Embed Size (px)
Citation preview

BIOFILM……

4. BIOFILM
4.1 Introduction
Biofilm is defined as a structured community of microorganisms
(bacteria, fungi, algae, and protozoa) wherein the cells are enclosed in a self-
produced polymeric matrix and adherent to an inert or living surface (Davey and
O'toole, 2000; Götz, 2002). This constitutes a protected mode of growth that
allows survival of bacteria in a hostile environment, particularly in high shear
environments (i.e., rapidly flowing milieus). Many microorganisms grow on
surfaces in well protected manner, in a closed structure hence, unaffected even
when high doses of antibiotics are used. Staphylococcus biofilm has been
extensively studied in human medicine and this pathogen is considered
significant in both device associated infections and tissue infections such as
pneumonia and osteomyelitis. In dairy cattle, the prevalence of bovine
staphylococcal mastitis associated with biofilm ranges from 7% to 40%. When
biofilm is formed in a low shear environment, they are generally more sensitive
to mechanical breakage. Biofilm-associated bacteria show an innate resistance to
antibiotics, disinfectants and clearance by host defence mechanisms.
4.2 Review of literature
4.2.1 Biofilm Structure and Composition
In harsh environment, biofilm tends to be the most important necessity in
protecting microbes as it proves to enable them to survive and disperse
(Melchior et al., 2009). The way in which biofilm spreads in an environment
relies on the movement of biofilm on a surface. The below diagram, Fig 4.1.,

depicts the different movement of biofilm on a surface like rippling, streaming,
rolling and seeding dispersal.
Fig 4.1: Mobility of biofilm
Biofilm is composed primarily of microbial cells and external
polysaccharide (EPS). This primary matrix material is accounting for 50% to
90% of the total organic carbon of biofilm (Rachid et al., 2000a). EPS may vary
in chemical and physical properties, but it is primarily composed of
polysaccharides. Some of them are neutral or polyanionic, as is the case of
gram-negative bacterial EPS. The presence of uronic acids (such as D-
glucuronic, D-galacturonic, and mannuronic acids) or ketal-linked pryruvates
confers the anionic property. This property is important because it allows
association of divalent cations such as calcium and magnesium, which have
been shown to cross-link with the polymer strands and provide greater binding
force in a developed biofilm. In the case of some gram-positive bacteria, such as

68
the staphylococci, the chemical composition of EPS may be quite different and
may be primarily cationic. Scientists found that the slime of coagulase-negative
bacteria consists of a teichoic acid mixed with small quantities of proteins.
EPS may associate with metal ions, divalent cations, other
macromolecules (such as proteins, DNA and lipids). EPS production is known
to be regulated by nutrient status of the growth medium; excess available carbon
and limitation of nitrogen, potassium, or phosphate promote EPS synthesis.
Slow bacterial growth will also enhance EPS production. Because EPS is highly
hydrated, it prevents desiccation in some natural biofilm. EPS may also
contribute to the antimicrobial resistance properties of biofilm by impeding the
mass transport of antibiotics through the biofilm, probably by binding directly to
these agents (Gotz, 2002). Structure may also be influenced by the interaction of
particles of nonmicrobial components from the host or environment. For
example, erythrocytes and fibrin may accumulate as the biofilm forms. S. aureus
biofilm is a complex composition consisting of PIA (Polysaccharide
intercellular adhesion), proteinaceous factors and eDNA (extracellular DNA).
4.2.2 Biofilm Formation
Biofilm formation is not restricted to certain group of microorganisms.
The various microbes involved in biofilm formation are Arcanobacterium
(Actinomyces) pyogenes, S. aureus, S. hyicus, Str. agalactiae, Corynebacterium
renale, or Corynebacterium pseudotuberculosis (Melchior et al., 2009; Quoc et
al., 2006; Yazdani et al., 2006).
Biofilm grows in a three stage process as shown in Fig 4.2. The initial
stage includes the attachment of bacteria to the substratum facilitated by cell
wall associated adhesions followed by surface motility and binary division result
in an aggregation of attached cells (Stoodley et al., 2010).

