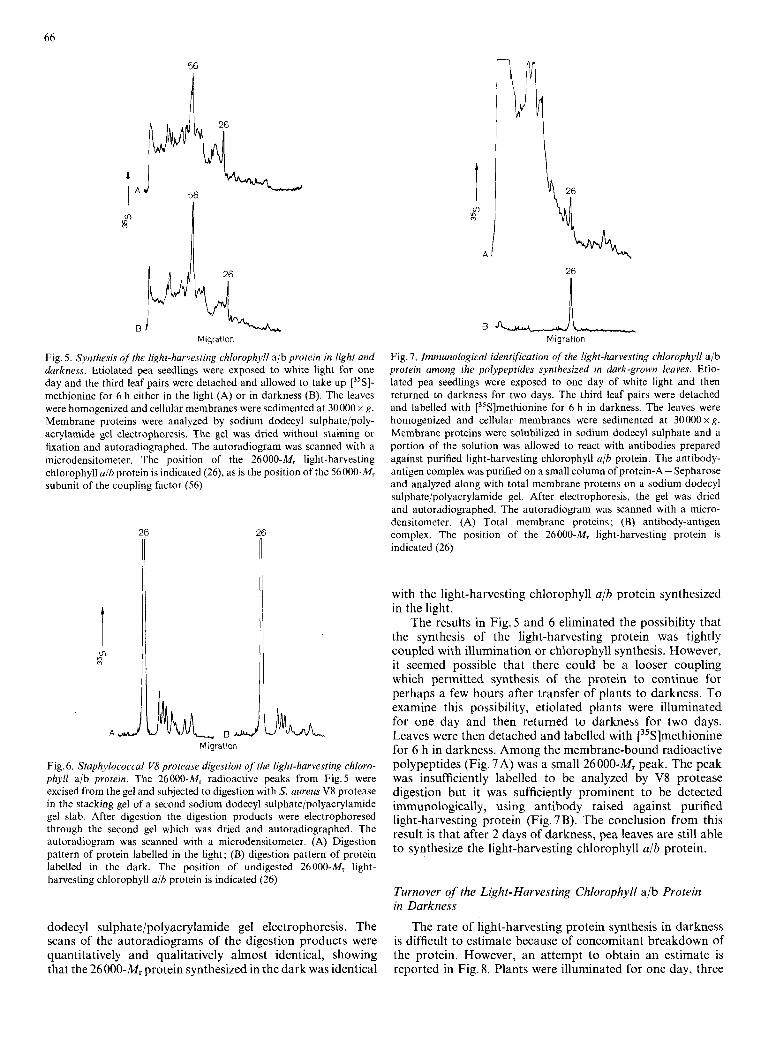
The failure of the light-harvesting protein to accumulatc in darkness could have been due either to inhibition of syn- thesis in the dark or to marked stimulation of breakdown. Inhibition of synthesis would imply the existence of a light- dependent step in the translation, transport, clcavage or mem- brane-attachment of thc protein. However, this possibility is made unlikely by the results in Fig. 5. Aftcr one day of conti- nuous illumination, pea leaves were detached and labelled with ["S]methionine for 6 11 in the light (Fig. 5 A ) or the dark (Fig. 5 €3). Mcmbrane proteins were extracted and analysed by sodium dodecyl sulphate/polyacrylamidc pel electropho- resis and autoradiography of thc dried gel. In both cases, radioisotope was incorporated into many mcmbranc poly- peptides, including the subunit of CFI and the 26000-M, light-harvesting protein.
