Embed Size (px)
Citation preview

ATTENTION-DEFICIT DISORDER (A ROSTAIN, SECTION EDITOR)
Biomarkers in the Diagnosis of ADHD – Promising Directions
Stephen V. Faraone & Cristian Bonvicini &Catia Scassellati
# Springer Science+Business Media New York 2014
Abstract The etiology and pathogenesis of attention-deficit/hyperactivity disorder (ADHD) are unclear and a more validdiagnosis would certainly be welcomed. Starting from theliterature, we built an hypothetical pyramid representing aputative set of biomarkers where, at the top, variants inDAT1 and DRD4 genes are the best candidates for theirassociations to neuropsychological tasks, activation in specificbrain areas, methylphenidate response and gene expressionlevels. Interesting data come from the noradrenergic system(norepinephrine transporter, norepinephrine, 3-methoxy-4-hydroxyphenylglycol, monoamine oxidase, neuropeptide Y)for their altered peripheral levels, their association with neu-ropsychological tasks, symptomatology, drugs effect andbrain function. Other minor putative genetic biomarkers couldbe dopamine beta hydroxylase and catechol-O-methyltrans-ferase. In the bottom, we placed endophenotype biomarkers.A more deep integration of “omics” sciences along with moreaccurate clinical profiles and new high-throughput computa-tional methods will allow us to identify a better list of bio-markers useful for diagnosis and therapies.
Keywords Biomarkers . Attention-deficit/hyperactivitydisorder . Dopaminergic pathway . Noradrenergic pathway .
Metabolism enzymes . CNS developmental network .
Environmental risk factors . Endophenotypes . SLC6A3 .
DRD4 . DBH . COMT . Reaction time variability .
Vigilance/sustained attention . Executive functions .
Workingmemory . EEG Theta/Beta ratio
Introduction
The aetiology and pathogenesis of attention-deficit/hyperac-tivity disorder (ADHD) is not yet fully understood [1, 2].ADHD is also a highly heritable disease, with estimatedheritability rates of up to 80% [3]. A recent review on geneticsof ADHD summarized that all variants associated with thepathology explain only a small fraction of heritability: pheno-type complexity and variants of small effect contribute to themissing heritability issue [4••]. To gain more insight into themechanisms leading from a genetic/biological basis of thedisease to the full clinical phenotype, endophenotypes are apromising strategy [5].
The DSM and ICD diagnoses of ADHD are based on aconsensus about clusters of clinical symptoms. However, thisdiagnostic procedure has been criticized for not allowingsufficiently reliable and valid diagnoses [6]. NIMH recentlyhas approved the Research Domain Criteria (RDoC) projectwhere a set of assumptions permit to found a new classifica-tion system, by integrating genetics, imaging and cognitiveinformation [6]. The RDoC approach suggests that a biomarkerapproach to diagnosis may be a more valid way to classifycomplexmental disorders such as ADHD.Biomarkers offer theopportunity to standardize and improve diagnostic assessmentwhile providing insights into etiological mechanisms.
What is a Biomarker?
The United States food and drug administration (FDA) de-fines a biomarker as an objective measure of normal process-es, pathological processes or pharmacological response [7].For psychiatry, biomarkers could be used to screen for, diag-noses, or predict the development of, not only psychiatricdisorders but also of personality or behavioral traits and
This article is part of the Topical Collection on Attention-Deficit Disorder
S. V. FaraoneDepartment of Psychiatry, SUNY Upstate Medical University,Syracuse, NY, USA
S. V. FaraoneDepartment of Neuroscience and Physiology, SUNY UpstateMedical University, Syracuse, NY, USA
C. Bonvicini : C. Scassellati (*)Genetic Unit - IRCCS “Centro S. Giovanni di Dio” Fatebenefratelli,Via Pilastroni 4, 25123 Brescia, Italye-mail: [email protected]
Curr Psychiatry Rep (2014) 16:497DOI 10.1007/s11920-014-0497-1

RDoC domains. Biomarkers could potentially be used toinform treatment decisions.
Because no single biomarker candidate may be sufficientfor accurate and reliable diagnosis, the current trend in psy-chiatry has shifted towards identifying sets of biomarkers [8,9••, 10••, 11]. For example, Pies et al. defined a set of bio-markers for schizophrenia comprising:-neuregulin-1 muta-tions; -abnormal smooth-pursuit eye movements; -reducedanterior cingulate volumes, enlarged lateral and third ventric-ular volumes and white matters abnormalities [12].
Despite their promise, the search for biomarkers of psychiatricdisorders has largely proven elusive. In a comprehensive review[10••], the authors concluded that according to their stringentcriteria, no biomarker is available for diagnosing ADHD.
The main goal of this review is to present evidence for aputative signature set of biomarkers for ADHD using geno-mic, neurophysiological/neuropsychological, neuroimaging,pharmacogenomic and proteomic knowledge to diagnoseADHD. In Table 1 we report data from genomics and meta-bolomics classified according to neurophysiological/neuro-psychological, neuroimaging, pharmacogenomic, biochemi-cal and gene expression findings. Table 2 presents data fromendophenotypes studies to define a further potential signatureset of biomarkers useful for ADHD diagnosis.
Potential Dopaminergic Biomarkers for ADHD
Genomics
Deficiencies of dopaminergic and noradrenergic neurotrans-mitter systems have been associated with ADHD and the mainsymptoms of ADHD, but the dopamine system has been themain focus of genomic research [69].
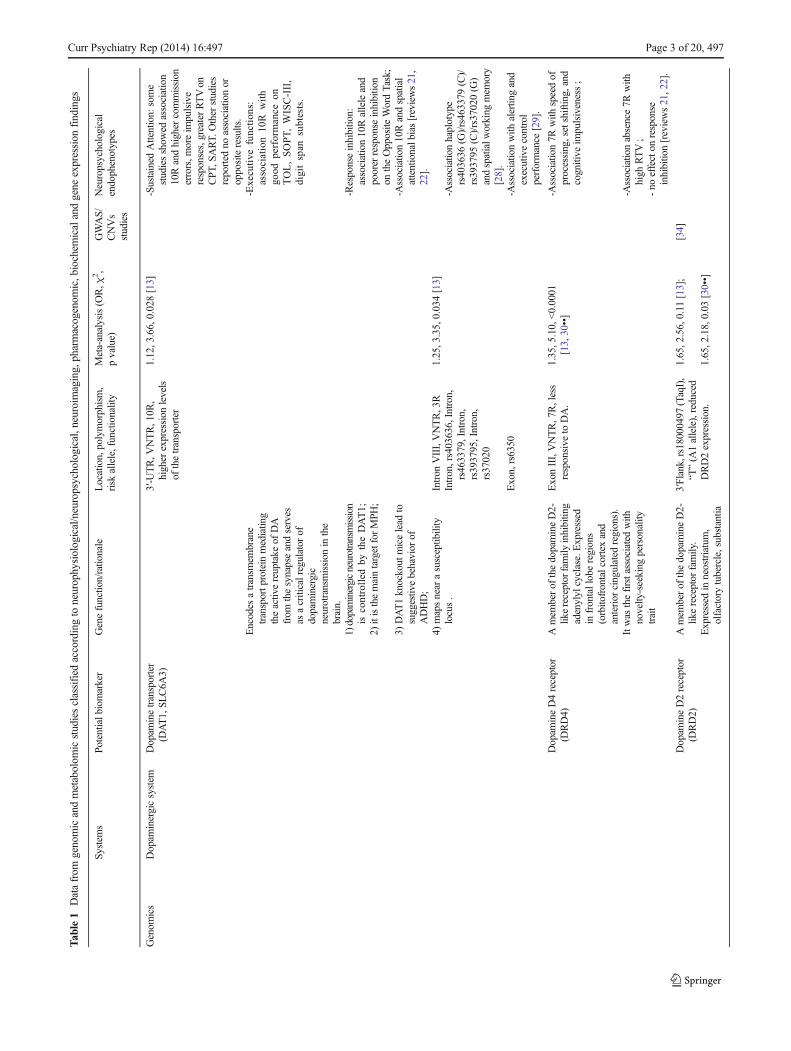
Dopamine Transporter Gene (DAT1, SLC6A3)
There are at least four main reasons for a central role of DAT1 inADHD genetics: 1) dopaminergic neurotransmission is con-trolled by the DAT1 protein; 2) the DAT1 protein is the maintarget for two ADHDmedications: methylphenidate (MPH) andamphetamine (AMP); 3) knockout mice for DAT1 show hyper-activity and deficits in inhibitory behavior [69]; 4) DAT1 hasbeenmapped near to a susceptibility locus for ADHD, 5p13 [70].The most studied DAT1 variant is a variable number of tandemrepeats (VNTR) of 40 base pairs located at the 3′-untranslatedregion (3′-UTR) of the gene. The ten repeat (10R) and ninerepeat (9R) alleles are the most common. It has been demon-strated that the DAT density measured in vivo using neuroimag-ing was significantly higher in ADHD children with homozy-gosity for the 10R in basal ganglia, a brain area participating ininhibitory behaviors. This suggests a functional effect of the 10Rallele [71]. A recent meta-analysis demonstrated an association
between the 10R allele and ADHD [13], although a reverseassociation has been seen for ADHD adults [72].
Kebir and Joober and Barnes et al. summarized DAT1 studiesof neuropsychological tasks [21, 22]. As regards the associationbetween attention tasks and the 10R allele, some studies showedhigher commission errors, more impulsive responses and greaterreaction time variability (RTV) using the continuous performancetest (CPT) and sustained attention to response test (SART). Otherstudies reported no association or opposite results [21]. Moreoverboys scoring high on a teacher-rated report of ADHD symptomsand who were homozygous for the 10R allele displayed poorerresponse inhibition on the opposite word task. Concerning execu-tive functions, the 10/10 genotype predicted better performance onthe Tower of London (TOL), the self-order pointing task, and theWechsler Intelligence Scale for Children-III (WISC-III) arithmeticand digit span subtests. Associations between a neurocognitivephenotype-spatial attentional bias and 10R allele was reported inchildren with ADHD [73, 74]. The attentional phenotype (left-sided inattention) was related to higher symptom severity [74].
Several studies have been conducted on neuroimaging and 3′UTR VNTR variant in ADHD. Genotype 10/10 individualswere found to generate more activations corresponding to inhi-bition tasks, showing also its interaction with diagnostic status(ADHD vs. controls) [15]. Moreover on non-ADHD siblings ofADHD probands and controls, an influence was observed be-tween the variant and the activation in striatum and cerebellumvermis, with its interaction with familial risk for ADHD [18].Similarly in another study, 10/10 individuals were more activat-ed in left striatum, right dorsal premotor cortex, and bilateraltemporoparietal junction [20]. In contrast, Congdon et al. ob-served greater neural activation associated with carriers of the 9R[23]. Finally caudate volumes were overall smaller in 10/10 than9/10 children, particularly in the left than right hemisphere [14].
Positron emission tomography (PET) and single photonemission tomography (SPECT) have been used to determineif ADHD patients show abnormal activity of the DAT instriatum. In a meta-analysis of these studies, Fusar-Poli et al.concluded that DAT activity was 14 % higher in ADHDpatients compared with controls [24]. A meta-analysis of neu-roimaging studies of humans found that the 9R allele wasassociated with increased DAT activity in striatum [75]. Thisstudy was conducted considering different neuropsychiatricillnesses and this strengths the issue that VNTR polymorphismregulates dopamine activity in the striatal brain regions not onlyin presence of ADHD but also of other disorders
A recent review on neuroimaging and pharmacogeneticsreported some studies on DAT1 [25••]. In particular an asso-ciation with 10R was found with higher regional cerebralblood flow in medial frontal and left basal ganglia regions,and with poorer treatment response, in children with ADHDreceiving MPH for 4 days before SPECT imaging session.Moreover this allele was associated with greater DAT avail-ability in basal ganglia following treatment and poorer
497, Page 2 of 20 Curr Psychiatry Rep (2014) 16:497
Tab
le1
Datafrom
genomicandmetabolom
icstudiesclassified
accordingto
neurophysiological/neuropsychological,neuroimaging,pharmacogenom
ic,biochem
icalandgene
expression
findings
System
sPo
tentialb
iomarker
Genefunctio
n/rationale
Location,polymorphism,
risk
allele,functionality
Meta-analysis(O
R,χ
2,
pvalue)
GWAS/
CNVs
studies
Neuropsychological
endophenotypes
Genom
ics
Dopam
inergicsystem
Dopam
inetransporter
(DAT1,SL
C6A
3)3′-U
TR,V
NTR,10R
,higher
expression
levels
ofthetransporter
1.12,3.66,0.028[13]
-Sustained
Attention:
some
studiesshow
edassociation
10Randhigher
commission
errors,m
oreim
pulsive
responses,greaterRTVon
CPT
,SART.
Other
studies
reported
noassociationor
oppositeresults.
Encodes
atransm
embrane
transportp
rotein
mediating
theactiv
ereuptake
ofDA
from
thesynapseandserves
asacriticalregulator
ofdopaminergic
neurotransmission
inthe
brain.
-Executive
function
s:association10
Rwith
good
performance
onTOL,SOPT,WISC-III,
digitspan
subtests.
1)dopaminergicneurotransmission
iscontrolled
bytheDAT1;
2)itisthemaintargetforMPH
;
-Responseinhibitio
n:association10Ralleleand
poorer
response
inhibitio
non
theOpposite
WordTask;
3)DAT1knockout
micelead
tosuggestiv
ebehavior
ofADHD;
-Association10Randspatial
attentionalbias[reviews21,
22].
4)mapsnearasusceptib
ility
locus.
Intron
VIII,VNTR,3R
1.25,3.35,0.034[13]
Intron,rs403636,Intron,
rs463379,Intron,
rs393795,Intron,
rs37020
-Associationhaplotype
rs403636
(G)/rs463379
(C)/
rs393795
(C)/rs37020(G
)andspatialw
orking
mem
ory
[28].
Exon,rs6350
-Associationwith
alertin
gand
executivecontrol
performance
[29].
Dopam
ineD4receptor
(DRD4)
Amem
berof
thedopamineD2-
likereceptor
family
inhibitin
gadenylyl
cyclase.Expressed
infrontalloberegions
(orbitofrontalcortexand
anterior
cingulated
regions).
ExonIII,VNTR,7R,less
responsive
toDA.
1.35,5.10,<0.0001
[13,30••]
-Association7R
with
speedof
processing,setshiftin
g,and
cognitive
impulsiveness;
Itwas
thefirstassociatedwith
novelty-seeking
personality
trait
-Associationabsence7R
with
high
RTV;
-no
effecton
response
inhibition[reviews21,22].
Dopam
ineD2receptor
(DRD2)
Amem
berof
thedopamineD2-
likereceptor
family.
3′Flank,rs18000497
(TaqI),
“T”(A
1allele),reduced
DRD2expression.
1.65,2.56,0.11
[13];
[34]
Expressed
inneostriatum,
olfactorytubercle,substantia
1.65,2.18,0.03
[30••]
Curr Psychiatry Rep (2014) 16:497 Page 3 of 20, 497