69
The second stage is characterized by cell multiplication and formation of
a mature structure consisting of many layers of cells, connected to each by
extracellular polysaccharides. Bacterial growth and division then leads to the
colonization of the surrounding area and the formation of the biofilm.
Fig 4.2: Stages of biofilm formation
The third stage involves maturation of biofilm, where many staphylococci
generate a glycocalyx, a slime layer that further protects the biofilm bacteria.
Mature biofilm, as shown in Figure 1, is a complex heterogeneous structure of
dormant and actively growing bacteria colonies along with further enzymes,
excretory products and small channels forming part of the overall structure.
When the biofilm reaches a critical mass, a dynamic equilibrium is reached at
which the outermost cell layer begins to generate planktonic organisms. These
bacteria are free to escape the biofilm and to colonize other surfaces (Saldarriaga
Fernandez et al., 2010).

70
4.2.3 Molecular level / QS system
To form an integral part of biofilm, a bacterial cell needs to communicate
with the population and generate a proper cell density for attachment. This is
achieved by means of extra cellular signal molecules which are already present
in the environment and act as auto inducers to start genetic programs (Camilli
and Bassler, 2006). This intercellular communication principle is used by many
bacterial species to monitor cell density, hence known as Quorum-sensing (QS),
and allows individual cells to behave as a community (Schauder and Bassler,
2001). Thus, quorum-sensing is important both to start biofilm formation and to
maintain the biofilm.
Quorum-sensing is regarded as key mechanism in biofilm development
and auto inducers involved offered an attractive new target for non-antibiotic
control of biofilm infections (Kjelleberg and Molin, 2002). Autoinducers action
in different bacterial species and the complex interaction between autoinducers
of different species demonstrated that the goal of controlling biofilm infections
is not easily achievable.
4.2.4 Genetics of biofilm formation
The formation and development of biofilm occurs in a sequential manner
certain genes coding for EPS needs to be turned on and also certain enzymes and
proteins are involved in each stage of biofilm development. The composition of
exopolysaccharide or extra cellular polymeric substance (EPS) is complex
containing mainly of cellulose and β-1, 6-linked N- acetylglucosamine. Their
synthesis is highly regulated by secondary signal of c-di-GMP whose
concentration in turn depends on the activity of proteins containing a
GGDEF/EAL domain. This protein is observed to be a common factor in
biofilm development (Lasa, 2006).
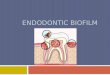
71
Fig 4.3: Scheme of typical biofilm formation steps (A). General
mechanism of the biofilm control (B). Biofilm control molecules action
(Milner et al., 1997)
The first stage in biofilm development is attachment of planktonic cells on
to a surface involving certain surface proteins; the first member to be identified
was Bap (biofilm Associated Protein), from S. aureus in bovine mastitis. The
Bap gene encoding the surface protein was carried in a putative composite
transposon inserted in a mobile Staphylococcal pathogenecity island and was
found only in bovine mastitis isolates. It was apparently present in only 5% of

