Embed Size (px)
Citation preview

Biochemical properties of overloaded fast-twitch skeletal muscle
K. M. BALDWIN, V. VALDEZ, R. E. HERRICK, A. M. MACINTOSH, AND R. R. ROY Department of Physiology and Biophysics, University of California, Irvine 92717; and Department of Kinesiology, University of California, Los Angeles, California 90027
BALDWIN, K. M., V. VALDEZ, R. E. HERRICK, A. M. MAC- INTOSH, AND R. R. ROY. Biochemicalproperties of overloaded fast-twitch skeZetaZ muscle. J. Appl. Physiol.: Respirat. Envi- ron. Exercise Physiol. 52(2): 467-472, 1982.-Previous studies suggest that fast-twitch skeletal muscle overloaded by surgical removal of synergists contains a greater percent of slow-twitch fibers than normal muscle. Therefore we examined subcellular systems known to represent biochemical properties of slow- twitch skeletal muscle by measuring myosin ATPase, Ca”’ regulation of myofibril ATPase, Ca”’ uptake of sarcoplasmic reticulum (SR), and marker enzymes of glycogenolysis in nor- mal soleus (NS) and in normal (NP) and surgically overloaded (OP) plantaris muscles of adult female rats. The OP muscles were 65% larger than NP muscles (P < 0.001). Specific activity of myosin and myofibril ATPase was approximately 25% lower in OP compared with NP muscle (P < 0.05). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis of myosin revealed the presence of more slow and less fast myosin light-chain components in OP muscles. Although SR of NP muscle took up u,ore Ca2+ than OP muscle during the initial 2-min phase of the reaction, total accumulation was similar for both groups. Marker regulatory enzymes of glycogenolysis collectively were reduced by 40% in OP compared with NP muscle (P < 0.001). Collectively the data are consistent with the concept that some muscle fiber types were converted from “fast” to “slow” in the OP muscle.
mately 125 g were provided with food and water ad libitum. They were assigned to one of three groups des- ignated as normal plantaris (NP) and soleus (NS) and overloaded plantaris (OP). The specific factors in the compensatory hypertrophy model that induce the en- largement remain unclear at present (14, 21). Therefore the term overload is used rather loosely. Two series of experiments were performed. The first series focused on obtaining tissue for myofibril ATPase and SR Ca”+ up- take assays. The second series focused on obtaining tissue for myosin analyses and glycogenolytic enzyme assays.
myosin ATPase; myofibril ATPase; sarcoplasmic reticulum; calcium metabolism; glycogenolytic enzymes
RODENT FAST-TWITCH skeletal muscle overloaded by sur- gical removal of synergists contains a greater percentage of histochemically identifiable slow-twitch fibers than normal muscle (13, 17, 18). However, the extent that this change in fiber types is manifested in terms of the overall biochemical profile of the overloaded muscle remains uncertain. Slow-twitch skeletal muscle (i.e., soleus) is characterized biochemically as having 1) a distinct myosin isozyme pattern with low ATPase activity that is labile after incubation at alkaline pH; 2) a sarcoplasmic reticulum (SR) with slow Ca2’ uptake kinetics and a low Ca”’ accumulation capacity; and 3) relatively low levels of marker glycogenolytic enzymes (7,19,25). A study was therefore undertaken to investigate the biochemical properties of the above components in normal soleus and in normal and overloaded fast-twitch plantaris muscle.
Rats subjected to surgical manipulation were anesthe- tized with ketamine hydrochloride (10 mg/lOO g im). The dorsal muscles of both lower legs were surgically exposed. After the hamstring group had been longitudinally cut the Achilles tendon was gently separated from the gas- trocnemius muscle and then dissected. To isolate the plantaris muscle, the gastrocnemius and soleus muscles were almost completely removed except for small por- tions near the proximal tendons. Following suturing, the animals were returned to their cages, housed in groups of four, and were studied 9-12 wk after surgery. Sham operations were not routinely performed, since we have previously observed no difference between sham and normal muscles with regard to the variables studied (unpublished observations).
Myofibril and myosin ATPase. Skeletal muscle my- ofibrils were prepared from fresh muscle samples as described previously (2, 3). The final pellet was sus- pended in 150 mM KC1 and 20 mM tris(hydroxymeth- yl)aminomethane hydrochloride (Tris-HCl) (pH 7.4), and the protein was adjusted to 6 mg/ml as determined by the biuret method (12).