Tab
le1
(contin
ued) Sy
stem
sPo
tentialb
iomarker
Genefunction/rationale
Location,polymorphism,
risk
allele,functionality
Meta-analysis(O
R,χ
2 ,pvalue)
GWAS/
CNVs
studies
Neuropsychological
endophenotypes
nigra,ventraltegmentalarea,
andnucleusaccumbens.
Itplaysarolein
regulatin
gthemesolim
bicreward
pathways.
Intron,rs2075654,Intron,
rs1079596
-Associatio
nwith
greater
commission
errorrates
[review21].
Dopam
ineD5receptor
(DRD5)
Amem
berof
thedopamine
D1-lik
ereceptorsfamily.
5′Flank,Dinucleotide
repeat,136
bp0.58,−
2.13,0.0329[30••]
Expressed
inam
ygdala,frontal
cortex,hippocampus,
striatum
,basalforebrain,
hypothalam
us,cerebellum,
andthalam
us.Itisim
plicated
inhippocam
palm
emory
form
ation.
5′Flank,Dinucleotide
repeat,148
bp1.26,2.91,0.0036
[30••]
-Associationwith
commission
errors,omission
errors,R
Ts,
RTV[36];
-Noassociationwith
cognitive
tests(Stop-signaltask,
Stroop
task,T
MT,
FDI,IQ
)[38].
Metabolom
ics
Phenylethylamine(PEA)
ADAreceptor
agonist,
synthesizedby
the
decarboxylationof
phenylalanine
Dopam
ine(D
A)
InADHDtheturnover
isreducedwith
excessivere-
uptake
andintra-synaptic
monoamineconcentrations
aredecreased.
Hom
ovanillicacid(H
VA)
MainDAmetabolite.
Genom
ics
Noradrenergicsystem
Norepinephrine
transporterprotein1
NET(SLC6A
2)
Codes
foraproteinresponsible
forthereuptake
ofNEfrom
thesynapticcleftb
ackinto
thepresynaptic
neuron.
Exon9,rs5569,“A”
1.06,1.17,0.279[13]
[34]
-Noassociationwith
CPT
performance
[review21].
Expressed
inthefrontallobes.
5′Flank,rs28386840
(A-3081T
),Tallele,
reducedlevelsof
NET
with
inthebrain.
-Associatio
nTallelewith
greaterdecrease
inmean
commission
errorsscores
[40,41].
Itisactiv
elyinvolved
inboth
noradrenergicand
dopaminergicreuptake
and
regulatio
nin
thisregion.
Intron,rs998424,Intron,
rs3785157
-Associatio
nwith
high
performance
onthe
similaritiessubtesto
fthe
WISCIII[42].
Intron,rs3785155
-Associatio
nwith
RTV[43].
Metabolom
ics
Norepinephrine(N
E)
Itisreleased
bynoradrenergic
neuronslocatedin
the
specificcentraln
ervous
system
apartfrom
dopaminergicneurons.
Normetanephrine(N
M)
Mainmetabolite
ofNE.
3-methoxy-4-
hydroxyphenylg
lycol
(MHPG
)
Mainmetabolite
ofNE.
497, Page 4 of 20 Curr Psychiatry Rep (2014) 16:497

Tab
le1
(contin
ued) Sy
stem
sPo
tentialb
iomarker
Genefunction/rationale
Location,polymorphism,
risk
allele,functionality
Meta-analysis(O
R,χ
2 ,pvalue)
GWAS/
CNVs
studies
Neuropsychological
endophenotypes
NeuropeptideY(N
PY)
Colocalizes
with
catecholam
ine
system
s.Itparticipatesinthe
regulationoffeeding,circadian
rhythm
s,reproductionand
thermoregulation.
[44]
Genom
ics
Adrenergicsystem
Alpha-2A-adrenergic
receptor
(ADRA2A
)5′Flank,rs1800544(M
spI),
“G”,functio
nal
0.99,0.01,0.542[13];
1.077,0.20,0.65[45]
-AssociationCallelewith
TOL,
TMT,RTVon
differenttasks
[review21].
Encodes
alpha-2A
-adrenergic
receptorsregulatin
gneurotransmitter
releasefrom
sympatheticnerves
andfrom
adrenergicneurons.
Metabolom
ics
Influences
executivefunctions
intheprefrontalcortex.
3′-U
TR,rs553668(D
raI),
“T”
0.94,0.12,0.638[13]
-Associationwith
performance
inTMT[review21].
Epinephrine
(EPI)
Neuroim
agingandanim
alstudiessupportalterationin
EPI
inADHD.
Metanephrine(M
)Mainmetabolite
ofEPI.
Metabolism
enzymes
Catechol-O-
methyltransferase
(COMT)
Enzym
ecatalyzing
the
inactiv
ationof
DAwithin
the
PFC.E
xpressed
infrontal
lobe
regionsof
brain.
ExonIV,rs4680(158Val/
Met),valine,higher
activ
ity
-,1.02,0.7267[48••]
-interaction:
COMT(val/val)-
MAOA(3R)andhigh
intelligence[49].
ItregulatessynapticDAlevels.
Genom
ics/
metabolom
ics
MonoamineoxidaseA
(MAOA)
Encodes
aproteininvolved
inthemetabolism
ofDA,5-H
T,andNE.
Prom
oter,30-bp
VNTR,2-
and3-Rareconsidered
‘low
-activity
’alleles
1.02,0.01,0.464[13]
-Associatio
n4-
and5R
alleles
with
morecommission
errors[54];
-Interactio
n:COMT(val/val)-
MAOA(3R)andhigh
intelligence[49].
Itisassociated
toim
pulsive,
aggressive
behavior.
Intron,rs12843268,“A
”,-Gender-specificassociationATT
haplotypeandpoor
motor
control(boys),visuo-spatial
working
mem
ory(girls)[55];
MAOAknockout
mouse
show
edincreasedlevelsof
aggressive
behavior
and
monoaminergic
neurotransmitterlevels.
Intron,rs3027400,“T”,
Exon,rs1137070,“T
”-A
ssociationof
rs12843268
andsymptom
sandreward
deficiency
orinsufficient
response
inhibitio
n[56].
Dopam
ineBeta
Hydroxylase
(DBH)
Catalyzes
theconversion
ofDA
into
NE.
Intron
V,rs2519152
(TaqI),
“T”/A2allele,associated
with
reducedDBH
activ
ity
1.12,0.43,0.206[13]
-AssociationA2/Tallelewith
poorer
performanceson
temporalo
rder
judgment
task,m
orecommission
and
omission
errorsandgreater
RTV(ontheSA
RT),more
errorson
theWCST
(problem
-solving)andthe
MFF
T(cognitiv
eim
pulsiveness).
Curr Psychiatry Rep (2014) 16:497 Page 5 of 20, 497

Tab
le1
(contin
ued) Sy
stem
sPo
tentialb
iomarker
Genefunction/rationale
Location,polymorphism,
risk
allele,functionality
Meta-analysis(O
R,χ
2 ,pvalue)
GWAS/
CNVs
studies
Neuropsychological
endophenotypes
Expressed
inthePF
C.
[reviews21,22].
CNSdevelopm
ent
pathways
5′Flank,rs1611115(-1021
C/T),Talleleassociated
with
reducedDBH
activ
ity,reductio
nof
DA-to-NEconversion
1.05,0.16,0.692[13]
-Executivefunction:Association
Calleleandglobalexecutive
functionandmoreerrorsof
commission
andom
ission
ontheCPT
[57].
Genom
ics/
metabolom
ics
Brain-D
erived
NeurotrophicFactor
(BDNF)
Encodes
aneurotrophin
prom
otingneurogenesis,
developm
ent,functio
nal
maintenance,and
plasticity
ofneurons.
Prom
oter,rs6265,“G
”,alleleValassociated
with
increasedactiv
ity-
dependentsecretionof
BDNF
1.01,0.02,0.406[13]
[34]
-Noassociationwith
Digitspan
[review21].
Expressed
inpons,
hippocam
pus,frontalcortex,
colliculi,
andolfactorytract.
Intron,rs2049046,Intron,
rs11030104
-Noassociationwith
Digitspan
[review21].
Genom
ics
Synaptosom
al-associated
protein25
(SNAP-25)
Encodes
aproteininvolved
inaxonalgrow
th,synaptic
plasticity,docking,and
fusion
ofsynapticvesicles
inpresynaptic
neurons.
3′-U
TR,rs3746544,
unknow
n1.15,4.71,0.03
[13]
Expressed
inhippocam
pus.
3′-U
TR,rs1051312,“T”
1.06,0.30,0.298[13]
Metabolom
ics
Environmentalrisk
factors
Ferritin(ironstore)
Coenzym
eof
tyrosine
hydroxylaseandMAO.Iron
deficiency
isassociated
with
ADHDsymptom
s.Zinc(Zn)
Cofactorformetabolism
relevant
toneurotransmitters,
anditaffectsDAmetabolism.
Zincdeficiency
isassociated
with
behavior
problemsin
ADHD.
Oxidativ
estress
Somestudiesshow
edsome
treatm
entefficacyforanti-
oxidantcom
pounds.
Potentialb
iomarker
Neurophysiological
endophenotypes
Variations
inbrainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
Biochem
ical/proteom
ics
endophenotypes
Geneexpression
Pharmacogenetics/drugseffect
Dopam
inetransporter
(DAT1,SL
C6A
3)-A
ssociatio
n10Rwith
smallercaudatevolumes
particularly
intheleft
than
righth
emisphere
[14].
-Associatio
n10Randmore
activations
correspondingto
inhibitio
ntasks[15];
-Association10Randincreased
MPH
response,but
also
contrastingresults
[reviews
16•,17••];
-Association10Randactiv
ation
instriatum
andcerebellu
mverm
is[18];
-Noassociation10RandMPH
response
[meta-analysis
[19••].
-Association10Randactiv
ation
inleftstriatum
,right
dorsal
497, Page 6 of 20 Curr Psychiatry Rep (2014) 16:497

