Embed Size (px)
Citation preview

Exp. Eye Res. (1997) 64, 1037–1041
Biochemical differences between Three Subcell-lines Derived
from SV40-Transformed Hamster Lens Cells
HANS BLOEMENDAL*, JACQUELINE H. ENZLIN, ANKE A. F. VAN RIJK
HANS J. JANSEN
Department of Biochemistry, University of Nijmegen, P.O. Box 9101, 6500 HB Nijmegen,
The Netherlands
(Received Oxford 19 August 1996 and accepted in revised form 12 February 1997)
Clones derived from SV40-transformed hamster lens cells have at least three different stable morphologies.Biochemical differences between the three cell types that become detectable after transfection of the αA-crystallin gene do exist at the level of αB-crystallin and small heat shock protein (HSP27) expression.Furthermore one cell type is capable of alternative splicing of the hamster αA-crystallin gene, whereasanother one cannot express αAIns-crystallin. # 1997 Academic Press Limited
Key words : SV40-transformed hamster lens cells ; αA-crystallin gene transfection; HSP27-expression;alternative splicing; αAIns-crystallin.
1. Introduction
More than 15 years ago we established a cell line
derived from SV40-transformed hamster epithelial
lens cells (Hale cells) which appeared to be a good
source of actin, vimentin and their corresponding
mRNAs (Bloemendal et al., 1980). This cell line has
been used routinely in our laboratory. By chance we
observed that those Hale cells, upon subculturing,
consisted of at least three different morphological
types. The individual types have been designated t1, t2
and t3 and, after subsequent passages, the three cell
types kept identical morphology.
Morphological changes of long-maintained cell lines
have frequently been reported, particularly in response
to external factors. It has also been described that
rodent cells transformed by SV40 show many alter-
ations in their growth pattern in vitro, including
morphological changes (Risser and Pollack, 1974).
We wondered whether the different morphological
features detected in our cell system were paralleled by
any difference at the molecular level. As a model we
verified whether or not there was a difference in the
capability of the cells to express the specific eye lens
protein subunits : αA- and αB-crystallin. Since, as
reported earlier (Bloemendal et al., 1980), the original
Hale cells had ceased to synthesize crystallins we
transfected the cells with a DNA construct comprising
the hamster vimentin promoter linked to the coding
region of the αA-crystallin gene (pVim-αA-crystallin)
and analysed the expression of αA- and αB-crystallin
and HSP27 without or after heat shock.
* Author for correspondence.
2. Materials and Methods
Cell Lines
Hale cells were grown as monolayers on Dulbecco’s
modified Eagle’s medium supplemented with 10%
fetal calf serum, glutamin (2 m), penicillin G
(100 units ml−"), streptomycin (100 µg ml−"), and
pyruvate (1 m). To clone different morphological
types, Hale cells were sown at a density of 50
cells}100 cm−# and single colonies were isolated and
subcultured as separate cell lines.
Transfection
The cell lines were transfected with 40 µg pVim-αA-
crystallin and 4 µg pSV#neo using the calcium-
phosphate precipitation method, essentially as de-
scribed previously (Pieper et al., 1987; Raats et al.,
1990). Twenty-four hours after transfection the cells
were transferred to a medium containing 0±4 mg ml−"
geneticin (Life Technologies Inc., Grand Island, NY,
U.S.A.). Expression of the transgene was verified by
immunofluorescence (Krimpenfort et al., 1988) using
an anti-αA-crystallin antibody (not shown).
Constructs
pVim-αA-crystallin contains a 3±1 kb BamHI-PstI
promoter fragment of the hamster vimentin gene
cloned in front of a 4±5 kb HindIII-KpnI fragment
which contains the hamster αA-crystallin gene.
pSV#neo contains the SV40 promoter in front of the
neomycine resistance gene.
