Embed Size (px)
Citation preview

Plant Physiol. (1980) 65, 838-8430032-0889/80/65/0838/06/$00.50/0
Biochemical and Cytological Analyses of RNA Synthesis inKinetin-treated Pea Root Parenchyma'
Received for publication September 17, 1979 and in revised form December 26, 1979
TERRY L. SHININGERDepartment of Biology, University of Utah, Salt Lake City, Utah 84112
ABSTRACT
Excised cortical parenchyma from the pea root (cv. Little Marvel)responds to kinetin/auxin treatment with an increased rate of RNAsynthesis well before reinmtiating DNA synthesis. Few cells synthesizeRNA in the 1st hour of culture. In the presence of kinetin/auxin, thenuclear labeling index increases 2.5-fold as compared to control cultures.The RNA synthesis response has an apparent lag period of 2-4 hours asshown by double label (13Hladenosine/I'4Cladenosine) experiments. Qual-itatively, the RNA synthesized at 4-6 hours sediments between 18S and5S. The RNA synthesized at 14-16 hours and 24-26 hours is primarilyribosomal RNA when kinetin is present. In the absence of kinetin, no clearpattern of RNA synthesis emerges.
The data are interpreted to mean that kinetin treatment elicits RNAsynthesis in a small proportion of the population initially and this mayinvolve messenger-like RNA. Later, more cells synthesize RNA and this isprimarily rRNA.
Cells of the pea root cortex become mitotically quiescent in thecourse of development in the root. When this tissue is excised,cytokinin and auxin are required for the cells to reinitiate repli-cation. In the excised pea root, cortical parenchyma DNA synthe-sis is reinitiated after 36-48 h of hormone treatment (16, 22).Mitosis and cytokinesis follow at 60 and 72 h, respectively (25).RNA synthesis, in contrast, increases within 9-12 h (21). Neitherquantitative nor qualitative changes in protein synthesis have beenobserved prior to resumed DNA or RNA synthesis (22; Shininger,in preparation). It is not known how cytokinin-auxin treatmentinteracts with the cells to reinitiate replication in these quiescentcells. Information on this problem is essential to any understand-ing of the mechanisms by which plant hormones regulate growthand development in the intact plant.The pea root cortex in contrast to suspension cultured cells is
an integrated, mitotically quiescent tissue and so there are possi-bilities for regulating replication at many levels. One possibility isthat within the cortex there are a few hormone-sensitive targetcells which regulate replication. These cells may respond to hor-mone treatment by initiating RNA or protein synthesis early,relative to the majority of the cortical cells. Such reactivationmight be detected autoradiographically in squash preparations.Alternatively, the reactivation of replication may be the result ofa general responsiveness of the cortical cells to cytokinin-auxintreatment. This system is also amenable to examining the actionof cytokinin on macromolecule synthesis.
Conclusions concerning the mode of action of cytokinin require
'This work supported by National Science Foundation Grant PCM 77-25 133.
(a) knowledge of the time lag between hormone application andthe earliest known response (RNA synthesis), (b) analysis of theRNA being synthesized, and (c) a measure of the cytokinin effecton protein synthesis.
MATERIALS AND METHODS
Segments (I mm) were excised at the 10-11th mm behind theroot tip of 66-h-old surface-sterilized, aseptically germinated seedsof Pisum sativum (cv. Little Marvel) as described (20). Thesesegments were cultured as such or converted to cortical explants(segments with the central vascular tissue punched out) and thencultured on standard S2M medium supplemented with kinetin (1,ug/ml) as indicated on filter paper wicks in the dark at 25 C (12,27).
After culturing (see figure legends) tissue was transferred tosmall Petri dishes containing filters dipped into culture mediumcontaining isotopically labeled precursor and the culture contin-ued for the times indicated. For DNA labeling, [3H]thymidine(New England Nuclear, 53 Ci/mmol, 5 ,uCi/ml) was used. ForRNA labeling, [2,8-3H]adenosine (New England Nuclear, 50 Ci/mmol, 50 ,uCi/ml) or ['4C]adenosine (New England Nuclear, 53mCi/mmol, 5 LCi/ml) was added to the medium. The labelingperiod was ended by transfer to ethanol acetic acid fixative (3:1,v/v) and the segments stored in 70% ethanol for the autoradi-ographic work. For biochemical work, the tissue was rinsed withice cold culture medium (3 times) containing unlabeled adenosine(I mM) then drained and frozen in an acetone/dry ice bath andstored at -78 C.RNA extraction was based on a procedure modified from that
of McKnight and Schimke (14). Frozen tissue was dropped intosterile SET buffer (10 mm Tris-HCl, pH 7.4, 5 ms EDTA, 1%SDS) containing heparin (I mg/ml) (Riker Laboratories) and 10pi/ml diethylpyrocarbonate in a ground glass homogenizer. Thetissue was ground with several strokes of the pestle until finelydispersed, and the homogenate then transferred to a 15-ml Corexcentrifuge tube and centrifuged (2 C, 3,000g max, 15 min) in aBeckman J21B centrifuge with the JA 20 angle rotor. The super-natant was extracted with the phenol chloroform method. Redis-tilled phenol, saturated with SET buffer (8 ml), and chloroform(8 ml) was added to the low speed supernatant (10 ml) and themixture shaken at room temperature for 30 min. The phases wereseparated by centrifugation (21 C, 23,700g max, 30 min). Theaqueous phase was extracted in the same way a second time andgenerally no precipitate was evident at the phase boundary afterthe second centrifugation. Nucleic acids were precipitated fromthe aqueous phase at -20 C ovemight by transferring the aqueousphase to a Corex tube containing an equal volume of 0.2 M NaClin 80%o ethanol. The precipitated nucleic acids were recovered bycentrifugation (2 C, 23,700g max, 30 min). The supernatant wasdiscarded, the tube drained, and then rinsed sequentially with 80%oethanol and anhydrous ethyl ether at -20 C. The samples werelyophylized and stored at -20 C.
