Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, JUlY 1989, p. 2223-22290019-9567/89/072223-07$02.00/0Copyright ©D 1989, American Society for Microbiology
Biological Activities and Chemical Composition of Purified TrachealCytotoxin of Bordetella pertussis
BRAD T. COOKSON, HWEI-LING CHO, LOREEN A. HERWALDT, AND WILLIAM E. GOLDMAN*
Department of Microbiology and Immunology, Washington University School of Medicine, St. Louis, Missouri 63110
Received 23 December 1988/Accepted 25 March 1989
Specific destruction of ciliated epithelial cells lining the large airways is the primary respiratory tractcytopathology associated with human Bordetella pertussis infections. We have purified a single low-molecular-weight glycopeptide, tracheal cytotoxin (TCT), that appears to cause this pathology. By using a combinationof solid-phase extraction and reversed-phase high-pressure liquid chromatography, about 700 nmol ofbiologically active peptide can be isolated from 1 liter of B. pertussis culture supernatant (approximately 60%yield). TCT at concentrations of 1 ,uM destroyed the ciliated cell population when incubated with respiratoryepithelium in vitro. This concentration of TCT is similar to the concentrations found in the culture supernatantof growing B. pertussis. Purified TCT also inhibited DNA synthesis of hamster trachea epithelial cells in aquantitative, dose-dependent fashion. Endotoxin was not detected in the purified material, and neither B.pertussis nor Escherichia coli endotoxin could duplicate the biological activities of TCT. Amino acid and aminosugar analyses of purified TCT revealed the presence of glucosamine, muramic acid, alanine, glutamic acid,and diaminopimelic acid in molar ratios of 1:1:2:1:1. This suggests that TCT, the released ciliostatic principleof B. pertussis, is a disaccharide tetrapeptide subunit of peptidoglycan.
Bordetella pertussis is a gram-negative bacterium thatcauses the human respiratory tract illness known as whoop-ing cough (or pertussis). The characteristic pathology asso-ciated with the disease, which features specific bacterialcolonization of and destruction of ciliated cells, was firstdescribed three-quarters of a century ago by Mallory andHorner (19). The consequences of this cytopathology can besevere, because the loss of ciliary activity significantlyimpairs mucus transport out of the respiratory tract. Accu-mulation of mucus, multiplying bacteria, and inflammatorydebris in the absence of a functional mucociliary "escalator"give rise to a situation where coughing is the only remainingmeans to clear the blocked airways. Therefore, damage tothe ciliated cells may generate the paroxysmal coughingepisodes characteristic of pertussis as well as predispose theinfected host to secondary pulmonary infections (24).The cytopathological events during B. pertussis infection
of the respiratory tract have been studied by using trachealorgan culture as an in vitro model system (7). With trachealrings dissected from hamsters, infection with B. pertussis issimilar to that in humans: a noninvasive, specific coloniza-tion of ciliated cells, followed by ciliostasis and the eventualextrusion of the ciliated cell population (23). Although thereare a variety of biologically active virulence-associatedmacromolecules produced by B. pertussis (for a review, seereference 32), only a low-molecular-weight fraction of bac-terial culture supernatant can mimic this ciliated-cell-specificpathology when incubated with tracheal organ cultures (15).
Furthermore, hamster trachea epithelial (HTE) cells (13),to which virulent B. pertussis can adhere, are unable tosynthesize DNA in the presence of viable B. pertussis. Thiseffect on HTE cells can be reproduced by the addition of thesame low-molecular-weight fraction of culture supernatantthat destroys tracheal organ culture (15). We speculate thatthis inhibitory activity of B. pertussis may delay the normalreplacement of destroyed ciliated cells in vivo by blockingdivision and differentiation of the underlying regenerative
* Corresponding author.
basal cell population (18). This notion is consistent with theobservation that the clinical course of pertussis is unchangedby antibiotic therapy initiated after the onset of coughingparoxysms (24). Viable B. pertussis organisms are no longerpresent, yet manifestations of active infection, including thecoughing episodes, can continue for weeks.To understand more fully the ciliated-cell-specific pathol-
ogy that results from B. pertussis infections, we sought topurify the tracheal cytotoxin (TCT) activity of culture super-natants to homogeneity. Here we report the isolation andcomposition of TCT-a single molecule produced by B.pertussis that specifically destroys ciliated respiratory epi-thelial cells.
MATERIALS AND METHODSBacteria. Virulent phase I B. pertussis Tohama I was
stored at -70°C in a solution containing Casamino Acids(Difco Laboratories, Detroit, Mich.), MgCI2, CaCl2, andNaCl with 50% glycerol (30). Bacteria were grown on solidmodified Stainer-Scholte medium (SSM) (16, 31) supple-mented with 10% fresh defibrinated sheep blood in Alsevers(GIBCO Laboratories, Grand Island, N.Y.) by incubating at37°C for 72 h in a humidified atmosphere containing 95%air-5% CO2. We used the resulting growth to inoculate liquidmedium consisting of SSM containing 10% Casamino Acids.After incubation on a rotary shaker (150 rpm) at 37°C for 24to 48 h in an atmosphere of 95% air-5% C02, the late-log-phase cells were harvested by centrifugation (15 min at2,200 x g), washed once with SSM, and suspended in freshSSM without Casamino Acids. Using these organisms weinoculated fresh cultures to approximately 107 bacteria perml (based on standardized A540) and incubated them on arotary shaker as described above.