72
350 bovine mastitis and absent in all human clinical S. aureus isolates tested so
far. It was also noticed that Bap positive isolates showed high resistance to
antibiotic treatment.
Studies on S. aureus biofilm development suggested that clinical S.
aureus isolates are not dependent only on PIA but also on non-polysaccharide
component of intracellular matrix. Certain protein mediated biofilm formation
has emerged as an alternative to PIA and many surface adhesions, such as Bap
(Cucarella et al., 2004a), Spa, FnBPA, FnBPB, and SasG. It was also seen that
polysaccharide degrading enzymes, such as dispersin B and mutation in the ica
gene locus that generates PIA are least effective on PIA-independent strains
(Ziebuhr et al., 2000).
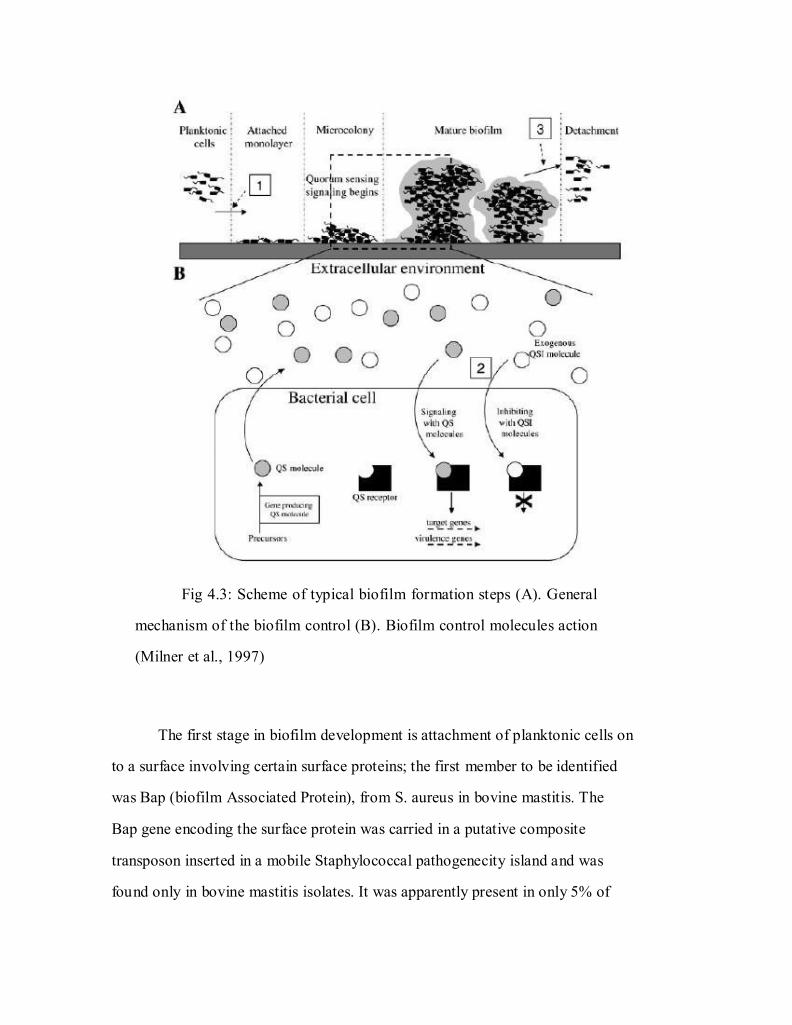
The ica ADBC operon which encodes PIA was first identified in S.
epidermidis and is also present in S. aureus and other Staphylococcal species
(Fitzpatrick et al., 2005; Lasa, 2006; Yazdani et al., 2006). A linear homoglycan
of β-1, 6-linked N-acetylglucosamine named PIA has been purified and
characterized as the main exopolysaccharide compound of S. epidermidis
biofilm matrix. PIA related exopolysaccharide PNAG, which resembles in
structure with that of PIA was described in S. aureus. Moreover, it has been
shown that PIA and PNAG are structurally and immunologically identical and
that both are synthesized by the action of four homologous proteins (IcaA, IcaD,
IcaB, IcaC) encoded by the genes organized in a single operon (icaADBC)
(Cucarella et al., 2004b; O'Gara, 2007; O'Neill et al., 2007; Tormo et al., 2005).
An overview of PIA biosynthesis by the ica operon is given in Fig: 4.4 a & b.
73
Fig 4.4a: Representation of the ica Operon
Fig 4.4b: Overview of PIA biosynthesis by ica operon
4.2.5 Identification tests
4.2.5.1 Tissue culture method
Tissue culture method was discovered by Christensen et.al in 1985. Since
this method has been most widely used and considered as standard test for
biofilm formation. The organisms grown in trypticase soy broth with 1%
glucose were tested for biofilm expression. The biofilm attached to the bottom
of the plate were fixed 2% sodium acetate and stained with 1% crystal violet.
Optical densities of the stained biofilm were then obtained at 570nm using
ELISA autoreader.