838
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.
KINETIN AND RNA SYNTHESIS
The A260/A20 ratio was determined on an aliquot after redis-solving the sample in sterile water. The trichloroacetic acid-pre-cipitable incorporation was determined by transferring an aliquotalong with carrier RNA to a tube containing 0.5 ml 10%1o trichlo-roacetic acid on ice for 30 min. The precipitate was collected ona Millipore filter with vacuum and washed sequentially withtrichloroacetic acid (10lo) and 95% ethanol at 4 C. After drying,the filter was transferred to a scintillation vial with the addition of5 ml Aquasol 2 (New England Nuclear) and counting performedin a liquid scintillation spectrometer (Beckman LS 233).
Oligo(dT)-cellulose chromatography of the RNA was per-formed as described by Alt et al. (1).
Sucrose Gradients. Linear sucrose gradients (12 ml) were madeby mixing 5% sucrose and 29% sucrose (both in SET buffer, pH7.1) in a gradient maker. Samples (75 ,ug nucleic acid) werebrought to a volume of 50 ,ul in 10 mm Tris (pH 7.1) heated at 68C for I min, cooled with the addition of 5 AIl 5 M NaCl, and loadedonto the gradients. Centrifugation was performed in a BeckmanL550 (36,000 rpm, 21 C, 11 h) using the SW41 rotor. At the endof the experiment, the gradients were fractionated into 0.3-mlfractions using an ISCO density gradient fractionator by pumping30%o sucrose through the bottom of the tube and continuouslyrecording the A254 of the effluent.Water and Aquasol 2 (5 ml) were added to bring the samples
into solution and form a single clear phase for scintillation count-ing.
Denaturing Agarose Gel Electrophoresis. Samples (6 ,ug nucleicacid in 2-4 ,l H20) were mixed with an equal volume of E buffer(50 mM boric acid, 5 mm Na2B407. 10 H20, 10 mm Na2SO4, 100fLM Na2EDTA, pH 8.2), 10%o glycerol, and 0.04% bromphenol blue(2), containing 10 mm methyl mercury hydroxide (CH3HgOH)prepared from methyl mercury chloride by the method of Sneedand Maynard (23).
Gels were prepared by dissolving 2 gm agarose (Bio-Rad) in 50ml E buffer, cooling to 60 C, adding 0.5 ml 1 M CH3HgOH, andpouring a 12.5 x 13.5 x 3-mm horizontal slab gel. The sampleswere applied and electrophoresis conducted for 16 h at 15, roomtemperature, with recirculating E buffer. At the end of the run,the gels were stained for 30 min with ethidium bromide (19) (1ytg/ml) in 0.5 M ammonium acetate, rinsed in 0.5 M ammoniumacetate, examined in UV light, and photographed with a yellowfilter. For autoradiography, the gels were dried after the positionsof the stained bands had been marked in the gel with india ink.Autoradiography was performed by exposing Kodak SB5 filmwith the dried gel at room temperature.
Scintillation counting of gels was performed after excising alane of the wet gel, and slicing into 1.5-mm slices. The radioactivitywas eluted with 10 ml 5% protosol in Econofluor at 37 C for 18 h.These samples were cooled to room temperature and counted.
Autoradiography. Cortical explants were labeled with [3Hjaden-osine or [3HJthymidine fixed 24 h in ethanol/acetic acid (3:1, v/v)and stored in 70%o ethanol at 4 C. Squashes were prepared aftertreating the tissue with pectinase (20). These preparations weredipped in nuclear track emulsion (Kodak NTB 2) diluted 1:1 withwater, exposed 9-14 days and developed with freshly made amidoldeveloper at 4 C. The cells were stained with toluidine blue,rinsed, dried, and examined under oil immersion at 540x magni-fication.
