Embed Size (px)
Citation preview

Biological Half-lives of Bromide and Sodium in the Rat AreConnected and Dependent
on the Physiological State**ARNOST BABICKY,1 STANISLAV PAVELKA,*,2,3
AND MILOSLAV VOBECKY1
1Institute of Analytical Chemistry and 2Institute of Physiology,Academy of Sciences of the Czech Republic, 142 20 Prague,
Czech Republic; and 3Department of Biochemistry, Faculty ofScience, Masaryk University, 61137 Brno, Czech Republic
Received May 5, 2004; Accepted May 31, 2004
ABSTRACT
The parallel course of the excretion rates of sodium and bromideions was demonstrated in adult male rats administered simultaneouslywith 24Na-sodium chloride and 82Br-bromide. These excretion rates wereinversely proportional to the magnitude of sodium intake in the ani-mals. The biological half-life of bromide, as a substitute for sodium orchloride, was investigated with the aid of the radionuclide 82Br in ani-mals situated in very different physiological states (i.e., in lactating andnonlactating female rats as well as in young rats of varying ages [2, 4, 6,and 10 wk of age]). The 82Br radioactivity retained in mothers and inwhole litters was measured in vivo at appropriate time intervals (up to240 h) after the application of 82Br-bromide to the mothers. The time-course of the changes in the 82Br radioactivity of the young was calcu-lated as the difference between the rate of 82Br intake in the mother’smilk and the 82Br excretion through the kidneys into the urine. The rateof 82Br excretion through the kidneys of the dam could be calculatedalso. Nonweaned young rats (12 d) had the highest half-life (269 h) andlactating dams had the lowest (44 h). The determined values demon-strated that nonweaned young apparently conserve sodium, because of
Biological Trace Element Research 49 Vol. 103, 2005
© Copyright 2005 by Humana Press Inc.All rights of any nature, whatsoever, reserved.0163-4984/05/10301–0049 $30.00
* Author to whom all correspondence and reprint requests should be addressed.** Presented in part at the 4th International Symposium on Trace Elements in
Human: New Perspectives held in Athens (Greece) on 9–11 October 2003

its relatively low concentration in mother’s milk, whereas lactatingdams, because of their large food intake, waste sodium.
Index Entries: Biological half-life; bromide; chloride; sodium;radionuclide; rat.
INTRODUCTION
A number of mechanisms are known by which a mammalian organ-ism maintains the constancy of the internal milieu, which is a prerequisitefor good health. The principal electrolytes of the extracellular fluid (ECF)(i.e., the blood plasma plus interestitial fluid) are sodium and chlorideions. The amount of sodium in the body primarily determines the size ofthe ECF volume. The constancy of electrolyte concentration is achieved byactive reabsorption of sodium in kidney tubules, whereas chloride followspassively the movement of sodium.
It has repeatedly been shown that elements from the same group ofthe periodic system exhibit some similarities of distribution and meta-bolic behavior. This is also true for the halide ions. In the 1930s, Mason (1)had recognized that bromide replaces part of the extracellular chloride,the molar sum of chloride, and bromide remaining constant at approx 110mmol/L. The distribution of chloride and that of bromide are indeedclosely analogous and almost exclusively extracellular (2). In the intes-tine, both ions are completely passively absorbed by the paracellularpathway. Bromide has often been used as inert indicator for the closelyrelated chloride in connection with physiological kinetic investigationsand in determinations of the extracellular space (e.g., ref. 3). The similar-ity of bromide to chloride entails an important pharmacokinetic interac-tion: both ions compete for tubular reabsorption. The biological half-lifeof bromide can be decreased by administering surplus halide (e.g., chlo-ride) ions (4). On the other hand, the already long half-life of bromide,which is 12 d in humans (5) and approx 3–8 d in the rat (6,7), might beincreased considerably by a salt-deficient diet. In the rat, the bromidehalf-life was prolonged to 25 d on a salt-free diet (6). This finding wasinterpreted by the authors as a marked dependence of the biological half-life of bromide on chloride concentration in the diet. However, keeping inmind the differences between the metabolism of sodium and chloride(sodium is reabsorbed in the kidneys actively whereas chloride is reab-sorbed passively) and the fact that sodium excretion rate depends on themagnitude of sodium intake, we hypothesized that the biological half-lifeof bromide depends on the magnitude of sodium intake rather than onthe intake of chloride.