METHODS
Myofibril ATPase was measured at 25°C in a 2-ml system containing 1 mM MgS04, 1 mM ATP, 2 mM, sodium azide, 1 mM ethyleneglycol-bis( P-aminoethyl- ether)-N, N’-tetraacetic acid (EGTA), 30 mM KCl, 30 mM Trig-HCI, CaC12 (0.01-2.09 mM), and 0.3 mg/ml protein (pH 7.0). Activity was determined by measuring inorganic phosphate (Pi) release over a 2-min period using the technique of Fiske and SubbaRow (10). In this system, Ca2+ was buffered with EGTA so that free Ca2’ was varied from a pCa2’ of 8.5 to approximately 3.0 (28). Although pCa2+ was varied throughout the physiological range thought to exist in intact muscle, the assay was designed to maintain pMg2+, pMg-ATP, and ionic strength at near constant level (28).
Experimental protocol and surgical procedure. Fe- Myosin was prepared from fresh muscle according to male rats of a Wistar strain initially weighing approxi- the method of Shiverick et al. (27). This procedure in-
OhSI- i’567~‘82/oooO-oooO$O1.25 Copyright 0 1982 the American Physiological Society 467
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from
468 BALDWIN ET AL.
volved several precipitation-dissolution cycles without the use of column chromatography. The purity of the preparation was verified by sodium dodecyl sulfate-poly- acrylamide gel electrophoresis (20). This technique also was used to assess the light-chain composition of the
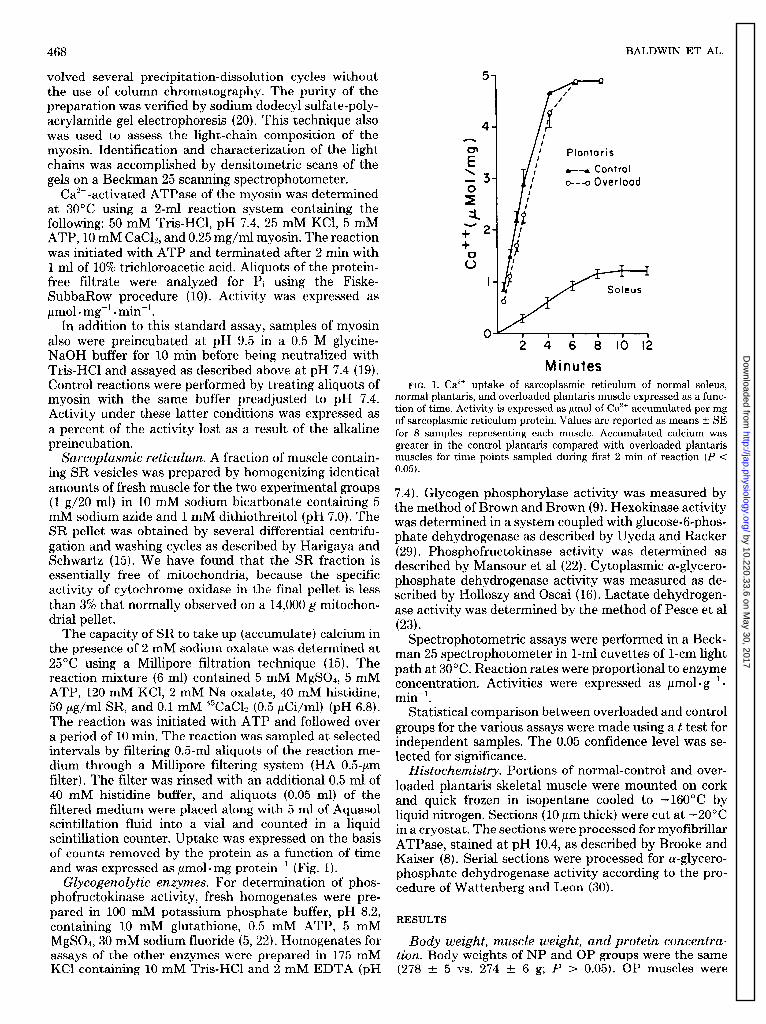
5 I0 / 0
myosin. Identification and characterization of the light // Plontoris chains was accomplished by densitometric scans of the gels on a Beckman 25 scanning spectrophotometer.