Tab
le1
(contin
ued)
Potentialb
iomarker
Neurophysiological
endophenotypes
Variations
inbrainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
Biochem
ical/proteom
ics
endophenotypes
Geneexpression
Pharmacogenetics/drugseffect
prem
otor
cortex,and
bilateral
temporoparietaljunction
[20];
-Associatio
n9R
andgreater
neuralactiv
ation[23];
-Associatio
n9R
andincreased
DATactiv
ityinstriatum
[24];
-Neuroim
aging/
Pharmacogenetics[review25••]:
a)Association10Ralleleand
higher
regionalcerebral
bloodflow
andwith
poorer
MPH
treatm
entresponse;
b)Associatio
n10Randgreater
DATavailabilityin
basal
ganglia
aftertreatm
entand
MPH
poorer
treatm
ent
response.
-Haplotype
studies:
a)Association10/6
and
dopaminergicreward-
processing
circuitry[26];
b)Associatio
n10/3
andspatial
inattention[27];
-Haplotype
studies:
a)Association10/6
and
dopaminergicreward-
processing
circuitry[26];
b)Associatio
n10/3
andspatial
inattention[27];
Dopam
ineD4receptor
(DRD4)
-Associatio
n7R
with
thinnerprefrontaland
parietalcortex;
-low
ermRNAexpression
levels[31].
-Associatio
n7R
andincreased
MPH
response,but
also
contrastingresults;
-Associatio
n7R
with
adelayin
cerebralcortical
maturation[32,33••].
-Associatio
n4R
andpositiv
elin
earim
provem
ento
nmathematicstestoutcom
eswith
increasing
MPH
dose,
absence4R
decreased
responsesto
higher
MPH
doses;
-Associatio
n4R
andpicking
behaviors,7R
with
social
Curr Psychiatry Rep (2014) 16:497 Page 7 of 20, 497

Tab
le1
(contin
ued)
Potentialb
iomarker
Neurophysiological
endophenotypes
Variations
inbrainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
Biochem
ical/proteom
ics
endophenotypes
Geneexpression
Pharmacogenetics/drugseffect
with
draw
al,w
ithincreasing
MPH
doses.
[reviews16•,17••].
Dopam
ineD2receptor
(DRD2)
-Noassociationwith
MPH
response
[35].
Dopam
ineD5receptor
(DRD5)
-Noassociationwith
MPH
response
[37].
Phenylethylamine(PEA)
-Reduced
urinelevelsinADHD
[meta-analysis39••].
-NormalizationafterAMPand
MPH
[39••].
Dopam
ine(D
A)
-Noalteratio
nin
urinelevelsin
ADHD[m
eta-analysis39••].
Hom
ovanillicacid(H
VA)
-Noalteratio
nin
urinelevelsin
ADHD[m
eta-analysis39••].
Norepinephrine
transporterprotein1
NET(SLC6A
2)
-Smallinfluence
onMPH
response
[review17••];
-Associatio
nGallelewith
greaterdecrease
inmean
omission
errorscores
after
MPH
administration
comparedto
Aallelecarriers
[40,41].
-Noassociationwith
the
cerebralvolumeor
thicknessof
thecortex
[42].
Norepinephrine(N
E)
-HigherurinarylevelsinADHD
[meta-analysis39••].
-Normalizationafterpolyphenol
complex
(Pyc),fenfluramine,
AMP[39••].
Normetanephrine(N
M)
-Noalteration
sin
urinary
levels
inADHD
after
Bon
ferron
icorrection
[meta-analysis
39••].
3-methoxy-4-
hydroxyphenylg
lycol
(MHPG
)
-Reduced
urinarylevelsin
ADHD[m
eta-analysis39••].
-Greater
reductions
afterMPH
andAMPtreatm
ent[meta-
analysis39••].
NeuropeptideY(N
PY)
-Associatio
nwith
gene
dose-
dependentincreases
inNPY
andem
otionprocessing
[44].
-Increased
plasma
concentrations
inADHD
[39••].
Alpha-2A-adrenergic
receptor
(ADRA2A
)-A
ssociatio
nGalleleandMPH
response
[review46];
497, Page 8 of 20 Curr Psychiatry Rep (2014) 16:497

Tab
le1
(contin
ued)
Potentialb
iomarker
Neurophysiological
endophenotypes
Variations
inbrainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
Biochem
ical/proteom
ics
endophenotypes
Geneexpression
Pharmacogenetics/drugseffect
-Haplotype
studies:association
betweenGGhaplotypeand
non-remission
ofADHD
symptom
swith
atom
oxetine
treatm
ent[47].
-Haplotype
studies:association
betweenGGhaplotypeand
non-remission
ofADHD
symptom
swith
atom
oxetine
treatm
ent[47].
Epinephrine
(EPI)
-Noperipheralalteratio
nsin
ADHD[m
eta-analysis39••].
Metanephrine(M
)-H
igherurinarylevelsin
ADHD,lostafterbonferroni
correctio
n[m
eta-analysis
39••].
Catechol-O-
methyltransferase
(COMT)
-Associatio
nwith
antisocialand
aggressive
behaviorsof
ADHD[50,51].
-AssociationValalleleandMPH
response
[52,53].
MonoamineoxidaseA
(MAOA)
-Reduced
plateletlevelsin
ADHD[m
eta-analysis[39••].
-NormalizationafterMPH
treatm
ent[39••].
Dopam
ineBeta
Hydroxylase
(DBH)
-Low
eractiv
ities
inserum/urine
inADHD,[39••].
Brain-D
erived
NeurotrophicFactor
(BDNF)
-HigherplasmalevelsinADHD
[58];
-AssociationValalleleandMPH
response
[59].
-noalterationin
serum
levelsin
ADHD[60].
Synaptosomal-associated
protein25
(SNAP-25)
-Associatio
nTallelewith
improved
MPH
responses;
-Associatio
nGallelewith
sleep
difficultiesandirritability
[37].
-Associatio
nTalleleandpoorer
MPH
response;
-Associatio
nCallelewith
tics
andotherabnorm
almovem
ents[37].
Ferritin(ironstore)
-Reportedreducedserum
levels
inADHD,lostafter
Curr Psychiatry Rep (2014) 16:497 Page 9 of 20, 497

treatment response inchildrenwithADHDwhocompleted8weeksof MPH treatment. In particular only a 28.6% of the subjects with10/10 genotype showed good response to MPH treatment [71].
Pharmacogenetic studies have demonstrated increased re-sponse toMPH among homozygous 10R, but there are conflict-ing results [16•, 17••]. However a recent meta-analysis [19••]shows that there is no significant association between thisVNTR and response to MPH treatment. The meta-analysis alsofound no effects on dimensions of hyperactivity/impulsivity andinattention. Thus this finding suggests that VNTR polymor-phism is not a reliable predictor of MPH treatment success inADHD, neither in relation to these specific symptom subscales.
A DAT1 VNTR at intron-8 that contains common 5-repeat(5R) and 6-repeat (6R) alleles has also been associated withADHD susceptibility [13]. Recent evidence suggests that jointconsideration in haplotypes may provide more information thancan be inferred from the analyses of single geneticmarkers. Thereare some neurophysiological studies on the haplotypes of thesetwoDAT1 variants: the 10/6 haplotype (formed by the 10R alleleof the 3’UTRVNTR and the 6R allele of the intron 8 VNTR) isassociated with dopaminergic reward-processing circuitry whosedeficits are linked toADHD [26]. The haplotype 10/3 (formed bythe 10R allele of the 3’UTRVNTR and the 3R allele of the intron8 VNTR) predicts spatial inattention suggesting a functionaleffect of this haplotype on cognitive performance in ADHD [27].
Other variants of DAT1 have been studied. rs6350 wasassociated to alerting and executive control performance usingthe attention network test [29]. Moreover a haplotype(rs403636 (G)/rs463379 (C)/rs393795 (C)/rs37020 (G)) wasassociated with spatial working memory in ADHD [28].
Dopamine D4 Receptor (DRD4)
The DRD4 gene is a good candidate for ADHD because of itshigh expression in brain regions implicated in attention andinhibition such as anterior cingulate cortex [76]. DRD4 wasthe first associated with novelty-seeking, a personality traitwhich is a common in ADHD. It has been demonstrated thatmRNA expression levels ofDRD4were lower in ADHD [31].
DRD4 is located on chromosome 11p15.5. A highly poly-morphic functional VNTR in the third exon has been frequent-ly investigated in association studies. It comprises 11 copies ofa 48-bp repeat sequence, where 4, 7, and 2R repeat alleles arethe most prevalent. A meta-analysis showed that the 7R allelewas associated with ADHD [13, 30••].
Recent reviews on neuropsychological endophenotypes[21, 22] concluded that, although more studies are needed,the 7R allele is associated with some cognitive markerssuch as speed of processing, set shifting, and cognitiveimpulsiveness, but not with response inhibition (the stopand go/no-go tasks). More robust evidence suggests thatthe absence of this allele is linked to high reaction timevariability (RTV).T
able1
(contin
ued)
Potentialb
iomarker
Neurophysiological
endophenotypes
Variations
inbrainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
Biochem
ical/proteom
ics
endophenotypes
Geneexpression
Pharmacogenetics/drugseffect
Bonferronicorrection[m
eta-
analysis39••].
Zinc(Zn)
-Reportedreducedserum/
plasma/urinelevelsinADHD
[meta-analysis39••].
Oxidativ
estress
-Normallevelsof
anti-oxidant
productionin
ADHD,but
theirresponse
tooxidative
stress
isinsufficient
[meta-
analysis61••].
Note:
ADHD:Attention-deficit/h
yperactiv
itydisorder;OR:Odd
Ratio;χ2:chi-square;VNTR:Variablenumbertandem
repeat;UTR:Untranslatedregion;MPH
:methylphenidate;DA:Dopam
ine;
WGAS:Genom
eWideAssociatio
nstudy;CNVs:Copynumbervariatio
ns;M
RI:Magnetic
resonanceim
aging;TMT:T
railmakingtest;R
TV:R
eactiontim
evariability;C
PT:C
ontin
uous
performance
test;
SART:S
ustained
attentiontoresponse
test;T
OL:T
owerof
Londontest;W
ISC-III:W
echslerintelligencescaleforchild
ren-III;SOPT:S
elforderedpointin
gTest;F
DI:Freedomfrom
distractibilityindex;
IQ:Intellig
ence
quotient;A
MP:
Amphetam
ine;MFF
T:M
atchingfamiliar
figurestest
497, Page 10 of 20 Curr Psychiatry Rep (2014) 16:497

