Embed Size (px)
Citation preview

Biochimica et Biophysica Acta 1780 (2008) 948–959
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbagen
Membrane interaction and perturbation mechanisms induced by twocationic cell penetrating peptides with distinct charge distribution
Isabel D. Alves a,b,⁎, Nicole Goasdoué a,b, Isabelle Correia a,b, Soline Aubry a,b, Cécile Galanth c,d,Sandrine Sagan a,b, Solange Lavielle a,b, Gérard Chassaing a,b
a UPMC Univ Paris 06, UMR 7613 Synthèse, Structure et Fonction de Molécules Bioactives, FR 2769, Paris F-75005, Franceb CNRS, UMR 7613, Paris F-75005, Francec UPMC Univ Paris 06, FRE 2852 Peptidome de la Peau d'Amphibiens, Paris F-75005, Franced CNRS, FRE2852, Paris F-75005, France
⁎ Corresponding author. UPMC, UMR 7613, case courParis, France. Fax: +33 1 44277150.
E-mail address: [email protected] (I.D. Alves).
0304-4165/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.bbagen.2008.04.004
a b s t r a c t
a r t i c l e i n f oArticle history:
Independently from the cell Received 26 October 2007Received in revised form 23 April 2008Accepted 24 April 2008Available online 2 May 2008Keywords:Cell penetrating peptideAmphipathic peptidePeptide-membrane interactionDifferential scanning calorimetryCircular dichroism31P NMR
penetrating peptide uptake mechanism (endocytic or not), the interaction of thepeptide with the lipid bilayer remains a common issue that needs further investigation. The cell penetratingor antimicrobial properties of exogenous peptides require probably different preliminary interactions withthe plasma membrane. Herein, we have employed 31P NMR, differential scanning calorimetry and CD tostudy the membrane interaction and perturbation mechanisms of two basic peptides with similar length butdistinct charge distribution, penetratin (non-amphipathic) and RL16, a secondary amphipathic peptide. Thepeptide effects on the thermotropic phase behavior of large multilamellar vesicles of dimyristoylpho-sphatidylcholine (DMPC), dimyristoylphosphatidylglycerol (DMPG) and dipalmitoleoyl phosphatidylethano-lamine (DiPoPE) were investigated. We have found that, even though both peptides are cationic, theirinteraction with zwitterionic versus anionic lipids is markedly distinct. Penetratin greatly affects thetemperature, enthalpy and cooperativity of DMPG main phase transition but does not affect those of DMPCwhile RL16 presents opposite effects. Additionally, it was found that penetratin induces a negative curvaturewhereas RL16 induces a positive one, since a decrease in the fluid lamellar to inverted hexagonal phasetransition temperature of DiPoPE (TH) was observed for penetratin and an increase for RL16. Contrary topenetratin, 31P NMR of samples containing DMPC MLVs and RL16 shows an isotropic signal indicative of theformation of small vesicles, concomitant with a great decrease in sample turbidity both below and at thephase transition temperature. Opposite effects were also observed on DMPG where both peptides provokestrong aggregation and precipitation. Both CPPs adopt helical structures when contacting with anionic lipids,and possess a dual behavior by either presenting their cationic or hydrophobic domains towards thephospholipid face, depending on the lipid nature (anionic vs zwitterionic, respectively). Surprisingly, theincrease of electrostatic interactions at the water membrane interface prevents the insertion of RL16hydrophobic region in the bilayer, but is essential for the interaction of penetratin. Modulation ofamphipathic profiles and charge distribution of CPPs can alter the balance of hydrophobic and electrostaticmembrane interaction leading to translocation or and membrane permeabilisation. Penetratin has a relativepure CPP behavior whereas RL16 presents mixed CPP/AMP properties. A better understanding of thoseprocesses is essential to unveil their cell translocation mechanism.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
In the past decade, cell penetrating peptides (CPPs), also calledprotein transduction domains (PTDs), have drawn major attention fortheir capability to translocate the cellular membranes and transportcargos such as peptides, proteins, oligonucleotides and nanoparticles,opening new possibilities in biomedical research and therapy [1].
ier 182, 4 place Jussieu, 75005
l rights reserved.
Additionally, some of them have been shown to display antimicrobialactivity, becoming good candidates as antibiotic substitutes. Despitethe widespread interest in such molecular carriers, the mechanismsunderlying the cellular translocation of CPP are yet incompletelyunderstood and have been in the center of controversial discussion.The common characteristic of CPPs is the high density of basic aminoacid residues (Arg and Lys). The large charge at physiological pHexcludes the passive diffusion of CPPs across the lipid bilayer (except ifthe lipids play the role of a phase transfer agent [2,3]). The absenceof chiral recognition precludes classical uptake mechanisms by areceptor or a transporter [4] and the classical endocytosis pathway is

949I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
controversial. Nonetheless, it has been evidenced by several labora-tories that interactions between CPPs and membrane lipids are likelyto play a major role in the translocation processes, even if the lipidsalone cannot explain the different cellular uptake characteristicsexhibited by these peptides [5]. Moreover, it should bementioned thatthe internalization of such peptides by endocytic pathways has notbeen discarded, a survey of the recent literature supports the con-clusion that a CPPmay enter by several mechanisms (endocytic or not)depending on their nature, the cargo transported, cell type and localenvironment such as lipid composition, etc.
Herein, we have chosen to investigate the molecular mechanismsinvolved in the interactions of twocationic peptideswith the lipidbilayer,penetratin (a non-amphipathic peptide, H-RQIKIWFQNRRMKWKK-NH2)and RL16 (H-RRLRRLLRRLLRRLRR-OH), a peptide designed from struc-ture/uptake relationships of penetratin [6]. Both peptides have beenproposed as vectors to address bioactive molecules [7]. RL16 possess anN-terminal/C-terminal pseudo-symmetrical sequence (RRLRRLLR) and asecondaryamphipathic character,whichmay inducepore formation [8].Penetratin, denoted pAntp, has a sequence corresponding to the 16residues of the third α-helix (residues 43–58) of the Antennapediahomeodomain protein of Drosophila [4,9]. This peptide has attractedwidespread interest because of its ability to directly target attachedoligopeptides and oligonucleotides to the cytoplasm and nuclearcompartments of cells [10]. The internalization of penetratin into cellsoccurs both at 4 and 37 °C, is receptor independent, highly efficient andis non-cell-type specific. Penetratin does not belong to the amphipathichelical peptide family according to its weak penetration into modelmembranes [11] even if a partial amphipathic character has beenattributed. It was suggested that penetratin translocates through thelipid bilayer by transiently forming invertedmicelles [6,10]. Endocytosis,macropinocytosis [12] and electroporation-like permeabilizationmechanism [13] may also participate in the uptake of penetratin.
Furthermore, it has been shown that some CPPs includingpenetratin present antimicrobial activities in bacteria [14] as naturalantimicrobial peptides (AMP). Regarding RL16, apart from the fact thatit permeabilizes eukaryotic cells and induces calcein release fromLUVs[8], its membrane interaction has not been analyzed.
We hypothesized that peptide translocation (CPP) and/ormembranepermeabilization (AMP) depend on both the peptide structures and thelipid composition of the phospholipid matrix. By investigating theirinteraction mode with lipids, important information can be obtained tohelp decipher their translocation/permeabilization mechanisms.
Herein, eukaryotic cell membrane perturbation upon incubationwith these peptides was monitored to address the possibility of poreformation and the antimicrobial activity against different bacterialstrains was assayed. The interaction of these peptides with zwitterionicand anionic large multilamellar vesicles (MLVs) was examined bymonitoring the consequences on the lipid phase transition temperatureby differential scanning calorimetry (DSC). Like in the vast majority ofDSC studies, we have chosen dimyristoylphosphatidylcholine (DMPC)because the lipid is stable tooxidation, readily hydrated inwater forminglamellar phases at physiological temperature and exhibits a single andhighly cooperative gel to liquid-crystalline phase transition at about23 °C, a temperature well above the freezing point of water but belowthe thermal denaturation temperature of peptides [15]. From thebiological point of view, since PCs are virtually absent in bacterialmembranes but are generally the most abundant phospholipids ineukaryotic plasma membranes, DMPC would be a good model for thesurface membrane of mammalian cells; even more because PCs aretypically found primarily in the outer monolayer of the lipid bilayer ofsuch membranes. Similarly, PGs are absent in eukaryotic plasmamembranes but are ubiquitous and often abundant in bacterialmembranes, so dimyristoylphosphatidylglycerol (DMPG) can serve asa goodmodel for the bacterialmembrane. Additionally, since the studiedpeptides are highly charged, (7 positive charges for penetratin and 11positive charges for RL16 at physiological pH, electrostatics may play a
role in their interactionwith the lipid bilayer and preferred orientation,at least for the initial contact. In order to get some insight into the role ofthe peptide on the lipid curvature,we havemonitored the peptide effecton lamellar (Lα) to hexagonal phase (HII) transition temperature (TH) ofdipalmitoleoyl phosphatidylethanolamine (DiPoPE). The presence ofnonlamellar-preferring lipids in membranes induces a curvature strain.When constrained to pack into planar bilayers, certain phospholipids(such as the PE) will form unstable structures as a consequence of theirmolecular shape and noncovalent bonding. This produces curvaturestrain, which may provide energy for subsequent membrane processes[16–18]. Additionally PE, along with PG are the major phospholipidcomponents in bacterial plasma membrane.
31P NMR has been widely used to characterize lipids and theirpeptide-induced changes in supramolecular organization. The even-tual formation of different lipid phases such as HII, cubic and differentisotropic phases has been monitored as a function of lipid and tem-perature. Additionally, due to the potential effects of these peptides onvesicle aggregation and bursting, which may eventually act as deter-gents, turbidity measurements were performed. Moreover, the sec-ondary structure of the peptides was investigated by CD in the absenceand presence of lipid and as a function of the P/L ratio. The biophysicaldata reported hereinwas used to delineate themechanism of action, interms of bilayer interaction, of these peptides.
2. Materials and methods
2.1. Materials
DMPC and DMPG were purchased from Genzyme (Switzerland) and were usedwithout further purification. DiPoPE was obtained from Avanti Polar Lipids (Alabaster,AL). The synthesis and purification of penetratin and RL16 was performed using Boc-solid phase strategy.
2.2. Cell culture
Chinese hamster ovary (CHO) K1 cells were cultured in Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 10% fetal calf serum (FCS), penicillin (100000 IU/L),streptomycin (100000 IU/L), and amphotericin B (1 mg/L) in a humidified atmospherecontaining 5% CO2 at 37 °C.
