Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/gca
ScienceDirect
Geochimica et Cosmochimica Acta 208 (2017) 354–367
Biological and physical modification of carbonatesystem parameters along the salinity gradient inshallow hypersaline solar salterns in Trapani, Italy
Yuta Isaji a,⇑, Hodaka Kawahata a, Junichiro Kuroda a, Toshihiro Yoshimura a,Nanako O. Ogawa b, Atsushi Suzuki c, Takazo Shibuya d,
Francisco J. Jimenez-Espejo b, Stefano Lugli e, Andrea Santulli f, Vinicio Manzi g,Marco Roveri g, Naohiko Ohkouchi b
aAtmosphere and Ocean Research Institute, University of Tokyo, 5-1-5 Kashiwanoha, Kashiwa, Chiba 277-8564, JapanbDepartment of Biogeochemistry, Japan Agency for Marine-Earth Science and Technology (JAMSTEC),
2-15 Natsushima, Yokosuka 237-0061, JapancGeological Survey of Japan, National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba Central 7, 1-1-1
Higashi, Tsukuba, Ibaraki 305-8567, JapandDepartment of Subsurface Geobiological Analysis and Research, Japan Agency for Marine-Earth Science and Technology (JAMSTEC),
2-15 Natsushima, Yokosuka 237-0061, JapaneDipartimento di Scienze Chimiche e Geologiche, Universita degli Studi di Modena e Reggio Emilia, Via Campi 103, 41125 Modena, Italy
f Istituto di Biologia Marina, Consorzio Universitario della Provincia di Trapani, Via Barlotta Giuseppe 4, 91100 Trapani, ItalygPhysics and Earth Science Department, University of Parma, Parco Area delle Scienze 157/A, 43124 Parma, Italy
Received 6 June 2016; accepted in revised form 10 April 2017; Available online 18 April 2017
Abstract
We investigated changes in the chemical characteristics of evaporating seawater under the influence of microbial activity byconducting geochemical analyses of the brines and evaporite sediments collected from solar salterns in Trapani, Italy. Themicrobial activity had a substantial effect on the carbonate system parameters. Dissolved inorganic carbon (DIC) was sub-stantially removed from the brine during the course of evaporation from the seawater to the point where calcium carbonateprecipitates, with an accompanying decrease in its carbon isotopic composition (d13CDIC) to as low as �10.6‰. Although theremoval of DIC was due to calcium carbonate precipitation, photosynthesis, and the degassing of CO2(aq) induced by evap-oration, the presence of 13C-depleted d13CDIC in ponds where calcium carbonate precipitates can be attributed to the disso-lution of atmospheric CO2 because of intensive CO2(aq) uptake by photosynthesis, and/or mineralization of organic matterby sulfate reduction. In contrast, d13CDIC increased up to 7.2‰ in the salinity range where halite precipitates, which can beascribed to the domination of the effect of degassing of CO2(aq) under conditions with reduced microbial activity. A gradualdecrease in microbial activity was also reflected in compound-specific d13C of photosynthetic pigments; isotopic fractionationassociated with DIC assimilation increased linearly as the evaporation proceeded, indicating DIC-limited conditions within
http://dx.doi.org/10.1016/j.gca.2017.04.013
0016-7037/� 2017 Elsevier Ltd. All rights reserved.
⇑ Corresponding author.E-mail addresses: [email protected] (Y. Isaji), [email protected] (H. Kawahata), [email protected]
(J. Kuroda), [email protected] (T. Yoshimura), [email protected] (N.O. Ogawa), [email protected] (A. Suzuki),[email protected] (T. Shibuya), [email protected] (F.J. Jimenez-Espejo), [email protected] (S. Lugli), [email protected] (A. Santulli), [email protected] (V. Manzi), [email protected] (M. Roveri), [email protected] (N. Ohkouchi).

Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 355
the microbial mats and gypsum crusts because of restricted DIC diffusion from the overlying brine and/or suppression of pri-mary production at higher salinity.� 2017 Elsevier Ltd. All rights reserved.
Keywords: Hypersaline environment; Carbonate system; Microbial mat; Biogeochemical cycle; Photosynthetic pigments; Compound-specificisotope analysis
1. INTRODUCTION
Salinity is one of the most critical environmental factorsdetermining habitability of aquatic environment. Althoughsalinity is potentially a strong limiting factor for habitabil-ity, hypersaline environments are populated by a surprisingdiversity of microorganisms, especially in shallow settings,where benthic microbial mats form (e.g. Oren, 2002; Leyet al., 2006; Oren et al., 2009). It therefore follows that var-ious biological processes are actively operating in the shal-low hypersaline environment, strongly influencing thebiogeochemical cycles and chemical characteristics of thesystem. In addition, seawater evaporation induces transi-tions in the state and composition of the microbial commu-nity through changes in various environmental factors(salinity, temperature, pH, light conditions, etc.). Thesechanges result in modifications of the biological processes,which in turn strongly affect the environment. For thesereasons, the chemical characteristics of the evaporating sea-water are determined not only by physical and chemicalprocesses induced by evaporation, but also by biologicalprocesses within the system. In this study, we focused onthe solar salterns of Trapani (Sicily, Italy) to increase ourunderstanding of the mutual interaction between physical,chemical, and biological processes with increasing salinityin hypersaline environment.
Solar salterns consist of a series of shallow ponds, nor-mally less than 1 m deep, affording a large surface areafor evaporation, with salinity increasing from seawater upto the saturation point of halite (NaCl). Different types ofevaporite minerals precipitate on the bottom of the pondsaccording to the degree of evaporation (Logan, 1987;Geisler-Cussey, 1997). Calcium carbonate (CaCO3, calciteor aragonite) starts to precipitate in ponds in which theevaporation of the original seawater exceeds 50%. Whenover 80% of the original seawater has been removed byevaporation, gypsum (CaSO4�2H2O) starts to precipitate.Benthic microbial mats usually form in the salinity rangewithin which calcium carbonate and gypsum precipitate.Halite starts to precipitate when evaporation exceeds 90%of the original seawater. Various K-Mg salts precipitateafter halite. There is no benthic microbial community inthese highly evaporated ponds, but there are planktonicmicroorganisms in halite crystallizer ponds (e.g. Antonet al., 2000; Rehakova et al., 2009). The evaporationchanges not only the chemical composition but also thephysical properties of the brine: e.g., it decreases the solu-bility and diffusion of dissolved inorganic carbon (DIC;Raven, 1991) and changes the activity coefficients of ionsthrough increasing ionic strength (Karcz and Zak, 1987).
One particular characteristic of these shallow hyper-saline environment is the formation of a highly productivemicrobial mat. The hypersaline microbial mats formed atthe bottom of the ponds are inhabited by highly diversegroups of microorganisms: cyanobacteria (e.g., Greenet al., 2008), chemotrophic and phototrophic sulfur-oxidizing bacteria (e.g., Ollivier et al., 1994; Imhoff,2001), and sulfate-reducing bacteria (e.g., Canfield andDes Marais, 1993; Risatti et al., 1994; Teske et al., 1998;Baumgartner et al., 2006). These groups are dominant inmany of the hypersaline microbial mats at various sites,and together with less abundant but highly diverse groupsof microorganisms they form a complex community struc-ture. This extreme diversity is produced by the broad nichespace provided by the light gradient and varying chemicalconditions within the mat, which itself is modified by bio-logical processes of the microorganisms (Ley et al., 2006).
Carbon, sulfur, and oxygen cycles within the mat clearlyillustrate the mutual interaction among the microbial com-munities via biological modification of the chemical condi-tions (Van Gemerden, 1993). For example, primaryproduction by photoautotrophs generates the organic car-bon that fuels the entire ecosystem, but at the same timereleases oxygen, which is toxic to anaerobes. The fixed car-bon is degraded by fermenters and mineralized to DIC byheterotrophs, sulfate-reducing bacteria and, in some cases,methanogens (e.g., Van Gemerden, 1993; Orphan et al.,2008). This efficient recycling of carbon inside the mataccounts in part for its high primary productivity (e.g.,Canfield and Des Marais, 1993; Des Marais, 2003;Kovac, 2009). On the other hand, sulfide produced by sul-fate reduction is toxic to aerobic microorganisms, but isoxidized back to sulfate, biotically by chemotrophic andphototrophic sulfur bacteria and abiotically by oxygen pro-duced during photosynthesis (e.g., Revsbech et al., 1983;Frund and Cohen, 1992; Canfield and Des Marais, 1993).The activity of these biological processes fluctuates on adaily cycle controlled by light availability (e.g., Canfieldand Des Marais, 1993).
Because most hypersaline evaporative settings in naturalenvironments harbor a microbial community, it is of criticalimportance to understand the responses of biological pro-cesses to increasing salinity. As described above, the physi-cal and biological processes associated with the evaporationof seawater have the potential to affect essential elementssuch as carbon, nitrogen, oxygen, and sulfur. Here, wespecifically focus on changes in the carbonate system, whichconstitutes a fundamental part of the biogeochemical cycle.The amount and chemical form of DIC, which is a resourcefor autotrophs, play a key role in biological processes. DIC

356 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
concentrations in continental aquatic systems are main-tained by water–atmosphere CO2 exchange, precipitationand dissolution of minerals, photosynthesis, respiration,and external inputs such as soil CO2 (e.g., Lazar andErez, 1992). The carbon cycle in the ocean, which has thelargest reservoir of DIC, also affects continental aquaticsystems by controlling the atmospheric CO2 level. Thereare distinct inorganic and biological controls on the carbonbudget and the relative proportions of the three major dis-solved carbon forms—aqueous carbon dioxide (CO2(aq)),bicarbonate (HCO3
�), and carbonate ion (CO32�)—in an
aquatic system. Thus, the interplay between changes in pre-cipitating salts and microbial communities is key to under-standing the changes in the carbonate system duringevaporative concentration processes.
