Embed Size (px)
Citation preview

Page 9
Biological Molecules — Introduction
• proteins: amino acids
protein folding, binding
synthesis
catalysis
cofactors
• nucleic acids: DNA
RNA
carcinogens, drugs
• carbohydrates: simple
oligosaccharides
• “small molecules”: natural products

Page 10
STRUCTUREAND
CHEMISTRYOF
PROTEINS
Page 11
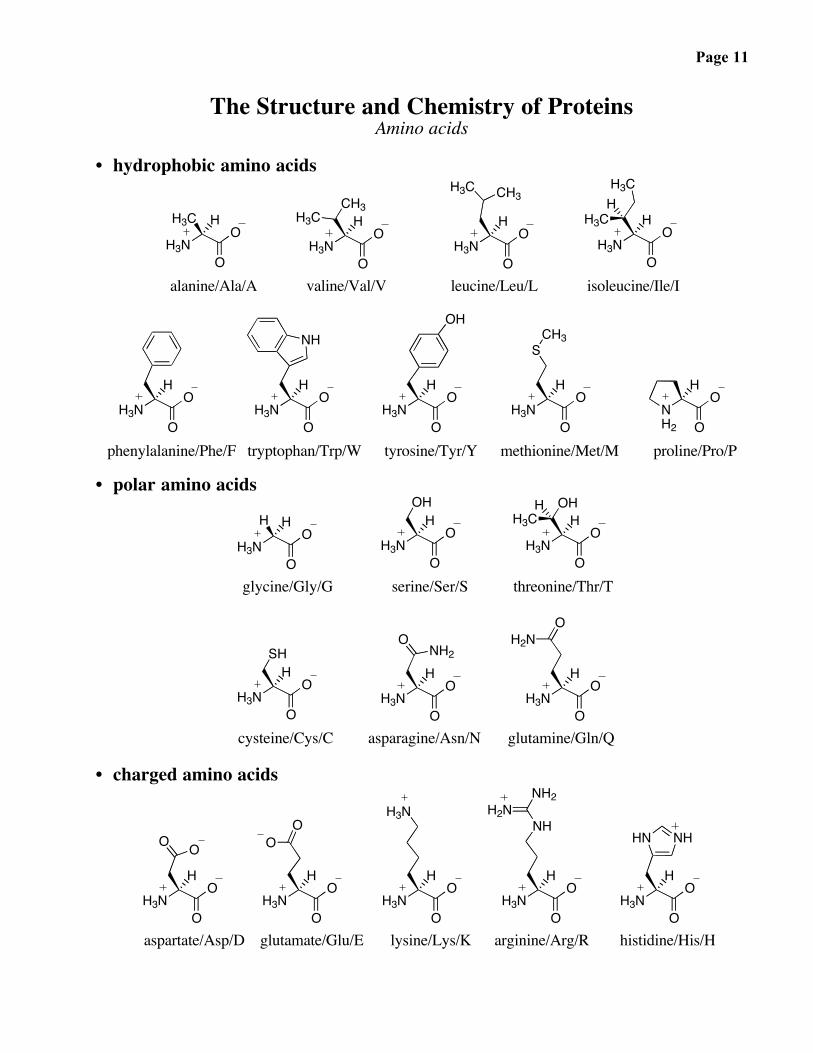
The Structure and Chemistry of ProteinsAmino acids
• hydrophobic amino acids
H3NO
OH3C H
H3NO
OH
H3NO
OH
H3NO
OH
CH3H3C
H3C CH3H3C
H3CH
alanine/Ala/A valine/Val/V leucine/Leu/L isoleucine/Ile/I
H3NO
OH
H3NO
OH
H3NO
OH
H3NO
OH
NH2 O
OH
NHOH
SCH3
phenylalanine/Phe/F tryptophan/Trp/W tyrosine/Tyr/Y methionine/Met/M proline/Pro/P
• polar amino acids
H3NO
OH H
H3NO
OH
H3NO
OH
OH OHH3C
H
glycine/Gly/G serine/Ser/S threonine/Thr/T
H3NO
OH
H3NO
OH
H3NO
OH
SHO
NH2
OH2N
cysteine/Cys/C asparagine/Asn/N glutamine/Gln/Q
• charged amino acids
H3NO
OH
H3NO
OH
H3NO
OH
H3NO
OH
H3NO
OH
O O
OO
H3NNH
H2NNH2
NHHN
aspartate/Asp/D glutamate/Glu/E lysine/Lys/K arginine/Arg/R histidine/His/H

Page 12
The Structure and Chemistry of ProteinsConformational analysis of small molecules
sp3-sp3 bonds: A quick review
• ethane
• butane

Page 13
Conformational analysis of small molecules — continued
• pentane

Page 14
Conformational analysis of small molecules — continued
• cyclohexane

Page 15
The Structure and Chemistry of ProteinsConformational parameters of peptides
• Valine (Val, V)
• Leucine (Leu, L)

Page 16
Conformational parameters of peptides — continued
• Isoleucine (Ile, I)
• Methionine (Met, M)

Page 17
A role for methionine flexibility in biology

Page 18
Conformational parameters of peptides — continued
sp2-sp3 bonds
• propene
• substituted propene
• peptide (amide) bond

Page 19
Conformational parameters of peptides — continued
• β-strands, β-sheet

Page 20
The Structure and Chemistry of ProteinsNon-covalent interactions and protein folding
Forces that are important for the folding of proteins, including packing of side chains,and for the binding of ligands to receptors (and substrates / transition states to enzymes)include:
• hydrogen bonds

Page 21
• electrostatic interactions (salt bridges)
• disulfide bonds

Page 22
• hydrophobic interactions

Page 23
Denaturing proteins
• hydrogen bonding
• electrostatics
• disulfides
• hydrophobic interactions
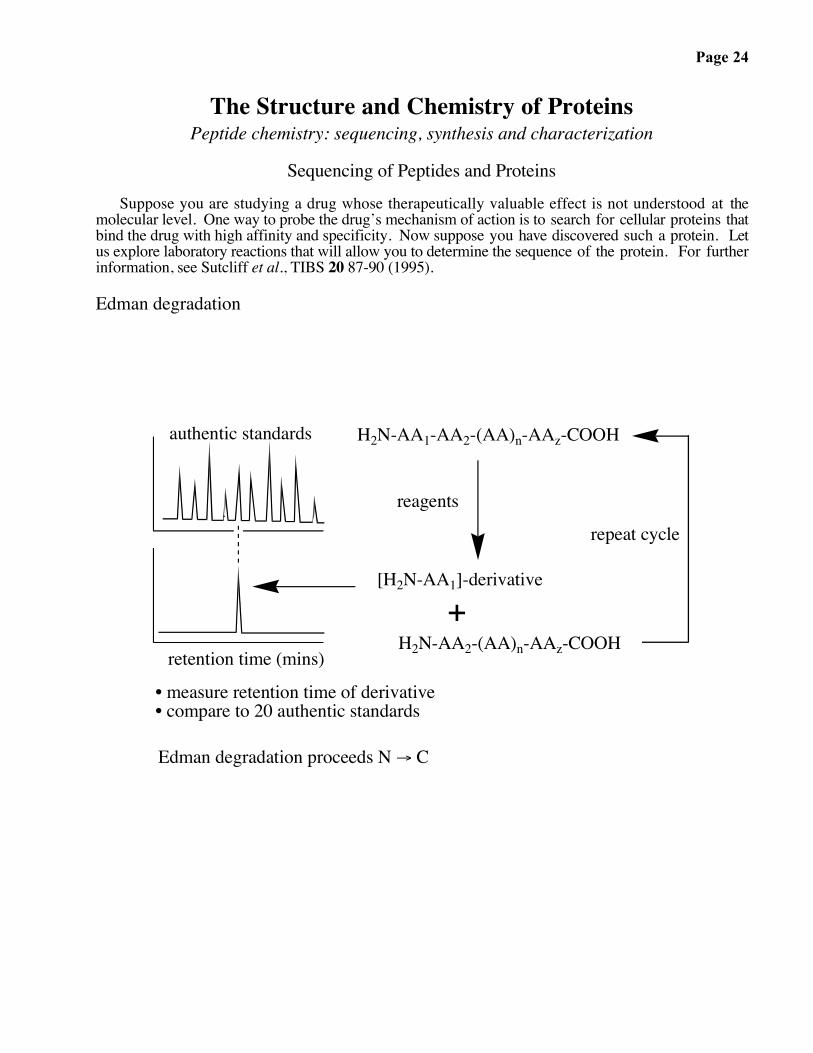
Page 24
The Structure and Chemistry of ProteinsPeptide chemistry: sequencing, synthesis and characterization
Sequencing of Peptides and ProteinsSuppose you are studying a drug whose therapeutically valuable effect is not understood at the
molecular level. One way to probe the drug’s mechanism of action is to search for cellular proteins thatbind the drug with high affinity and specificity. Now suppose you have discovered such a protein. Letus explore laboratory reactions that will allow you to determine the sequence of the protein. For furtherinformation, see Sutcliff et al., TIBS 20 87-90 (1995).
Edman degradation
H2N-AA1-AA2-(AA)n-AAz-COOH
[H2N-AA1]-derivative
reagents
• measure retention time of derivative• compare to 20 authentic standards
repeat cycle
Edman degradation proceeds N → C
H2N-AA2-(AA)n-AAz-COOH+
authentic standards
retention time (mins)

Page 25
Edman Degradation: Peptide Bond Cleavage
-H+
-H+
+H+
thiazolinone derivative
peptide (n-1)
thiourea derivative-NH2 group at N-terminus of peptide or protein: strong nucleophile
phenyl isothiocyanate:strong electrophile
H2NN
O
H OR1 H
R2HN
Ph H R2
HR1 O
N
H
O
NN
N
H
H
S
H
S
N
H
H
NH
NH
H
R2O
R1
O ON
O
N
R2
H
H
H
R1
HNN
H
S
H
H
O H
NN
H
S
N H
R1
N
O
N
R2
H
H
HS
NH
NH
H
R2O
H
O
R1
H
N
H
Ph
Ph
PhPh
Ph
tetrahedralintermediate
δ-
δ+
+
C
S
(lower pH)
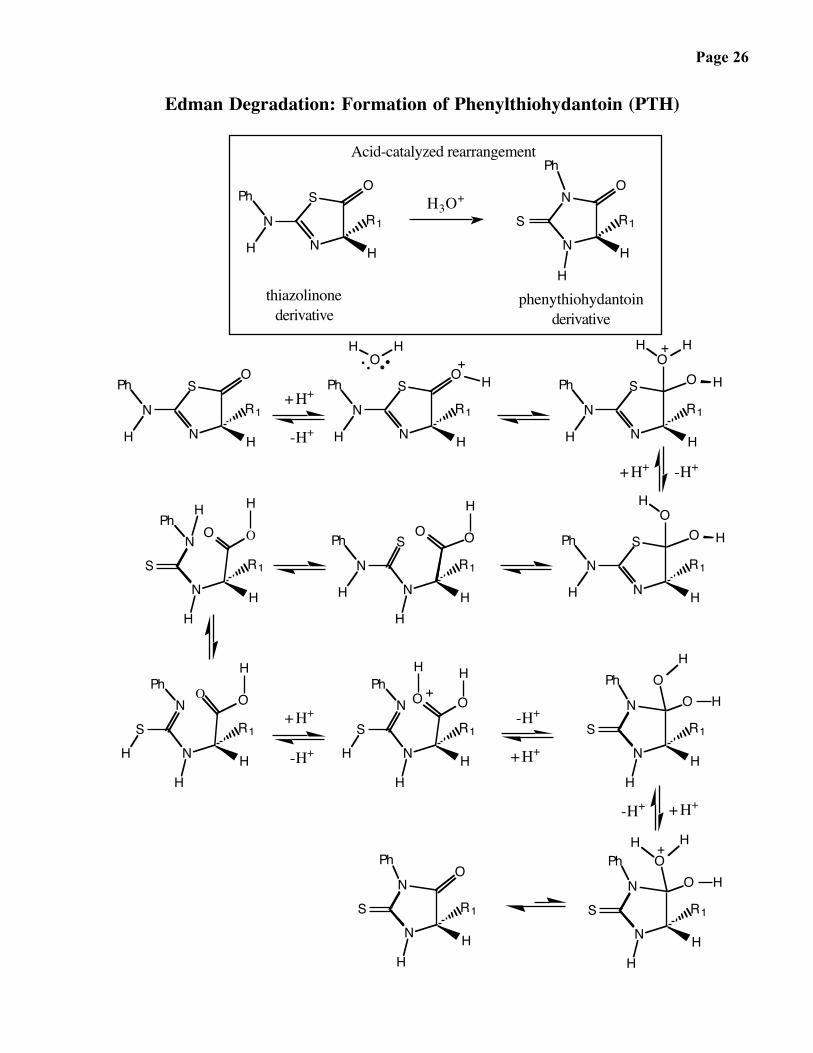
Page 26
Edman Degradation: Formation of Phenylthiohydantoin (PTH)
Acid-catalyzed rearrangement
H3O+
phenythiohydantoinderivative
thiazolinonederivative
N
SO
R1
H
N
H
Ph
SN
HH
R1
ON
Ph
+
-H+
+H+
+
+H+
-H+
+
-H+
+H+
+
+H+-H+
N
Ph
H N H
R1
OS
N
SO
R1
H
N
H
Ph H
HO
H
OH
O H
N
Ph
H N H
R1
S
OHH
N
S O
R1
H
N
H
Ph HO
N
Ph
H N H
R1
O
H
SO
N
N
H
O
R1
H
PhH
S
SH
PhN
N H
R1
H
N
N
H
R1
H
Ph
SH
O H
S
N
Ph
N
N H
R1
H
OH
OH
N
H
O
R1
H
Ph
S
H
H
S
Ph
N
N H
R1
O
H
-H++H+
H H
O O
H
O O
H H

Page 27
Peptide Sequencing

Page 28
Mass Spectrometry
IONIZATION SOURCE
1. Matrix-Assisted Laser Desorption Ionization (MALDI)

Page 29
Mass Spectrometry (continued)
2. Electrospray Ionization (ESI)
MASS ANALYZERS
1. Time-of-Flight (TOF)

Page 30
Mass Spectrometry (continued)
2. Quadrupole (Q)
TANDEM MASS SPECTROMETRY (MS/MS)

Page 31
Mass Spectrometry (continued)
Peptide Fragmentation
AA residuemass
Ala 71.0Val 99.1Gly 57.0Ser 87.0

Page 32
Synthesis of Peptides and Proteins
To study a newly sequenced protein in detail, you will need sufficient quantities of it. The ability tosynthesize proteins provided novel access to them when the methodology was first introduced severaldecades ago. In more recent years, molecular biological techniques for the overproduction ofrecombinant proteins have had a significant impact on the availability of proteins. Methods developedfor the laboratory synthesis of proteins during the past two years have now greatly expanded our abilityto carry out structure-function analyses, primarily by allowing the synthesis of proteins with elementsother than the naturally occurring twenty amino acids.
• laboratory:
NH
R2
O
HN
HN
O
key reaction:(DCC)
+
N C N
(DCU)
DMFR1R1 NH2
R2
O
OH+
• selectivity problem
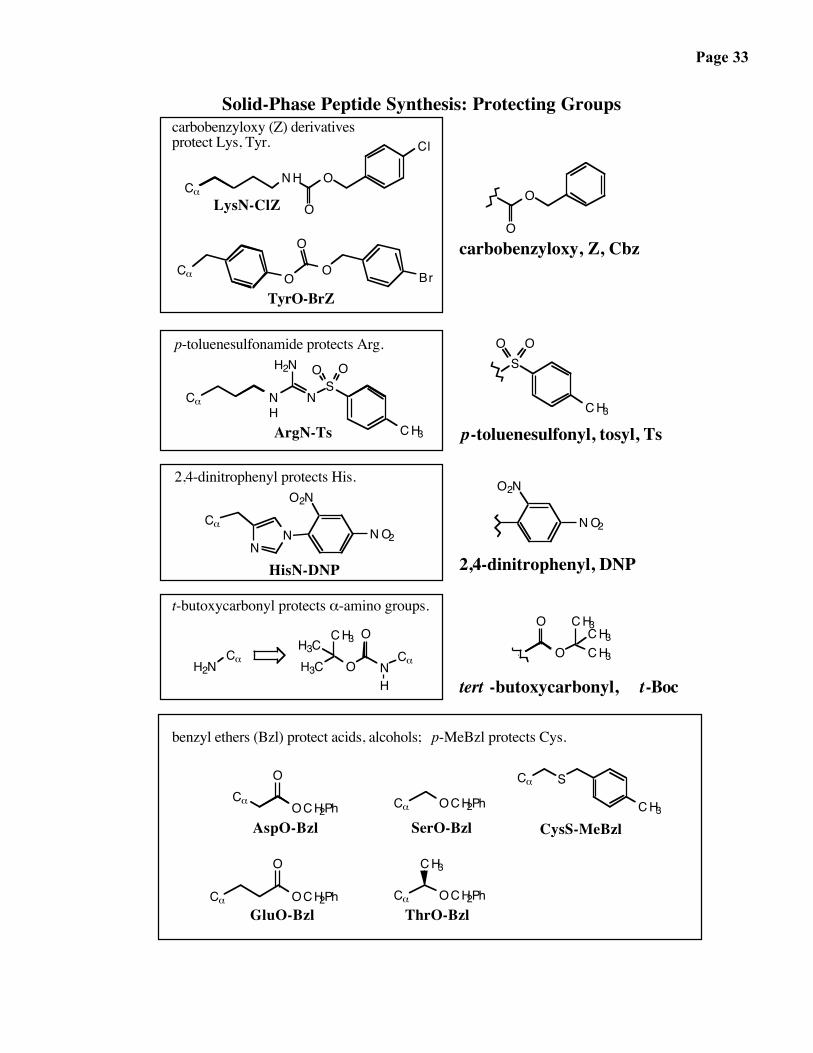
Page 33
Solid-Phase Peptide Synthesis: Protecting Groups
CαNH O
O
Cl
O
O
Br
O
OO
NH
Cα
NS
H2N O O
Cα
CH3
CH3
SOO
Cα
NN
O2N
NO2NO2
O2N
H2NCα
NCαOH3C
H
CH3 OH3C
Cα
Cα OCH2Ph
OCH2PhCαOCH2Ph
O
OCH2Ph
CH3
Cα S
Cα
O
2,4-dinitrophenyl protects His.
p-toluenesulfonamide protects Arg.
HisN-DNP
ArgN-Ts
carbobenzyloxy (Z) derivatives protect Lys, Tyr.
TyrO-BrZ
carbobenzyloxy, Z, Cbz
LysN-ClZ
t-butoxycarbonyl protects α-amino groups.
2,4-dinitrophenyl, DNP
AspO-Bzl
GluO-Bzl
SerO-Bzl
ThrO-Bzl
CysS-MeBzl
benzyl ethers (Bzl) protect acids, alcohols; p-MeBzl protects Cys.
O
O
CH3CH3CH3
CH3
p-toluenesulfonyl, tosyl, Ts
tert -butoxycarbonyl, t-Boc
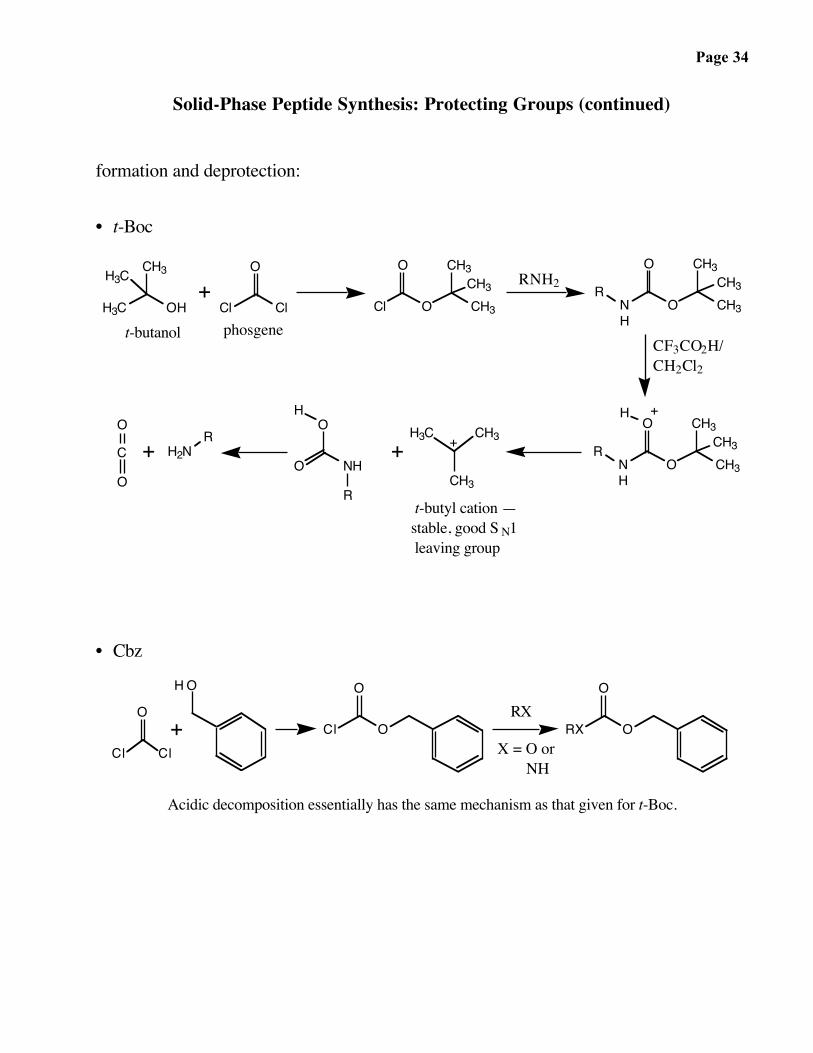
Page 34
Solid-Phase Peptide Synthesis: Protecting Groups (continued)
formation and deprotection:
• t-Boc
t-butyl cation —stable, good S N1 leaving group
++ +
+
CF3CO2H/CH2Cl2
RNH2
phosgenet-butanol
+H3C OH
CH3H3C
Cl Cl
O
CH3OCl
O CH3CH3 CH3
CH3O
NH
O CH3R
HO
O NH
RCH3O
CH3CH3
NH
O H3C
CH3
CH3C
O
O
H2NR
H
R
• Cbz
X = O or NH
RX+
Cl Cl
O
HO
Cl O
O O
ORX
Acidic decomposition essentially has the same mechanism as that given for t-Boc.
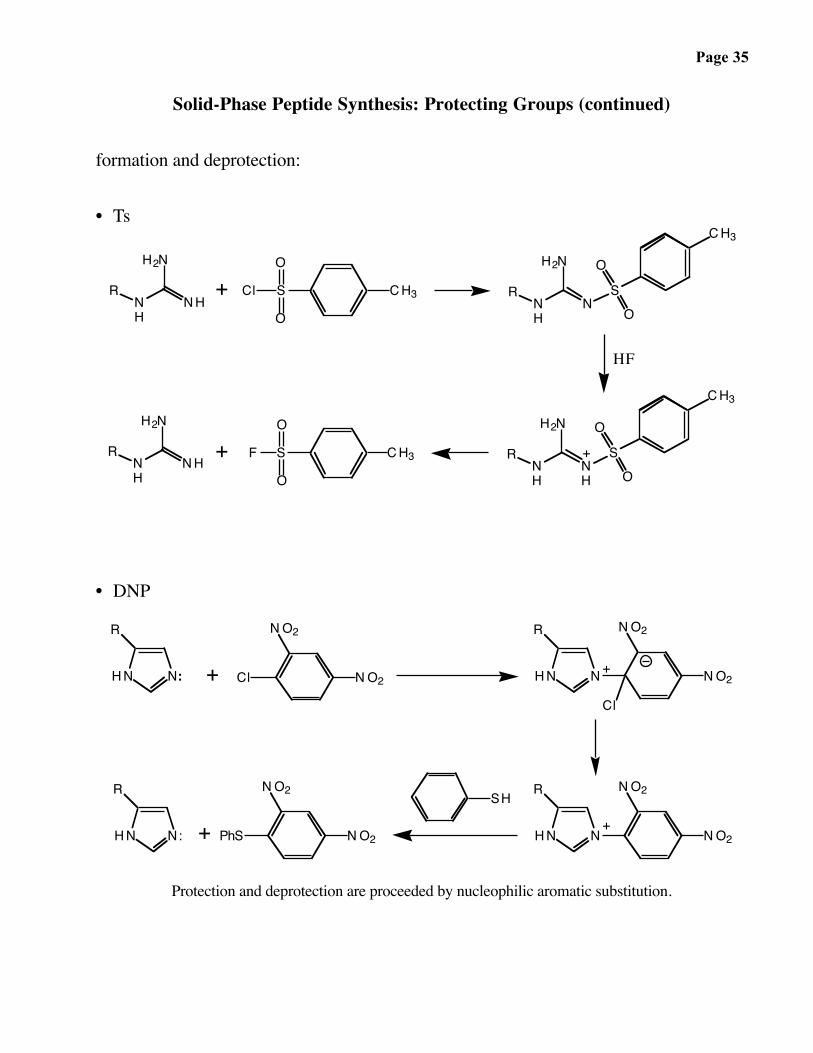
Page 35
Solid-Phase Peptide Synthesis: Protecting Groups (continued)
formation and deprotection:
• Ts
+
HF
++
NH
NH
H2N
R S
O
O
ClNH
R
H2N
NSO
O
S
O
ONH
NH
H2N
RNH
R
H2N
NHSF
O
O
CH3
CH3
CH3
CH3
• DNP
+
+
+
+
HN N:
R NO2
NO2Cl
Cl
NO2
NO2R
HN N
NHN
R NO2
NO2PhS
SH
NO2
NO2R
HN N:
Protection and deprotection are proceeded by nucleophilic aromatic substitution.

Page 36
Solid-Phase Peptide Synthesis: General Strategy
Growing peptide chain is attached to a solid polymeric support:
polystyrenelinker
P peptide-NH2 peptideP NH2
Unlike ribosomal peptide synthesis, chemical synthesis proceeds from C-terminus to N-terminus; after each round of synthesis, a new N-terminalresidue is added on.
reagents reagentspeptide-NH2Ppeptide-NH2P
P peptide-NH2
peptide-NH2PP peptide-NH2peptide-NH2P
Peptides are attached to solid support, which is loadedinto a column -- immobilization.
Reagents are pumped through the column in liquidform. This format allows rapid changes from one set ofsynthetic reagents to another. You will notice in theproceeding discussions that peptide synthesis requiressequential usage of many reagents that are chemicallyincompatible.
Reaction takes place on the surface of a solid.