2.3. LDH leakage
Membrane integrity was measured using the Promega CytoTox-ONE™ assay. Inbrief, CHO cells were seeded in 96-well plates 1 day before treatment with serum-freeDMEM for 30 min. Untreated cells were defined as zero and lactate dehydrogenase(LDH) released by lysating in 0.2% Triton X-100 in the supplied buffer defined as 100%leakage. Melittin, a peptide known to disturb the bilayer integrity, was employed as anadditional positive control. The cells were incubated with 1, 10, 50 and 100 μM of thepeptide for periods of time ranging from 1–6 h.
2.4. Antimicrobial activity
Gram-positive eubacteria (Bacillus Megaterium and Staphylococcus aureus) andGram-negative eubacteria (Escherichia coli 363 and Klebsiella pneumoniae) werecultured as described previously [19]. The minimal inhibitory concentrations (MICs)of peptides were determined in 96-well microtitration plates by growing the bacteria inthe presence of 2-fold serial dilutions of peptide. Aliquots (10 μL) of each serial dilutionwere incubated for 16 h at 37 °C with 100 μL of a suspension of a midlogarithmic phaseculture of bacteria at a starting absorbance A630=0.01 in Poor–Broth nutrient medium(1% bactotryptone and 0.5% NaCl, w/v). Inhibition of growth was assayed by measuringthe absorbance at 630 nm. The MIC was defined as the lowest concentration of peptidethat inhibited the growth of ≥99% of the cells. Bacteria that was incubated with thepeptide corresponding to the MIC was plated out on solid culture medium containing1% noble agar to distinguish between lytic and non-lytic effects, the peptide being lyticif after overnight incubation with the peptide (at the MIC concentration) the bacteriadevelopment was inhibited and non-lytic when the bacteria was able to re-grow uponpeptide incubation. All assays were performed in triplicate plus positive controlswithout the peptide and negative controls with 0.7% formaldehyde.
2.5. Hemolytic activity
Red blood cells (RBC) were isolated from human plasma and washed three timeswith PBS (10mMphosphate buffer,140mMNaCl, 3mMKCl, pH 7.4). RBCs (108 cells/mL)in PBS were incubated with peptide concentrations ranging from 1 to 200 µM for30 min at 37 °C for 60 min. After centrifugation (15 min, 900 g, 4 °C), haemolysis wasmeasured by monitoring the optical density of the supernatant at 405 nm andcomparing with supernatants of lysed RBCs after addition of 1% (v/v) Triton X-100.

Table 1Mammalian cell membrane integrity perturbation. The results are presented aspercentages relative to a positive control (Triton X-100 at 0.18%), which represents100% membrane integrity perturbation
Peptide Concentration (µM) Incubation time (h)
1 2 3 4 5 6
Penetratin 1 0 0 0 0 0 010 0 0 0 0 0 050 0 0 0 0 0 0100 0 0 0 0 0 0
RL16 1 0 0 0 0 0 1010 0 0 2 5 12.5 4050 15 30 50 70 70 70100 40 70 70 70 70 70
Mellitin 1 40 70 90 100 100 10010 60 85 100 100 100 10050 100 100 100 100 100 100100 100 100 100 100 100 100
950 I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
2.6. Preparation of MLVs and LUVs
Lipid films were made by dissolving the appropriate amounts of lipid in a mixtureof chloroform and methanol, 2/1 (v/v), followed by solvent evaporation under nitrogento deposit the lipid as a film on the wall of a test tube. Final traces of solvent wereremoved in a vacuum chamber attached to a liquid nitrogen trap for 3–4 h. Films werehydrated with 10 mM Tris, 0.1 M NaCl, 2 mM EDTA, pH 7.6 (Tris buffer) (for DSCexperiments) or with 10 mM phosphate buffer, pH 7.6 (for CD experiments) andvortexed extensively at a temperature superior to the phase transition temperature ofthe lipid to obtain MLVs. The peptide was either co-dissolved with the lipid or addedafter the formation of the MLVs. To form LUVs, the lipid (MLVs) was subjected to fivefreeze/thawing cycles and the homogeneous lipid suspension passed (19 times)through a mini-extruder equipped with two stacked 0.1 μm polycarbonate filters(Avanti, Alabaster, AL). The size of the LUVs formed was monitored by light scatteringand found to have a diameter of 90±10 nm. Additionally, the UV absorbance wasmeasured for the CD experiments.
2.7. Differential scanning calorimetry
The calorimetry was performed on a high-sensitivity differential scanningcalorimeter (Calorimetry Sciences Corporation). A scan rate of 0.5 °C/min was usedand there was a delay of 10 min between sequential scans in a series that allow forthermal equilibration. Data analysis was performed with the fitting program CPCALCprovided by CSC and plotted with Igor. The total lipid concentrations used were about1 mg/ml, considering full hydration of the phospholipids mixtures except for theexperiments with DiPoPE where 10 mg/ml were used. Samples containing the peptidealone, dissolved in buffer at peptide concentrations corresponding to those at thehigher peptide/lipid molar ratios studied (P/L 1:10), exhibited no thermal events overthe temperature range of 0–100 °C. This indicates that the peptides do not denatureover this temperature range and that the endothermic events observed in this studyarise solely from phase transitions of the phospholipids vesicles. A minimum of at leastthree to four heating and cooling scans were performed for each analysis dependingwhether or not the spectra was reproducible.
2.8. 31P nuclear magnetic resonance (NMR) spectroscopy
Multilamellar lipid samples were prepared using typically 10 mg of lipid, asdescribed previously, in Tris buffer containing 10% D2O. The peptide was either co-dissolved with the lipid before MLV formation or added after liposome formation. TheNMR spectra were collected at several temperatures: below, at and above the phasetransition temperature of the lipid. The 31P-NMR spectra were acquired on a BrukerDRX-500 spectrometer operating at 202.36 MHz for 31P equipped with a 10 mmmultinuclear probe and a Silicon Graphics workstation. Chemical shifts weredetermined relative to external 85% H3PO4. Prior to FT, a 30 Hz exponential linebroadening was applied to the FID. The samples for the NMRwere prepared from 10mgof lipid dispersed in 2.6 ml of H2O. FIDs were acquired over 24.3 kHz with 32 K datapoints and 26000 scans. A pulse width of 12 μs (~60° rotation angle), a relaxation delayof 2 s, and full-power proton decoupling with a RF field of 3125 Hz was employed. Thetemperature was controlled by using a Bruker variable-temperature unit.
2.9. Measurements of vesicle aggregation and dissociation
Changes in aggregation state of vesicles were monitored by absorbance measure-ments at 436 nm. Aliquots of peptide stock solutions were added to 250 μL suspensionsof LUV (lipid concentration, 1250 μM) in 10 mM sodium phosphate buffer and to MLVs(at similar lipid concentration) in 10 mM Tris, 0.1 M NaCl, 2 mM EDTA. The absorbancewas measured using a UVICON 930 spectrophotometer before and after addition ofpeptide. All the measurements were carried out at room temperature.
2.10. Circular dichroism (CD) spectroscopy
The CD spectra of the peptides were recorded with a Jobin Yvon CD6 dichrographlinked to a PC microprocessor. The instrument outputs were calibrated with D(+)-10-camphorsulfonic acid. The spectra were scanned at 25 °C in a quartz optical cellwith a 1 mm path length, unless specified otherwise. Spectra were recorded between185–260 nm with 0.5 nm step. Typically, four or eight scans were accumulated andaveraged after buffer (or LUV) spectra subtraction and baseline correction. The CDspectrum of each peptidewas recorded in 10mMphosphate buffer (pH 7.6), the peptideconcentrationwas varied from 20 to 120 μM. Additionally, the spectrumwas acquired inthe presence of DMPC LUVs with a lipid concentration of 1250 μM. CD measurementsare reported as Δε (M−1 cm−1). The relative helix content was deduced according toZhong and Johnson [20] as the percent of helix=[Δε222 nm°10]/n, where Δε222 nm is thedichroic increment at 222 nm and n is the number of residues in the peptide.
3. Results
3.1. Mammalian cell membrane integrity
In order to test formammalian cellmembrane perturbation and theeventual formation of permanent or transient pores, membrane
leakage of lactate dehydrogenase (LDH) in CHO cells was measuredafter cell exposure to 1,10, 50 and100 μMofpeptide for 1–6h. LDH is anenzyme that exists in the cytosol of eukaryotic cells, its detectionoutside the cells is an index of damaged cell membranes. Penetratinhas no effect on LDH leakage (Table 1). LDH release could be detectedupon RL16 cell incubation over longer periods of time and higherconcentrations than those employed formelittin. Since RL16 is toxic forCHO cells [8], this points to the idea that this peptide may lyse the cellmembrane or lead to pore formation. However, the level of membraneperturbation is nonetheless, much weaker than that observed formelittin either because pores formed are of smaller size (so that LDHrelease is reduced) or of shorter duration. Penetratin as reported inother studies does not affect membrane integrity [8,21].
3.2. Antimicrobial and hemolytic activity
The antibacterial activity of penetratin and RL16 was assayedagainst Gram-negative and Gram-positive bacteria (Table 2). RL16 hasa broad-spectrum of antibacterial activities, inhibiting the growth ofboth Gram-positive and Gram-negative bacteria with minimalinhibitory concentrations in the micromolar range. Bacteria incubatedovernight, with a RL16 concentration corresponding to the MIC,produced no colony-forming units in the case of B. megaterium andE. coli, indicating that RL16 is bactericidal. In the case of S. aureus, thebacteria were able to re-grow after being incubated with RL16 at theMIC concentration, demonstrating the bacteriostatic activity of RL16on this bacterial strain. Penetratin was found to be bactericidal only inthe case of B. megaterium.
As for the hemolytic activity of those peptides, penetratin did notshow any hemolytic activity up to 250 µM. RL16 did show hemolyticactivity, which increased as a function of the concentration used: 0% at7 µM, 3% at 15 µM, 7% at 16 µM, 18% at 30 µM, 28% at 125 µM and 49%at 250 µM.