In order to test our hypothesis, we determined in the present studiessimultaneously the biological half-lives of sodium (with the aid ofradionuclide 24Na) and bromide (with the aid of radionuclide 82Br). From
50 Babicky, Pavelka, and Vobecky
Biological Trace Element Research Vol. 103, 2005

the point of view of the animal’s organism, with regard to the necessaryeffort at maintaining constancy of the internal milieu, the most criticalperiods in the life of mammals are early postnatal development and theperiod of lactation. To demonstrate the dependence of the biological half-lives of bromide and sodium on the physiological state of the animals, wedetermined the values of these half-lives in young rats of varying ages aswell as in lactating and nonlactating female rats.
MATERIALS AND METHODS
Animals and Diets
Experiments were performed on Wistar rats of both sexes and vari-ous ages. Animals of the experimental groups in the first series of experi-ments were fed a special low-sodium pelleted diet (Bergman, Kocanda,Czech Republic) with a mean concentration of bromine of 4.0 ± 0.7mg/kg, chlorine of 751 ± 233 mg/kg, and sodium of 296 ± 35 mg/kg. Allrats in the second and the third experimental series and control group ofrats in the first series of experiments were fed a standard pelleted diet(Bergman, Kocanda, Czech Republic) with a mean concentration ofbromine of 9.7 mg/kg, that of chlorine of 2900 mg/kg, and that of sodiumof 1696 ± 138 mg/kg. All of the animals drank tap water. During the entireexperimental period, diet and drinking water were provided to the ani-mals ad libitum. Concentrations of bromine, chlorine, and sodium in thediets were determined by short-term instrumental neutron activationanalysis (8). The radionuclides 24Na and 82Br were prepared by irradiat-ing the NaCl and KBr targets with neutrons in the core of the LWR-15research nuclear reactor (Nuclear Research Institute, Rez near Prague,Czech Republic) (9).
Simultaneous Determination of Biological Half-lives of Sodium and Bromide
The first experimental series included one control and five experi-mental groups, each containing five adult male rats. The control animalsdrank distilled water and the rats in experimental groups drank distilledwater with the addition of sodium chloride with increasing final concen-trations of 0 to 50 to 100 to 150 to 200 mmol NaCl/L. On d 14, all of the ani-mals, including rats of the control group, were given simultaneouslyapprox 3 MBq 82Br in the form of potassium bromide and 4.5 MBq 24Na inthe form of sodium chloride in 0.3 mL saline by subcutaneous injection.The retained radioactivity was measured in vivo and the biological half-lives of bromide and sodium in these animals were determined by using aprocedure described next (in the case of lactating dams).
Biological Half-lives of Electrolytes in the Rat 51
Biological Trace Element Research Vol. 103, 2005

Determination of Biological Half-life of Bromide in Lactating Rat Dams and Their Young
In the second experimental series, six lactating rats kept with eightyoung each were used. On d 12 after delivery, the dams were givenapprox 3 MBq 82Br in the form of potassium bromide (microgramamounts) in 0.3 mL saline by subcutaneous injection. The radioactivityretained in the mothers and in the whole litters was measured in vivo bymeans of a computerized γ-spectrometric system equipped with a HPGedetector (Canberra). Details of the whole-body measurement of radioac-tivity are given in an accompanying article (10). The time-course of thechanges in the whole-body 82Br radioactivity of the young was calcu-lated as the difference between the rate of the 82Br intake throughmother’s milk and the 82Br excretion through the kidneys according tothe equation
(1)
where RYt is the amount of 82Br radioactivity retained in the young at timet expressed as the percent of the dose applied to the mother, AY is the max-imum amount of the 82Br radioactivity that can be transferred to the youngthrough mother’s milk under the given experimental conditions expressedas the percent of the dose applied to the mother, Tex is the half-life of the82Br excretion through the kidneys of the young (given in hours), Ti is thehalf-life of the 82Br intake by the young through mother’s milk (given inhours), and t is the time that elapsed from the moment of the 82Br applica-tion to the mother (given in hours).