RESULTS AND DISCUSSION
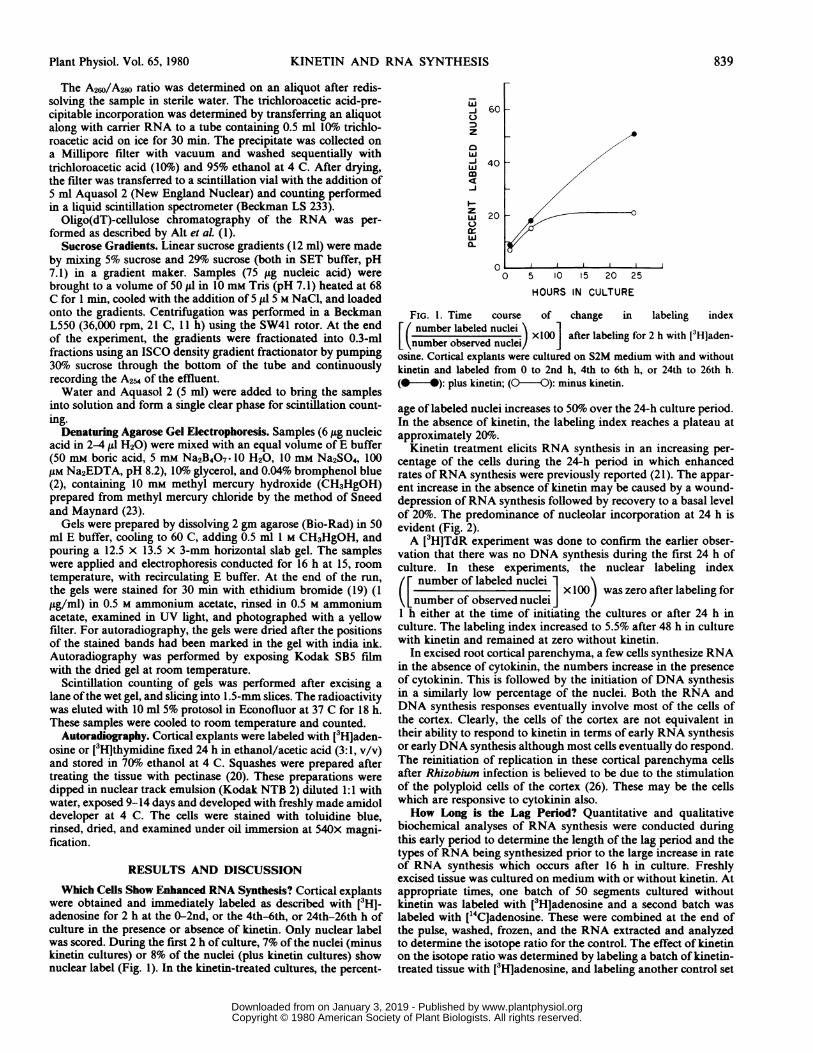
Which Cells Show Enhanced RNA Synthesis? Cortical explantswere obtained and immediately labeled as described with [3HJ-adenosine for 2 h at the 0-2nd, or the 4th-6th, or 24th-26th h ofculture in the presence or absence of kinetin. Only nuclear labelwas scored. During the first 2 h of culture, 7% of the nuclei (minuskinetin cultures) or 8% of the nuclei (plus kinetin cultures) shownuclear label (Fig. 1). In the kinetin-treated cultures, the percent-
.i
60
z0w-J 40
zw 20
a-
00 5 10 15 20 25
HOURS IN CULTURE
FIG. 1. Time course of change in labeling indexE( number labeled nuclei'\
number oabsered nuclei) xlOO after labeling for 2 h with [3Hjaden-number observed nuclei/osine. Cortical explants were cultured on S2M medium with and withoutkinetin and labeled from 0 to 2nd h, 4th to 6th h, or 24th to 26th h.(@ -): plus kinetin; (0--0): minus kinetin.
age of labeled nuclei increases to 50%1o over the 24-h culture period.In the absence of kinetin, the labeling index reaches a plateau atapproximately 20%o.
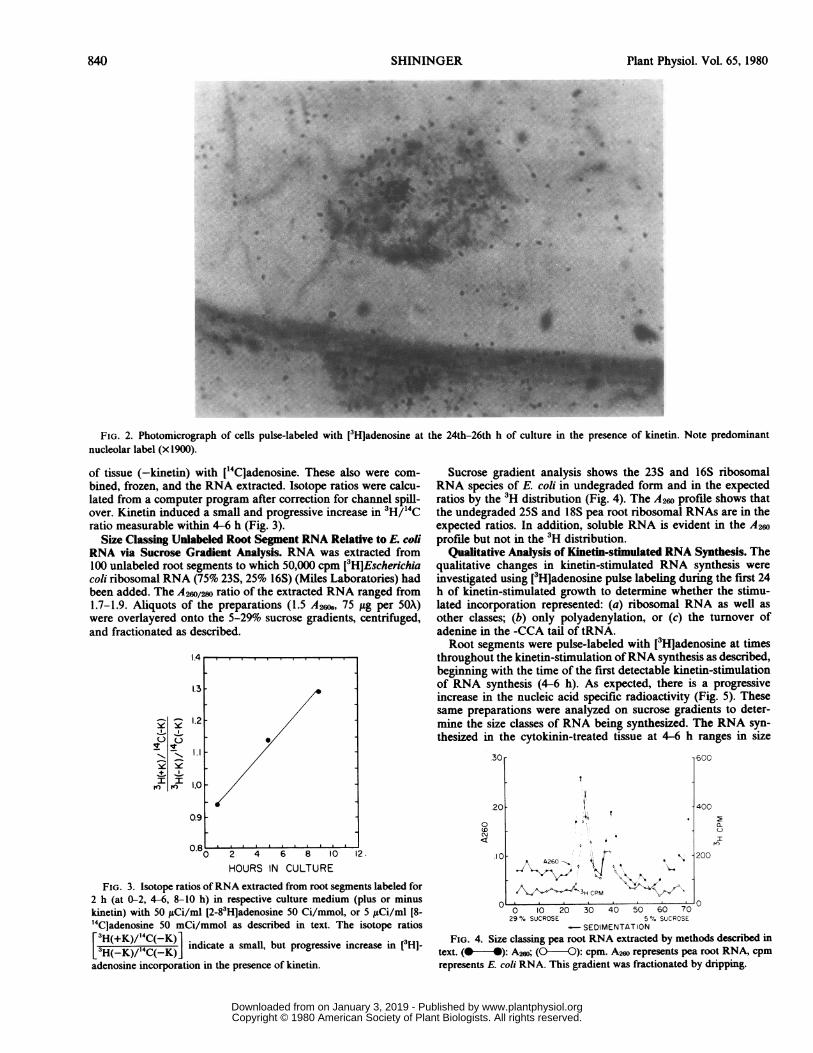
Kinetin treatment elicits RNA synthesis in an increasing per-centage of the cells during the 24-h period in which enhancedrates of RNA synthesis were previously reported (21). The appar-ent increase in the absence of kinetin may be caused by a wound-depression of RNA synthesis followed by recovery to a basal levelof 20%. The predominance of nucleolar incorporation at 24 h isevident (Fig. 2).A [3H]TdR experiment was done to confirm the earlier obser-
vation that there was no DNA synthesis during the first 24 h ofculture. In these experiments, the nuclear labeling index
number of labeled nuclei 1 \i x 100 was zero after labeling for
number of observed nucleiJJI h either at the time of initiating the cultures or after 24 h inculture. The labeling index increased to 5.5% after 48 h in culturewith kinetin and remained at zero without kinetin.