3.3. Comparing the interaction of the peptides with zwitterionic vsanionic lipids
DSC heating endotherms illustrating the effects of RL16 andpenetratin on the thermotropic phase behavior of large multilamellarvesicles (MLVs) of DMPC are presented in Fig.1. Aqueous dispersions ofDMPC exhibit two endothermic events, a less energetic pre-transitionnear 13 °C that arises from the conversion of the Lβ’ to the Pβ’ and amore energetic main transition from the conversion of Pβ’ to Lα ataround 24 °C. The interaction of RL16 with DMPC clearly alters thethermotropic behavior of the latter, the peptide reduces the phasetransition temperature (Tm), the cooperativity, the enthalpy of the pre-transition (which is abolished at a P/L of 1/25) and of the main
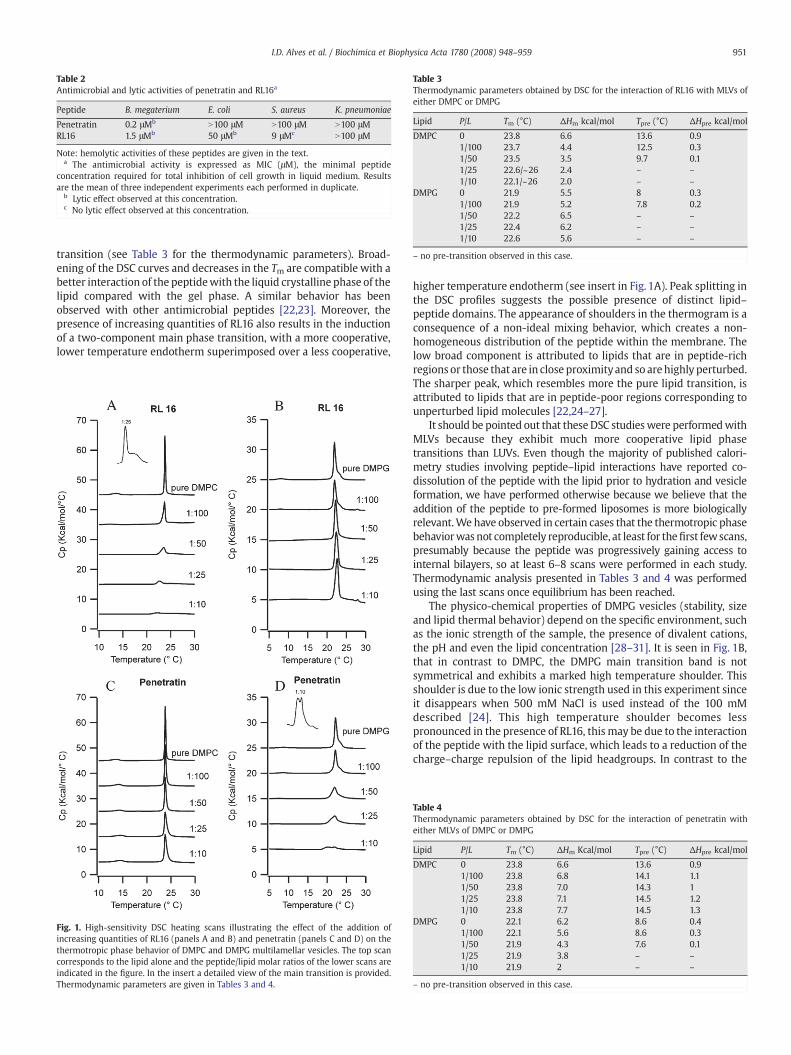
Table 3Thermodynamic parameters obtained by DSC for the interaction of RL16 with MLVs ofeither DMPC or DMPG
Lipid P/L Tm (°C) ΔHm kcal/mol Tpre (°C) ΔHpre kcal/mol
DMPC 0 23.8 6.6 13.6 0.91/100 23.7 4.4 12.5 0.31/50 23.5 3.5 9.7 0.11/25 22.6/~26 2.4 – –
1/10 22.1/~26 2.0 – –
DMPG 0 21.9 5.5 8 0.31/100 21.9 5.2 7.8 0.21/50 22.2 6.5 – –
1/25 22.4 6.2 – –
1/10 22.6 5.6 – –
– no pre-transition observed in this case.
Table 2Antimicrobial and lytic activities of penetratin and RL16a
Peptide B. megaterium E. coli S. aureus K. pneumoniae
Penetratin 0.2 μMb N100 μM N100 μM N100 μMRL16 1.5 μMb 50 μMb 9 μMc N100 μM
Note: hemolytic activities of these peptides are given in the text.a The antimicrobial activity is expressed as MIC (μM), the minimal peptide
concentration required for total inhibition of cell growth in liquid medium. Resultsare the mean of three independent experiments each performed in duplicate.
b Lytic effect observed at this concentration.c No lytic effect observed at this concentration.
951I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
transition (see Table 3 for the thermodynamic parameters). Broad-ening of the DSC curves and decreases in the Tm are compatible with abetter interaction of the peptidewith the liquid crystalline phase of thelipid compared with the gel phase. A similar behavior has beenobserved with other antimicrobial peptides [22,23]. Moreover, thepresence of increasing quantities of RL16 also results in the inductionof a two-component main phase transition, with a more cooperative,lower temperature endotherm superimposed over a less cooperative,
Fig. 1. High-sensitivity DSC heating scans illustrating the effect of the addition ofincreasing quantities of RL16 (panels A and B) and penetratin (panels C and D) on thethermotropic phase behavior of DMPC and DMPG multilamellar vesicles. The top scancorresponds to the lipid alone and the peptide/lipid molar ratios of the lower scans areindicated in the figure. In the insert a detailed view of the main transition is provided.Thermodynamic parameters are given in Tables 3 and 4.
higher temperature endotherm (see insert in Fig. 1A). Peak splitting inthe DSC profiles suggests the possible presence of distinct lipid–peptide domains. The appearance of shoulders in the thermogram is aconsequence of a non-ideal mixing behavior, which creates a non-homogeneous distribution of the peptide within the membrane. Thelow broad component is attributed to lipids that are in peptide-richregions or those that are in close proximityand so arehighly perturbed.The sharper peak, which resembles more the pure lipid transition, isattributed to lipids that are in peptide-poor regions corresponding tounperturbed lipid molecules [22,24–27].
It should be pointed out that these DSC studieswere performedwithMLVs because they exhibit much more cooperative lipid phasetransitions than LUVs. Even though the majority of published calori-metry studies involving peptide–lipid interactions have reported co-dissolution of the peptide with the lipid prior to hydration and vesicleformation, we have performed otherwise because we believe that theaddition of the peptide to pre-formed liposomes is more biologicallyrelevant.We have observed in certain cases that the thermotropic phasebehaviorwas not completely reproducible, at least for thefirst fewscans,presumably because the peptide was progressively gaining access tointernal bilayers, so at least 6–8 scans were performed in each study.Thermodynamic analysis presented in Tables 3 and 4 was performedusing the last scans once equilibrium has been reached.
The physico-chemical properties of DMPG vesicles (stability, sizeand lipid thermal behavior) depend on the specific environment, suchas the ionic strength of the sample, the presence of divalent cations,the pH and even the lipid concentration [28–31]. It is seen in Fig. 1B,that in contrast to DMPC, the DMPG main transition band is notsymmetrical and exhibits a marked high temperature shoulder. Thisshoulder is due to the low ionic strength used in this experiment sinceit disappears when 500 mM NaCl is used instead of the 100 mMdescribed [24]. This high temperature shoulder becomes lesspronounced in the presence of RL16, this may be due to the interactionof the peptide with the lipid surface, which leads to a reduction of thecharge–charge repulsion of the lipid headgroups. In contrast to the
Table 4Thermodynamic parameters obtained by DSC for the interaction of penetratin witheither MLVs of DMPC or DMPG
Lipid P/L Tm (°C) ΔHm Kcal/mol Tpre (°C) ΔHpre kcal/mol
DMPC 0 23.8 6.6 13.6 0.91/100 23.8 6.8 14.1 1.11/50 23.8 7.0 14.3 11/25 23.8 7.1 14.5 1.21/10 23.8 7.7 14.5 1.3
DMPG 0 22.1 6.2 8.6 0.41/100 22.1 5.6 8.6 0.31/50 21.9 4.3 7.6 0.11/25 21.9 3.8 – –
1/10 21.9 2 – –
– no pre-transition observed in this case.

952 I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
observations with DMPC, the interaction of RL16with DMPGMLVs hasminor effects on the energetics of the main phase transition (Fig. 1Band Table 3), indicating that in this case RL16 does not penetratedeeply in the lipid hydrophobic core. The only effect observedconcerns the pre-transition that is abolished at P/L of 1/50, confirmingthe interaction of this peptide with the head groups. Since RL16contains ten basic residues, a preferential interaction of the peptidewith an anionic bilayer relative to a zwitterionic one would beexpected. Surprisingly, it seems that the negative charges on themembrane surface sequester the peptide in the lipid headgroupregion, and thus prevent RL16 from entering into the hydrophobic partof the bilayer, a similar behavior has been observed for melittin [32].
Regarding penetratin interaction with DMPC (Fig. 1C), no majoreffects were observed in the main phase transition: no change in thephase transition temperature, small change in the enthalpy and a smalldecrease in the cooperativity of the transition. These results are con-sistent with aweak interaction of penetratinwith this lipid. Regardingthe interaction with anionic lipids (DMPG), a much stronger effect ofpenetratin on the pre- and main phase transitions of the lipid (Fig. 1D,Table 4) was observed. The energetics of the pre-transition decreasedcontinuously with the peptide concentration until it vanished at P/Lratio of 1/25. The pre-transition, which is observed for some saturatedphospholipids like DPPG and DPPC, is related to the untilting of thehydrocarbon side chains and usually is even more sensitive to thepresence of foreignmolecules than themain transition. The untiltingofthe hydrocarbon chainsmay be explained by a simple neutralization ofthe headgroup charge by the cationic peptide, which will result inreduced electrostatic headgroup repulsion and concomitantly in asmaller headgroup area observed in the case of DMPG. Strong effectson the pre-transition, including abolition, have been observed forothermembrane active peptides such asmelittin and δ-lisin. Regardingthe DMPG main phase transition, a large effect was observed uponpenetratin interaction, with a N60% decrease in the enthalpy and asmall decrease in Tm. The enthalpy of themain transition is mainly dueto the disruption of intra- and inter-molecular van der Waals inter-actions that lead to chain melting, indicating some level of peptideinsertion within the acyl chain region. Like in the case of RL16, inter-action with DMPC bilayer, a splitting in the transition peak wasobserved, probably due to the presence of distinct domains in thesystem (peptide rich and peptide poor) as a consequence of in-homogeneous peptide distributionwithin the lipid. The higher affinityof penetratin for negatively charged membranes has already beenobserved [6,13] and maybe a consequence of an accumulation in theaqueous lipid–water interface by electrostatic attraction rather than aspecific binding to anionic lipids [13].