Solid-phase extraction. Bacteria growing in the mid- tolate-log phase (-4 x 109 bacteria per ml) were centrifuged at13,200 x g for 20 min at 4°C. The supernatant was filteredthrough 0.2-,um-pore-size cellulose-acetate membrane filterunits (Corning Glass Works, Coming, N.Y.) and acidified topH 3 with 100% trifluoroacetic acid (TFA) to a final concen-
2223
Vol. 57, No. 7
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2224 COOKSON ET AL.
tration of 0.5%. Portions of this solution were poured into aglass column (1.5 by 22 cm), which served as a samplereservoir. At the tapered end of the column we attached aC18 Sep-Pak (Waters Associates, Milford, Mass.) that hadbeen prepared to receive the sample by wetting twice, firstwith 100% methanol and then with 0.1% TFA in water. Allsolutions were forced through Sep-Paks by pressurized air.After complete extraction of the sample of interest, theSep-Pak was washed with two 10-ml volumes of 0.1% TFAin water. We then eluted some of the retained molecules bythe addition of 3 ml of 20% n-propyl alcohol in 0.1% TFAand concentrated them to dryness in a rotary evaporator(Speed Vac Concentrator; Savant, Farmingdale, N.Y.). Theresulting C18 fraction was characterized by reversed-phasehigh-pressure liquid chromatography (HPLC) analysis (seebelow) and used for the subsequent solid-phase extraction.We used quaternary methylamine (QMA) Sep-Pak car-
tridges (ACCELL media; Waters Associates) attached to theend of a 10-ml plastic syringe for anion-exchange solid-phaseextraction. The exchange matrix was sequentially wettedwith 10 mM ammonium acetate (pH 5.5) containing 20%methanol (running buffer), 10 mM ammonium acetate (pH5.5) containing 1 M NaCl and 20% methanol (elution buffer),100% methanol, and again with running buffer. After loadingthe C18 fraction onto the QMA Sep-Pak cartridge in runningbuffer, we washed it with 5 ml of the same buffer and elutedthe retained molecules with 5 ml of elution buffer. Thiseluate was concentrated to a volume of approximately 2.5 mlby rotary evaporation, acidified by the addition of 10% TFAto a final concentration of 0.5%, and desalted by subjectingit to C18 solid-phase extraction as described above. Afterconcentration to dryness by rotary evaporation, the anion-exchange-extracted sample was stored at -70°C beforereversed-phase HPLC analysis.
Reversed-phase HPLC. We performed HPLC with WatersAssociates equipment, except where noted. Aquapore Octyl(C8) RP-300 cartridge columns (Brownlee Labs, Rainin In-strument Co., Woburn, Mass.) maintained at ambient tem-perature were used for all chromatographic separations: weemployed a 130- by 4.6-mm (inner diameter) column con-taining 7-,um spherical particles of C8-derivatized silica witha solvent delivery rate of 1 ml/min. UV-absorbing compo-nents of the column effluent were detected at 214 nm byusing a Spectroflow 757 variable-wavelength UV detector(Kratos, Ramsey, N.J.), and peak areas and retention timeswere recorded by using a Hitachi Model D-2000 Chromato-Integrator (Hitachi, Ltd., Tokyo, Japan). We collectedpeaks of interest in borosilicate glass tubes and stored themat -70°C after removal of the volatile mobile phase by rotaryevaporation. For making mobile phases, we filtered allsolvents with a Millipore all-glass filtration apparatus withDurapore 0.45-,um-pore-size membranes (Millipore Corp.,Bedford, Mass.) and degassed the acetonitrile, methanol,and purified water before mixing. All organic reagents wereHPLC/Spectro grade (Pierce Chemical Co., Rockford, Ill.),and water was distilled and deionized (NANOpure; Barn-stead Co., Newton, Mass.).Two distinct chromatographic systems were employed to
purify TCT from solid-phase extracts: reversed-phase HPLCby gradient elution with methanol in a triethylamine acetate(TEA-acetate)-buffered mobile phase and gradient elutionwith acetonitrile in a TFA-buffered mobile phase. The TEA-acetate buffer was prepared as described by Matrisian et al.(21), except that a 0.01 M acetic acid solution was brought topH 5.55 by the addition of triethylamine. The aqueousportion of the mobile phase (solution A) consisted of 0.01 M
TEA-acetate (pH 5.55), and solution B consisted of solutionA containing methanol at a final concentration of 30%(voUvol). In the TEA-acetate-buffered system, a convexgradient from 0 to 7.5% methanol developed over a period of30 min. This gradient was preceded by a 5-min isocraticloading and washing interval with A and followed by a 3-minisocratic elution with 3A:1B. Subsequent purging with B for5 min removed any adsorbed molecules remaining on thecolumn, and equilibration with A for at least 20 min beforeanother sample injection insured reproducible chromatogra-phy.For the TFA-buffered system, solution A consisted of
0.1% TFA in water (pH approximately 3), and solution Bconsisted of solution A plus acetonitrile at a final concentra-tion of 30% (vol/vol). A linear gradient from 0 to 9.0%acetonitrile proceeded over an interval of 30 min. Otheraspects of the TFA system were similar to those of theTEA-acetate system. In both buffer systems we found it tobe extremely important that the final volume of the Bsolution contained the correct concentration of bufferingagents, i.e., TFA or TEA-acetic acid.