Here, we focus on the water chemistry as well as on theconcentration and isotope signature of DIC in salterns,with the aim of gaining a comprehensive understanding ofcarbon dynamics in the shallow hypersaline environment.We also investigate changes in primary productivity alongthe salinity gradient by performing a compound-specificisotope analysis of photosynthetic pigments. We expectthe resulting insights to also be beneficial as basic informa-tion for understanding the massive evaporation eventsknown to have occurred repeatedly worldwide in the geo-logical past (e.g., Hay et al., 2006; Warren, 2010).
2. MATERIALS AND METHODS
2.1. Study site
We studied three commercial solar salterns located inTrapani (Western Sicily, Italy): the Sosalt (SS), Culcasi(CU), and Chiusicella (CH) salterns (Fig. 1). These solarsalterns, each consisting of multiple ponds with differentsalinities, differ in scale; Sosalt is the largest, with a totalsurface area of 800 ha and an annual production of saltreaching 1 � 105 tons, and Chiusicella is the smallest inboth surface area (7 ha) and number of ponds.
Progressively increasing salinities characterize each ser-ies of ponds, and the corresponding evaporite minerals that
Fig. 1. Locations of the solar salterns investigated in this study.The aerial image is from Google Earth.
precipitate at the bottom. The ponds where calcium carbon-ate precipitates (carbonate ponds) are characterized by theformation of a dense benthic microbial mat. This microbialmat consists of a slimy layer a few millimeters thick, whichis composed of thin yellow, green, and pink layers on thesurface, and black, loose deposits buried underneath(Fig. 2a and b). The gypsum ponds have a thick layer ofgypsum precipitates, which consists of striking stratifiedsolid layers of different colors—yellowish transparent,green, and pink layers, from the surface of the precipitateto a depth averaging around 5 cm—with loose black depos-its below (Fig. 2c and d). Large halite crystals(Fig. 2e and f) form in the subsequent halite ponds. Thereare apparently no benthic microbial communities in thehalite ponds.
2.2. Sampling protocols
Normal seawater, brine, and deposits in the ponds werecollected during the daytime in September 2015 (Table 1).Seawater and brine samples were collected in 100-mL poly-acrylonitrile (PAN) bottles. Those samples collected for themeasurement of total alkalinity (TA), DIC concentration,and DIC carbon isotopic composition (d13CDIC) wereimmediately poisoned with 200 mL of saturated HgCl2 solu-tion to prevent further biological activity. The lid wasclosed without headspace until the analysis to prevent fur-ther gas exchange with the atmosphere. The temperatureand pH of brine and seawater were measured in situ usinga pH meter with a combination electrode (GST-5741C;DKK-TOA Corporation, Tokyo, Japan). The effect of tem-perature on pH was calibrated using the equation ofGieskes (1969). The pH values are given using the seawaterhydrogen ion (SWS) scale. Brine and seawater samples werekept cool in a refrigerator until analysis.
Samples of pond deposits were collected by hand or byusing a hammer and chisel. Microbial mats were collectedfrom three ponds (SS-3, CU-1, and CU-2); small gypsumcrystals were found in the deposits from SS-3. Gypsumcrusts were collected from three ponds (SS-1, SS-2, andCH-1), and halite crystals from two ponds (SS-4 and CU-5). Samples were stored in a freezer until analysis.
2.3. Brine and seawater sample analysis
2.3.1. Salinity
Salinity was measured by using a digital laboratory sali-nometer at the National Institute of Advanced IndustrialScience and Technology, Japan (AIST) (Digi-Auto model5, Tsurumi-Seiki Co., Kanagawa, Japan). Standard seawa-ter (International Association for the Physical Sciences ofthe Ocean [IAPSO]) was used as a reference. Analytical pre-cision was within ±0.01 salinity unit.
2.3.2. Ion concentrations
Brine samples were diluted on a weight basis with ultra-pure water prior to analysis. Concentrations of Na+, Mg2+,and K+ were measured by ion chromatography at theJapan Agency for Marine-Earth Science and Technology(JAMSTEC) (Dionex ICS-1600, Thermo Fisher Scientific,

Fig. 2. Photographs showing the appearance of the ponds and the bottom deposits. (a) Carbonate pond; (b) bottom deposit of carbonatepond showing a slimy layer a few millimeters thick, which is composed of thin yellow, green, and pink layers on the surface, and black, loosedeposits buried underneath; (c) gypsum pond; (d) gypsum crust from gypsum pond showing yellowish transparent, green, and pink layers,from the surface, and loose black deposits below; (e) halite pond; (f) halite crystals from halite pond. (For interpretation of the references tocolor in this figure legend, the reader is referred to the web version of this article.)
Table 1Concentration of dissolved inorganic carbon (DIC) and its carbon isotopic composition (d13CDIC), total alkalinity (TA), and pH of seawaterand brine from the Culcasi (CU), Sosalt (SS), and Chiusicella (CH) solar salterns in Trapani, Sicily, Italy. Numbers in the sample names referto individual ponds at the salterns. SWS, seawater pH scale.
Sample Type of evaporite DIC (mmol L�1) TA (mmol L�1) d13CDIC (‰) pH (SWS)
CU-0 Seawater 2.04 2.68 2.2 8.2CU-1 Carbonate 1.25 3.40 �5.0 8.5CU-2 Carbonate 1.25 4.03 �10.6 8.5SS-3 Carbonate 1.00 3.85 �8.6 8.3SS-1 Gypsum 1.09 4.22 �8.2 8.2SS-2 Gypsum 1.31 4.73 �9.9 8.0CH-1 Gypsum 4.38 11.90 �3.2 7.5SS-4 Halite 3.06 9.24 �5.2 7.3CU-5 Halite 5.95 21.10 7.2 7.0
Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 357
Inc., Waltham, Massachusetts, USA), as were Cl�, Br�,and SO4
2� (Dionex ICS-2100, Thermo Fisher Scientific,Inc.). Elemental boron (B) and Ca2+ were measured usinginductively coupled plasma-optical emission spectrometryat JAMSTEC (ICP-OES, SII SPS5510, SII NanoTechnol-ogy Inc., Chiba, Japan). The analytical precisions (±2r)of replicate measurements were within ±2% for Na+ andCl�, and ±10% for the other elements.
2.3.3. Total alkalinity
Total alkalinity was measured using a total alkalinitytitrator at AIST (ATT-05, Kimoto Electric Co., Osaka,Japan). Samples were titrated with 0.1 mol L�1 HCl, andTA was calculated by the Gran method. Samples collectedfrom CH-1, SS-4, and CU-5 were diluted on a weight basiswith ultrapure water prior to the measurement. The analyt-ical precision of replicate measurements was within ±1.5%.
2.3.4. DIC concentration and carbon isotopic composition
For measurement of DIC, brine samples (10–50 mL)were transferred to a glass vial, and the air inside was
completely evacuated using a high-vacuum glass line. Next,the samples were reacted with H3PO4 and left for 12 h sothat DIC was completely converted to CO2 gas. Theevolved gas was then introduced into a high-vacuum glassline and separated cryogenically. The gas pressure in theglass line was recorded and converted to DIC concentra-tion. The precision of the DIC measurement was within0.3%. Afterwards, the purified CO2 gas was introduced intoan isotope-ratio mass spectrometer (Delta Plus XL,Thermo Fisher Scientific, Inc.) to measure the carbonisotopic composition of the DIC. Isotopic compositionsare expressed as conventional d13C relative to Vienna PeeDee Belemnite. The analytical precision was within 0.1‰.
2.4. Deposit sample analyses
2.4.1. Carbon isotopic composition of sediment TOC
Sediment samples collected from the bottom of theponds were subsampled for measurement of d13C of organicmatter (d13CTOC). Samples collected from the carbonateponds were separated into two parts: the upper slimy layer

358 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
and the loose black deposit underneath (Fig. 2b). The gyp-sum crusts were separated into four parts: the yellowishtransparent, green, and pink gypsum layers, and the looseblack deposits below (Fig. 2d). Subsampled deposits werefreeze-dried and ground to powder. Together with groundhalite samples, they were transferred to pre-cleanedsmooth-wall tin capsules and treated with 0.1 mol L�1
HCl to remove CaCO3. After the samples were dried theywere analyzed for d13CTOC at JAMSTEC using a modifiedFlash EA1112 automatic elemental analyzer connected to aThermo Finnigan Delta plus XP isotope ratio mass spec-trometer (IRMS) via a ConFlo III Interface (Ogawaet al., 2010). Isotopic compositions are expressed as conven-tional d13C values relative to Vienna Pee Dee Belemnite.The analytical precision was within 0.3‰.
2.4.2. Compound-specific pigment isotopic composition
The surface deposits from CU-1 and SS-3 (microbialmat), the yellowish transparent, green, and pink layers fromSS-1 and CH-1 (gypsum crust), and halite crystals from SS-4 and CU-5 were analyzed for compound-specific isotopecompositions of pigments. First, the deposits were freeze-dried and ground to powder. Organic matter was extractedwith acetone three times by sonication for 15 min in anultrasonic ice bath. The acetone fraction was then extractedwith n-hexane three times. The n-hexane fraction was driedcompletely under N2 gas and dissolved in 100 mL ofN,N-dimethylformamide for high-performance liquid chro-matography (HPLC) injection. All procedures were carriedout in a dark room.