Page 37
Solid-Phase Peptide Synthesis: the Reactions
Polymeric support: functionalized polystyrene derivative:
immobilized SN2 acceptor
protected C-terminal amino acid (Cys, in this case);
carboxyl free to react.
polymer charged with C-terminal amino acid residue.
Ph Ph
Cl
BocNHO-Cs+
O
SCH2PhMe
PhPh
O
SCH2PhMe
O
BocNH
CF3CO2H / CH2Cl214 min.2. neutralize with
10%
in DMF
3. wash with DMFO
SCH2PhMe
O
CF3CO2- H3N
N
H2NO
O
SCH2PhMe
BocNHOH
O
CO2CH2PhCO2CH2Ph
O
OBocNH
NHCyC
NHCy
O
SCH2PhMe
OHN
BocNH
CO2CH2Ph
O
polymeric support+
DCC
DMF
DCU
P
P
P
1. wash with CH 2Cl2
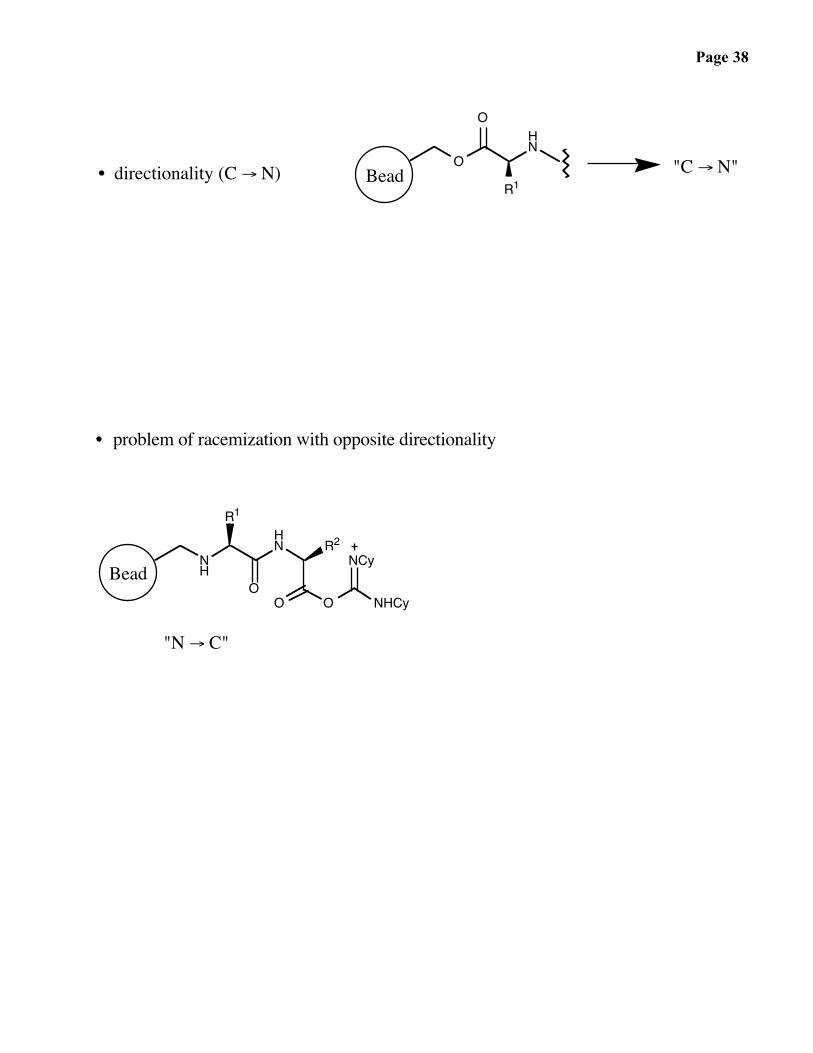
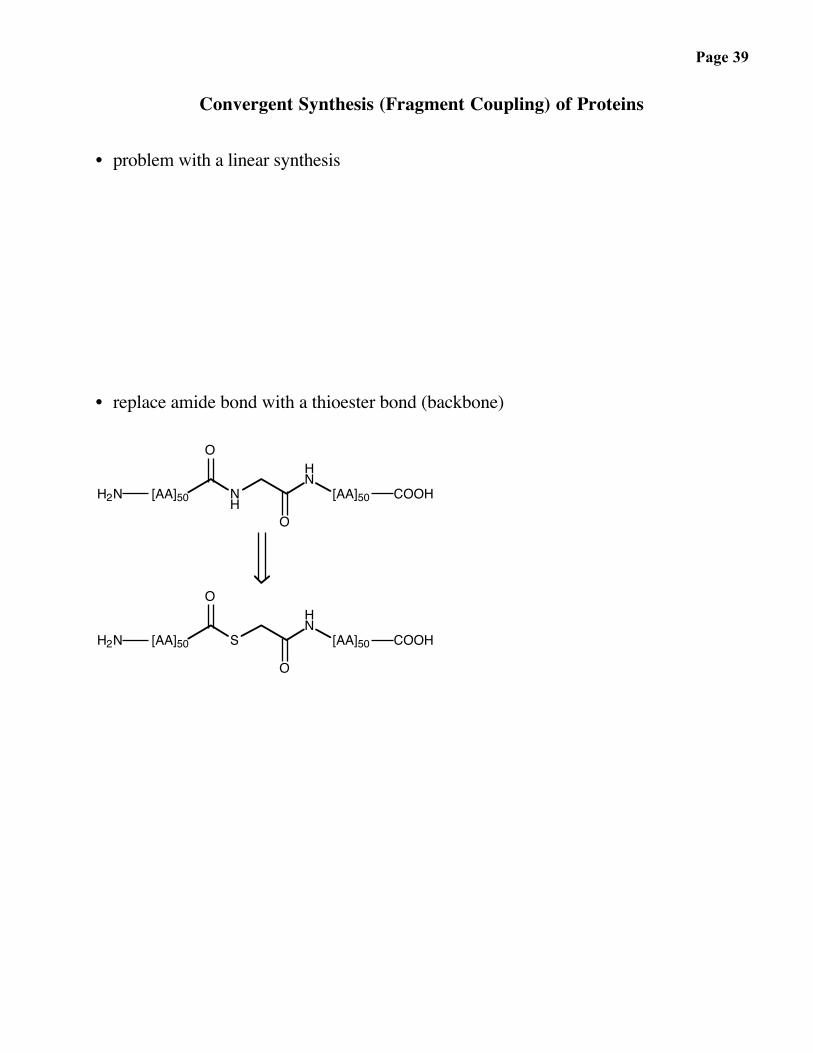
Page 38
O
HN
O
R1• directionality (C → N) Bead "C → N"
• problem of racemization with opposite directionality
NH
HN
O
R1
OO
R2NCy
NHCy
Bead
"N → C"
Page 39
Convergent Synthesis (Fragment Coupling) of Proteins
• problem with a linear synthesis
• replace amide bond with a thioester bond (backbone)
[AA]50 NH
HN
[AA]50
O
O
COOHH2N
[AA]50 S
HN
[AA]50
O
O
COOHH2N

Page 40
• synthesis of the fragments
S
HN[AA]
O
R1
O
NHBoc
Rω
O
HSNHBoc
R1
O
HN[AA]
O
R1
O
NHBoc
Rω
Bead
start with:
Bead
conventional starting unit
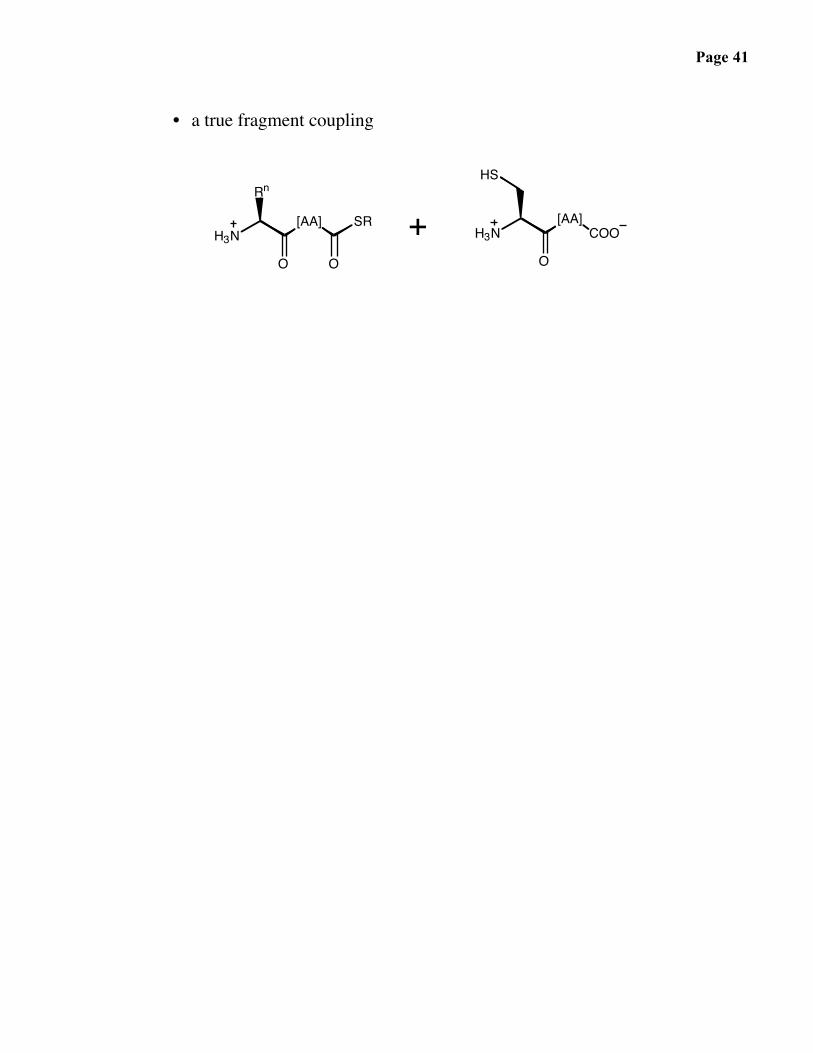
Page 41
• a true fragment coupling
O
[AA]COO
O
[AA]
Rn
SR
O
+HS
H3N H3N
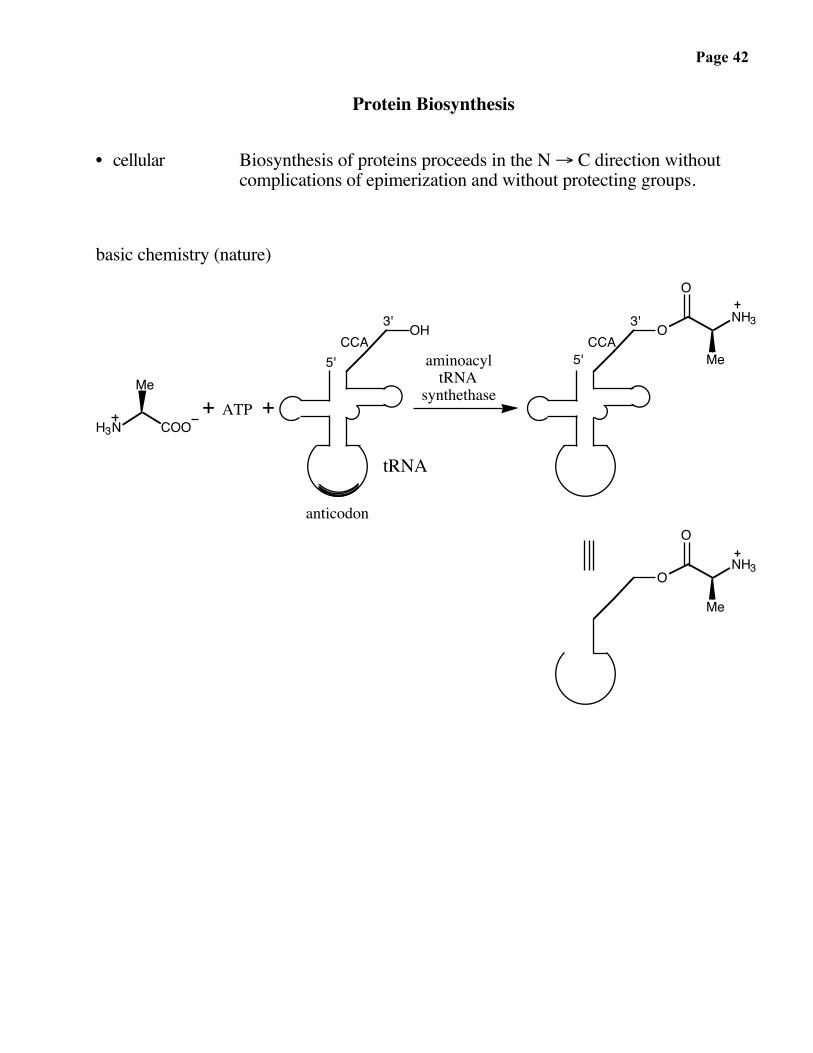
Page 42
Protein Biosynthesis
• cellular Biosynthesis of proteins proceeds in the N → C direction withoutcomplications of epimerization and without protecting groups.
basic chemistry (nature)
H3N COO
Me
OH O
+ ATP +
O
NH3
MeaminoacyltRNA
synthethase
anticodon
tRNA
O
3'
O
NH3
Me
CCACCA3'
5' 5'

Page 43
Mechanism of Peptide Bond Formation in Biological Systems
H3N COO
R H
O
N
N N
N
NH2
HO OH
OP
O
O
OP
O
O
OP
O
O
O
O
N
N N
N
NH2
HO OH
OP
O
O
O
O
H3N
R H
O
HO OH
N
N N
N
NH2
O
O OH
N
N N
N
NH2
O
H3N
HRH3N
NH
OtRNA
O
HR
amino acid(carboxylate)
ATP (phosphoric acid anhydride)
R' H
pyrophosphate
aminoacyl-tRNAsynthase
O
tRNA
tRNA (alcohol)
aminoacyl-AMP (mixed anhydride)
AMP
OtRNAH2N
O
aminoacyl-tRNA synthase
tRNA
aminoacyl-tRNA(ester)
tRNA
R' H
ribosomal complex
dipeptidyl-tRNA (amide)aminoacyl-tRNA

Page 44
The Ribosome
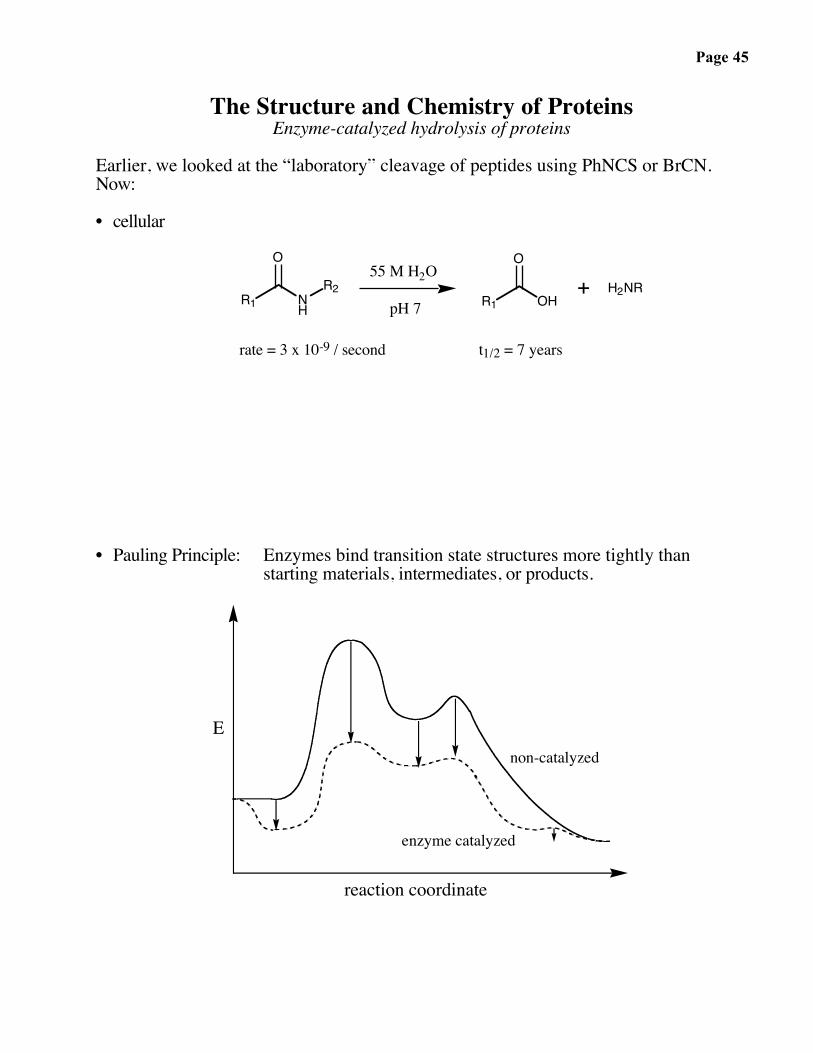
Page 45
The Structure and Chemistry of ProteinsEnzyme-catalyzed hydrolysis of proteins
Earlier, we looked at the “laboratory” cleavage of peptides using PhNCS or BrCN.Now:
• cellular
R1
O
NH
R255 M H2O
pH 7 R1
O
OH+ H2NR
rate = 3 x 10-9 / second t1/2 = 7 years
• Pauling Principle: Enzymes bind transition state structures more tightly thanstarting materials, intermediates, or products.
reaction coordinate
E
enzyme catalyzed
non-catalyzed
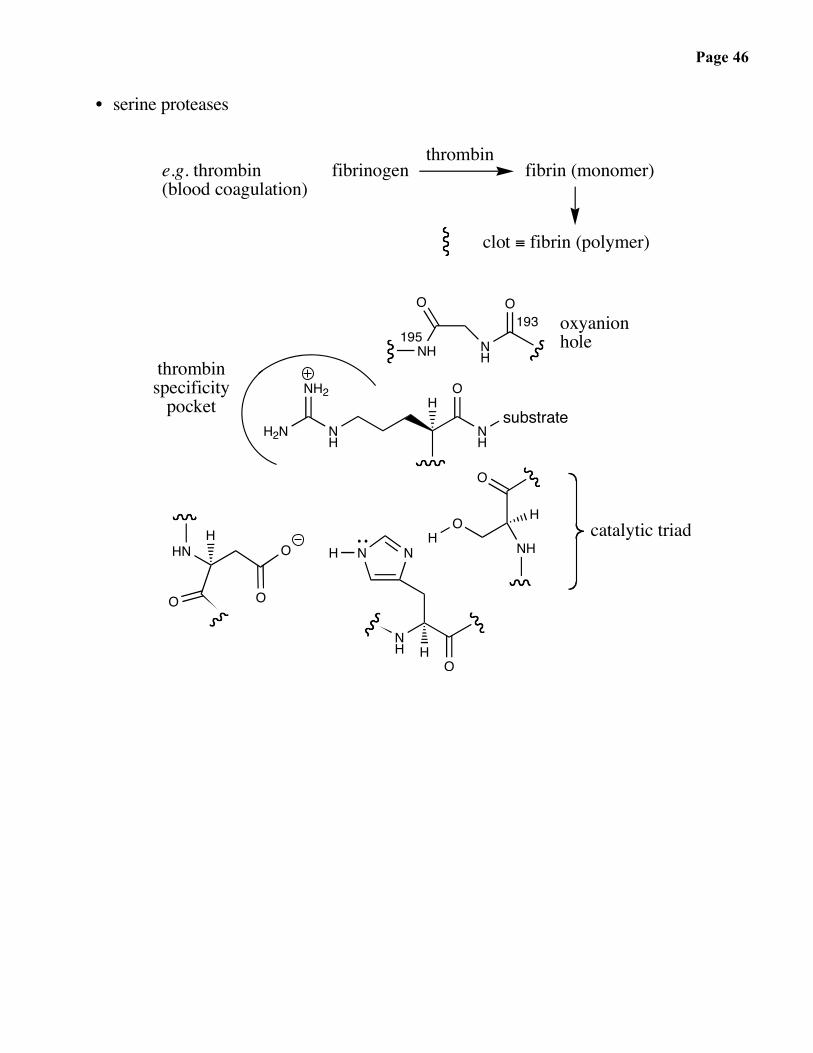
Page 46
• serine proteases
NH
O
NH
O
H2N NH
NH
substrate
NH2H
O
HO H
NHNNH
HNH
HN O
O
H
fibrinogenthrombin
clot ≡ fibrin (polymer)
e.g. thrombin(blood coagulation)
fibrin (monomer)
catalytic triad
193 oxyanionhole
O
O
O
thrombinspecificity
195

Page 47
• serine proteases (continued)
N NH
His
O
O
NH
N N
O
H
H H
O
OAsp
NH
NH2
H2N
Ser
N NH
His
O
O
N
N N
O
H H
O
OAsp
Arg
Ser
H H
N NH
His
O
O
N N
O
H H
O
OAsp
Arg Ser
OHH
N NH
His
O
O
O
N N
O
H
H H
O
OAsp
Arg
H
Ser

Page 48
• serine protease inhibitors: natural
H2N NH
HN
NH2 O
OOHNNH
O
N
NH
O
H3CNH
O
cyclotheonamide
+
OH
• serine protease inhibitors: designed
H
O
R
NH
O
R
NH
BOH
OH
R
NH
Cl

Page 49
• aspartyl proteases
O
HN
Asp
OOH
Asp
OO
H
OH
H2N
Asp
OOH
Asp
OO
OO
HO
N
Asp
OO
Asp
OO
O
H H
H H
O
HN
Asp
OO
Asp
OOH
O
H H

Page 50
• aspartyl proteases — a medically relevant case: HIV protease
host
chromosomeDNA
H3N
HIV genome
transcription → mRNAtranslation → protein
env integraseCOO
reversetranscriptase PR pol
PR
virus particle
proteinproducts
etc.
Page 51
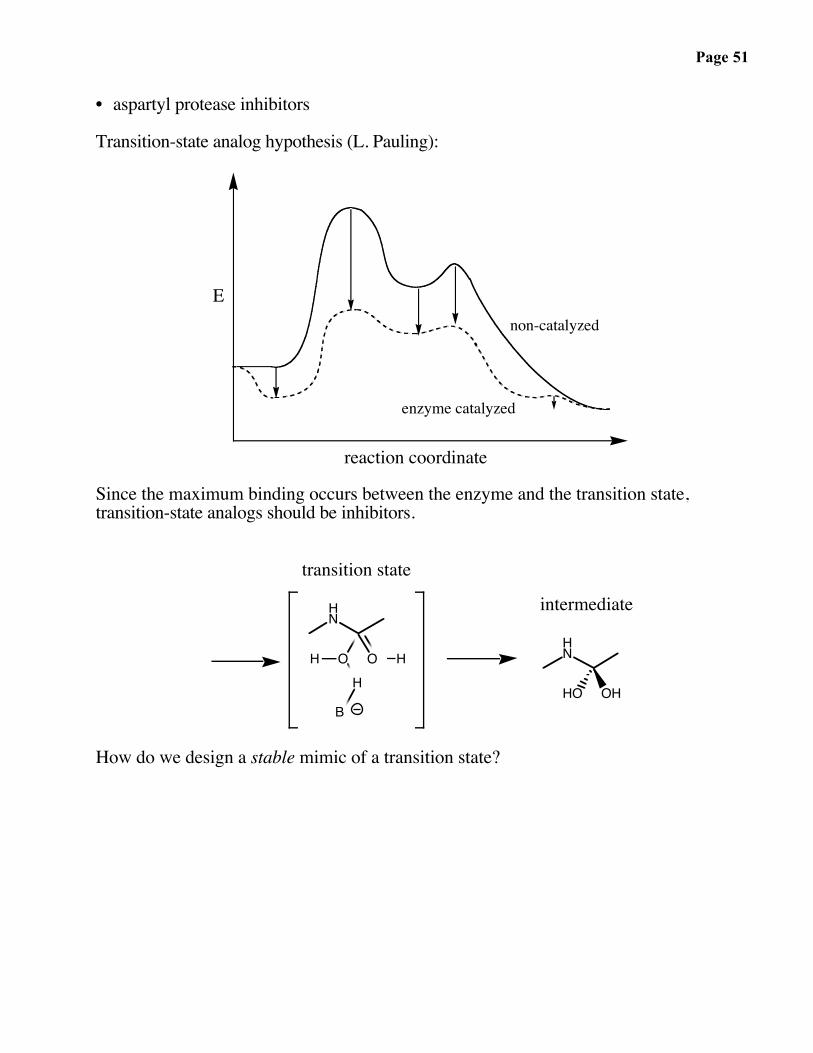
• aspartyl protease inhibitors
Transition-state analog hypothesis (L. Pauling):
reaction coordinate
E
enzyme catalyzed
non-catalyzed
Since the maximum binding occurs between the enzyme and the transition state,transition-state analogs should be inhibitors.
HN
O O HH
B
HHN
OHHO
transition state
intermediate
How do we design a stable mimic of a transition state?
Page 52
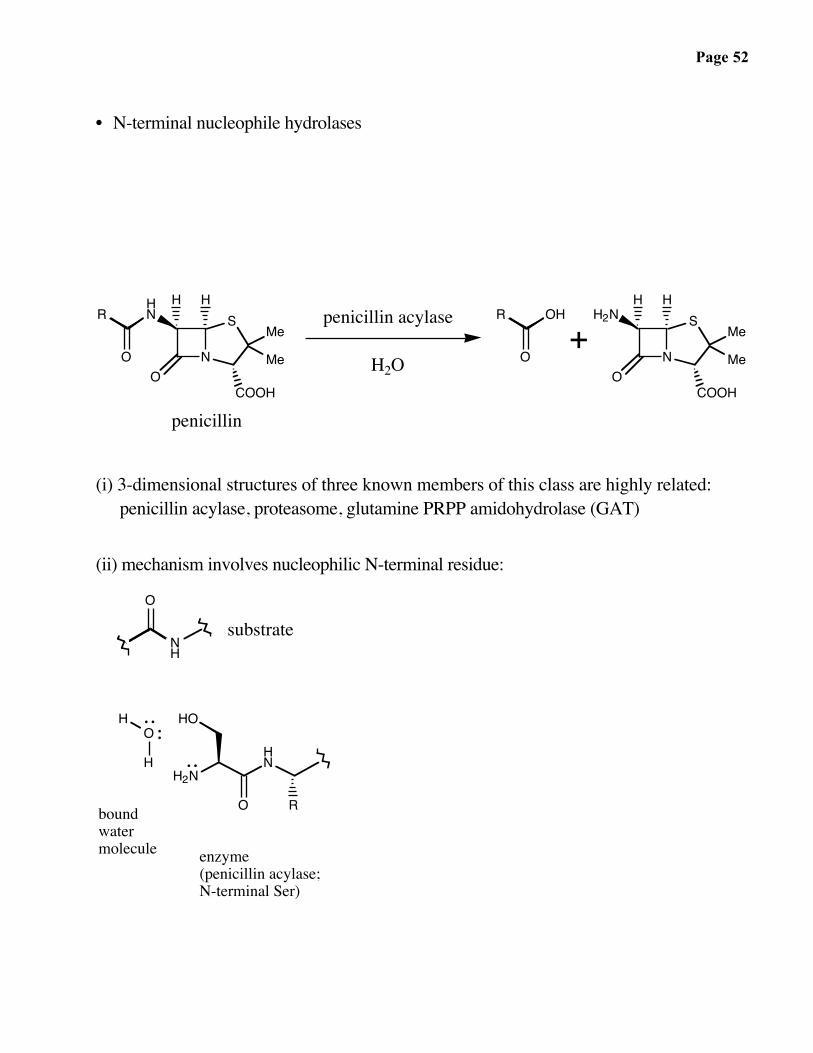
• N-terminal nucleophile hydrolases
HNR
O NO
SHH
Me
Me
COOH
penicillin
penicillin acylase
H2O
H2N
NO
SHH
Me
Me
COOH
OHR
O+
(i) 3-dimensional structures of three known members of this class are highly related:penicillin acylase, proteasome, glutamine PRPP amidohydrolase (GAT)
(ii) mechanism involves nucleophilic N-terminal residue:
NH
O
H2NHN
O
HO
R
O
H
H
substrate
boundwatermolecule enzyme
(penicillin acylase;N-terminal Ser)

Page 53
Page 54
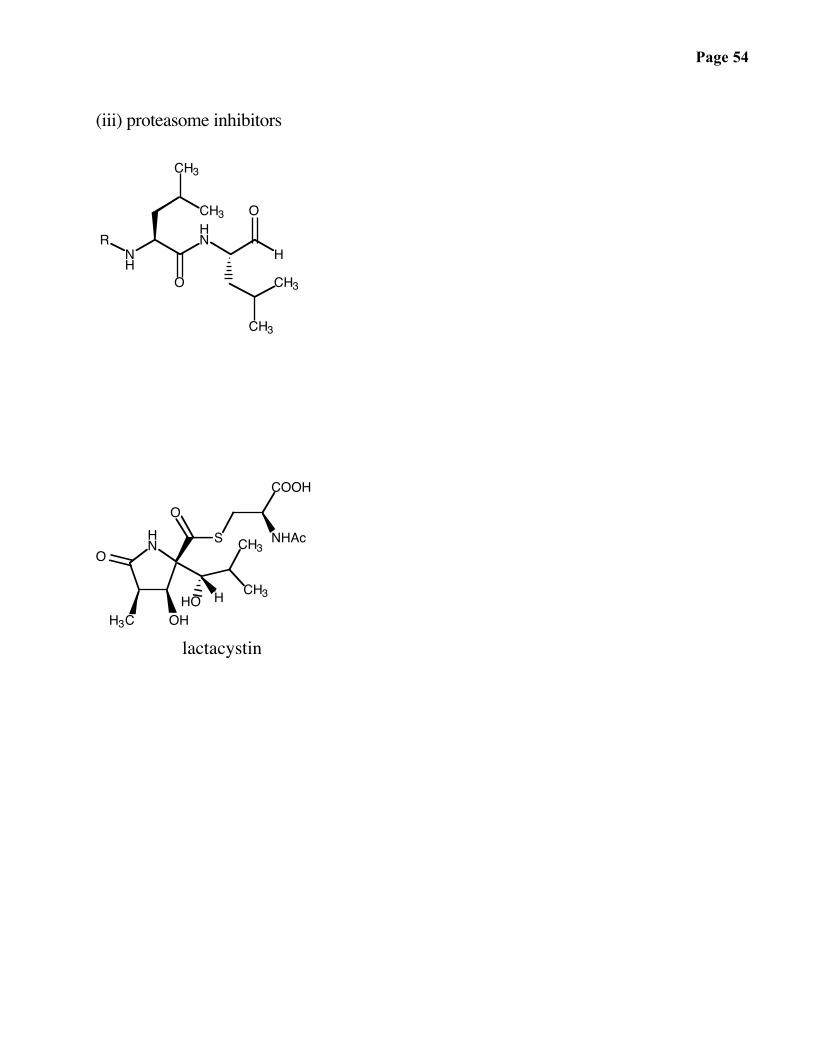
(iii) proteasome inhibitors
RNH
HN
H
O
O
CH3
CH3
CH3
CH3
HN
H3C OH
OCH3
CH3HO
S
COOH
O
NHAc
lactacystin
H

Page 55
(iv) Autoproteolysis of inactive precursor yields active enzyme:
NH
O
OR
HO CH3
OH
O
R
H2N
O
HO
inactive formpreproteasome
CH3autoproteolysis
+
prosequence active proteasome(N-terminal residue)

Page 56
(v) relevance to protein splicing
NH
OHO
O
INTEINNH
HN
O
NH2
O O
HO
extein 2extein 1
(Ser) (Ser)(Asn)
SerThrCys
H3N extein 1 SerThrCys
extein 2 COOAsn
INTEIN
protein splicing(auto splicing)
H3N SerThrCys
NH
O
Ospliced INTEIN
H3N extein 1 SerThrCys
extein 2 COO
Mechanism for the case of serine:
+