The time-course of the changes in the whole-body 82Br radioactivity ofthe lactating dam can be expressed by the equation
(2)
where RDt is the 82Br radioactivity retained in the mother at time texpressed as the percent of the applied dose AD, Tel is the half-life of the82Br elimination from the maternal body (given in hours), and t has thesame meaning as in Eq. (1).
The rate of 82Br radioactivity elimination in the mother’s milk, Mt, canbe given as the amount of 82Br radioactivity taken by the young:
(3)
If all other methods of bromide elimination from the maternal body(e.g., salivary gastroenteric circulation) are neglected, the amount of 82Br
M At
Tt Y
i= − −⎛
⎝⎜⎞⎠⎟
⎡
⎣⎢
⎤
⎦⎥1
0 693exp
.
R At
TDt D= −⎛
⎝⎜⎞⎠⎟
⎡
⎣⎢
⎤
⎦⎥exp
.0 693
el
R At
Tt
TYt Y
i= −⎛
⎝⎜⎞⎠⎟
⎡
⎣⎢
⎤
⎦⎥ − −⎛
⎝⎜⎞⎠⎟
⎡
⎣⎢
⎤
⎦⎥
⎧⎨⎪
⎩⎪
⎫⎬⎪
⎭⎪exp
.exp
.0 693 0 693
ex
52 Babicky, Pavelka, and Vobecky
Biological Trace Element Research Vol. 103, 2005

radioactivity excreted from the body of lactating dam through the kidneyscan be given as the difference between the applied dose of 82Br radioactiv-ity (AD) and the 82Br content in mother’s milk plus the retained radioac-tivity in the body. On the basis of the values calculated in this way, the rateof 82Br radioactivity excretion through the kidneys of the dam can beexpressed by the equation
(4)
where Ut is the amount of 82Br radioactivity in the urine of the lactatingdam at time t expressed as the percentage of the dose applied to the dam,UD is the maximum amount of 82Br radioactivity that can be excreted in theurine of the dam under the given experimental conditions expressed as thepercentage of the applied dose, Texc is the half-life of 82Br excretion throughthe kidneys of the lactating dam (given in hours), and t has the same mean-ing as in Eq. (1). It is evident that UD = AD – AY.
The relevant parameters of all the equations were found by fitting themeasured or calculated values to the curves using the least squares method.
Determination of Biological Half-life of Bromide in Animals Situated in Diverse Physiological States
In the third series of experiments, four groups of rats were used. Thefirst group contained eight young prematurely weaned at the age of 16 dand one litter (eight young) of the same age kept with the mother. All ofthese young rats received 82Br on the 21st postnatal day. The second groupconsisted of five males and five females at the age of 4 wk; the third groupcomprised the same number of animals at the age of 6 wk. Finally, fivefemales at the age of 10 wk were used. The biological half-lives of bromidein these animals were determined by using the same procedure as in thecase of lactating dams.
RESULTS
Simultaneous Determination of Biological Half-lives of Sodium and Bromide
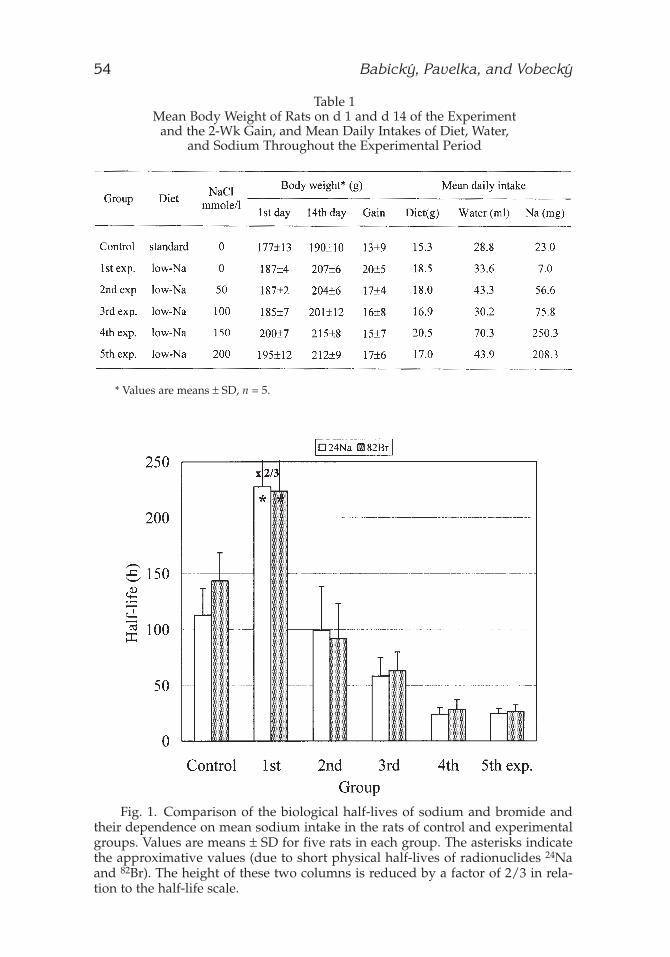
The results of the first experimental series are summarized in Table 1and Fig. 1. Table 1 presents the mean body weight of rats of control andfive experimental groups at the beginning and on d 14 of the experimentand the 2-wk gains. The differences in the body weight gain among thesegroups were not significant at the level of p = 0.05. The mean daily intakesof food, drinking water (containing various concentrations of NaCl), andsodium ions throughout the experimental period are also given. An excep-
U Ut
Tt D= − −⎛
⎝⎜⎞⎠⎟
⎡
⎣⎢
⎤
⎦⎥1
0 693exp
.