3.4. Peptide effect on the lipid membrane curvature investigated by DSC
The effect of a peptide effect on the membrane monolayer cur-vature strain provides important information on the peptide mechan-ism of action. The effect can be determined by the shift in the lamellar
Fig. 2. High-sensitivity DSC heating scans illustrating the effect of the addition of RL16multilamellar vesicles. Panel A represents the lipid alone, and panels B and C upon additionphase transition temperature (TH) observed is 40.4, 46.7 and 38.1 °C for the lipid alone, and
(Lα) to hexagonal (HII) phase transition temperature (TH), resultingfrom the introduction of a peptide into a pure phospholipid mem-brane providing important information on the peptide mechanism ofaction [25,32–34]. The shift in TH is a consequence of the stability ofthe HII phase and changes in monolayer curvature. The effect of RL16and penetratin on membrane spontaneous curvature was studied bymonitoring their capacity to alter TH of DiPoPE, a standard lipid usedfor these experiments, owing to its favorable thermodynamic be-havior [26,35]. It is well documented that the Lα-to-HII phase transi-tion has a low energy profile, the best transition being that of DiPoPE.The DSC thermograms of samples composed of DiPoPE combinedwithRL16 and penetratin, are shown in Fig. 2. As one can see, the transitionobserved is quite small, even at such high lipid concentration.We haveobserved that there was some lipid precipitation due to the highconcentration employed. As observed in Fig. 2, the lipid aloneexhibited a TH of 40.4 °C. Both peptides lead to a decrease in thecalorific capacity (Cp) suggesting that they perturb the lipid phasetransition by intercalating between the fatty acid chains. As for theireffects on the phase transition temperature, RL16 leads to an increasein TH to 46.7 °C meaning that RL16 disfavors a negative curvature, soinduces a positive curvature strain in the lipid bilayers. Regardingpenetratin, an opposite effect was observed, there is a decrease in THto 38.1 °C, implying that penetratin induces a negative curvature in thelipid bilayer. This information is important to elucidate the penetratinmode of action (this will be further discussed below).
3.5. The effect of the peptides on the structural organization of the lipidsas determined by 31P NMR
In order to get some insight on the peptide effects on the lipidproperties and their supramolecular organization, 31P NMR wasemployed. MLVs of DMPC were prepared and the peptide was addedeither prior to liposome formation (before forming the lipid film) orafter. Most of the reported studies so far, have chosen to pre-mix thelipid and the peptide prior to liposome formation since a morehomogeneous sample is obtained. However, in our mind, the additionof the peptide after liposome formation seems to better mimic thebiological event. Moreover, adding the peptide to the liposomes allowsa better monitoring of the kinetics. When the peptide did not lead toliposome precipitation, the addition of the peptide before or after MLVformation gave similar NMR profiles, although the kinetics of theprocess is slower for the peptide added after MLV formation, since ithas to get access to the internal lipid layers. The NMR signal wasobtained at several temperatures: below, at and above the phasetransition temperature of the lipid.
The presence of RL16 in DMPC MLVs (peptide being pre-mixedwith the lipids before MLV formation), below and at the lipid phasetransition temperature, leads to the appearance of an isotropic signallocated at 0 ppm, this signal is indicative of an isotropic lipid mor-phology, such as micelles, very small vesicles or cubic phases inwhichthe lipids are rapidly diffusing in all directions in space (Fig. 3). The
(panels B) and penetratin (panels C) on the thermotropic phase behavior of DiPoPEof RL16 and penetratin, respectively at a P/L of 1/25. The lamellar (Lα) to hexagonal (HII)in the presence of RL16 and penetratin, respectively.
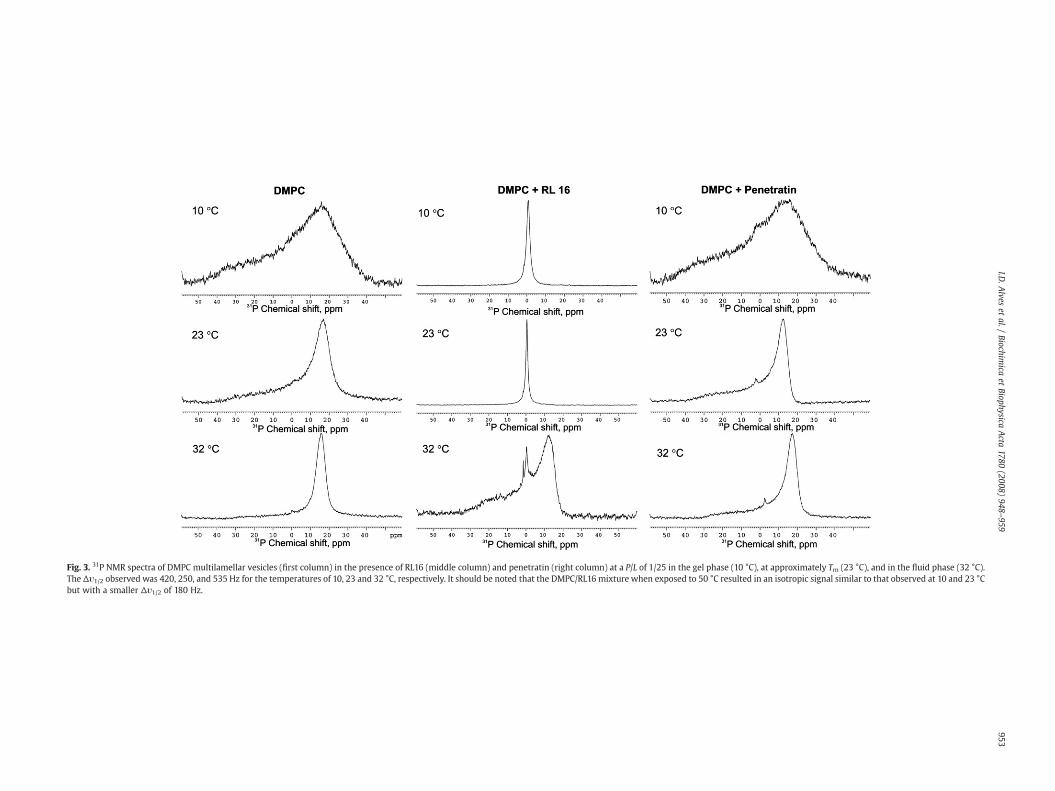
Fig. 3. 31P NMR spectra of DMPC multilamellar vesicles (first column) in the presence of RL16 (middle column) and penetratin (right column) at a P/L of 1/25 in the gel phase (10 °C), at approximately Tm (23 °C), and in the fluid phase (32 °C).The Δυ1/2 observed was 420, 250, and 535 Hz for the temperatures of 10, 23 and 32 °C, respectively. It should be noted that the DMPC/RL16 mixture when exposed to 50 °C resulted in an isotropic signal similar to that observed at 10 and 23 °Cbut with a smaller Δυ1/2 of 180 Hz.
953I.D
.Alves
etal./
Biochimica
etBiophysica
Acta
1780(2008)
948–959
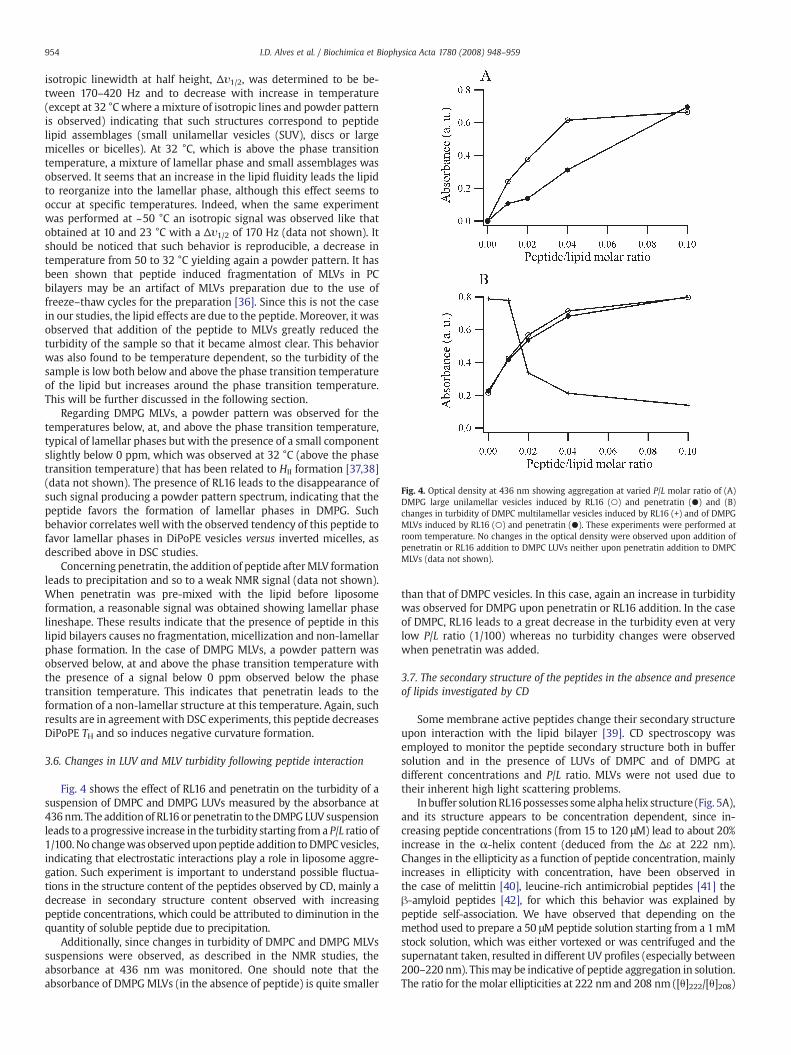
Fig. 4. Optical density at 436 nm showing aggregation at varied P/L molar ratio of (A)DMPG large unilamellar vesicles induced by RL16 (○) and penetratin (●) and (B)changes in turbidity of DMPC multilamellar vesicles induced by RL16 (+) and of DMPGMLVs induced by RL16 (○) and penetratin (●). These experiments were performed atroom temperature. No changes in the optical density were observed upon addition ofpenetratin or RL16 addition to DMPC LUVs neither upon penetratin addition to DMPCMLVs (data not shown).