Page 57

Page 58
ENZYMESAND
COFACTORS

Page 59
Enzymes and CofactorsThe role of enzymes in metabolic and catabolic pathways
Energy from glucose: two case studies of enzyme-catalyzed reactions
O
OH
OH
HOOHHO H3C
OH
O
O
pyruvic acid
glycolysis
glucoseC6(H2O)6
"carbo hydrate"
Krebs cycle
O2 , mitochondria6 CO2 6 H2O+
production of energy36 ATP generated
2
glycolysis
Triosephosphate isomerase (TIM) was defined by J. R. Knowles as a “perfect enzyme”. The rate ofreaction is controlled by the rate of encounter of substrate with the enzyme (diffusion controlled). Thus,it is impossible to evolve an enzyme that can accelerate the rate further.
Page 60
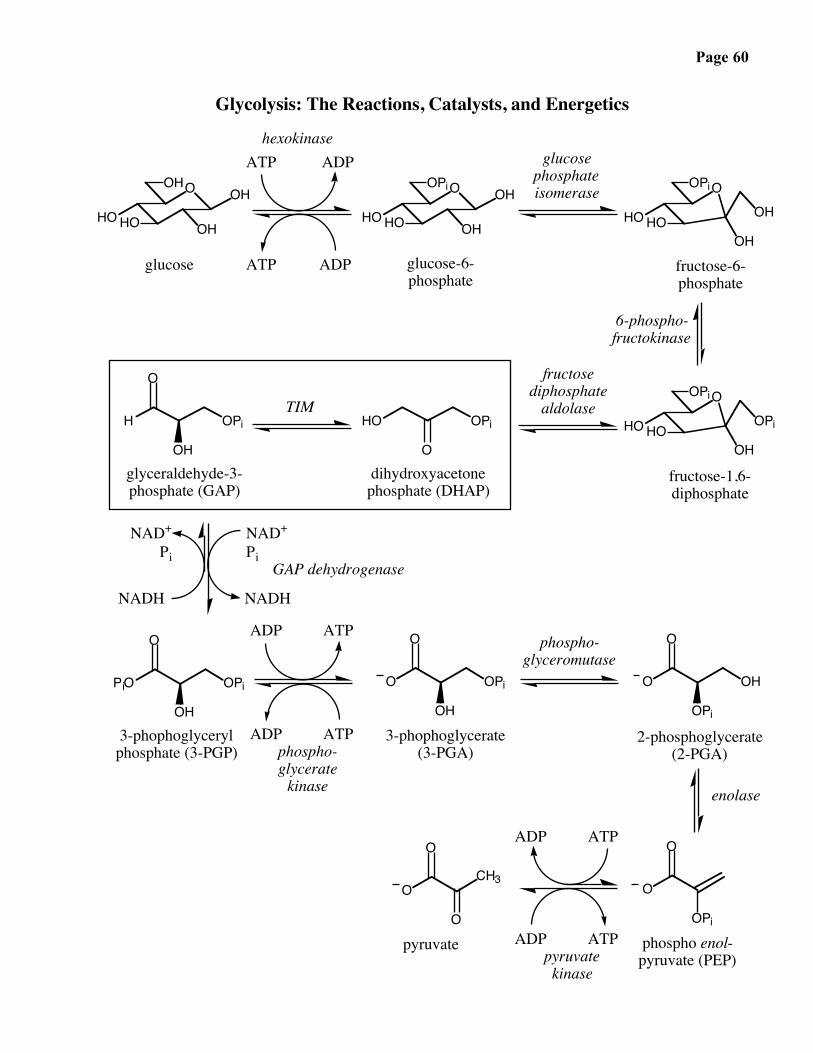
Glycolysis: The Reactions, Catalysts, and Energetics
OOHOH
OHHOHO
OOPi OH
OHHOHO
OOPi
HOHO OH
OH
OOPi
HOHO OPi
OHO
OPiHO
OH
OPiH
O
OH
OPiPiO
O
glucose
OH
OPiO
ADP
ATP
ATP
O
ADP
OPi
OHO
glucose-6-phosphate
glucosephosphateisomerase
hexokinase
O
fructose-6-phosphate
6-phospho-fructokinase
fructose-1,6-diphosphate
fructosediphosphate
aldolase
OPi
O
glyceraldehyde-3-phosphate (GAP)
dihydroxyacetonephosphate (DHAP)
O
3-phophoglycerylphosphate (3-PGP)
3-phophoglycerate(3-PGA)
CH3O
2-phosphoglycerate(2-PGA)
O
phospho enol-pyruvate (PEP)
pyruvateO
TIM
NAD+
Pi
NADH
GAP dehydrogenase
phospho-glyceromutase
enolase
NADH
NAD+
Pi
phospho-glycerate
kinase
ADP ATP
ADP ATP
pyruvatekinase
ADP ATP
ADP ATP
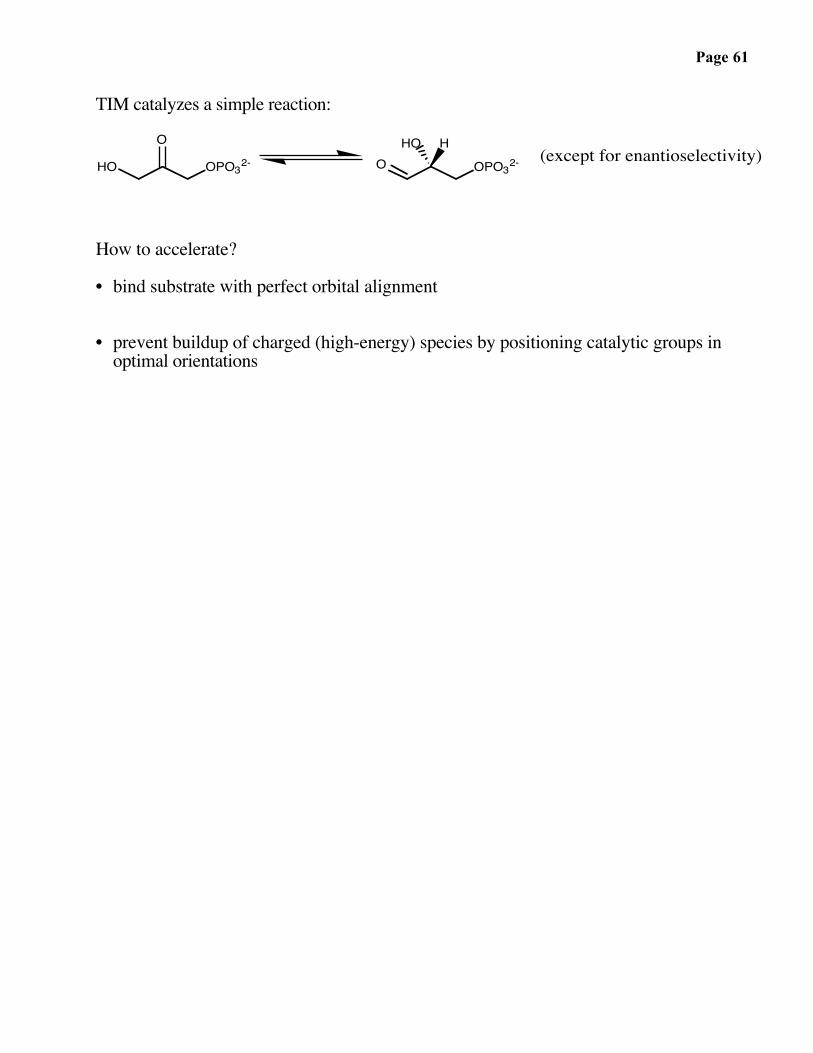
Page 61
TIM catalyzes a simple reaction:
HO OPO32-O
O OPO32-HO H
(except for enantioselectivity)
How to accelerate?
• bind substrate with perfect orbital alignment
• prevent buildup of charged (high-energy) species by positioning catalytic groups in optimal orientations
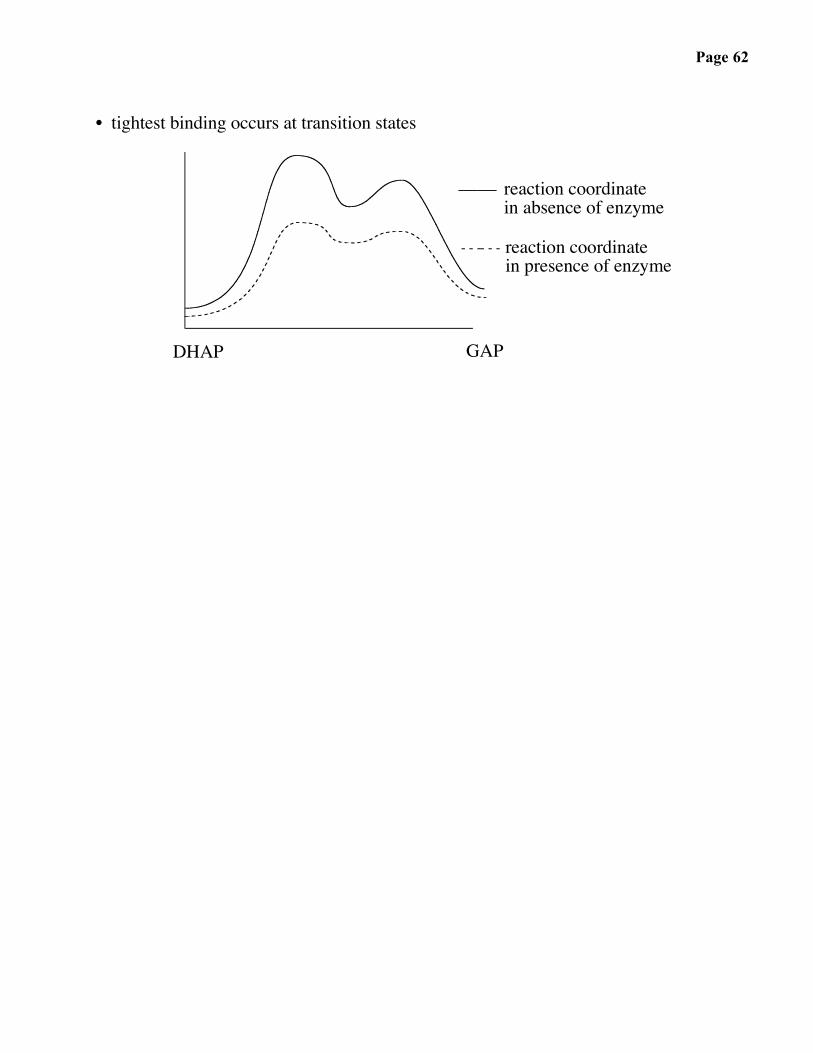
Page 62
• tightest binding occurs at transition states
DHAP GAP
reaction coordinatein absence of enzyme
reaction coordinate in presence of enzyme

Page 63
The Shikimic Acid Pathway
HO
OPO32-
OH
2-O3PO
OCOO
O
OH
OH2-O3PO
HO COO
OH
OHO
HO COO
OH
OHO
COO
OH
OHHO
COO
OH
2-O3PO
COO
OH
OH
O2-O3PO
COO
COO
OH
O
COO
COO
OH
COO
OOCO
tryptophan
D-erythrose-4-phosphatePEP
3-dehydroquinate 3-dehydroshikimate
DHQ synthase
3-deoxy-arabino-heptulosonate-7-phosphate
DAHP synthase
EPSPsynthase
shikimate
5-enolpyruvylshikimate-3-phosphate
DHQ dehydratase
shikimate kinase
shikimate oxidoreductase
shikimate-3-phosphate
+
p-aminobenzoicacid
prephenate
chorismatemutase
chorismatesynthase
chorismate
isoprenoidquinones
tyrosine
phenylalanine
anthranilate
H

Page 64
The Shikimic Acid Pathway
Plants and microorganisms synthesize aromatic amino acids via the shikimic acidpathway:
H3C COOH
O
COO
OP
OP
O
OH
OH
OP OH
OH COO
O
OH
O
OH
PO OH
OHOOC
O
OP
OH
COO
glycolysis
+
HOHO
aldolreaction
(-P = -PO32-)
DAHP
H
2nd enzyme:
DHQ synthase
In contrast to TIM, DHQ synthase uses a cofactor. The emphasis here is on nature’s useof a hidden reaction to trigger a series of non-catalyzed steps.

Page 65
The Mechanism of DHQ Synthase
O
HO O
OH H
OH
OOC
HO
OOC OPO32-O
HO
OH
OPO32-
H
HO
O
HO
OOCOOC
HO
O
OH
O
OH
O
OOC
OH
H
HO
O OOC
HO
O
OH
:B
+ H2PO42-
DHQ
: B
DAHP
NADHNAD+WHERE IS THE BASE COMING FROM?

Page 66
The Design of Mechanistic Probes
O
HO
OH
OOCOH
OP
O
O
O
• How do we know that an oxidation has taken place?
Keeping in mindthat the naturalsubstrate is:
design, synthesize:
O
HO
OH
OOCOH
PO
O
O
• What about elimination? Stereochemistry?
design, synthesize:
O
HO
H
OOCOH
OP
O
O
O
O
HO
H
OOCOH
O
P OO
O

Page 67
• Additional evidence that the phosphate serves as the base…
O
HO
OH
OOC
PO
OO
O
H
O
HO
OH
OOC
O
H
P
O
OO
D2O
D2O

Page 68
• What about the aldol reaction?
design, synthesize:
O
O
OOC
HO
OH
NO
OOH
OOC
HO
OHhv
(non-enzymatic)O
DHQ+

Page 69
Concerted Reactions
Reactant bonds are broken and product bonds are formed at the same time,without the formation of intermediates.
Evidence: lack of detectable intermediates
Example: SN2 reaction
Pericyclic Reactions
Reactions that involve a cyclic shift of electrons (cyclic transition state)
1. Electrocyclic reactions
• ring is formed• one new sigma bond• one fewer pi bonds
2. Cycloaddition reactions
• ring is formed• 2 new sigma bonds• 2 fewer pi bonds
3. Sigmatropic rearrangement
• allylic sigma bond at one end of a pi electron system appears to migrate to theother end
• total number of pi bonds remains unchanged

Page 70
The Claisen Rearrangement
OH
O COO
COO
OH
OOC
COO
O
• cellular
O O
Δ• laboratory
chorismate
mutase
Phe
Tyr
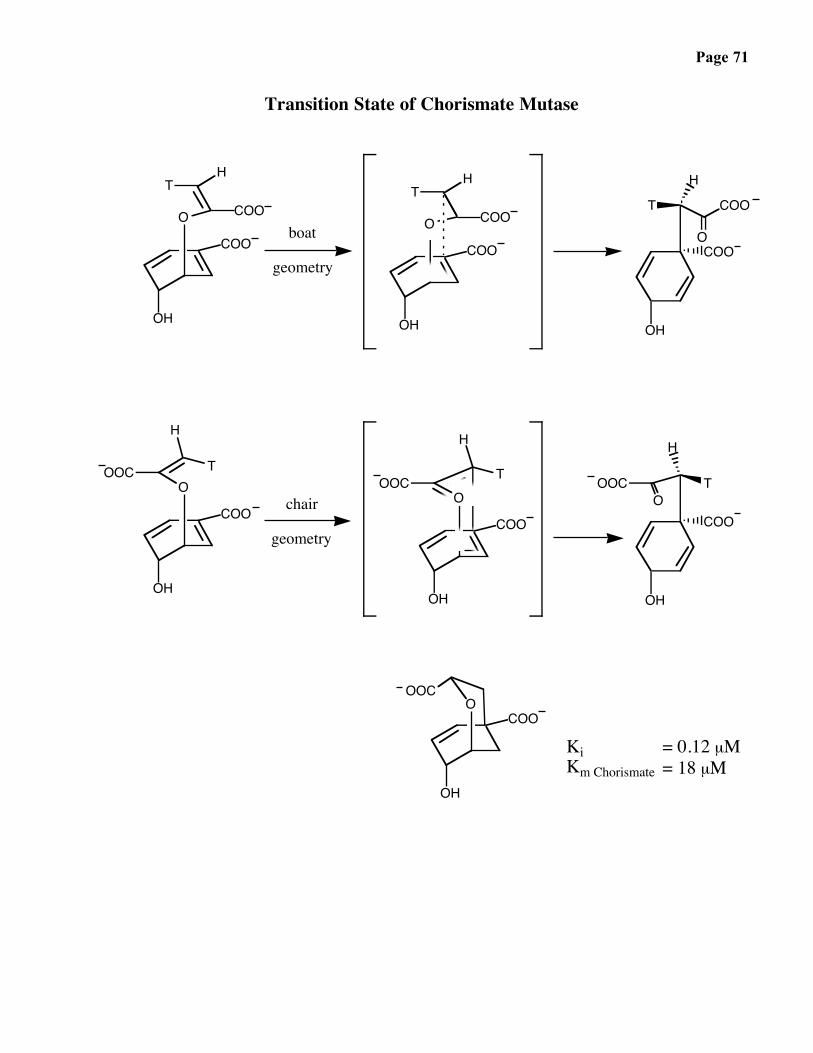
Page 71
Transition State of Chorismate Mutase
OH
COO
O COO
TH
OH
COO
O COO
TH
OH
T
H
COO
COO
O
OH
COO
O
OH
COO
O
OH
COO
TOOCO
OOCOOC
T T
HHH
OH
COOO
boat
geometry
OOC
chair
geometry
KiKm Chorismate
= 0.12 µM= 18 µM

Page 72
Catalytic Antibodies
Can we make an artificial enzyme that catalyzes a reaction of our choosing?
Can we generate a protein that stabilizes a transition state of our choosing?

Page 73
Cofactors: Nature’s Reagents
Protein sidechains lack: -
-
Me Me
H OH
Me Me
O• consider the reactions: + 2 e-, 2 H+
S SHN NH
SHHN+ 2 e-, 2 H+
• the only reasonable electron acceptor among the 20 natural amino acids:
2O O O
Me Me
H OH
Me Me
O• problem: the redox potentials are mismatched:
+ RS-SR
Therefore, there is too muchwasted energy, and the carbonylcannot be reduced.
+ 2 RSH(a weak reducing agent)
Me Me
H OH
N
H O
NH2
Me Me+
N
O
NH2++
O
H H• as we will soon discuss, nature solved this problem with the NAD+ / NADH system:
NAD+ NADH
matchedpotentials
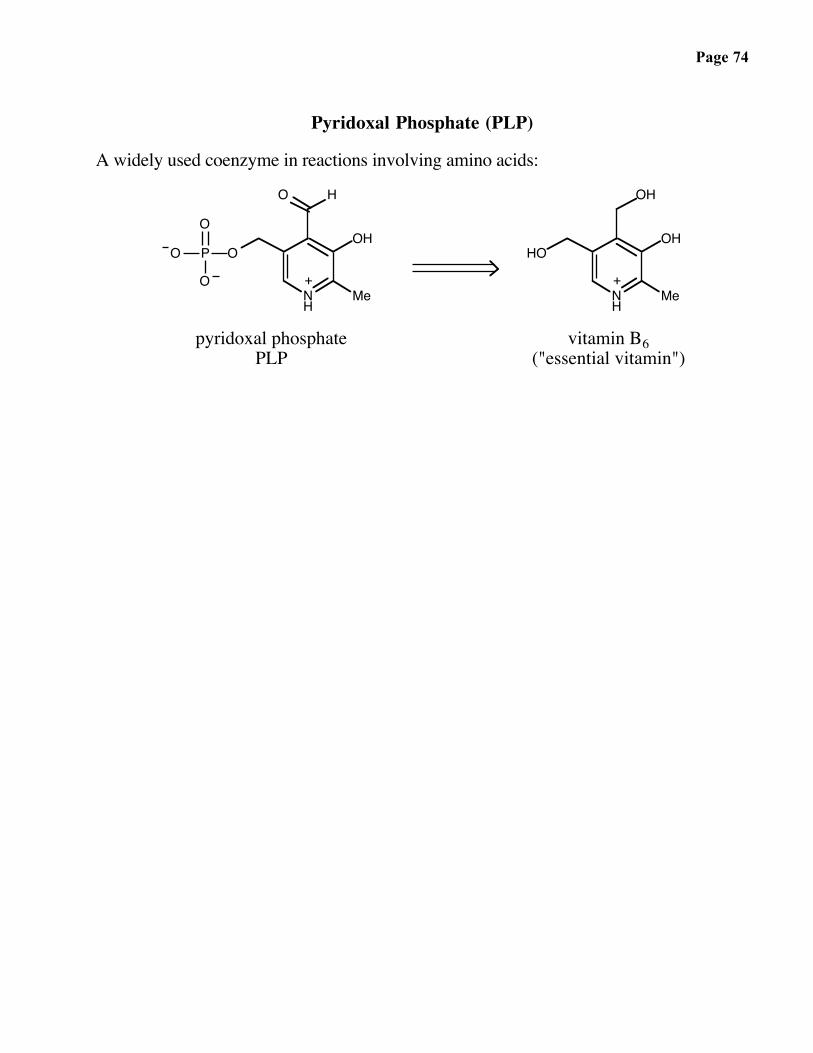
Page 74
Pyridoxal Phosphate (PLP)
A widely used coenzyme in reactions involving amino acids:
NH
HO
OP
O
O
O
OH
Me NH
OH
HOOH
Me
pyridoxal phosphatePLP
vitamin B6("essential vitamin")
++

Page 75
Summary of PLP-Catalyzed Reactions on α−C: (α−H loss)
NH
HC
CCH2CON
HO
H
O
COO
H
H2C
N
C
HCO
NH
NH
HC
CCH2CON
O
H
O
H3N CO2
CH2OH
CH2OH
CO2H3NCH2OH
CO2O
H2CNH2
NH
+
.. ..
+
+
or+
+
COO
H
H2C
N
C
HCO
NH
H
racemization
hydrolysis(or aminolysis)
transamination
α-H loss
COO
H
H2C
N
C
CH2
O
NH
HCO
NH
+
hydrolysis
PLP(or PLP-Enz)
PAP
OH
OH
Me
Me
O
O
P
P
OH
Me
O
OH
Me
O
OH
Me
O
OOH
Me
P
P
P
OH
Me
OP
α Cprotonation
"early"protonation
P

Page 76
Summary of PLP-Catalyzed Reactions on α−C: (sidechain loss)
NH
HC
CCH2CON
HO
H
O
CO
N
C
HCO
NH
NH
HC
CCON
O
H2CNH2
NH
+
.. ..
+
CO
N
C
HCO
NH
H
hydrolysis(or aminolysis)
sidechain loss
CO
N
C
CH2
O
NH
HCO
NH
+
hydrolysis
PLP(or PLP-Enz)
PAP
OH
OH
Me
Me
O
O
P
P
OH
Me
O
OH
Me
O
OH
Me
O
OOH
Me
P
P
P
OH
Me
OP
α Cprotonation
"early"protonation
H HCH2
O
CH2
O
H
H3N CO2
H H
amino acidmetabolism
H
O CO2P
H
amino acidmetabolism

Page 77
Summary of PLP-Catalyzed Reactions on α−C: (decarboxylation)
NH
HC
CCH2CON
HO
H
O
O
H
H2C
N
C
HC
NH
NH
HC
CH2C
N
O
H
H2CNH2
NH
+
.. ..
+
+
H
O
H
H2C
N
C
HC
NH
H
hydrolysis(or aminolysis)
O
H
H2C
N
C
CH2
NH
HCO
NH
+
hydrolysis
PLP(or PLP-Enz)
PAP
OH
OH
Me
Me
O
O
P
P
OH
Me
O
OH
Me
O
OH
Me
O
OOH
Me
P
P
P
OH
Me
OP
α Cprotonation
"early"protonation
P
CO2 lossH H
H
CH2OH
CH2H3N
simpledecarboxylation
CH2OH
O H
oxidativedecarboxylation

Page 78
Question. How does an enzyme direct which bond will be cleaved?
• stereoelectronic effects
HR2
R1
HN
rotate
bind
+
R3
N
OHH
Me
2-O3POCH2

Page 79
Formation and Turnover of a Catalytically Active Enzyme-PLP Complex
NH
CH3
lys
NH2
OH OH
OHO
H
NH
NH
NHH
lysN
Sub
EnzH
NH
H
NH
SubNH2
NHH
Prod
Enz
ProdNH2
.
+
+
+++
The enzyme uses a transiminationmechanism to bind and releasesubstrates and products becausethe activation barriers are relativelylow. Contract this to a hypotheticalaldehyde-imine route, which would proceed via the sequence A → B → D → E → B → A, and which would have two high barriers per turnover. These high barriers would slow the enzymatic reaction down, and thus the enzyme has evolved to avoid such a route.
Because of the high C → B barrier, the enzyme undergoes many turnovers (C → D → E → C or C → E → D → C)before becoming inactivated owing to loss of the cofactor. When the cofactor is lost, however, reactivation of the enzymeis slow (much slower than turnover) because of the high A → B barrier.
CEDC
B
A
rxn. coordinate (note: not all intermediates and/or transition states are shown)
ΔG
The active form of the enzyme possesses an imine linkage between an active site lysine residue and the PLP cofactor (form C). The binding of the substrate to the enzyme proceeds via a transimination reaction(form C → form D) in which the covalent bond between the enzyme and cofactor is broken and replaced by acofactor-substrate imine bond (form D). The trans-imination reaction does not proceed via high-energyintermediates. Form D then undergoes the conversion steps to generate the product imine E. The product is then released via transimination with the active sitelysine, resulting in regeneration of the catalytically activeform of the protein (C).
Turnover phase
Preparation phase
... H-B
This phase involves the preparation of active enzyme. The noncovalent enzyme-PLP complex (forms A and B) is unable to catalyze the conversion of substrate toproduct ("inactive"); only the covalent complex, which has an imine linkage between the cofactor and a lysine residue of the protein (form C), is active. The enzymecatalyzes all steps shown in this scheme. Form A,being an aromatic aldehyde, presumably requiresprotonation (form B) in order to condense with the activesite lysine residue (to give form C). Form B is a high-energy intermediate.
conversionsteps
E
D
CBA
PLP-productimine
covalent product-cofactor complex
+
lysineresidue
on enzyme
PLP-lys iminecovalent enzyme-cofactor complex
+
PLP-substrate imine
covalent substrate-cofactor complex
PLP
+
Not sufficiently electrophilic toreact directly with amines; requires activation (protonation).
+
P
substrate

Page 80
Examples of PLP-Catalyzed Reactions
• bacterial cell wall: A highly cross-linked glycoprotein that serves as an essentialprotective layer:
O
O
O
O
O
O
O
O
O
O2C CO2
NH3 NH3
meso DAP
polypeptide
carbohydrateO O O
usedforcross-linking
NH
HN
R
O R
O fungalproteases
hydrolysis
NH
HN
R
O R
O
no reaction
D-amino acid
fungalproteases
Predators could in principle break down the cell wall using proteases. Bacteria foil thisstrategy by using D-amino acids.
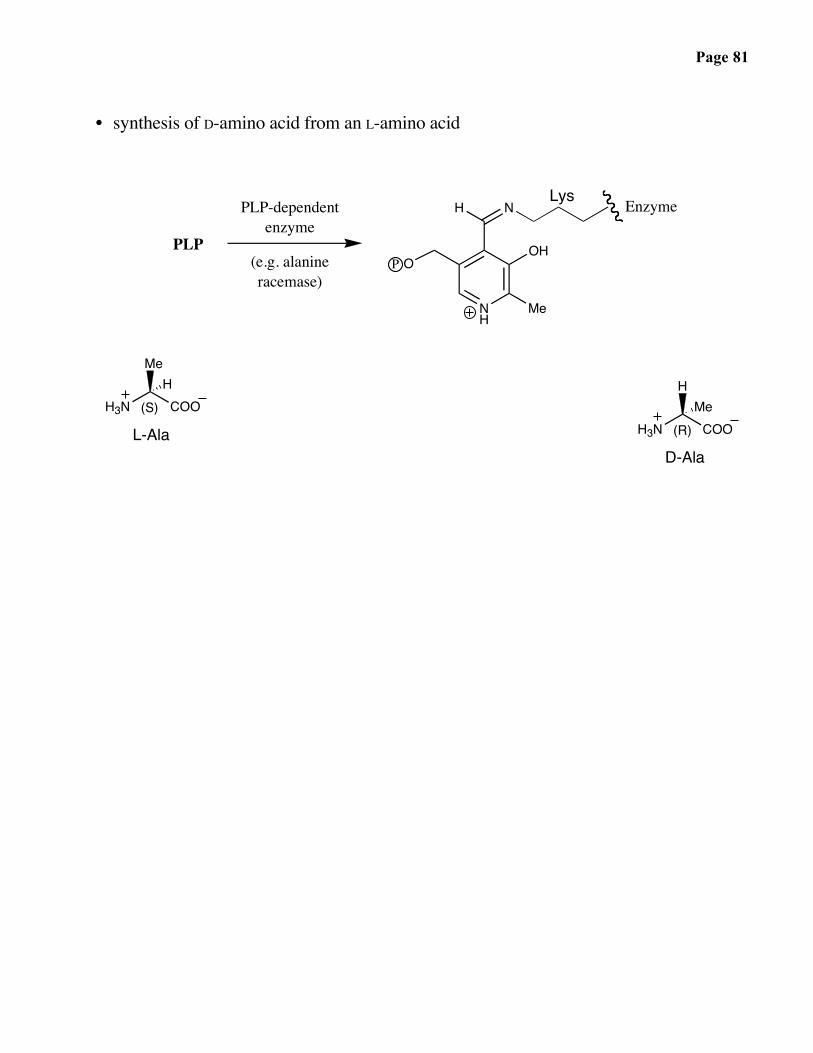
Page 81
• synthesis of D-amino acid from an L-amino acid
NH
OH
Me
O
N
H3N COO
Me
PLP-dependentenzyme
(e.g. alanineracemase)
PLP
H Enzyme
P
Lys
H
L-Ala(S)
H3N COO
HMe
D-Ala(R)

Page 82
Reaction Coordinate Diagram of Racemization
NH
NH
Me COO-H
+
NH
NH
Me COO-
+
H
H3N+ COO-
MeFree (S)-Ala
H3N+ COO-
Free (R)-AlaMe
++
NH
N
Me COO-
H3N+
Me
O-
OHEnergy
Reaction Coordinate
(R)-alanine-PLP adduct
(S)-alanine-PLP adduct
Deprotonatedalanine-PLPintermediate
HIGH ENERGYTRANSITION STATE
uncatalyzed reaction