The 26000-M, peak from each profile was exciscd I'rorn the gel and digested with Stnphy?l'lococ,cu.v uurm.v V 8 protease (Fig. 6). When about 50 2; of the radioactive protein molecules had been digested, the products were analyzed by sodium
66
56
Migration
Fig. 5. Synthesis of the light-harvesting chlorophyll ajb protein in light and darkness. Etiolated pea seedlings were exposed to white light for one day and the third leaf pairs were detached and allowed to take up [35S]- methionine for 6 h either in the light (A) or in darkness (B). The leaves were homogenized and cellular membranes were sedimented at 30000 x g. Membrane proteins were analyzed by sodium dodecyl sulphatelpoly- acrylamide gel electrophoresis. The gel was dried without staining or fixation and autoradiographed. The autoradiogram was scanned with a microdensitometer. The position of the 26000-Mr light-harvesting chlorophyll ajh protein is indicated (26), as is the position of the 56000-Mr subunit of the coupling factor (56)
Migration
Fig. 6. Staphylococcal V8 protease digestion of the light-harvesting chloro- phyll a/b protein. The 26000-Mr radioactive peaks from Fig.5 were excised from the gel and subjected to digestion with S. aureus V8 protease in the stacking gel of a second sodium dodecyl sulphate/polyacrylamide gel slab. After digestion the digestion products were electrophoresed through the second gel which was dried and autoradiographed. The autoradiogram was scanned with a microdensitometer. (A) Digestion pattern of protein labelled in the light; (B) digestion pattern of protein labelled in the dark. The position of undigested 26000-M, light- harvesting chlorophyll a/b protein is indicated (26)
dodecyl sulphate/polyacrylamide gel electrophoresis. The scans of the autoradiograms of the digestion products were quantitatively and qualitatively almost identical, showing that the 26000-M, protein synthesized in the dark was identical
26
Migration
Fig. I . Immunological identification of the light-harvesting chlorophyll ajb protein among the polypeptides synthesized in dark-grown leaves. Etio- lated pea seedlings were exposed to one day of white light and then returned to darkness for two days. The third leaf pairs were detached and labelled with [35S]methionine for 6 h in darkness. The leaves were homogenized and cellular membranes were sedimented at 30000 x g. Membrane proteins were solubilized in sodium dodecyl sulphate and a portion of the solution was allowed to react with antibodies prepared against purified light-harvesting chlorophyll a/b protein. The antibody- antigen complex was purified on a small column of protein-A - Sepharose and analyzed along with total membrane proteins on a sodium dodecyl sulphate/polyacrylamide gel. After electrophoresis, the gel was dried and autoradiographed. The autoradiogram was scanned with a micro- densitometer. (A) Total membrane proteins; (B) antibody-antigen complex. The position of the 26000-Mr light-harvesting protein is indicated (26)
with the light-harvesting chlorophyll a/b protein synthesized in the light.
The results in Fig. 5 and 6 eliminated the possibility that the synthesis of the light-harvesting protein was tightly coupled with illumination or chlorophyll synthesis. However, it seemed possible that there could be a looser coupling which permitted synthesis of the protein to continue for perhaps a few hours after transfer of plants to darkness. To examine this possibility, etiolated plants were illuminated for one day and then returned to darkness for two days. Leaves were then detached and labelled with [35S]methionine for 6 h in darkness. Among the membrane-bound radioactive polypeptides (Fig. 7A) was a small 26000-Mr peak. The peak was insufficiently labelled to be analyzed by V8 protease digestion but it was sufficiently prominent to be detected immunologically, using antibody raised against purified light-harvesting protein (Fig. 7 B). The conclusion from this result is that after 2 days of darkness, pea leaves are still able to synthesize the light-harvesting chlorophyll a/b protein.