In excised root cortical parenchyma, a few cells synthesize RNAin the absence of cytokinin, the numbers increase in the presenceof cytokinin. This is followed by the initiation of DNA synthesisin a similarly low percentage of the nuclei. Both the RNA andDNA synthesis responses eventually involve most of the cells ofthe cortex. Clearly, the cells of the cortex are not equivalent intheir ability to respond to kinetin in terms of early RNA synthesisor early DNA synthesis although most cells eventually do respond.The reinitiation of replication in these cortical parenchyma cellsafter Rhizobium infection is believed to be due to the stimulationof the polyploid cells of the cortex (26). These may be the cellswhich are responsive to cytokinin also.How Long is the Lag Period? Quantitative and qualitative
biochemical analyses of RNA synthesis were conducted duringthis early period to determine the length of the lag period and thetypes of RNA being synthesized prior to the large increase in rateof RNA synthesis which occurs after 16 h in culture. Freshlyexcised tissue was cultured on medium with or without kinetin. Atappropriate times, one batch of 50 segments cultured withoutkinetin was labeled with [3H]adenosine and a second batch waslabeled with ['4Cladenosine. These were combined at the end ofthe pulse, washed, frozen, and the RNA extracted and analyzedto determine the isotope ratio for the control. The effect of kinetinon the isotope ratio was determined by labeling a batch of kinetin-treated tissue with [3H]adenosine, and labeling another control set
Plant Physiol. Vol. 65, 1980 839
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 65, 1980
FIG. 2. Photomicrograph of cells pulse-labeled with [3Hladenosine at the 24th-26th h of culture in the presence of kinetin. Note predominantnucleolar label (x 1900).
of tissue (-kinetin) with [1"Cladenosine. These also were com-bined, frozen, and the RNA extracted. Isotope ratios were calcu-lated from a computer program after correction for channel spill-over. Kinetin induced a small and progressive increase in 3H/'4Cratio measurable within 4-6 h (Fig. 3).
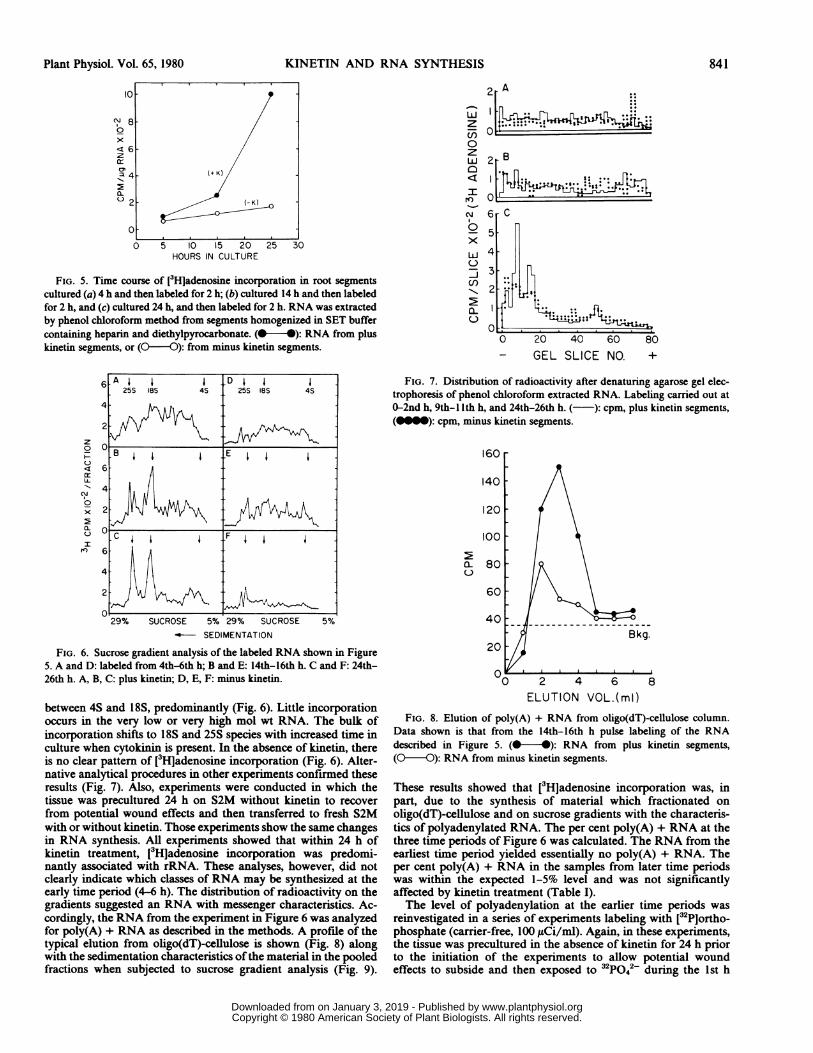
Size Classing Unlabeled Root Seginent RNA Relative to E. coliRNA via Sucrose Gradient Analysis. RNA was extracted from100 unlabeled root segments to which 50,000 cpm [3HjEscherichiacoli ribosomal RNA (75% 23S, 25% 16S) (Miles Laboratories) hadbeen added. The A26o/2ws ratio of the extracted RNA ranged from1.7-1.9. Aliquots of the preparations (1.5 A26N8, 75 i.g per 50X)were overlayered onto the 5-29% sucrose gradients, centrifuged,and fractionated as described.
