Embed Size (px)
Citation preview

ARTICLE IN PRESS
0142-9612/$ - se
doi:10.1016/j.bi
�CorrespondE-mail addr
Biomaterials 27 (2006) 1851–1858
www.elsevier.com/locate/biomaterials
Review
Bioluminescent imaging: Emerging technology for non-invasive imagingof bone tissue engineering
Jan de Boera,�, Clemens van Blitterswijka, Clemens Lowikb
aInstitute for Biomedical Technology, University of Twente, Prof. Bronkhorstlaan 10D, 3723 MB, Bilthoven, The NetherlandsbDepartment of Endocrinology, Leiden University Medical Centre, Leiden, The Netherlands
Received 27 April 2005; accepted 26 September 2005
Available online 20 October 2005
Abstract
Bone tissue engineering is a multidisciplinary research area in which new strategies are developed to treat patients with large bone
defects as occurring during e.g. hip revisions, upon trauma or in spinal fusions. In vivo evaluation of bone formation in animal models is
highly relevant for graft evaluation but is time-consuming, invasive and difficult to quantify. As a consequence, most in vivo studies
ignore the dynamic nature of bone regeneration and the molecular processes underlying it. In vivo bioluminescent imaging (BLI) is a
relatively young research field with great potential that may overcome these problems. BLI encompasses non-invasive imaging of
luciferase gene activity using cooled charge coupled device cameras in luciferase transgenic animals or in grafted, luciferase transgenic
cells. The imaging procedure is technically simple and quantifiable. Because luciferase expression can be put under the control of tissue-
specific regulatory elements, BLI allows non-invasive imaging of processes highly relevant to bone tissue engineering like differentiation,
apoptosis, vasculogenesis and inflammation. In this review, we describe the basic principle of BLI and discuss transgenic animals and
constructs currently available for application in bone tissue engineering. Furthermore, we reflect on technical developments that will
make BLI even more promising for future application in bone tissue engineering research.
r 2005 Elsevier Ltd. All rights reserved.
Keywords: Bone tissue engineering; Molecular imaging
Contents
1. Bone tissue engineering: a technological meltingpot . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1851
2. Clogging the pipeline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1852
3. Bioluminescent imaging (BLI) in rodents: an emerging technology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1852
4. Imaging of biological processes relevant to bone regeneration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1854
4.1. Cell numbers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1854
4.2. Osteogenic differentiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1854
4.3. Inflammation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1855
4.4. Apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1856
4.5. Vasculogenesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1856
4.6. Signal transduction. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1856
5. Current limitations and future directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1857
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1857
e front matter r 2005 Elsevier Ltd. All rights reserved.
omaterials.2005.09.034
ing author. Tel.: +3130 2295154; fax: +31 30 2280225.
ess: [email protected] (J. de Boer).
1. Bone tissue engineering: a technological meltingpot
Bone tissue engineering is a relatively young, rapidlyevolving and innovative research field, which combines a

ARTICLE IN PRESSJ. de Boer et al. / Biomaterials 27 (2006) 1851–18581852
multitude of scientific disciplines such as materials science,stem cell biology and molecular biology in order toengineer improved grafts for the treatment of large bonedefects. Cross-disciplinarity is the rule rather than theexception. Bone tissue engineers are increasingly aware thatmaterials traditionally used for bone regeneration, such asceramics, titanium and polymers are not merely there todeliver and support grafted cells or provide mechanicalstrength. They can play a direct role in modulating tissueformation by interacting with endogenous or implantedcells, as vehicle for controlled release of bioactivecompounds and by creating the proper microenvironmentfor tissue growth [1–5]. Recent years have seen a surge inknowledge of the molecular pathways controlling bonegrowth and regeneration. Every step in the fascinatingcellular cascade leading from a multipotent mesenchymalstem cell to a mineralising osteoblast is controlled bydifferent molecular cues, i.e. growth factors, extracellularmatrix proteins, cytokines and hormones. Translation ofmolecular knowledge into improved bone grafts is anexciting new challenge in bone tissue engineering research[6]. A classical ‘‘from-bench-to-bed’’ example is theidentification and clinical application of bone morphoge-netic proteins [7,8]. Other examples of applied molecularknowledge include incorporation into or coating ofbiomaterials with bioactive groups such as chitosan [9]and RGD peptides [10] for enhanced cell binding, andcontrolled release of bone-inducing growth factors frommaterials [11–13]. Besides their direct application in boneregeneration technology, genes involved in osteogenesis arealso useful molecular markers for analysis of the variousstages of bone formation.