74
4.2.5.2 Tube method
Tube method was discovered in the year 1982 (Goldstein and Roberts,
1982). In this method, ability of S. aureus adherence to the walls of borosilicate
glass tube is noted. This method is considered to be the most primitive and first
method to check for biofilm forming strains of S. aureus and practiced till now
in diagnostic laboratory. The tubes are dried and stained with 0.1% crystal
violet. Visible stained film was seen on the tube wall and also on the bottom of
the tube. Ring formation observed at the broth interface is not considered as
positive result for biofilm formation.
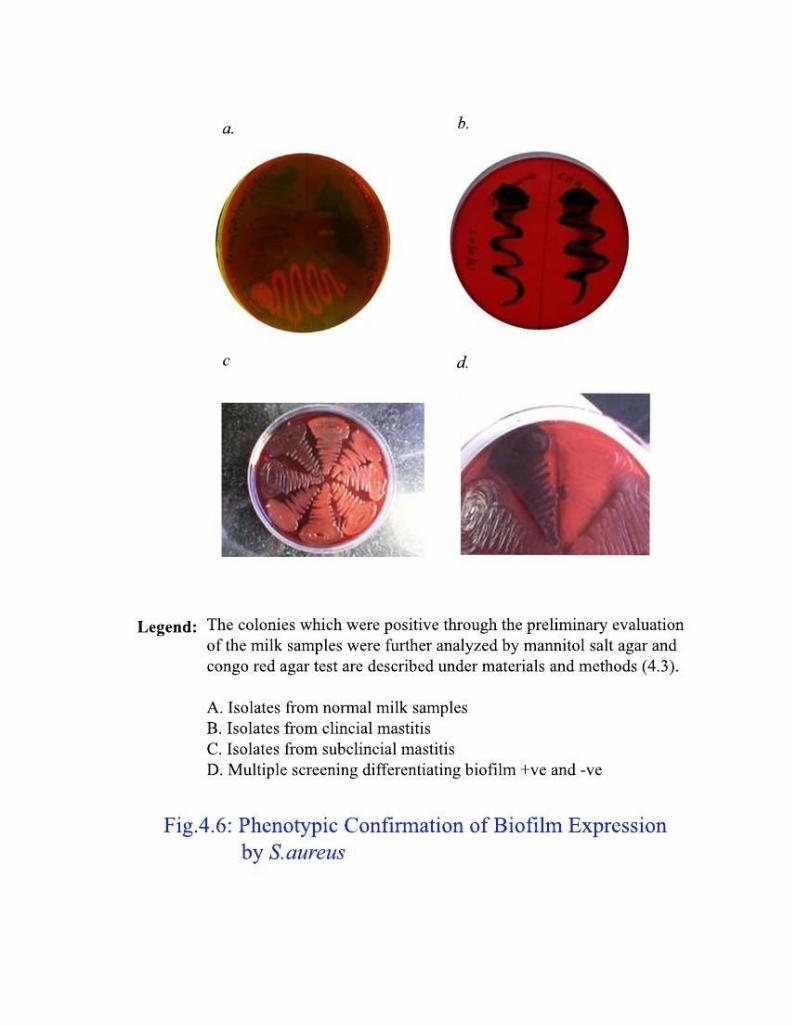
4.2.5.3 Congo Red Agar test
Phenotyping the pathogenic and biofilm producing S. aureus can be
performed by Congo Red Agar test. The slime producing phenotype appears as
black colonies and can be differentiated from other non slime producers
visually. This is an easy and cost effective method and can be performed
routinely in microbiological laboratory to identify biofilm producing pathogenic S. aureus.
4.2.6 Molecular Screening
Gene level identification of biofilm forming strains of Staphylococcus
aureus is the most confirmatory test and relevant, unlike qualitative tests like
Congo red agar method which at times shows false positive results. Candidate
genes like icaD and icaA representing ica operon encoding the biofilm pathway
enzymes can be targeted in molecular screening. By quantitative PCR, the
expression level could be measured and the effect of the drug could be
evaluated. This is important since, the biofilm level is increased when the
organism is under stress like exposure to antibiotic.

75
4.2.7 Eradication / elimination
Elimination of bovine mastitis is very problematic, firstly, as it is an
endemic disease i.e. a disease that is constantly present to a greater or lesser
degree in a population of a certain class or in population living in a particular
location. And secondly, apart from this there are no tests available which can
detect the mastitis disease at the early stage in single step. Visualization of the
udder and milk is not the confirmatory test for Mastitis. Depending on timing of
infection, eradication of S. aureus infection can be achieved. But during
lactation the antibiotic treatment is avoided and many of the S. aureus affected
cows become chronic and have to be culled. Preventive measures can be
adopted for the contagious and environmental mastitis.
4.2.7.1 Resistance to antimicrobials and predator living cells
Biofilms are more resistant to antibiotics compared to planktonic bacterial
cells. The cause of the resistance is not yet completely understood but several
hypotheses were proposed. One of the earliest hypotheses was about the
inability of the antibiotics to penetrate the biofilm matrix, which may contribute
to antibiotic resistance. Other hypotheses suggested that antimicrobials are
degraded in the biofilm before showing its effect, like, penicillin being degraded
by penicillinase enzyme.
Apart from biofilm being resistant to synthetic chemical or antimicrobial
compounds, they are also found to be resistant to protozoa or phagocytic blood
cells (Paape et al., 2000). The Fig 4.5. shows neutrophil attacking the biofilm
mass by grazing through the surface, where the cells located deep inside the
biofilm remain unaffected and replace the lost cells thus, maintaining the
integrity of the biofilm.