exc
Biological Half-lives of Electrolytes in the Rat 53
Biological Trace Element Research Vol. 103, 2005
54 Babicky, Pavelka, and Vobecky
Table 1Mean Body Weight of Rats on d 1 and d 14 of the Experiment
and the 2-Wk Gain, and Mean Daily Intakes of Diet, Water, and Sodium Throughout the Experimental Period
* Values are means ± SD, n = 5.
Fig. 1. Comparison of the biological half-lives of sodium and bromide andtheir dependence on mean sodium intake in the rats of control and experimentalgroups. Values are means ± SD for five rats in each group. The asterisks indicatethe approximative values (due to short physical half-lives of radionuclides 24Naand 82Br). The height of these two columns is reduced by a factor of 2/3 in rela-tion to the half-life scale.
tionally high consumption of water was noticed in the rats of the fourthexperimental group, resulting in about a 10-fold higher intake of sodiumin these animals in comparison with the control rats. Figure 1 compares theaverage values of simultaneously determined biological half-lives ofsodium and bromide ions in the rats of the individual groups. The half-lives of both electrolytes, in parallel, dramatically decreased with increas-ing intake of sodium in the animals.
Biological Half-life of Bromide in the Lactating Dams and Their Young
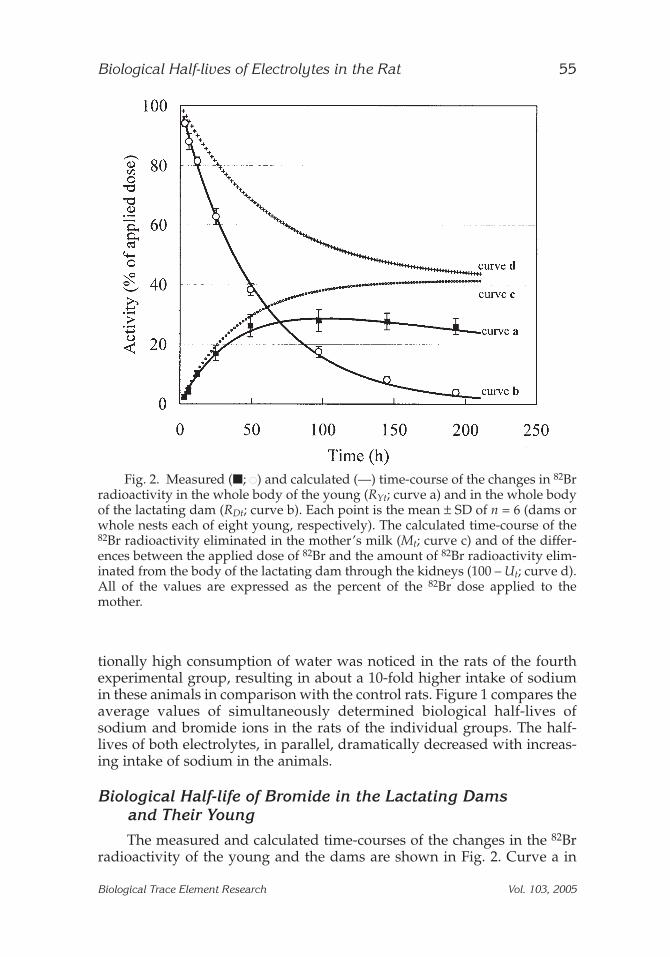
The measured and calculated time-courses of the changes in the 82Brradioactivity of the young and the dams are shown in Fig. 2. Curve a in
Biological Half-lives of Electrolytes in the Rat 55
Biological Trace Element Research Vol. 103, 2005
Fig. 2. Measured (■; ●) and calculated (—) time-course of the changes in 82Brradioactivity in the whole body of the young (RYt; curve a) and in the whole bodyof the lactating dam (RDt; curve b). Each point is the mean ± SD of n = 6 (dams orwhole nests each of eight young, respectively). The calculated time-course of the82Br radioactivity eliminated in the mother’s milk (Mt; curve c) and of the differ-ences between the applied dose of 82Br and the amount of 82Br radioactivity elim-inated from the body of the lactating dam through the kidneys (100 – Ut; curve d).All of the values are expressed as the percent of the 82Br dose applied to themother.