Ca”+-activated ATPase of the myosin was determined
4++ Control o--q Overlood
at 30°C using a 2-ml reaction system containing the following: 50 mM Tris-HCl, pH 7.4, 25 mM KCl, 5 mM ATP, 10 mM CaC12, and 0.25 mg/ml myosin. The reaction was initiated with ATP and terminated after 2 min with 1 ml of 10% trichloroacetic acid. Aliquots of the protein- free filtrate were analyzed for Pi using the Fiske- SubbaRow procedure (10). Activity was expressed as pm01 l mg-l . min?
In addition to this standard assay, samples of myosin also were preincubated at pH 9.5 in a 0.5 M glycine- I 1 1 1 I 1
NaOH buffer for 10 min before being neutralized with 2 4 6 8 IO 12
Tris-HCl and assayed as described above at pH 7.4 (19). Minutes Control reactions were performed by treating aliquots of FIG. 1. Ca” uptake of sarcoplasmic reticulum of normal soleus,
myosin with the same buffer preadjusted to pH 7.4. normal plantaris, and overloaded plantaris muscle expressed as a func-
Activity under these latter conditions was expressed as tion of time. Activity is expressed as pmol of Ca2’ accumulated per mg
a percent of the activity lost as a result of the alkaline of sarcoplasmic reticulum protein. Values are reported as means t SE
- . for 8 samples representing each muscle. Accumulated calcium was preincu bation.
Sarcoplasmic reticulum. A fraction of muscle contain- greater in the control plantaris compared wi th overloaded plantaris muscles for time points sampled during first 2 min of reaction (P <
ing SR vesicles was prepared by homogenizing identical 0.05).
amounts of fresh muscle for the two experimental groups (1 g/20 ml) in 10 mM sodium bicarbonate containing 5
7.4). Glycogen phosphorylase activity was measured by
mM sodium azide and 1 mM dithiothreitol (pH 7.0). The the method of Brown and Brown (9). Hexokinase activity
SR pellet was obtained by several differential centrifu- was determined in a system coupled with glucose-6-phos-
gation and washing cycles as described by Harigaya and phate dehydrogenase as described by Uyeda and Racker
Schwartz (15). We have found that the SR fraction is (29). Phosphofructokinase activity was determined as
essentially free of mitochondria, because the specific described by Mansour et al (22). Cytoplasmic cx-glycero-
activity of cytochrome oxidase in the final pellet is less phosphate dehydrogenase activity was measured as de-
than 3% that normally observed on a 14,000 g mitochon- scribed by Holloszy and Oscai (16). Lactate dehydrogen-
drial pellet. ase activity was determined by the method of Pesce et al
The capacity of SR to take up (accumulate) calcium in (23)
the presence of 2 mM sodium oxalate was determined at Spectrophotometric assays were performed in a Beck-
25°C using a Millipore filtration technique (15). The man 25 spectrophotometer in l-ml cuvettes of l-cm light
reaction mixture (6 ml) contained 5 mM MgS04, 5 mM path at 30°C. Reaction rates were proportional to enzyme
ATP, 120 mM KCl, 2 mM Na oxalate, 40 mM histidine, concentration. Activities were expressed as pmol l g-’ l
50 pg/ml SR, and 0.1 mM *“CaC& (0.5 ,&i/ml) (pH 6.8). min-1
The reaction was initiated with ATP and followed over Statistical comparison between overloaded and control
a period of 10 min. The reaction was sampled at selected groups for the various assays were made using a t test for
intervals by filtering 0.5-ml aliquots of the reaction me- independent samples. The 0.05 confidence level was se-
dium through a Millipore filtering system (HA 0.5~pm lected for significance.
filter). The filter was rinsed with an additional 0.5 ml of Histochemistry. Portions of normal-control and over-
40 mM histidine buffer, and aliquots (0.05 ml) of the loaded plantaris skeletal muscle were mounted on cork
filtered medium were placed along with 5 ml of Aquasol and quick frozen in isopentane cooled to -160°C by
scintillation fluid into a vial and counted in a liquid liquid nitrogen. Sections (10 pm thick) were cut at -2O’C
scintillation counter. Uptake was expressed on the basis in a cryostat. The sections were processed for myofibrillar
of counts removed by the protein as a function of time ATPase, stained at pH 10.4, as described by Brooke and
and was expressed as ,umol*mg protein-’ (Fig. 1). Kaiser (8). Serial sections were processed for a-glycero-
GZycogenoZytic enzymes. For determination of phos- phosphate dehydrogenase activity according to the pro-
phofructokinase activity, fresh homogenates were pre- cedure of Wattenberg and Leon (30).