_
. IT
IV Y
II1 14
1.4 _
1.3
1.2-
1.0
0.9
0.8A-.-0 2 4 6 8 loHOURS IN CULTURE
Sucrose gradient analysis shows the 23S and 16S ribosomalRNA species of E. coli in undegraded form and in the expectedratios by the 3H distribution (Fig. 4). The A260 profile shows thatthe undegraded 25S and 18S pea root ribosomal RNAs are in theexpected ratios. In addition, soluble RNA is evident in the A260profile but not in the 3H distribution.
Qualitative Analysis of Kinetin-stimulated RNA Synthesis. Thequalitative changes in kinetin-stimulated RNA synthesis wereinvestigated using [3HJadenosine pulse labeling during the first 24h of kinetin-stimulated growth to determine whether the stimu-lated incorporation represented: (a) ribosomal RNA as well asother classes; (b) only polyadenylation, or (c) the turnover ofadenine in the -CCA tail of tRNA.Root segments were pulse-labeled with [3Hladenosine at times
throughout the kinetin-stimulation ofRNA synthesis as described,beginning with the time of the first detectable kinetin-stimulationof RNA synthesis (4-6 h). As expected, there is a progressiveincrease in the nucleic acid specific radioactivity (Fig. 5). Thesesame preparations were analyzed on sucrose gradients to deter-mine the size classes of RNA being synthesized. The RNA syn-thesized in the cytokinin-treated tissue at 4-6 h ranges in size
30[
20[0CM
12.
FIG. 3. Isotope ratios ofRNA extracted from root segments labeled for2 h (at 0-2, 4-6, 8-10 h) in respective culture medium (plus or minuskinetin) with 50 [iCi/ml [2-83HJadenosine 50 Ci/mmol, or 5 ,uCi/ml [8-4Cladenosine 50 mCi/mmol as described in text. The isotope ratios
[H(-K)/'4C(-K) indicate a small, but progressive increase in [3HJ-adenosine incorporation in the presence of kinetin.
o-/13H CP
X^ g3 CPM '
600
400
I
20010
0 10 20 30 40 50 60 7029 % SUCROSE 5 % SUCRCSE
-SEDIMENTATIONFIG. 4. Size classing pea root RNA extracted by methods described in
text. (@-4): A26o; (Q-O ): cpm. A26o represents pea root RNA, cpmrepresents E. coli RNA. This gradient was fractionated by dripping.
a '
840 SHININGER
**
I
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.
KINETIN AND RNA SYNTHESIS
N
x
LUzcn0zLLJ0
N
cw
x
LJLUI
U)
0l~0
HOURS IN CULTURE
FIG. 5. Time course of [3Hjadenosine incorporation in root segmentscultured (a) 4 h and then labeled for 2 h; (b) cultured 14 h and then labeledfor 2 h, and (c) cultured 24 h, and then labeled for 2 h. RNA was extractedby phenol chloroform method from segments homogenized in SET buffercontaining heparin and diethylpyrocarbonate. (@ *): RNA from pluskinetin segments, or (O-0): from minus kinetin segments.
z0
LL-
c1
0x
a.
I
29% SUCROSE 5% 29% SUCROSE 5%-- SEDIMENTATION
2 A
...:*i:~~~~~~~~~~~:1,C.
2 -B
6r C
5
4
3
2
00 20 40 60 80- GEL SLICE NO. +
FIG. 7. Distribution of radioactivity after denaturing agarose gel elec-trophoresis of phenol chloroform extracted RNA. Labeling carried out at0-2nd h, 9th- IIth h, and 24th-26th h. ( ): cpm, plus kinetin segments,(mm): cpm, minus kinetin segments.
160
140
120
100
02a- 80
60
40
20FIG. 6. Sucrose gradient analysis of the labeled RNA shown in Figure5. A and D: labeled from 4th-6th h; B and E: 14th-16th h. C and F: 24th-26th h. A, B, C: plus kinetin; D, E, F: minus kinetin.