Pigment isolation and purification was accomplishedusing dual step HPLC. The HPLC system comprised a bin-ary pump (G1312B; Agilent, Santa Clara, California,USA), an on-line degasser (G1379B; Agilent), an autosam-pler (G1367C; Agilent), a column temperature controller(Cool Pocket Column Chiller; Thermo Fisher Scientific),an on-line photodiode-array detector (G4212B; Agilent),and a fraction collector (G1364C; Agilent). The pigmentswere isolated using an Agilent Zorbax Eclipse XDB C-18column (4.6 mm � 250 mm; 5-lm silica particle size) witha guard column (4.6 mm � 12.5 mm; 5-lm silica particlesize). The pigments were eluted isocratically with 75% ace-tonitrile:pyridine (100:0.5, v/v) and 25% ethyl acetate:pyri-dine (100:0.5, v/v) for 5 min, followed by a linear gradientof ethyl acetate:pyridine to 50% over 50 min. The flow ratewas set to 1 mL min�1 and the column temperature to30 �C. Pigments were detected by the photodiode-arraydetector. The structure assignment of each compound wasaccomplished by comparing the photo-absorption spectraand the retention times with those of authentic standards.Chlorophyll a (Chl a), bacteriochlorophyll a (BChl a),and b-carotene were collected using the fraction collector.We carefully collected the entire peak for each compoundto avoid analytical isotopic fractionation.
The collected pigments were dried completely underargon gas. Prior to the second HPLC purification step,Chl a and BChl a were dissolved in 1.5 mL hexane andreacted with 2 mol L�1 HCl to converted them to pheo-phytin a (Pheo a) and bacteriopheophytin a (BPheo a),respectively. The hexane fraction was collected and dried
completely under argon gas, and dissolved in 100 mL ofN,N-dimethylformamide for HPLC injection. The columnused for second purification step was an Agilent ZorbaxEclipse PAH column (4.6 mm � 250 mm; 5-lm particlesize). Pigments were eluted isocratically with 80% acetoni-trile:pyridine (100:0.5, v/v) and 20% ethyl acetate:pyridine(100:0.5, v/v) for 5 min, followed by a linear gradient ofethyl acetate:pyridine to 60% over 25 min, and a linear gra-dient of ethyl acetate:pyridine to 100% over 10 min. Theflow rate was set to 1 mL min�1 and the column tempera-ture to 15 �C.
The stable carbon isotopic compositions of the pigmentswere measured using a modified EA/IRMS (Ogawa et al.,2010). Purified pigments were dissolved in dichloro-methane, transferred to pre-cleaned smooth-wall tincapsules, and dried before analysis. The analytical preci-sions were within 0.3‰ for Chl a and BChl a, and 0.6‰for b-carotene.
3. RESULTS
Data from the brine sample analyses are summarized inTables 1 and S1. The brines in the solar salterns originatefrom seawater from the same region, and precipitation ofevaporites is the major process occurring within the salt-erns. Thus, we report and discuss the data from differentsolar salterns together, under the assumption that the bio-logical processes in the three systems are comparable. Wedid not determine the salinity or the concentration ofNa+ and Cl� of the brine samples collected from the pondswith higher salinities (CH-1, SS-4, and CU-5) because halitecrystals precipitated in the sample bottles after the sampleswere collected.
3.1. Variations in concentrations of inorganic elements
The concentrations of solutes in the brines are deter-mined mainly by condensation due to evaporation, removalby precipitation of evaporite minerals, and the effects ofbiological activity. One way to determine the behavior ofthe solutes is to normalize their concentrations by thedegree of evaporation (DE; e.g., Babel and Schreiber,2014). The DE of the brine can be estimated from the con-centrations of the solutes that behave conservatively uponevaporation. In the salinity range of our samples, Mg2+,K+, Br�, and B behave as conservative solutes. Here, weestimated the DE of each brine sample from the Mg2+ con-centration as follows:
DEMg ¼ ½Mg2þbrine�½Mg2þseawater�
where ½Mg2þseawater� and ½Mg2þbrine� are the molar concentrationsof Mg ions in the seawater and brine samples, respectively.Normalization of the solute concentrations by DEMg can-cels out the effect of condensation due to evaporation,therefore allowing the examination of the addition orremoval of solutes to or from the brine.
The composition of the major ions in the sample of sea-water (CU-0), which is the source of the brines in the solarsalterns studied, was comparable to that reported for
Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 359
average modern seawater (Babel and Schreiber, 2014). Themajor evaporite minerals precipitated from seawater in thesalinity range of our samples are calcium carbonate, gyp-sum, and halite. Accordingly, ions such as Ca2+, HCO3
�,SO4
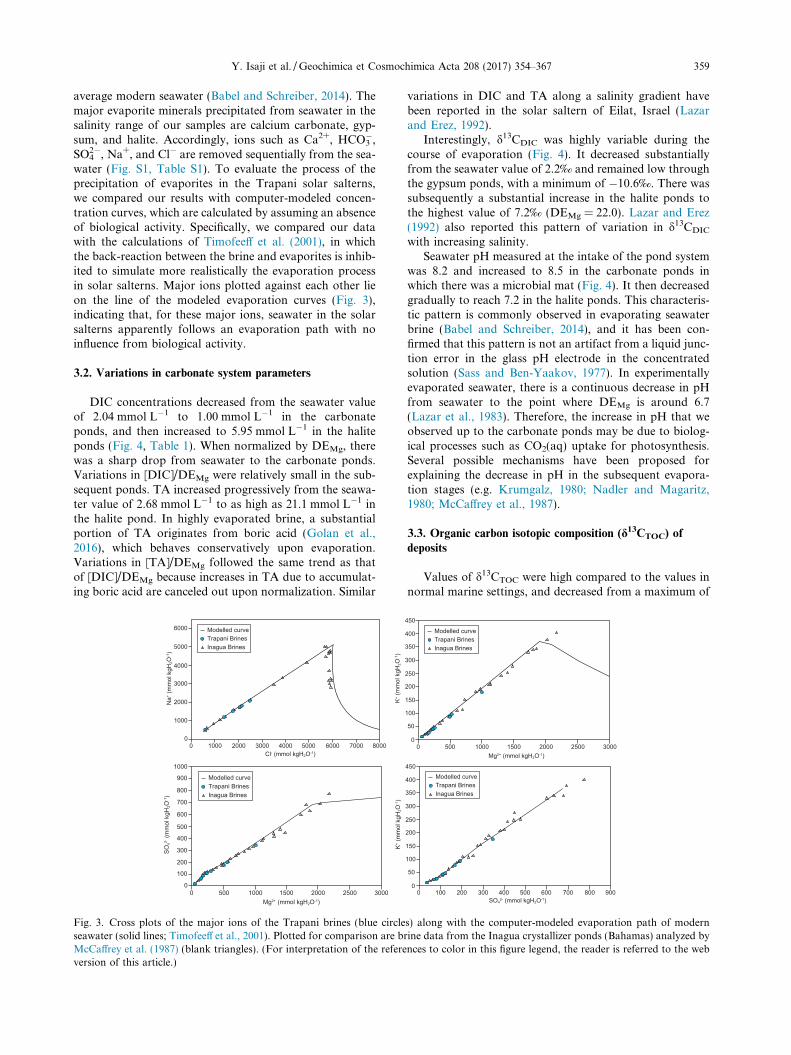
2�, Na+, and Cl� are removed sequentially from the sea-water (Fig. S1, Table S1). To evaluate the process of theprecipitation of evaporites in the Trapani solar salterns,we compared our results with computer-modeled concen-tration curves, which are calculated by assuming an absenceof biological activity. Specifically, we compared our datawith the calculations of Timofeeff et al. (2001), in whichthe back-reaction between the brine and evaporites is inhib-ited to simulate more realistically the evaporation processin solar salterns. Major ions plotted against each other lieon the line of the modeled evaporation curves (Fig. 3),indicating that, for these major ions, seawater in the solarsalterns apparently follows an evaporation path with noinfluence from biological activity.
3.2. Variations in carbonate system parameters
DIC concentrations decreased from the seawater valueof 2.04 mmol L�1 to 1.00 mmol L�1 in the carbonateponds, and then increased to 5.95 mmol L�1 in the haliteponds (Fig. 4, Table 1). When normalized by DEMg, therewas a sharp drop from seawater to the carbonate ponds.Variations in [DIC]/DEMg were relatively small in the sub-sequent ponds. TA increased progressively from the seawa-ter value of 2.68 mmol L�1 to as high as 21.1 mmol L�1 inthe halite pond. In highly evaporated brine, a substantialportion of TA originates from boric acid (Golan et al.,2016), which behaves conservatively upon evaporation.Variations in [TA]/DEMg followed the same trend as thatof [DIC]/DEMg because increases in TA due to accumulat-ing boric acid are canceled out upon normalization. Similar
Fig. 3. Cross plots of the major ions of the Trapani brines (blue circleseawater (solid lines; Timofeeff et al., 2001). Plotted for comparison are bMcCaffrey et al. (1987) (blank triangles). (For interpretation of the refereversion of this article.)
variations in DIC and TA along a salinity gradient havebeen reported in the solar saltern of Eilat, Israel (Lazarand Erez, 1992).