Tab
le2
Dataextractedfrom
endophenotypes
studiesto
define
apotentialsignature
seto
fbiom
arkersuseful
forADHDdiagnosis
Potentialb
iomarker
Rationale
Meta-analysis
Review
Neuropsychological
endophenotypes
Neurophysiological
endophenotypes
Variatio
nsin
brainanatom
y(structuralM
RIfinding)
Variatio
nsin
brainfunctio
n(functionalM
RIfinding)
EEGTheta/Betaratio
(TBR)
ElevatedTheta/Betaratio
(TBR)in
ADHD.
[62••]
[63]
-Prognostic
measure
[62••];
-potential
biom
arker[63].
Reactiontim
evariability
(RTV)
IncreasedRTVinADHD.[100••]
Stablefeatureof
ADHD
butn
ospecific.
Selectiv
eattentionandvigilance/
sustainedattention
Attentionim
pairmentin
ADHD.
[64–66].
[10••]
-Robustfindings[10••];
-Noassociation[64–66].
Verbaland
visuo-spatial
working
mem
ory
Anomaliesin
prefrontal
corticalregiones
and
thebasalg
angliain
ADHD.A
lteratio
nsin
dopaminergic,
noradrenergicand
fronto-striatalsystems.
[64–67]
[10••]
-Robustfindings
[10••,67,68];
-Noassociation[64–66]
Caudate
Neuroimagingstudies
supporteddysfunctionof
severalareasofthebrain.
[102••]
Reduced
gray
mattervolume
Reduced
activ
ityand
functio
nalconnectivity
Thalamus
Reduced
gray
mattervolume
Reduced
activ
ityand
functio
nalconnectivity
Anteriorcingulated
Reduced
gray
mattervolume,
reducedwhitematterintegrity
Reduced
activ
ityand
functio
nalconnectivity
Prefrontalcortex
Reduced
gray
mattervolume,
reducedwhitematterintegrity
Reduced
activ
ityand
functio
nalconnectivity
Premotor
andSM
Acortex
Reduced
gray
mattervolume
Reduced
activ
ityand
functio
nalconnectivity
Superiorparietalcortex
Reduced
gray
mattervolume
Reduced
activ
ityand
functio
nalconnectivity
Precuneus,posteriorcingulate,
lateralp
arietalcortex,medial
frontalcortex(default-mode
network)
Reduced
activ
ityand
functio
nalconnectivity
Cerebellum
(posterior
inferior
verm
is)
Reduced
gray
mattervolume,
reducedwhitematterintegrity
Reduced
activ
ityand
functio
nalconnectivity
Corpuscallo
sum
(splenium/
isthmus)
Reduced
whitematterintegrity
Fasciculus
longitu
dinalis
superior
Reduced
whitematterintegrity
Anteriorcorona
radiate
Reduced
whitematterintegrity
Note:ADHD:A
ttention-deficit/h
yperactiv
itydisorder;M
RI:Magnetic
resonanceim
aging;
SMA:S
upplem
entary
motor
area
Curr Psychiatry Rep (2014) 16:497 Page 11 of 20, 497

Neuroimaging studies found associations of 7R allele withthinner prefrontal and parietal cortex as well as with delay incerebral cortical maturation [32, 33••].
Pharmacogenetic studies suggest an enhanced response toMPH among 7R carriers even though conflicting results havebeen obtained [16•, 17••]. Moreover some studies have sug-gested that different DRD4 genotypes exhibit different MPHdose–response curves [16•, 17••]. One study demonstrated thatindividuals with one or two copies of the 4R allele had positivelinear improvement on mathematics test outcomes with increas-ing MPH dose, while those lacking a copy showed greaterimprovement at lower doses, and deterioration at higher doses.A second study found that those lacking any 4R allele haddecreasing responses to higherMPHdoses compared with othergenotypes. Both these studies are consistent with the ideathat the 7R codes for a defective dopamine receptor thatresponds less effectively at higher doses, an idea with somepotential for clinical relevance if replicated in larger samples.One interesting finding suggests that DRD4 might play arole in predicting susceptibility to medication side effects. Inthe Preschool ADHD Treatment Study, MPH treated ADHDchildren homozygous for the 4R were three-times morelikely to develop picking behaviors, while those with at leastone copy of the 7R were four-times more likely to developsocial withdrawal with increasing doses. Thus, 7R carriersappear to be a distinct subgroup of ADHD patients whichexperiences differential medication effects.
Other variants in the promoter region have also been stud-ied, focusing on a 120-base pair duplication (120-bp dup),−521C/T (rs1800955), −616C/G (rs747302), −615A/G, and−376C/T, located in the 5′ untranslated region. However twometa-analyses conducted on 120-bp dup and −521C/Tshowed no association with ADHD [13, 30••].
Dopamine D2 receptor (DRD2)
The DRD2 gene is located at chromosome 11q23.1. Therationale for its involvement in ADHD is based on the evi-dence that it is expressed in those brain areas relevant for thepathology and it is implicated in the regulation of mesolimbicreward pathways [77]. A recent genome wide associationstudy reported a nominal association of this gene withADHD susceptibility [34]. The TaqIA (rs1800497) variantof DRD2 was believed to connect with urinary level of theDA metabolite homovanillic acid (HVA) and DRD2 expres-sion levels. According to Wu et al.’s meta-analysis, an asso-ciation of this polymorphism was reported [30••]. However,due to excessive heterogeneity, the authors concluded that thisresult was invalid. Pharmacogenetic studies have found noassociation of this variant with MPH response [35]. Twoother DRD2 variants, rs2075654 and rs1079596, havebeen associated with greater commission error in ADHD[21].
Dopamine D5 Receptor (DRD5)
DRD5 maps to chromosome 4p15.3. In situ hybridizationstudies demonstrated that the expression of the gene is higherin hippocampus, a brain area involved in ADHD pathogenesis[78]. Moreover functional studies showed that DRD5 is impli-cated in synaptic strength in hippocampal memory formation[79]. These studies support the rationale on the involvement ofDRD5 gene in ADHD. The association between ADHD and ahighly polymorphic dinucleotide repeat of DRD5 ((CA)n),located in 18.5 kb at the end of the 5′ flanking region, has beenthemost studied. This variant comprises 12 alleles ranging from134 to 156 bps in length, among which the 148-bp and 136-bpalleles are the most common. The 148-bp allele was a riskfactor for ADHD according meta-analyses, while that of 136-bp allele was a protective factor for ADHD [13, 30••].
Interestingly, the 148 bp allele associated to commis-sion errors, omission errors, RTs, and RTV [36], whereasno association was detected with cognitive test scores [38].Pharmacogenetic studies found no association of MPHresponse with the 148-bp allele, whereas the 151-bp allelehas been linked to a favorable MPH response [37].
Metabolomics
Phenylethylamine (PEA)
In dopaminergic neurons of the nigrostriatal system, the dopamine(DA) receptor agonist PEA is synthesized by the decarboxylationof phenylalanine. PEA stimulates the release of DA and, accord-ing to a meta-analysis [39••], urinary levels of PEA are signifi-cantly lower in patients with ADHD compared with controls.Interestingly decreased levels of PEA have also been associatedwith symptoms of inattentiveness and administration ofAMP andMPH markedly increased urinary excretion of PEA [39••].
DA and HVA
DA and its mainmetabolite HVA showed no difference in urineexcretion in ADHD patients compared with controls [39••].
Potential Noradrenergic Biomarkers for ADHD
Genomics
Norepinephrine Transporter Gene (NET1, SLC6A2)
The SLC6A2 gene codes for the norepinephrine transporter,which is responsible for the reuptake of norepinephrine (NE)from the synaptic cleft back into the presynaptic neuron and istargeted by atomoxetine, a medication for ADHD. NET1 ismost highly expressed in the frontal lobes where it plays a role
497, Page 12 of 20 Curr Psychiatry Rep (2014) 16:497

in noradrenergic and dopaminergic reuptake [80]. A frequent-ly studied SNP (rs5569 or G1287A) located in exon 9 was notsignificant in a meta-analysis of candidate gene studies [13],but a nominal association was observed in a genome-wideassociation study [34]. No association was found between thisvariant and CPT performance [21]. A recent review on phar-macogenetics concluded that NET1 influence on response toMPH is small [17••], although a recent genome-wide associ-ation study found two SNPs in SLC6A2 gene associated withMPH response [81]. In a study rs3785143 was associated withatomoxetine response [47].
Moreover childrenwith theG/G genotype at G1287A showeda greater decrease in mean omission error scores after MPHadministration compared to A allele carriers. Another polymor-phism A-3081T at 5’flank, demonstrated to be functional, wasassociated with T allele with greater decrease in mean commis-sion errors scores, meaning improved impulsive behavior [40,41]. Other variants such as rs998424 and rs3785157 were linkedto high performance on the similarities subtest of the WISCIII,but not with the cerebral volume or thickness of the cortex [42].Another SNP rs3785155 was associated to RTV [43].
Metabolomics
Norepinephrine (NE)
Concerning NE levels, a meta-analysis [39••] indicated higherurinary levels in ADHD compared with controls, and a nor-malization of NE concentrations has been observed aftertreatment with polyphenol complex and with fenfluramineand AMP [39••].
3-Methoxy-4-Hydroxyphenylglycol (MHPG)
MHPG is the main metabolite of NE. Its urinary levelswere lower in ADHD patients and stimulant trialsshowed that decreases in ADHD symptoms with treat-ment were associated with greater reductions in urinaryMHPG excretion [39••].
Neuropeptide Y (NPY)
NPY frequently colocalizes with catecholamine systems. Itparticipates in the regulation of feeding, circadian rhythms,reproduction, and thermoregulation. Oades et al. found in-creased plasma NPY concentrations in ADHD children com-pared with controls [82]. Moreover a recent genome-widecopy number variation analysis [44] found that NPY wasincluded in a rare 3 Mb duplication on chromosome 7p15.2to 15.3 and an association of this duplication was found withincreased NPY plasma concentrations. Moreover an associa-tion was observed between gene dose-dependent increases inNPY and emotion processing [44].
Potential Adrenergic Biomarkers for ADHD
Genomics
Alpha-2A-adrenergic receptor (ADRA2A)
ADRA2A in the prefrontal cortex influences executive func-tions impaired in ADHD [83]. It is also a target for twoADHDmedications: guanfacine and clonidine. A-1291 C>G createsan MspI site in the promoter region and it is a functionalpolymorphism. Two meta-analyses confirmed no associationwith ADHD [13, 45]. However the C allele was associatedwith TOL, trail making test (TMT), and RTV on the stop-signal task as well as the CC genotype was associated withhigh RTV on the CPT test [21]. A recent pharmacogeneticreview [17••] summarized some studies reporting that G alleleis associated with greater reduction of inattentive symptomsover time. Froehlich et al. also found a main effect of thisgenetic variant on MPH response [46]. However, the G allelewas associated with hyperactive-impulsive symptoms on pla-cebo and across doses.
Another variant rs553668, for which the disorder meta-analysis was negative [13], was associated with the perfor-mance on TMT suggesting an involvement of this gene onexecutive functions [21]. Yang et al. described an associationbetween GG haplotype (rs1800544/rs553668) and non-remission of ADHD symptoms with atomoxetine treatment[47].
Metabolomics
Epinephrine (EPI) and Metanephrine (M)
EPI and its main metabolite M levels, according to a recentmeta-analysis [39••], were respectively no different or higherin ADHD patients. The difference for M was lost afterBonferroni correction.
Potential Metabolism Enzymes as Biomarkers for ADHD
Genomics/Metabolomics
Catechol-O-Methyltransferase (COMT)
COMT is an enzyme responsible for the degradation of DAand NE. It is highly expressed in frontal lobe where it regu-lates synaptic DA levels [84]. Studies examining the associa-tion between COMT and ADHD have largely focused on afunctional SNP in exon 4 that creates an amino acid substitu-tion (valine/methionine). This variant affects COMT enzymeactivity, such that homozygotes for the valine allele shows 3–4times greater activity than homozygotes for the methionine
Curr Psychiatry Rep (2014) 16:497 Page 13 of 20, 497