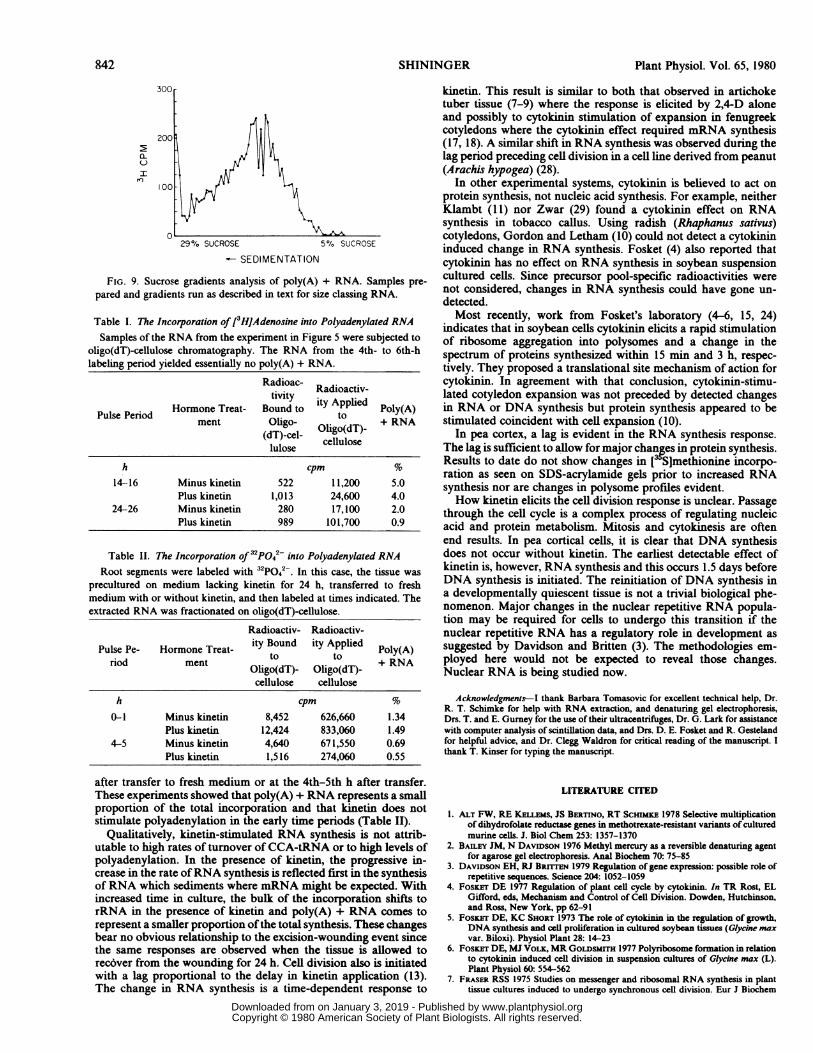
between 4S and 18S, predominantly (Fig. 6). Little incorporationoccurs in the very low or very high mol wt RNA. The bulk ofincorporation shifts to 18S and 25S species with increased time inculture when cytokinin is present. In the absence of kinetin, thereis no clear pattern of [3H]adenosine incorporation (Fig. 6). Alter-native analytical procedures in other experiments confirmed theseresults (Fig. 7). Also, experiments were conducted in which thetissue was precultured 24 h on S2M without kinetin to recoverfrom potential wound effects and then transferred to fresh S2Mwith or without kinetin. Those experiments show the same changesin RNA synthesis. All experiments showed that within 24 h ofkinetin treatment, [3HJadenosine incorporation was predomi-nantly associated with rRNA. These analyses, however, did notclearly indicate which classes of RNA may be synthesized at theearly time period (4-6 h). The distribution of radioactivity on thegradients suggested an RNA with messenger characteristics. Ac-cordingly, the RNA from the experiment in Figure 6 was analyzedfor poly(A) + RNA as described in the methods. A profile of thetypical elution from oligo(dT)-cellulose is shown (Fig. 8) alongwith the sedimentation characteristics ofthe material in the pooledfractions when subjected to sucrose gradient analysis (Fig. 9).
0 ) 2 4 6ELUTION VOL.(ml)
8
FIG. 8. Elution of poly(A) + RNA from oligo(dT)-cellulose column.Data shown is that from the 14th-16th h pulse labeling of the RNAdescribed in Figure 5. (-*): RNA from plus kinetin segments,(0-O): RNA from minus kinetin segments.
These results showed that [3H]adenosine incorporation was, inpart, due to the synthesis of material which fractionated onoligo(dT)-cellulose and on sucrose gradients with the characteris-tics of polyadenylated RNA. The per cent poly(A) + RNA at thethree time periods of Figure 6 was calculated. The RNA from theearliest time period yielded essentially no poly(A) + RNA. Theper cent poly(A) + RNA in the samples from later time periodswas within the expected 1-5% level and was not significantlyaffected by kinetin treatment (Table I).The level of polyadenylation at the earlier time periods was
reinvestigated in a series of experiments labeling with [32PJortho-phosphate (carrier-free, 100 ,uLCi/ml). Again, in these experiments,the tissue was precultured in the absence of kinetin for 24 h priorto the initiation of the experiments to allow potential woundeffects to subside and then exposed to 32P042- during the 1st h
;3
Plant Physiol. Vol. 65, 1980 841
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 65, 1980
00
100 j p
29% SUCROSE 5% SUCROSE
SEDIMENTATION
FIG. 9. Sucrose gradients analysis of poly(A) + RNA. Samples pre-
pared and gradients run as described in text for size classing RNA.
Table I. The Incorporation of [3HJAdenosine into Polyadenylated RNASamples of the RNA from the experiment in Figure 5 were subjected to
oligo(dT)-cellulose chromatography. The RNA from the 4th- to 6th-hlabeling period yielded essentially no poly(A) + RNA.