2. Clogging the pipeline
Newly designed bone grafts have to be tested for theirbiological performance. To assess applicability, each newgraft should be tested in a system as close as possible to theclinical reality, but for practical reasons the test systemshould allow rapid screening of many variables at low costand in a short period of time. Evidently, these viewsconflict and in practice, product development starts with invitro biological assays, to proceed with in vivo studies inrodent models, then larger animal models and finallyclinical trials in human beings. The great advantage of invitro biological assays is that they allow a degree ofautomation and are therefore suitable for medium to highthroughput biomaterials research [14], gene discovery[15,16] and cellular assays [17]. However, in vitro assaysare mostly limited to cellular research because it is difficultto mimic the complexity of tissue formation, which is a 3-dimensional (3D) interplay between different cell types, in atest tube. In vivo bone formation by stem cells is mostlystudied by seeding the cells onto biomaterials andsubsequent ectopic implantation in the back of immune-deficient mice. Ectopic implantation is a simple surgicalhandling and immune-deficient mice allow implantation of
xenogenic cells and materials. In this milieu, mesenchymalstem cells produce bone matrix, blood vessels will infiltratethe tissue and bone marrow formation is supported, whichallows investigation of a number of highly relevant aspectsof bone tissue engineering. A clinically even more relevantimplantation site is bone itself but orthotopic implanta-tions are technically more difficult to perform and moreexpensive because one has to rely on larger animal modelssuch as rats, goats or rabbits. Although biologically morerelevant, orthotopic and ectopic bone formation modelslack the versatility of in vitro assays due to the limitedbiological readout and time-consuming analysis. Whereasin vitro osteogenic differentiation can be easily followedthrough time by quantitative PCR, flow cytometry orELISA, in vivo analysis of bone formation mostly dependson histological examination at single time points andtedious in situ hybridisation or immunohistochemistry.Evaluation at different time intervals surmounts toexcessive animal usage and time-consuming histologicalanalysis. Due to these limitations, many in vivo studiesignore the dynamic molecular and cellular processesleading to repair of bone defects. Bioluminescent imagingprovides a technological platform that allows non-invasiveimaging of marker gene expression. Thus, all aspects ofbone formation can be followed in individual animalsfrom the time of implantation until the moment whennewly formed bone is remodelled and fully integrated (seeTable 1).
3. Bioluminescent imaging (BLI) in rodents: an emerging
technology
The recent surge in BLI research is spurred by thedemand of cancer biologists to image tumour growth andmaturation and the efficacy of therapy in vivo (for reviewson BLI and molecular imaging see [18–20]. Biolumines-cence refers to the enzymatic generation of visible light byliving organisms. The most commonly used bioluminescentreporter gene for research purposes is luciferase from theNorth American firefly Photinus pyralis. The enzymecatalyses the transformation of its substrate D-luciferininto oxoluciferin, in an oxygen and ATP-dependentprocess, leading to emission of photons (see Fig. 1).Biological sources of light have sufficient intensity to crossanimal tissues, including bone, provided that the light has awavelength of more than 550 nm. The firefly luciferase/luciferin reaction emits photons at wavelengths from 500 to620 nm and is therefore suitable for external detection.Apart from fulfilling this criterion, another importantfeature is that the substrate luciferin diffuses within fewminutes from intravenous or peritoneal injection through-out the entire body of small animals and is rapidly taken upby cells [21]. The quantitative relationship between geneexpression and the detected signal is influenced by severalfactors. First of all, the luciferin substrate must be availablewithin the cells, which is influenced by factors such astechnical skills of the experimentor to inject the substrate

ARTICLE IN PRESS
Table 1
Transgenic reporters currently available for BLI of bone tissue engineering
Name Regulatory element Application Reference
Transgenic mice
hOC-luc Human osteocalcin promoter Matrix deposition [31]
colI(a1)-luc Bone-specific enhancer of the mouse collagen (a1) I gene Matrix deposition [33]
BSP-luc Mouse BSP promoter Matrix deposition [36]
VEGFR2-luc Mouse VEGFR2 gene promoter Angiogenesis [46]
iNOS-luc Mouse iNOS gene promoter Inflammation [40]
NFkB-luc 3 NFkB site from the Igk light chain promoter Inflammation [39]
ER-DEVD-luc Caspase cleavage sequence Apoptosis [41]
Transgenic constructs
Retro-luc Cytomegalovirus promoter Cell tracking [28]
BRE-luc BMP responsive element of the ID1 gene Osteo-induction, BMP CDD [30]
WRE-luc TCF/LEF binding sites Wnt signalling [54]
Hes1-luc Hes1 gene promoter Differentiation [52]
Fech-luc Ferrochelatase promoter Hypoxia [48]
HRE-luc 5 hypoxia responsive elements from the VEGF gene Hypoxia [49]
Fig. 1. Biological principle of bioluminescent imaging of luciferase expression. (A) Firefly luciferase converts the substrate luciferin into oxyluciferin and
light. Tissue-specific transcription factors bind to regulatory sequences cloned directly adjacent to the luciferase gene, resulting in tissue specific expression
of luciferase, which can be visualised by injection of luciferin and detection of the emitted light by CCCD cameras. (B) Image of a transgenic mouse during
pro-estrous expressing the luciferase gene under the control of an estrogen responsive element. Note the intense staining of the ovaries, which subsides
during estrous.
J. de Boer et al. / Biomaterials 27 (2006) 1851–1858 1853
but also by the degree of vascularisation of the targettissue. For example, we recently used BLI as a tool tomeasure enhanced luciferin perfusion in a vascularisedtissue construct [22]. Furthermore, light scattering occursas it travels through the body. As such, it is difficult tocompare the relative light intensities emitted from differentorgans because light scattering depends on the character-
istics of the tissue. Recent technical advances for imagingweak visible light sources using cooled charged coupleddevice (CCCD) cameras, peltier cooled detectors andmicro-plate channel intensifiers allow detection and quan-tification of bioluminescent emission from inside the tissuesof the animals. This technical development made it possibleto monitor gene expression in living animals via a luciferase

ARTICLE IN PRESS
Fig. 2. BLI of transgenic cancer cells. 1� 105 luciferase transgenic MD-
231 cells were injected in the left hart ventricle and bone metastases were
imaged using BLI. Photon intensity was indicated using pseudo-colouring.