pared in 100 mM potassium phosphate buffer, pH 8.2, containing 10 mM glutathione, 0.5 mM ATP, 5 mM
RESULTS
MgSO+ 30 mM sodium fluoride (5,22). Homogenates for Body weight, muscle weight, and protein concentra- assays of the other enzymes were prepared in 175 mM tion. Body weights of NP and OP groups were the same KC1 containing 10 mM Tris-HCl and 2 mM EDTA (pH (278 t 5 vs. 274 t 6 g; P > 0.05). OP muscles were
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from
BIOCHEMICAL PROPERTIES OF OVERLOADED MUSCLE 469
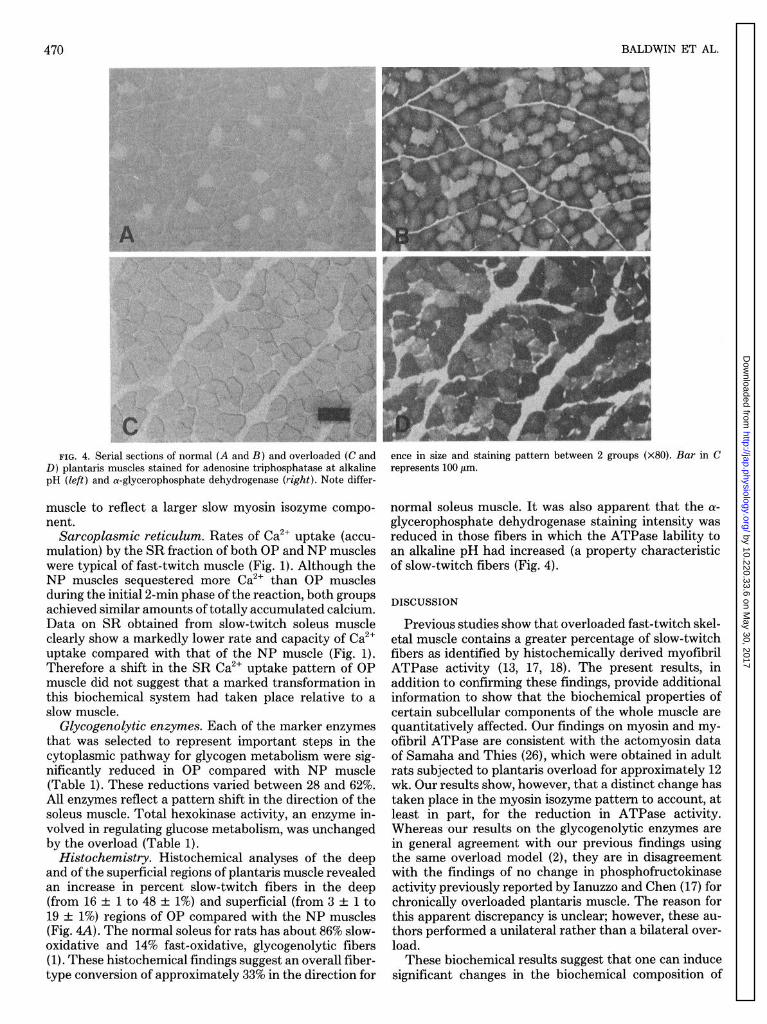
significantly larger than NP muscles (425 t 18 vs. 255 t 9 mg; P < 0.05). Total protein concentrations were similar between NP and OP muscles (195 t 8 vs. 202 t 15 mg/g; P > 0.05). Thus the increase in muscle weight is largely attributed to an increase in the protein content per muscle. This is also verified by the obviously larger fibers seen in OP compared with NP muscle (Fig. 4).