Interestingly, d13CDIC was highly variable during thecourse of evaporation (Fig. 4). It decreased substantiallyfrom the seawater value of 2.2‰ and remained low throughthe gypsum ponds, with a minimum of �10.6‰. There wassubsequently a substantial increase in the halite ponds tothe highest value of 7.2‰ (DEMg = 22.0). Lazar and Erez(1992) also reported this pattern of variation in d13CDIC
with increasing salinity.Seawater pH measured at the intake of the pond system
was 8.2 and increased to 8.5 in the carbonate ponds inwhich there was a microbial mat (Fig. 4). It then decreasedgradually to reach 7.2 in the halite ponds. This characteris-tic pattern is commonly observed in evaporating seawaterbrine (Babel and Schreiber, 2014), and it has been con-firmed that this pattern is not an artifact from a liquid junc-tion error in the glass pH electrode in the concentratedsolution (Sass and Ben-Yaakov, 1977). In experimentallyevaporated seawater, there is a continuous decrease in pHfrom seawater to the point where DEMg is around 6.7(Lazar et al., 1983). Therefore, the increase in pH that weobserved up to the carbonate ponds may be due to biolog-ical processes such as CO2(aq) uptake for photosynthesis.Several possible mechanisms have been proposed forexplaining the decrease in pH in the subsequent evapora-tion stages (e.g. Krumgalz, 1980; Nadler and Magaritz,1980; McCaffrey et al., 1987).
3.3. Organic carbon isotopic composition (d13CTOC) of
deposits
Values of d13CTOC were high compared to the values innormal marine settings, and decreased from a maximum of
s) along with the computer-modeled evaporation path of modernrine data from the Inagua crystallizer ponds (Bahamas) analyzed bynces to color in this figure legend, the reader is referred to the web

Fig. 4. Variations in dissolved inorganic carbon (DIC) concentra-tions, total alkalinity (TA), d13CDIC, and pH (blue circles), andDIC and TA normalized to the degree of evaporation calculatedfrom magnesium concentrations (DEMg; red circles). The bars atthe top of the figure show the precipitation ranges for calciumcarbonate, gypsum, and halite. (For interpretation of the referencesto color in this figure legend, the reader is referred to the webversion of this article.)
Fig. 5. Variations in d13C of dissolved inorganic carbon (DIC, bluestars), total organic carbon (TOC, gray circles), chlorophyll a
(green circles), and b-carotene (red circles), as well as thefractionation factor e calculated from d13C of TOC and pigments.(For interpretation of the references to color in this figure legend,the reader is referred to the web version of this article.)
360 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
�8.6‰ in the carbonate ponds to a minimum of �22.7‰ inthe halite ponds (Fig. 5, Table S2). The differences betweend13CTOC and d13CDIC increased as the salinity increased.Depth profiles of d13CTOC do not show any common trendsamong ponds (Fig. 6, Table S2). The lack of a commontrend may be because the isotopic signals of organisms liv-ing in the lower layers of the deposits are superimposed onthe signals of upper-layer organisms, reflecting the fact thatthe lower-layer deposits consist of upper-layer deposits thatsubsequently became buried and then occupied by lower-layer dwellers. This preservation of the original mat-surface d13C signature in deeper layers has been reportedpreviously (e.g., Des Marais et al., 1992).
3.4. Distribution of pigments and their carbon isotopic
composition
The distribution of pigments was similar for the sametypes of samples from different solar salterns. Therefore,only representative chromatograms are shown: from thesurface slimy layer of the microbial mats in the carbonate
ponds, from the yellowish transparent, green, and pink lay-ers of the gypsum crusts in the gypsum ponds, and from thehalite crystals in the halite ponds (Fig. S2).
The major pigments detected in the surface slimy layerof the carbonate ponds were the Chl a series, the BChl aseries, and various carotenoids. The Chl a series includesChl a and its degradation products, Pheo a and pyropheo-phytin a (PPheo a). The BChl a series includes BChl a andits degradation products, BPheo a and bacteriopyropheo-phytin a (BPPheo a). Among the various carotenoidsdetected, the peaks with a retention time around 23 minwere identified as b-carotene and its degradation products(b-carotene series), based on a comparison with the authen-tic standard. The relative concentrations of the original pig-ments (i.e., Chl a and BChl a) were substantially higherthan their counterpart degradation products. We measuredthe d13C of Chl a, BChl a, and b-carotene.
In the gypsum crust, the main pigments in the yellowishtransparent layer and the green layer were the Chl a seriesand smaller peaks of carotenoids, including theb-carotene series. The pink layer contained Chl a, BChl a,and the b-carotene series, with BChl a highest in concentra-tion. The Chl a in the pink layer was probably originallyfrom the cyanobacteria or algae in the upper yellowishand green layers, because they migrate upward as the photicand oxic zones moves upward with the growth of the gyp-sum crust. We therefore measured d13C of Chl a in the yel-lowish transparent layer and the green layer, which isdominated by cyanobacteria and algae, and BChl a in thepink layer dominated by purple sulfur bacteria. We alsomeasured d13C of b-carotene in the yellowish transparentlayer of the gypsum crust.
The pigment distribution in the halite crystals from thehalite ponds was completely different from that in the car-bonate and gypsum ponds, with the b-carotene series inhighest concentrations and extremely low Chl a concentra-tions. Because there was not enough Chl a for isotopic

Fig. 6. Depth profiles of d13C of chlorophyll a (Chl a) originating from cyanobacteria and/or algae (green circles), bacteriochlorophyll a(BChl a) from purple sulfur bacteria (red circles), and total organic carbon (TOC, gray squares) in the microbial mats of the carbonate ponds(CU-1 and SS-3) and the gypsum crusts of the gypsum ponds (SS-1 and CH-1). Blue circles indicate the d13C of dissolved inorganic carbon(DIC) in the surface brine. CU, Culcasi; SS, Sosalt; CH, Chiusicella. (For interpretation of the references to color in this figure legend, thereader is referred to the web version of this article.)
Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 361
measurement, we determined d13C of only the b-caroteneseries in halite samples.
The depth variations of pigment d13C values showedsimilar patterns in all ponds: d13C of BChl a was lower thanthat of Chl a in both microbial mats and gypsum crusts(Fig. 6, Table 2). In a comparison between ponds, thed13C values of Chl a, BChl a, and b-carotene were highestin CU-1, at �11.5‰, �19.5‰ and �22.1‰, respectively(Fig. 5). The d13C values of all pigments showed decreasingtrends as evaporation proceeded, and reached minimumvalues of �20.6‰ and �26.3‰ in CH-1 for Chl a and BChla, respectively, and �28.5‰ in CU-5 for b-carotene. Thed13CTOC of the surface sediment samples showed similartrends.
We calculated an isotopic fractionation factor from thed13CDIC in the surface brine and that of TOC, Chl a, andb-carotene: e � (Rorg/RDIC) � 1000 (‰), where Rorg is13C/12C for TOC, Chl a, or b-carotene. We specifically usedthe d13C of Chl a and b-carotene originating from the sur-face slimy layer of the microbial mats, the yellowish trans-parent layer from the gypsum crusts, and the halite crystals,which contain pigments derived from microorganismsassumed to assimilate DIC mainly from the surface brine.Overall, the values of e were lower in the lower salinityponds, and increased more or less linearly as evaporationproceeds (Fig. 5). Between TOC, Chl a, and b-carotene, eof b-carotene was highest, ranging from 14.9‰ to 36.0‰,followed by that of Chl a, ranging between 6.4‰ and17.3‰, and the lowest, that of TOC, ranging from 3.5‰to 30.2‰.
We did not quantify the concentration of each pigmentbecause of the somewhat patchy distribution of the coloredlayers on the pond bottoms. However, our rough estimatesindicate that the concentrations of Chl a and BChl a were
on the order of micrograms per gram of dry sediment forthe microbial mat and the gypsum crust.
4. DISCUSSION
4.1. Changes in primary production with increasing salinity
For estimating the primary productivity, the d13C of pig-ments has an advantage over d13CTOC because the pigmentsderive exclusively from photoautotrophs; thus, other fac-tors such as heterotrophic activity, which potentially affectsd13CTOC, are excluded. Compound-specific isotope analysisof chlorophylls has been successfully used to improveunderstanding of modern and past biogeochemical cycles,especially in the euphotic zone (e.g., Sachs and Repeta,1999; Ohkouchi et al., 2005; Kusch et al., 2010; Tyleret al., 2010; Isaji et al., 2015).
Among the pigments measured in this study, Chl a andb-carotene are synthesized by aerobic photoautotrophssuch as cyanobacteria and algae, and BChl a is producedby the purple sulfur bacteria present in the pink layer ofthe microbial mat. In solar salterns, both planktonic andbenthic cyanobacteria and algae are generally present. InSpain, the reported concentrations of Chl a in the brineof a solar saltern were around 2–15 mg L�1 (Joint et al.,2002). On the other hand, the concentrations of Chl a inthe benthic deposits of the Trapani solar salterns were onthe order of micrograms per gram of dry sediment, whichis much higher than the concentration of the overlyingbrine reported in Joint et al. (2002). Although we did notmeasure the concentration of Chl a in the brine in thisstudy, we speculate that the dominant primary produceris the benthic community in ponds where benthic microbialmat is formed. Chl a and b-carotene in the halite ponds

Table 2d13C of chlorophyll a (Chl a), bacteriochlorophyll a (BChl a), and b-carotene extracted from deposits collected from the Culcasi (CU), Sosalt(SS), and Chiusicella (CH) solar salterns in Trapani, Sicily, Italy. Numbers in the sample names refer to individual ponds at the salterns.Analytical errors are based on replicate measurements of standard material.