Turnover of the Light-Harvesting Chlorophyll alb Protein in Darkness
The rate of light-harvesting protein synthesis in darkness is difficult to estimate because of concomitant breakdown of the protein. However, an attempt to obtain an estimate is reported in Fig. 8. Plants were illuminated for one day, three

67
56 56
E
F Migration
Fig. 8. Synthesis of the light-harvesting chlorophyll a/b protein and the small subunit of ribulose I,5-bisphosphate carboxylase in darkness. Etio- lated pea seedlings were exposed to white light for one day (C,F), three days (A,D) or one day followed by two days of darkness (B,E). The third leaf pairs were detached and allowed to take up [3sS]methionine for 6 h in darkness. The leaves were homogenized and cellular mem- branes were sedimented at 30000 x g . Membrane proteins (A - C) and soluble proteins (D - F) were analyzed by sodium dodecyl sulphate/ polyacrylamide gel electrophoresis. The gel was dried and autoradio- graphed. The autoradiogram was scanned with a microdensitometer. The positions of the light-harvesting protein (26000-M,) and the small subunit (14000-Mr) are indicated (26 and 14). Each membrane protein track was loaded with 80000 counts min-' and each soluble protein track was loaded with the same volume of sample that had been loaded on the corresponding membrane protein track. This ensured that the two samples originated from the same amount of tissue
/I
Migration
Fig. 9. Long-term and short-term labelling of membrane-bound proteins in pea leaves exposed to light and darkness. Etiolated pea seedlings were illuminated for 24 h and then the leaves of some plants were painted with ~-[~~S]methionine. Some of the labelled and unlabelled plants were main- tained under illumination for a further 16 h, while the remainder of the labelled and unlabelled plants were returned to darkness for this period. The labelled leaves were then detached from the plants and analyzed for radioactive membrane proteins. The unlabelled leaves were detached and were allowed to take up isotope for 4 h through their cut stems under a continuation of the light regime that they had experienced in the preceding 16 h. These leaves were then also homogenized and analyzed for radioactive membrane proteins, by dodecylsulphate/poly- acrylamide gel electrophoresis and autoradiography. Scans A and B : long term labelling. Scans C and D: short-term labelling. Scans A and C : labelling in light. Scans B and D: labelling in the dark. The locations of the 26000-Mr light-harvesting chlorophyll a/b protein and the 56000-M, p subunit of the chloroplast coupling factor are indicated (26 and 56). Each track was loaded with protein corresponding to 50000counts min-'
days, or one day followed by two days of darkness. Leaves were detached and labelled in darkness for 6 h with [35S]- methionine. Membrane-bound and soluble proteins were isolated and analyzed by sodium dodecyl sulphate/polyacryl- amide gel electrophoresis. Radioactive proteins were detected by autoradiography. Scans of the autoradiogram are given in Fig. 8. (As Fig. 4B shows, there is an approximately linear relationship between the area under the peak of such a scan and the amount of radioactivity present in the protein band.)
In leaves labelled after one or three days of illumination, both the light-harvesting protein and the small subunit were prominent relative to the general background of radioactive polypeptides (Fig. 8 A, C, D, F). However, in leaves labelled two days after transfer to darkness (Fig. 8 B, E), the two poly- peptides were comparatively minor radioactive species. As far as the small subunit is concerned, these results are con- sistent with the rates of accumulation of the protein. Between the first and third day of illumination, the small subunit accumulated 4.5 times more rapidly in the light than in the dark (Table 2) and was labelled 5.0 times more rapidly in leaves that had been illuminated for three days (Fig.8D) than in leaves that had been returned to darkness after one day of illumination (Fig. 8 E).
In the case of the light-harvesting protein, however, the situation is rendered more complex by the occurrence of breakdown. A direct comparison of the level of the protein with the level of labelling is impossible. Nevertheless, it is
clear from the results in Fig. 7 and 8 B that the light-harvesting chlorophyll a/b protein is synthesized in leaves returned to darkness for two days. Furthermore, the extent of labelling of the protein in these leaves is at least 30% of the extent of labelling of the small subunit of ribulose bisphosphate car- boxylase (Fig. 8 E). Thus, over a two-day period, leaves trans- ferred from light to darkness gradually lose their ability to synthesize both the light-harvesting protein and the small subunit of the carboxylase but the small subunit can accumu- late under these conditions because, unlike the light-harvesting protein, it does not appear to be subject to rapid turnover in darkness.
If the light-harvesting chlorophyll a/b protein is turned over in darkness, the level of radioactivity found in this protein should reach a higher specific activity in the light than in darkness. However, as shown in Fig. 5 , the labelling of detached leaves for 6 h produces essentially the same degree of labelling of the protein, irrespective of illumination. The reason for this apparent inconsistency in the results lies in the fact the a 6-h labelling period is actually rather short, since it takes at least 3 h for a substantial amount of radioactivity to enter a pea leaf that is labelled through the stem or petiole. This is especially true of labelling in darkness.