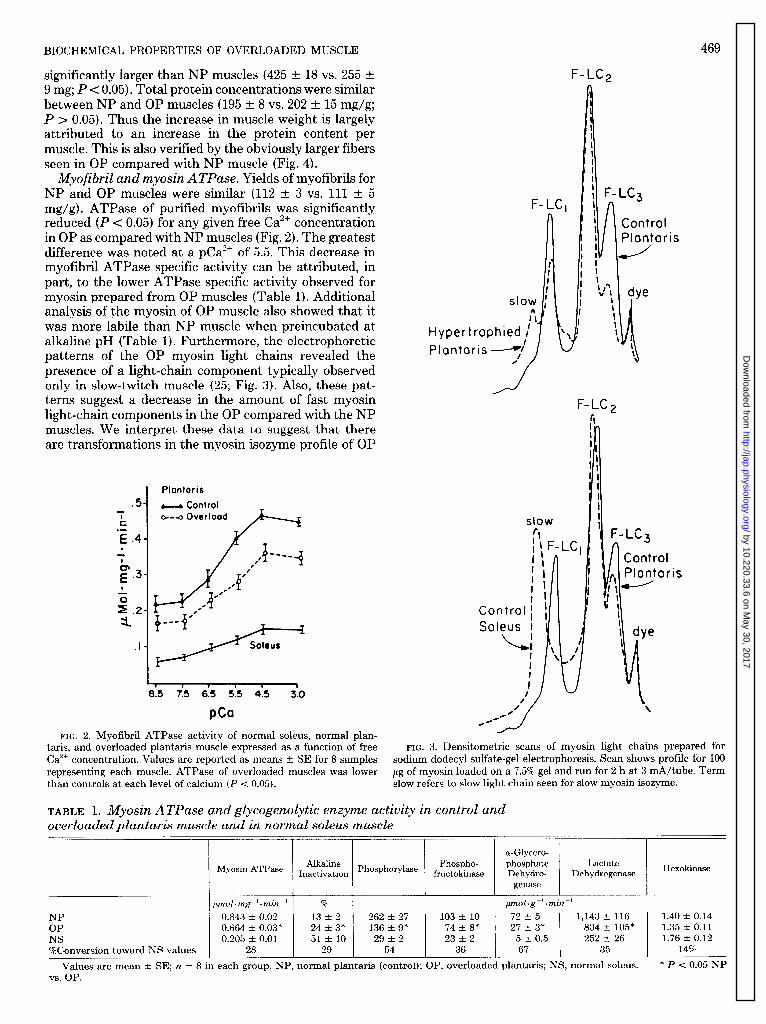
Myofibril and myosin ATPase. Yields of myofibrils for NP and OP muscles were similar (112 t 3 vs. 111 t 5 mg/g). ATPase of purified myofibrils was significantly reduced (P < 0.05) for any given free Ca”’ concentration in OP as compared with NP muscles (Fig. 2). The greatest difference was noted at a pCa2’ of 5.5. This decrease in myofibril ATPase specific activity can be attributed, in part, to the lower ATPase specific activity observed for myosin prepared from OP muscles (Table 1). Additional analysis of the myosin of OP muscle also showed that it was more labile than NP muscle when preincubated at alkaline pH (Table 1). Furthermore, the electrophoretic patterns of the OP myosin light chains revealed the presence of a light-chain component typically observed only in slow-twitch muscle (25; Fig. 3). Also, these pat- terns suggest a decrease in the amount of fast myosin light-chain components in the OP compared with the NP muscles. We interpret these data to suggest that there are transformations in the myosin isozyme profile of OP
.5 - 1 k .- E .4 . -i 1 r.3 . 1
Plontoris
- Control 01-e Ovcrlood
I 1 1 I v 1
8.5 7.5 6.5 5.5 4.5 3.0
PCQ
FIG. 2. Myofibril ATPase activity of normal soleus, normal plan- taris, and overloaded plantaris muscle expressed as a function of free Ca2+ concentration. Values are reported as means t SE for 8 samples representing each muscle. ATPase of overloaded muscles was lower than controls at each level of calcium (P < 0.05).
F-LC,
F-LC,
is
,Ol
aris
FIG. 3. Densitometric scans of myosin light chains prepared for sodium dodecyl sulfate-gel electrophoresis. Scan shows profile for 100 pg of myosin loaded on a 7.5% gel and run for 2 h at 3 mA/tube. Term slow refers to slow light chain seen for slow myosin isozyme.
TABLE 1. Myosin ATPase and gZycogenolytic enzyme activity in control and overloaded plantaris muscle and in normal soleus muscle
a-Glycero-
Myosin ATPase Alkaline Phospho- phosphate Lactate Inactivation Phosphorylase fructokinase Dehydro- Dehydrogenase
genase
pmol- mg-‘-min-’ % pmol-g-l.min-’
NP ' 0.843 t 0.02 13 t 2 262 t 27 103 t, 10 72 t 5 1,143 t 116 OP 0.664 t, 0.03* 24 t 3* 136 t, 9* 74 t8* 27 f; 3* 834 t 105* NS 0.205 t, 0.01 51t 10 29 t 2 23 t 2 5 t 0.5 252 t, 26 %Conversion toward NS values 28 29 54 36 67 35
Values are mean t SE; n = 8 in each group. NP, normal plantaris (control); OP, overloaded plantaris; NS, normal soleus. vs. OP.