Analytical methods. The Protein Chemistry Laboratory atWashington University School of Medicine conducted theamino acid and amino sugar analyses. Samples were hydro-lyzed under vacuum in 6 N HCI for 24 h at 110°C for aminoacid analyses or in 4 N HCl for 6 h at 100°C for amino sugaranalyses. They identified amino group-containing compo-nents in the hydrolyzed samples by using standard tech-niques employing ion-exchange HPLC.We examined test samples for contaminating endotoxin by
the chromogenic Limulus amebocyte lysate assay (QCL-1000; Whittaker Bioproducts, Inc., Walkersville, Md.). Pu-rified Escherichia coli lipopolysaccharide serotype 026:B6(Sigma Chemical Co., St. Louis, Mo.) was used as a controlfor lysate reactivity, and purified B. pertussis lipopolysac-charide (List Biological Laboratories, Inc., Campbell, Calif.)was used to construct a standard curve. Each assay wascarried out according to the recommendations of the sup-plier.We determined protein concentrations with the Bio-Rad
protein Assay (Bio-Rad Laboratories, Richmond, Calif.)with bovine serum albumin as a standard.
Assessment of biological activity. HTE cells were isolatedand cultured as described previously (13), except that weused 9 ,ug of gentamicin per ml instead of penicillin andstreptomycin. HTE cells were harvested from nearly con-fluent monolayers, seeded at 1.5 x 104 cells per microtiterwell in bicarbonate-buffered minimal essential medium con-taining 2.5% fetal bovine serum (HyClone, Logan, Utah),and incubated for 24 h at 37°C in a humidified environmentcontaining 95% air-5% CO2. We removed the medium fromeach well with gentle suction and replaced it with theappropriate control or test samples dissolved in 5 mMHEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid)-buffered minimal essential medium (pH 7.3). PurifiedTCT was added in the presence of bovine serum albumin asa protein protectant and carrier; control samples also in-cluded bovine serum albumin. After incubation for 4 h at370C in a humidified environment (without C02), the DNA-synthetic ability of sample-treated cells and untreated con-trol cells was quantitated by measuring [3H]thymidine incor-poration after serum stimulation as described previously(15).Tracheas dissected from young (6 to 12 weeks) male
golden Syrian hamsters (Charles River Breeding Laborato-ries, Inc., Wilmington, Mass.) were used to prepare tracheal
INFECT. IMMUN.
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BORDETELLA PERTUSSIS TRACHEAL CYTOTOXIN 2225
A.
E
CM
cu
-)0c
o0Cu
B.
EC
a)co00C
E.00
.0(U
I
I IIII I,,,,. , I. gI g,II,,I ,I III.I II
10 20 30 40
retention time (minutes)
,,.,,,,,,.,,,,,,.,,*,.,,,,,,,I,,,,,.,,
10 20 30 40
retention time (minutes)FIG. 1. TFA-buffered reversed-phase HPLC analysis of solid-phase extracts. The peak with toxic activity is indicated by the arrows. (A)
Retained components from a C18 extract of 25 ml of B. pertussis culture supernatant (0.128 A214 units full scale). (B) Anionic componentsobtained by ion-exchange extraction of the C18 extract in A (0.064 A214 units full scale).
rings manually (6) or with a tissue chopper (custom design byRichard McDonald; details available upon request). Cultureswere grown in nutrient mixture F-12 (GIBCO) buffered with5 mM HEPES (pH 7.3) in a humidified environment at 37°C.To evaluate the biological activity of sterile test samples, weobserved rings for pathology at 24-h intervals with aninverted light microscope under x 10 and x 20 power.