Page 83
Oxidation-Reduction Examples Involving Transfer of Electrons
Me Me
O
Me Me
NH
MeMe
H2N H
Oxidation number of carbon atoms
• assign –1 for every negative charge• assign –1 for every bond to a less electronegative atom
• assign 0 for every lone pair• assign 0 for every bond to carbon
• assign +1 for every positive charge• assign +1 for every bond to a more electronegative atom
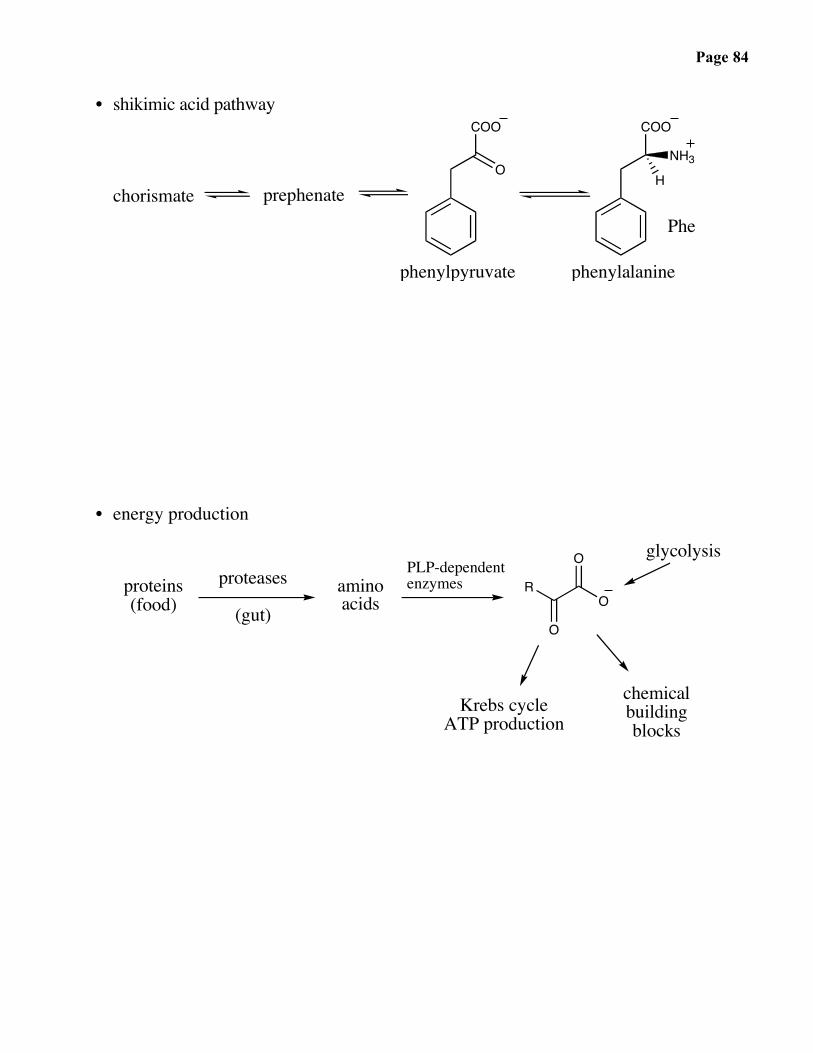
Page 84
• shikimic acid pathwayCOO
O
COO
H
NH3
Phechorismate prephenate
phenylpyruvate phenylalanine
• energy production
R
O
O
Oproteins(food)
proteases
(gut)aminoacids
chemicalbuildingblocks
Krebs cycleATP production
PLP-dependentenzymes
glycolysis

Page 85
• preventing buildup of lactic acid
When the heart muscle is overworked, the normally oxidizing milieu becomes reducingdue to a lack of oxygen, resulting in the conversion of pyruvic acid to lactic acid:
glucose pyruvateMe COOH
OHlactic acid
Lactic acid causes muscle pain due to acidification and can cause tissue damage.
To prevent pain or damage, aspartate transaminase comes to the rescue by removingpyruvate (and thus lactic acid).
• aspartate transaminase
Me CO2
O
H3N CO2
CO2H
+AT
PLPH3N CO2
Me + OOH
O
O
O
Asp Ala oxaloacetatepyruvate
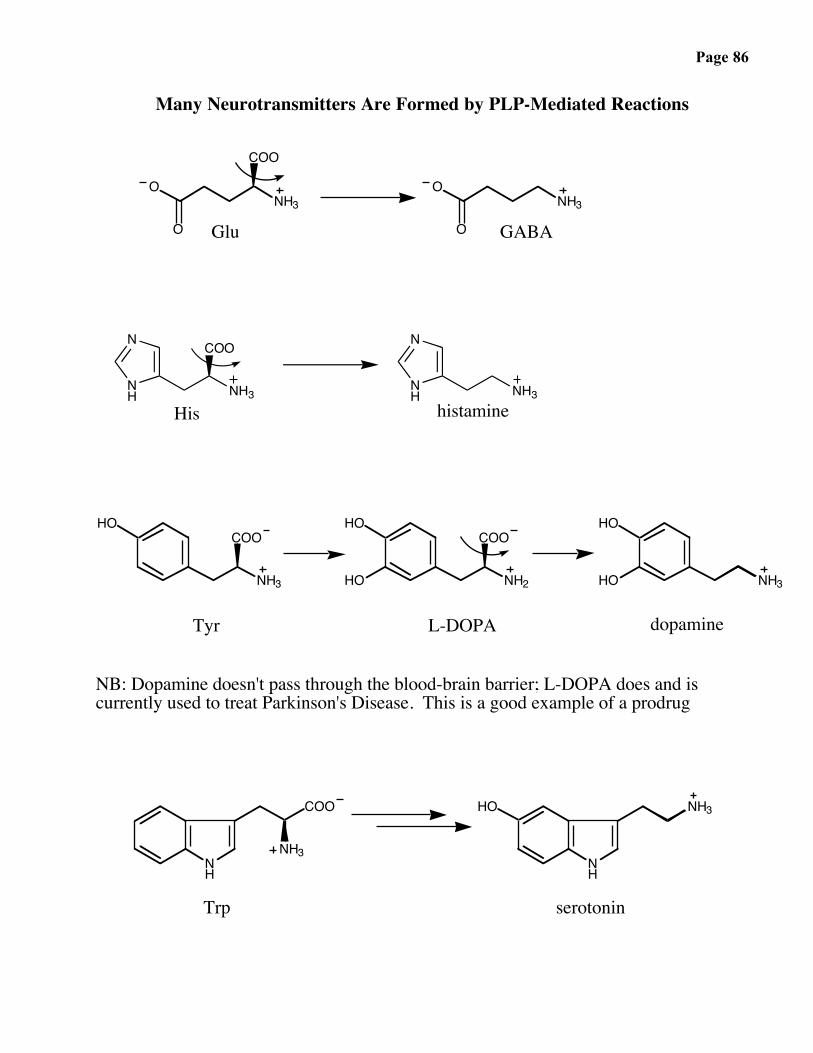
Page 86
Many Neurotransmitters Are Formed by PLP-Mediated Reactions
ONH3
O
COO
ONH3
OGlu GABA
NH3
COON
NH NH3
N
NH
His histamine
NH3
COOHO
NH2
COOHO
HO NH3
HO
HO
Tyr dopamineL-DOPA
NB: Dopamine doesn't pass through the blood-brain barrier; L-DOPA does and iscurrently used to treat Parkinson's Disease. This is a good example of a prodrug
NH
NH3
COO
NH
NH3HO
serotoninTrp
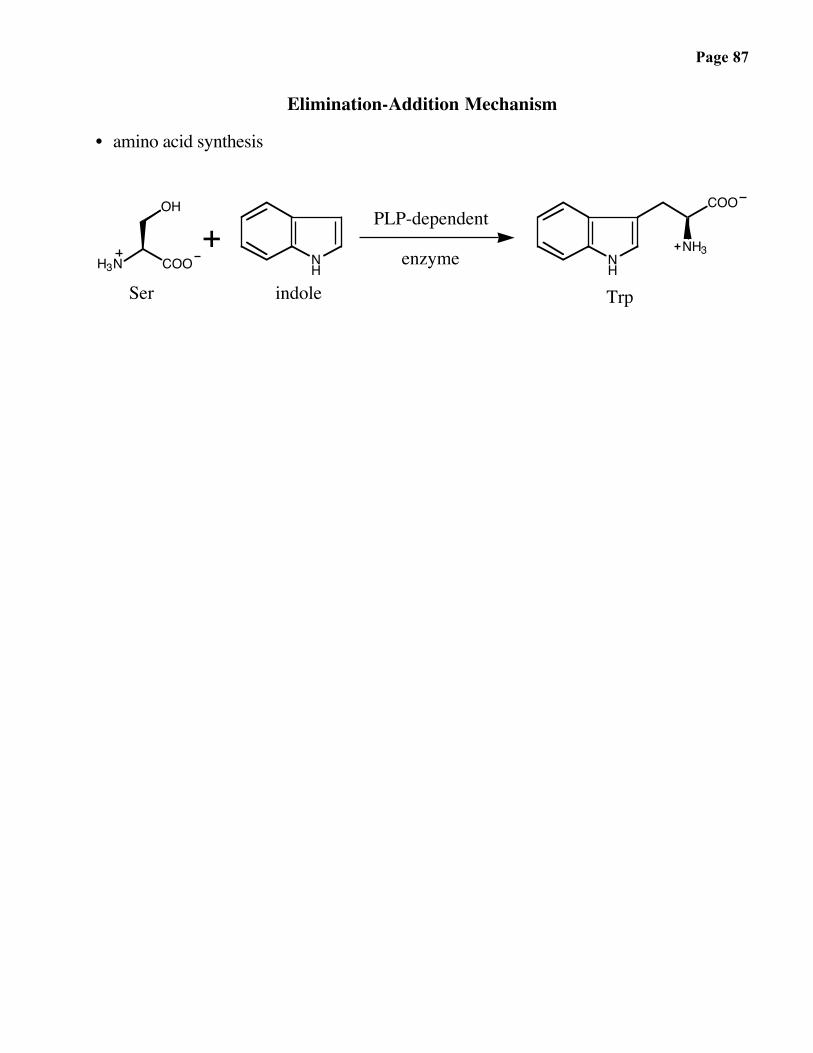
Page 87
Elimination-Addition Mechanism
• amino acid synthesis
NH
NH
COO
NH3H3N COO
OH
indole
PLP-dependent
enzyme
Trp
+Ser

Page 88
Inhibitors of PLP-dependent Enzymes
Inhibitors of PLP-dependent enzymes have found numerous medical applications. Theyhave been discovered by considering the mechanisms of the enzyme-catalyzed reactions.
e.g. Suicide inhibitors: stable compounds that become hyperreactive at theenzyme active site.
Ornithine decarboxylase catalyzes the PLP-dependent conversion of ornithine intoputrescine:
Putrescine is used by the cell to package the one-meter long nuclear DNA. Thedicationic form can neutralize the negative charge associated with DNA’s phosphategroups.
DFMO: a rationally designed anticancer (gout) agent
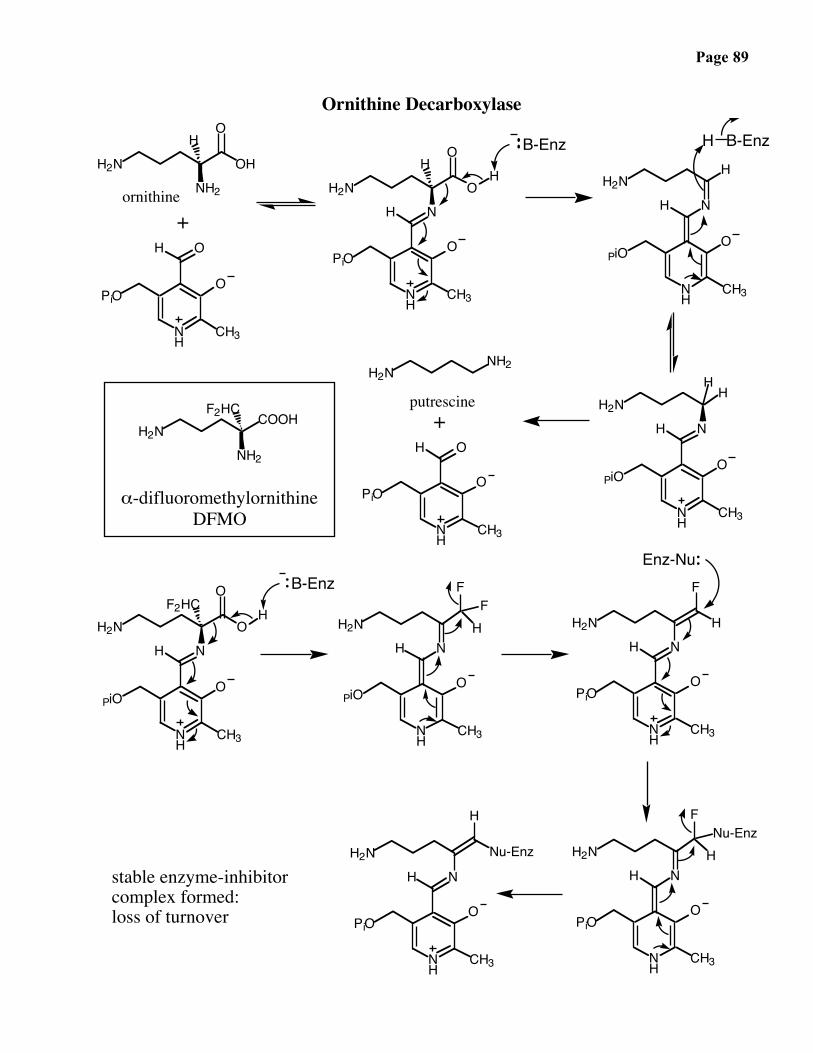
Page 89
Ornithine Decarboxylase
H2N
O
OHNH2
NH
OH
PiOO
CH3
+
H
NH
NH
PiOO
CH3
H2N
O
OH
HB-Enz
NH
NH
PiOO
CH3
H2NH
H B-Enz
NH
NH
PiOO
CH3
H2NH
H
NH
OH
PiOO
CH3
H2NNH2
+H2NCOOH
NH2
F2HC
α-difluoromethylornithineDFMO
putrescine
ornithine
NH
NH
PiOO
CH3
H2N
O
OH
F2HCB-Enz
NH
NH
PiOO
CH3
H2N
NH
NH
PiOO
CH3
H2N
FFH
NH
NH
PiOO
CH3
H2N
F
H
Enz-Nu
NH
NH
PiOO
CH3
H2N
FNu-EnzHNu-Enz
H
stable enzyme-inhibitorcomplex formed:loss of turnover
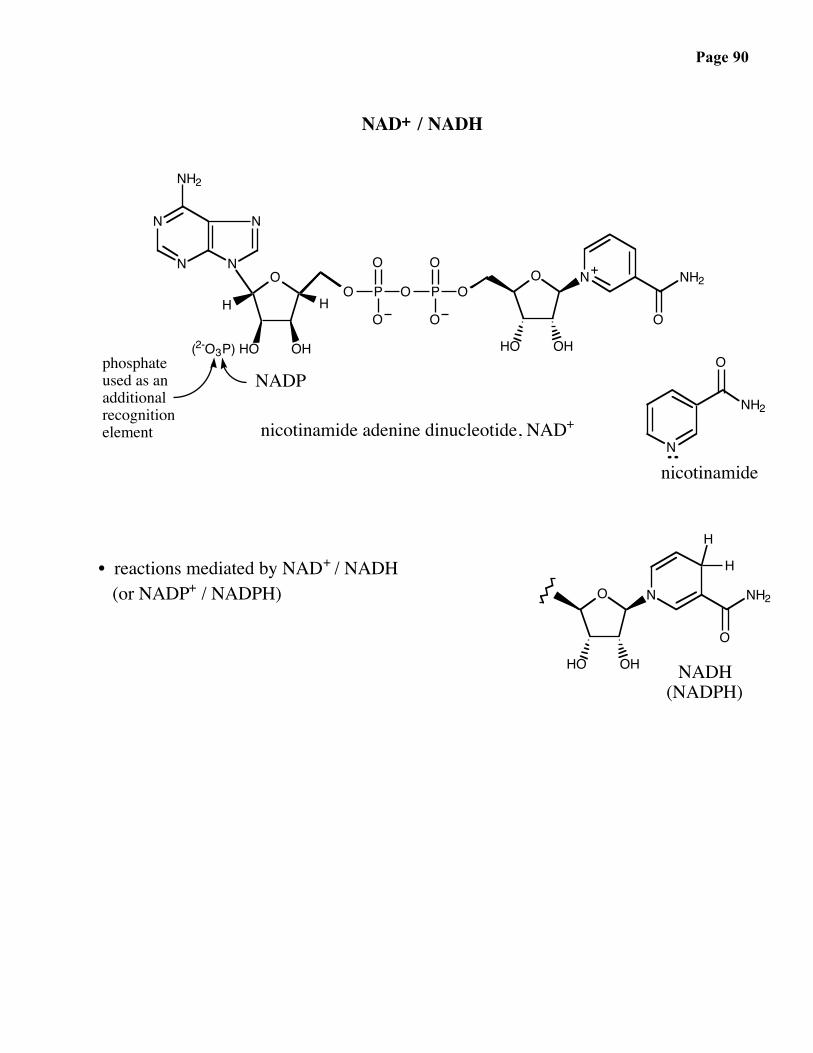
Page 90
NAD+ / NADH
O
OH(2-O3P) HO
H HO P
O
O
O P
O
O
OO
HO OH
N NH2
O
N
N N
N
NH2
N
O
NH2
O
HO OH
N NH2
O
H
H
phosphateused as anadditionalrecognitionelement
+
NADP
nicotinamide adenine dinucleotide, NAD+
nicotinamide
NADH(NADPH)
• reactions mediated by NAD+ / NADH (or NADP+ / NADPH)

Page 91
Summary of Reactions Mediated by NAD+ / NADH or NADP+ / NADPHC. T. Walsh, Enzymatic Reaction Mechanisms, Freeman, third edition.
H3CC
CH3
H OH
H3CC
CH3
O
H3CC
CH3
H +NH3
H3CC
CH3
O
H3CC
C
H3C OHCO2
HHH3C
CC
CO2
HH3C
O
H3CC
CH3
O
H3CC
C
H3C HCH3
CH3H
H3C
H3C
CH3
CH3
H3CC
N
H3C HCH3
CH3H
NH3C
H3C
CH3
CH3
2 e-
+ NH4+ + H+
+ 2 H+ +
+ 2 e-
+
RC
OR'
H OH
RC
OR'
O
malate dehydrogenaselactate dehydrogenasealcohol dehydrogenase
glutamate dehydrogenase
CO2isocitrate dehydrogenase6-phosphogluconate dehydrogenasealdehyde dehydrogenase
+
1
2 H+ + 2 e- dihydrosteroid dehydrogenase(steroid reductase)
CHR
O
2
3
4
5
P
O
OHO
O
+ 2 H+ + 2 e- dihydrosteroid dehydrogenase(Steroid reductase)
6
+
COR
O
2 e-+ +2 H+
P
O
O
O
glucose oxidase
7 + glyeraldehyde-3-phosphate dehydrogenase
ExamplesReaction typeCategory
2 e-+ +2 H+

Page 92
Alcohol Dehydrogenase
S
NH
O
H
N
NH
O H
H
S
H
HN
O
• enzymes (e. g. yeast alcohol dehydrogenase) use a second essential cofactor: Zn2+
HisNH R
ONH2
Zn2+CH3CH2OH

Page 93
The now classic experiements of F. Westheimer revealed that the reaction isstereospecific at both the alcohol and the nicotinamide ring. As both the reagent(NAD+) and the substrate (CH3CH2OH) are “achiral”, the results illuminated thestereospecific manner in which enzymes interact with their substrate.
N
O
NH2
R
N
O
NH2
R
D H
H3C
O
D
H3C
O
H
++ CH3CD2OH
YADH
YADH
+
N
O
NH2
R
H H
H3C
O
D+ YADH
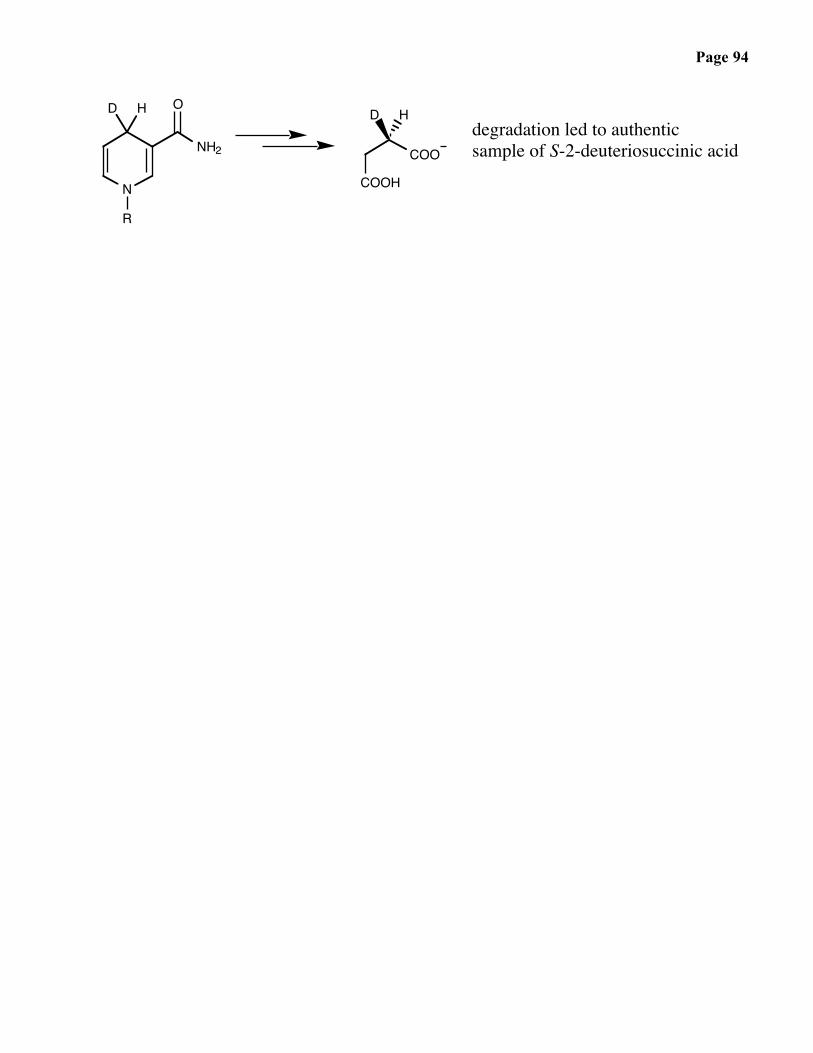
Page 94
COOH
COO
HDdegradation led to authenticsample of S-2-deuteriosuccinic acid
N
O
NH2
R
D H

Page 95
FAD / FADH2
N
RN
NH
N O
O
Me
Me
NH
RN
NH
HN O
O
Me
Me
N
N
NH
N O
O
Me
Me
O
OH
OH
OH
O
N
N N
N
NH2
HO OH
+ 2H+, 2e-
FADFMN
FADH2FMNH2
• flavins are versatile redox enzymes: Me
S
riboflavinvitamin B2
NH
flavin adeninedinucleotide
FAD
H
reducedribose
flavin mononucleotide FMN
flavoproteins bind eitherFAD or FMN, sometimescovalently:
P
O
O
OPO
O
O
O
• FADH2 / NAD+ couple

Page 96
• FADH2 / O2 couple
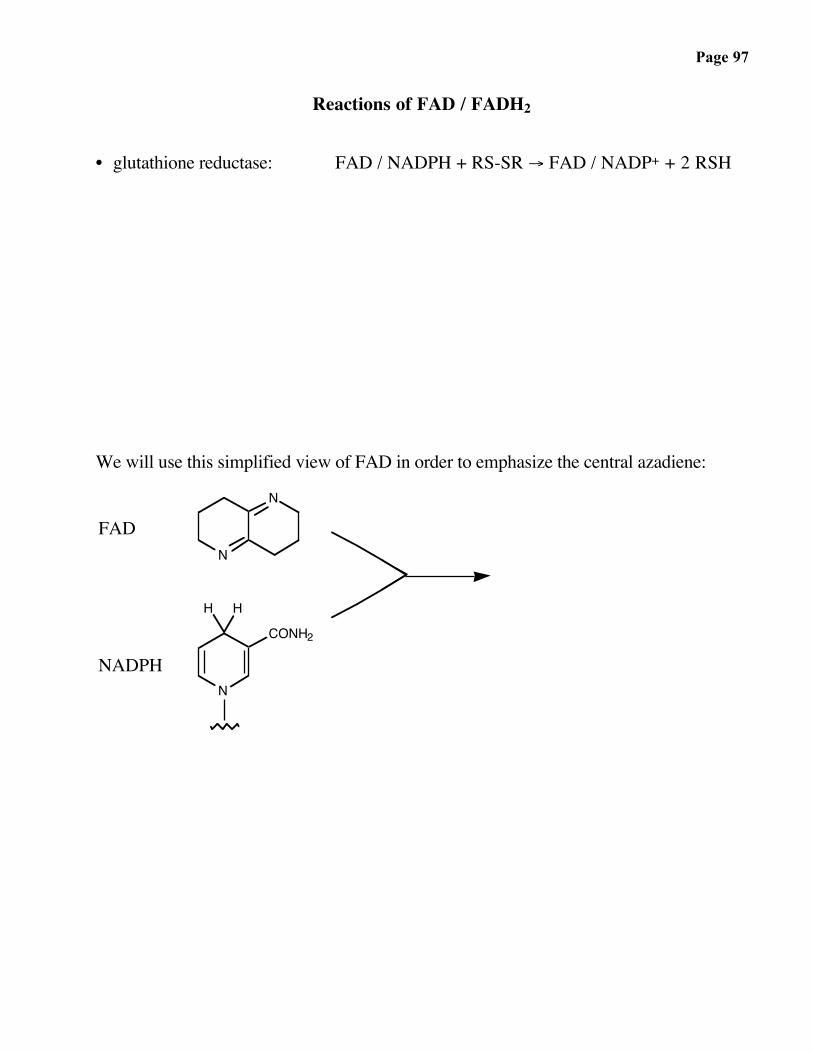
Page 97
Reactions of FAD / FADH2
• glutathione reductase: FAD / NADPH + RS-SR → FAD / NADP+ + 2 RSH
We will use this simplified view of FAD in order to emphasize the central azadiene:
N
N
N
H H
CONH2
FAD
NADPH
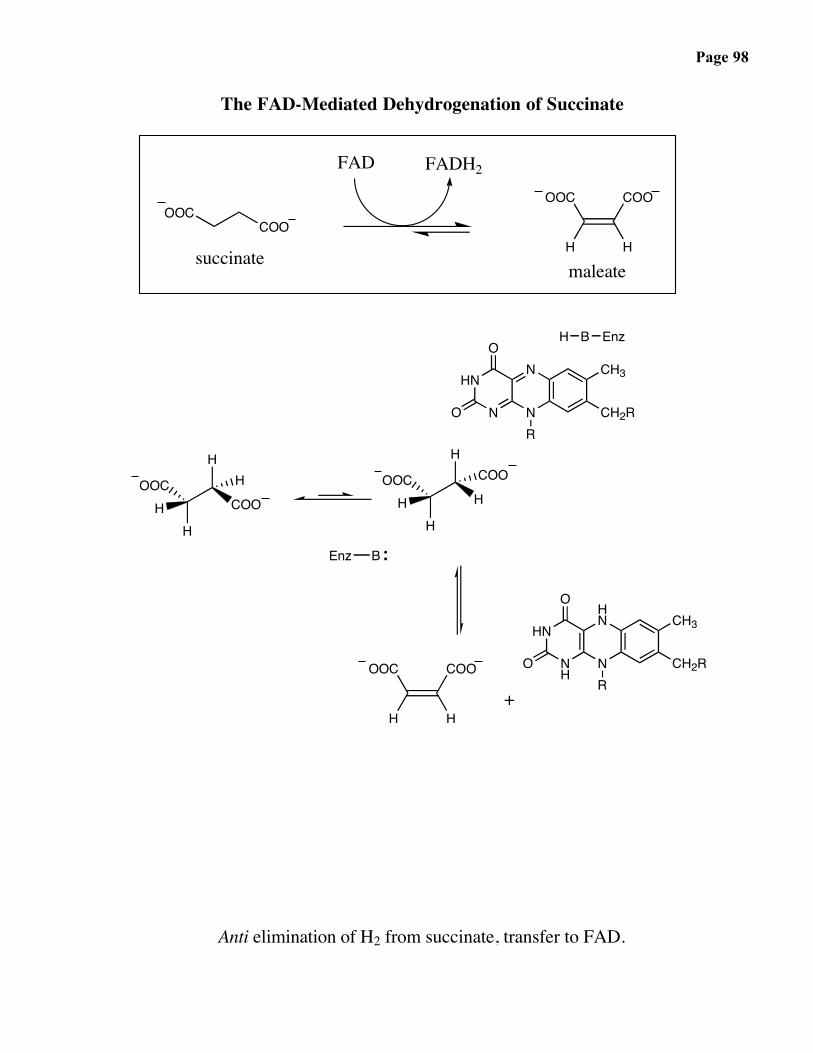
Page 98
The FAD-Mediated Dehydrogenation of Succinate
OOCCOO
N
N
HN
N
CH3
CH2RR
O
O
H B Enz
Enz B
HN
N
HN
NH
CH3
CH2RR
O
OOOC
H
COO
H
succinatemaleate
FADH2OOC
H
COO
H
+
FAD
OOC
HH
HHCOO
OOC
HH
HCOOH
Anti elimination of H2 from succinate, transfer to FAD.