Hexokinase
1.40 t 0.14 1.35 t, 0.11 1.76 -+ 0.12
14% *P< 0.05 NP
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from
470 BALDWIN ET AL.
FIG. 4. Serial sections of normal (A and B) and overloaded (C and D) plantaris muscles stained for adenosine triphosphatase at alkaline pH (left) and a-glycerophosphate dehydrogenase (right). Note differ-
muscle to reflect a larger slow myosin isozyme compo- nent.
Sarcoplasmic reticulum. Rates of Ca2+ uptake (accu- mulation) by the SR fraction of both OP and NP muscles were typical of fast-twitch muscle (Fig. 1). Although the NP muscles sequestered more Ca2+ than OP muscles during the initial 2-min phase of the reaction, both groups achieved similar amounts of totally accumulated calcium. Data on SR obtained from slow-twitch soleus muscle clearly show a markedly lower rate and capacity of Ca2+ uptake compared with that of the NP muscle (Fig. 1). Therefore a shift in the SR Ca2+ uptake pattern of OP muscle did not suggest that a marked transformation in this biochemical system had taken place relative to a slow muscle.
Glycogenolytic enzymes. Each of the marker enzymes that was selected to represent important steps in the cytoplasmic pathway for glycogen metabolism were sig- nificantly reduced in OP compared with NP muscle (Table 1). These reductions varied between 28 and 62%. All enzymes reflect a pattern shift in the direction of the soleus muscle. Total hexokinase activity, an enzyme in- volved in regulating glucose metabolism, was unchanged by the overload (Table 1).
Histochemistry. Histochemical analyses of the deep and of the superficial regions of plantaris muscle revealed an increase in percent slow-twitch fibers in the deep (from 16 & 1 to 48 & 1%) and superficial (from 3 +_ 1 to 19 f 1%) regions of OP compared with the NP muscles (Fig. 4A). The normal soleus for rats has about 86% slow- oxidative and 14% fast-oxidative, glycogenolytic fibers (1). These histochemical findings suggest an overall fiber- type conversion of approximately 33% in the direction for
ence in size and staining pattern between 2 groups (x80). Bar in C represents 100 pm.
normal soleus muscle. It was also apparent that the (Y- glycerophosphate dehydrogenase staining intensity was reduced in those fibers in which the ATPase lability to an alkaline pH had increased (a property characteristic of slow-twitch fibers (Fig. 4).
DISCUSSION
Previous studies show that overloaded fast-twitch skel- etal muscle contains a greater percentage of slow-twitch fibers as identified by histochemically derived myofibril ATPase activity (13, 17, 18). The present results, in addition to confirming these findings, provide additional information to show that the biochemical properties of certain subcellular components of the whole muscle are quantitatively affected. Our findings on myosin and my- ofibril ATPase are consistent with the actomyosin data of Samaha and Thies (26), which were obtained in adult rata subjected to plantaris overload for approximately 12 wk. Our results show, however, that a distinct change has taken place in the myosin isozyme pattern to account, at least in part, for the reduction in ATPase activity. Whereas our results on the glycogenolytic enzymes are in general agreement with our previous findings using the same overload model (2), they are in disagreement with the findings of no change in phosphofructokinase activity previously reported by Ianuzzo and Chen (17) for chronically overloaded plantaris muscle. The reason for this apparent discrepancy is unclear; however, these au- thors performed a unilateral rather than a bilateral over- load.
These biochemical results suggest that one can induce significant changes in the biochemical composition of
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from

BIOCHEMICAL PROPERTIES OF OVERLOADED MUSCLE 471
skeletal muscle while subjecting “intac t” muscle to an overload. The data on myosin, including ATPase specific activity, alkaline lability, light-chain electrophoretic pat- terns, and histochemical ATPase staining, provide evi- dence to suggest that a different isozyme is synthesized in some OP muscle fibers. This change in the biochemical expression of myosin is also manifest when the myofibril protein complex was evaluated for ATPase activity under physiological concentrations of calcium (Fig. 1).