RESULTS
Extraction of TCT from culture supernatants. To devise aneffective purification scheme, we chose solid-phase extrac-tion to prepare samples for more rigorous isolation proce-dures. In the first step of a rapid method, we used areversed-phase matrix (C18) to concentrate tracheal cyto-toxic activity from the culture supernatant of B. pertussisgrowing in the mid- to late-log phase. Once the supernatanthad been acidified and passed through the C18 Sep-Pak,nonspecifically bound substances were washed away andadsorbed molecules were selectively eluted by the additionof 0.1% TFA in water containing 20% n-propyl alcohol.Under these conditions, a peptide-rich fraction can be ob-tained that is free of large polypeptides, salts, and free aminoacids (2-4, 29). HPLC analysis of this relatively complexextract revealed the presence of one peak with cytotoxicactivity (Fig. 1A).Amino acid analysis of partially purified material sug-
gested that TCT may have significant anionic or acidiccharacter. Therefore, anion exchange was chosen to frac-
tionate acidic molecules from basic and/or neutral moleculespresent in the TCT-enriched eluate from the C18 extraction.The C18 extract was loaded onto the strong anion-exchangeQMA Sep-Pak in low-salt running buffer (pH 5.5) andwashed with the same to remove any nonspecifically boundcomponents. Adsorbed molecules with presumed anioniccharacter were then eluted from the QMA Sep-Pak withhigh-salt elution buffer, also at pH 5.5. Optimal extraction oftoxic activity into the eluted fraction occurred at pH 5.5; alower pH resulted in decreased recovery, whereas a higherpH increased the proportion of contaminating moleculesharvested without a significant increase in yield. Figure 1Bshows a typical chromatogram of the UV-absorbing compo-nents in the effluent from this ion-exchange solid-phaseextraction. Comparison of the HPLC profiles of the C18 (Fig.1A) and QMA (Fig. 1B) extracts indicates that the complex-ity of the fraction containing activity has been significantlyreduced and that several components with HPLC retentiontimes similar to the peak with cytotoxic activity have beenremoved.
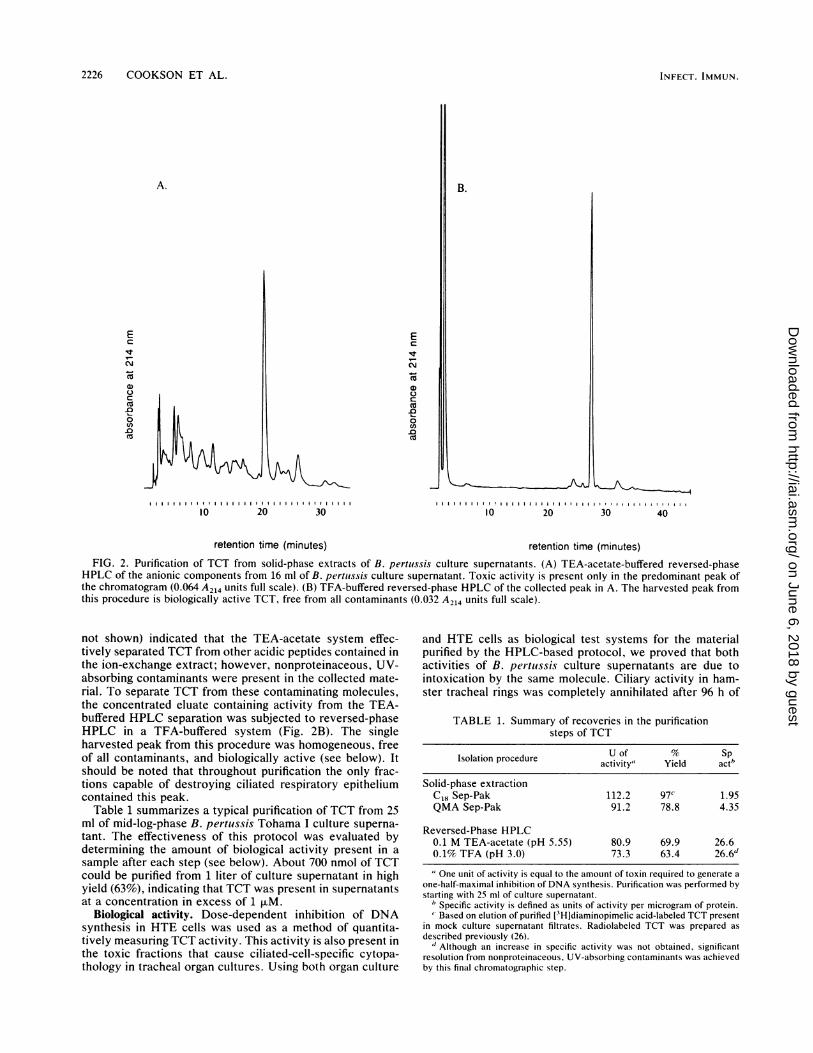
Purification of TCT from culture supernatant extracts. TCTwas completely purified from the ion-exchange extract de-scribed above by using two successive reversed-phaseHPLC separations. First, the QMA eluate was loaded onto aC8 column in TEA-acetate buffer (pH 5.55) and separated bygradient elution with methanol. A typical chromatogramresulting from this purification step is shown in Fig. 2A.Amino acid analysis of the biologically active fraction (data
VOL. 57, 1989
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
2226 COOKSON ET AL.
A.