Fig. 9 shows the results of a labelling experiment of much longer duration (16 h). Since detached pea leaves synthesized chlorophyll at the same rate as attached leaves for only about 6 h (J. Bennett, unpublished results), this experiment was

performed with attached leaves. Seven-day-old etiolated peas were illuminated for 24 h and then the leaves of some plants were painted with ~-[~'S]methionine (1 mCi m1-l in 0.01 Triton X-100 solution). Half the plants were maintained in the light while the remainder were returned to darkness. After 16 h, the labelled leaves were harvested and membrane pro- teins were analyzed by dodecylsulphate/polyacrylamide gel electrophoresis followed by autoradiography (Fig. 9 A. B). At the same time, leaves of unlabelled plants were detached and were allowed to take up the radioisotope through their cut stems; leaves from illuminated plants were labelled in the light, while leaves from plants that had been returned to darkness for 16 h were labelled in the dark. After 4 h, these leaves were also harvested and their membrane proteins were analyzed (Fig.9C,D). The results show that over 16 h the light-harvesting chlorophyll a/b protein can be labelled to a high specific activity in the light but not in the dark, in spite of the ability of leaves that have been in darkness for 16 h to label the same protein in the short term (4 h). This inability to accumulate radioactive light-harvesting chlorophyll a/b protein confirms the occurrence of turnover within the photo- synthetic membrane.
DISCUSSION After initiation by 24 h of illumination, the synthesis of
the light-harvesting chlorophyll a/h protein continues in darkness in the absence of chlorophyll synthesis for at least 2 days. However, the protein fails to accumulate because of rapid turnover within the photosynthetic membrane. Two forms of the protein appear to be susceptible to proteolysis in darkness. Firstly, as transfer of plants from light to darkness is followed by the destruction of 38"/, of the chlorophyll a, 74% of the chlorophyll b and 84% of the light-harvesting protein present in the leaves at the end of the light period, a large percentage of the apo-protein already in the form of chlorophyll-protein complexes would appear to be unstable in darkness, at least in immature leaves. Secondly, as the light-harvesting apo-protein and the small subunit of ribulose 1,5-bisphosphate carboxylase are synthesized in darkness at readily detectable rates while only the latter accumulates, newly synthesized apo-protein molecules must also be subject to breakdown within the photosynthetic membrane. This is confirmed by the long-term labelling experiment in which the apo-protein could not be labelled to a high specific activity in darkness in spite of continuous synthesis.
The destruction of the unstable light-harvesting chloro- phyll a/b protein complexes is accompanied by a rise in the chlorophyll alchlorophyll b ratio. A rise in this ratio after transfer of plants from light to darkness has been reported previously. Kupke and Dorrier [31] suggested that chloro- phyll b reverts to chlorophyll a in darkness, whereas Thorne and Boardman [32] suggested that chlorophyll b may be de- graded more rapidly than chlorophyll a. The results in this paper support the second explanation and indicate that the light-harvesting chlorophyll a/h complex is more susceptible to breakdown than the other complexes of the thylakoid that contain only chlorophyll a. Further work will be required before we can provide a complete list of the thylakoid compo- nents that are susceptible to breakdown in darkness in imma- ture leaves. One problem in gathering this information is that the concentration of thylakoid polypeptides is very low after just 24 h of illumination but it is not possible to extend the pre-illumination period to increase the polypeptide con-
centration because longer periods of illumination serve only to stabilize the light-harvesting apparatus against break- down. It should be noted that Thorne and Boardman [32] found that the most marked rise in the chlorophyll alchloro- phyll h ratio after transfer to darkness took place in leaves that had received the shortest period of pre-illumination. This reflects the fact that the least mature thylakoids contain the largest proportion of potentially unstable light-harvesting chlorophyll ajb complexes.