Sample Type of evaporite Layer Compound d13C (‰) Error (2r)
CU-1-1 Carbonate Top slimy Chl a �11.5 0.23BChl a �19.7 0.23b-carotene �22.1 0.39
SS-3-1 Chl a �20.1 0.15BChl a �23.4 0.15b-carotene �23.7 0.39
SS-1-1 Gypsum Yellowish Chl a �17.9 0.23b-carotene �25.9 0.37
SS-1-2 Green Chl a �17.0 0.23SS-1-3 Pink BChl a �20.6 0.23CH-1-1 Yellowish Chl a �20.6 0.15
b-carotene �23.6 0.57CH-1-2 Green Chl a �18.6 0.23CH-1-3 Pink BChl a �26.3 0.23SS-4 Halite Bulk crystal b-carotene �25.8 0.15CU-5 b-carotene �28.5 0.37
362 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
probably originate from planktonic photoautotrophs,because these ponds contain no benthic microbial mat.Specifically, Dunaliella salina is the likely candidate,because this organism is known to be a dominant primaryproducer in halite ponds (e.g., Rehakova et al., 2009),and it also accumulates high amounts of b-carotene(Oren, 2005).
There are several possible factors that could account forthe lower d13C of BChl a compared to that of Chl a in thebenthic community (Fig. 6). One is the difference in thesource of DIC utilized by each photoautotroph. Becausethe purple sulfur bacteria inhabit the deeper layer of themat, some proportion of DIC assimilated by them is sup-plied through mineralization of the organic matter withinthe mat. The values of d13CTOC indicate that d13C of themineralized DIC is lower than that of DIC in the surfacebrine, because isotopic fractionation associated with degra-dation of organic matter is negligible (e.g., Meyers andEadie, 1993). Another factor is the chemical species ofDIC assimilated by the photoautotroph. Cyanobacteriaare capable of assimilating HCO3
� through active transport(e.g., Kaplan et al., 1980; Badger and Price, 2003). BecauseHCO3
� is enriched in 13C compared to CO2(aq) by 8.4‰under the temperature of 30 �C (Mook et al., 1974), activeassimilation of HCO3
� by cyanobacteria may have resultedin the relatively high d13C of Chl a. As for the purple sulfurbacteria, d13C depletion in BChl a compared to Chl a hasbeen reported from the saline meromictic Lake Kaiike inJapan (Ohkouchi et al., 2005). Because purple sulfur bacte-ria, cyanobacteria, and algae use identical biochemicalpathways for carbon assimilation and chlorophyll biosyn-thesis, differences in their d13C can be ascribed to physiolog-ical factors such as growth rate, cell size, or geometry(Pancost et al., 1997; Popp et al., 1998; Bidigare et al.,1999; Ohkouchi et al., 2008).
The isotopic fractionation factor (e) calculated from Chla and b-carotene increased almost linearly with increasingsalinity (Fig. 5). Because the source photoautotrophs ofthese pigments do not change substantially along the
salinity gradient (i.e., cyanobacteria and/or algae), thereare several possible explanations for this observation. Thehabitat of the photoautotrophs is one of the major factorscontrolling e along the salinity gradient, because whileplanktonic photoautotrophs such as D. salina inhabitingthe halite ponds can utilize DIC in the surface brine, ben-thic cyanobacteria and algae in the microbial mats and gyp-sum crusts utilize DIC that diffuses from the overlyingbrine. DIC diffusion into the benthic microbial mat canbe limited by a diffusive boundary layer over the mat sur-face (Jørgensen, 1994b) or within the mat (Wieland et al.,2001). As for the gypsum crust, there is likely only limitedexchange of brine through the pore water. These limits ondiffusion could result in DIC-limited conditions inside themicrobial mat, which is expressed as a relatively small ewithout depletion of DIC in the overlying brine. Indeed,limited DIC diffusion into the microbial mat has also beenconsidered as a possible reason for relatively high d13CTOC
and therefore low values of e (e.g., Des Marais andCanfield, 1994; Schouten et al., 2001). From this perspec-tive, the relatively large e in the halite ponds is becausethe habitat of the dominant photoautotroph, D. salina, isin the surface brine, where conditions are not DIC-limited.
Another possible factor controlling e along the salinitygradient comes from the observation that e increases whencompared among ponds of the same type: the carbonateponds (CU-1, SS-3), the gypsum ponds (SS-1, CH-1), andthe halite ponds (SS-4, CU-5). The only exception is thedecrease in e calculated from d13C of b-carotene in the car-bonate ponds. If the diffusion rate of DIC into the mat orthe crust does not change substantially with increasingsalinity, then the increase in e can be interpreted as reflect-ing a lower proportion of DIC assimilated by photoau-totrophs under the higher salinities. In other words,increasing salinity may have suppressed primary produc-tion. This suggestion is consistent with the findings of pre-vious studies that photosynthetic activity decreases withincreasing salinity (Oren, 2009) in microbial mats (e.g.,Pinckney et al., 1995; Wieland and Kuhl, 2006) and gypsum

Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 363
crusts (e.g., Caumette et al., 1994; Canfield et al., 2004), aswell as in the planktonic community of halite ponds (Jointet al., 2002). Salinity may directly control the primary pro-ductivity by affecting the physiology of photoautotrophs, orindirectly by affecting the elemental cycles of nutrients suchas nitrogen, phosphorus, and iron.
4.2. Effect of biological activities on the chemical evolution of
evaporating seawater
4.2.1. Influence of sulfate reduction on brine
One of the highest rates of sulfate reduction knownoccurs in hypersaline microbial mats (Canfield and DesMarais, 1991). Sulfate reduction removes SO4
2� from brineby reduction to H2S or HS� and subsequent precipitationas various metal sulfides (e.g., Wieland et al., 2005;Valdivieso-Ojeda et al., 2014). Although this process musthave removed some portion of SO4
2� from the brine in thisstudy, concentrations of SO4
2� plotted against other majorions lie on the line of the ideal curve (Fig. 3). To evaluatethe influence of the sulfate reduction in the brine, weroughly estimated the amount of sulfate removed on thebasis of sulfate reduction rates in hypersaline microbialmats and gypsum crusts of the solar salterns of GuerreroNegro, Mexico (Canfield and Des Marais, 1991, 1993),Eilat, Israel (Frund and Cohen, 1992; Jørgensen, 1994a;Canfield et al., 2004; Sørensen et al., 2004), and Salins-de-Giraud, France (Caumette et al., 1994). Assuming an aver-age pond water depth of 50 cm, we calculated the amountof SO4
2� reduced by 20 cm2 of the mat, which correspondsto 1 L of overlying brine. The calculated value varied sub-stantially for both the microbial mats and the gypsumcrusts; both varied between tens of micromoles to mil-limoles per liter per day. In contrast, the SO4
2� in the over-lying brine in the Trapani solar salterns ranged from 79.2 to233.2 mmol L�1 from the carbonate to the gypsum ponds(Table S1).
Although sulfate reduction rates were not measured inthe Trapani solar salterns, this rough estimation impliesthat sulfate reduction can influence the SO4
2� concentrationof the brine if the reduction is on the order of millimoles perliter per day. The fact that the SO4
2� concentration tracksthe ideal evaporation curve therefore indicates that SO4
2�
is supplied by other processes to compensate any lossthrough reduction, or that the sulfate reduction rate inthe Trapani solar salterns is low. One possible source ofSO4
2� for the former explanation is oxidation of the reducedsulfur species by chemotrophic and phototrophic sulfurbacteria or by oxygen produced by photosynthesis in theupper layer of deposits and surface brine (e.g., Revsbechet al., 1983; Frund and Cohen, 1992; Canfield and DesMarais, 1993). Note that decreases in SO4
2� and increasesin total alkalinity (TA) through sulfate reduction are can-celed out by sulfide oxidation, but increases in DIC are not.
4.2.2. Effect of biological processes on the brine carbonate
system
In the shallow hypersaline environment where benthicmicrobial mats form, carbonate system parameters, i.e.,DIC, TA, pH, and pCO2, are affected by various processes
such as calcium carbonate precipitation and dissolution(Eq. (1) below), photosynthesis and respiration (Eq. (2)),sulfate reduction (Eq. (3)), sulfide oxidation (Eq. (4)), andCO2(g) exchange with the atmosphere. The following equa-tions describe these processes.
Ca2þ þ 2HCO�3 ¢CaCO3ðsÞ þ CO2ðgÞ þH2O ð1Þ
106CO2 þ 16HNO3 þH3PO4 þ 122H2O
¢ ðCH2OÞ106ðNH3Þ16ðH3PO4Þ þ 138O2 ð2ÞSO2�
4 þ 2CH2O ! HS� þ 2HCO�3 þHþ ð3Þ
HS� þ 2O2 ! SO2�4 þHþ ð4Þ
Our values for [DIC]/DEMg and [TA]/DEMg indicatethat DIC and TA are removed from the brine during thecourse of evaporation from the seawater to the carbonateponds, accompanied by a substantial drop in d13CDIC
(Fig. 4). In this salinity range, calcium carbonateprecipitation is one of the major process affecting DIC con-centrations, decreasing the d13CDIC of the brine by prefer-entially removing 13C from DIC reservoirs. The typicalenrichment factors for carbon isotope fractionationbetween calcium carbonate and DIC are +1.0‰ and+2.7‰ for calcite and aragonite, respectively (Romaneket al., 1992). Theoretically, the decrease in d13CDIC inCU-1 from the seawater value of 2.2‰ to �5.1‰ couldbe reached if almost all DIC precipitated as calcium car-bonate. However, calcium carbonate precipitation fromseawater to DEMg = 6.7 results in a TA loss of around60% of the source seawater, in the absence of biologicalactivity (Lazar et al., 1983). Therefore, calcium carbonateprecipitation alone cannot explain the drop in d13CDIC from2.2‰ to �5.1‰ (from seawater to CU-1) or from �5.1‰ to�10.6‰ (from CU-1 to CU-2).