Radioac- .Radioc. Radioactiv-tivity iyApe
Pulse Period Hormone Treat- Bound to ity Applied Poly(A)ment Oligo- Olg(T-+ RNA
(dT)-cel- celluloselulose
h cpm %
14-16 Minus kinetin 522 11,200 5.0Plus kinetin 1,013 24,600 4.0
24-26 Minus kinetin 280 17,100 2.0Plus kinetin 989 101,700 0.9
Table II. The Incorporation of 32P042- into Polyadenylated RNARoot segments were labeled with 32P042 In this case, the tissue was
precultured on medium lacking kinetin for 24 h, transferred to freshmedium with or without kinetin, and then labeled at times indicated. Theextracted RNAwas fractionated on oligo(dT)-cellulose.
Radioactiv- Radioactiv-
Pulse Pe- Hormone Treat- ity Bound ity Applied Poly(A)riod ment tto + RNAOligo(dT)- Oligo(dT)-
cellulose cellulose
h cpm %
0-1 Minus kinetin 8,452 626,660 1.34Plus kinetin 12,424 833,060 1.49
4-5 Minus kinetin 4,640 671,550 0.69Plus kinetin 1,516 274,060 0.55
after transfer to fresh medium or at the 4th-5th h after transfer.These experiments showed that poly(A) + RNA represents a smallproportion of the total incorporation and that kinetin does notstimulate polyadenylation in the early time periods (Table II).
Qualitatively, kinetin-stimulated RNA synthesis is not attrib-utable to high rates of turnover of CCA-tRNA or to high levels ofpolyadenylation. In the presence of kinetin, the progressive in-crease in the rate ofRNA synthesis is reflected first in the synthesisof RNA which sediments where mRNA might be expected. Withincreased time in culture, the bulk of the incorporation shifts torRNA in the presence of kinetin and poly(A) + RNA comes torepresent a smaller proportion of the total synthesis. These changesbear no obvious relationship to the excision-wounding event sincethe same responses are observed when the tissue is allowed torecover from the wounding for 24 h. Cell division also is initiatedwith a lag proportional to the delay in kinetin application (13).The change in RNA synthesis is a time-dependent response to
kinetin. This result is similar to both that observed in artichoketuber tissue (7-9) where the response is elicited by 2,4-D aloneand possibly to cytokinin stimulation of expansion in fenugreekcotyledons where the cytokinin effect required mRNA synthesis(17, 18). A similar shift in RNA synthesis was observed during thelag period preceding cell division in a cell line derived from peanut(Arachis hypogea) (28).
In other experimental systems, cytokinin is believed to act onprotein synthesis, not nucleic acid synthesis. For example, neitherKlambt (11) nor Zwar (29) found a cytokinin effect on RNAsynthesis in tobacco callus. Using radish (Rhaphanus sativus)cotyledons, Gordon and Letham (10) could not detect a cytokinininduced change in RNA synthesis. Fosket (4) also reported thatcytokinin has no effect on RNA synthesis in soybean suspensioncultured cells. Since precursor pool-specific radioactivities werenot considered, changes in RNA synthesis could have gone un-detected.Most recently, work from Fosket's laboratory (4-6, 15, 24)
indicates that in soybean cells cytokinin elicits a rapid stimulationof ribosome aggregation into polysomes and a change in thespectrum of proteins synthesized within 15 min and 3 h, respec-tively. They proposed a translational site mechanism of action forcytokinin. In agreement with that conclusion, cytokinin-stimu-lated cotyledon expansion was not preceded by detected changesin RNA or DNA synthesis but protein synthesis appeared to bestimulated coincident with cell expansion (10).
In pea cortex, a lag is evident in the RNA synthesis response.The lag is sufficient to allow for major chan3Ees in protein synthesis.Results to date do not show changes in [ S]methionine incorpo-ration as seen on SDS-acrylamide gels prior to increased RNAsynthesis nor are changes in polysome profiles evident.How kinetin elicits the cell division response is unclear. Passage
through the cell cycle is a complex process of regulating nucleicacid and protein metabolism. Mitosis and cytokinesis are oftenend results. In pea cortical cells, it is clear that DNA synthesisdoes not occur without kinetin. The earliest detectable effect ofkinetin is, however, RNA synthesis and this occurs 1.5 days beforeDNA synthesis is initiated. The reinitiation of DNA synthesis ina developmentally quiescent tissue is not a trivial biological phe-nomenon. Major changes in the nuclear repetitive RNA popula-tion may be required for cells to undergo this transition if thenuclear repetitive RNA has a regulatory role in development assuggested by Davidson and Britten (3). The methodologies em-ployed here would not be expected to reveal those changes.Nuclear RNA is being studied now.
Acknowledgments-I thank Barbara Tomasovic for excellent technical help, Dr.R. T. Schimke for help with RNA extraction, and denaturing gel electrophoresis,Drs. T. and E. Gurney for the use of their ultracentrifuges, Dr. G. Lark for assistancewith computer analysis of scintillation data, and Drs. D. E. Fosket and R. Gestelandfor helpful advice, and Dr. Clegg Waldron for critical reading of the manuscript. Ithank T. Kinser for typing the manuscript.