76
Fig 4.5: Neutrophils on surface of biofilm.
4.2.7.2 Antibiotics and biofilm: A strategy to overcome difficulties
For the eradication of biofilm formed by various microbes, a wide range
of antibiotics are used. But it is observed that plank tonic cells showed
sensitivity than biofilm, thus it necessitated to develop drugs that targeted that
quorum sensing system and disrupted the formation and activation of auto
inducers. Olsen et.al used 12 different antibiotics to check their effect on biofilm
composed of Arcanobacterium (Actinomyces) pyogenes, S. aureus, S. hyicus,
Str. agalactiae, Corynebacterium renale, or Corynebacterium
pseudotuberculosis microbes.
4.2.7.3 Combo drug
Combo drug denotes different combination of antibiotics used to eradicate
biofilm formed during Mastitis. This mechanism is possible due to synergy
between the antibacterial agents. Scientists observed that combination dosage of
farnesol and gentamicin treated on static biofilm depleted bacterial population
by more than 2 log units

77
4.3 Materials and Methods
4.3.1 Materials
The Staphylococcus culture (Staphylococcus aureus-MTCC3160)
obtained from Microbial Type Culture Collection and Gene Bank, Institute of
Microbial Technology, Chandigarh was used as positive control for the analysis
of biofilm expression.
4.3.1.1 Congo Red Agar
Composition
Congo Red
Muller
Agar
Sucrose
Agar
gm/l
0.8
Hinton 21
36
15
The final volume was made up to 1l and the pH was adjusted to 7.4 ± 0.2 at
250C. Sterilization was carried out by autoclaving at 1210C for 15 min.
4.3.1.2 Reverse Transcriptase - Polymerase Chain reaction
Random primer 1.25mM/µl,
dNTPs 2.5mM
MMLV Reverse transcriptase 40units/µl
10 picomoles of forward and reverse primers,
1.25 units of Taq

78
4.3.2 Methods
The biofilm phenotypic expression of S. aureus isolates were evaluated by
Congo red agar and quantified. Total RNA was extracted from the bacterial
cultures and subjected to reverse transcriptase PCR using icaD specific primer
representing the biofilm synthesis by the ica operon.
4.3.2.1 Congo Red Agar
The mannitol salt agar positive colonies screened for biofilm producing
capacity by Congo Red Agar. Congo red agar was prepared by adding the dye to
Muller Hinton agar. The test organisms were streaked onto Congo red agar
plates and incubated at 370C for 24 h. The biofilm producing colonies exhibited black colonies while those failed were of yellow color colonies.
4.3.2.2 Quantification of Biofilm
S. aureus culture was initiated with 50µl of 104CFU cells in a 96-well
polystyrene plate containing 150µl of Congo red broth and incubated for 24h at
370C without shaking. After 24h, the plates were washed vigorously three times with PBS and dried for 1h at 56˚C prior to staining with a 0.4% crystal Violet
solution. The absorbance of the adhered, stained cells producing biofilm was
measured at 492nm using Multiskan plate reader (Thermo systems, USA).
4.3.2.3 RNA isolation
Total RNA isolation was performed by TRIZOL method with slight
modifications. The frozen bacterial cells were thawed to room temperature and
centrifuged at 10000X for 10min at 4˚C. The supernatant was discarded and the
cell pellet was resuspended in 150µl of trizol. The cell was homogenized using
Tissue Tearor (Biospec Products INC, USA) at 30,000rpm for 30 sec and
incubated at room temperature for 5 min. To this, 100µl of Chloroform was