E
C\
a)
-0
cn(0
10 20 30
retention time (minutes)
E
cxv-C\
a)cJCZ
n
co
B.
10 20 30 40
retention time (minutes)
FIG. 2. Purification of TCT from solid-phase extracts of B. pertussis culture supernatants. (A) TEA-acetate-buffered reversed-phaseHPLC of the anionic components from 16 ml of B. pertuissis culture supernatant. Toxic activity is present only in the predominant peak ofthe chromatogram (0.064 A214 units full scale). (B) TFA-buffered reversed-phase HPLC of the collected peak in A. The harvested peak fromthis procedure is biologically active TCT, free from all contaminants (0.032 A214 units full scale).
not shown) indicated that the TEA-acetate system effec-tively separated TCT from other acidic peptides contained inthe ion-exchange extract; however, nonproteinaceous, UV-absorbing contaminants were present in the collected mate-rial. To separate TCT from these contaminating molecules,the concentrated eluate containing activity from the TEA-buffered HPLC separation was subjected to reversed-phaseHPLC in a TFA-buffered system (Fig. 2B). The singleharvested peak from this procedure was homogeneous, freeof all contaminants, and biologically active (see below). Itshould be noted that throughout purification the only frac-tions capable of destroying ciliated respiratory epitheliumcontained this peak.
Table 1 summarizes a typical purification of TCT from 25ml of mid-log-phase B. pertussis Tohama I culture superna-tant. The effectiveness of this protocol was evaluated bydetermining the amount of biological activity present in a
sample after each step (see below). About 700 nmol of TCTcould be purified from 1 liter of culture supernatant in highyield (63%), indicating that TCT was present in supernatantsat a concentration in excess of 1 F.M.
Biological activity. Dose-dependent inhibition of DNAsynthesis in HTE cells was used as a method of quantita-tively measuring TCT activity. This activity is also present inthe toxic fractions that cause ciliated-cell-specific cytopa-thology in tracheal organ cultures. Using both organ culture
and HTE cells as biological test systems for the materialpurified by the HPLC-based protocol, we proved that bothactivities of B. pertiussis culture supernatants are due tointoxication by the same molecule. Ciliary activity in ham-ster tracheal rings was completely annihilated after 96 h of
TABLE 1. Summary of recoveries in the purificationsteps of TCT
Isolation procedure Uait Yield Sactb
Solid-phase extractionCl8 Sep-Pak 112.2 97C 1.95QMA Sep-Pak 91.2 78.8 4.35
Reversed-Phase HPLC0.1 M TEA-acetate (pH 5.55) 80.9 69.9 26.60.1% TFA (pH 3.0) 73.3 63.4 26.6"
" One unit of activity is equal to the amount of toxin required to generate aone-half-maximal inhibition of DNA synthesis. Purification was performed bystarting with 25 ml of culture supernatant.
"Specific activity is defined as units of activity per microgram of protein.'*Based on elution of purified [3H]diaminopimelic acid-labeled TCT present
in mock culture supernatant filtrates. Radiolabeled TCT was prepared asdescribed previously (26).
" Although an increase in specific activity was not obtained, significantresolution from nonproteinaceous, UV-absorbing contaminants was achievedby this final chromatographic step.
INFECT. IMMUN.
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BORDETELLA PERTUSSIS TRACHEAL CYTOTOXIN 2227
0 100U)
C 80
z 60
0 40 /
C
0D0 2o0
~~~~0~~~~110.1 1 10Concentration (>M)
FIG. 3. Dose-response analysis of purified TCT: inhibition ofHTE cell DNA synthesis. All points were calculated by comparingcells treated with TCT with cells treated with the control sample intriplicate. Each point represents the average of values <10% fromthe standard deviation of the mean.
incubation with purified TCT at 1 F.M and after 48 to 72 hwhen incubated with 3 ,uM TCT. Figure 3 is a representativedose-response curve showing inhibition of DNA synthesis inHTE cells by purified TCT. A threshold for an inhibitoryresponse was observed, and maximal inhibition occurred atconcentrations of about 1 F.M. This is in contrast to thelower specific activity and more shallow dose-responsecurve reported with less purified material (14, 15). Weattribute this improved biological activity to the removal ofcontaminating molecules interfering with this assay for tra-cheal cytotoxic activity. Importantly, TCT is found in B.pertussis culture supernatants at concentrations sufficient toelicit the destruction of respiratory epithelium (see above).To establish that the observed biological responses were
in fact due to TCT and not to contaminating endotoxin, testsamples were evaluated for the presence of endotoxin byusing the Limulus amebocyte lysate assay. No detectableendotoxin (<10 fg per 6.5 ,ug of TCT) was present in samplesthat had been subjected to either TFA- or TEA-bufferedreversed-phase HPLC. In C18 solid-phase extracts of B.pertussis culture supernatants, we detected endotoxin con-centrations as high as 10 ng/ml. However, purified B. per-tussis or E. coli endotoxin at concentrations as high as 0.1p.g/ml neither inhibited DNA synthesis of HTE cells norcaused ciliated-cell-specific cytopathology in organ cultures(data not shown). This strongly suggests that contaminatingendotoxin is not responsible for the biological activitiesdescribed above for purified TCT.