Heat Shock and Harvesting of Hale Cells
Cells of the three types (both untransfected and
transfected) including a sample of the original Hale
0014–4835}97}06103705 $25.00}0}ey970294 # 1997 Academic Press Limited
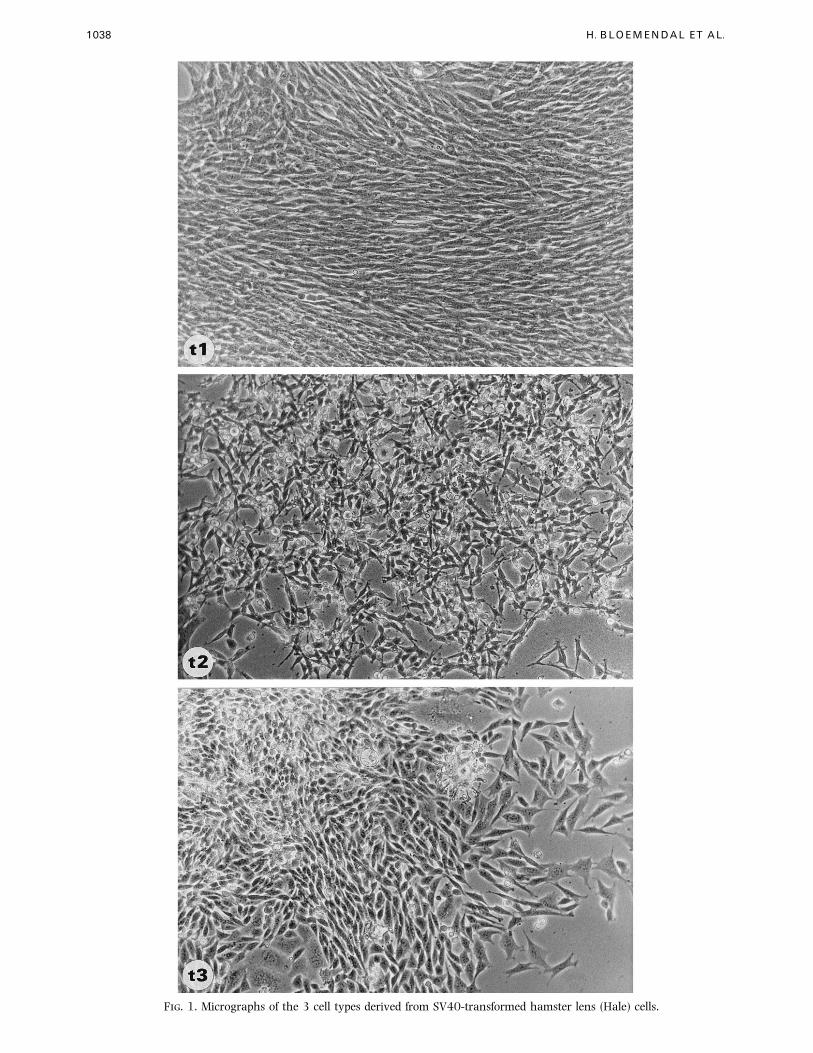
1038 H. BLOEMENDAL ET AL.
F. 1. Micrographs of the 3 cell types derived from SV40-transformed hamster lens (Hale) cells.

BIOCHEMICAL DIFFERENCES BETWEEN THREE SUBCELL-LINES 1039
cells were subjected to a heat shock in a thermostatted
water-bath at 42±5°C for 1 hr. Thereafter the cells
were kept at 37°C for about 24 hr. The medium was
removed by aspiration and the cells were washed
twice with phosphate-buffered saline (PBS) and supple-
mented with 100 µl of a solution containing 20%
glycerol, 6% sodium dodecyl-sulphate (SDS) and
0±12 Tris–HCl at pH 6±8. The cells were scraped
from the bottom of the dishes and transferred to an
Eppendorf centrifuge tube and spun. DNA was broken
by shearing.
Protein Estimation
The protein content of samples was estimated with
the aid of the BCA protein reagent (Pierce, Illinois,
U.S.A.).
PAGE and Blotting
PAGE and blotting of Hale protein extracts was
performed as described previously (Laemmli and Faore,
1973; Broers et al., 1986).
3. Results and Discussion
The morphological appearance of the three different
clones derived from SV40-transformed hamster lens
cells is shown in Fig. 1 (A–C). Hale t3 cells look rather
similar when compared to the original cell line. On the
other hand the most obvious difference exists between
Hale t1 and Hale t2. Whereas the former cell type
reveals contact inhibition the latter cell type has a
strong tendency to grow in layers.