79
added and centrifuged at 13,000X for 20 min. The upper aqueous phase was
transferred to a fresh 1.5ml micro centrifuge tube and added equal amount of
isopropanal. Incubated at -200C for one hour, then centrifuged at 13,000X for 15
min at 40C. The supernatant was discarded and the pellet was washed with 500µl
of 75% ethanol by centrifugation at 10,000X for 5min at 40C. The supernatant
was discarded and the pellet was dissolved in 20µl of double autoclaved water.
The quality and quantity of the isolated RNA was checked by 2% agarose gel
electrophoresis.
DNA contamination if any seen with RNA preparations was removed by
the DNase treatment. The reaction volume was set up to 20µl containing 9U of
DNase. It was incubated at 370C for 30-45 min then 20mM of 2µl EGTA was
added and further incubated at 650C for 10min. Sodium acetate (1/10 V) and
absolute ethanol (2V) was added and incubated at -20˚C for 1 h. Then it was
centrifuged at 12000xg for 20 min at 40C, the supernatant was discarded and the pellet was washed with 500µl of 75% ethanol. Air-dried pellet was dissolved in
20µl of DEPC treated water and used for RT-PCR.
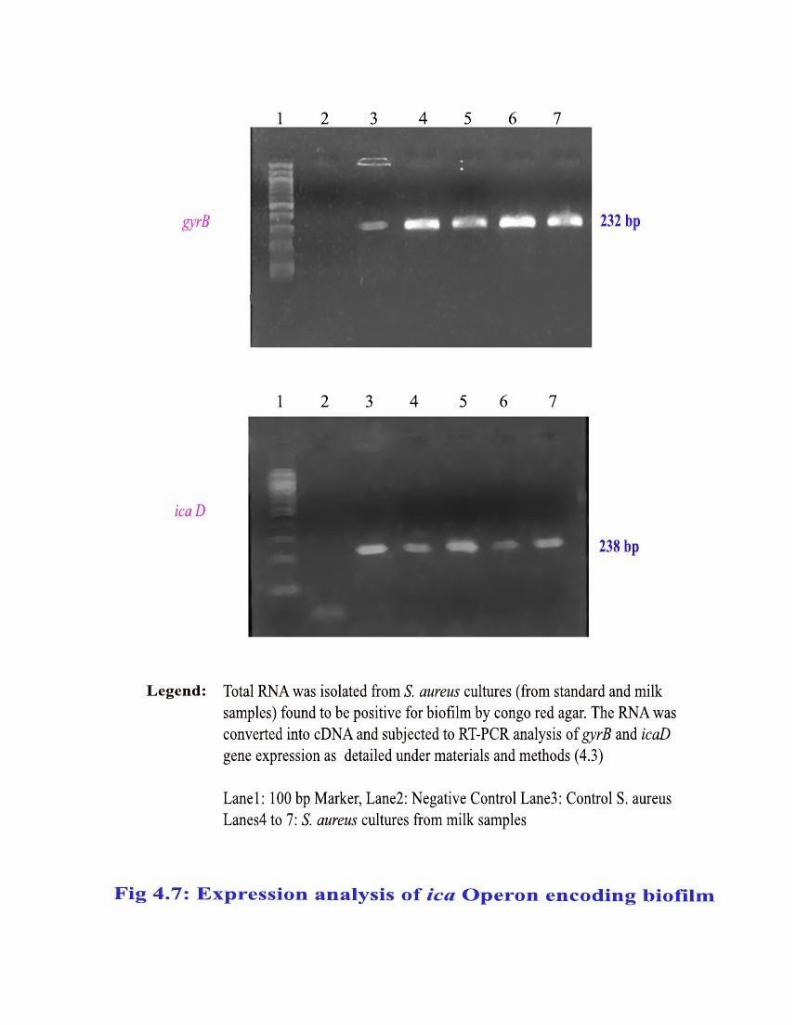
4.3.2.4 Expression analysis of ica Operon
The biofilm expression by S. aureus cultures was analyzed by amplifying
the ica D mRNA representing the ica Operon gyrB as housekeeping gene. The
primers for icaD and gyrB genes of S. aureus were designed based on the
available sequence (Gene Bank Accession no: NC_002592) using Genetool lite
software (Gene twist, USA). Total RNA was converted into cDNA using a
reaction mixture containing MMLV Reverse Transcriptase, RNase Inhibitor,
Random hexamer as primers, dNTPs and 1X Buffer. The cDNA synthesis was
carried out at 420C for 60 min and extension at 750C for 10 min. The prepared cDNA was used as template to amplify gyrB and ica D genes. The primers

80
gyrBF and gyrBR amplified a 232 bp of gyrB gene while icaDF and icaDR
primers amplified a 238 bp product of icaD gene. The temperature profile of
icaD gene was: initial denaturation for one cycle at 940C for 2 min, followed by
30 cycles of 940C for 15 sec, the annealing at 52.5ºC for 15 sec, extension at
720C for 30 sec and a final extension for one cycle at 720C for 1 minute. The
same temperature profile was used for gyrB gene except that the annealing
temperature was at 630C. The amplicons were purified with a gel extraction kit (GF-1 GEL DNA recovery Kit, Vivantis, Singapore) and subjected to
sequencing in both directions using big dye terminator-sequencing kit (Applied
Biosystems, USA).