Fig. 2 depicts the time-course of the whole-body 82Br radioactivity of theyoung (RYt) and represents the differences between the amount of 82Brtransferred through mother’s milk into the body of the young and theamount of 82Br excreted through the kidneys of the young. Curve b showsthe time-course of the whole-body 82Br radioactivity of the lactating dam(RDt) and represents the differences between the applied dose of 82Br (AD)and the totals of the amount of 82Br eliminated in the milk (Mt) and theamount of 82Br excreted through the kidneys of the dam (Ut). In both cases,a good agreement can be seen between the experimentally found valuesand the calculated curves. Parameters that comply with Eq. (1) are: AY =41.5 ± 3.1%, Ti = 27.6 ± 1.5 h, and Tex = 269.1 ± 22.4 h. The parameter com-plying with Eq. (2) is Tel = 37.3 ± 2.0 h. Curve c shows the calculated time-course of the 82Br radioactivity elimination in mother’s milk, and curve ddepicts the differences between the applied dose of 82Br (AD) and theamount of 82Br eliminated from the body of the lactating dam through thekidneys into the urine (100 – Ut). The calculated value of UD = 58.2 ± 3.3%is in very good agreement with the theoretical value of 58.5%. The calcu-lated value of the half-life of the 82Br excretion through the kidneys of thelactating dam is Texc = 44.3 ± 4.5 h.
Biological Half-life of Bromide in Animals Situated in Diverse Physiological States
All of the determined half-life values of the 82Br excretion through thekidneys into the urine are summarized in Table 2. The values were calcu-lated from the in vivo measured time-courses of changes in the retained82Br radioactivity in animals of various ages and situated in diverse phys-iological states, in the same way as in the case of lactating dams.
56 Babicky, Pavelka, and Vobecky
Biological Trace Element Research Vol. 103, 2005
Table 2Half-lives (h) of 82Br-Bromide Excretion Through the Rat Kidneys
* Young prematurely weaned on the 16th postnatal day.** Lactating dams.

DISCUSSION
Data on the rate of urinary excretion of bromide obtained in thesestudies confirm the well-recognized augmentation of urinary bromidewhen dietary chloride increases. According to Hellerstein et al. (3), total-body chloride can be followed for a considerable time after a single doseof bromide. This conclusion follows from the evidence that bromide isexcreted slowly and almost entirely in the urine. Differences between bro-mide and chloride ions are rather subtle. The most important one lies intheir passive transport: bromide is transported more readily. This leads tolarger bromide/chloride ratios in saliva and gastric juice than in plasmaand, conversely, to lower bromide/chloride ratios in urine (4). Because therat tissues show consistent agreement between the ratios of chloride tobromide in serum, liver, skin, and muscle (3), we suppose that the use of82Br-bromide for following the excretion rate of chloride is justified. Inaddition, because of the close relation of sodium and chloride, the excre-tion rate of sodium could also be followed by using 82Br-bromide. Theresults of the simultaneous determination of biological half-lives ofsodium and bromide in the rats maintained on various sodium intakes(Fig. 1) verify this hypothesis.