Note the metastasis throughout the body of the mouse.
J. de Boer et al. / Biomaterials 27 (2006) 1851–18581854
reporter gene linked to specific promoters [21,23] (Fig. 1).For instance, we generated mice transgenic for theluciferase gene under the control of an estrogen-responsiveelement (ERE-luc) [24]. Luciferase activity in individualmice could be followed throughout the estrous cycle and apeak of luciferase activity was observed during proesterusin reproductive tissues (Fig. 1). Using ERE-luc mice, wewere able to monitor estrogen receptor activity dynamics ina tissue-specific and quantitative way, demonstrating thepower of BLI. Besides imaging of transgene expression intransgenic animals, BLI can also be used to follow in realtime the fate of luciferase transfected cells injected in livinganimals, an application which is widely used in cancerresearch [25]. One of the many advantages of BLI is that itis non-invasive, and therefore allows investigations in thesame animal at different time points. Accordingly, thenumber of animal groups in the experimental setting isgreatly reduced and the data variation is reduced becausethe signal is sequentially detected in the same animal.
Detection of the bioluminescent activity in vivo of theluciferase enzyme as a transcriptional reporter facilitatesrapid and repetitive screening for both the presence andfunction of transgenes in intact living mice [26]. Below, wewill discuss recent developments in the field of BLI thathave brought the technique within the reach of bone tissueengineering research.
4. Imaging of biological processes relevant to bone
regeneration
4.1. Cell numbers
Bone tissue engineering often includes seeding ofosteogenic cells onto biomaterials and co-implantation inan ectopic or orthotopic site. Survival of the cells relies onboth cell and biomaterial properties and it would be veryvaluable to assess survival and proliferation of implantedcells for a prolonged period of time in the same animal. Forthis purpose, transgenic cells can be prepared expressingthe luciferase gene under the control of a constitutivelyactive promoter. Using viral transduction, virtually everycell type used in bone tissue engineering, including MSCs,can be stably transduced with a luciferase gene and thusfollowed in real time upon implantation. Luciferasetransgenic cells have been extensively used in cancerresearch, for instance to monitor the efficacy of drugtreatment on the survival of implanted cancer cells or toanalyse tumour metastasis. For instance, we implantedluciferase transgenic mammary carcinoma cells into the leftcardiac ventricle of mice and during a 36 days follow-upstudy we were able to trace bone marrow progression andmetastasis of �0.5mm3 (see Fig. 2), which is below thelimit at which tumours need angiogenesis for furthergrowth and well before radiological osteolysis can beobserved [27]. One report demonstrates the proof ofconcept for the use of luminescent cells in the field oftissue engineering. Blum et al. [28] used a stably transfected
fibroblast cell line expressing a luciferase reporter constructunder the control of a constitutively active viral promoter.The transgenic cells were seeded on a titanium mesh usedfor bone tissue engineering purposes and implantedsubcutaneously in nude mice. Using BLI, Blum et al. wereable to monitor cell survival over a period of 28 days.
4.2. Osteogenic differentiation
The tissue- and time-specific expression of genes thatdelineate the process of osteogenic differentiation can bemonitored by driving the expression of luciferase byspecific regulatory elements. For instance, Korchynski etal. [29] identified a DNA sequence in the promoter of theID1 gene, designated the bone morphogenetic proteinresponsive element (BRE), which responds uniquely toBMP stimulation. Monteiro et al. [30] used the BRE todrive lacZ in transgenic mice and luciferase in embryonicstem cells. They were able to detect lacZ expression inmany tissues in which BMP is known to be active and theyshowed a BMP dose–responsive upregulation of luciferaseactivity in ES cells. Because activation of BMP signallingmarks the onset of osteogenic differentiation, BRE-luciferase mice and cells can in the future be used to assaythe earliest events of osteogenic differentiation.A later phase of differentiation, i.e. osteoblast matrix