Composition. To characterize further the biologically ac-tive material, we subjected samples of TCT to amino acidand amino sugar analyses (Table 2). The previously reported
TABLE 2. Composition of purified B. pertussistracheal cytotoxin
Amino acid or Molar No. ofamino sugar ratio' residues
Glucosamine 0.83 1Muramic acid 0.78 1Alanine 1.7 2Glutamic acid 1.0 1Diaminopimelic acid 0.72 1
" Relative to glutamic acid.
composition of partially purified toxic activity prepared byconventional chromatography (12, 15) indicated the pres-ence of contaminating amino acids, amino sugars, or pep-tides. HPLC analysis of such preparations revealed multiplepeaks, confirming the presence of impurities not removed bygel filtration chromatography and high-voltage paper electro-phoresis (data not shown). A single UV-absorbing peak washarvested in the final step of the HPLC-based purificationprotocol (Fig. 2B), suggesting that the composition of TCTreflects its identity as a single glycopeptide. Its constituentresidues are typical components of B. pertussis peptidogly-can (9) and of bacterial peptidoglycan in general (27). Themolar ratios of amino acids and amino sugars in purifiedsamples are consistent with the notion that TCT is a disac-charide-tetrapeptide subunit of peptidoglycan.
DISCUSSION
In this report, we have described the purification of asingle peptidoglycan-related molecule called TCT from thesupernatant of growing B. pertussis. TCT causes ciliated-cell-specific respiratory tract pathology in vitro that is indis-tinguishable from the primary pathology seen in humanpertussis infections. In addition, TCT inhibits DNA synthe-sis of HTE cells in a quantitative, dose-dependent fashion, afinding that may also have pathogenic significance in vivo.The purification scheme utilizes both solid-phase extrac-
tion and reversed-phase HPLC technologies to harvest bio-logically active TCT; the technique is rapid and delivers ahigh yield of TCT that is free from all contaminants. Solid-phase extraction was a useful method to employ because it issimple, fast, inexpensive, and reproducible while offering anintermediate level of chemical selectivity with high loadcapacity. This allowed us to concentrate the tracheal cyto-toxic activity present in 1 liter of bacterial culture superna-tant by over 1,000-fold (reversed-phase matrix) and to par-tition it with molecules of acidic character from basic andneutral populations (ion-exchange matrix). TCT was sepa-rated from this mixture of acidic molecules by reversed-phase HPLC in buffer systems at a pH above (pH 5.55) andbelow (pH 3) the PKa of the constituent acidic residues ofTCT. These buffers presumably induced changes in thepolarity of the molecule, correspondingly altered its chro-matographic character, and permitted us to isolate TCT tohomogeneity. Thus, we determined that a single moleculefrom the culture supernatant of B. pertussis was capable ofcausing the primary respiratory tract pathology associatedwith pertussis.The speed and efficiency with which the separation can be
performed makes the method useful as an analytical tool.This has allowed us to examine other members of theBordetella genus, which infect a variety of warm-bloodedhosts, for the ability to produce TCT. We found thatrepresentative strains of each Bordetella species releasemeasurable amounts of TCT (10; B. T. Cookson and W. E.Goldman, J. Cell. Biochem. 11B:124, 1987). This is consis-tent with the observations that all virulent Bordetella strainscause remarkably similar illnesses and that they specificallycolonize and destroy ciliated cells in the respiratory epithe-lium of their respective hosts. Furthermore, it supports thenotion that TCT, a single molecule that can mimic theprimary cytopathology of these infections, is a conservedvirulence determinant of all Bordetella species.
This analytical system can resolve various classes ofpeptidoglycan fragments in addition to TCT, the majorfragment released by growing B. pertussis (26). Although the
VOL. 57, 1989
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2228 COOKSON ET AL.