Page 99
Reactions of FAD / FADH2
• ketone monooxygenase
O
O
O
FAD
H+
enzyme
+ H2O + NADP+H+ + O2 + NADPH +

Page 100
• lactate oxidase
H3C
OH
H3C OH
OFAD
enzyme+ CO2 + H2O O2 + OH
O

Page 101
ATP
O
N
N N
Nnegative charge yieldskinetic stability, yetP-O-P linkage yields ahigh-energy bond
NH2
HO OH
OP
O
O
OP
O
O
OP
O
O
O
• sites of reactivity
• reactions
• stereochemistry

Page 102

Page 103
Mechanism and Stereochemistry of Phosphate Attack
2 possibilities
(1) SN2-like, in-line displacement
(2) Addition – Elimination

Page 104
ATP: The Most Widely Used Energy Currency in Living Systems
chemistry: phosphoryl transfer reactions
αβγ
Ado
ON
N
HO OH
ON
N
NH2
PO
OOP
O
OOP
O
OO
HO
possible displacement reactions at P:
a)γ β α
+
γ β αb) +
c)γ αβ
+
βγ α+d)
αβγe)
γ β αf)
Nu
+
+
γ β αPHOO
OO P
O
OO AdoO
PHOO
OO P
O
OO P
O
ONu
OP
AdoO
O
OHO O-AdoP
O
OOP
O
ONu
NuPO
OOP
O
OHO PO
O
OO-Ado
O-AdoPO
OOP
O
OO
PO
ONu
PO
ONu
PHOO
ONu PO
O
OO P
O
OO-Ado
PHOO
OO P
O
OO P
O
OO-Ado
γ β αPHOO
OO P
O
OO P
O
OO-Ado
γ β αPHOO
OO P
O
OO P
O
OO-Ado
γ β αPHOO
OO P
O
OO P
O
OO-Ado
γ β αPHOO
OO P
O
OO P
O
OO-Ado
γ β αPHOO
OO P
O
OO P
O
OO-Ado
b, c, and e have not been observed to date.d is rarely observed but important.a and f are the most commonly observed.

Page 105
Displacement Reactions Involving ATP:
2terpenesisoprenoidscholesterol. etc
geranyl pyrophosphate
+
γ β α
γ
β α+
αβγ
γ γ + αβ
γγ
Phosphoryl transfer:
dimethylallyl pyrophosphate
2
isopentenylpyrophosphate
mevalonate
O P OPO
OO
O
OO P OHP
O
OO
O
O
O-PPi
O-PPi H O-PPi
OOCO
PO
OH
OO
O P
H3C
O
OPOH
P O P O-Ado
OO-Ado
O
O P
O
OO
O
H3C
OP OO
O
OH
OOOC
O
OPO
OO-AdoO
PO
O
O
OOCO P
O
OH
OO
H3C
P O P O-AdoO
O
O
OO
PO
OO
O
PO
OOP
OO
O
+
+
+
B:

Page 106
Another example of phosphoryl transfer:
αβγβ α
glucose
B :
glucose-6-phosphate
γ
O
HOHO OH
OHO
P O P O-AdoO
O
O
OOP
OO
O
O
O PP
OO
OO
PO
OO-AdoOH O
OH
OHHO HO
O
+
Phosphorylation of glucose is the first step in glycolysis.
+
nicotinamide ribonucleotide
βα γ α
nicotinamide adenine dinucleotide (NAD)
+
γβ
N
O
OHHO
O
NH2
O
OPO
O
OAdo
PO
OO
P O
PO
OOP
O
P
PO
O
OO
O
O
OOH
O
O
Ado
O PO
OO
O
NH2
NO
HO OH
+
O
Nucleotidyl transfer:

Page 107
:B
phosphoribosyl pyrophosphate (PRPP)
β αγ
P O
HO OH
O
OH
O P OP P O-AdoO
O
O
OO
OO
OO
OHHO
P O +
Pyrophosphoryl transfer:
PRPP is a biosynthetic precursor to a broad range of biological compounds, includingnucleotides (used in DNA, RNA and cofactors) and several amino acids. This highlightsanother important aspect of phosphate chemistry -- pyrophosphate and phosphate aregood leaving groups for substitution reactions at carbon.
(PRPP)
..
nicotinamide +
nicotinamide ribonucleotide
ΔG˚' =-7 kcal/mol
2
+
P O
HO OH
O
O
PO
OO P
O
OO
N
NH2
O O
NH2
N
O P OHPO
OO
O
O
O
O P
OHHO
P O
O
OOH
Note also:• Doubly linked diphosphates such as NAD never arise via path c.• FAD biosynthesis is similar.• RNA synthesis involves nucleotidyl transfer to the 3'-OH of preceeding
ribonucleotide unit.

Page 108
• adenosyl transfer mode of reactivity: the biosynthesis of S-adenosyl methionine
O
N
N N
N
NH2
HO OH
O
N
N N
N
NH2
HO OH
OP
O
O
OP
O
O
OP
O
O
O
S
NH2
O OH
S
NH2
O OH
• adenosyl transfer mode of reactivity: the biosynthesis of vitamin B12
O
N
N N
N
NH2
HO OH
O
N
N N
NNu
NH2
HO OH
NuOP
O
O
OP
O
O
OP
O
O
O
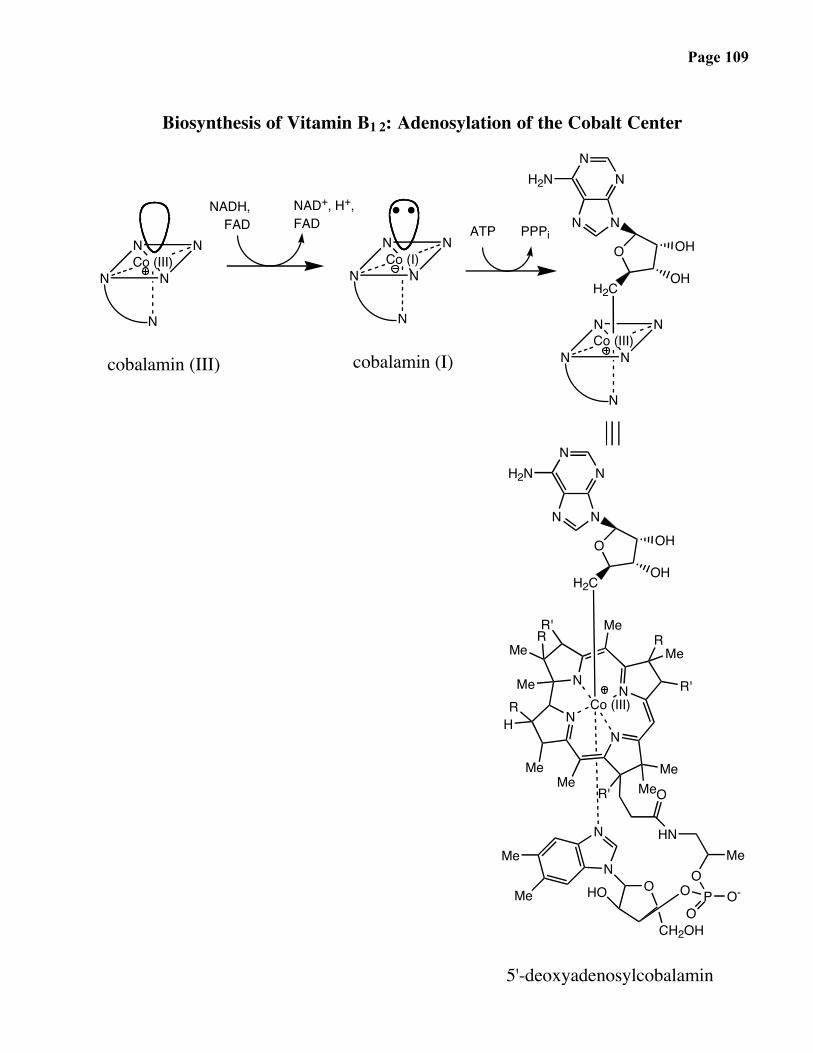
Page 109
Biosynthesis of Vitamin B1 2: Adenosylation of the Cobalt Center
N NCo (III)
N
N N
N NCo (I)
N
N N
N NCo (III)
N
N N
H2C
NNH2N
N N
O OH
OH
cobalamin (III) cobalamin (I)
NADH,FAD
NAD+, H+,FAD ATP PPPi
Co (III)
H2C
NNH2N
N N
O OH
OH
N
Me
N
NN R'
RMe
MeMeR'
RH
Me
RMe
R' Me
Me
N
NMe
Me OHO
CH2OH
P O-
O
OO
MeHN
O
5'-deoxyadenosylcobalamin

Page 110
Biosynthesis of ATP: An Example
H
O
OH
OP
GAP dehydrogenasephosphoglycerate
kinaseO
O
OH
+ NAD+ + Pi + ADP OP + ATP + NADH
GAP (recall from glycolysis) ΔG° = -3.0 kcal / mol
How to activate the phosphate in the reaction ADP + Pi ATP
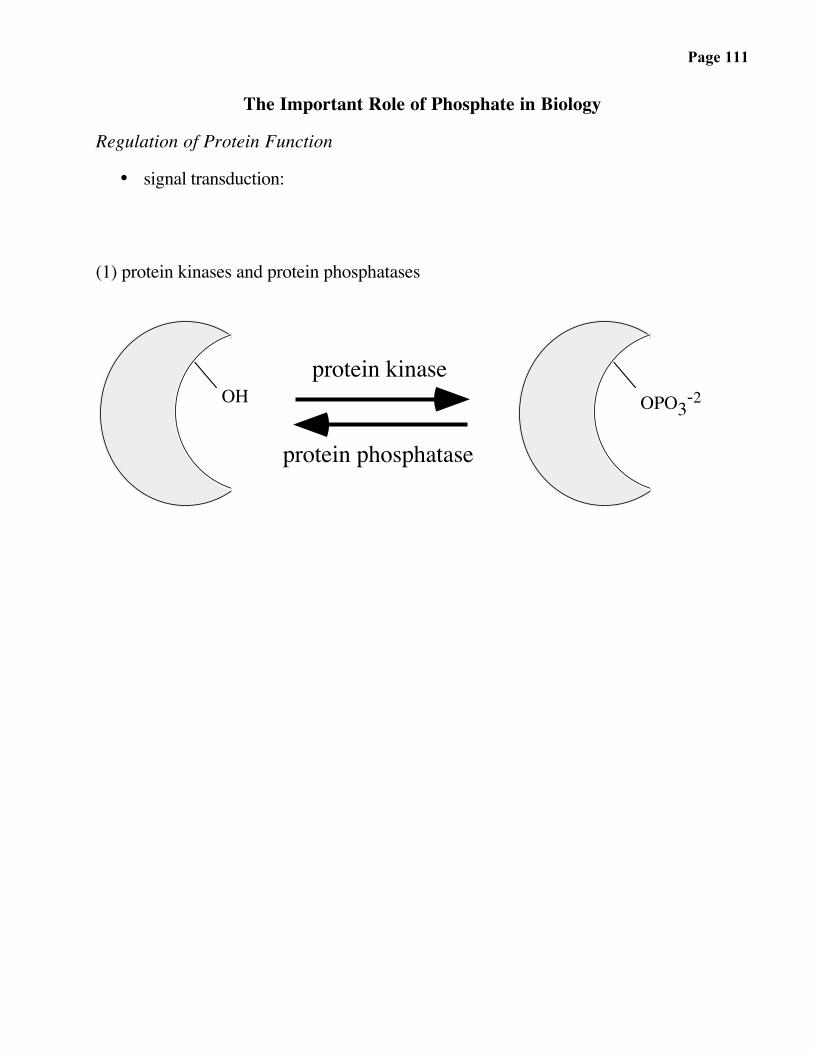
Page 111
The Important Role of Phosphate in Biology
Regulation of Protein Function
• signal transduction:
(1) protein kinases and protein phosphatases
OH OPO3-2protein kinase
protein phosphatase

Page 112
Consequence of Phosphorylation
(1) Protein-protein association
(2) Conformational change

Page 113
Proximity and Orientation Effects in Chemistry and Biology
• proximity effects
CHEMISTRY:
MeOHH
OH
H
O
O
H
+
very slow(i)
H
O OMe
OHfast
(ii) HH
O OMe
HH
O
O MeOH
BIOLOGY:

Page 114
• orientation effects
CHEMISTRY:
very slow
S
SO2
O
O O
Me
Me(-)
S
SO2
O-
O O
Me
Me
(remember 180ο con-
straint for SN2 reaction)
BIOLOGY:
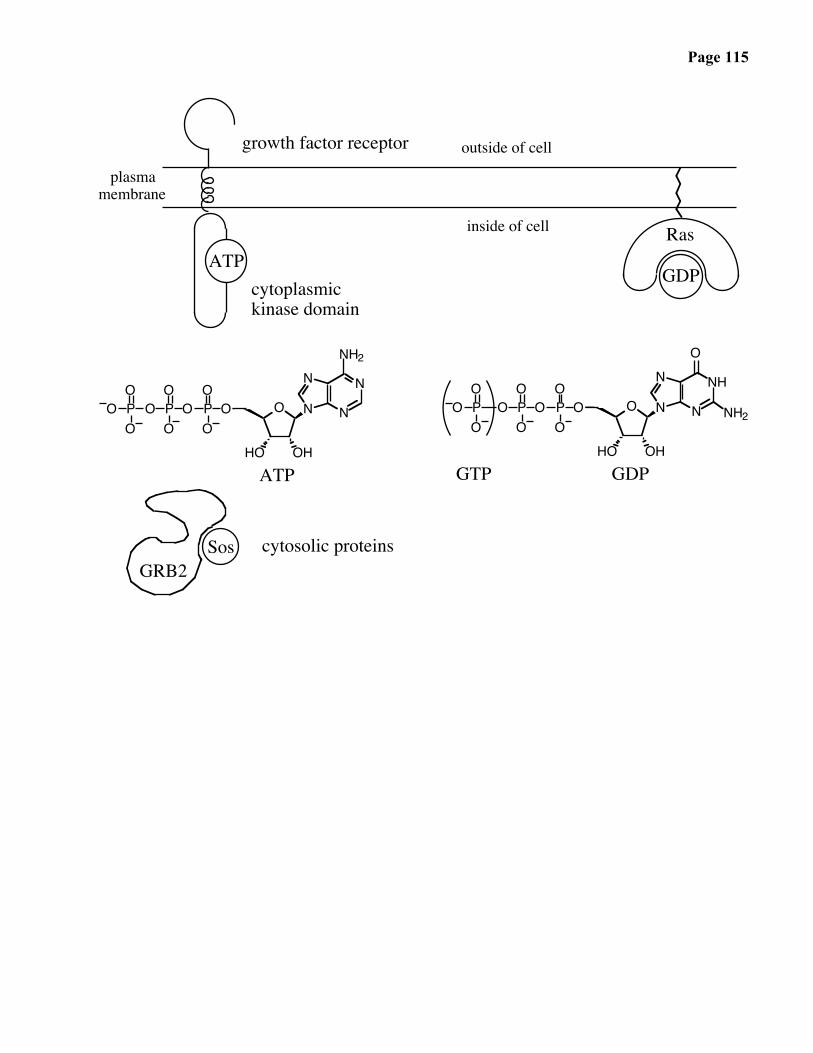
Page 115
O
N
N N
N
NH2
HO OH
OPO
OOP
O
OOP
O
OO O
N
N N
NH
O
HO OH
ATP
OPO
OOP
O
OOP
O
OO
growth factor receptor
plasmamembrane
cytoplasmickinase domain
inside of cell
ATP
NH2
outside of cell
GDPGTP
cytosolic proteins
GDP
Ras
GRB2Sos
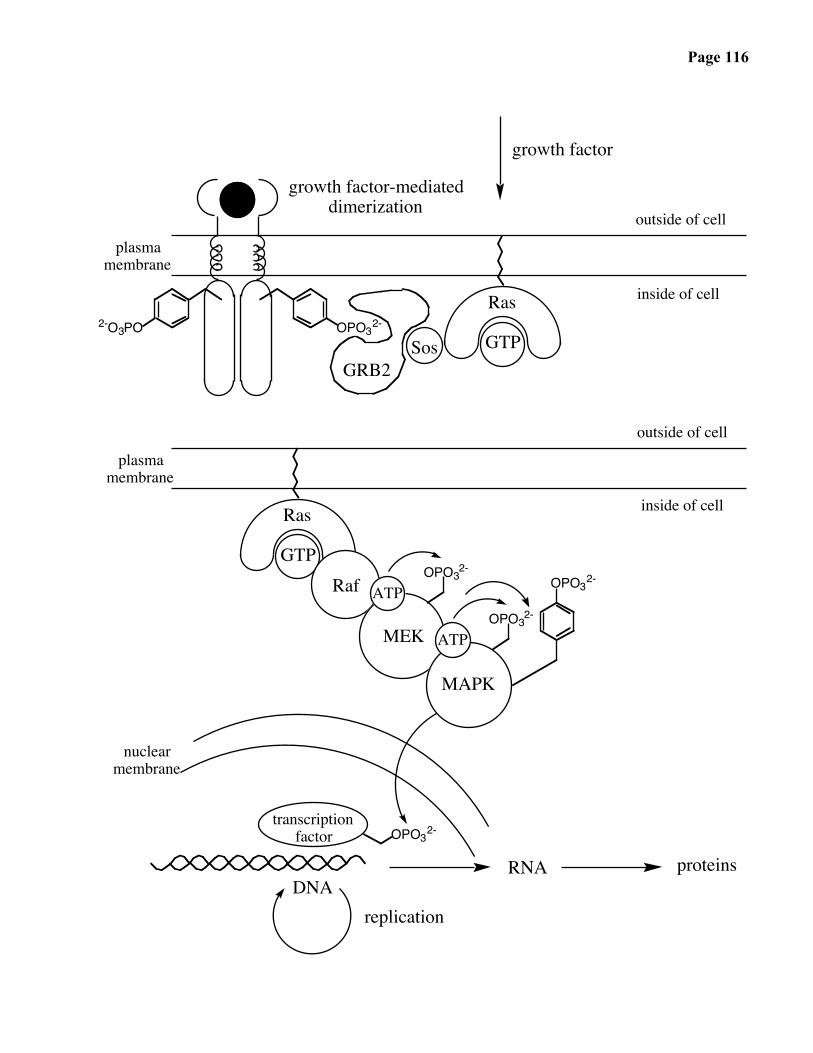
Page 116
growth factor-mediateddimerization
plasmamembrane
inside of cell
outside of cell
GTP
Ras
GRB2Sos
2-O3PO OPO32-
plasmamembrane
inside of cell
GTP OPO3
2-
Ras
OPO32-
MAPK
Raf
outside of cell
OPO32-
MEK
ATP
ATP
nuclearmembrane
OPO32-
transcriptionfactor
DNAreplication
RNA proteins
growth factor

Page 117
STRUCTUREAND
CHEMISTRYOF
NUCLEICACIDS
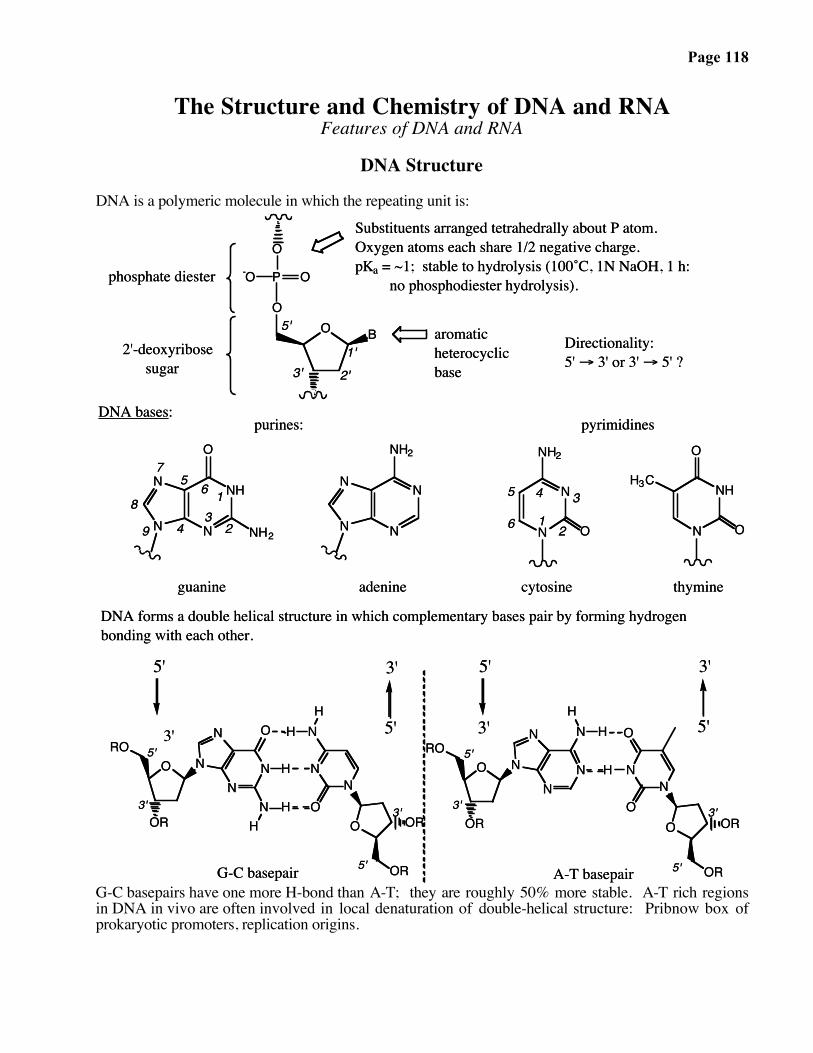
Page 118
The Structure and Chemistry of DNA and RNAFeatures of DNA and RNA
DNA Structure
DNA is a polymeric molecule in which the repeating unit is:
6
5 4 3
21
9
8
4
65
231
7
A-T basepairG-C basepair
5'
3'
3'
5'5'
3'5'
guanine
Directionality:5' → 3' or 3' → 5' ?
pyrimidines
DNA forms a double helical structure in which complementary bases pair by forming hydrogen bonding with each other.
purines:DNA bases:
Substituents arranged tetrahedrally about P atom. Oxygen atoms each share 1/2 negative charge.pKa = ~1; stable to hydrolysis (100˚C, 1N NaOH, 1 h: no phosphodiester hydrolysis).
aromaticheterocyclicbase
5'
3' 2'1'2'-deoxyribose
sugar
phosphate diester
O B
O
N
O
P O-O
N
N
NH
O
NH2 N N
NH2
N
N
NN
O
NH2
N
O
O
NHH3C
O N
N
O
NN
N
O
N
N
O
N
HH
H
HRO
OR
OR
H
OR
OOR
ROO
NH
NH
H O
N
OR OR
NO
N
N
N
thymineadenine cytosine
3'
3'3'3'3'
5'5'
5'5'
6
5 4 3
21
9
8
4
65
231
7
A-T basepairG-C basepair
5'
3'
3'
5'5'
3'5'
guanine
Directionality:5' → 3' or 3' → 5' ?
pyrimidines
DNA forms a double helical structure in which complementary bases pair by forming hydrogen bonding with each other.
purines:DNA bases:
Substituents arranged tetrahedrally about P atom. Oxygen atoms each share 1/2 negative charge.pKa = ~1; stable to hydrolysis (100˚C, 1N NaOH, 1 h: no phosphodiester hydrolysis).
aromaticheterocyclicbase
5'
3' 2'1'2'-deoxyribose
sugar
phosphate diester
O B
O
N
O
P O-O
N
N
NH
O
NH2 N N
NH2
N
N
NN
O
NH2
N
O
O
NHH3C
O N
N
O
NN
N
O
N
N
O
N
HH
H
HRO
OR
OR
H
OR
OOR
ROO
NH
NH
H O
N
OR OR
NO
N
N
N
thymineadenine cytosine
3'
3'3'3'3'
5'5'
5'5'
G-C basepairs have one more H-bond than A-T; they are roughly 50% more stable. A-T rich regionsin DNA in vivo are often involved in local denaturation of double-helical structure: Pribnow box ofprokaryotic promoters, replication origins.

Page 119
The Electronic Structure of DNA
N
N
N
N
H
NRH
H
O
N
NH
O
NH2
N
NH
• Basicity of different sites
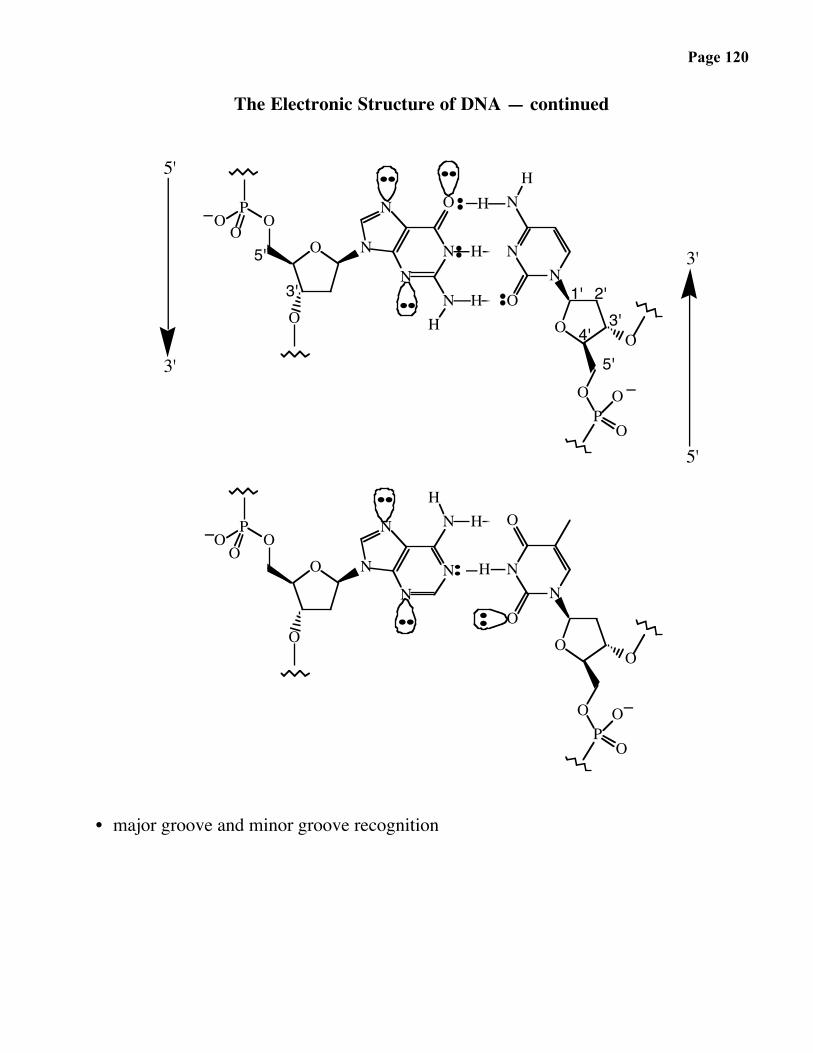
Page 120
The Electronic Structure of DNA — continued
O
O
OP
OO
NN
N
N
N HH
NN
O
O
H
OO
OPO
O
O
O
OP
OO
NN
N
N
O H
NN
N
OO
O
OPO
O
H
H
N HH
5'
3' 1' 2'3'
4'5'
5'
3'
3'
5'
• major groove and minor groove recognition

Page 121
Role of the 2'-OH in RNA
• stability
O
OP
O
O-
O
B
O B
O
100 °C
1 N NaOH
NOREACTION
O
OP
O
O-
O
B
O B
O
OH
OH
100 °C
1 N NaOH
O
O32-PO
B
OH
HO
O
HO
B
OPO32-
HO
+
quantitative hydrolysis
• catalysis