954 I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
isotropic linewidth at half height, Δυ1/2, was determined to be be-tween 170–420 Hz and to decrease with increase in temperature(except at 32 °C where a mixture of isotropic lines and powder patternis observed) indicating that such structures correspond to peptidelipid assemblages (small unilamellar vesicles (SUV), discs or largemicelles or bicelles). At 32 °C, which is above the phase transitiontemperature, a mixture of lamellar phase and small assemblages wasobserved. It seems that an increase in the lipid fluidity leads the lipidto reorganize into the lamellar phase, although this effect seems tooccur at specific temperatures. Indeed, when the same experimentwas performed at ~50 °C an isotropic signal was observed like thatobtained at 10 and 23 °C with a Δυ1/2 of 170 Hz (data not shown). Itshould be noticed that such behavior is reproducible, a decrease intemperature from 50 to 32 °C yielding again a powder pattern. It hasbeen shown that peptide induced fragmentation of MLVs in PCbilayers may be an artifact of MLVs preparation due to the use offreeze–thaw cycles for the preparation [36]. Since this is not the casein our studies, the lipid effects are due to the peptide. Moreover, it wasobserved that addition of the peptide to MLVs greatly reduced theturbidity of the sample so that it became almost clear. This behaviorwas also found to be temperature dependent, so the turbidity of thesample is low both below and above the phase transition temperatureof the lipid but increases around the phase transition temperature.This will be further discussed in the following section.
Regarding DMPG MLVs, a powder pattern was observed for thetemperatures below, at, and above the phase transition temperature,typical of lamellar phases but with the presence of a small componentslightly below 0 ppm, which was observed at 32 °C (above the phasetransition temperature) that has been related to HII formation [37,38](data not shown). The presence of RL16 leads to the disappearance ofsuch signal producing a powder pattern spectrum, indicating that thepeptide favors the formation of lamellar phases in DMPG. Suchbehavior correlates well with the observed tendency of this peptide tofavor lamellar phases in DiPoPE vesicles versus inverted micelles, asdescribed above in DSC studies.
Concerning penetratin, the addition of peptide afterMLV formationleads to precipitation and so to a weak NMR signal (data not shown).When penetratin was pre-mixed with the lipid before liposomeformation, a reasonable signal was obtained showing lamellar phaselineshape. These results indicate that the presence of peptide in thislipid bilayers causes no fragmentation, micellization and non-lamellarphase formation. In the case of DMPG MLVs, a powder pattern wasobserved below, at and above the phase transition temperature withthe presence of a signal below 0 ppm observed below the phasetransition temperature. This indicates that penetratin leads to theformation of a non-lamellar structure at this temperature. Again, suchresults are in agreementwith DSC experiments, this peptide decreasesDiPoPE TH and so induces negative curvature formation.
3.6. Changes in LUV and MLV turbidity following peptide interaction
Fig. 4 shows the effect of RL16 and penetratin on the turbidity of asuspension of DMPC and DMPG LUVs measured by the absorbance at436 nm. The addition of RL16 or penetratin to theDMPG LUV suspensionleads to a progressive increase in the turbidity starting from a P/L ratio of1/100. No changewas observeduponpeptide addition toDMPC vesicles,indicating that electrostatic interactions play a role in liposome aggre-gation. Such experiment is important to understand possible fluctua-tions in the structure content of the peptides observed by CD, mainly adecrease in secondary structure content observed with increasingpeptide concentrations, which could be attributed to diminution in thequantity of soluble peptide due to precipitation.
Additionally, since changes in turbidity of DMPC and DMPG MLVssuspensions were observed, as described in the NMR studies, theabsorbance at 436 nm was monitored. One should note that theabsorbance of DMPGMLVs (in the absence of peptide) is quite smaller
than that of DMPC vesicles. In this case, again an increase in turbiditywas observed for DMPG upon penetratin or RL16 addition. In the caseof DMPC, RL16 leads to a great decrease in the turbidity even at verylow P/L ratio (1/100) whereas no turbidity changes were observedwhen penetratin was added.
3.7. The secondary structure of the peptides in the absence and presenceof lipids investigated by CD
Some membrane active peptides change their secondary structureupon interaction with the lipid bilayer [39]. CD spectroscopy wasemployed to monitor the peptide secondary structure both in buffersolution and in the presence of LUVs of DMPC and of DMPG atdifferent concentrations and P/L ratio. MLVs were not used due totheir inherent high light scattering problems.
Inbuffer solutionRL16possesses somealphahelix structure (Fig. 5A),and its structure appears to be concentration dependent, since in-creasing peptide concentrations (from 15 to 120 μM) lead to about 20%increase in the α-helix content (deduced from the Δε at 222 nm).Changes in the ellipticity as a function of peptide concentration, mainlyincreases in ellipticity with concentration, have been observed inthe case of melittin [40], leucine-rich antimicrobial peptides [41] theβ-amyloid peptides [42], for which this behavior was explained bypeptide self-association. We have observed that depending on themethod used to prepare a 50 μM peptide solution starting from a 1mMstock solution, which was either vortexed or was centrifuged and thesupernatant taken, resulted in different UV profiles (especially between200–220 nm). Thismay be indicative of peptide aggregation in solution.The ratio for the molar ellipticities at 222 nm and 208 nm ([θ]222/[θ]208)
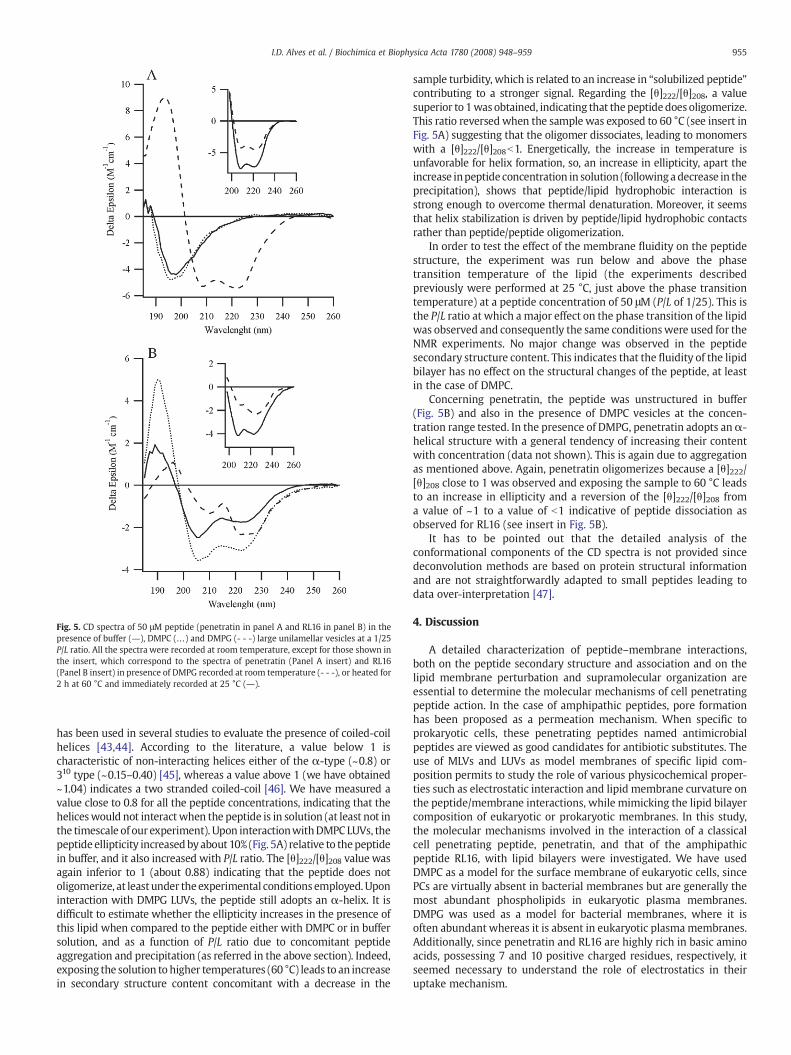
Fig. 5. CD spectra of 50 μM peptide (penetratin in panel A and RL16 in panel B) in thepresence of buffer (―), DMPC (…) and DMPG (- - -) large unilamellar vesicles at a 1/25P/L ratio. All the spectra were recorded at room temperature, except for those shown inthe insert, which correspond to the spectra of penetratin (Panel A insert) and RL16(Panel B insert) in presence of DMPG recorded at room temperature (- - -), or heated for2 h at 60 °C and immediately recorded at 25 °C (―).
955I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
has been used in several studies to evaluate the presence of coiled-coilhelices [43,44]. According to the literature, a value below 1 ischaracteristic of non-interacting helices either of the α-type (~0.8) or310 type (~0.15–0.40) [45], whereas a value above 1 (we have obtained~1.04) indicates a two stranded coiled-coil [46]. We have measured avalue close to 0.8 for all the peptide concentrations, indicating that theheliceswould not interact when the peptide is in solution (at least not inthe timescale of our experiment). Upon interactionwithDMPC LUVs, thepeptide ellipticity increased byabout 10% (Fig. 5A) relative to thepeptidein buffer, and it also increased with P/L ratio. The [θ]222/[θ]208 value wasagain inferior to 1 (about 0.88) indicating that the peptide does notoligomerize, at least under the experimental conditions employed. Uponinteraction with DMPG LUVs, the peptide still adopts an α-helix. It isdifficult to estimate whether the ellipticity increases in the presence ofthis lipid when compared to the peptide either with DMPC or in buffersolution, and as a function of P/L ratio due to concomitant peptideaggregation and precipitation (as referred in the above section). Indeed,exposing the solution to higher temperatures (60 °C) leads to an increasein secondary structure content concomitant with a decrease in the
sample turbidity, which is related to an increase in “solubilized peptide”contributing to a stronger signal. Regarding the [θ]222/[θ]208, a valuesuperior to 1was obtained, indicating that thepeptidedoes oligomerize.This ratio reversed when the sample was exposed to 60 °C (see insert inFig. 5A) suggesting that the oligomer dissociates, leading to monomerswith a [θ]222/[θ]208b1. Energetically, the increase in temperature isunfavorable for helix formation, so, an increase in ellipticity, apart theincrease inpeptide concentration in solution (followingadecrease in theprecipitation), shows that peptide/lipid hydrophobic interaction isstrong enough to overcome thermal denaturation. Moreover, it seemsthat helix stabilization is driven by peptide/lipid hydrophobic contactsrather than peptide/peptide oligomerization.
In order to test the effect of the membrane fluidity on the peptidestructure, the experiment was run below and above the phasetransition temperature of the lipid (the experiments describedpreviously were performed at 25 °C, just above the phase transitiontemperature) at a peptide concentration of 50 μM (P/L of 1/25). This isthe P/L ratio at which a major effect on the phase transition of the lipidwas observed and consequently the same conditionswere used for theNMR experiments. No major change was observed in the peptidesecondary structure content. This indicates that the fluidity of the lipidbilayer has no effect on the structural changes of the peptide, at leastin the case of DMPC.