production, can be monitored by luciferase under thecontrol of the osteocalcin promoter. Using mice transgenicfor the human osteocalcin promoter driving the luciferasegene [31], Iris et al. [32] observed luciferase expression inliving mice upon bone formation induced by fracturehealing or bone marrow ablation. Interestingly, luciferaseexpression was highly induced upon injection of vitaminD3, a compound known to directly activate the osteocalcingene (Fig. 3). Similarly, mice transgenic for the bone-specific regulatory element of the collagen type I gene (Col(a1) I-luc mice) were treated with parathyroid hormone orprostaglandin E2, which are known bone anabolic agents[33]. In both cases, enhanced luciferase expression wasobserved in tissue extracts demonstrating that Col (a1)
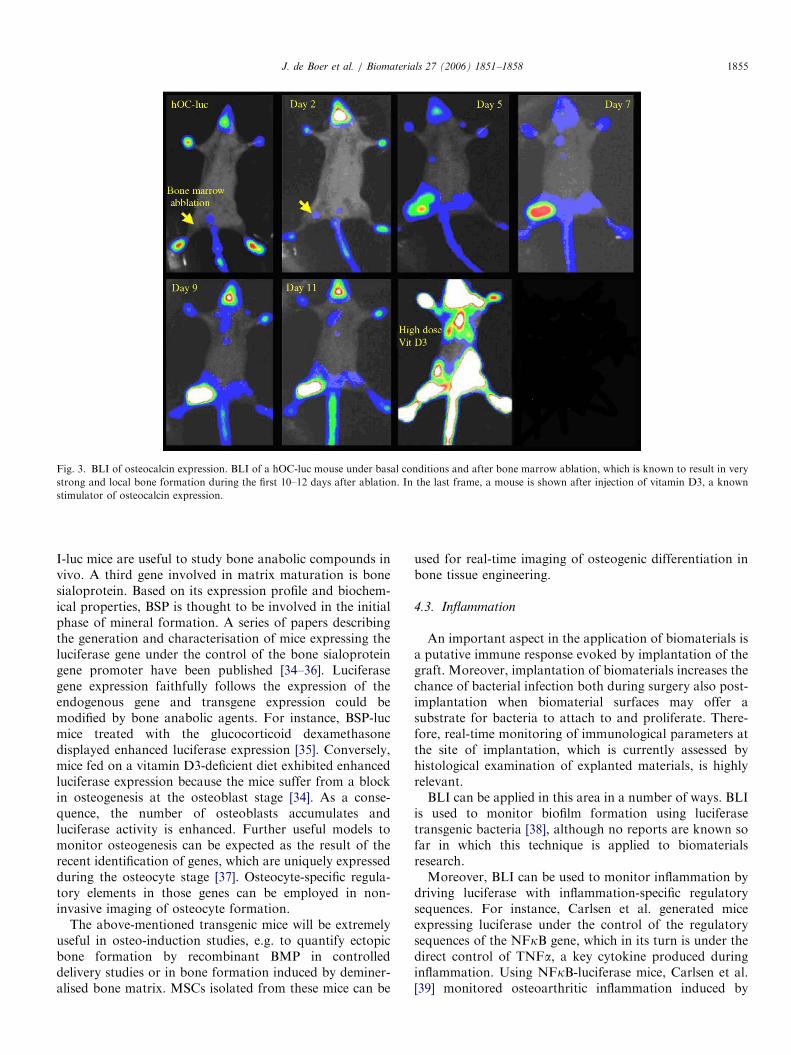
ARTICLE IN PRESS
Fig. 3. BLI of osteocalcin expression. BLI of a hOC-luc mouse under basal conditions and after bone marrow ablation, which is known to result in very
strong and local bone formation during the first 10–12 days after ablation. In the last frame, a mouse is shown after injection of vitamin D3, a known
stimulator of osteocalcin expression.
J. de Boer et al. / Biomaterials 27 (2006) 1851–1858 1855
I-luc mice are useful to study bone anabolic compounds invivo. A third gene involved in matrix maturation is bonesialoprotein. Based on its expression profile and biochem-ical properties, BSP is thought to be involved in the initialphase of mineral formation. A series of papers describingthe generation and characterisation of mice expressing theluciferase gene under the control of the bone sialoproteingene promoter have been published [34–36]. Luciferasegene expression faithfully follows the expression of theendogenous gene and transgene expression could bemodified by bone anabolic agents. For instance, BSP-lucmice treated with the glucocorticoid dexamethasonedisplayed enhanced luciferase expression [35]. Conversely,mice fed on a vitamin D3-deficient diet exhibited enhancedluciferase expression because the mice suffer from a blockin osteogenesis at the osteoblast stage [34]. As a conse-quence, the number of osteoblasts accumulates andluciferase activity is enhanced. Further useful models tomonitor osteogenesis can be expected as the result of therecent identification of genes, which are uniquely expressedduring the osteocyte stage [37]. Osteocyte-specific regula-tory elements in those genes can be employed in non-invasive imaging of osteocyte formation.
The above-mentioned transgenic mice will be extremelyuseful in osteo-induction studies, e.g. to quantify ectopicbone formation by recombinant BMP in controlleddelivery studies or in bone formation induced by deminer-alised bone matrix. MSCs isolated from these mice can be
used for real-time imaging of osteogenic differentiation inbone tissue engineering.
4.3. Inflammation
An important aspect in the application of biomaterials isa putative immune response evoked by implantation of thegraft. Moreover, implantation of biomaterials increases thechance of bacterial infection both during surgery also post-implantation when biomaterial surfaces may offer asubstrate for bacteria to attach to and proliferate. There-fore, real-time monitoring of immunological parameters atthe site of implantation, which is currently assessed byhistological examination of explanted materials, is highlyrelevant.BLI can be applied in this area in a number of ways. BLI
is used to monitor biofilm formation using luciferasetransgenic bacteria [38], although no reports are known sofar in which this technique is applied to biomaterialsresearch.Moreover, BLI can be used to monitor inflammation by
driving luciferase with inflammation-specific regulatorysequences. For instance, Carlsen et al. generated miceexpressing luciferase under the control of the regulatorysequences of the NFkB gene, which in its turn is under thedirect control of TNFa, a key cytokine produced duringinflammation. Using NFkB-luciferase mice, Carlsen et al.[39] monitored osteoarthritic inflammation induced by

ARTICLE IN PRESSJ. de Boer et al. / Biomaterials 27 (2006) 1851–18581856
injection of bacterial lipopolysacharide, and quantified thetherapeutic potential of dexamethasone treatment of thearthritic lesion. Using a similar approach, iNOS-luciferasemice were used to image zymosan-induced arthritis in theknee joint [40]. Like NFkB, iNOS is induced duringinflammation and therefore both mouse models might beused for biocompatibility studies of newly designedmaterials.