81
4.0 Results
The S. aureus isolates biochemically confirmed were further evaluated for
their biofilm expression. The phenotypic confirmation was obtained by congo
red agar and at molecular level, the ica operon encoding the genes responsible
for the biofilm synthesis was screened.
4.4.1 Congo Red Agar
In this selective medium, congo red in muller hinton agar interacts with
biofilm and generates black colonies. The S. aureus isolates that are negative for
biofilm expression exhibit pale yellow colonies. Out of 180 confirmed S. aureus
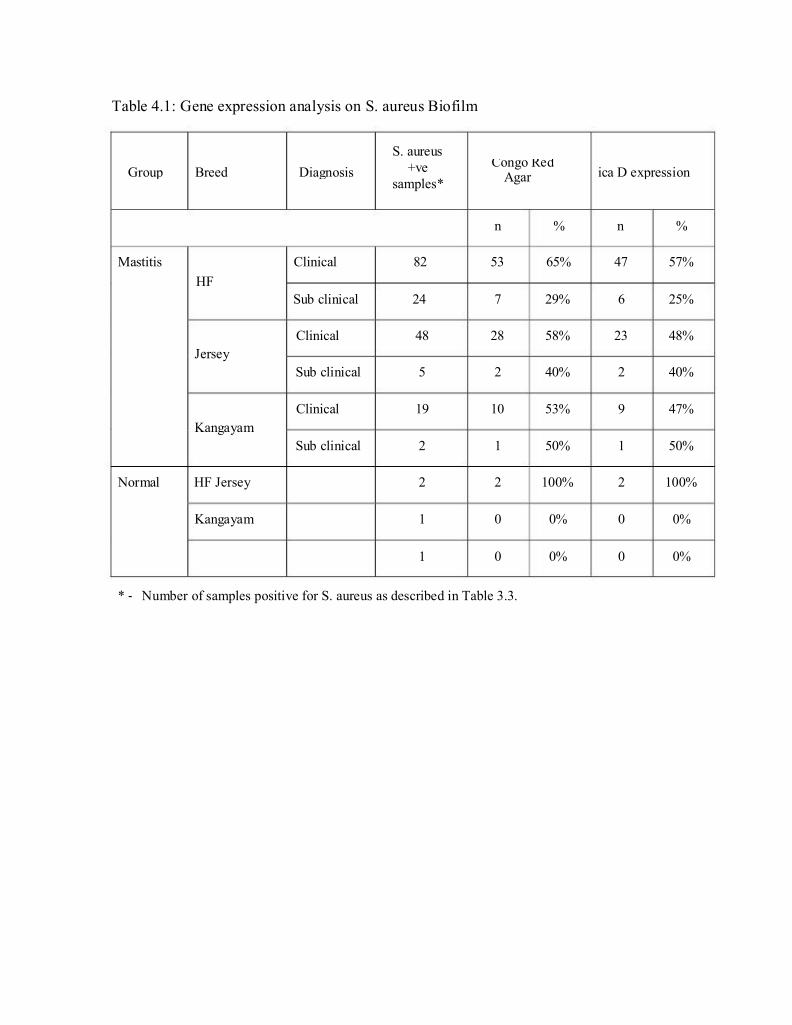
isolates 101 (56%) were found to express biofilm (Table 4.1 and Fig. 4.6). This
constitutes 31.5% of the total mastitis samples. Among the very few S. aureus
isolates obtained from normal milk samples, the two isolates from HF breed
were found to have biofilm expression. Others were negative for biofilm
synthesis.
4.4.2 Expression analysis of ica Operon encoding biofilm
The enzymes in major synthetic pathway for biofilm are encoded by ica
iperon which is comprised of four genes namely ica A, D, B & C. The ica D was
considered as representative gene for the ica operon and analyzed by RT-PCR
among the S. aureus isolates. The expression of icaD was compared with that of
the housekeeping gene gyrB as internal control. Out of 101 isolates confirmed
by congo red agar, 88 (87%) were found to express icaD mRNA (Table 4.1 &
Fig.4.7). This is about 49% (88/180) of the S. aureus isolates and 27.5% of the
study sample size of 320 milk specimens. The S. aureus isolates obtained from
the normal HF milk samples were positive for icaD expression.
82
Group Breed Diagnosis +ve Congo Red ica D expression
Table 4.1: Gene expression analysis on S. aureus Biofilm
S. aureus
samples*
Agar
n % n %
Mastitis
HF
Clinical 82 53 65% 47 57%
Sub clinical 24 7 29% 6 25%
Jersey
Kangayam
Normal HF Jersey
Kangayam
Clinical 48
Sub clinical 5
Clinical 19
Sub clinical 2
2
1
1
28 58%
2 40%
10 53%
1 50%
2 100%
0 0%
0 0%
23 48%
2 40%
9 47%
1 50%
2 100%
0 0%
0 0%
* ‐ Number of samples positive for S. aureus as described in Table 3.3.