LITERATURE CITED
1. ALT FW, RE KELEms, JS BERTINO, RT ScHimsE 1978 Selective multiplicationof dihydrofolate reductase genes in methotrexate-resistant variants of culturedmurine cells. J. Biol Chem 253: 1357-1370
2. BAILEY JM, N DAVIDSON 1976 Methyl mercury as a reversible denaturing agentfor agarose gel electrophoresis. Anal Biochem 70: 75-85
3. DAVIDSON EH, RJ BRITTEN 1979 Regulation of gene expression: possible role ofrepetitive sequences. Science 204: 1052-1059
4. FosKET DE 1977 Regulation of plant cell cycle by cytokinin. In TR Rost, ELGifford, eds, Mechanism and Control of Cell Division. Dowden, Hutchinson,and Ross, New York, pp 62-91
5. FossET DE, KC SHORT 1973 The role of cytokinin in the regulation of growth,DNA synthesis and cell proliferation in cultured soybean tissues (Glycein maxvar. Biloxi). Physiol Plant 28: 14-23
6. FosKrr DE, MJ VoLx, MR GOLDSMITH 1977 Polyribosome formation in relationto cytokinin induced cell division in suspension cultures of Glycine max (L).Plant Physiol 60: 554-562
7. FRASER RSS 1975 Studies on messenger and ribosomal RNA synthesis in planttissue cultures induced to undergo synchronous cell division. Eur J Biochem
842 SHININGER
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.

KINETIN AND RNA SYNTHESIS
50: 529-5378. FRASER RSS 1975 Synchronous cell division in cultured explants of Jerusalem
artichoke tubers: the effects of 5-fluorouracil on messenger RNA synthesis andthe induction of cell division. J Exp Bot 26: 555-568
9. FRAsER RSS, UE LEONING 1974 RNA synthesis during synchronous cell divisionin cultured explants of Jerusalem artichoke tuber. J Exp Bot 25: 847-859
10. GORDON ME, DS LsmAm 1975 Regulators of cell division in plant tissues. XII.Physiological aspects of cytokinin-induced radish cotyledon growth. Aust JPlant Physiol 2: 129-154
1 1. KLAMBT D 1974 Einfluss von Auxin and Cytokinin auf die RNA-Synthesis insterilem Tabakgewebe. Planta 118: 7-16
12. LIBBENGA K, JG ToRRLzY 1973 Hormone-induced endoreduplication prior tomitosis in cultured pea root cortex cells. Am J Bot 60: 293-299
13. MATrHYSSE AG, JG ToRREY 1967 Nutritional requirements for polyploid mitosesin cultured pea root segments. Physiol Plant 20: 661-672
14. McKNIsHT GS, RT SCHIMKE 1974 Ovalbumen messenger RNA: evidence thatthe initial product of transcription is the same size as polysomal ovalbumenmessenger. Proc Nat Acad Sci USA 71: 4327-4331
15. MUREN RC, DE FosKEr 1977 Cytokinin-mediated translational control ofproteinsynthesis in cultured cells of Glycine max. J Exp Bot 28: 775-784
16. PHILLIPS R, JG ToRREY 1973 DNA synthesis, cell division and specific cytodif-ferentiation in cultured pea root cortical explants. Dev Biol 31: 336-347
17. RIIVEN, AHGC 1972 Inhibitors and the dependence of kinetin induced expansionon RNA synthesis in fenugreek cotyledons. J Exp Bot 23: 1039-1049
18. RIJVEN AHGC, V PARUASH 1971 Action of kinetin on cotyledons of fenugreek.
843
Plant Physiol 47: 59-6419. SHARP PA, B SUGDEN, J SAMBROOK 1973 Selection of two restriction endonucle-
ase activities in Haemophilus parainfluenzae using analytical agarose-ethidiumbromide electrophoresis. Biochemistry 12: 3055-3063
20. SHININGER TL 1975 Is DNA synthesis required for the induction of differentia-tion in quiescent root cortical parenchyma? Dev Biol 45: 137-150
21. SHININGER TL, LD POLLEY 1977 Biochemical responses of pea root tissue tocytokinin: enhanced rates of RNA synthesis. Plant Physiol 59: 831-835
22. SIMPSON SF, JG ToRREY 1977 Hormonal control of deoxyribonucleic acid andprotein synthesis in pea root cortical explants. Plant Physiol 59: 4-9
23. SNEED MC, JL MAYNARD 1922 The preparation of methylmercuric acetate andthe isolation of methylmercuric hydroxide. J Am Chem Soc 44: 2942-2947
24. TEPFER DA, DE FosICET 1976 Cytokinin-stimulated polysome formation incultured soybean cells. Plant Physiol 57: S-75
25. ToRREY JG 1961 Kinetin as a trigger for mitosis in mature endomitotic plantcells. Exp Cell Res 23: 281-299
26. ToRREY JG 1976 Root hormones and plant growth. Annu Rev Plant Physiol 27:435-459
27. ToRREY JG, DE FosKET 1970 Cell division in relation to cytodifferentiation incultured pea root segments. Am J Bot 57: 1072-1080
28. VERMA DPS, A MARCUS 1973 Regulation ofRNA synthesis in plant cell culture:delayed synthesis of ribosomal RNA during transition from the stationaryphase to active growth. Dev Biol 30: 104-114
29. ZWAR JA 1973 Effects of cytokinin on the nucleic acids of tobacco pith. J ExpBot 24: 701-7 10
Plant Physiol. Vol. 65, 1980
www.plantphysiol.orgon January 3, 2019 - Published by Downloaded from Copyright © 1980 American Society of Plant Biologists. All rights reserved.





