The cells were stably transfected with the construct
pVim-αA-crystallin. Moreover part of the transfected
cells were subjected to a heat shock. All cell extracts
were analysed by one-dimensional SDS-PAGE followed
by blotting of the gel slabs with anti-αA- and anti-αB-
crystallin [Fig. 2(A) and (B)].
Several conclusions can be drawn from the results
depicted in this figure. Firstly, in Hale t1 cells
expression of αA-crystallin is not detectable (lanes 5
and 6), whereas transfection of pVim-αA-crystallin
clearly results in expression of this α-crystallin subunit
(lanes 3 and 4). Secondly, there is a clear difference in
the extent of expression of αAIns-crystallin, one of the
products of alternative splicing of the αA-crystallin
gene in rodents (Cohen et al., 1978a; 1978b), which
is expressed in Hale t3 cells [Fig. 2(A), lanes 1 and 2,
arrow], but not or only to a negligible extent in Hale
t1 cells [Fig. 2(A), lanes 3 and 4]. Thirdly, heat shock
has no effect upon this expression (compare the
intensities of the αAIns-crystallin bands in lanes 1 and
2). On the other hand in αA-crystallin-transfected cells
heat shock enhances the expression of αB-crystallin
expression both in t3 [Fig. 2(B), lane 2] and in t1 cells
[Fig. 2(B), lane 4].
In Fig. 3 the effect of transfection, heat shock and
α 1 2 3 4 5 6kDa
986450
36
30
16
α 1 2 3 4 5 6kDa
986450
36
30
16
F. 2. Immunoblotting of expression products obtainedafter transfection of different Hale-derived cells with pVim-αA-crystallin. Panels (A) and (B) : (α) α-crystallin ; (1) Halet3 transfected ; (2) Hale t3 transfected and heat-shocked; (3)Hale t1 transfected ; (4) Hale t1 transfected and heat-shocked; (5) Hale t1 cells ; (6) Hale t1 cells heat-shocked.Panel (A) : staining with anti-αA-crystallin ; Panel (B) :staining with anti-αB-crystallin. Note : the marker α isobtained from calf lens α-crystallin and shows the migrationdifference with rodent α-crystallin observed earlier by DeJong, Zweers and Cohen (1978).
the combination thereof on αB-crystallin and HSP27
expression in the three cell types is compared. The
transfection per se presumably affects αB-crystallin
expression in Hale t3 cells [Fig. 3(A), lanes 5 and 7],
but not in Hale t1 and Hale t2 cells [Fig. 3(A), lanes 1
and 3 and Fig. 3(B), lanes 3 and 5]. After heat shock
the concentration of αB-crystallin is raised in trans-
fected Hale t3 cells [Fig. 3(A), lane 8]. Also in the
transfected Hale t1 cells the expression of αB-crystallin
then becomes evident [Fig. 3(A), lane 4]. In the
untransfected Hale t1 cells αB-crystallin expression is
negligible whether or not the cells have been subjected
to heat shock [Fig. 3(A), lanes 1 and 2]. In Hale t2 cells
even after transfection and heat shock there is no αB-
crystallin expression whatsoever [Fig. 3(B)]. This leads
to the assumption that, either the transfection of the
construct per se is a certain form of stress for some cell
types (here Hale t3) but not for others (here Hale t1),
or the expression of the αA-crystallin gene promotes
simultaneous expression of the αB-crystallin gene in
some cell types.