Differences in the determined excretion rates of bromide in animalssituated in diverse physiological states (Table 2) seem, at first sight, to besurprisingly large. However, if all of the features characteristic for the par-ticular stage of life of the rat are taken into account, the differences can beexplained by an effort of the animal’s organism to preserve the constancyof the internal milieu. During the period of early postnatal life, the youngrat grows very rapidly. Because the concentration of sodium and, there-fore, of chloride in rat milk is relatively low, almost all of these electrolytesreceived through mother’s milk have to be retained in the body of theyoung, in order to preserve the constancy of the electrolyte composition ofthe ECF. During the weaning period, when the young gradually pass froma pure milk diet to the standard laboratory diet with greater content ofthese electrolytes, the need to retain all of them slowly ceases. It is knownthat an increase in the salt content in food leads to a decrease in the secre-tion of aldosterone and, therefore, to a decrease in sodium reabsorption inthe tubules, and vice versa. This explanation is also supported by theresults of experiments with prematurely weaned young whose consump-tion of solid food was greater than that of the young of the same age butkept with their mother (Table 2). During further periods of life, the excre-tion rate of electrolytes depends on the metabolic activity and, conse-quently, on food and salt intake. A good example of this can be foundduring the period of lactation. In the course of the first 3 wk of nursing, thegrowth of the young is secured mostly by the intake of mother’s milk. Dur-ing this period of nursing, a lactating dam with a litter of eight youngalmost doubles her body weight, the greater part of this increment appear-ing in the mass of her young. The dam has to substantially enhance her
Biological Half-lives of Electrolytes in the Rat 57
Biological Trace Element Research Vol. 103, 2005

intake of food. The excretion rate of these electrolytes has to be substan-tially accelerated because only a smaller part of the electrolytes receivedbecause of the enhanced food intake passes into milk.
In conclusion, the very large differences among the values of the bio-logical half-life of bromide determined in the rats situated in diverse phys-iological states obviously reflect a struggle of the animal’s organism formaintaining constancy of the internal milieu. Using the 82Br-bromideseems to be adequate not only for the determination of the biological half-life of bromide but also of some other related electrolytes, such as chlorideand sodium.
ACKNOWLEDGMENT
This work was supported by the Academy of Sciences of the CzechRepublic (research project no. AVOZ 5011922).
REFERENCES
1. M. F. Mason, Halide distribution in body fluids in chronic bromide intoxication, J. Biol.Chem. 113, 61–73 (1936).
2. S. Ullberg, L. E. Appelgren, C. J. Clemedson, et al., A comparison of the distribution ofsome halide ions in the body, Biochem. Pharmacol. 13, 407–412 (1964).
3. S. Hellerstein, C. Kaiser, D. D. Darrow, and D. C. Darrow, The distribution of bromideand chloride in the body, J. Clin. Invest. 39, 282–287 (1960).
4. A. G. Rauws, Pharmacokinetics of bromide ion—an overview, Food Chem. Toxicol. 21,379–382 (1983).
5. R. Söremark, The biological half-life of bromide ions in human blood, Acta Physiol.Scand. 50, 119–123 (1960).
6. A. G. Rauws and M. J. van Logten, The influence of dietary chloride on the bromideexcretion in the rat, Toxicology 3, 29–32 (1974).
7. S. Pavelka, A. Babicky, M. Vobecky, J. Lener, and E. Svandová, Bromide kinetics and dis-tribution in the rat. I. Biokinetics of 82Br-bromide, Biol. Trace Element Res. 76, 57–66(2000).
8. M. Vobecky, A. Babicky, S. Pavelka, and J. Lener, Determination of bromine and iodinein the rat thyroid by short-term INAA, J. Trace Microprobe Tech. 18, 467–473 (2000).
9. M. Vobecky, A. Babicky, J. Lener, and S. Pavelka, Biological half-life of bromine in therat thyroid, Physiol. Res. 46, 385–389 (1997).
10. M. Vobecky, S. Pavelka, and A. Babicky, Bromide transfer through mother’s milk andits impact on the suckling rat, Biol. Trace Element Res. 103, 37–48 (2005).
58 Babicky, Pavelka, and Vobecky
Biological Trace Element Research Vol. 103, 2005





