released material represents only a small percentage of themacromolecular peptidoglycan of B. pertussis organisms,our experiments show that TCT is produced at levels suffi-cient to evoke significant biological effects. Since TCT isundetectable by pulse-chase-type experiments designed tolook at turnover of macromolecular peptidoglycan (25, 28),detecting soluble fragments released by bacterial microor-ganisms may require more direct methods (like those de-scribed here). The use of HPLC to separate fragments ofpurified peptidoglycan has been reported by workers inseveral laboratories (8, 11, 20), but its use has been limited tothe analysis of enzymatic digestions of high-molecular-weight cell wall material precipitated by hot sodium dodecylsulfate. TCT, for instance, is refactory to commonly usedprotein precipitation procedures (unpublished observations).This suggests that implementation of solid-phase extractionto harvest peptide extracts would greatly expedite the anal-ysis of solutions thought to contain soluble peptidoglycanfragments. This technique could be widely applicable toavenues of research such as the investigation of othermucosal pathogens for production of TCT or TCT-likemolecules, the detection of peptidoglycan in host tissue (seebelow), the analysis of antibiotic-induced bacterial autolyticproducts, and the study of recycling of peptidoglycan fornew cell wall synthesis in bacteria.The peptidoglycan-derived composition of TCT estab-
lishes that it is one of the muramyl peptides (muramicacid-containing glycopeptides), a family of molecules withdiverse biological activities. Well-documented effects ofthese molecules include pyrogenicity, adjuvanticity, arthri-togenicity, stimulation of leukocytes to produce interleukin-1, and induction of slow-wave sleep (for reviews, see refer-ences 1, 5, and 17). Of particular interest is the observationthat Neisseria gonorrhoeae releases anhydromuramic acid-containing disaccharide peptides (27, 28) that are capable ofcausing ciliated-cell-specific damage to fallopian tube mu-cosa (22). Results reported here and our collaborative workwith Rosenthal et al. (26) indicate that TCT has an analogousdisaccharide-tetrapeptide arrangement. Presumably TCT isderived from macromolecular Bordetella peptidoglycan, theend product of a multigene biosynthetic pathway. We there-fore conclude that TCT is not a clonable gene product andthat biochemical approaches will be necessary to elucidatefurther the pathobiological roles of TCT in disease. We haveaccomplished the first step of such an approach by develop-ing a rapid method of isolating completely purified andbiologically active TCT in high yield. Homogeneous materialwill allow us to look at host cell receptors and/or target sitesthrough which TCT exerts its toxic effects on respiratoryepithelium. Armed with powerful separation and analysistools and the appropriate biological model systems, we arenow exploring the toxic effects of TCT on epithelia.
ACKNOWLEDGMENTS
This work has been supported by Public Health Service grantA122243 from the National Institutes of Health, by grant 1708 fromthe Council for Tobacco Research, USA, Inc., and by Public HealthService training grants GM-07200 and AI-07172 from the NationalInstitutes of Health to Washington University School of Medicine.
LITERATURE CITED1. Adam, A., and E. Lederer. 1984. Muramyl peptides: immuno-
modulators, sleep factors, and vitamins. Med. Res. Rev. 4:111-152.
2. Bennett, H. P. J., C. A. Browne, and S. Solomon. 1981. Purifi-cation of the two major forms of rat pituitary corticotropin using
only reversed-phase liquid chromatography. Biochemistry 20:4530-4538.
3. Bennett, H. P. J., A. M. Hudson, L. Kelly, C. McMartin, andG. E. Purdon. 1978. A rapid method, using octadecasilyl-silica,for the extraction of certain peptides from tissues. Biochem. J.175:1139-1141.
4. Bohlen, P., F. Castillo, N. Ling, and R. Guillemin. 1980. Purifi-cation of peptides: an efficient procedure for the separation ofpeptides from amino acids and salt. Int. J. Pept. Protein Res.16:306-310.
5. Chedid, L. 1983. Muramyl peptides as possible endogenousimmunopharmacological mediators. Microbiol. Immunol. 27:723-732.
6. Collier, A. M. 1976. Techniques for establishing tracheal organcultures. TCA (Tissue Cult. Assoc.). Man. 2:333-334.
7. Collier, A. M., L. P. Peterson, and J. B. Baseman. 1977.Pathogenesis of infection with Bordetella pertussis in hamstertracheal organ culture. J. Infect. Dis. 136:S196-S203.
8. Dougherty, T. J. 1985. Analysis of Neisseria gonorrhoeaepeptidoglycan by reverse-phase high-pressure liquid chromatog-raphy. J. Bacteriol. 163:69-74.
9. Folkening, W. J., W. Nogami, S. A. Martin, and R. S. Rosenthal.1987. Structure of Bordetella pertussis peptidoglycan. J. Bacte-riol. 169:4223-4227.
10. Gentry-Weeks, C. R., B. T. Cookson, W. E. Goldman, R. B.Rimler, S. Porter, and R. Curtiss III. 1988. Dermonecrotic toxinand tracheal cytotoxin: putative virulence factors of Bordetellaavium. Infect. Immun. 56:1698-1707.
11. Glauner, B., and U. Schwarz. 1983. The analysis of mureincomposition with high-pressure-liquid chromatography, p. 29-34. In R. Hakenbeck, J.-V. Holtje, and H. Labischinski (ed.),The target of penicillin. Proceedings of the International FEMSSymposium. Walter de Gruyter, Berlin.
12. Goldman, W. E. 1986. B(ordetella pertussis tracheal cytotoxin:damage to the respiratory epithelium, p. 65-69. In L. Leive,P. F. Bonventre, J. A. Morello, S. D. Silver, and H. C. Wu(ed.), Microbiology-1986. American Society for Microbiology,Washington, D.C.