4.4. Apoptosis
An unwanted side effect of implanted materials iscytotoxicity to the cells surrounding the implant. Apopto-sis, i.e. programmed cell death, can be used as a marker forcytotoxicity.
Laxman et al. developed an elegant tool to monitorapoptosis based on the activity of caspases, which aresequence-specific proteases specifically activated uponinitiation of apoptosis. They generated a recombinant geneencoding a chimeric protein of luciferase fused to a domainof the estrogen receptor (ER). The ER domain efficientlysilences luciferase reporter activity [41] but the inactivechimeric protein can be activated upon proteolytic cleavageof a caspase recognition site separating luciferase and thesilencing domain. Laxman et al. [41] describe how theywere able to monitor apoptosis in tumourigenic cellsimplanted in nude mice and similar studies could beperformed with implanted materials. The paper furtherunderscores the versatility of this approach. Using thesame methodology, one could monitor the activity of anysequence-specific protease such as e.g. metalloproteasesinvolved in arthritis, simply by changing the caspaserecognition sequence with other protease sensitive se-quences. Another apoptosis model makes use of the factthat the p53 protein accumulates upon apoptosis andactivates gene expression through p53-responsive elements.The specificity of these regulatory sequences has beendemonstrated in vivo by PET imaging of DNA damage-induced apoptosis [42] and in a bioluminescent xenograftmodel of human colon carcinoma cells [43].
4.5. Vasculogenesis
Bone is a well-vascularised tissue and vasculogenesisduring bone regeneration serves two purposes. First, itprovides nutrition to the implanted cells and adjacenttissue and, secondly, it presents the molecular cues requiredfor the final step of endochondral bone formation: invasionby MSCs of the fracture area and replacement ofhypertrophic cartilage tissue by bone. A number oftechniques exist that monitor vasculogenesis indirectly byimaging of perfusion of the implant by an injected tracer,i.e. using MRI [44] or CT [45]. Direct BLI of vasculogenesisis feasible using endothelial specific promoters drivingluciferase expression. For instance, Zhang et al. [46]describe vascular endothelial growth factor receptor 2(VEGFR2)-luciferase mice. In a cutaneous wound healing
model, VEGFR2-luc expression was monitored in thewound tissue. Dexamethasone suppressed luciferase activ-ity, concomitant with delayed healing and impairedangiogenesis. Alternatively, endothelial specificity can beprovided by the Tie2 promoter. Using Tie2-driven GFPexpression in whole mounts of mouse retina, Otani et al.[47] could clearly delineate the eye vasculature. In thefuture, Tie2-luciferase and VEGR2-luciferase mice mightbe employed to monitor blood vessel ingrowth in bonegrafts.In close relation to vasculogenis, the nutritional status of
implanted cells might be assessed by BLI of hypoxia, amajor problem in tissue engineering of large bone grafts.Both the ferrochelatase and VEGF gene promoter respondto hypoxia-inducible factor and are clearly regulated byhypoxic conditions [48,49]. In one report, a hypoxia-responsive element was used to drive expression ofluciferase and enhanced luciferase activity was seen inischemic tissue [50].
4.6. Signal transduction
As discussed before, bone tissue engineering increasinglyincorporates molecular cues into the graft, i.e. growthfactors, small molecules and adhesion proteins to promotetissue regeneration. BLI can be used to monitor thebiological activity of these molecules in vivo because anumber of DNA sequences are known which activate genetranscription as a direct consequence of the compound’sbiological activity. Examples include the BMP-responsiveelement identified in the ID-1 gene; the TCF/LEF-bindingsites responding to Wnt signaling [51]; the promoter of theHes1 gene, which is a direct target gene of Notch signalling[52]; and the responsive elements to either the estrogen,glucocorticoid or retinoic acid receptors [19 and referencestherein]. For example, the previously discussed BRE-luciferase construct can be employed to trace the biologicalactivity of BMP in demineralised bone matrix or recombi-nant BMP. A number of reports describe incorporation ofBMP into materials for local release but, so far, the in vivorelease profile and biological activity are black boxes andefficacy can only be scored in post-mortem evaluation ofbone formation. Using BRE-luciferase transgenic mice orcells one could on-line monitor the biological efficacy ofreleased BMPs. Similarly, the Wnt signalling cascade playsan important role in bone biology and strategies aimedat enhancing Wnt signalling during bone regenerationare anticipated. We recently discovered that humanmesenchymal stem cell proliferation is enhanced by theWnt-activating drug lithium [53] and, to this end, weincorporated lithium into calcium phosphate coatings(unpublished data). Wnt activates target genes throughthe well-characterised TCF/LEF-binding sites (WRE) andnumerous papers describe the use of WRE-luciferaseconstructs to monitor Wnt activity in vitro [54,55]. Tomonitor the biological activity of our coatings in vivo, weare currently generating WRE-luciferase transgenic mice.