Concerning penetratin, the peptide was unstructured in buffer(Fig. 5B) and also in the presence of DMPC vesicles at the concen-tration range tested. In the presence of DMPG, penetratin adopts anα-helical structure with a general tendency of increasing their contentwith concentration (data not shown). This is again due to aggregationas mentioned above. Again, penetratin oligomerizes because a [θ]222/[θ]208 close to 1 was observed and exposing the sample to 60 °C leadsto an increase in ellipticity and a reversion of the [θ]222/[θ]208 froma value of ~1 to a value of b1 indicative of peptide dissociation asobserved for RL16 (see insert in Fig. 5B).
It has to be pointed out that the detailed analysis of theconformational components of the CD spectra is not provided sincedeconvolution methods are based on protein structural informationand are not straightforwardly adapted to small peptides leading todata over-interpretation [47].
4. Discussion
A detailed characterization of peptide–membrane interactions,both on the peptide secondary structure and association and on thelipid membrane perturbation and supramolecular organization areessential to determine the molecular mechanisms of cell penetratingpeptide action. In the case of amphipathic peptides, pore formationhas been proposed as a permeation mechanism. When specific toprokaryotic cells, these penetrating peptides named antimicrobialpeptides are viewed as good candidates for antibiotic substitutes. Theuse of MLVs and LUVs as model membranes of specific lipid com-position permits to study the role of various physicochemical proper-ties such as electrostatic interaction and lipid membrane curvature onthe peptide/membrane interactions, while mimicking the lipid bilayercomposition of eukaryotic or prokaryotic membranes. In this study,the molecular mechanisms involved in the interaction of a classicalcell penetrating peptide, penetratin, and that of the amphipathicpeptide RL16, with lipid bilayers were investigated. We have usedDMPC as a model for the surface membrane of eukaryotic cells, sincePCs are virtually absent in bacterial membranes but are generally themost abundant phospholipids in eukaryotic plasma membranes.DMPG was used as a model for bacterial membranes, where it isoften abundant whereas it is absent in eukaryotic plasmamembranes.Additionally, since penetratin and RL16 are highly rich in basic aminoacids, possessing 7 and 10 positive charged residues, respectively, itseemed necessary to understand the role of electrostatics in theiruptake mechanism.

956 I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
From the biophysical tools employed in this study, wewould like toemphasize the diversity and importance in the information providedby DSC, which has emerged as a most valuable tool to study peptide–membrane interactions [6,28–30,48–51]. DSC analysis providesimportant information regarding the level of interaction of thepeptide with the lipid and the eventual formation of domainsconsecutive to rearrangement of lipid components into lipid domainsinduced by the presence of a peptide/protein [52]. However, in orderto be able to gain a maximum of information and to propose a modelfor the mechanism of action of membrane active peptides, acombination of DSC with other biophysical tools is essential and wasemployed herein. It should be pointed out that the concentrations ofpeptide employed in the biophysical studies were higher than the MICvalues obtained for those peptides due to the sensitivity limits of suchtechniques.
4.1. Interaction of RL16 with a lipid bilayer
RL16 is a perfect amphipathic peptide whose sequence is derivedfrom sequence/uptake relationships of penetratin. The fact that thispeptide is perfectly amphipathic leads us to evaluate its potentialantimicrobial activity. RL16 possess antimicrobial activity in bothB. megaterium (MIC=1.5 μM), S. aureus (MIC=6–12 μM) and E. coli(MIC=50 μM), being bactericidal in both B. megaterium and E. coli andbacteriostatic in S. aureus. Additonally, the peptide presents hemolyticactivity at high concentration. This behaviour has been also observedon eukaryote cells since, namely on MDCK cell permeabilizationstudies measured by annexin movement upon calcium uptake,showing that membrane permeability is affected [8]. Deleteriouseffects in mammalian cell membrane integrity require prolongedincubation (N4 h) at 10 μM, suggesting that the peptide could lead tothe formation of pores, although much less deleterious than melittinwhere membrane integrity was readily perturbed after 10 minincubation. Calorimetric studies indicate that RL16 has a strongperturbation in the lipid chain packing of DMPC, suggesting that thepeptidemay insert deeply in the lipid fatty acyl chain region. In DMPG,RL16 does not insert into the acyl chain region but rather remains at theheadgroup region where it establishes electrostatic interactions.Although, an attraction between cationic peptides and anionic lipidswould have been expected, anionic lipids are not essential for such aninteraction. Indeed, it seems that the peptide, by establishingelectrostatic interactions between the arginine residues and the lipidheadgroups remains anchored on the bilayer surface and inhibits itsinsertion into the membrane core region. Other cationic peptides likemagainin and PGLa also do not rely on the presence of anionic lipids inthe bilayer for their interactions [48,53,54], moreover melittin lyticactivity has been shown to decrease in the presence of anionic lipids[32]. The antimicrobial activity of this peptide could not be explainedby its interactionwith PG so it should rely on the interactionwith otherlipids. It should be noted that all bacteria strains are rich in PG but apeptide may preferentially interact with certain strains (and possessdistinct MIC values), which is a consequence of the distinct lipidcomposition of their bacterial membrane, other than PG.
31P NMR studies show that the addition of RL16 to MLVs of DMPCleads to the disappearance of a powder pattern (typical of lamellarphases) and the appearance of an isotropic signal both below, right atthe phase transition temperature of the lipid and well above (Fig. 4),which according to its Δυ1/2 correspond to the formation of smallobjects such as vesicles or discs. Right above the phase transitiontemperature, at 32 °C, a mixture of a powder pattern and isotropicsignal is observed, indicating that an increase in the fluidity of thebilayer would favor the formation of lamellar phases that may coexistwith small vesicles. A very similar behavior has been observed byDufourc et al. on the action of melittin on DPPC [55], where thepresence of an isotropic signal was observed onmixtures of DPPCwithmelittin, both below and above the phase transition temperature and
amixture of isotropic signal with powder pattern observed around thephase transition temperature. A similar behavior has also been ob-served with several other antimicrobial peptides, such as a magaininanalog (MSI-78) [26], gramicidin S [51], but also in the case of a cell-penetrating peptide, TAT, whose internalization has been postulatedto occur through endocytosis [56]. Membrane fragmentation has beenobserved upon interaction of amyloidogenic fragments where vesicleswere observed with a similar size to the one observed in this study(~100 nm) [57]. The formation of an isotropic signal was observed formixtures of TAT and to a less extent Arg8 with DMPC, but not withDMPG bilayers [50]. The 31P-NMR results were interpreted on thebasis of small vesicle or disc formation, which at the phase transitiontemperature probably coexist with membrane discs. Additionally, thepresence of RL16 greatly reduces the turbidity of the DMPC MLVssample at all the temperatures tested, except at 32 °C. As for DMPG,the peptide appears to favor lamellar rather than inverted hexagonalphases as indicated by the presence of a powder pattern in the 31PNMR spectrum. Such results are consistent with an increase in thehexagonal phase transition temperature of DiPoPE induced by RL16,which disfavors an hexagonal phase or promotes a positive curvaturein the lipid bilayer. The induction of positive curvature strain byantimicrobial peptides has been observed [26,58–60].
Concerning the secondary structure, RL16 adopts an α-helix asexpected for an amphipathic peptide, which is promoted in thepresence of lipids, additionally the peptide oligomerizes in thepresence of DMPG. These peptide oligomers dissociate when exposedto high temperatures (60 °C) leading to an increase in the helicalcontent. Such results indicate that the peptide structure would prevailevenwhen peptide/peptide interactions are disrupted suggesting thatlipophilic forces between the peptide and the lipid acyl chain mayensure the peptide helical organization.
A model for the mechanism of action is proposed, which takes intoaccount all this data (Fig. 6). According to the model, the peptide insolutionwould have some degree of α-helix content with an eventualaggregation taking place, the presence of a zwitterionic lipid (such asDMPC) would lead to an increase in peptide structure driven byhydrophobic contacts between the peptide leucine rich face and thelipid hydrophobic core. RL16 would have its hydrophilic face exposingthe extra vesicle medium (buffer). When the peptide accesses theinterior vesicles of MLVs, electrostatic repulsions may originatebetween charged arginines of contacting peptides present in adjacentlipid layers eventually leading to the disruption ofMLVs and formationof small lipid assemblages. In those assemblages the peptide couldorient either parallel to the disc surface or perpendicular with itshydrophobic side facing the lipid and hydrophilic exposed to thesolvent. This process could be accompanied by the transient formationof lipid micelles, reflecting a weak detergent effect of the peptide. Inpresence of anionic lipids (such as DMPG), RL16 would ratherestablish electrostatic interactions between the arginines and thenegatively charged lipid headgroups, such an interaction wouldsimultaneously lead to peptide oligomerization in order to preventunfavorable hydrophobic exposure of the peptide hydrophobic faceand concomitant liposome aggregation (see Fig. 6).
In terms of a mechanistic model, RL16 seems to act according to adetergent-like carpet mechanism (formation of small lipid assem-blages). According to this model, the peptides accumulate at thebilayer surface like a carpet, and above a threshold concentration ofmonomers, the membrane is permeated and disintegrated in adetergent-like manner. Two other permeation mechanisms havebeen proposed so far: the barrel stavemodel inwhich the amphipathicpeptides aggregate and insert into the lipid bilayer forming a channel,the toroidal, detergent or carpet model in which the pore is linedby both peptide and lipid headgroups [49,61]. It may be rathercomplicated, in some instances, to differentiate between the carpetand the toroidal pore mechanisms. In the carpet-model, peptidesremain associated with the phospholipids head group throughout the

Fig. 6.Model to explain the behavior of RL16 upon interactionwith DMPC and DMPGmultilamellar vesicles. The peptide possesses two faces: the hydrophilic face is presented in lightgray and the hydrophilic in black. See text for a description of the model.