For comparison the blots shown in Fig. 3(A) and (B)
were also incubated with an antibody directed against
the small heat shock protein [HSP27] [Fig. 3(C) and
(D)]. HSP27 is expressed in untransfected Hale t1 and
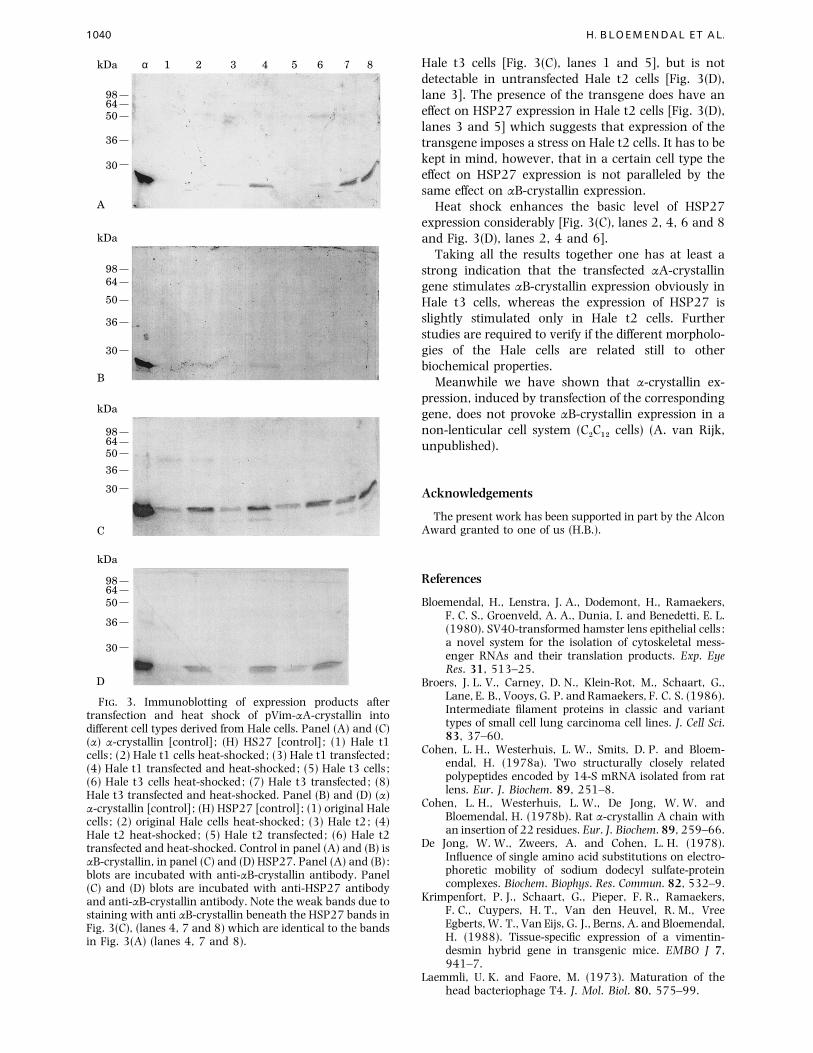
1040 H. BLOEMENDAL ET AL.
α 1 2 3 4 5 6kDa
986450
36
30
7
A
α 1 2 3 4 5 6kDa
9864
50
36
30
B
H 1 2 3 4 5 6kDa
986450
36
30
C
7 8
H 1 2 3 4 5 6kDa
986450
36
30
D
8
F. 3. Immunoblotting of expression products aftertransfection and heat shock of pVim-αA-crystallin intodifferent cell types derived from Hale cells. Panel (A) and (C)(α) α-crystallin [control] ; (H) HS27 [control] ; (1) Hale t1cells ; (2) Hale t1 cells heat-shocked; (3) Hale t1 transfected ;(4) Hale t1 transfected and heat-shocked; (5) Hale t3 cells ;(6) Hale t3 cells heat-shocked; (7) Hale t3 transfected ; (8)Hale t3 transfected and heat-shocked. Panel (B) and (D) (α)α-crystallin [control] ; (H) HSP27 [control] ; (1) original Halecells ; (2) original Hale cells heat-shocked; (3) Hale t2; (4)Hale t2 heat-shocked; (5) Hale t2 transfected ; (6) Hale t2transfected and heat-shocked. Control in panel (A) and (B) isαB-crystallin, in panel (C) and (D) HSP27. Panel (A) and (B) :blots are incubated with anti-αB-crystallin antibody. Panel(C) and (D) blots are incubated with anti-HSP27 antibodyand anti-αB-crystallin antibody. Note the weak bands due tostaining with anti αB-crystallin beneath the HSP27 bands inFig. 3(C), (lanes 4, 7 and 8) which are identical to the bandsin Fig. 3(A) (lanes 4, 7 and 8).
Hale t3 cells [Fig. 3(C), lanes 1 and 5], but is not
detectable in untransfected Hale t2 cells [Fig. 3(D),
lane 3]. The presence of the transgene does have an
effect on HSP27 expression in Hale t2 cells [Fig. 3(D),
lanes 3 and 5] which suggests that expression of the
transgene imposes a stress on Hale t2 cells. It has to be
kept in mind, however, that in a certain cell type the
effect on HSP27 expression is not paralleled by the
same effect on αB-crystallin expression.