83
4.5 Discussion
Many common bacterial pathogens exist in animals as biofilm and their
infections are generally chronic and often difficult to treat. Biofilm associated
microorganisms behave differently from planktonic organisms with respect to
growth rates and ability to resist antimicrobial treatments and therefore pose a
major health problem. S. aureus with phenotypic expression of biofilm is known
for its disease severity with increased resistance to the antibiotics and blocking
the lumen of the duct. The synthesis of biofilm is by two different pathways, ica
operon dependent and independent (Gotz, 2002).
Earlier studies have reported that 20-30% of mastitis is associated with S.
aureus infection. In this study 320 milk samples from clinical and subclinical
mastitis cows were screened for S. aureus infection. The conventional
microbiological and biochemical methods employed confirmed 56% of them
positive for S. aureus. The selection medium, congo red agar identified 31.5% of
the S. aureus isolates exhibiting biofilm. The molecular screening recognized
27.5% of the samples had biofilm expressed by functional ica operon. The
incidence recorded by this study is in accordance with the published reports.
The biofilm produced by S. aureus confers antibiotic resistance, blocking
the secretion duct and the inflammation, emphasizing the fact that preventing the
biofilm synthesis is the primary requirement in controlling S. aureus infection
associated mastitis. Since, synthesis of biofilm depends on two different
pathways, it is important to consider the genes associated with the pathways. On
evaluating the role of the candidate genes belonging to the pathway in a given
geographical location, drugs could be developed/identified and targeted.

84
Bacteriological cure rate for the treatment with antimicrobial agents is
ranging between 0% and 80% as indicated by earlier epidemiological studies.
Bacterial species with biofilm synthesis exhibit an innate resistance to
antibiotics, disinfectants and clearance by host defense mechanisms. Despite its
higher incidence, expression of the ica locus and biofilm formation, seems to be
highly variable among staphylococci isolates. Sub-inhibitory concentrations of
tetracyclines and quinuprestin-dalfopristin, as well as high temperature and
osmolarity increase the ica promotor activity. In contrast, penicillin, oxacillin,
chloramphenicol, clindamycin, gentamicin, ofloxacin, vancomycin and
teicoplanin seem to have no effect (Rachid et al., 2000b; Ziebuhr et al., 2000).
Comparatively, bacteria growing in a biofilm can become 10–1000 times
more resistant to the effect of antimicrobial agents than planktonic growing
bacteria of the same strain (Amorenaa et al., 1999; O’Toole, 2003). Several
mechanisms are known to be responsible for resistance of biofilms to
antimicrobial agents, including delayed penetration of the antimicrobial agents
through the biofilm matrix, altered growth rate of biofilm organisms and
physiological changes due to the biofilm mode of growth.
The relapsing nature of biofilm infections can be extrapolated from the
dynamic features of biofilm formation and shedding of cells from one biofilm to
form a new biofilm. Added to this there is a high prevalence of the ica genes
among S. aureus mastitis isolates. This emphasizes the need for extending
antibiotic therapy to interrupt the dynamics of biofilm formation. antibiotics had
a greater effect on young biofilms and those grown in milk, than on older
biofilms and those grown in broth (Amorenaa et al., 1999). These experiments
did show also that gentamicin and erythromycin were the least effective

85
antibiotics against S. aureus biofilms. Mastitis pathogens are able to form
biofilms, we must accept that current NCCLS guidelines on testing sensitivity of
pathogens to antibiotics have a poor predictive value, and that current
therapeutic regimes might not be appropriate.