Degradation of the light-harvesting protein requires a protease that is able to attack a hydrophobic polypeptide embedded in the lipid bilayer of the thylakoid. That such an enzyme may exist is suggested by recent work on the rapidly turned-over 32000-Mr thylakoid protein known as peak D [33]. This particular protein is a major product of chloroplast protein synthesis [33 - 351. It is also hydrophobic. It is there- fore possible that the protease which degrades peak D protein is also able to degrade the light-harvesting protein. The rela- tionship between this protease, whose existence can only be inferred at present, and the two proteases found recently in oat etioplasts and immature chloroplasts [36] remains to be investigated. The preferred substrate of the oat proteases appears to be the proteins of the prolammellar bodies [36].
It has frequently been pointed out that etiolation is not a natural stage in leaf development [37]. There may therefore be little justification in seeking a physiological role for the breakdown of the light-harvesting chlorophyll a/b protein observed in this study. It may be simply a consequence of subjecting leaves to an unnaturally long period of darkness, then suddenly to illumination and finally, and equally sud- denly, to a further period of darkness. However, there remains the possibility that breakdown of the apo-protein in darkness is the mechanism by which the leaf co-ordinates the accumula- tion of the pigment and protein components of the light- harvesting apparatus. There are two general mechanisms by which such a co-ordination could be achieved: firstly, both chlorophyll and apo-protein could be synthesized only in the light ; secondly, chlorophyll could be synthesized in the light, apo-protein could be synthesized in light and darkness, and the excess apo-protein could be degraded in darkness, being recognised as surplus to requirements by its inability to become incorporated into functional aggregates.
There are at least two reasons for doubting that the first mechanism operates in vivo. Firstly, there is the evidence presented in this paper that synthesis of the apo-protein occurs in darkness. Over a period of 2 days in darkness there is certainly a marked decline in the rate of labelling of the light-harvesting apo-protein but as this decline is also seen in the rate of labelling of the small subunit of ribulose 1,Sbis- phosphate carboxylase, it cannot be taken to reflect the exis- tence of a specific photocontrol over apo-protein synthesis. Secondly, such a photocontrol would have to operate in the cytoplasm where the light-harvesting protein precursor is synthesized. There would have to be a photoreceptor in the cytoplasm or some sort of signal would need to leave the plastid to control translation of only the light-harvesting protein mRNAs. The advantage of the second mechanism described above is that it would operate entirely within the thylakoid and provide a direct link between the rate of chlorophyll synthesis and the rate of light-harvesting protein accumulation.
There is, however, an apparent weakness in the operation of the second mechanism. According to the results presented here, a large percentage of the light-harvesting chlorophyll u/b complexes in immature thylakoids are subject to degradation.

I have interpreted the loss of these complexes as indicating that there is a pool of complexes that have recently been syn- thesized but not yet incorporated into a photosynthetic unit. This apparently wasteful degradation will need to be studied further before the second mechanism can be accepted as a sufficient explanation for the co-ordination of the synthesis of chlorophyll with the accumulation of the light-harvesting apo-protein.
One aspect that needs particularly close examination is the finding that intermittently illuminated leaves are photo- chemically active yet are devoid of the light-harvesting chloro- phyll nib complex [7,20,21]. The fact that intermittent illumi- nation (such as 2 min of white light every 2 h for 2 days [20]) fails to elicit synthesis of chlorophyll b or the apo-protein and permits synthesis of only a low level of chlorophyll n, has been taken to indicate that there is an intimate connection be- tween light, the synthesis of chlorophyll b and the accumula- tion of the apo-protein. Recent work in this laboratory has been directed towards explaining the absence of the light-har- vesting apparatus from intermittently illuminated pea leaves. The results of this study are reported in the following paper. The conclusion is that the absence of the light-harvesting complex is unlikely to be due to control of transcription or translation. The most likely explanation is that the apo-pro- tein is synthesized continuously under intermittent illumina- tion but is subject to complete degradation in the immature bithylakoids that form under intermittent illumination. I understand that turnover also occurs in the chlorophyll-6-less mutant of barley (G. Bellemare, S. Bartlett, and N.-H. Chua, personal communication). This mutant lacks the major light- harvesting chlorophyll a/b protein [38].