allele. However meta-analysis [48••] indicates no associationbetween ADHD and the 158Val/Met.
A review summarized studies on 158Val/Met and neuropsy-chological endophenotypes [21]. While no association wasobserved between this variant and performance on cognitivetasks, a significant finding was reported between the Met alleleand impairment in sustained attention and fewer commissionerrors. In the neurophysiological studies, this variant was asso-ciated with antisocial and aggressive behaviors of ADHD [50,51]. Pharmacogenetic studies reported a positive associationbetween Val allele and response to MPH [52, 53].
Monoamine Oxidase A (MAOA)
TheMAOA gene encodes a protein involved in themetabolism ofDA, 5-HT, and NE. It has been associated with impulsive,aggressive behavior [85]. Moreover an MAOA knockout mouseshowed increased levels of aggressive behavior and monoamin-ergic neurotransmitter levels [86]. Recent studies have focusedlargely on a functional 30-bp VNTR 1.2 kb upstream of the gene,previously associatedwith impulsivity and aggression. Accordingthe classification system [87], the 2- and 3R are considered ‘low-activity’ alleles, the others are high-activity’ alleles. A recentmeta-analysis reported no association with ADHD [13].According to neuropsychological studies, the 4 and 5R allelesare associated with commission errors on the TOVA test, and aMPH administration attenuated this association [54].
Moreover a gender-specific association has been reportedwiththe ATT haplotype (rs12843268/rs3027400/rs1137070) [55]. Inboys, the ATT haplotype was associated with motor control; ingirls with visuo-spatial working memory. This suggests thatMAOA genotype influences cognitive and motor functioning inADHD and sex moderates these effects. This may be related tobiological differences between males and females in serotoninneurotransmission. Finally Nymberg et al. reported a sex-specificassociation between ADHD symptomatology and reward defi-ciency or insufficient response inhibition depending onrs12843268 genotype [56]. Concerning blood levels, a recentmeta-analysis indicated reduced levels in ADHD compared withcontrols [39••]. MAO levels were associated with impulsivenessand MPH treatment normalized the reduced levels.
Dopamine Beta Hydroxylase (DBH)
TheDBH gene encodes an enzyme that catalyzes the conversion ofDA into NEwhich is particularly expressed in the prefrontal cortex[88]. A–1021C/T variant (rs1611115) accounts for up to 50 % ofthe variation of plasma DBH activity, but it is the intron 5 TaqIvariant (rs2519152) that has been most often tested. A meta-analyses of these variants reported no associationwithADHD [13].
Two recent reviews of neuropsychological endophenotypessummarized studies of DBH [21, 22]. In particular the A2/Tallele was found associated with poorer performances on a
temporal order judgment task, more commission and omissionerrors and greater RTV on the SART, more errors on theWisconsin Card Sorting Test (WCST) (problem-solving) andthe Matching Familiar Figures Test (MFFT) (cognitiveimpulsiveness).
Finally an association between neuropsychological mea-sures of executive function in children with ADHD and the –1021C/T variant has been reported [57].
A meta-analysis [39••] found that ADHD patients showedlower activities of DBH in serum and urine and it was sug-gested that decreased DBH levels correlate with ADHDsymptoms [39••, 89, 90••].
Potential CNS Development Biomarkers for ADHD
Genomics/Metabolomics
Brain Derived Neurotrophic Factor (BDNF)
BDNF has been mapped to chromosome 11p13. It belongs to afamily of neurotrophins, involved in promoting neurogenesis,neuronal survival, and synaptic plasticity [91]. A genome widestudy supported a nominal association with ADHD susceptibil-ity [34]. A common variant resulting in a valine to methionineamino acid substitution at codon 66 (Val66Met; rs6265) regu-lates the intracellular tracking and activity-dependent secretionof BDNF in brain. A meta-analysis found no association of thisvariant with ADHD [13]. This variant and two others,rs2049046 and rs11030104, were not associated with the DigitSpan test [21]. Moreover better MPH response in ADHD chil-dren who are Val/Val homozygous was reported [59].
Concerning peripheral levels, higher plasma concentrationswere observed in ADHD patients as compared with controls[58], whereas no alterations were observed in serum levels [60].
Synaptosomal-Associated Protein 25 (SNAP-25)
SNAP-25 codes for a protein involved in axonal growth,synaptic plasticity, and in the docking and fusion of synapticvesicles in presynaptic neurons necessary for the regulation ofneurotransmitter release [92]. The major variants studied werers362987 (intron 4), rs363006 (intron 6), rs3746544 (3′UTR),and rs1051312 (3′UTR). A main effect on ADHD suscepti-bility was observed only with rs3746544 [13].
Concerning pharmacogenetic studies, homozygotes forthe T allele of T1065G had moderately improved MPHresponses, whereas homozygotes for the G allele developedsleep difficulties and irritability. Moreover, homozygotes forthe T allele at T1069C exhibited poorer MPH response,whereas homozygotes for the C allele developed tics andother abnormal movements [37].
497, Page 14 of 20 Curr Psychiatry Rep (2014) 16:497

Potential Biomarkers for Environmental Risk factors
Metabolomics
Iron
It is a coenzyme of tyrosine hydroxylase andMAO, which arecritical in the synthesis and degradation, respectively. Ferritinis a major intracellular iron storage protein and serum ferritinlevels are an indication of iron stores in the body and brain.Children with iron deficiency exhibited ADHD symptoms,such as inattention, hyperactivity, and/or impulsivity [review90••]. Ameta-analysis indicated that serum ferritin levels werelower for ADHD patients compared with controls, althoughthe finding was lost after Bonferroni correction [39••].
Zinc
Zinc is another essential cofactor for neurotransmitter me-tabolism which affects DA metabolism. Zinc deficiency isassociated with behavior problems in children with ADHD[review 90••]. Although a systematic review of randomizedcontrolled clinical trials demonstrated that using zinc, eitheralone or in combination with stimulants, did not improveADHD symptoms [93], a meta-analysis reported reducedserum/plasma/urine levels in ADHD [39••].
Oxidative Stress
The normal oxidation-reduction reactions that create energy in thecell create toxic oxidants or reactive oxygen species. These by-products of normal oxidation-reduction reactions are highly un-stable. Antioxidants counteract the effects of oxidants. Whenantioxidants are not sufficient, oxidative stress occurs and dam-ages cellular proteins, lipids, carbohydrates, and nucleic acids.Brain tissue is especially susceptible to oxidative stress. Indirectevidence for oxidative stress in ADHD comes from studies show-ing some treatment efficacy for anti-oxidant compounds such asomega-3 fatty acids, pycnogenol and N-acetylcysteine [94–97].
A meta-analysis of oxidative stress in ADHD suggestedthat patients with ADHD have normal levels of anti-oxidantproduction but that their response to oxidative stress is insuf-ficient, leading to oxidative damage [61••].
Potential Endophenotypes as Biomarkers for ADHD
Besides indentifying a genetic marker involved in neurophysio-logical/neuropsychological, neuroimaging, pharmacogenetics,biochemical features, a putative signature set of biomarkerscould be integrated with a research on endophenotypes(Table 2).
EEG Theta/Beta Ratio (TBR)
TBR is a measurement of the power (wave amplitude squared)produced by brain waves altering between 3.5 and 28.0 Hz inchildren with ADHD [98]. A genetic overlap between ADHDsymptoms and EEG theta power has been reported [99] andthis supports a recent review suggesting TBR as a potentialbiomarker for ADHD [63]. However a recent meta-analysisconcluded that excessive TBR cannot be considered a reliablediagnostic measure of ADHD but it could have a prognosticvalue [62••]. In June 2013 FDA approved NeuropsychiatricEEG-Based Assessment Aid (NEBA) System, a test recordingthe type and number of brain waves that nerve cells give offeach second.
In summary, as also reported in Thome et al. the neurophys-iological markers must be viewed as promising candidates forbiomarkers of ADHD but further studies are needed [10••].
Reaction Time Variability (RTV)
Individuals with ADHD are described frequently as ubiqui-tously slower and more variable than their unaffected peers,and ADHD-related RTV is considered by many to reflect aunique, stable, and etiologically important characteristic of thedisorder. Results of a recent meta-analysis indicate that, con-trary to contemporary characterizations of individuals withADHD as slower and more variable, ADHD individualsmay be better characterized as more variable but not slowerafter accounting for their increased response variability.However RTV lacks specificity among clinical disorders,and thus is not a viable diagnostic marker of ADHD [100••].
Selective Attention and Vigilance/Sustained Attention
A prominent multidimensional model of attention differentiatesbetween alertness, divided attention, selective attention,vigilance/sustained attention and shifting. According toThome et al. the most robust findings indicated that childrenwith ADHD displayed difficulties in measures of selectiveattention and vigilance/sustained attention [10••]. Impairmentsof vigilance/sustained attention are the most replicated neuro-psychological finding inADHD.However a considerable num-ber of investigations were unable to find difference betweenADHD patients and controls [64–66]. In addition effect sizesindicated only small to moderate differences between ADHDpatients and controls.
Executive Functions
They are an umbrella term encompassing various functions ofhigher cognitive functioning including planning, problemsolving, concept formation, fluency, cognitive flexibility, andworking memory. The most consistent finding in the domain
Curr Psychiatry Rep (2014) 16:497 Page 15 of 20, 497