ARTICLE IN PRESSJ. de Boer et al. / Biomaterials 27 (2006) 1851–1858 1857
5. Current limitations and future directions
The current BLI systems use photographic principles tocapture light emitted from the animals at sites whereluciferase is expressed. Photons are detected at the surfaceof the animal in 2D, also known as planar imaging, thesimplest technique for detecting optical reporter moleculesin vivo. Photon attenuation, however, is strongly nonlinearas a function of depth and of the optical heterogeneity oftissue, which hampers signal quantification. Planar imagingis further complicated by the inability to resolve depth andby tissue scattering and absorption, which limits spatialresolution. A way to circumvent the problems of 2D planarBLI would be a combination of multi-view angle imaging(bioluminescence tomography) with a priori informationon tissue heterogeneity. This can now be achieved using thenewly developed 3D BLI systems from Xenogen andBerthold Technologies. If these 3D images are combinedwith 3D spatial (tissue) information obtained by CT orMRI, algorithms can be developed to more preciselycorrect for photon scattering and absorption due to tissueheterogeneity and depth.
Besides advances in the optimisation in imaging equip-ment and software, recent years have also seen a steadyincrease in the number of transgenic reporter constructs,which express luciferase under the control of specificregulatory elements and the generation of transgenic cellsand animals from them. The emergence of BLI in rodentslikely spurs the generation of many more and they will beapplicable in three ways. First, reporter constructs can beintroduced into any cell type used in bone tissue engineer-ing by means of e.g. viral transduction, which allowsus to acquire molecular data from clinically relevantcell types in vivo, i.e. human mesenchymal stem cells.Secondly, cells can be isolated from transgenic mice.Efficient protocols exist to isolate mesenchymal stemcells from bone marrow and fat tissue, and embryonicstem cells from pre-implantation blastocysts. Finally,parameters relevant to the bone regeneration processcan be monitored in transgenic animals. Because oftheir small size, it is very difficult to use mice for orthotopicimplantation but rats are frequently used for this purpose.The current advances in rat genetics, such as the comple-tion of the rat genome project [56] and strongly improvedtechnology to generate transgenic rats [57], bring BLIof orthotopic bone regeneration within reach. Otheradvances in the field that are likely to further reducethe threshold to apply BLI in bone tissue engineeringare improved imaging (software, cameras) with moresensitive and 3D imaging modes. Finally, luciferasefrom other species becomes available which emit lightat different wavelengths. Simultaneous imaging of differentparameters thus becomes possible [58,59]. In conclusion,we believe that BLI is knocking on the door of bonetissue engineering and can strongly contribute to ourunderstanding of the in vivo dynamics of bone tissueengineering.
References
[1] Lutolf MP, Hubbell JA. Synthetic biomaterials as instructive
extracellular microenvironments for morphogenesis in tissue engi-
neering. Nat Biotechnol 2005;23:47–55.
[2] Frost MC, Reynolds MM, Meyerhoff ME. Polymers incorporating
nitric oxide releasing/generating substances for improved biocompat-
ibility of blood-contacting medical devices. Biomaterials 2005;26:
1685–93.
[3] Liu Y, de Groot K, Hunziker EB. Osteoinductive implants: the mise-
en-scene for drug-bearing biomimetic coatings. Ann Biomed Eng
2004;32:398–406.
[4] Olivier V, Faucheux N, Hardouin P. Biomaterial challenges and
approaches to stem cell use in bone reconstructive surgery. Drug
Discov Today 2004;9:803–11.
[5] Liu X, Ma PX. Polymeric scaffolds for bone tissue engineering. Ann
Biomed Eng 2004;32:477–86.
[6] Heng BC, Cao T, Stanton LW, Robson P, Olsen B. Strategies for
directing the differentiation of stem cells into the osteogenic lineage in
vitro. J Bone Miner Res 2004;19:1379–94.
[7] Urist MR. Bone: formation by autoinduction. Science 1965;150:
893–9.
[8] Khan SN, Lane JM. The use of recombinant human bone
morphogenetic protein-2 (rhBMP-2) in orthopaedic applications.
Expert Opin Biol Ther 2004;4:741–8.
[9] Khor E, Lim LY. Implantable applications of chitin and chitosan.
Biomaterials 2003;24:2339–49.
[10] Shakesheff K, Cannizzaro S, Langer R. Creating biomimetic micro-
environments with synthetic polymer–peptide hybrid molecules.
J Biomater Sci Polym Ed 1998;9:507–18.
[11] Richardson TP, Peters MC, Ennett AB, Mooney DJ. Polymeric
system for dual growth factor delivery. Nat Biotechnol 2001;19:
1029–34.
[12] Langer R, Moses M. Biocompatible controlled release polymers for
delivery of polypeptides and growth factors. J Cell Biochem 1991;
45:340–5.
[13] Bezemer JM, Grijpma DW, Dijkstra PJ, van Blitterswijk CA, Feijen
J. Control of protein delivery from amphiphilic poly(ether ester)
multiblock copolymers by varying their water content using
emulsification techniques. J Control Release 2000;66:307–20.