Whereas DIC is affected by processes such as calciumcarbonate precipitation and dissolution, photosynthesisand respiration, sulfate reduction, and CO2(g) exchangewith the atmosphere, TA is primarily affected by precipita-tion and dissolution of calcium carbonate, sulfate reduc-tion, and sulfide oxidation. We suggested in Section 4.2.1that the loss of SO4
2� through sulfate reduction was com-pensated for by sulfide oxidation, or that the rate of sulfatereduction was low. These observations implies that the netchange in TA must have also been near zero. Cyanobacte-ria, the main photoautotroph in the hypersaline microbialmat, are capable of assimilating HCO3
� during photosyn-thesis (e.g., Kaplan et al., 1980; Badger and Price, 2003).This process does not affect TA, however, because theyrelease OH� when utilizing HCO3
� as the carbon source(e.g., Prins and Elzenga, 1989).
On the basis of these observations, we calculated theamount of calcium carbonate precipitated from the changesin TA, taking the transition from CU-0 (seawater) to CU-1(DE = 2.7) as the model case. It is calculated that1.40 mmol L�1 of TA is lost from the seawater to CU-1.According to Eq. (1), the precipitation of one mole of cal-cium carbonate utilizes two moles of HCO3
� and releasesone mole of CO2(g). The buffering effect of seawaterreduces the actual amount of CO2(g) liberated to the atmo-sphere to around 0.6 mol per mole of calcium carbonate

364 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
precipitated (Ware et al., 1992; Frankignoulle et al., 1994).Under the assumption that this rule is applicable to thehypersaline solutions in saltern ponds, 1.40 mmol L�1 ofTA loss is equivalent to the precipitation of 0.70 mmol L�1
calcium carbonate and production of 0.70 mmol L�1 ofCO2(g), of which 0.28 mmol L�1 is re-dissolved into thesolution, during the transition from seawater to the brinein CU-1. This results in a decrease of 1.12 mmol L�1 ofDIC, because calcium carbonate precipitation decreasesDIC and TA equally but the re-dissolution of CO2(g)increases only DIC. However, the actual decrease in DICfrom CU-0 to CU-1 was 1.57 mmol L�1, according to thechanges from seawater to CU-1. This value indicates that0.45 mmol L�1 of DIC was lost by processes other than cal-cium carbonate precipitation (Fig. 7).
There are several possible processes responsible for theloss of DIC other than calcium carbonate precipitation.One is carbon fixation by photoautotrophs, which preferen-tially removes 13C-depleted DIC from the brine. The iso-topic fractionation factor (e) calculated from the d13C ofpigments indicates that photosynthesis was more active inthe lower salinity ponds (Fig. 5). The degassing of CO2(aq)due to a decrease in solubility induced by evaporation alsoremoves 13C-depleted DIC from the brine (Li and Tsui,1971; Stiller et al., 1985; Raven, 1991; Barkan et al.,2001). This process was active throughout the evaporationpath. Thus, although these processes that remove 13C-depleted DIC from the brine could balance the DIC budget,there must be other processes supplying 13C-depleted DICto explain the 13C-depleted d13CDIC of the carbonate ponds.We propose several processes that may account for thesupply of 13C-depleted DIC:
Fig. 7. (a) Cross plot of dissolved inorganic carbon (DIC) concentrationbased on magnesium ion concentrations ([DIC]/DEMg and [TA]/DEMg, rutilization of HCO3
� and re-dissolution of CO2(g) by calcium carbonatcarbonate pond (CU-1, DEMg = 2.7), respectively. Dotted red arrow C[TA]/DEMg in CU-1. Solid arrows in the bottom right corner indicate the dprecipitation, (2) photosynthesis and respiration, (3) sulfate reduction,degassing of CO2(aq). (b) Close-up view of (a). Numbers next to the sreferences to color in this figure legend, the reader is referred to the web
(1) Because intensive photosynthesis and degassing dueto evaporation remove CO2(aq) from the brine, someDIC must have been supplied by equilibrium withatmospheric CO2, which is relatively depleted in 13C(e.g. Keeling, 1958). Moreover, Baertschi (1952) sug-gested that 13C-depleted CO2 might be selectively dis-solved into brine from the atmosphere under alkalineconditions (‘‘the Baertschi effect”). As also suggestedby Lazar and Erez (1992), this process may beresponsible for the 13C-depleted DIC in the brine ofthe carbonate ponds.
(2) A previous study excluded sulfate reduction releasingDIC in the form of HCO3
� as the main cause of rela-tively low d13CDIC of the brine because there was nosubstantial increase in [TA]/DEMg with increasingDEMg (Fig. 4; Lazar and Erez, 1992). In the Trapanisolar salterns, the changes in SO4
2� concentrationsuggest that the loss of SO4
2� through sulfate reduc-tion was compensated for by sulfide oxidation, or elsethe rate of sulfate reduction was low (Fig. 3). As sug-gested in Section 4.2.1, sulfate reduction coupled withsulfide oxidation cancels out increases in TA, but notDIC. Therefore, we cannot exclude the possibilitythat the 13C-depleted DIC was supplied by sulfatereduction. It may originate from the benthic micro-bial mat within the pond, or from the highly produc-tive microbial communities in less evaporated,upstream ponds (Joint et al., 2002). Note that theactual amount of DIC that diffuses into the overlyingbrine is less than the amount mineralized from organ-ic matter, because of restricted diffusion of DICwithin the mat (Wieland et al., 2001) and on the
s and total alkalinity (TA) normalized to the degree of evaporationespectively). Dotted black arrows A and B indicate changes due toe precipitation during the transition from seawater (CU-0) to theindicates the supply of DIC necessary to explain [DIC]/DEMg andirection and slope of the following processes: (1) calcium carbonate(4) sulfide oxidation, (5) dissolution of atmospheric CO2, and (6)ymbols are DEMg values. CU, Culcasi. (For interpretation of theversion of this article.)

Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 365
mat surface (Jørgensen, 1994b). Indeed, Canfield andDes Marais (1993) suggested that a large proportionof mineralized DIC is fixed back into organic matterwithin the mat during the day, whereas it diffuses intothe overlying brine at night.
In the gypsum and halite ponds, d13CDIC graduallyincreased to reach 7.2‰ in CU-5. This increase indicatesthat the biological processes that reduced the d13CDIC ofthe carbonate and gypsum ponds are suppressed in thehalite ponds, and that d13CDIC is primarily controlled bythe degassing of CO2(aq) (Stiller et al., 1985). A reducedinfluence of biological activity on d13CDIC is consistent withthe discussion about the d13C of pigments indicating thatphotosynthesis is more active in the lower salinity ponds.
Another observation is that DIC concentrations and TAin CH-1 and CU-5 were higher than those found in thesolar saltern of Eilat, Israel (Lazar and Erez, 1992). Thesehigher concentrations might be because the biological pro-cesses that potentially accumulate DIC and TA were moreintense in the upper-stream, lower salinity ponds of the Tra-pani solar salterns. Indeed, as in CU-5, DIC and TA in theupstream ponds (CU-1 and CU-2) were also slightly higherthan the values reported by Lazar and Erez (1992), suggest-ing that biological processes in the lower salinity pondsaccumulated in and modified the chemical composition ofhigher salinity ponds. Alternatively, the higher DIC andTA in the Trapani ponds may be due to dissolution ofcalcium carbonate supplied by aerial transport, becauseexperimentally evaporated seawater is known to beundersaturated with respect to aragonite (Lazar et al.,1983).
5. CONCLUSIONS AND IMPLICATIONS
In this study, we demonstrated that the isotopic frac-tionation factor (e) calculated from d13C of chlorophyll aand b-carotene, which originate from cyanobacteria andalgae, increased linearly along an increasing salinity gradi-ent in solar saltern ponds in Trapani, Italy. We ascribe thisobservation to the DIC-limited conditions within themicrobial mats and gypsum crusts caused by restrictedDIC diffusion from the overlying brine, and/or suppressionof primary production with increasing salinity. Variationsin the carbonate system parameters also indicate changingmicrobial activity along the salinity gradient. We proposethat dissolution of atmospheric CO2 into the brine throughintensive CO2(aq) uptake by photosynthesis and mineral-ization of organic matter by sulfate reduction may be theprocesses responsible for 13C-depleted DIC in the carbon-ate and gypsum ponds. In contrast, we attribute increasesin d13CDIC in subsequent ponds to the dominance of degas-sing of CO2(aq) with reduced microbial activity.
One important reason for elucidating the carbon cycle ofhypersaline environments is that such environments mayhave had a substantial impact on the global carbon cycleduring massive evaporation events that repeatedly occurredworldwide in the geological past (e.g., Hay et al.,2006; Warren, 2010). This study and previous studies
(e.g., Stiller et al., 1985; Lazar and Erez, 1992) havedemonstrated that CO2 exchange between brine and theatmosphere is an important factor controlling the brine car-bonate system during the evaporation of seawater. There isstill some debate about the environmental setting (e.g., shal-low vs. deep) and the extent of evaporation (e.g., total des-iccation vs. non-desiccation) of the massive evaporationevents in the past (e.g., the Messinian Salinity Crisis; Hsuet al., 1973; Roveri et al., 2014). However, furtherconstraints on the evaporation model during these events,as well as the elucidation of the behavior of the carbonatesystem under various evaporative settings, will enhanceour understanding of the role of hypersaline environmentsin the global carbon cycle.