We proposed previously that there is a relationship between glycogenolytic enzyme levels and the specific activity of actomyosin ATPase in different types of ro- dent muscle (6). The present findings on overloaded muscle support this concept in that these two systems maintained the normal relationship; i.e., in fibers in which myosin ATPase staining was reduced so was cx-glycero- phosphate dehydrogenase (Fig. 4). The significance of this relationship, although unclear at present, suggests that there may be some functional advantage to match the maximal rate of glycogen degradation with the max- imal rate of ATP degradation during certain conditions of intense brief muscular activity. On the other hand, previous findings with developing (4) and conditioned (3) skeletal muscle suggest that the respiratory capacity of skeletal muscle fiber types is regulated independently of glycogenolytic enzyme activity and myofibril ATPase. In support of this observation, we have recently reported that the oxidative capacity for the substrates pyruvate and palmitate in OP muscle is not markedly affected by the same conditions of overload as used in the present study (5).
REFERENCES
1. ARIANO, M. A., R. B. ARMSTRONG, AND V. R. EDGERTON. Hindlimb muscle fiber populations of five mammals. J. Histochem. Cytochem. 21: 51-55, 1973.
2. BALDWIN, K. M., W. G. CHEADLE, 0. M. MARTINEZ, AND D. A. COOKE. Effect of functional overload on enzyme levels in different types of skeletal muscle. J. Appl. Physiol.: Respirat. Environ. Exercise Physiol. 42: 312-317, 1977.
3. BALDWIN, K. M., D. A. COOKE, AND W. G. CHEADLE. Time course adaptations in cardiac and skeletal muscle to different running programs. J. Appl. Physiol.: Respirat. Environ. Exercise Physiol. 42: 267-272, 1977.
4. BALDWIN, K. M., A. M. HOOKER, P. J. CAMPBELL, AND R. E. LEWIS. Enzyme changes in neonatal skeletal muscle: effect of thyroid deficiency. Am. J. Physiol. 235 (Cell Physiol. 4): C97-C102, 1978.
5. BALDWIN, K. M., V. VALDEZ, L. F. SCHRADER, AND R. E. HERRICK. Effect of functional overload on substrate oxidation capacity of skeletal muscle. J. Appl. Physiol.: Respirat. Environ. Exercise Physiol. 50: 1272-1276, 1981.
6. BALDWIN, K. M., W. W. WINDER, AND J. 0. HOLLOSZY. Adaptation of actomyosin ATPase in different types of muscle to endurance exercise. Am. J. Physiol. 229: 422-426, 1975.
7. BALDWIN, K. M., W. W. WINDER, R. L. TERJUNG, AND J. 0. HOLLOSZY. Glycolytic enzymes in different types of skeletal muscle: adaptation to exercise. Am. J. Physiol. 225: 962-966, 1973.
8. BROOKE, M. H., AND K. K. KAISER. Muscle fiber types: how many and what kind? Arch. Neurol. 23: 369-374,1974.
9. BROWN, B. I., AND D. H. BROWN. Subcellular distribution of en- zymes in type II glycogenosis and the occurrence of an oligo-a-1,4- glucan glucohydrolase in human tissues. Biochim. Biophys. Acta 110: 124-133, 1965.
10. FISKE, C. H., AND Y. J. SUBBAROW. The calorimetric determination of phosphorus. J. Biol. Chem. 66: 375-400, 1925.
11. FITTS, R. H., W. W. WINDER, M. H. BROOKE, K. K. KAISER, AND J. 0. HOLLOSZY. Contractile, biochemical, and histochemical prop-
Recently Fitts et al. (11) exposed rodents to a hyper- thyroid state and found that SR calcium uptake capacity of slow-twitch soleus muscle was not regulated in parallel with ATPase of the contractile machinery. They showed that only the former was influenced by their experimental conditions. Although we have used different experimen- tal conditions and have focused on fast-twitch muscle, our data also suggest that the SR and the contractile machinery that controls speed of shortening in skeletal muscle can be affected independently of one another. In agreement with this differential response to overload by these two subcellular components, Roy et al. (24), using the same overload protocol as used for these experiments, found the maximum velocity of fiber shortening was reduced by over 55% in OP compared with NP muscle, whereas the isometric contraction time was prolonged in OP muscle by only approximately 12%. These biochem- ical results, combined with the physiological data (24)) are consistent with the physiological idea that the SR and myofibrillar components can regulate somewhat in- dependently different aspects of the contractile process.
In conclusion, we have provided some biochemical and histochemical evidence to suggest that skeletal muscle has the capacity to adjust its biochemical-physiological properties in response to an overload that increases mus- cle mass. The specific factors that regulate this plasticity under these conditions remain to be elucidated.
This research AM-21406.
was supported by National Institute of Health Grant
Received 10 April 1981; accepted in final form 15 September 1981.
erties of thyrotoxic rat soleus muscle. Am. J. Physiol. 238 (Cell Physiol. 7): C15-C20, 1980.