[14] Anderson DG, Levenberg S, Langer R. Nanoliter-scale synthesis of
arrayed biomaterials and application to human embryonic stem cells.
Nat Biotechnol 2004;22:863–6.
[15] Qi H, Aguiar DJ, Williams SM, La Pean A, Pan W, Verfaillie CM.
Identification of genes responsible for osteoblast differentiation from
human mesodermal progenitor cells. Proc Natl Acad Sci U S A
2003;100:3305–10.
[16] Korchynskyi O, Dechering KJ, Sijbers AM, Olijve W, ten Dijke P.
Gene array analysis of bone morphogenetic protein type I receptor-
induced osteoblast differentiation. J Bone Miner Res 2003;18:
1177–85.
[17] Bailey SN, Sabatini DM, Stockwell BR. Microarrays of small
molecules embedded in biodegradable polymers for use in mamma-
lian cell-based screens. Proc Natl Acad Sci USA 2004;101:16144–9.
[18] Herschman HR. Molecular imaging: looking at problems, seeing
solutions. Science 2003;302:605–8.
[19] Maggi A, Ottobrini L, Biserni A, Lucignani G, Ciana P. Techniques:
reporter mice—a new way to look at drug action. Trends Pharmacol
Sci 2004;25:337–42.
[20] Edinger M, Cao YA, Hornig YS, Jenkins DE, Verneris MR,
Bachmann MH, et al. Advancing animal models of neoplasia
through in vivo bioluminescence imaging. Eur J Cancer 2002;38:
2128–36.
[21] Honigman A, Zeira E, Ohana P, Abramovitz R, Tavor E, Bar I, et al.
Imaging transgene expression in live animals. Mol Ther 2001;4:
239–49.

ARTICLE IN PRESSJ. de Boer et al. / Biomaterials 27 (2006) 1851–18581858
[22] Levenberg S, Rouwkema J, Macdonald M, Garfein ES, Kohane DS,
Darland DC, et al. Engineering vascularized skeletal muscle tissue.
Nat Biotechnol 2005;23:879–84.
[23] Zhang W, Feng JQ, Harris SE, Contag PR, Stevenson DK, Contag
CH. Rapid in vivo functional analysis of transgenes in mice using
whole body imaging of luciferase expression. Transgenic Res
2001;10:423–34.
[24] Ciana P, Raviscioni M, Mussi P, Vegeto E, Que I, Parker MG, et al.
In vivo imaging of transcriptionally active estrogen receptors. Nat
Med 2003;9:82–6.
[25] Soling A, Rainov NG. Bioluminescence imaging in vivo—application
to cancer research. Expert Opin Biol Ther 2003;3:1163–72.
[26] Contag PR. Whole-animal cellular and molecular imaging to
accelerate drug development. Drug Discov Today 2002;7:555–62.
[27] Wetterwald A, van der Pluijm G, Que I, Sijmons B, Buijs J,
Karperien M, et al. Optical imaging of cancer metastasis to bone
marrow: a mouse model of minimal residual disease. Am J Pathol
2002;160:1143–53.
[28] Blum JS, Temenoff JS, Park H, Jansen JA, Mikos AG, Barry MA.
Development and characterization of enhanced green fluorescent
protein and luciferase expressing cell line for non-destructive
evaluation of tissue engineering constructs. Biomaterials 2004;25:
5809–19.
[29] Korchynskyi O, ten Dijke P. Identification and functional character-
ization of distinct critically important bone morphogenetic protein-
specific response elements in the Id1 promoter. J Biol Chem
2002;277:4883–91.
[30] Monteiro RM, de Sousa Lopes SM, Korchynskyi O, ten Dijke P,
Mummery CL. Spatio-temporal activation of Smad1 and Smad5 in
vivo: monitoring transcriptional activity of Smad proteins. J Cell Sci
2004;117:4653–63.
[31] Clemens TL, Tang H, Maeda S, Kesterson RA, Demayo F, Pike JW,
et al. Analysis of osteocalcin expression in transgenic mice reveals a
species difference in vitamin D regulation of mouse and human
osteocalcin genes. J Bone Miner Res 1997;12:1570–6.
[32] Iris B, Zilberman Y, Zeira E, Galun E, Honigman A, Turgeman G,
et al. Molecular imaging of the skeleton: quantitative real-time
bioluminescence monitoring gene expression in bone repair and
development. J Bone Miner Res 2003;18:570–8.
[33] Opas EE, Gentile MA, Rossert JA, de Crombrugghe B, Rodan GA,
Schmidt A. Parathyroid hormone and prostaglandin E2 preferentially
increase luciferase levels in bone of mice harboring a luciferase
transgene controlled by elements of the pro-alpha1(I) collagen
promoter. Bone 2000;26:27–32.
[34] Chen JJ, Jin H, Ranly DM, Sodek J, Boyan BD. Altered expression
of bone sialoproteins in vitamin D-deficient rBSP2.7Luc transgenic
mice. J Bone Miner Res 1999;14:221–9.
[35] Chen J, Sodek J, Thomas HF, Ranly DM. Dexamethasone stimulates
luciferase gene expression through the rat bone sialoprotein gene
promoter in transgenic mice. Connect Tissue Res 1996;35:33–9.