Page 122
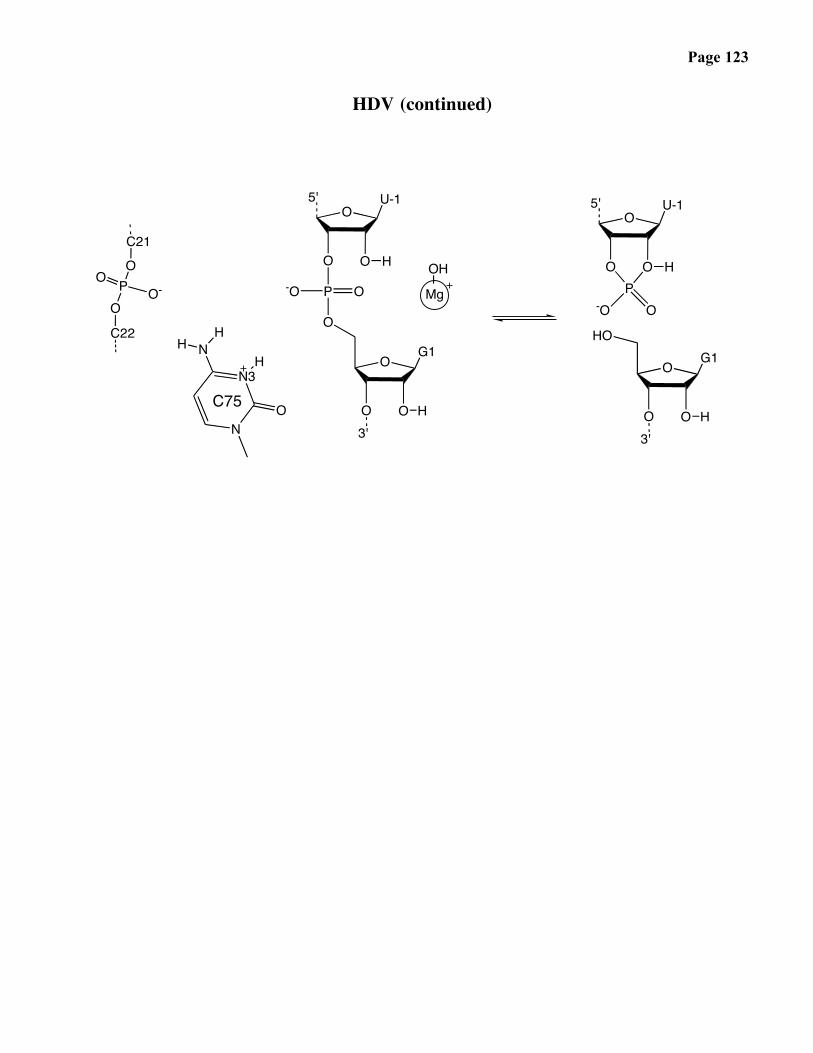
Ribozymes
• self-cleaving ribozymes: HDV
- double-rolling-circle-mechanism of replication- ribozyme cleaves genomic RNA into linear monomers
Page 123
HDV (continued)
O5'
O O
U-1
H
P
O
-O O
O
O3'
O
G1
HN
N3
O
NH
H
C75
H
Mg
OH
O-PO
O
O
C21
C22
O5'
O O
U-1
HP
HO
-O O
O
O3'
O
G1
H
Page 124
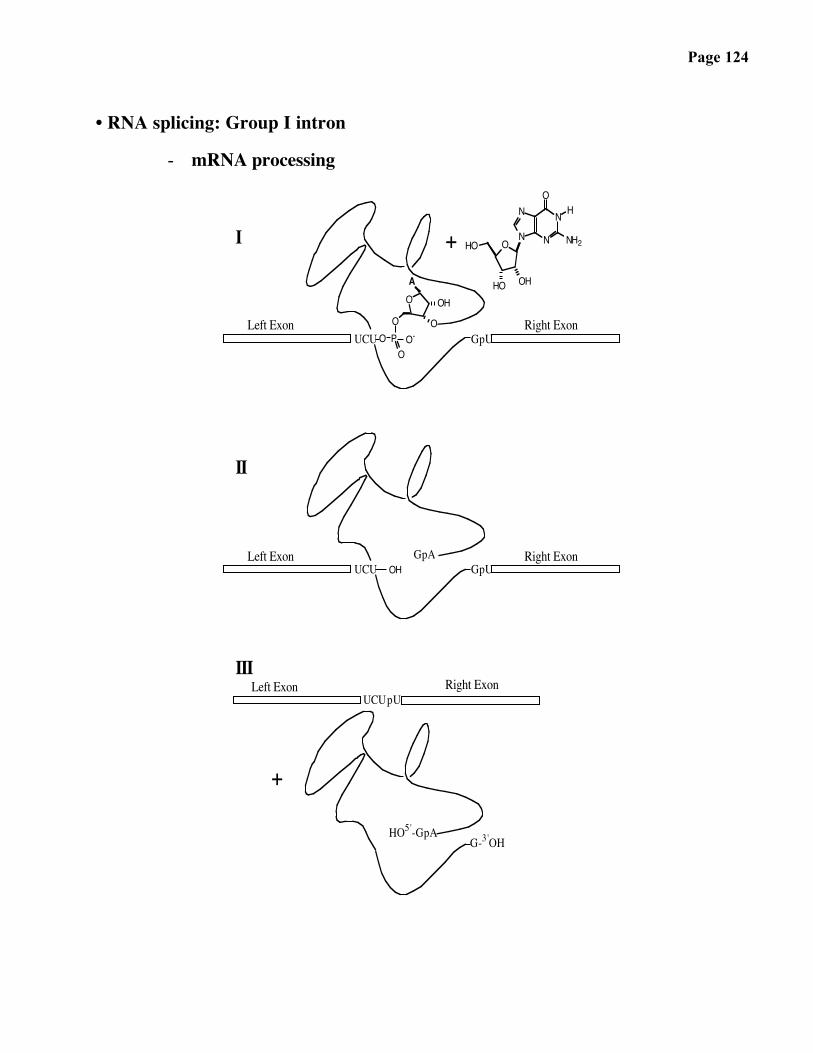
• RNA splicing: Group I intron
- mRNA processing
OA
OPOOO-
OH
O
N
NN
NO NH2
O
HO
HO OH
H
OH
UCU
III
+
I
Right ExonLeft ExonpUUCU
HO5'-GpAG-3'OH
+
Right ExonGpU
Left ExonUCU
II
Right ExonGpU
GpA
Left Exon
Page 125
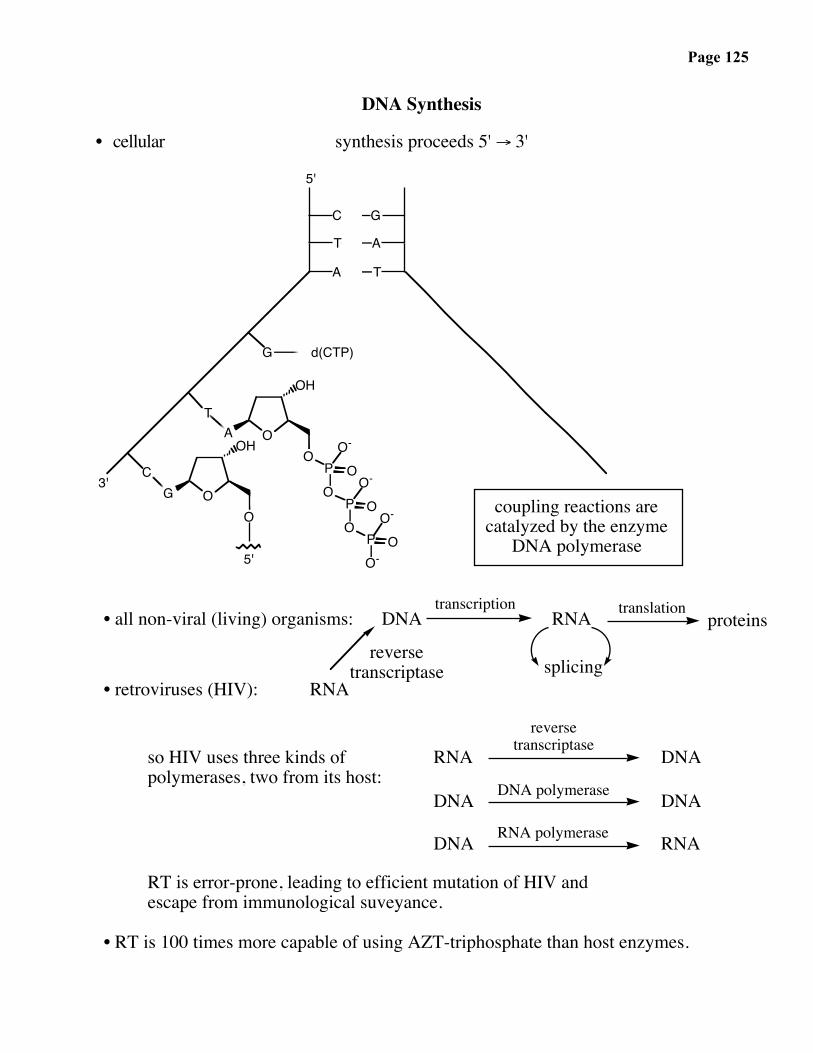
DNA Synthesis
• cellular synthesis proceeds 5' → 3'
C
T
A
G
A
T
G
T
C
d(CTP)
A O
OH
OPO
PO
O-
O
OO-
PO-
OO-
G O
OH
coupling reactions arecatalyzed by the enzyme
DNA polymerase
3'
O
5'
5'
• all non-viral (living) organisms: DNA RNAtranscription
proteinstranslation
splicing• retroviruses (HIV): RNA
reversetranscriptase
so HIV uses three kinds ofpolymerases, two from its host:
RNAreverse
transcriptase
DNA polymerase
DNA
DNA DNARNA polymeraseDNA RNA
RT is error-prone, leading to efficient mutation of HIV andescape from immunological suveyance.
• RT is 100 times more capable of using AZT-triphosphate than host enzymes.

Page 126
AZT (Azidothymidine)

Page 127
Biological DNA Synthesis
OO
PO
PO
PHO
Bn+1
OH
O O-O O-O O-
O
OHpolymerase
B:
Bn
P
O- O
P
O-
P
O
OBn
P
O-O
P
O-O
-O
O
OH
Bn+1
HO O O-
O
OO
P
S- O
OH
Bn+1
HO O O OO
OO
PBn+1
OH
O
-S
O
O
Bn
OHO
N
H
NH
O
O
H3C
NN
NN
N
N
H3C
O
O
N
NH
H
POPOPOO
Bn
O-
O
Displacement at phosphorus occurs by direct, in-line displacement; contrast to substitution reactions at carbonyls (formation of tetrahedral intermediate).
Why does this drug causecessation of DNA synthesisafter it is incorporated into DNA?
Inhibits DNA synthesis;selective for viralpolymerase, HIVreverse transcriptase.
Anti-AIDS drug AZT (azidothymidine)
5'
3'
3'
5'
dNTP-α-phosophorothioate
Two important points:1. α-phosphate of dNTP becomes incorporated into phosphodiester bond in DNA; use of isotopically labelled phosphorus at that position leads to incorporation of the isotope into DNA → "end-labelling".2. Nucleophilic attack of 3'-OH occurs with inversion of stereochemical configuration at phosphorus! How experiment is done:
β
α
γ β α
pyrophosphategood leaving group
5'
3'3'
5'
dNTPmonomeric unit in biological DNA synthesis.
γ
AZT
in vivo
-
5'
3'++ 3'
5'
-
5'
3'
5'
3'
enzymaticDNA synthesis
3'
5'
α
3'
5'αβγ

Page 128
The Structure and Chemistry of DNA and RNALaboratory DNA synthesis
DNA Synthesis
• laboratory synthesis proceeds 3' → 5'
O B*
O
O
O
O
O
OMe
MeO
O B*
O
PO
O
OMe
MeO
SolidSupport
(DMT)
N
initial base(* protected)
N≡C

Page 129
The Assembly Steps of Solid-Phase DNA Synthesis:
phosphoramiditeβ-cyanoethyl
3'
5'
3'
5'
controlledporeglass
oxidation
coupling
pyridine / H2O
controlledporeglass
activation
detritylation
controlledporeglass
controlledporeglass
OO
O
O
ODMT-O
B* B*HO
O
O
O
OO
O
B*
NP
O
NC
DMT-OO
NN
O
N
O
N
DMT-O
NC
PO
NN
B*
NN
OO
O
O
OO
PB*
B*
O
OO
DMT-OO
NCNC
ODMT-O
PO O
B*
B*O
O
O
OO
O
I2
CH3CN
CH3CN
Cl2HCCO2H
H
capping
Page 130
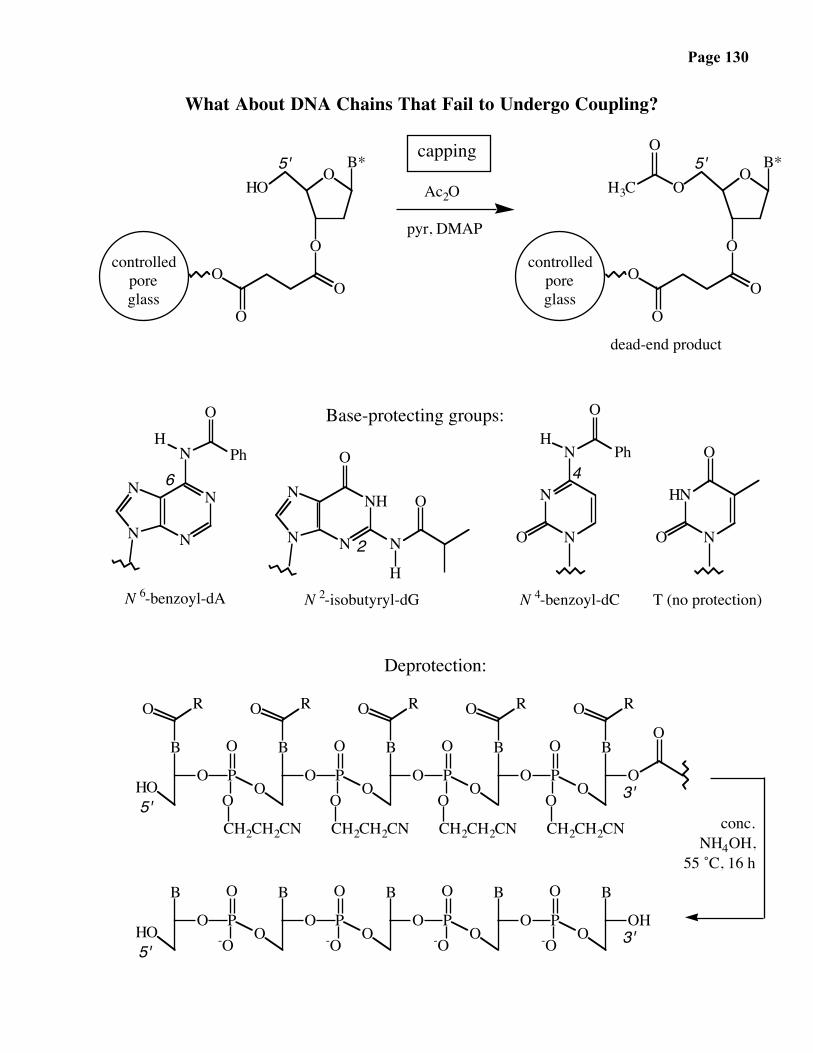
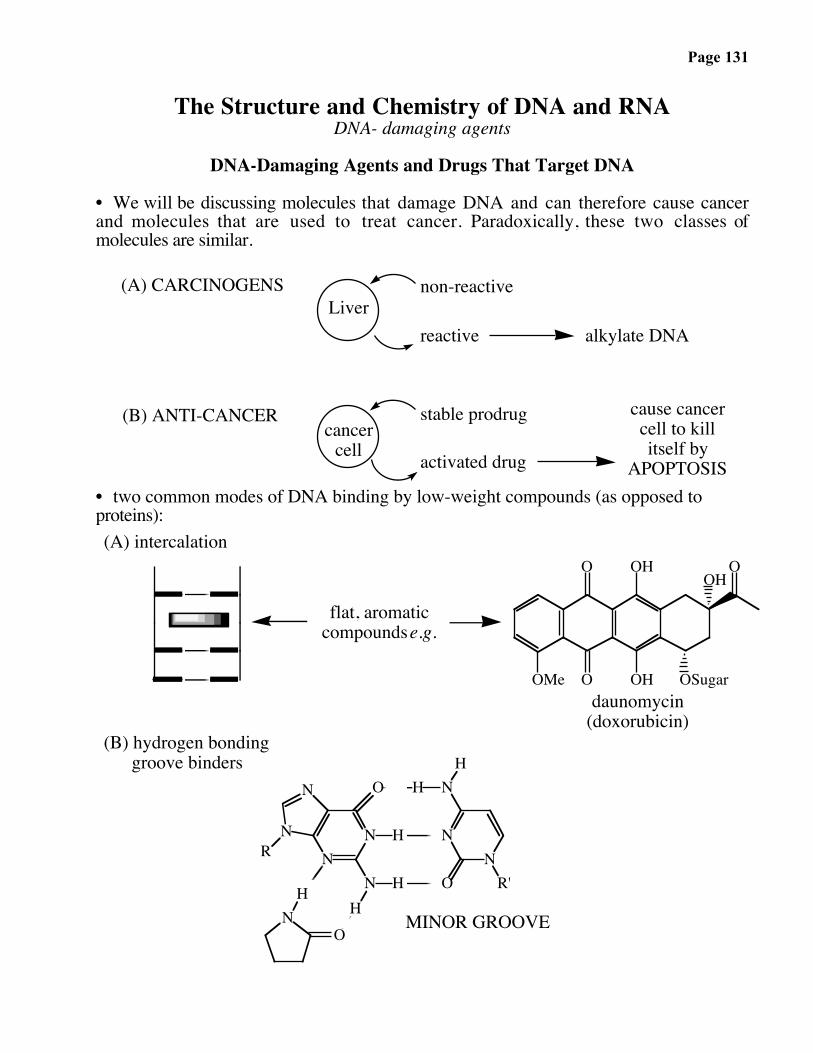
What About DNA Chains That Fail to Undergo Coupling?
4
2
6
3'
3'
5'
conc.NH4OH,
55 ˚C, 16 h
Deprotection:
5'
T (no protection)N 4-benzoyl-dCN 2-isobutyryl-dGN 6-benzoyl-dA
dead-end product
Base-protecting groups:
5'
controlledporeglass
Ac2O
pyr, DMAP
capping
controlledporeglass
5' B*HO
O
O
O
OO O
O
O
O
OO
B*H3C
O
N
N
N
NH
O
N
NO
N N
O
H
N
PhHH
Ph
N
O
O
HOO
O
B
O
CH2CH2CN
O
N
P
O
O
O
HNN
N
N
OO
B
O
CH2CH2CN
B
P
O
P
O
OO
B
O
CH2CH2CN
O
CH2CH2CN
OO
B
P
OO
RO O R RO O R RO
O P
O
-OO
B
OHO
B
P-O
O
OO P
O
-OO
B
O
B
P-O
O
O
B
HO
Page 131
The Structure and Chemistry of DNA and RNADNA- damaging agents
DNA-Damaging Agents and Drugs That Target DNA
• We will be discussing molecules that damage DNA and can therefore cause cancerand molecules that are used to treat cancer. Paradoxically, these two classes ofmolecules are similar.
(A) CARCINOGENSLiver
non-reactive
reactive alkylate DNA
(B) ANTI-CANCERcancer
cell
stable prodrug
activated drug
cause cancercell to killitself by
APOPTOSIS• two common modes of DNA binding by low-weight compounds (as opposed toproteins):
O
OOMe
OH
OH OSugar
OHO
NN
N
N
O
RH
N HHN
O
H
NN
O R'
(A) intercalation
NHH
flat, aromaticcompounds e.g.
daunomycin(doxorubicin)
(B) hydrogen bonding groove binders
MINOR GROOVE

Page 132
• Oxidative metabolism in the liver is used to convert fat (lipid) soluble compounds into water-soluble ones for excretion. Enzymes named P450 oxidases are used for this
purpose:
P450 oxidase
O2
Ohydratase
H2O
OH
OH
hydrogen-bondingcapacity of OH'sincreases watersolubility
• Aflatoxin — produced by a mold (Aspergillus flavus) that grows on grain, especiallyrice and peanuts, and causes cancer:
O
O O
OMeO
O
H
H
• Benzo[α]pyrene — In the mid-1700’s, Sir Percival Polts, a British surgeon, noted thehigh occurence of skin cancer among chimney sweeps, now known to be due to thisend product of combustion (e. g. a charcobroiled steak). 1300 tons per year areemitted into the air, particularly by coal-burning power plants.
member of classof compounds
called polyaromatichydrocarbons (PAH's)

Page 133
Aflatoxin
O
O O
OMeOO
H
H O
O O
OMeOO
H
HO
N
NH
O
NH2
N
N
N
NH
O
NH2
N
N
O
OHH
HO
enzymaticoxidation
+
N7-guanyl adduct
O
HOPO
OO
OPO
OODNA
strandcleavage

Page 134
How Does N7 Alkylation Lead to DNA Strand Cleavage?
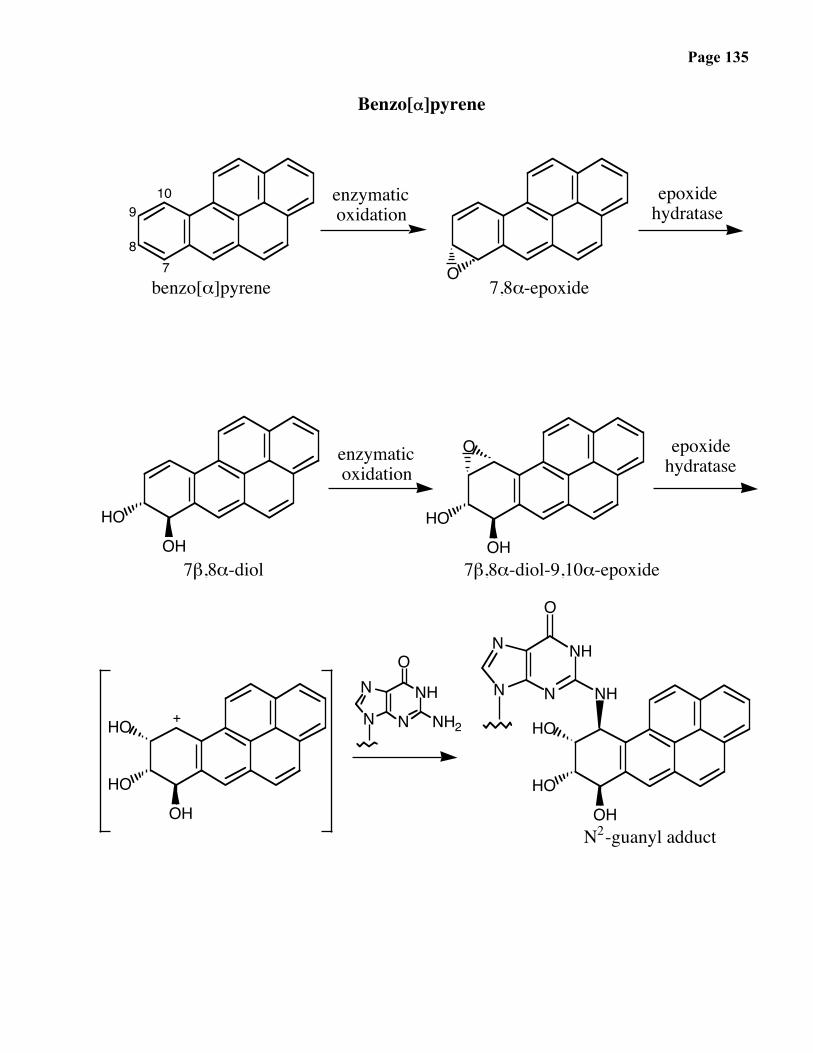
Page 135
Benzo[α]pyrene
O
HOOH
O
OHHO
HOOH
7
OHHO
HO
10
NNHN
N
8
9
O
benzo[α]pyrene
epoxidehydratase
7β,8α-diol
NH2
7,8α-epoxide
HO
N
NHN
N
enzymaticoxidation
O
N2-guanyl adduct
7β,8α-diol-9,10α-epoxide
enzymaticoxidation
NH+
epoxidehydratase
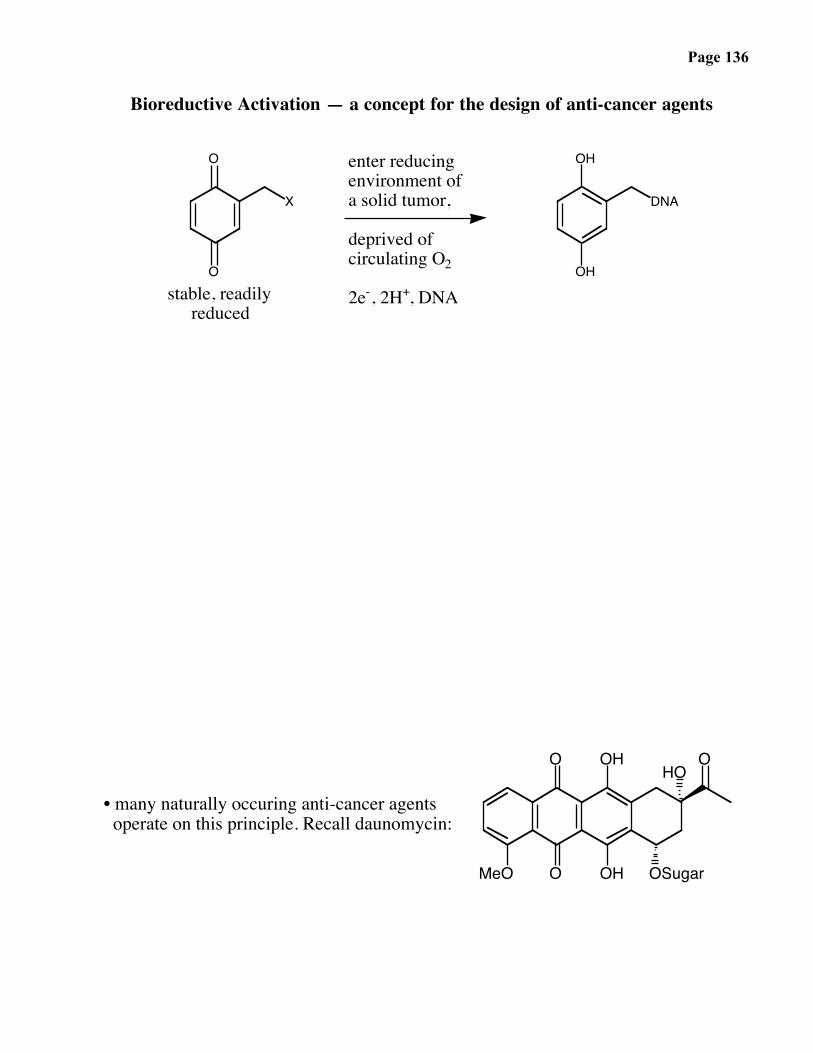
Page 136
Bioreductive Activation — a concept for the design of anti-cancer agents
O
O
X
OH
OH
DNA
enter reducingenvironment ofa solid tumor,
deprived ofcirculating O2
2e-, 2H+, DNAstable, readilyreduced
O
OMeO OH
OH
OSugar
HO
• many naturally occuring anti-cancer agents operate on this principle. Recall daunomycin:
O
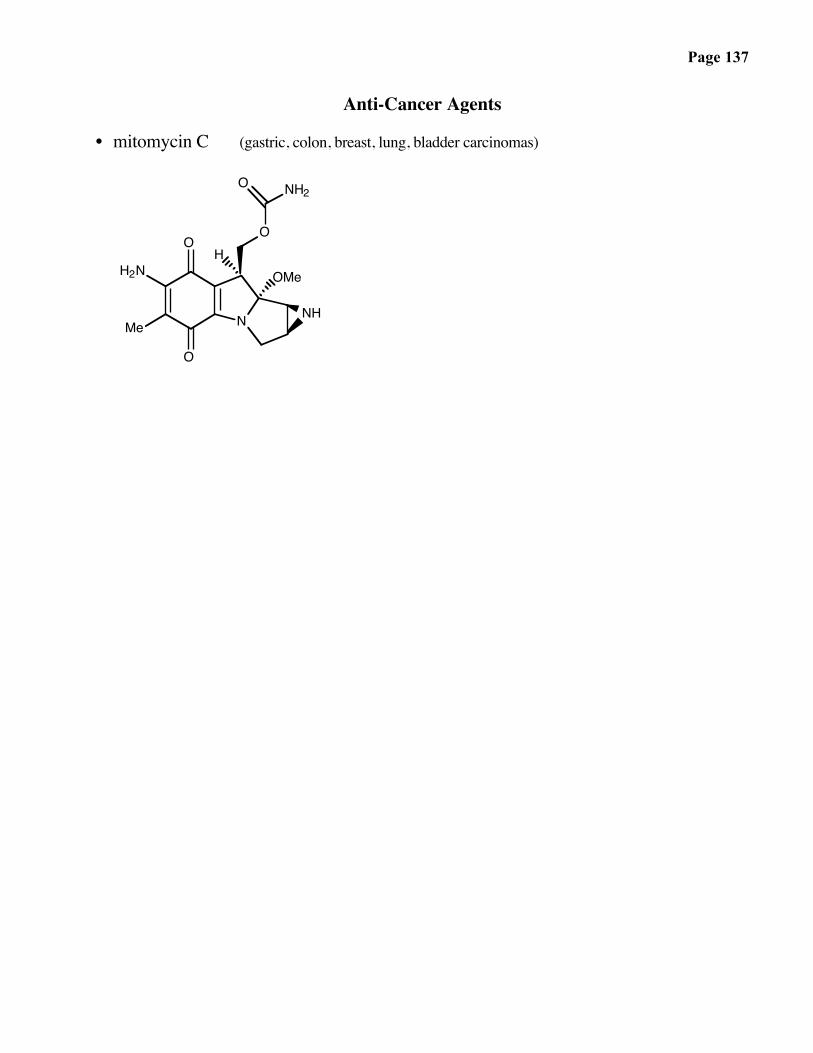
Page 137
Anti-Cancer Agents
• mitomycin C (gastric, colon, breast, lung, bladder carcinomas)
O
O
Me
H2N
N NH
OMe
O
NH2O
H