ACKNOWLEDGEMENTS
We are grateful to SoSalt Spa and R.N.O Saline di Trapani ePaceco for permission to sample Trapani solar works. We grate-fully acknowledge Dr. K. Nagashima of the Japan Agency forMarine-Earth Science and Technology (JAMSTEC) for assistancein X-ray diffraction analysis. We thank Dr. H. Suga of JAMSTECfor assistance with the laboratory work, and Dr. K. Ozaki of Geor-gia Institute of Technology for help with the sampling in the solarsalterns. We also thank Dr. R. H. Byrne, an associate editor, Dr. B.Lazar, and two anonymous reviewers for their insightful com-ments. This study was partly supported by a Japan Society forthe Promotion of Science (JSPS) Research Fellowship (16J07844)to Y. Isaji, Grants-in-Aid to H. Kawahata (Nos. 19340146,22224009, and 15H02139), and JAMSTEC. All data used in thisarticle are available from the corresponding author.
APPENDIX A. SUPPLEMENTARY MATERIAL
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.gca.2017.04.013.
REFERENCES
Anton J., Rossello-Mora R., Rodrıguez-Valera F. and Amann R.(2000) Extremely halophilic bacteria in crystallizer ponds fromsolar saltern. Appl. Environ. Microbiol. 66, 3052–3057.
Babel M. and Schreiber B. C. (2014) Geochemistry of evaporitesand evolution of seawater. In Treatise on Geochemistry (eds. H.Holland and K. Turekian), second ed. Elsevier, Oxford, pp.483–560.
Badger M. R. and Price G. D. (2003) CO2 concentratingmechanisms in cyanobacteria: molecular components, theirdiversity and evolution. J. Exp. Bot. 54, 609–622.
Baertschi P. (1952) Die Fraktionierung der Kohlenstoffisotopen beider absorption von Kohlendioxyd. Helv. Chim. Acta 35, 1030–1036.
Barkan E., Luz B. and Lazar B. (2001) Dynamics of the carbondioxide system in the Dead Sea. Geochim. Cosmochim. Acta 65,355–368.
Baumgartner L. K., Reid R. P., Dupraz C., Decho A. W., BuckleyD. H., Spear J. R., Przekop K. M. and Visscher P. T. (2006)Sulfate reducing bacteria in microbial mats: changing para-digms, new discoveries. Sediment. Geol. 185, 131–145.

366 Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367
Bidigare R. R., Hanson K. L., Buessler K. O., Wakeham S. G.,Freeman K. H., Pancost R. D., Millero F. J., Steinberg P.,Popp B. N., Latasa M., Landry M. R. and Laws E. A. (1999)Iron-stimulated changes in 13C fractionation and export byequatorial Pacific phytoplankton: toward a paleogrowth rateproxy. Paleoceanography 14, 589–595.
Canfield D. E. and Des Marais D. J. (1991) Aerobic sulfatereduction in microbial mats. Science 251, 1471–1473.
Canfield D. E. and Des Marais D. J. (1993) Biogeochemical cyclesof carbon, sulfur, and free oxygen in a microbial mat. Geochim.
Cosmochim. Acta 57, 3971–3984.Canfield D. E., Sørensen K. B. and Oren A. (2004) Biogeochem-
istry of a gypsum-encrusted microbial ecosystem. Geobiology 2,133–150.
Caumette P., Matheron R., Raymond N. and Relexans J. C. (1994)Microbial mats in the hypersaline ponds of Mediterraneansalterns (Salins-de-Giraud, France). FEMS Microbiol. Ecol. 13,273–286.
Des Marais D. J. (2003) Biogeochemistry of hypersaline microbialmats illustrates the dynamics of modern microbial ecosystemsand the early evolution of the biosphere. Biol. Bull. 204, 160–167.
Des Marais D. J. and Canfield D. E. (1994) The carbon isotopebiogeochemistry of microbial mats. In Microbial Mats: Struc-
ture, Development and Environmental Significance, vol. 35 (eds.L. J. Stal and P. Caumette). Springer, Berlin, pp. 289–298,NATO ASI Series G.
Des Marais D. J., Bauld J., Palmisano A. C., Summons R. E. andWard D. M. (1992) The biogeochemistry of carbon in modernmicrobial mats. In The Proterozoic Biosphere: A Multidisci-
plinary Study (eds. J. W. Schopf and C. Klein). CambridgeUniversity Press, Cambridge, pp. 299–308.
Frankignoulle M., Canon C. and Gattuso J. P. (1994) Marinecalcification as a source of carbon dioxide: positive feedback ofincreasing atmospheric CO2. Limnol. Oceanogr. 39, 458–462.
Frund C. and Cohen Y. (1992) Diurnal cycles of sulfate reductionunder oxic conditions in cyanobacterial mats. Appl. Environ.Microbiol. 58, 70–77.
Geisler-Cussey D. (1997) Modern depositional facies developed inevaporative environments (marine, mixed, and nonmarine). InSedimentary Deposition in Rift and Foreland Basins in France
and Spain (Paleogene and Lower Neogene) (eds. G. Busson andB. C. Schreiber). Columbia University Press, New York, pp. 3–42.
Gieskes J. M. (1969) Effect of temperature on the pH of seawater.Limnol. Oceanogr. 14, 679–685.
Golan R., Gavrieli I., Ganor J. and Lazar B. (2016) Controls onthe pH of hyper-saline lakes-A lesson from the Dead Sea. EarthPlanet. Sci. Lett. 434, 289–297.
Green S. J., Blackford C., Bucki P., Jahnke L. L. and Prufert-Bebout L. (2008) A salinity and sulfate manipulation ofhypersaline microbial mats reveals stasis in the cyanobacterialcommunity structure. ISME J. 2, 457–470.
Hay W. W., Migdisov A., Balukhovsky A. N., Wold C. N., FlogelS. and Soding E. (2006) Evaporites and the salinity of the oceanduring the Phanerozoic: implications for climate, ocean circu-lation and life. Palaeogeogr. Palaeoclimatol. Palaeoecol. 240, 3–46.
Hsu K. J., Ryan W. B. F. and Cita M. B. (1973) Late Miocenedesiccation of the Mediterranean. Nature 242, 240–244.
Imhoff J. F. (2001) True marine and halophilic anoxygenicphototrophic bacteria. Arch. Microbiol. 176, 243–254.
Isaji Y., Kawahata H., Ohkouchi N., Ogawa N. O., MurayamaM.,Inoue K. and Tamaki K. (2015) Varying responses to Indianmonsoons during the past 220 kyr recorded in deep-seasediments in inner and outer regions of the Gulf of Aden. J.Geophys. Res. Oceans. 120, 7253–7270.
Joint I., Henriksen P., Garde K. and Riemann B. (2002) Primaryproduction, nutrient assimilation and microzooplankton graz-ing along a hypersaline gradient. FEMS Microbiol. Ecol. 39,245–257.
Jørgensen B. B. (1994a) Sulfate reduction and thiosulfate trans-formations in a cyanobacterial mat during a diel oxygen cycle.FEMS Microbiol. Ecol. 13, 303–312.
Jørgensen B. B. (1994b) Diffusion processes and boundary layers inmicrobial mats. In Microbial Mats: Structure, Development and
Environmental Significance, vol. 35 (eds. L. J. Stal and P.Caumette). Springer, Berlin, pp. 243–253, NATO ASI Series G.
Kaplan A., Badger M. R. and Berry J. A. (1980) Photosynthesisand the intracellular inorganic carbon pool in the bluegreenalga Anabaena variabilis: response to external CO2 concentra-tion. Planta 149, 219–226.
Karcz I. and Zak I. (1987) Bedforms in salt deposits of the DeadSea brines. J. Sediment. Petrol. 57, 723–735.
Keeling C. D. (1958) The concentration and isotopic abundances ofatmospheric carbon dioxide in rural areas. Geochim. Cos-
mochim. Acta 13, 322–334.Kovac N. (2009) Chemical characterization of stromatolitic
‘‘Petola” layer (Secovlje Salt-Pans, Slovenia) using FT-IRspectroscopy. Annales. Ser. Hist. Nat. 19, 95–102.
Krumgalz B. S. (1980) Salt effect on the pH of hypersalinesolutions. In Hypersaline Brines and Evaporitic Environments.
Developments in Sedimentology, vol. 28 (ed. A. Nissenbaum).Elsevier, Amsterdam, pp. 73–83.
Kusch S., Kashiyama Y., Ogawa N. O., Altabet M., Butzin M.,Friedrich J., Ohkouchi N. and Mollenhauer G. (2010) Impli-cations for chloro- and pheopigment synthesis and preservationfrom combined compound-specific d13C, d15N, and D14Canalysis. Biogeosci. Discuss. 7, 6265–6294.
Lazar B. and Erez J. (1992) Carbon geochemistry of marine-derived brines: I. 13C depletions due to intense photosynthesis.Geochim. Cosmochim. Acta 56, 335–345.
Lazar B., Starinsky A., Katz A., Sass E. and Ben-Yaakov S. (1983)The carbonate system in hypersaline solutions: alkalinity andCaCO3 solubility of evaporated seawater. Limnol. Oceanogr.
28, 978–986.Ley R. E., Harris J. K., Wilcox J., Spear J. R., Miller S. R., Bebout
B. M., Maresca J. A., Bryant D. A., Sogin M. L. and Pace N.R. (2006) Unexpected diversity and complexity of the GuerreroNegro hypersaline microbial mat. Appl. Environ. Microbiol. 72,3685–3695.