of executive functions is that patients with ADHD displayimpairments in working memory. Working memory measuresappear therefore to be the most sensitive indicator of executivedysfunctioning in ADHD [10••]. Additional studies supportthis finding [67, 68]. However, as for vigilance/sustainedattention, no difference was found between patients and con-trols with small to moderate effect sizes [64–66].
In summary, although promising results were obtained forselective attention and vigilance/sustained attention and work-ing memory, no sensitive or specific profile based on neuro-psychological tests assessing cognition in ADHD has as yetbeen found [101••].
Brain Differences in ADHD
Kasparek et al. reported alterations in several areas of thebrain, particularly the anterior cingulum, the dorsolateral aswell as ventrolateral prefrontal cortex, the orbitofrontal cortex,the superior parietal regions, the caudate nucleus, the thala-mus, the amygdala and the cerebellum. Imaging studies pointto the persistence of changes in both brain structure andfunction into adulthood, although there might be a tendencyfor improvement of caudate nucleus pathology. Growing ev-idence suggests that MPH treatment can lead to improvementof brain changes seen in neuroimaging by its positive effect onneuroplasticity [102••].
Conclusion
The study of psychiatric biomarkers is very complex becauseof the heterogeneous nature of psychiatric disorders which areoften referred to as syndromes with several subtypes, notuniform problems. Therefore, a single biomarker is very un-likely to provide enough information to identify cellular andmetabolic pathways involved in a particular individual.Identification of a signature set of biomarkers for disordersubtypes each based on their underlying biological pathwayswill be the most effective for diagnosis and treatmentselection.
Starting from the literature, we have built an hypotheticalpyramid representing a putative set of biomarkers where at thetop there are potentially useful biomarkers for ADHD and atthe bottom those that are probably not useful (Fig. 1).Following Pies et al.’s [12] example, a potential genetic-biomarker could be represented by the variants in DAT1 andDRD4 genes as the best candidates as biomarkers for ADHD.Indeed the 10R allele at the 3′UTR of SLC6A3, is associatedwith specific neuropsychological tasks [21, 22], generatesmore activation in specific brain areas [33••], and is associatedto MPH response [16•, 17••]. Similarly the 7R allele at exon 3ofDRD4 is involved in specific neuropsychological tasks [21,22], brain structure [32, 33••], MPH response [16•, 17••], andexpression levels of DRD4 [31].
Fig. 1 Hypothetical pyramid representation of possible signature sets ofbiomarkers for ADHD diagnosis. Starting from the literature, we built anhypothetical pyramid describing a putative set of biomarkers where, atthe top, variants in DAT1 and DRD4 genes are the best candidates asuseful biomarkers, for their associations to neuropsychological tasks,activation in specific brain areas, methylphenidate response and geneexpression levels. A further level is represented by the noradrenergicsystem (Norepinephrine transporter, Norepinephrine, 3-methoxy-4-hydroxyphenylglycol, Monoamine Oxidase, Neuropeptide Y) for theiraltered peripheral levels, their association with neuropsychologicaltasks, symptomatology, drugs effect and brain function. Other minorputative genetic biomarkers could be Dopamine Beta Hydroxylase and
Catechol-O-methyltransferase. In the bottom, we placed endophenotypebiomarkers. In particular working memory, selective attention vigilance/sustained attention, Theta/beta ratio, Reaction Time Variability andneuropsychological endophenotypes are, in this order, probably notuseful biomarkers for ADHD diagnosis. Notes: Dopamine Transportergene (DAT1, SLC6A3); Dopamine D4 receptor (DRD4);Norepinephrine transporter (NET1, SLC6A2); Norepinephrine (NE);3-methoxy-4-hydroxyphenylglycol (MHPG); Monoamine oxidase(MAO); Neuropeptide Y (NPY); Dopamine beta hydroxylase (DBH);Catechol-O-methyltransferase (COMT); Theta/beta ratio (TBR);Reaction time variability (RTV)
497, Page 16 of 20 Curr Psychiatry Rep (2014) 16:497

Interesting data also come from the noradrenergic system.In fact, according to metabolomic studies, NET1, NE, itsmetabolite MHPG and MAO along with NPY could representgood candidates as biomarkers for ADHD, based on theirperipheral levels altered in ADHD [39••], their associationwith neuropsychological tasks [21, 40–43, 54–56], symptom-atology, drugs effect [39••], and brain function [44].
Other minor putative genetic biomarkers could be the twoenzymes DBH and COMT. Although the meta-analyses ofTaq1 and -1021 T/C in DBH gene and 158Val/Met inCOMT gene have not identified susceptibility alleles, variantsof these genes are associated with neuropsychological perfor-mance [21, 22, 57], neurophysiological features [50, 51], brainfunction [103], peripheral levels [39••, 89], and MPH re-sponse [52, 53].
Concerning the identification of endophenotypebiomarkers, to date it is not possible to characterize an accu-rate biomarker for ADHD. As described in [10••, 101••, 104],no biomarker has sufficiently high specificity and sensitivityfor the ADHD diagnosis. Currently proposed biomarkers arelimited by age, gender, and drug effects and studies have beenlimited by small sample sizes and poor control for multipletesting.
This work supports the necessity to take into account the deepintegration of “omics” sciences such as “pharmacogenomics”,“phenomics”, “epigenomics,” “proteomics”, “transcriptomics”,and “metabolomics”. In fact, a better understanding of theinteraction network of genes, proteins, and biochemical pro-cesses in relation to more accurate clinical profiles, by usingnew high-throughput computational methods, will allow usto identify a list of biomarkers both for the optimization ofdiagnostic assessment as well as for the personalization oftherapies.
Acknowledgments This research was supported by grants from theFondazione Mariani (RF2006) and from the Italian Ministry of Health(Ricerca Corrente).
Compliance with Ethics Guidelines
Conflict of Interest Cristian Bonvicini Ph.D., Catia Scassellati declarethat they have no conflict of interest.
In the past year, Stephen V. Faraone received consulting income,travel expenses and/or research support from Ironshore, Shire, AkiliInteractive Labs, Alcobra, VAYA Pharma, and SynapDx and researchsupport from the National Institutes of Health (NIH). His institution isseeking a patent for the use of sodium-hydrogen exchange inhibitors inthe treatment of ADHD. In previous years, he received consulting fees orwas on Advisory Boards or participated in continuing medical educationprograms sponsored by: Shire, Alcobra, Otsuka, McNeil, Janssen,Novartis, Pfizer and Eli Lilly. Dr. Faraone receives royalties from bookspublished by Guilford Press: Straight Talk about Your Child’s MentalHealth and Oxford University Press: Schizophrenia: The Facts.
Human and Animal Rights and Informed Consent This article doesnot contain any studies with human or animal subjects performed by anyof the authors.
References
Papers of particular interest, published recently, have beenhighlighted as:• Of importance•• Of major importance
1. Biederman J, Faraone SV. Attention-deficit hyperactivity disorder.Lancet. 2005;366(9481):237–48.
2. Faraone SV.Attention-deficit hyperactivity disorder and the shiftingsands of psychiatric nosology. Br J Psychiatr. 2013;203(2):81–3.
3. Faraone SV, Mick E. Molecular genetics of attention deficit hy-peractivity disorder. Psychiatr Clin N Am. 2010;33(1):159–80.
4.•• Akutagava-Martins GC, Salatino-Oliveira A, Kieling CC, Rohde LA,Hutz MH. Genetics of attention-deficit/hyperactivity disorder: currentfindings and future directions. Expert Rev Neurother. 2013;13(4):435–45. This is an exhaustive review on the genetics studies inADHD. It summarized that all variants associated with the pathologyexplain only a small fraction of heritability: phenotype complexity andvariants of small effect contribute to the missing heritability issue.
5. Gould TD, Manji HK. The molecular medicine revolution andpsychiatry: bridging the gap between basic neuroscience researchand clinical psychiatry. J Clin Psychiatry. 2004;65(5):598–604.
6. Insel T (2013) Director’s Blog: transforming diagnosis. http://www.nimh.nih.gov/about/director/2013/transforming-diagnosis.shtml
7. Biomarkers Definitions Working Group. Biomarkers and surro-gate endpoints preferred definitions and conceptual framework.Clin Pharmacol Ther. 2001;69:89–95.
8. Singh I, Rose N. Biomarkers in psychiatry. Nature. 2009;460(7252):202–7.
9.•• Sokolowska I, Ngounou Wetie AG, Wormwood K, Thome J,Darie CC, Woods AG. The potential of biomarkers in psychiatry:focus on proteomics. J Neural Transm. 2013Dec 20. doi:10.1007/s00702-013-1134-6. This is a review on the importance to use theproteomics to identify biomarkers for different psychiatricpathologies including ADHD.
10.•• Thome J, Ehlis AC, Fallgatter AJ, Krauel K, Lange KW, RiedererP, et al. Biomarkers for attention-deficit/hyperactivity disorder(ADHD). A consensus report of the WFSBP task force onbiological markers and the World Federation of ADHD.World J Biol Psychiatry. 2012;13(5):379–400. This is a com-prehensive review on clinical, imaging, genetic and proteomicmarkers and ADHD. The review concluded that, although somepromising candidates such as olfactory sensitivity, substantialechogenicity, no biomarkers for ADHD are available.
11. Lakhan SE, Vieira K, Hamlat E. Biomarkers in psychiatry: draw-backs and potential for misuse. Int Arch Med. 2010;3:1.
12. Pies R. Beyond reliability: biomarkers and validity in psychiatry.Psychiatry (Edgmont). 2008;5(1):48–52.
13. Gizer IR, Ficks C, Waldman ID. Candidate gene studies ofADHD: a meta-analytic review. Hum Genet. 2009;126(1):51–90.
14. ShookD, Brady C, Lee PS, Kenealy L, Murphy ER, GaillardWD,et al. Effect of dopamine transporter genotype on caudatevolume in childhood ADHD and controls. Am J Med GenetB. 2011;156B(1):28–35.
15. BraetW, JohnsonKA, Tobin CT, Acheson R,McDonnell C, HawiZ, et al. fMRI activation during response inhibition and errorprocessing: the role of the DAT1 gene in typically developingadolescents and those diagnosed with ADHD. Neuropsychologia.2011;49(7):1641–50.
16.• McGough JJ. Attention deficit hyperactivity disorder pharma-cogenetics: the dopamine transporter and D4 receptor.Pharmacogenomics. 2012;13(4):365–8. This is a review on
Curr Psychiatry Rep (2014) 16:497 Page 17 of 20, 497