12. GORNALL, A. G., C. J. BARDAWILL, AND M. M. DAVID. Determi- nation of serum proteins by means of the biuret reaction. J. Biol. Chem. 177: 751-756, 1949.
13. GUTH, L., AND H. YELLIN. The dynamic nature of the so-called “fiber types” of mammalian skeletal muscle. Exp. Neurol. 31: 277- 300, 1971.
14. GUTMANN, E., S. SCHIAFFINO, AND V. HANZLIKOVA. Mechanism of compensatory hypertrophy in skeletal muscle of the rat. Exp. Neurol. 31: 451-464, 1971.
15. HARIGAYA, S., AND A. SCHWARTZ. Rate of calcium binding and uptake in normal animal and failing human cardiac muscle: mem- brane vesicles (relaxing system) and mitochondria. Circ. Res. 25: 782-794, 1969.
16. HOLLOSZY, J. O., AND L. B. OSCAI. Effect of exercise on a-glycero- phosphate dehydrogenase activity of skeletal muscle. Arch. Bio- them. Biophys. 130: 653-656, 1969.
17. IANUZZO, C. D., AND V. CHEN. Metabolic character of hypertro- phied rat muscle. J. Appl. Physiol.: Respirat. Environ. Exercise Physiol. 46: 738-742, 1979.
18. IANUZZO, C. D., P. D. GOLLNICK, AND R. B. ARMSTRONG. Compen- satory adaptations of skeletal muscle fiber types to a long-term functional overload. Life Sci. 19: 1517-1524, 1976.
19. JABLECKI, C., AND C. KAUFMAN. Myosin adenosine triphosphatase activity during work-induced growth of slow and fast skeletal mus- cle in the normal rat. J. Biol. Chem. 248: 1056-1062, 1973.
20. LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage Td. Nature London 227: 680-685,197O.
21. MACKOVA, E., AND P. HNIK. Compensatory muscle hypertrophy induced by tenotomy of synergists is not true working hypertrophy. Physiol. Bohemoslov. 22: 43-47, 1973.
22. MANSOUR, T. E., N. WAKID, AND H. M. SPROUSE. Studies on heart phosphofructokinase: purification, crystallization and properties of sheep heart phosphofructokinase. J. Biol. Chem. 241: 1512-1521,
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from

472 BALDWIN ET AL.
of cardiac mvosin: annlication to hynertronhied mvocardium. Biochim. Bioihys. A&393: 124-133, 1975. - ”
28. TIBBITS. G. F.. R. J. BARNARD. K. M. BALDWIN. N. CUCAW. AND
1966. 23. PESCE, A., R. H. MCKAY, F. STOLZENBACH, R. D. CAHN, AND N.
D. KAPLAN. Comnarative enzvmoloav of LDH. J. Biol. Chem. 239: 1753-1761, 1964. _
-”
24. ROY, R. R., I. D. MEADOWS, K. M. BALDWIN, AND V. R. EDGERTON. Functional significance of compensatory overloaded rat fast muscle. J. Appl. Physiol.: Respirat. Environ. Exercise Physiol. 52: 473- 478, 1982.
25. SAMAHA, F. J., L. GUTH, AND R. W. ALBERS. Differences between slow and fast muscle myosin. J. Biol. Chem. 24: 219-224, 1970.
26. SAMAHA, F. J., AND W. H. THIES. Actomyosin changes in muscles with altered function. Exp. Neurol. 51: 310-315, 1976.
27. SHIVERICK, K. T., L. L. THOMAS, AND N. R. ALPERT. Purification
N. K. ROBERTS. Influence of ‘exercise on excitation-contrsction coupling in rat myocardium. Am. J. Physiol. 240 (Heart Circ. Physiol. 9): H472-H480, 1981.
29. UYEDA, F., AND E. RACKER. Regulatory mechanisms in carbohy- drate metabolism. VII. hexokinase and phosphofructokinase. J. Biol. Chem. 240: 4682-4688,1965.
30. WATTENBERG, L. W., AND J. L. LEONG. Effects of coenzyme QlO and menadione on succinate dehydrogenase activity as measured by tetrazolium salt reduction. J. Histochem. Cytochem. 8: 296-303, 1960.
by 10.220.33.6 on May 30, 2017
http://jap.physiology.org/D
ownloaded from





