957I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
membrane disintegration process. This is similar to the transition thatoccurs in the toroidal-pore model, and it has been suggested that theformation of “holes” or toroidal poresmay occur as an early step in themembrane disintegration process, i.e. toroidal pores are regarded asan intermediate state prior to micellization [49]. It seems likely thatmost AMPs, by virtue of their amphipathic character, will act asdetergents at sufficiently high concentrations. Nonetheless, in manycases (e.g., magainin, MSI-78) it is clear that pore formation is suf-ficient to inducemembrane lysis, without the progression to completemembrane disruption. At this time, we cannot distinguish betweenthese two mechanisms, other experiments are in progress to obtaininformation about the orientation of the peptide relative to the lipidbilayer that may allow one to shed light on this question. Although,the reversible formation of lamellar phases and small lipid assem-blages observed by NMR as well as the slight effects in membraneintegrity, when comparing to melittin, a peptide known to form holesin bilayers, may suggest that this peptide would eventually formtransient pores in the bilayer. It should be kept in mind that, suchtransient pores are either kinetically (transiently formed) and/ordynamically (less numerous and of smaller size) distinct from thoseobserved uponmelittin action. This will also explain the bacteriostaticactivity, without lysis, of RL16 on S. aureus. So, bacteria growthinhibition could be explained either by a coating of the bacteriasurface by the peptide, which inhibits its growing or by a transientdisturbance of the bacterial membrane, which leads to the formationof pores of rather short size and lifetime.
4.2. Interaction of penetratin with a lipid bilayer
Penetratin is a cationic non amphipathic peptide with a greatuptake capacity whose internalization mechanism remains highlycontroversial. Calorimetric studies show that peptide induced nosubstantial effect in the lipid phase transition and pre-transition ofDMPC, suggesting very weak or no interaction with this lipid. In thecase of DMPG, a strong perturbation on both the pre-transition andmain phase transitionwas observed suggesting that electrostatics playa role in peptide/lipid interaction and that penetratin is able to insertinto the acyl chain region. A preferential interaction of penetratinwithanionic versus zwitterionic lipids has been often proposed [13,39].Concerning the peptide secondary structure, penetratin is unstruc-tured in solution (buffer) and also in the presence of DMPC, but adoptsan α-helical structure in presence of DMPG. The electrostatic inter-action between the arginine residues in the peptide and the lipid
headgroups may trigger peptide structure which creates hydrophobicpatches in the peptide allowing partial insertion into the lipid acylchain region. Some studies have suggested that penetratin could alsoadopt a β-sheet structure under certain conditions such as certain P/Lratios, salt concentration [39] or upon vesicle aggregation [62]. Eventhoughwe have screened a large P/L ratio range, we have not observedβ-sheet formation. β-sheet formation being linked to peptide aggre-gation, it is plausible that wewere not able to observe it due to peptideprecipitation. Penetratin leads to a powder pattern in the 31P-NMRspectra in DMPC MLVs and induces the formation of a non-lamellarstructure in DMPG only above the lipid phase transition temperature.The peptide does not affect the membrane integrity of mammaliancells nor bacteria (except for B. megaterium, which is a bacterial strainrather susceptible to different types of molecules) suggesting thatinternalization of penetratin does not occur via pore formation ordetergent-type mechanism where membrane disruption takes place.DSC data indicates that penetratin favors the formation of non-lamellar lipid structures possessing negative curvature such asinverted micelles. The translocation of penetratin by an invertedmicelle mechanism has been proposed [6] and a role of transient non-bilayer structure (?) formation has been proposed for membranetranslocation of proteins across the lipid bilayer [63]. However, thereis still uncertainty about the uptake mechanism into cells, with somestudies reported a direct translocation through liposomes [64,65]while others go against that [66,67]. Our studies point to the inductionof a negative lipid curvature by penetratin on model systems ofspecific lipid composition, if this is indeed the case also in cells stillneeds further investigation.
Studying these two CPPs, which are both highly charged but differin their amphipathicity, allows to demonstrate that the mode ofinteraction of cationic peptides with lipids and the degree of mem-brane structure perturbation is divergent. And the results gained so farprovide important details on the mechanism of action of penetratinand RL16 and may be relevant for other amphipathic peptides. Theinteractions of such peptides are partly dictated by two factors: i) thespatial charge distribution in the peptide sequence and ii) the balancebetween from lipophilic forces within the peptide sequence and thelipid core region and hydrophilic and electrostatic contacts from thepeptide charged residues and the lipid headgroup region. The initialinteractions are the hallmark, which determines the internalizationprocess. More generally, two points come out from this work thatcould be useful for further studies. Firstly, testing the effects of CPPs onmembrane perturbation in distinct cell membranes provides valuable

958 I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
information regarding preferential interaction with a membranepossessing a particular lipid composition, such information has to betaken into account when selecting the lipids employed in biophysicalstudies lipid model systems. Secondly, we would like to stress theimportance of combining several biophysical techniques to study thevarious aspects of peptide–lipid interactions both from the peptideand lipid perspective which allow one to get a better insight in theunderstanding of mechanisms of transfer across a lipid bilayer.
In conclusion, the biophysical and biological results obtained withthese two CPPs provide important details that help distinguishbetween penetration (CPP activity) and lytic (AMP activity) properties.The borderline between these very distinct properties depends onslight differences in charge distribution and amphipathic character ofthe peptide and on the phospholipid structures as revealed by thesestudies involving two typical phospholipids. The next question raisedis to know whether penetrating and antimicrobial activities dependon the ability of the peptide to select lipids belonging to eithereukaryotic or prokaryotic membranes and to identify those potentiallipid molecules. Such experiments are currently underway.
Acknowledgements
This work was supported by the Association Nationale pour laRecherche (ANR-Prob DOM). We thank the laboratory of Dr. Guitatin the Museum National d’Histoire Naturelle for the use of theircalorimeter. Dr. Yves Jacquot and Vanessa Point are thanked for thepeptide synthesis.
References
[1] G.P. Dietz, M. Bähr, Peptide-enhanced cellular internalization of proteins inneuroscience, Brain Res. Bull. 68 (2005) 103–114.
[2] J.B. Rothbard, T.C. Jessop, P.A. Wender, Adaptive translocation: the role of hydrogenbonding and membrane potential in the uptake of guanidinium-rich transportersinto cells, Adv. Drug Deliv. Rev. 57 (2005) 495–504.
[3] F. Perret, M. Nishihara, T. Takeuchi, S. Futaki, A.N. Lazar, A.W. Coleman, N. Sakai, S.Matile, Anionic fullerenes, calixarenes, coronenes, and pyrenes as activators ofoligo/polyarginines in model membranes and live cells, J. Am. Chem. Soc. 127(2005) 1114–1115.
[4] D. Derossi, S. Calvet, A. Trembleau, A. Brunissen, G. Chassaing, A. Prochiantz, Cellinternalization of the third helix of the Antennapedia homeodomain is receptor-independent, J. Biol. Chem. 271 (1996) 18188–18193.
[5] P.E.G. Thorén, D. Persson, E.K. Esbjörner, M. Goksör, P. Lincoln, B. Nordén,Membrane binding and translocation of cell-penetrating peptides, Biochem. 43(2004) 3471–3489.
[6] D. Derossi, G. Chassaing, A. Prochiantz, Trojan peptides: the penetratin system forintracellular delivery, Trends in Cell Biol. 8 (1998) 84–87.
[7] C. Gerard, A. Prochiantz, Peptides usable as vectors for the intracellular addressingof bioactive molecules, PCT Int. Appl. WO 9712912 (1997) 31.
[8] A. Lamaziere, F. Burlina, C. Wolf, G. Chassaing, G. Trugnan, J. Ayala-Sanmartin, Non-metabolic membrane tabulation and permeability induced by bioactive peptides,PLoS ONE 2 (2007) e201.
[9] D. Derossi, A.H. Joliot, G. Chassaing, A. Prochiantz, The third helix of theAntennapedia homeodomain translocates through biological membranes, J. Biol.Chem. 271 (1996) 18188–18193.
[10] A. Prochiantz, Getting hydrophilic compounds into cells: lessons from homeo-proteins, Curr. Opin. Neurobiol. 6 (1996) 629–634.
[11] G. Drin, H. Demene, J. Temsamani, R. Brasseur, Translocation of the pAntp peptideand its amphipathic analogue AP-2AL, Biochemistry 40 (2001) 1824–1834.
[12] S.D. Conner, S.L. Schmid, Regulated portals of entry into the cell, Nature 422 (2003)37–44.
[13] H. Binder, G. Lindblom, Charge-dependent translocation of the Trojan peptidepenetratin across lipid membranes, Biophys. J. 85 (2003) 982–995.
[14] C. Palm, S. Netzereab, M. Hällbrink, Quantitatively determined uptake of cell-penetrating peptides in non-mammalian cells with an evaluation of degradationand antimicrobial effects, Peptides 27 (2006) 1710–1716.
[15] R.N. McElhaney, Differential scanning calorimetric studies of lipid–protein inter-actions in model membrane systems, Biochim. Biophys. Acta 864 (1986) 361–421.
[16] B. de Kruijff, Lipids beyond the bilayer, Nature 386 (1997) 129–130.[17] R.M. Epand, R.F. Epand, Calorimetric detection of curvature strain in phospholipid
bilayers, Biophys. J. 66 (1994) 1450–1456.[18] A.V. Botelho, T. Huber, T.P. Sakmar, M.F. Brown, Curvature and hydrophobic forces
drive oligomerization and modulate activity of rhodopsin in membranes, BiophysJ. 91 (2006) 4464–4477.
[19] M. Amiche, A. Séon, H. Wroblewski, P. Nicolas, Isolation of dermatoxin from frogskin, an antibacterial peptide encoded by a novel member of the dermaseptingenes family, Eur. J. Biochem. 267 (2000) 4583–4592.
[20] L. Zhong, W.C. Johnson Jr., Environment affects amino acid preference forsecondary structure, Proc. Natl. Acad. Sci. U.S.A. 89 (1992) 4462–4465.
[21] S. El-Andaloussi, P. Järver, H.J. Johansson, U. Langel, Cargo-dependent cytotoxicityand delivery efficacy of cell-penetrating peptides: a comparative study, Biochem J.407 (2007) 285–292.
[22] U.H. Dürr, U.S. Sudheendra, A. Ramamoorthy, LL-37, the only human member ofthe cathelicidin family of antimicrobial peptides, Biochim. Biophys. Acta 1758(2006) 1408–1425.
[23] R.F. Epand, A. Ramamoorthy, R.M. Epand, Membrane lipid composition and theinteraction of pardaxin: the role of cholesterol, Protein Pept Lett. 13 (2006) 1–5.
[24] V.V. Andrushchenko, H.J. Vogel, E.J. Prenner, Interactions of tryptophan-richcathelicidin antimicrobial peptides with model membranes studied by differentialscanning calorimetry, Biochim. Biophys. Acta 1768 (2007) 2447–2458.