pharmacogenetics and ADHD focusing on DAT1 and DRD4genes. The author concludes that there is no evidence atpresent to suggest any role for pharmacogenetics in the clin-ical management of ADHD for SLA6A3, DRD4 or any othergenetic polymorphism.
17.•• Bruxel EM, Akutagava-Martins GC, Salatino-Oliveira A, Contini V,Kieling C, Hutz MH, et al. ADHD pharmacogenetics across the lifecycle: new findings and perspectives. Am J Med Genet BNeuropsychiatr Genet. 2014. doi:10.1002/ajmg.b.32240. This is asystematic review of the literature on ADHD pharmacogenetics toprovide a critical discussion of the existent findings, new approaches,limitations, and recommendations for future research. It focuses onthe number of studies continues to grow, and the heterogeneity inmethodological strategies employed by different studies.
18. Durston S, Fossella JA, Mulder MJ, Casey BJ, Ziermans TB,Vessaz MN, et al. Dopamine transporter genotype conveysfamilial risk of attention-deficit/hyperactivity disorder throughstriatal activation. J Am Acad Child Adolesc Psychiatry.2008;47(1):61–7.
19.•• Kambeitz J, Romanos M, Ettinger U. Meta-analysis of the asso-ciation between dopamine transporter genotype and response tomethylphenidate treatment in ADHD. Pharmacogenomics J.2014;14(1):77–84. This is a a meta-analysis of studies investigat-ing the moderating effect of the SLC6A3 VNTR on response tomethylphenidate treatment in subjects with ADHD. The resultsindicate that SLC6A3 VNTR is not a reliable predictor of methyl-phenidate treatment success in ADHD.
20. Bedard AC, Schulz KP, Cook EH, Fan J, Clerkin SM, Ivanov I,et al. Dopamine transporter gene variation modulates activation ofstriatum in youth with ADHD. Neuroimage. 2010;53(3):935–42.
21. Kebir O, Joober R. Neuropsychological endophenotypes in atten-tion-deficit/hyperactivity disorder: a review of genetic associationstudies. Eur Arch Psychiatry Clin Neurosci. 2011;261(8):583–94.
22. Barnes JJM, Dean AJ, Nandam LS, O’Connell RG, BellgroveMA. The molecular genetics of executive function: role of mono-amine system genes. Biol Psychiatry. 2011;69(12):E127–43.
23. Congdon E, Constable RT, Lesch KP, Canli T. Influence ofSLC6A3 and COMT variation on neural activation during re-sponse inhibition. Biol Psychol. 2009;81(3):144–52.
24. Fusar-Poli P, Rubia K, Rossi G, Sartori G, Balottin U. Striataldopamine transporter alterations in ADHD: pathophysiology oradaptation to psychostimulants? A meta-analysis. Am J Psychiatr.2012;169(3):264–72.
25.•• Falcone M, Smith RM, Chenoweth MJ, Bhattacharjee AK, KelsoeJR, TyndaleRF, et al. Neuroimaging in psychiatric pharmacogeneticsresearch: the promise and pitfalls. Neuropsychopharmacology.2013;38(12):2327–37. This review summarizes progress in effortsto incorporate neuroimaging into genetics and treatment research onmajor psychiatric disorders, such as schizophrenia, majordepressivedisorder, bipolar disorder, attention-deficit/hyperactivity disorder,and addiction.
26. Paloyelis Y, MehtaMA, Faraone SV, Asherson P, Kuntsi J. Striatalsensitivity during reward processing in attention-deficit/hyperac-tivity disorder. J Am Acad Child Adolesc Psychiatry. 2012;51(7):722–32.
27. Bellgrove MA, Johnson KA, Barry E, Mulligan A, Hawi Z, GillM, et al. Dopaminergic haplotype as a predictor of spatial inatten-tion in children with attention-deficit/hyperactivity disorder. ArchGen Psychiatr. 2009;66(10):1135–42.
28. Shang CY, Gau SSF. Association between the DAT1 gene andspatial workingmemory in attention deficit hyperactivity disorder.Int J Neuropsychopharmacol. 2014;17(1):9–21.
29. Konrad K, Dempfle A, Friedel S, Heiser P, Holtkamp K,Walitza S,et al. Familiality and molecular genetics of attention networks inADHD. Am J Med Genet B Neuropsychiatr Genet. 2010;153B(1):148–58.
30.•• Wu J, Xiao HF, Sun HJ, Zou L, Zhu LQ. Role of dopaminereceptors in ADHD: a systematic meta-analysis. Mol Neurobiol.2012;45(3):605–20. This review summarized the ongoing re-search of DA receptor genes in ADHD pathogenesis and gatheredthe past published data with meta-analysis and revealed the highrisk of DRD5, DRD2, and DRD4 polymorphisms in ADHD.
31. Taurines R, Grunblatt E, Schecklmann M, Schwenck C,Albantakis L, Reefschlager L, et al. Altered mRNA expressionof monoaminergic candidate genes in the blood of children withattention deficit hyperactivity disorder and autism spectrum dis-order. World J Biol Psychiatry. 2011;12:104–8.
32. Shaw P, Eckstrand K, Sharp W, Blumenthal J, Lerch JP,Greenstein D, et al. Attention-deficit/hyperactivity disorder ischaracterized by a delay in cortical maturation. Proc Natl AcadSci U S A. 2007;104(49):19649–54.
33.•• Wu Z, Yang L, Wang Y. Applying Imaging Genetics to ADHD:the Promises and the Challenges. Mol Neurobiol. 2014 Apr 1. doi:10.1007/s12035-014-8683-z. This is an exhaustive review onneuroimaging genetics in ADHD. The authors conclude thatimaging genetics has somewhat showed its potential to provide amore precise understanding of how the genes shape the brainvariations and further the clinical features of ADHD.
34. Lasky-Su J, Neale BM, Franke B, Anney RJL, Zhou KX, MallerJB, et al. Genome-wide association scan of quantitative traits forattention deficit hyperactivity disorder identifies novel associa-tions and confirms candidate gene associations. Am J MedGenet B. 2008;147B(8):1345–54.
35. Winsberg BG, Comings DE. Association of the dopamine trans-porter gene (DAT1) with poor methylphenidate response. J AmAcad Child Adolesc Psychiatry. 1999;38(12):1474–7.
36. Manor I, Corbex M, Eisenberg J, Gritsenkso I, Bachner-MelmanR, Tyano S, et al. Association of the dopamine D5 receptor withattention deficit hyperactivity disorder (ADHD) and scores on acontinuous performance test (TOVA). Am J Med Genet B.2004;127B(1):73–7.
37. Froehlich TE, McGough JJ, Stein MA. Progress and promise ofattention-deficit hyperactivity disorder pharmacogenetics. CNSDrugs. 2010;24(2):99–117.
38. Loo SK, Rich EC, Ishii J, McGough J, McCracken J, Nelson S,et al. Cognitive functioning in affected sibling pairs with ADHD:familial clustering and dopamine genes. J Child PsycholPsychiatry. 2008;49(9):950–7.
39.•• Scassellati C, Bonvicini C, Faraone SV, Gennarelli M. Biomarkersand attention-deficit/hyperactivity disorder: a systematic reviewand meta-analyses. J Am Acad Child Adolesc Psychiatry.2012;51(10):1003–19. This is an exhaustive meta-analysis pro-viding evidence for several peripheral biomarkers as being asso-ciated with ADHD both in diagnosis and in treatment efficacy.
40. Park MH, Kim JW, Yang YH, Hong SB, Park S, Kang H, et al.Regional brain perfusion before and after treatment with methyl-phenidate may be associated with the G1287A polymorphism ofthe norepinephrine transporter gene in children with attention-deficit/hyperactivity disorder. Neurosci Lett. 2012;514(2):159–63.
41. Kim BN, Kim JW, Cummins TDR, BellgroveMA, Hawi Z, HongSB, et al. Norepinephrine genes predict response time variabilityand methylphenidate-induced changes in neuropsychologicalfunction in attention deficit hyperactivity disorder. J ClinPsychopharmacol. 2013;33(3):356–62.
42. Bobb AJ, Addington AM, Sidransky E, Gornick MC, Lerch JP,Greenstein DK, et al. Support for association between ADHD andtwo candidate genes: NET1 and DRD1. Am J Med Genet B.2005;134B(1):67–72.
43. Kollins SH, Anastopoulos AD, Lachiewicz AM, FitzGerald D,Morrissey-Kane E, Garrett ME, et al. SNPs in dopamine D2receptor gene (DRD2) and norepinephrine transporter gene(NET) are associated with continuous performance task (CPT)
497, Page 18 of 20 Curr Psychiatry Rep (2014) 16:497

phenotypes in ADHD children and their families. Am J MedGenet B. 2008;147B(8):1580–8.
44. Lesch KP, Selch S, Renner TJ, Jacob C, Nguyen TT, Hahn T, et al.Genome-wide copy number variation analysis in attention-deficit/hyperactivity disorder: association with neuropeptide Y genedosage in an extended pedigree. Mol Psychiatry. 2011;16(5):491–503.
45. Shiffrin ND, Gruber J, Glatt SJ, Faraone SV. No association betweenMspI allele of the ADRA2A polymorphism and ADHD: meta-analysis of family-based studies. Psychiatr Genet. 2013;23(4):174–5.
46. Froehlich TE, Epstein JN, Nick TG, Castro MSM, Stein MA,Brinkman WB, et al. Pharmacogenetic predictors of methylphe-nidate dose-response in attention-deficit/hyperactivity disorder. JAm Acad Child Adolesc Psychiatry. 2011;50(11):1129–39.
47. Yang L, Qian Q, Liu L, Li H, Faraone SV, Wang Y. Adrenergicneurotransmitter system transporter and receptor genes associatedwith atomoxetine response in attention-deficit hyperactivity disor-der children. J Neural Transm. 2013;120(7):1127–33.
48.•• Sun HJ, Yuan FF, Shen XM, Xiong GL, Wu J. Role of COMT inADHD: a systematic meta-analysis. Mol Neurobiol. 2014;49(1):251–61. This is a meta-analysis on findings investigating associationsbetweenCOMTgene andADHD.The results indicated no association.
49. Qian QJ, Yang L, Wang YF, Zhang HB, Guan LL, Chen Y, et al.Gene-gene interaction between COMT and MAOA potentiallypredicts the intelligence of attention-deficit hyperactivity disorderboys in China. Behav Genet. 2010;40(3):357–65.
50. Thapar A, Langley K, Fowler T, Rice F, Turic D, Whittinger N,et al. Catechol O-methyltransferase gene variant and birth weightpredict early-onset antisocial behavior in children with attention-deficit/hyperactivity disorder. Arch Gen Psychiatr. 2005;62(11):1275–8.
51. Caspi A, Langley K, Milne B, Moffitt TE, O’Donovan M, OwenMJ, et al. A replicated molecular genetic basis for subtypingantisocial behavior in children with attention-deficit/hyperactivitydisorder. Arch Gen Psychiatr. 2008;65(2):203–10.
52. Cheon KA, Jun JY, Cho DY. Association of the catechol-O-methyltransferase polymorphism with methylphenidate responsein a classroom setting in children with attention-deficit hyperac-tivity disorder. Int Clin Psychopharmacol. 2008;23(5):291–8.
53. Kereszturi E, Tarnok Z, Bognar E, Lakatos K, Farkas L, GadorosJ, et al. Catechol-O-methyltransferase Val158Met polymorphismis associated with methylphenidate response in ADHD children.Am J Med Genet B. 2008;147B(8):1431–5.
54. Manor I, Tyano S, Mel E, Eisenberg J, Bachner-Melman R, KotlerM, et al. Family-based and association studies of monoamineoxidase A and attention deficit hyperactivity disorder (ADHD):preferential transmission of the long promoter-region repeat andits association with impaired performance on a continuous perfor-mance test (TOVA). Mol Psychiatry. 2002;7(6):626–32.
55. Rommelse NNJ, Altink ME, Arias-Vasquez A, Buschgens CJM,Fliers E, Faraone SV, et al. Differential association betweenMAOA, ADHD and neuropsychological functioning in boys andgirls. Am J Med Genet B. 2008;147B(8):1524–30.
56. Nymberg C, Jia T, Lubbe S, Ruggeri B, Desrivieres S, Barker G,et al. Neural mechanisms of attention-deficit/hyperactivity disor-der symptoms are stratified by MAOA genotype. Biol Psychiatry.2013;74(8):607–14.
57. Kieling C, Genro JP, Hutz MH, Rohde LA. The -1021 C/T DBHpolymorphism is associated with neuropsychological performanceamong children and adolescents with ADHD. Am J Med Genet BNeuropsychiatr Genet. 2008;147B(4):485–90.
58. Shim SH, Hwangbo Y, Kwon YJ, Jeong HY, Lee BH, Lee HJ,et al. Increased levels of plasma brain-derived neurotrophic factor(BDNF) in children with attention deficit-hyperactivity disorder(ADHD). Prog Neuro-Psychopharmacol Biol Psychiatry.2008;32(8):1824–8.
59. KimH, HeoHI, KimDH, Ko IG, Lee SS, Kim SE, et al. Treadmillexercise and methylphenidate ameliorate symptoms of attentiondeficit/hyperactivity disorder through enhancing dopamine syn-thesis and brain-derived neurotrophic factor expression in sponta-neous hypertensive rats. Neurosci Lett. 2011;504(1):35–9.
60. Scassellati C, Zanardini R, Tiberti A, PezzaniM,Valenti V, EffedriP, et al. Serum brain-derived neurotrophic factor (BDNF) levels inattention deficit-hyperactivity disorder (ADHD). Eur ChildAdolesc Psichiatry. 2014;23(3):173–7.
61.•• Joseph N, Zhang-James Y, Perl A, Faraone SV. Oxidative Stressand ADHD: A Meta-Analysis. J Atten Disord. 2013;20(10):1–10doi:10.1177/1087054713510354. The authors performed a meta-analysis of studies on oxidative stress and antioxidant status inmedication naive patients with ADHD and controls. The resultssuggest that patients with ADHD have normal levels of antioxi-dant production, but that their response to oxidative stress isinsufficient, leading to oxidative damage.
62.•• Arns M, Conners CK, Kraemer HC. A decade of EEG theta/betaratio research in ADHD: ameta-analysis. J AttenDisord. 2013;17(5):374–83. This is a meta-analysis on the TBR in ADHD. The resultsindicated that excessive TBR cannot be considered a reliable diag-nosticmeasure of ADHD, however a substantial sub-group of ADHDpatients do deviate on this measure and TBR has prognostic value inthis sub-group, warranting its use as a prognostic measure ratherthan a diagnostic measure.
63. Yener GG, Basar E. Brain oscillations as biomarkers in neuropsy-chiatric disorders: following an interactive panel discussion andsynopsis. Suppl Clin Neurophysiol. 2013;62:343–63.
64. Frazier TW, Demaree HA, Youngstrom EA. Meta-analysis of intel-lectual and neuropsychological test performance in attention-deficit/hyperactivity disorder. Neuropsychology. 2004;18(3):543–55.
65. Martinussen R, Hayden J, Hogg-Johnson S, Tannock R. A meta-analysis of working memory impairments in children with atten-tion-deficit/hyperactivity disorder. J Am Acad Child AdolescPsychiatry. 2005;44(4):377–84.
66. Willcutt EG, Doyle AE, Nigg JT, Faraone SV, Pennington BF.Validity of the executive function theory of attention-deficit/hy-peractivity disorder: a meta-analytic review. Biol Psychiatry.2005;57(11):1336–46.
67. Kasper LJ, Alderson RM, Hudec KL. Moderators of workingmemory deficits in children with attention-deficit/hyperactivitydisorder (ADHD): A meta-analytic review. Clin Psychol Rev.2012;32(7):605–17.
68. van Ewijk H, Heslenfeld DJ, Luman M, Rommelse NN, HartmanCA, Hoekstra P, et al. Visuospatial working memory in ADHDpatients, unaffected siblings, and healthy controls. J Atten Disord.2014;18(4):369–78.
69. Cortese S. The neurobiology and genetics of attention-deficit/hyperactivity disorder (ADHD): what every clinician shouldknow. Eur J Paediatr Neurol. 2012;16(5):422–33.
70. Friedel S, Saar K, Sauer S, Dempfle A, Walitza S, Renner T, et al.Association and linkage of allelic variants of the dopamine trans-porter gene in ADHD. Mol Psychiatry. 2007;12(10):923–33.
71. Cheon KA, RyuYH, Kim JW, ChoDY. The homozygosity for 10-repeat allele at dopamine transporter gene and dopamine trans-porter density in Korean children with attention deficit hyperac-tivity disorder: relating to treatment response to methylphenidate.Eur Neuropsychopharmacol. 2005;15(1):95–101.
72. Franke B, Faraone SV, Asherson P, Buitelaar J, Bau CHD, Ramos-Quiroga JA, et al. The genetics of attention deficit/hyperactivitydisorder in adults, a review. Mol Psychiatry. 2012;17(10):960–87.
73. Bellgrove MA, Hawi Z, Kirley A, Gill M, Robertson IH.Dissecting the attention deficit hyperactivity disorder (ADHD)phenotype: sustained attention, response variability and spatialattentional asymmetries in relation to dopamine transporter(DAT1) genotype. Neuropsychologia. 2005;43(13):1847–57.
Curr Psychiatry Rep (2014) 16:497 Page 19 of 20, 497