Li Y. H. and Tsui T. F. (1971) The solubility of CO2 in water andsea water. J. Geophys. Res. 76, 4203–4207.
Logan B. W. (1987) The MacLeod evaporite basin, westernAustralia. Holocene environments, sediments and geologicalevolution. AAPG Memoir. 44, 1–140.
McCaffrey M. A., Lazar B. and Holland H. D. (1987) Theevaporation path of seawater and the coprecipitation of Br�
and K+ with halite. J. Sediment. Petrol. 57, 928–937.Meyers P. A. and Eadie B. J. (1993) Sources, degradation and
recycling of organic matter associated with sinking particles inLake Michigan. Org. Geochem. 20, 47–56.
Mook W. G., Bommerson J. C. and Staverman W. H. (1974)Carbon isotope fractionation between dissolved bicarbonateand gaseous carbon dioxide. Earth Planet. Sci. Lett. 22, 169–176.
Nadler A. and Magaritz M. (1980) Studies of marine solutionbasins – isotopic and compositional changes during evapora-tion. In Hypersaline Brines and Evaporitic Environments.
Developments in Sedimentology, vol. 28 (ed. A. Nissenbaum).Elsevier, Amsterdam, pp. 115–129.
Ogawa N. O., Nagata T., Kitazato H. and Ohkouchi N. (2010)Ultra sensitive elemental analyzer/isotope ratio mass spectrom-

Y. Isaji et al. /Geochimica et Cosmochimica Acta 208 (2017) 354–367 367
eter for stable nitrogen and carbon isotope analyses. In Earth,
Life, and Isotopes (eds. N. Ohkouchi, I. Tayasu and K. Koba).Kyoto University Press, Kyoto, pp. 339–353.
Ohkouchi N., Nakajima Y., Okada H., Ogawa N. O., Suga H.,Oguri K. and Kitazato H. (2005) Biogeochemical processes inthe saline meromictic Lake Kaiike, Japan: implications frommolecular isotopic evidences of photosynthetic pigments. Env-iron. Microbiol. 7, 1009–1016.
Ohkouchi N., Nakajima Y., Ogawa N. O., Chikaraishi Y., SugaH., Sakai S. and Kitazato H. (2008) Carbon isotopic compo-sition of the tetrapyrrole nucleus in chloropigments from asaline meromictic lake: a mechanistic view for interpreting theisotopic signature of alkyl porphyrins in geological samples.Org. Geochem. 39, 521–531.
Ollivier B., Caumette P., Garcia J. L. and Mah R. A. (1994)Anaerobic bacteria from hypersaline environments. Microbiol.
Mol. Biol. Rev. 58, 27–38.Oren A. (2002) Diversity of halophilic microorganisms: environ-
ments, phylogeny, physiology, and applications. J. Ind. Micro-
biol. Biotechnol. 28, 56–63.Oren A. (2005) A hundred years of Dunaliella research: 1905–2005.
Saline Systems 1, 1.Oren A. (2009) Saltern evaporation ponds as model systems for the
study of primary production processes under hypersalineconditions. Aquat. Microb. Ecol. 56, 193–204.
Oren A., Sørensen K. B., Canfield D. E., Teske A. P., Ionescu D.,Lipski A. and Altendorf K. (2009) Microbial communities andprocesses within a hypersaline gypsum crust in a salternevaporation pond (Eilat, Israel). Hydrobiologia 626, 15–26.
Orphan V. J., Jahnke L. L., Embaye T., Turk K. A., Pernthaler A.,Summons R. E. and Des Marais D. J. (2008) Characterizationand spatial distribution of methanogens and methanogenicbiosignatures in hypersaline microbial mats of Baja California.Geobiology 6, 376–393.
Pancost R. D., Freeman K. H., Wakeham S. G. and Robertson C.Y. (1997) Controls on carbon isotope fractionation by diatomsin the Peru upwelling region. Geochim. Cosmochim. Acta 61,4983–4991.
Pinckney J., Paerl H. W. and Bebout B. M. (1995) Salinity controlof benthic microbial mat community production in a Bahamianhypersaline lagoon. J. Exp. Mar. Biol. Ecol. 187, 223–237.
Popp B. N., Laws E. A., Bidigare R. R., Dore J. E., Hanson K. L.and Wakeham S. G. (1998) Effect of phytoplankton cellgeometry on carbon isotope fractionation. Geochim. Cos-
mochim. Acta 62, 69–77.Prins H. B. A. and Elzenga J. T. M. (1989) Bicarbonate utilization:
function and mechanism. Aqua. Bot. 34, 59–83.Raven J. A. (1991) Implications of inorganic carbon utilization:
ecology, evolution, and geochemistry. Can. J. Bot. 69, 908–924.Rehakova K., Zapomelova E., Prasil O., Vesela J., Medova H. and
Oren A. (2009) Composition changes of phototrophic microbialcommunities along the salinity gradient in the solar salternevaporation ponds of Eilat, Israel. Hydrobiologia 636, 77–88.
Revsbech N. P., Jørgensen B. B., Blackburn T. H. and Cohen Y.(1983) Microelectrode studies of the photosynthesis and O2,H2S, and pH profiles of a microbial mat. Limnol. Oceanogr. 28,1062–1074.
Risatti J. B., Capman W. C. and Stahl D. A. (1994) Communitystructure of a microbial mat: the phylogenetic dimension. Proc.Natl. Acad. Sci. USA 91, 10173–10177.
Romanek C., Grossman E. and Morse J. (1992) Carbon isotopicfractionation in synthetic calcite, effects of temperature andprecipitation rate. Geochim. Cosmochim. Acta 56, 419–430.
Roveri M., Flecker R., Krijgsman W., Lofi J., Lugli S., Manzi V.,Sierro F. J., Bertini A., Camerlenghi A., De Lange G., GoversR., Hilgen F. J., Hubscher C., Meijer P. and Stoica M. (2014)The Messinian Salinity Crisis: past and future of a greatchallenge for marine sciences. Mar. Geol. 352, 25–58.
Sachs J. P. and Repeta D. J. (1999) Oligotrophy and nitrogenfixation during eastern Mediterranean sapropel events. Science286, 2485–2488.
Sass E. and Ben-Yaakov S. (1977) The carbonate system inhypersaline solutions: dead Sea brines. Mar. Chem. 5, 183–199.
Schouten S., Hartgers W. A., Lopez J. F., Grimalt J. O. andDamste J. S. S. (2001) A molecular isotopic study of 13C-enriched organic matter in evaporitic deposits: recognition ofCO2-limited ecosystems. Org. Geochem. 32, 277–286.
Sørensen K. B., Canfield D. E. and Oren A. (2004) Salinityresponses of benthic microbial communities in a solar saltern(Eilat, Israel). Appl. Environ. Microbiol. 70, 1608–1616.
Stiller M., Rounick J. S. and Shasha S. (1985) Extreme carbon-isotope enrichments in evaporating brines. Nature 316, 434–435.
Teske A., Ramsing N. B., Habicht K., Fukui M., Kuver J.,Jørgensen B. B. and Cohen Y. (1998) Sulfate-reducing bacteriaand their activities in cyanobacterial mats of Solar Lake (Sinai,Egypt). Appl. Environ. Microbiol. 64, 2943–2951.
Timofeeff M. N., Lowenstein T. K., Brennan S. T., Demicco R. V.,Zimmermann H., Horita J. and Von Borstel L. E. (2001)Evaluating seawater chemistry from fluid inclusions in halite:examples from modern marine and nonmarine environments.Geochim. Cosmochim. Acta 65, 2293–2300.
Tyler J., Kashiyama Y., Ohkouchi N., Ogawa N. O., YokoyamaY., Chikaraishi Y., Staff R. A., Ikehara M., Ramsey C. B.,Bryant C., Brock F., Gotanda K., Haraguchi T., Yonenobu H.and Nakagawa T. (2010) Tracking aquatic change usingchlorine-specific carbon and nitrogen isotopes: the last glacial-interglacial transition at Lake Suigetsu, Japan. Geochem.
Geophys. Geosyst. 11, Q09010.Valdivieso-Ojeda J. A., Huerta-Diaz M. A. and Delgadillo-
Hinojosa F. (2014) High enrichment of molybdenum inhypersaline microbial mats of Guerrero Negro, Baja CaliforniaSur, Mexico. Chem. Geol. 363, 341–354.
Van Gemerden H. (1993) Microbial mats: a joint venture. Mar.
Geol. 113, 3–25.Ware J. R., Smith S. V. and Reaka-Kudla M. L. (1992) Coral reefs:
sources or sinks of atmospheric CO2? Coral Reefs 11(3), 127–130.
Warren J. K. (2010) Evaporites through time: tectonic, climatic andeustatic controls in marine and nonmarine deposits. Earth-Sci.Rev. 98, 217–268.
Wieland A. and Kuhl M. (2006) Regulation of photosynthesis andoxygen consumption in a hypersaline cyanobacterial mat(Camargue, France) by irradiance, temperature and salinity.FEMS Microbiol. Ecol. 55, 195–210.
Wieland A., de Beer D., Damgaard L. R., Kuhl M., VanDusschoten D. and Van As H. (2001) Fine-scale measurementof diffusivity in a microbial mat with nuclear magneticresonance imaging. Limnol. Oceanogr. 46, 248–259.
Wieland A., Zopfi J., Benthien M. and Kuhl M. (2005) Biogeo-chemistry of an iron-rich hypersaline microbial mat (Camargue,France). Microb. Ecol. 49, 34–49.
Associate editor: Robert Byrne





