[25] K. Lohner, E.J. Prenner, Differential scanning calorimetry and X-ray diffractionstudies of the specificity of the interaction of antimicrobial peptides withmembrane-mimetic systems, Biochim. Biophys. Acta 1462 (1999) 141–156.
[26] K.J. Hallock, D-K. Lee, A. Ramamoorthy, MSI-78, an analogue of the magaininantimicrobial peptides, disrupts lipid bilayer structure via positive curvaturestrain, Biophys. J. 84 (2003) 3052–3060.
[27] V. Dhople, A. Krukemeyer, A. Ramamoorthy, The human beta-defensin-3, anantibacterial peptide with multiple biological functions, Biochim. Biophys. Acta.1758 (2006) 1499–1512.
[28] M.T. Lamy-Freund, K.A. Riske, The peculiar thermo-structural behavior of theanionic lipid DMPG, Chem. Phys. Lipids 122 (2003) 19–32.
[29] Y.P. Zhang, R.N. Lewis, R.N. McElhaney, Calorimetric and spectroscopic studies ofthe thermotropic phase behavior of the n-saturated 1,2-diacylphosphatidylglycer-ols, Biophys. J. 72 (1997) 779–793.
[30] K.A. Riske, H.-G. Dobereiner,M.T. Lamy-Freund, Gel–fluid transition in dilute versusconcentrated DMPG aqueous dispersions, J. Phys. Chem.B 106 (2002) 239–246.
[31] R.M. Epand, S.W. Hui, Effect of electrostatic repulsion on the morphology andthermotropic transitions of anionic phospholipids, FEBS Lett. 209 (1986) 257–260.
[32] D.K.Hincha, J.H. Crowe, The lytic activity of thebee venompeptidemelittin is stronglyreduced by the presence of negatively charged phospholipids or chloroplastgalactolipids in the membranes of phosphatidylcholine large unilamellar vesicles,Biochim. Biophys.Acta 1284 (1996) 162–170.
[33] K.J. Hallock, D.K. Lee, J. Omnaas, H.I. Mosberg, A. Ramamoorthy, Membrane com-position determines pardaxin's mechanism of lipid bilayer disruption, Biophys. J.83 (2002) 1004–1013.
[34] J-P.S. Powers, A. Tan, A. Ramamoorthy, R.E.W. Hancok, Solution structure andinteraction of the antimicrobial polyphemusins with lipid membranes, Biochem.44 (2005) 15504–15513.
[35] R.M. Epand, R.F. Epand, Modulation of membrane curvature by peptides,Biopolymers 55 (2000) 358–363.
[36] C.D. Pointer-Keenan, D.K. Lee, K. Hallok, A. Tan, R. Zand, A. Ramamoorthy, A,Investigation of the interaction of myelin basic protein with phospholipid bilayersusing solid-state NMR spectroscopy, Chem Phys Lipids 132 (2004) 47–54.
[37] A.J. Verkleij, B. de Kruijff, P.H.J.T. Ververgaert, J.F. Tocanne, L.L.M. Van Deenen, Theinfluence of pH, Ca2+ and protein on the thermotropic behavior of the negativelycharged phospholipids, phosphatidylglycerol, Biochim. Biophys. Acta 339 (1974)432–437.
[38] L. Girault, A. Boudou, E.J. Dufourc, 113Cd-, 31P-NMR and fluorescence polarizationstudies of cadmium (II) interactions with phospholipids in model membranes,Biochim. Biophys. Acta 1414 (1998) 140–154.
[39] M. Magzoub, L.E. Eriksson, A. Graslund, Conformational states of the cell-penetrating peptide penetratin when interacting with phospholipids vesicles:effects of surface charge and peptide concentration, Biochim. Biophys. Acta 1563(2002) 53–63.
[40] E. Perez-Paya, R.A. Hougthen, S.E. Blondelle, The role of amphipathicity in thefolding, self-association and biological activity of multiple subunit small proteins,J. Biol. Chem. 270 (1995) 1048–1056.
[41] M.M. Javadpour, M.D. Barkley, Self-assembly of designed antimicrobial peptides insolution and micelles, Biochemistry 36 (1997) 9540–9549.
[42] E. Terzi, G. Hölzemann, J. Seelig, Self-association of beta-amyloid peptide (1–40) insolution and binding to lipid membranes, J. Mol. Biol. 252 (1995) 633–642.
[43] A.S. Ladokhin, S.H. White, Folding of amphiphatic α-helices on membranes:energetics of helix formation by melittin, J. Mol. Biol. 285 (1999) 1363–1369.
[44] N. Choy, V. Raussens, V. Narayanaswami, Inter-molecular coiledcoil formation inhuman apolipoprotein E C-terminal domain, J. Mol. Biol. 334 (2003) 527–539.
[45] M. Crisma, E. Andreetto, M. De Zotti, A. Moretto, C. Peggion, F. Formaggio, C.Toniolo, Crystal-state 3D-structural characterization of novel, Aib-based, turn andhelical peptides, J. Pept. Sci. 13 (2007) 190–205.
[46] T.M. Cooper, R.W. Woody, The effect of conformation on the CD of interactinghelices: a theoretical study of tropomyosin, Biopolymers 30 (1990) 657–676.
[47] S.M. Kelly, T.J. Jess, N.C. Price, How to study proteins by circular dichroism, Biochim.Biophys. Acta 1751 (2005) 119–139.
[48] T. Wieprecht, O. Apostolov, M. Beyermann, J. Seelig, Membrane binding and poreformation of the antibacterial peptide PGLa: thermodynamic and mechanisticaspects, Biochemistry 39 (2000) 442–452.
[49] Y. Shai, Mode of action of membrane active antimicrobial peptides, Biopolymers66 (2002) 236–248.
[50] S. Afonin, A. Frey, S. Bayerl, D. Fischer, P. Wadhwani, S. Weinkauf, A.S. Ulrich, Thecell-penetrating peptide TAT(48–60) induces a non-lamellar phase in DMPCmembranes, Chemphyschem. 13 (2006) 2134–2142.
[51] E.J. Prenner, R.N.A.H. Ruthven, R.E. McElhaney, The interaction of the antimicrobialpeptide gramicidin S with lipid bilayer model and biological membranes, Biochim.Biophys. Acta 1462 (1999) 201–221.

959I.D. Alves et al. / Biochimica et Biophysica Acta 1780 (2008) 948–959
[52] R.M. Epand, Detecting the presence of membrane domains using DSC, Biophys.Chem. 126 (2007) 197–200.
[53] B. Bechinger, M. Zasloff, S.J. Opella, Structure and orientation of the antibioticpeptide magainin in membranes by solid-state nuclear magnetic resonancespectroscopy, Protein Sci. 2 (1993) 2077–2084.
[54] M. Dathe, T. Wieprecht, Structural features of helical antimicrobial peptides: theirpotential to modulate activity on model membranes and biological cells, Biochim.Biophys. Acta Biomembr. 1462 (1999) 71–78.
[55] T. Pott, M. Paternostre, E.J. Dufourc, A comparative study of the action of melittin onsphingomyelin and phosphatidylcholine bilayers, Eur. Biophys. J. 27 (1998) 237–245.
[56] J.M. Gump, S.F. Dowdy, TAT transduction: the molecular mechanism andtherapeutic prospects, Trends Mol. Med. 13 (2007) 443–448.
[57] J.R. Brender, U.H. Dürr, D. Heyl, M.B. Budarapu, A. Ramamoorthy, Membranefragmentation by an amyloidogenic fragment of human Islet Amyloid Polypeptidedetected by solid-state NMR spectroscopy of membrane nanotubes, Biochim.Biophys. Acta 1768 (2007) 2026–2029.
[58] K. Matsuzaki, K. Sugishita, N. Ishibe, M. Ueha, S. Nakata, K. Miyajima, R.M. Epand,Relationship of membrane curvature to the formation of pores by magainin 2,Biochem. 37 (1998) 11856–11863.
[59] U.H. Dürr, U.S. Sudheendra, A. Ramamoorthy, LL-37, the only human member ofthe cathelicidin family of antimicrobial peptides, Biochim. Biophys. Acta 1758(2006) 1408–1425.
[60] A. Ramamoorthy, S. Thennarasu, A. Tan, D.K. Lee, C. Clayberger, A.M. Krensky, Cellselectivity correlates with membrane-specific interactions: a case study on the
antimicrobial peptide G15 derived from granulysin, Biochim. Biophys. Acta 1758(2006) 154–163.
[61] R.M. Epand, Y. Shai, J.P. Segrest, G.M. Anantharamaiah, Mechanisms for themodulation of membrane bilayer properties by amphipathic helical peptides,Biopolymers 37 (1995) 319–338.
[62] D. Persson, P.E. Thoren, E.K. Esbjorner, M. Goksor, P. Lincoln, B. Norden, Vesiclesize-dependent translocation of penetratin analogs across lipid membranes,Biochim. Biophys. Acta. 1665 (2004) 142–155.
[63] A.G. Rietveld, M.C. Koorengevel, B. de Kruijff, Non-bilayer lipids are required forefficient protein transport across the plasma membrane of Escherichia coli,EMBO J. 14 (1995) 5506–5513.
[64] P.E. Thorén, D. Persson, M. Karlsson, B. Nordén, The antennapedia peptidepenetratin translocates across lipid bilayers — the first direct observation, FEBSLett. 482 (2000) 265–268.
[65] D. Terrone, S.L. Sang, L. Roudaia, J.R. Silvius, Penetratin and related cell-penetratingcationic peptides can translocate across lipid bilayers in the presence of atransbilayer potential, Biochemistry 42 (2003) 13787–13799.
[66] E. Bárány-Wallje, S. Keller, S. Serowy, S. Geibel, P. Pohl, M. Bienert, M. Dathe, A criticalreassessment of penetratin translocation across lipid membranes, Biophys. J. 89(2005) 2513–2521.
[67] M.F. Ross, A. Filipovska, R.A. Smith, M.J. Gait, M.P. Murphy, Cell-penetratingpeptides do not cross mitochondrial membranes even when conjugated to alipophilic cation: evidence against direct passage through phospholipid bilayers,Biochem. J. 383 (2004) 457–468.











![Biochimica et Biophysica Acta - immed.org considerations/09.07.2017 updates/Membrane... · G.L. Nicolson, M.E. Ash / Biochimica et Biophysica Acta 1859 (2017) 1704–1724 1705 [8]](https://img.pdfslide.us/doc/110x75/5c684f1e09d3f2f5638b5509/biochimica-et-biophysica-acta-immed-considerations09072017-updatesmembrane.jpg)

