74. Bellgrove MA, Barry E, Johnson KA, Cox M, Daibhis A,Daly M, et al. Spatial attentional bias as a marker of geneticrisk, symptom severity, and stimulant response in ADHD.Neuropsychopharmacology. 2008;33(10):2536–45.
75. Faraone SV, Spencer TJ, Madras BK, Zhang-James Y, BiedermanJ. Functional effects of dopamine transporter gene genotypes onin vivo dopamine transporter functioning: a meta-analysis. MolPsychiatry. 2013;19:880–889.
76. Noaín D, Avale ME, Wedemeyer C, Calvo D, Peper M,Rubinstein M. Identification of brain neurons expressing thedopamine D4 receptor gene using BAC transgenic mice. Eur JNeurosci. 2006;24(9):2429–38.
77. Usiello A, Baik JH, Rouge-Pont F, Picetti R, Dierich A, LeMeurM, et al. Distinct functions of the two isoforms of dopamine D2receptors. Nature. 2000;408(6809):199–203.
78. Beischlag TV, Marchese A, Meador-Woodruff JH, Damask SP,O’Dowd BF, Tyndale RF, et al. The human dopamine D5 receptorgene: cloning and characterization of the 5′-flanking and promoterregion. Biochemistry. 1995;34(17):5960–70.
79. Li S, Cullen WK, Anwyl R, Rowan MJ. Dopamine-dependentfacilitation of LTP induction in hippocampal CA1 by exposure tospatial novelty. Nat Neurosci. 2003;6(5):526–31.
80. Stahl SM. Neurotransmission of cognition, part 3. Mechanism ofaction of selective NRIs: both dopamine and norepinephrine in-crease in prefrontal cortex. J Clin Psychiatry. 2003;64(3):230–1.
81. Mick E, Neale B, Middleton FA, McGough JJ, Faraone SV.Genome-wide association study of response to methylphenidatein 187 children with attention-deficit/hyperactivity disorder. Am JMed Genet B. 2008;147B(8):1412–8.
82. Oades RD, Daniels R, Rascher W. Plasma neuropeptide-Y levels,monoaminemetabolism, electrolyte excretion and drinking behav-ior in children with attention-deficit hyperactivity disorder.Psychiatry Res. 1998;80(2):177–86.
83. Waldman ID, Nigg JT, Gizer IR, Park L, Rappley MD, Friderici K.The adrenergic receptor alpha-2A gene (ADRA2A) and neuropsy-chological executive functions as putative endophenotypes forchildhood ADHD. CognAffect Behav Neurosci. 2006;6(1):18–30.
84. Hong J, Shu-Leong H, Tao X, Lap-Ping Y. Distribution ofcatechol-O-methyltransferase expression in human central ner-vous system. Neuroreport. 1998;9(12):2861–4.
85. Brunner HG, Nelen M, Breakefield XO, Ropers HH, Vanoost BA.Abnormal-behavior associated with a point mutation in the structuralgene for monoamine oxidase-A. Science. 1993;262(5133):578–80.
86. Cases O, Lebrand C, Giros B, Vitalis T, De Maeyer E, Caron MG,et al. Plasma membrane transporters of serotonin, dopamine, andnorepinephrine mediate serotonin accumulation in atypical loca-tions in the developing brain ofmonoamine oxidase A knock-outs.J Neurosci. 1998;18(17):6914–27.
87. Deckert J, Catalano M, Syagailo YV, Bosi M, Okladnova O, DiBella D, et al. Excess of high activity monoamine oxidase A genepromoter alleles in female patients with panic disorder. Hum MolGenet. 1999;8(4):621–4.
88. Gaspar P, Berger B, Febvret A, Vigny A, Henry JP. Catecholamineinnervation of the human cerebral cortex as revealed by compar-ative immunohistochemistry of tyrosine hydroxylase and dopa-mine-beta-hydroxylase. J Comp Neurol. 1989;279(2):249–71.
89. Rogeness GA, Maas JW, Javors MA, Macedo CA, Fischer C,Harris WR. Attention deficit disorder symptoms and urine cate-cholamines. Psychiatry Res. 1989;27(3):241–51.
90.•• Caylak E. Biochemical and genetic analyses of childhood attentiondeficit/hyperactivity disorder. Am J Med Genet B. 2012;159B(6):613–27. This is an review on biochemistry and genetics of ADHD.The authors reported many published studies about biochemicalabnormalities in ADHD as well as many published genetic studiesimplicating several genes (dopaminergic: DRD4, DAT1, DRD5,COMT; noradrenergic: DBH, ADRA2A; serotonergic: 5-HTT,
HTR1B,HTR2A; cholinergic: CHRNA4, and central nervous systemdevelopment pathway: SNAP25, BDNF) in the etiology of ADHD.
91. Mattson MP. Glutamate and neurotrophic factors in neuronalplasticity and disease. Ann N YAcad Sci. 2008;1144:97–112.
92. Söllner T, Whiteheart SW, Brunner M, Erdjument-Bromage H,Geromanos S, Tempst P, et al. SNAP receptors implicated invesicle targeting and fusion. Nature. 1993;362(6418):318–24.
93. Ghanizadeh A, Berk M. Zinc for treating of children and adoles-cents with attention-deficit hyperactivity disorder: a systematicreview of randomized controlled clinical trials. Eur J Clin Nutr.2013;67(1):122–4.
94. Bloch MH, Qawasmi A. Omega-3 fatty acid supplementation forthe treatment of children with attention-deficit/hyperactivity dis-order symptomatology: systematic review and meta-analysis. JAm Acad Child Adolesc Psychiatry. 2011;50(10):991–1000.
95. Chovanova Z, Muchova J, SivonovaM, DvorakovaM, ZitnanovaI, Waczulikova I, et al. Effect of polyphenolic extract, Pycnogenol(R), on the level of 8-oxoguanine in children suffering fromattention deficit/hyperactivity disorder. Free Radic Res.2006;40(9):1003–10.
96. Dvorakova M, Jezova D, Blazicek P, Trebaticka J, Skodacek I,Suba J, et al. Urinary catecholamines in children with attentiondeficit hyperactivity disorder (ADHD): Modulation by a polyphe-nolic extract from pine bark (Pycnogenol((R))). Nutr Neurosci.2007;10(3–4):151–7.
97. Garcia RJ, Francis L, Dawood M, Lai ZW, Faraone SV, Perl A.Attention deficit and hyperactivity disorder scores Are elevated andrespond to N-acetylcysteine treatment in patients with systemiclupus erythematosus. Arthritis Rheum US. 2013;65(5):1313–8.
98. Snyder SM, Hall JR. A meta-analysis of quantitative EEG powerassociated with attention-deficit hyperactivity disorder. J ClinNeurophysiol. 2006;23(5):440–55.
99. Tye C, Rijsdijk F, McLoughlin G. Genetic overlap betweenADHD symptoms and EEG theta power. Brain Cogn. 2014;87:168–72.
100.•• Kofler MJ, Rapport MD, Sarver DE, Raiker JS, Orban SA,Friedman LM, et al. Reaction time variability in ADHD: a meta-analytic review of 319 studies. Clin Psychol Rev. 2013;33(6):795–811. This is a meta-analysis of 319 studies of RT variabilityin children, adolescents, and adults with ADHD relative to typi-cally developing (TD) groups, clinical control groups, and them-selves. The results of the meta-analysis indicate that RT variabilityreflects a stable feature of ADHD and other clinical disorders thatis robust to systematic differences across studies.
101.•• Lange KW, Hauser J, Lange KM, Makulska-Gertruda E, Takano T,Takeuchi Yet al. Utility of cognitive neuropsychological assessmentin attention-deficit/hyperactivity disorder. Atten Defic HyperactDisord. 2014 Mar 18. doi:10.1007/s12402-014-0132-3. This is aninteresting review on the utility of neuropsychological tests inADHD. The authors concluded that cognitive neuropsychologicalassessment appears to be at present of limited clinical use andconfined to individual descriptions.
102.•• Kasparek T, Theiner P, Filova A. Neurobiology of ADHD fromchildhood to adulthood: findings of imaging methods. J AttenDisord. 2013;20(10):1–13. doi:10.1177/1087054713505322. Theauthors concluded that changes in neuronal plasticity may be behindpersisting brain changes in ADHD. Current treatment approachesseem to improve these neuroplastic processes, and, therefore, mayhave a positive effect on the neuropathology of ADHD.
103. Bellgrove MA, Mattingley JB, Hawi Z, Mullins C, Kirley A, GillM, et al. Impaired temporal resolution of visual attention anddopamine beta hydroxylase genotype in attention-deficit/hyperac-tivity disorder. Biol Psychiatry. 2006;60(10):1039–45.
104. Wasserman T, Wasserman LD. The sensitivity and specificity ofneuropsychological tests in the diagnosis of attention deficit hy-peractivity disorder. Appl Neuropsychol Child. 2012;1(2):90–9.
497, Page 20 of 20 Curr Psychiatry Rep (2014) 16:497





