Page 138
The Reaction of Mitomycin C with DNA
N
OH2N
H3CO
OCH3
NH
O
ONH2
O
O
NH
OCH3
NH2
OH3C N:
H2NO
N
OH2N
H3CO
H
NH
O
ONH2
NH2
O
O
NH
H
OH3C N
H2NO
N
OH2N
H3CO
NH2
NH2
O
ONH2
O
O
NH2
HN
OH3C N
H2NO
N N
NO
H2NNDNA
NH
ON
DNAHN
NN:
OH2N
H3CO NH2
O
ONH2
NH
NH2O
O
NH2
HN
N
H2NO
O
N N
NO
DNA NH
NDNA
H2N
ON
NH2
HN
OH3C N
H2NO
N N
NO
DNA
HN
N
HN
N N
N
NDNA
NH
ONHN
NN
OH2N
H3CO NH2
O
HN
DNA
HN
NH
NDNA
ON
NH2
HN
OH3C N
H2NO
N N
NO
DNA
HN
N
H3C
path a:elimination
Mitomycin C (MC):a clinically used antitumor drug
FADH2 FADH2
+enzyme-catalyzed
+
MC semiquinone radical
+
. .
O2O2
monoadduct: dead end product
path b:oxidation
. .
+
. .. .
O2 O2
MC-DNA crosslink

Page 139
• cisplatin (ovarian cancer, lymphomas, squamous cell carcinomas of the head and neck)
Pt
ClH3N
H3N Cl

Page 140
The Growing Class of Enediyne AntibioticsFrom the thesis of Dr. John A. Porco
O
OMeNHEt
OSMe
Me
OH
HO
OO
OMe
O
Me
OH
NH
OO
I OMeO
O
MeO
O SSSMe
H
NHCO2Me
OH
HOMe
ONHCO2Me
HO
SSSMeO
O
HOH
O
O NH
O
O
OOR2
Me
R1O
OH
Me
MeSMe
OH
NHR3OMe
O
OO
O
OO
OO
HO
HO
Me
NHMe
OH
Me
MeO
H
H N
OHMeO
HO2C
H
H O
Me
O
O
OH
HO
dynemicin A
calicheamicin γ1
esperamicins A1, A1β, A2
neocarzinostatin chromophore
H

Page 141
Carbon Cations, Radicals, and Anions
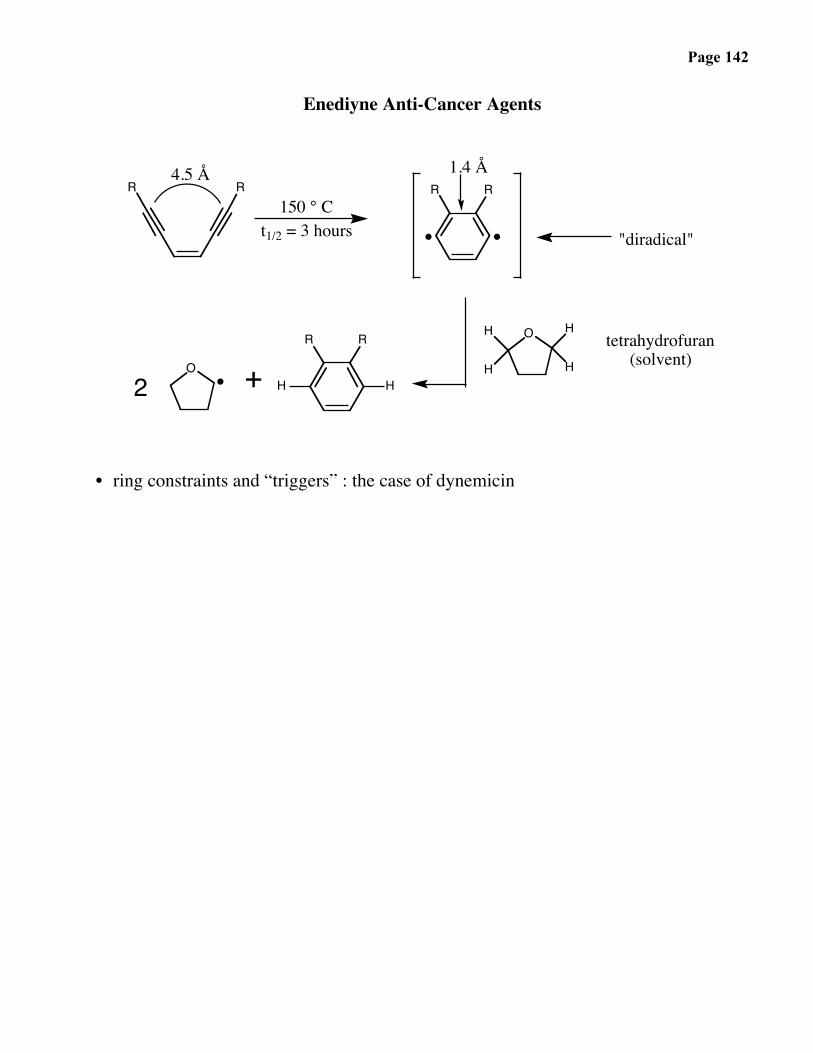
Page 142
Enediyne Anti-Cancer Agents
OHH
R R
R R R R
+O
2
H
H
H
H
4.5 Å
150 ° Ct1/2 = 3 hours
1.4 Å
"diradical"
tetrahydrofuran(solvent)
• ring constraints and “triggers” : the case of dynemicin

Page 143

Page 144
Proposed Priming / Cleaving Mechanism forDNA-Damaging Properties of Dynemicin A
O
HO
MeO
OH
HO
HNH
H
H
HOO
O
HO
Me
OH
OH
Me
HOHO
OH
H
H
NHH
OHH
H
MeO
HO
O
Nu
OH
OH
MeOH
OHHO
H
H
NHH
OH
H
MeO
HO
O
Me
OHO
MeO
H
H OH
HNH
H
H
OHO
HO HO
OH
OH
HO
Me
O
O
H
H
NHH
HO
OHH
H
MeO
HOO
Me
R
O
HO
MeO
H
HOH
HNH
H
H
OH
HO HO
OH
HO
R
C H
C
HH
anthraquinone forstep C
B
bioreduction
A
nucleophilicaddition
(NuH) C
diylformation
E
.
DNA
O2 DNA+
.
epoxideopening
R = Nuc (Step C)

Page 145
Enediyne Anti-Cancer Agents
• ring constraints and “triggers” : the case of calicheamicin

Page 146

Page 147
BIOSYNTHESISOF
NATURALPRODUCTS
Page 148
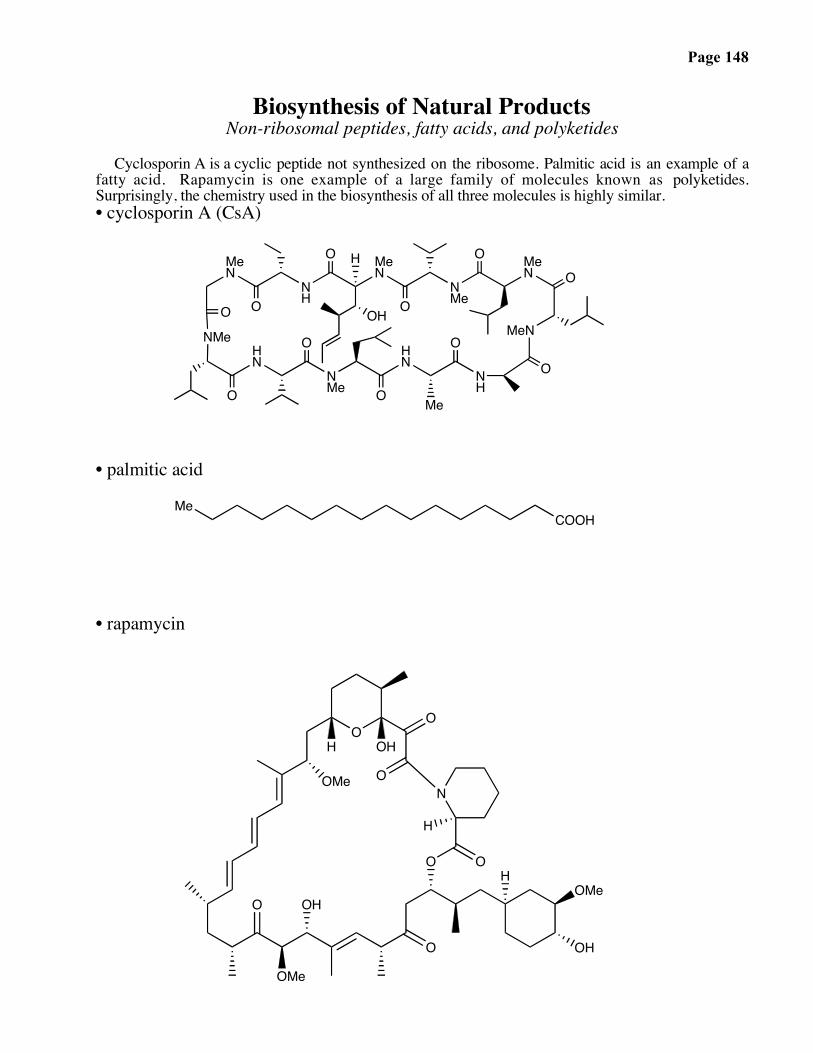
Biosynthesis of Natural ProductsNon-ribosomal peptides, fatty acids, and polyketides
Cyclosporin A is a cyclic peptide not synthesized on the ribosome. Palmitic acid is an example of afatty acid. Rapamycin is one example of a large family of molecules known as polyketides.Surprisingly, the chemistry used in the biosynthesis of all three molecules is highly similar.
O
O
O
OH
OMe
OMe
OH
HO O
N
O
OMe
H OH
O
H
MeCOOH
MeN
NH
MeN
NMe
MeN
H
NH
HN
NMe
OHO
O OO
O
O
O OHN
MeN
O
O
• rapamycin
NMe
O
• palmitic acid
• cyclosporin A (CsA)
Me

Page 149
Biosynthesis of Natural Products
All three classes of molecules are synthesized on complexes of proteins of extremelyhigh molecular weight. One component is the acyl-carrier protein, which provides thecovalent transport and attachment of acyl intermediates. The long “arm” ofphosphopantetheine (note the relationship to Coenzyme A) provides the neededflexibility as the nascent chains are moved about to different enzyme active sites in thecomplex:
ONH
NH
OH
O O
SH
N N
NN
H2N O
HO OPO32-
O ONH
NH
OH
O
Coenzyme A
O
SH
nucleophilic carrier site
P
O-
O
O
P
O-
O
P
O-
O
ACP
Ser
O

Page 150
Ribosomal
Non-Ribosomal
Page 151
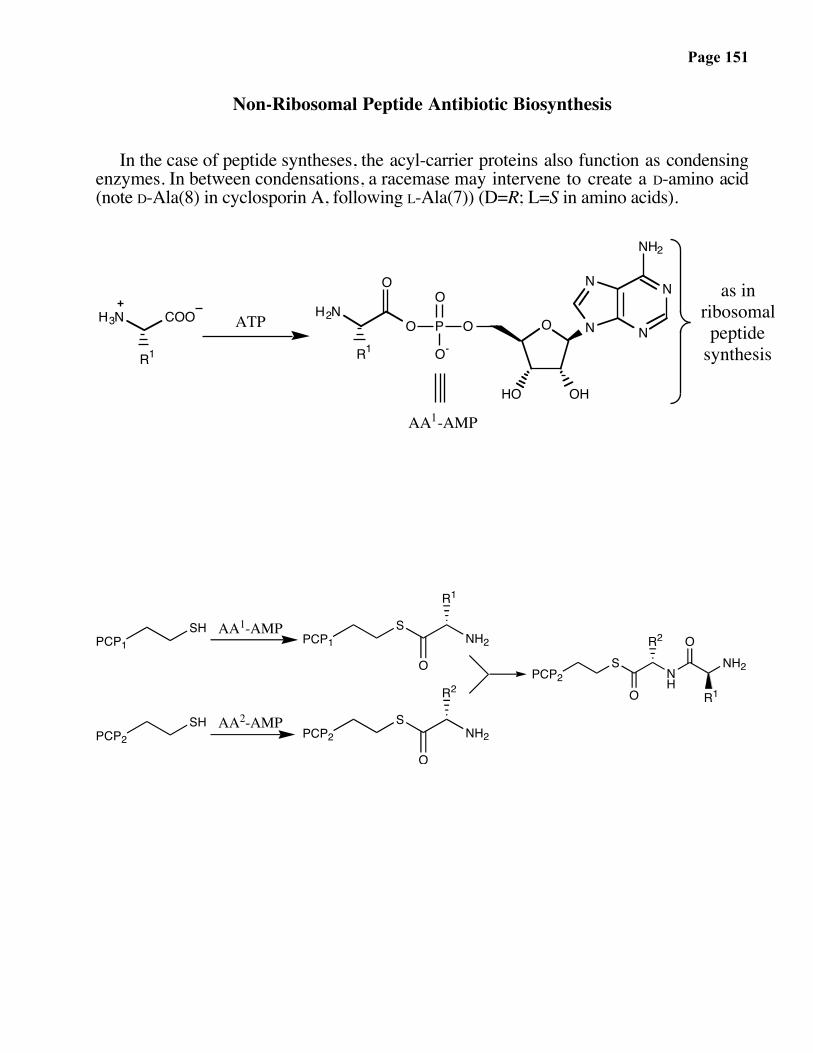
Non-Ribosomal Peptide Antibiotic Biosynthesis
In the case of peptide syntheses, the acyl-carrier proteins also function as condensingenzymes. In between condensations, a racemase may intervene to create a D-amino acid(note D-Ala(8) in cyclosporin A, following L-Ala(7)) (D=R; L=S in amino acids).
H3N COO
R1
ATP O
N
N N
N
NH2
HO OH
OP
O
O-O
O
R1
H2Nas in
ribosomalpeptide
synthesis
AA1-AMP
PCP1SH AA1-AMP
PCP1S
O
NH2
R1
PCP2SH AA2-AMP
PCP2S
O
NH2
R2PCP2
S
ONH
R2 ONH2
R1
Page 152
PCP8S
O
NH2
Me
PCP8S
O
NH
Me OHN
R7 O
R6
racemase (module 8)
PCP8S
O
NH2
Me
PCP8S
O
NH
Me OHN
R7 O
R6
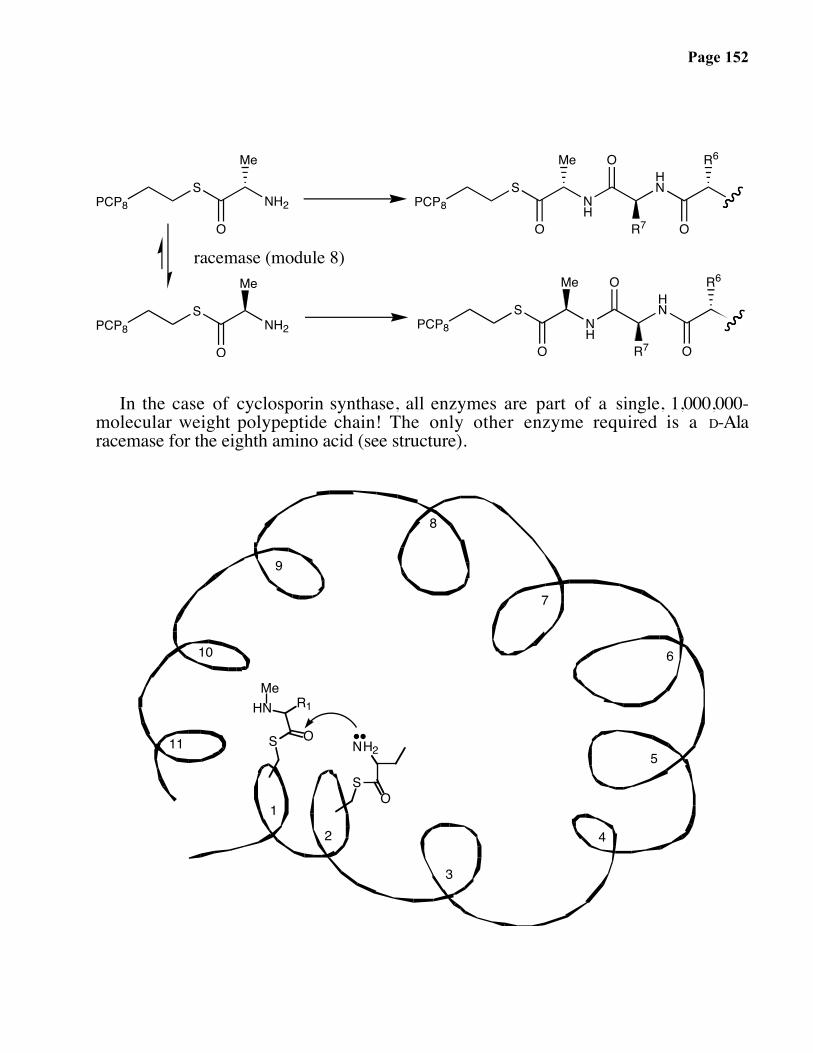
In the case of cyclosporin synthase, all enzymes are part of a single, 1,000,000-molecular weight polypeptide chain! The only other enzyme required is a D-Alaracemase for the eighth amino acid (see structure).
S O
R1HNMe
1
2
3
4
5
6
7
8
9
10
11
SO
NH2
Page 153
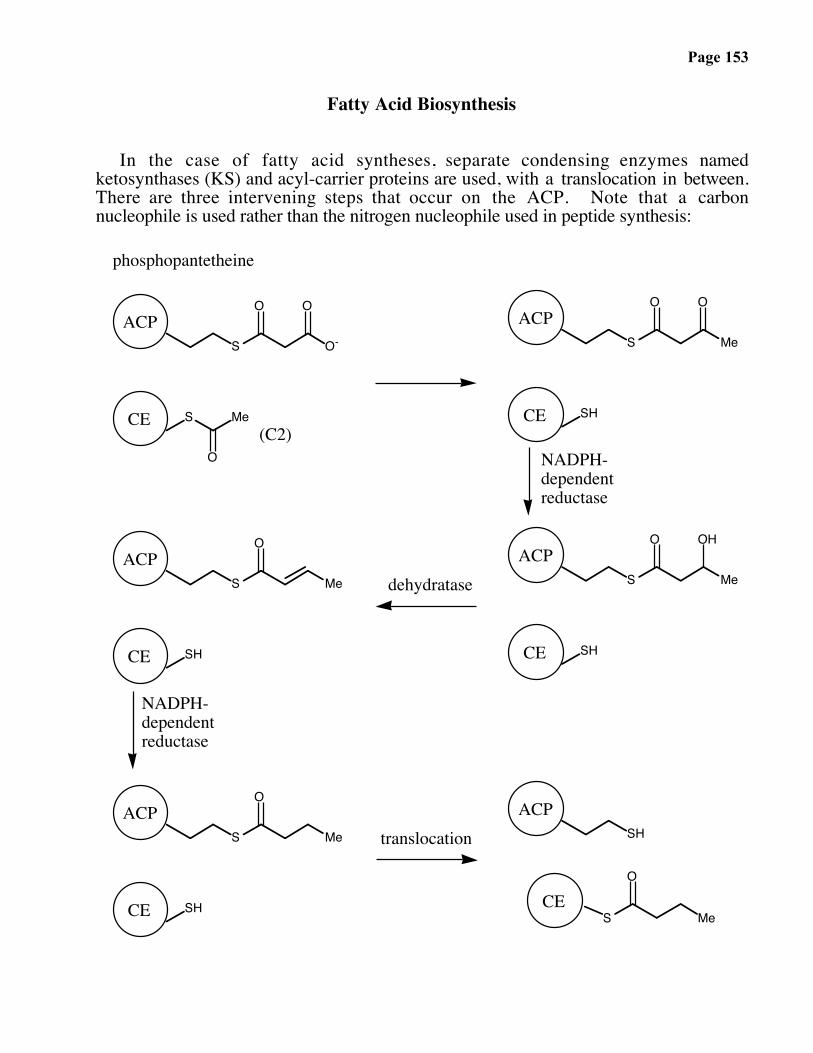
Fatty Acid Biosynthesis
In the case of fatty acid syntheses, separate condensing enzymes namedketosynthases (KS) and acyl-carrier proteins are used, with a translocation in between.There are three intervening steps that occur on the ACP. Note that a carbonnucleophile is used rather than the nitrogen nucleophile used in peptide synthesis:
S O-
O O
S Me
O
S Me
O O
SH
S Me
O OH
SH
S Me
O
ACP
CE
ACP
SH
SH
phosphopantetheine
CE
NADPH-dependentreductase
ACP
S Me
CE
O
ACP
CE
translocation
SH
dehydratase
ACP
NADPH-dependentreductase
S
O
Me
ACP
CE
CE(C2)

Page 154

Page 155
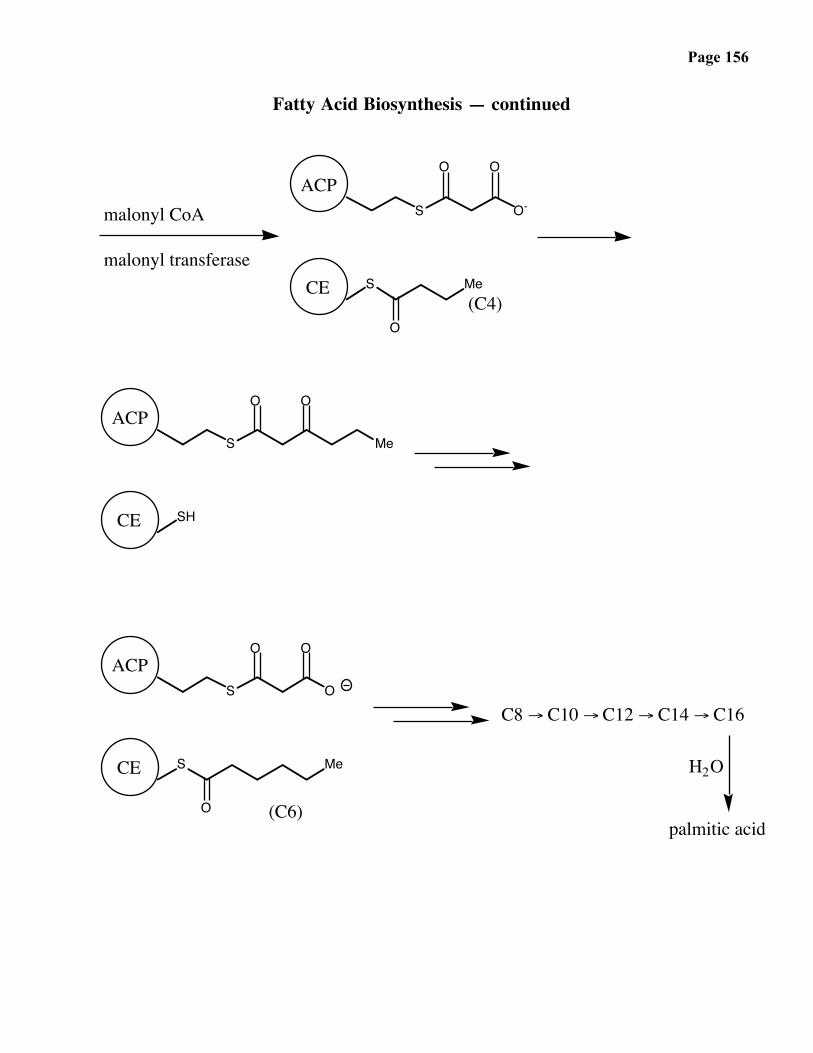
Page 156
Fatty Acid Biosynthesis — continued
S O-
O O
S
O
Me
S
O O
SH
S O
ACP
O O
malonyl CoA
malonyl transferaseCE
(C4)
ACP
CE
S
O
ACP
CE
(C6)
Me
C8 → C10 → C12 → C14 → C16
H2O
palmitic acid
Me

Page 157
Unsaturated Fatty Acids

Page 158
Fatty Acid Synthases (FAS’s)
Polyketide Synthases (PKS’s)
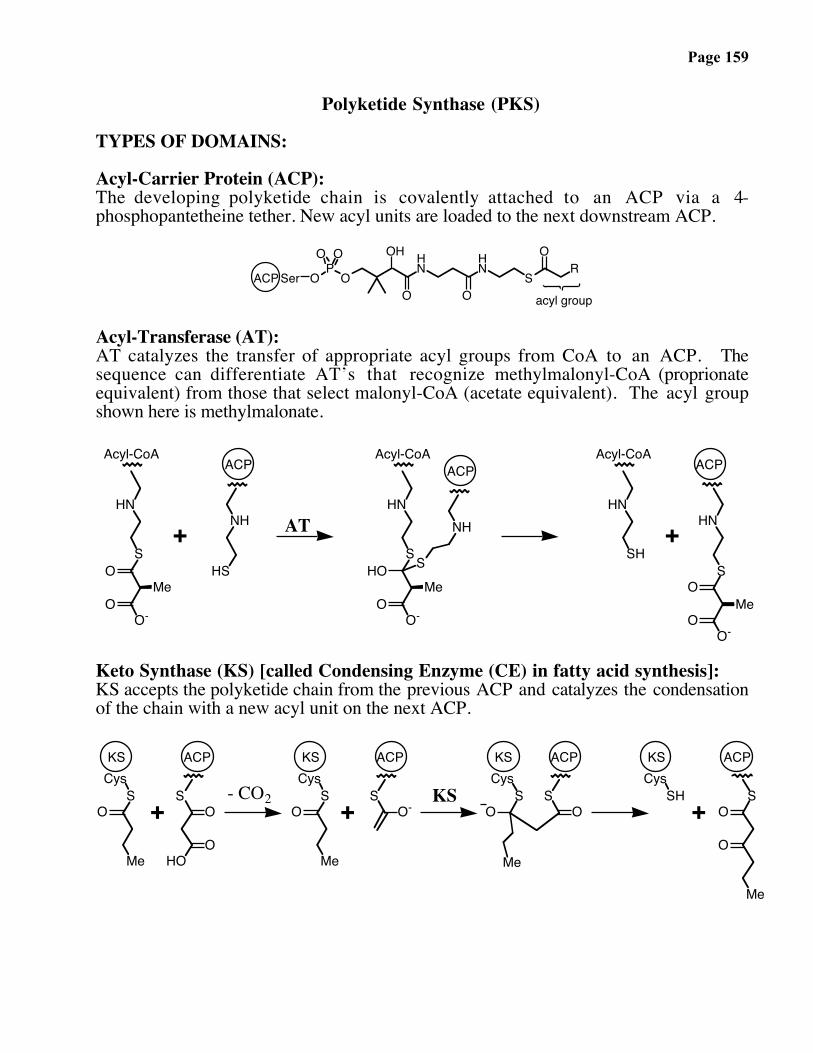
Page 159
Polyketide Synthase (PKS)
TYPES OF DOMAINS:
Acyl-Carrier Protein (ACP):The developing polyketide chain is covalently attached to an ACP via a 4-phosphopantetheine tether. New acyl units are loaded to the next downstream ACP.
O
OH HN
O
HN
S
OR
OP
O
O O
SerACPacyl group
Acyl-Transferase (AT):AT catalyzes the transfer of appropriate acyl groups from CoA to an ACP. Thesequence can differentiate AT’s that recognize methylmalonyl-CoA (proprionateequivalent) from those that select malonyl-CoA (acetate equivalent). The acyl groupshown here is methylmalonate.
HN
S
O-
OMe
O
NH
HS
HN
S
O-
HOMe
O
NH
S
HN
SH+
ACP
HN
S
Acyl-CoA Acyl-CoA
O
OO-
ACPAcyl-CoA
+
Me
ACP
AT
Keto Synthase (KS) [called Condensing Enzyme (CE) in fatty acid synthesis]:KS accepts the polyketide chain from the previous ACP and catalyzes the condensationof the chain with a new acyl unit on the next ACP.
OS
Cys
Me
OS
HOO
OS
Cys
Me
OS
OS
Cys
Me
S
KS
+ O-SH
CysKS
+
KS
+ OS
ACPKS ACP
O
ACP
Me
ACP
- CO2 KS