To reach the thylakoid membrane, the light-harvesting chlorophyll nib protein must be translated in the cytoplasm as a larger precursor which is transported into the chloroplast and cleaved to the mature molecular weight [lo- 321. It has recently been shown that transport and/or cleavage of the precursor is markedly stimulated in .vitro by light and that light acts by generating ATP within the chloroplast for an energy-requiring aspect of this process [ 121. Incorporation of mature light-harvesting protein into the membrane fraction of pea leaves in darkness shows that photophosphorylation is not the only source of ATP in vivo for transport and cleavage of the light-harvesting protein precursor.
Finally, there is the problem of explaining the marked decline in the synthesis of both the light-harvesting protein and the small subunit of ribulose 1,5-bisphosphate carboxyl- ase when pea leaves are transferred from light to darkness for two days. Almost certainly the explanation lies in the exten- sive loss of these mRNAs by breakdown [13-151. Thus, after transfer of pea plants from light to darkness, the rate of synthesis of the light-harvesting protein declines gradually as mRNA is degraded or rendered inactive, while much pre- existing and all newly synthesized light-harvesting protein is turned over within the photosynthetic membrane in the absence of chlorophyll synthesis. The result is the loss of light-harvesting protein from the leaves. In contrast, the small subunit of ribulose 1,5-bisphosphate carboxylase is allowed to accumulate because its synthesis in the dark is not counteracted by breakdown.
When barley leaves are exposed to a single pulse of red light and then returned to darkness, the level of the light- harvesting chlorophyll nib protein mRNA rises to a maxi- mum over the next 4 h and then declines [13]. This delay before the onset of net breakdown of the mRNA provides a
the light-harvesting chlorophyll a/h protein a t much the same rate in light and in darkness for up to 16 h; that is, synthesis of the message continues for several hours after transfer of the plants from light to dark. A comparison of the results in Fig. 8 and Fig. 9 suggest that the period between 16 h and 48 h after transfer from light to dark is one of considerable mRNA breakdown.
From the results in this paper and from the work of other laboratories [13,15,17], it is now clear that the level of light- harvesting chlorophyll a/b protein in leaves of higher plants is determined by four factors: (a) phytochrome-dependent synthesis of mRNA; (b) mRNA breakdown; (c) photocon- trol of chlorophyllide synthesis; and (d) breakdown of thc protein within the photosynthetic membrane.
Note Added duving Prepvation ,for the Press
Since the submission of the revised manuscript, a paper [39] has come to my attention which presents evidence for the requirement of chlorophyll LI for the stabilization of the light-harvesting apo-protein and is in close agreement with the conclusions described here.
I wish to thank Richard Williams for excellent technical assistance and the Science Research Council for financial support.
REFERENCES
1 . Thornber, J. P., Markwell, J. P. & Reinman, S. (1979) Pkotochcm.
2. Bennett, J. (3979) T r e d s Biochem. Sci. 4 , 268-271. 3. Apel, K. (1977) Biochim. Biophys. Acta, 462, 390-402. 4. Hoober, J. K., Millington, R. H . & D’Angelo (1980) Arch. Biochem.
5 . Chua, N.-H. & Blomberg, F. (1979) J . Biol. Chem. 254, 215-223. 6 . Knox, R. S. (1975) in Bioenerptics of Photosyntltesis (Govindjee, ed.)
I. Butler, W. L. (1978) Annu. Rev. Plant Physiol. 29, 345-378. 8. Bennett, J., Steinback, K. E. & Arntzcn, C. J . (1980) Proc. Nut1
9. Kung, S. D., Thornber, J. P. & Wildman, S . G . (1972) FEBS Lelt.
Photohiol. 29, 1205 - 1216.
Biophys. 202,221 -234.
pp. 183-221, Academic Press, New York.
Acad. Sci. U S A , 77, 5253 - 5257.
24, 185-188. 10. Apel, K. & Kloppstech, K. (1978) Eur. J . Biochrm. 85, 581 -588 . 11. Schmidt, G. W., Cashmore, A. R., Broadhurst, M. K., Bartlett, S.