13. Goldman, W. E., and J. B. Baseman. 1980. Selective isolationand culture of a proliferating epithelial cell population from thehamster trachea. In Vitro 16:313-319.
14. Goldman, W. E., and L. A. Herwaldt. 1985. Bordetella pertussistracheal cytotoxin. Dev. Biol. Stand. 61:103-111.
15. Goldman, W. E., D. G. Klapper, and J. B. Baseman. 1982.Detection, isolation, and analysis of a released Bordetellapertussis product toxic to cultured tracheal cells. Infect. Im-mun. 36:782-794.
16. Hewlett, E., and J. Wolff. 1976. Soluble adenylate cyclase fromthe culture medium of Bordetella pertussis: purification andcharacterization. J. Bacteriol. 127:890-898.
17. Kotani, S., M. Tsujimoto, T. Koga, S. Nagao, A. Tanaka, and S.Kawata. 1986. Chemical structure and biological activity rela-tionship of bacterial cell walls and muramyl peptides. Fed. Proc.45:2534-2540.
18. Lane, B. P., and R. Gordon. 1974. Regeneration of rat trachealepithelium after mechanical injury. 1. The relationship betweenmitotic activity and cellular differentiation. Proc. Soc. Exp.Biol. Med. 145:1139-1144.
19. Mallory, R. B., and A. A. Hornor. 1912. Pertussis: the histolog-ical lesion in the respiratory tract. J. Med. Res. 27:115-123.
20. Martin, S. A., R. S. Rosenthal, and K. Biemann. 1987. Fast atombombardment mass spectrometry and tandem mass spectrome-try of biologically active peptidoglycan monomers from Neis-seria gonorrhoeae. J. Biol. Chem. 262:7514-7522.
21. Matrisian, L. M., B. R. Larsen, J. S. Finch, and B. E. Magun.1982. Further purification of epidermal growth factor by high-performance liquid chromatography. Anal. Biochem. 125:339-351.
22. Melly, M. A., Z. A. McGee, and R. S. Rosenthal. 1984. Ability ofmonomeric peptidoglycan fragments from Neisseria gonor-rhoeae to damage human fallopian-tube mucosa. J. Infect. Dis.149:378-386.
23. Muse, K. E., A. M. Collier, and J. B. Baseman. 1977. Scanning
INFECT. IMMUN.
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

BORDETELLA PERTUSSIS TRACHEAL CYTOTOXIN
electron microscopic study of hamster tracheal organ culturesinfected with Bordetella pertussis. J. Infect. Dis. 136:768-777.
24. Olson, L. C. 1975. Pertussis. Medicine (Baltimore) 54:427-469.25. Rosenthal, R. S. 1979. Release of soluble peptidoglycan from
growing gonococci: hexaminidase and amidase activities. In-fect. Immun. 24:869-878.
26. Rosenthal, R. S., W. Nogami, B. T. Cookson, W. E. Goldman,and W. J. Folkening. 1987. Major fragment of soluble pepti-doglycan released from growing Bordetella pertussis is trachealcytotoxin. Infect. Immun. 55:2117-2120.
27. Schleifer, K. H., and 0. Kandler. 1972. Peptidoglycan types ofbacterial cell walls and their taxonomic implications. Bacteriol.Rev. 36:407-477.
28. Sinha, R. K., and R. S. Rosenthal. 1980. Release of soluble
peptidoglycan from growing gonococci: demonstration of anhy-dro-muramyl-containing fragments. Infect. Immun. 29:914-925.
29. Smith, A. I., A. B. Keith, J. A. Edwardson, J. A. Biggins, andJ. R. McDermott. 1982. Characterization of corticotropin-likeimmunoreactive peptides in rat brain using high performanceliquid chromatography. Neurosci. Lett. 30:133-138.
30. Smith, C. J., J. G. Coote, and R. Parton. 1986. R-plasmidmediated chromosome mobilization in Bordetella pertussis. J.Gen. Microbiol. 132:2685-2692.
31. Stainer, D. W., and M. J. Scholte. 1971. A simple chemicallydefined medium for the production of phase I Bordetella pertus-sis. J. Gen. Microbiol. 63:211-220.
32. Weiss, A. A., and E. L. Hewlett. 1986. Virulence factors ofBordetella pertussis. Annu. Rev. Microbiol. 40:661-686.
VOL. 57, 1989 2229
on June 6, 2018 by guesthttp://iai.asm
.org/D
ownloaded from








![MIL-STD-2223 [TEST METHODS FOR INSULATED ELECTRIC WIRE]€¦ · Title: MIL-STD-2223 [TEST METHODS FOR INSULATED ELECTRIC WIRE] Author: USA Information Systems, Inc. Subject: TEST](https://img.pdfslide.us/doc/110x75/5f65847b5b737f035a79a784/mil-std-2223-test-methods-for-insulated-electric-wire-title-mil-std-2223-test.jpg)




