Heat shock enhances the basic level of HSP27
expression considerably [Fig. 3(C), lanes 2, 4, 6 and 8
and Fig. 3(D), lanes 2, 4 and 6].
Taking all the results together one has at least a
strong indication that the transfected αA-crystallin
gene stimulates αB-crystallin expression obviously in
Hale t3 cells, whereas the expression of HSP27 is
slightly stimulated only in Hale t2 cells. Further
studies are required to verify if the different morpholo-
gies of the Hale cells are related still to other
biochemical properties.
Meanwhile we have shown that α-crystallin ex-
pression, induced by transfection of the corresponding
gene, does not provoke αB-crystallin expression in a
non-lenticular cell system (C#C"#
cells) (A. van Rijk,
unpublished).
Acknowledgements
The present work has been supported in part by the AlconAward granted to one of us (H.B.).
References
Bloemendal, H., Lenstra, J. A., Dodemont, H., Ramaekers,F. C. S., Groenveld, A. A., Dunia, I. and Benedetti, E. L.(1980). SV40-transformed hamster lens epithelial cells :a novel system for the isolation of cytoskeletal mess-enger RNAs and their translation products. Exp. EyeRes. 31, 513–25.
Broers, J. L. V., Carney, D. N., Klein-Rot, M., Schaart, G.,Lane, E. B., Vooys, G. P. and Ramaekers, F. C. S. (1986).Intermediate filament proteins in classic and varianttypes of small cell lung carcinoma cell lines. J. Cell Sci.83, 37–60.
Cohen, L. H., Westerhuis, L. W., Smits, D. P. and Bloem-endal, H. (1978a). Two structurally closely relatedpolypeptides encoded by 14-S mRNA isolated from ratlens. Eur. J. Biochem. 89, 251–8.
Cohen, L. H., Westerhuis, L. W., De Jong, W. W. andBloemendal, H. (1978b). Rat α-crystallin A chain withan insertion of 22 residues. Eur. J. Biochem. 89, 259–66.
De Jong, W. W., Zweers, A. and Cohen, L. H. (1978).Influence of single amino acid substitutions on electro-phoretic mobility of sodium dodecyl sulfate-proteincomplexes. Biochem. Biophys. Res. Commun. 82, 532–9.
Krimpenfort, P. J., Schaart, G., Pieper, F. R., Ramaekers,F. C., Cuypers, H. T., Van den Heuvel, R. M., VreeEgberts, W. T., Van Eijs, G. J., Berns, A. and Bloemendal,H. (1988). Tissue-specific expression of a vimentin-desmin hybrid gene in transgenic mice. EMBO J 7,941–7.
Laemmli, U. K. and Faore, M. (1973). Maturation of thehead bacteriophage T4. J. Mol. Biol. 80, 575–99.

BIOCHEMICAL DIFFERENCES BETWEEN THREE SUBCELL-LINES 1041
Pieper, F. R., Slobbe, R. L., Ramaekers, F. C. S., Cuypers,H. T. and Bloemendal, H. (1987). Upstream regions ofthe hamster desmin and vimentin genes regulateexpression during in vitro myogenesis. EMBO J. 12,3611–18.
Raats, J. M., Pieper, F. R., Vree Egberts, W. T. M., Verrijp,K. N., Ramaekers, F. C. S. and Bloemendal, H. (1990).
Assembly of amino-terminally deleted desmin in vimen-tin-free cells. J. Cell Biol. 111, 1971–85.
Risser, R. and Pollack, R. (1974). A nonselective analysis ofSV40 transformation of mouse 3T3 cells. Virology 59,474–89. [see also : Readings in mammalian cell culture,second edition (1981) (Ed. R. Pollack). Cold SpringHarbor Laboratory.














![Hamster[1] (3)rt](https://img.pdfslide.us/doc/110x75/5453f07faf7959856d8b512d/hamster1-3rt-5584af5997318.jpg)