& Chua, N.-H. (1980) Cold Spring Harbor Symp. Quanl. Biol., in the press.
12. Grossman, A., Bartlett, S. & Chua, N.-H. (1980) Nature (Lond.) 285,
13. Apel, K. (1979) Eur. J . Biochem. Y7, 183-188. 14. Tobin, E. M. (1978) Proc. Nut1 Acud. Sci. U S A , 75, 4749-4753. 15. Tobin, E. M. & Suttie, J. L. (1980) Plant Phy.sio1. 65, 641 -647. 16. Jones, 0. T. (1979) in The Porphyrins (Dolphin, D., ed.) vol. VI,
pp. 179-232, Academic Press, New York. 17. Griffiths, W. T. & Mapleston, R. E. (1978) in Intrrnationczl Sym-
posium on Chlorop1a.st Development (Akoyunoglou, G., ed.) pp. 99 104, Elsevier/North-Holland Biomedical Press, Amsterdam.
18. Machold, O., Meister, A., Sagromsky, H., Hoeyer-Hansen, G. & Wettstein, D. v. (1977) Photosynthetica, I I , 200 - 206.
19. Henriques, F. & Park, R. B. (1975) Plant Physiol. 5.5, 763-767. 20. Armond, P. A,, Arntzen, C. J., Briantais, J. M. & Vernotte, C. (1976)
21. Akoyunoglou, G. & Argyroudi-Akoyunoglou, J . H. (1969) Physiol.
22. Hcnriques, F. & Park, R. B. (1976) Proc. Nut1 .4cud. Sci. U S A , 73,
23. Arnon, D. I. (1949) Plant Physiol. 24, 1-15. 24. Bonner, W. M. & Laskey, R. A. (1974) Eur. J . Biochem. 46, 83-88. 25. Bennett, J . (1977) Nature (Lond.) 26Y, 344-346.
625 - 628.
Arch. Biochem. Biophys. 175, 54-63.
Phnl . 22, 288-295.
4560 - 4564.
plausible explanation for the ability of pea leaves to synthesize 26. Bennett, J. (1979) Eur J Blochem. 99, 133- 137

70
27. Laemmli, U. K. (1970) Nut. New Biol. 227, 680-685. 28. O’Farrell, P. H. (1975) J . Biol. Chem. 250, 4007-4021. 29. Burke, J. J., Ditto, C. L. & Arntzen, C. J. (1978) Arch. Biochem.
30. Anderson, J. M., Waldron, J. C. & Thorne, S. W. (1978) FEBS Lett.
31. Kupke, D. W. & Dorrier, T. E. (1962) Plant Ph,ysiol. 42, lxiii. 32. Thorne, S. W. & Boardman, N. K. (1971) Plant Physiol. 47, 252-
33. Eaglesham, A. R. J. & Ellis, R. J. (1974) Biochim. Biophys. Acta,
Biophys. 187,252 - 263.
92, 227-233.
261.
335, 396 - 407.
J. Bennett, Department of Biological Sciences, University of Warwick, Coventry, Warwickshire, Great Britain, CV4 7AL
34. Edelman, M. & Reisfeld, A. (1978) in International Symposium on Chloroplast Development (Akoyunoglou, G., ed.) pp. 641 - 652, Elsevier/North-Holland Biomedical Press, Amsterdam.
35. Weinbaum, S. A., Gressel, J., Reisfeld, A. & Edelman, M. (1979) Plant Physiol. 64, 828 - 832.
36. Hampp, R. & Filippis, L. F. d. (1980) Plant Physiol. 65, 663-668. 37. Boffey, S. A, , Sellden, G. & Leech, R. M. (1980) Plant Physiol. 65,
38. Burke, J. J . , Steinback, K. E. & Arntzen, C. J. (1979) Piant Physiol.
39. Apel, K. & Kloppstech, K. (1980) Planta (Berl.) 150, 426-430.
680 - 684.
63, 237-243.