Page 160
Polyketide Synthase (PKS)
Keto Reductase (KR):KR catalyzes a NADPH-dependent reduction of the β-ketone to an alcohol.
OS
ACP
O
Me
+N
O
NH2
H H
R
OS
ACP
HO
Me
+N
O
NH2
R
H
+KR
Dehydratase (DH):After reduction by KR, DH eliminates water to form an α,β-unsaturated thioester.
ACP
OS
ACP
Me
OS
Me
HHH
HO HDH H
Enoyl Reductase (ER):ER hydrogenates the double bond to form a methylene unit at the β-carbon.
OS
ACP
Me
+N
O
NH2
H H
R
-OS
ACP
Me
+N
O
NH2
R
H
+ER
HH H
H
OS
ACP
Me
HH
HH
Thio Esterase (TE):TE releases the fatty acid or polyketide. In the PKS, the TE may also be responsible for the cyclizationof the macrolide ring.
OS
HO
Ser
+TE
ACP
Me
-OS
ACP
Me
OSer SH
+
ACP
O
SerTE
O
Me
O-O
Me
+ OH
SerTE
TE
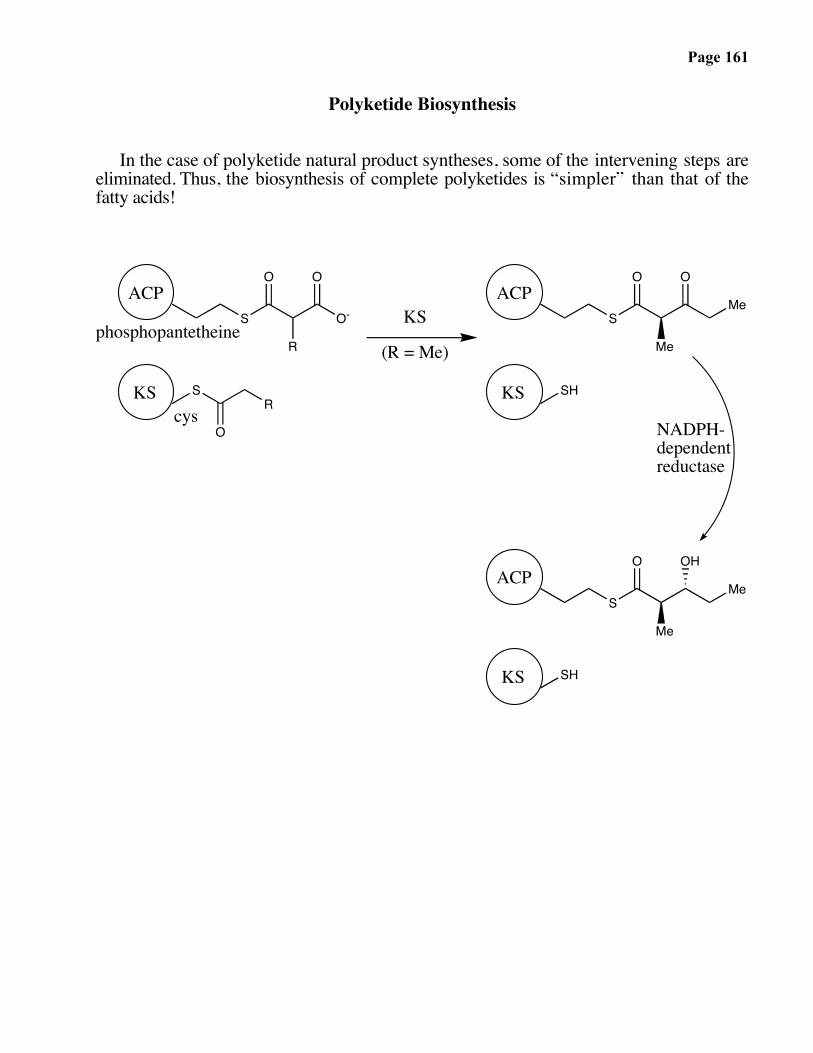
Page 161
Polyketide Biosynthesis
In the case of polyketide natural product syntheses, some of the intervening steps areeliminated. Thus, the biosynthesis of complete polyketides is “simpler” than that of thefatty acids!
S O-
O O
S
O
R
R
S
O O
SH
ACP
phosphopantetheineMe
Me
KS
(R = Me)
S
O OH
ACP
SH
ACPMe
Me
NADPH-dependentreductase
KS
KS
KScys

Page 162

Page 163
Polyketide Diversity

Page 164
Polyketide Biosynthesis
Each cycle may differ in terms of the intervening steps (ketoreduction, dehydration,enone reduction), and in terms of the stereochemistry. This yields enormous diversity.
OH
OH
Me
OH O
Me
O
1 step 0 steps 3 steps
acetate acetatepropionate
butyrate
• Recently, polyketide synthase gene clusters have been identified. Likecyclosporinsynthase, they are high molecular-weight, multi-enzyme polypeptidechains. They can be expressed in convenient bacterial strains and rationally “mixedand matched” to produce “non-natural natural products”. In the future,combinatorial biosynthesis should be possible.
Gene table for the polyketide above:
Module # Starting Material ACP AT KS KR DH ER TE
0
1
2
3
4
5

Page 165

Page 166
Biosynthesis of the polyketide:
HO
OH
Me
OHO
Me
O
direction of synthesis

Page 167

Page 168
6-Deoxyerythronolide B Synthase (DEBS)
SO
MeHO
Et
SO
SO
Me
SO
SO
SO
O
Et S
MeHO
Et
MeHO
OMe
Et
Et
Et
Et
HOMe
HOMe
HOMe
HOMe
HOMe
HOMe
HOMe
O
O
O
Me
Me
Me
Me
Me
Me
HO
HOMe
MeHO
Me
ATACPKSAT KRACP
Module 1 KSAT KRACP
Module 2
DEBS I
O
OH
O
O
KSATDHERKRACP
Module 4
eryAI
Me
Me
Me
Me
Et
KSAT
(KR)ACP
Module 3 KSAT KRACP
Module 5 KSAT KRACP
Me
Me
Module 6
OH
OH
6-Deoxyerythronolide B
DEBS II
eryAII
DEBS III
eryAIII
TE
HO
Donadio, S.; Staver, M. J.; McAlpine, J. B.; Swanson, S. J.; Katz, L. Science 1991, 675-679.
O O
Saccharopolyspora erthyraeaStreptomyces coelicolor
DEBSI TE+Bacillus subtilis ??
MeMeOH
Et
Kao, C. M.; Luo, G.; Katz, L.; Cane, D. E.; Khosla, C. J. Am. Chem. Soc. 1995, 9105-9106.Cortes, J.; Wiesmann, K. E. H.; Roberts, G. A.; Brown, M. J. B.; Staunton, J.; Leadley, P. F. Science1995, 268, 1487-1488.
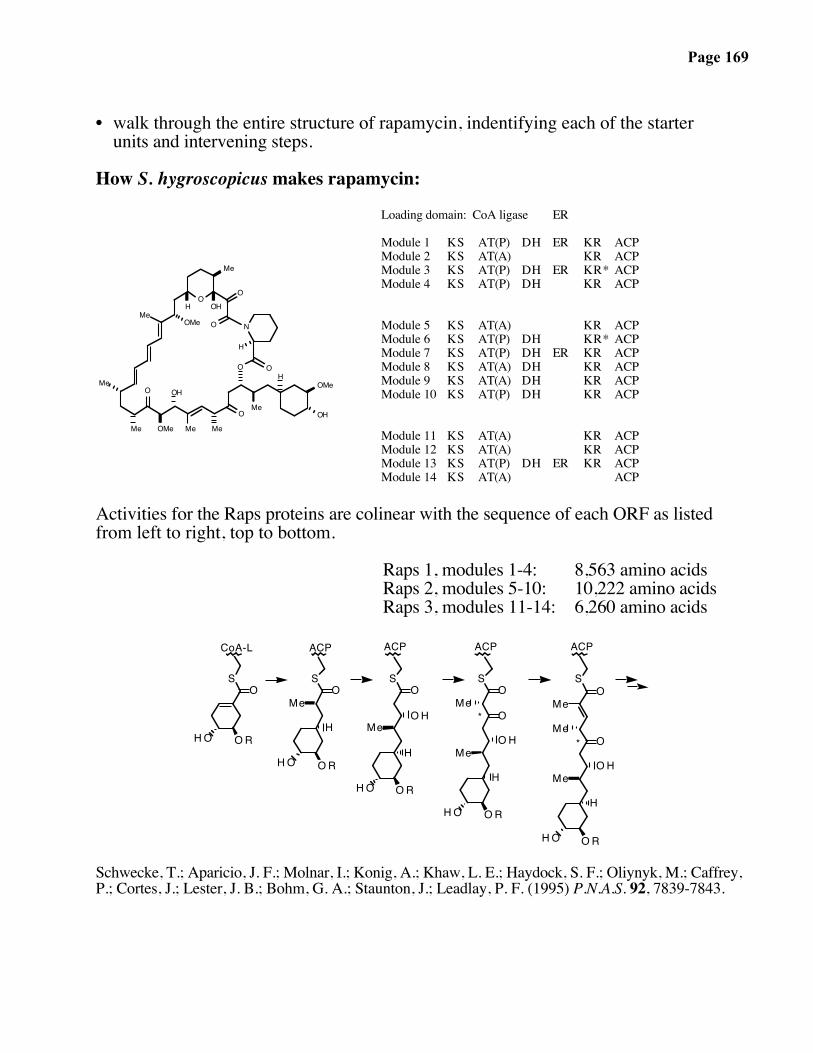
Page 169
• walk through the entire structure of rapamycin, indentifying each of the starter units and intervening steps.
How S. hygroscopicus makes rapamycin:
Me
H
O
O
Me
NMe
OMe
Me
OMe
OHMe
H OH
O
O
H
O
Me
Me
OHO
OMe
O
Loading domain: CoA ligase
Module 1Module 2Module 3Module 4
Module 5Module 6Module 7Module 8Module 9Module 10
Module 11Module 12Module 13Module 14
KSKSKSKS
KSKSKSKSKSKS
KSKSKSKS
AT(P)AT(A)AT(P)AT(P)
AT(A)AT(P)AT(P)AT(A)AT(A)AT(P)
AT(A)AT(A)AT(P)AT(A)
DH
DHDH
DHDHDHDHDH
DH
ER
ER
ER
ER
ER
KRKRKR*KR
KRKR*KRKRKRKR
KRKRKR
ACPACPACPACP
ACPACPACPACPACPACP
ACPACPACPACP
Activities for the Raps proteins are colinear with the sequence of each ORF as listedfrom left to right, top to bottom.
Raps 1, modules 1-4: 8,563 amino acidsRaps 2, modules 5-10: 10,222 amino acidsRaps 3, modules 11-14: 6,260 amino acids
ORHO
SO
ORHO
H
S
Me
ORHO
H
Me
S
OH
O O
ORHO
H
Me
S
OH
O
O
ORHO
H
MeOH
O
S
Me
Me
MeO
CoA-L ACP ACPACP
*
ACP
*
Schwecke, T.; Aparicio, J. F.; Molnar, I.; Konig, A.; Khaw, L. E.; Haydock, S. F.; Oliynyk, M.; Caffrey,P.; Cortes, J.; Lester, J. B.; Bohm, G. A.; Staunton, J.; Leadlay, P. F. (1995) P.N.A.S. 92, 7839-7843.
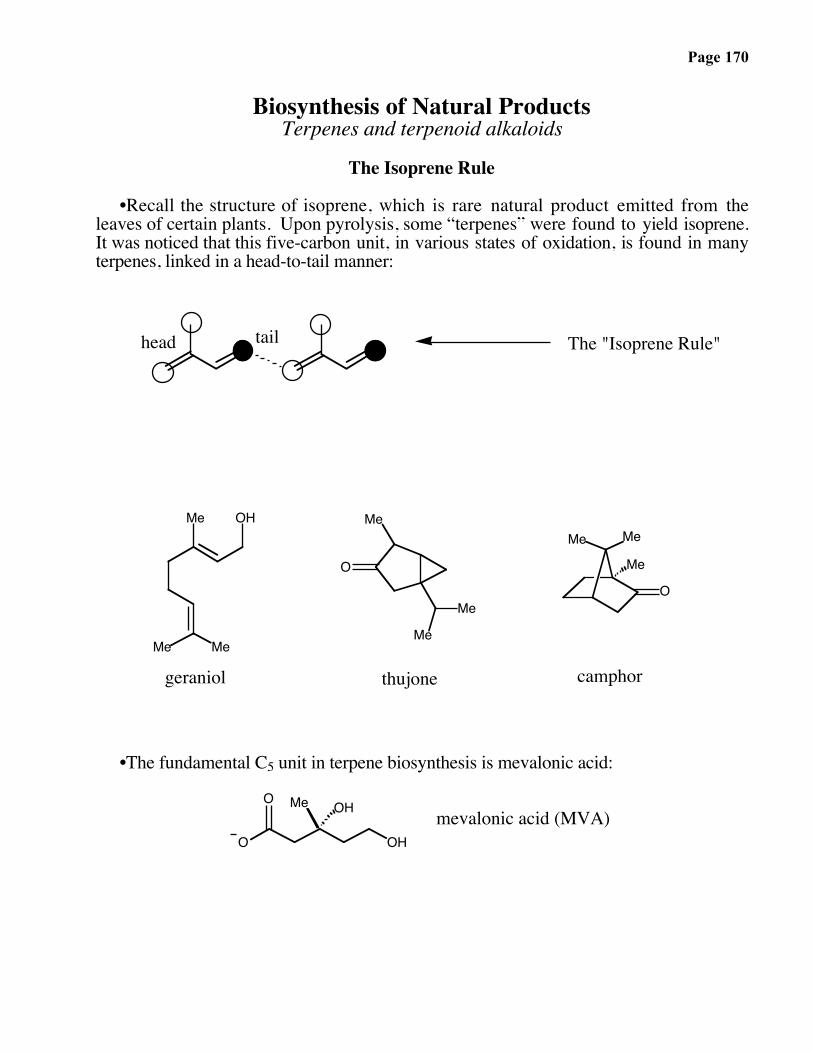
Page 170
Biosynthesis of Natural ProductsTerpenes and terpenoid alkaloids
The Isoprene Rule
•Recall the structure of isoprene, which is rare natural product emitted from theleaves of certain plants. Upon pyrolysis, some “terpenes” were found to yield isoprene.It was noticed that this five-carbon unit, in various states of oxidation, is found in manyterpenes, linked in a head-to-tail manner:
MeMe
Me OH
O
Me
Me
Me
Me Me
O
The "Isoprene Rule"
Me
geraniol thujone camphor
tailhead
•The fundamental C5 unit in terpene biosynthesis is mevalonic acid:
O OH
O Me OH mevalonic acid (MVA)

Page 171
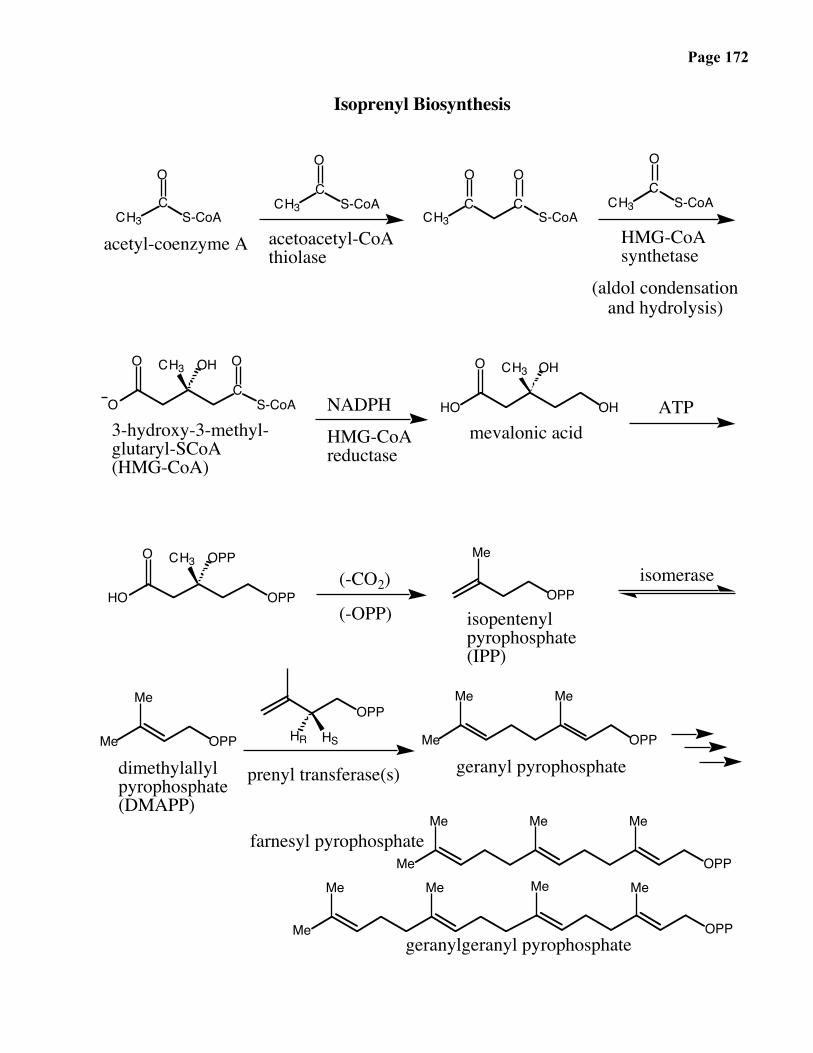
Page 172
Isoprenyl Biosynthesis
O
CCH3 S-CoA
CS-CoACH3
O
CS-CoA
O
CCH3
OO
CCH3 S-CoA
CS-CoA
O
O
CH3 OHO O OHCH3
HO
OPP
OH
HO
CH3 OPPO
OPP
Me
Me OPP
MeOPP
HSHR Me OPP
Me Me
HMG-CoAreductase
acetyl-coenzyme A
(-OPP)
(-CO2)
geranyl pyrophosphateprenyl transferase(s)dimethylallylpyrophosphate(DMAPP)
NADPH
isopentenyl pyrophosphate(IPP)
3-hydroxy-3-methyl-glutaryl-SCoA(HMG-CoA)
HMG-CoAsynthetase
acetoacetyl-CoAthiolase
mevalonic acidATP
isomerase
(aldol condensationand hydrolysis)
Me
Me MeMe
Me Me
Me Me
OPP
Me
OPP
geranylgeranyl pyrophosphate
farnesyl pyrophosphate

Page 173
Terpene Biosynthesis
• Mevacor, a chlosterol-lowering, life-saving drug inhibits the biosynthesis of MVA:
HO
Me
Me
Me
OO
OHO
H
HO
Me
Me
Me
OOH
COO-HO
in vivo H
H2O
active drugmevacor (prodrug)
MVA / HMG mimic

Page 174
Prenyl Transferases
isopentenylpyrophosphate
IPP
OPP
Me isopentenylpyrophosphateisomerase
dimethylallylpyrophosphateDMAPP
Me OPP
Me
IPP DMAPP+Me OPP
Me Meprenyltransferase
geranylpyrophosphate
GPP
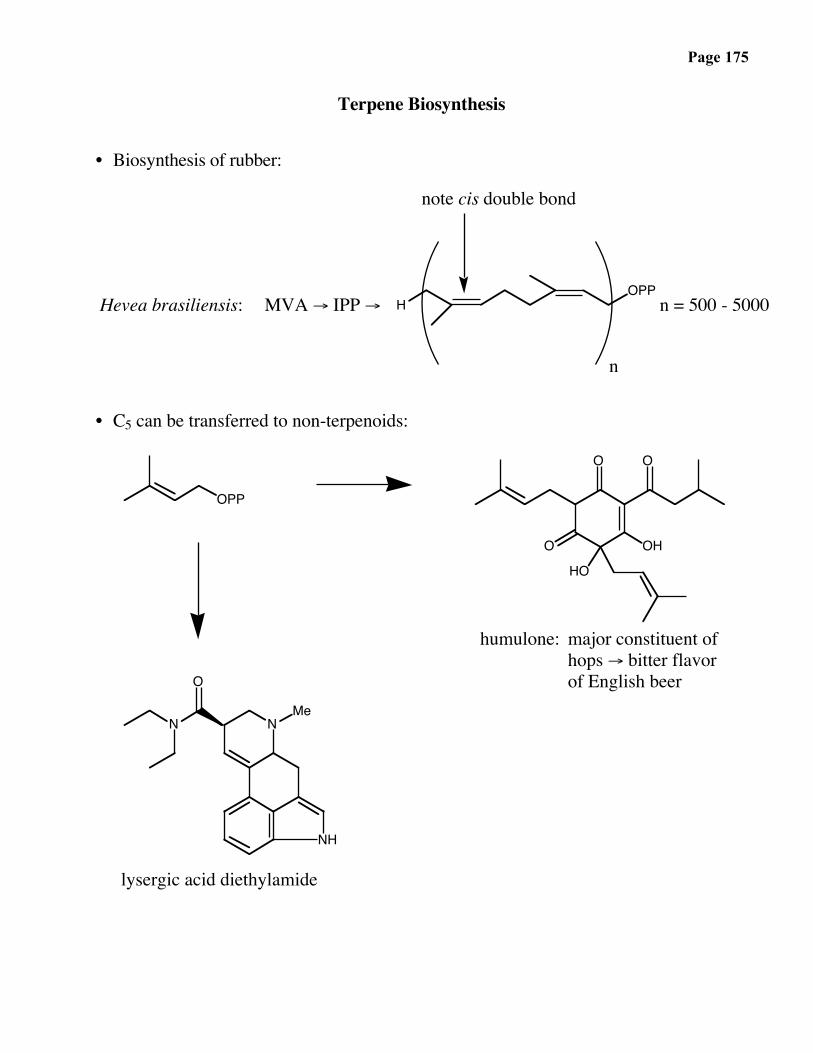
Page 175
Terpene Biosynthesis
• Biosynthesis of rubber:
Hevea brasiliensis: MVA → IPP →
n
n = 500 - 5000OPP
H
note cis double bond
• C5 can be transferred to non-terpenoids:
OPP
O O
O OH
HO
N
O
N
lysergic acid diethylamide
major constituent ofhops → bitter flavorof English beer
humulone:
NH
Me

Page 176

Page 177
Cyclization of IPP Oligomers by Cyclases
• isomerase-cyclase
OPPisomerase
MePPO
cyclase
Me
+

Page 178
Me
+
• FPP cyclases

Page 179
Pentalene Cyclases

Page 180
Stereochemistry — Isotope Labeling
• SN2' reactions can proceed with syn or anti stereochemistry:
MeMe
LG
Me
Me
Me MeNu
MeMe
Me Me
Nu
MeMe
syn
or
anti
• linaloyl pyrophosphate
D
D
MeMe
PPO Me
cyclaseD
MeMeOH
Me
HD

Page 181
How Do Enzymes Stabilize Reactive Carbocations?

Page 182
Squalene Oxide Cyclase
O
squalene oxide
MeMeHO
Me
H
Me
H
Me
cyclase
lanosterol
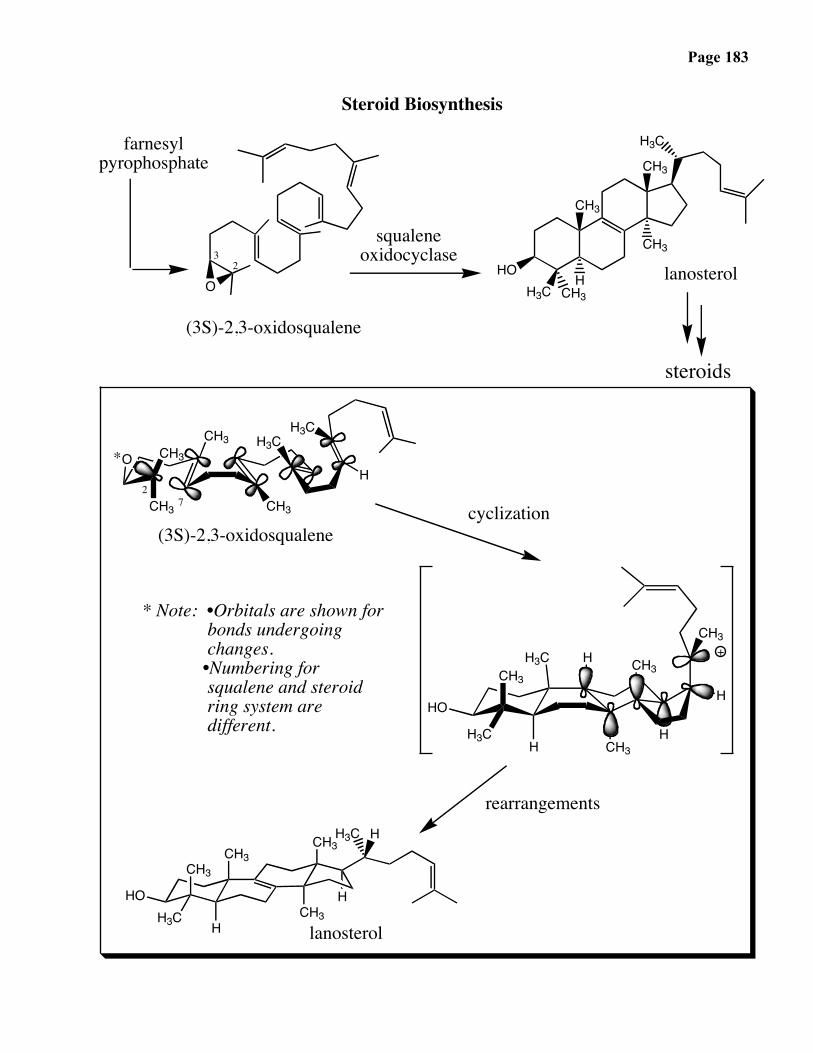
Page 183
Steroid Biosynthesis
O
H3C
H3C CH3
CH3
CH3
CH3HO
H
CH3
CH3CH3
CH3
H3CO
H3C
H
CH3
H3CCH3
H3C
HO
3
H
CH3H
H
2
H
farnesylpyrophosphate
CH3
squaleneoxidocyclase
* Note: •Orbitals are shown for bonds undergoing changes. •Numbering for squalene and steroid ring system are different.
CH3CH3
CH3H3C
CH3HO
(3S)-2,3-oxidosqualene
rearrangements
lanosterol
H
H3C
steroids
7
(3S)-2,3-oxidosqualene
*
2
cyclization
H
H
+
lanosterol

Page 184
(1) Substrate Folding
(2) Initiation of A-ring Cyclization
(3) C-ring Cyclization
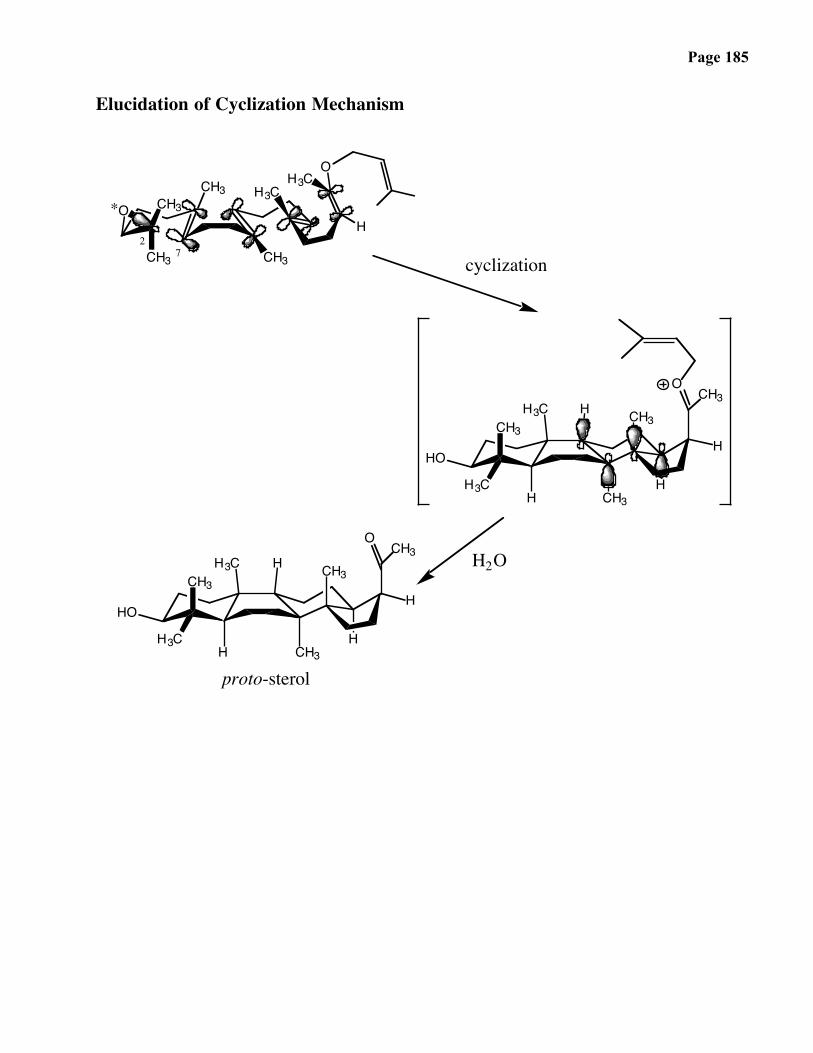
Page 185
Elucidation of Cyclization Mechanism
CH3
CH3CH3
CH3
H3CO
OH3C
H
CH3
H3CCH3
H3C
HO
H
CH3H
H
H
CH3O
7
*
2
cyclization
H2O
proto-sterolCH3
H3CCH3
H3C
HO
H
CH3H
H
H
CH3O

Page 186
(4) Hydride and Methyl Shifts
Question: What does the enzyme have to do once the protosterol cation is formed?

Page 187
The Daphniphyllum Alkaloids
HNH
OO
O
N
NH
OAc
OO
O
O
O
N
OO
H
NAcO
HO
COOMe
H
AcO MeN
COOMe
H
O
OH
seco-daphniphylline
daphnilactone A daphnilactone B
yuzurimine daphnigracine
daphniphylline

Page 188
HNH
proto-daphniphylline
HNH
proto-daphniphylline

Page 189
Daphniphyllum Biosynthesis
• cellular
HNH
proto-daphniphylline

Page 190
The Biosynthesis of proto-DaphniphyllineR. B. Ruggeri and C. H. Heathcock, Pure & Appl. Chem. 61, 289 (1989).
CHO
CHO CHO
N
NH
HO
H3C
OPO32-
CHO
HN
NH
NH-Lys (enz)
NH
Lys-HN NH
O
NH
Lys-HN NH
O
H
PAP
H2N-Lys (enz)
enal-enaminecycloaddition
++
+

Page 191
The Biosynthesis of proto-Daphniphylline (continued)
NH
Lys-HN NH
O
H
NH2
O
H
N
H
NH
N HNH
Lys-PLP
H
+ (from previous page)H2O
proto-daphniphylline

Page 192
The Biomimetic Synthesis of proto-DaphniphyllineS. Piettre and C. H. Heathcock, Science 248, 1532 (1990).
CHO
CHO
CHOCHO O
O
O
H
OH
NH
N
H
NH
NH380°C
KOHroom temp.
intramolecularMichael reaction
HNH
HOAc, 80°Diels-Alder
(E) - dihydrosqualene-dialdehyde
(Z) - dihydrosqualene-dialdehyde
proto -daphniphylline(50%)
one-pot reactionfrom dihydrosqualenedialdehyde





