[36] Gopalakrishnan R, Thomas PE, Benson MD, Wang D, Franceschi
RT. A homeodomain protein binding element in the bone sialopro-
tein promoter is critical for tissue-specific expression in bone.
Connect Tissue Res 2003;44(Suppl 1):154–60.
[37] van Bezooijen RL, Roelen BA, Visser A, van der Wee-Pals L, de Wilt
E, Karperien M, et al. Sclerostin is an osteocyte-expressed negative
regulator of bone formation, but not a classical BMP antagonist.
J Exp Med 2004;199:805–14.
[38] Kadurugamuwa JL, Sin L, Albert E, Yu J, Francis K, DeBoer M,
et al. Direct continuous method for monitoring biofilm infection in a
mouse model. Infect Immun 2003;71:882–90.
[39] Carlsen H, Moskaug JO, Fromm SH, Blomhoff R. In vivo imaging of
NF-kappa B activity. J Immunol 2002;168:1441–6.
[40] Zhang N, Weber A, Li B, Lyons R, Contag PR, Purchio AF, et al.
An inducible nitric oxide synthase-luciferase reporter system for in
vivo testing of anti-inflammatory compounds in transgenic mice.
J Immunol 2003;170:6307–19.
[41] Laxman B, Hall DE, Bhojani MS, Hamstra DA, Chenevert TL, Ross
BD, et al. Noninvasive real-time imaging of apoptosis. Proc Natl
Acad Sci U S A 2002;99:16551–5.
[42] Doubrovin M, Ponomarev V, Beresten T, Balatoni J, Bornmann W,
Finn R, et al. Imaging transcriptional regulation of p53-dependent
genes with positron emission tomography in vivo. Proc Natl Acad Sci
USA 2001;98:9300–5.
[43] Wang W, El-Deiry WS. Bioluminescent molecular imaging of
endogenous and exogenous p53-mediated transcription in vitro and
in vivo using an HCT116 human colon carcinoma xenograft model.
Cancer Biol Ther 2003;2:196–202.
[44] Miller JC, Pien HH, Sahani D, Sorensen AG, Thrall JH. Imaging
angiogenesis: applications and potential for drug development. J Natl
Cancer Inst 2005;97:172–87.
[45] Miles KA. Perfusion CT for the assessment of tumour vascularity:
which protocol? Br J Radiol 2003;76(Spec No. 1):S36–42.
[46] Zhang N, Fang Z, Contag PR, Purchio AF, West DB. Tracking
angiogenesis induced by skin wounding and contact hypersensitivity
using a Vegfr2-luciferase transgenic mouse. Blood 2004;103:617–26.
[47] Otani A, Kinder K, Ewalt K, Otero FJ, Schimmel P, Friedlander M.
Bone marrow-derived stem cells target retinal astrocytes and can
promote or inhibit retinal angiogenesis. Nat Med 2002;8:1004–10.
[48] Liu YL, Ang SO, Weigent DA, Prchal JT, Bloomer JR. Regulation of
ferrochelatase gene expression by hypoxia. Life Sci 2004;75:2035–43.
[49] Shibata T, Giaccia AJ, Brown JM. Development of a hypoxia-
responsive vector for tumor-specific gene therapy. Gene Ther
2000;7:493–8.
[50] Mogford JE, Roy NK, Cross KJ, Mustoe TA. Use of hypoxia-
inducible factor signal transduction pathway to measure O2 levels
and modulate growth factor pathways. Wound Repair Regen
2003;11:496–503.
[51] Barker N, Morin PJ, Clevers H. The Yin-Yang of TCF/beta-catenin
signaling. Adv Cancer Res 2000;77:1–24.
[52] Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R, Israel A.
Signalling downstream of activated mammalian Notch. Nature
1995;377:355–8.
[53] De Boer J, Wang HJ, Van Blitterswijk C. Effects of Wnt signaling on
proliferation and differentiation of human mesenchymal stem cells.
Tissue Eng 2004;10:393–401.
[54] Roose J, Molenaar M, Peterson J, Hurenkamp J, Brantjes H, Moerer
P, et al. The Xenopus Wnt effector XTcf-3 interacts with Groucho-
related transcriptional repressors. Nature 1998;395:608–12.
[55] Rosin-Arbesfeld R, Cliffe A, Brabletz T, Bienz M. Nuclear export of
the APC tumour suppressor controls beta-catenin function in
transcription. EMBO J 2003;22:1101–13.
[56] Gibbs RA, Weinstock GM, Metzker ML, Muzny DM, Sodergren EJ,
Scherer S, et al. Genome sequence of the Brown Norway rat yields
insights into mammalian evolution. Nature 2004;428:493–521.
[57] Lois C, Hong EJ, Pease S, Brown EJ, Baltimore D. Germline
transmission and tissue-specific expression of transgenes delivered by
lentiviral vectors. Science 2002;295:868–72.
[58] Bhaumik S, Gambhir SS. Optical imaging of Renilla luciferase
reporter gene expression in living mice. Proc Natl Acad Sci U S A
2002;99:377–82.
[59] Bhaumik S, Lewis XZ, Gambhir SS. Optical imaging of Renilla
luciferase, synthetic Renilla luciferase, and firefly luciferase reporter
gene expression in living mice. J Biomed Opt 2004;9:578–86.





























