Embed Size (px)
Citation preview

354 G. BERNOCCHI ET AL.THE JOURNAL OF EXPERIMENTAL ZOOLOGY 280:354–367 (1998)
© 1998 WILEY-LISS, INC.
Bioactive Peptides and SerotoninImmunocytochemistry in the Cerebral Gangliaof Hibernating Helix aspersa
G. BERNOCCHI,* C. VIGNOLA, E. SCHERINI, D. NECCHI, AND M.B. PISUDipartimento di Biologia Animale, Centro di Studio per l�Istochimica delC.N.R., Universita´ degli Studi di Pavia, I-27100 Pavia, Italy
ABSTRACT The role of some neuromodulators and neurotransmitters in the functioning of mol-luskan cerebral neurons and in their metabolic changes during hibernation has been considered.The cerebral ganglion of mollusks is a center for the integration of different inputs from the sensoryareas of the head and for the generation of motor command impulses. During hibernation, animalsare deprived of many external sensory stimuli and do not have locomotion and feeding. Immunocy-tochemistry for bioactive peptides (BAPs), such as SP (Substance P), CCK8 (Cholecystokinin 8/Gastrin), CGRP (Calcitonin-Gene-Related Peptide) and ET (Endothelin), and serotonin was per-formed on cerebral ganglia of active and hibernating Helix aspersa. The distribution of theimmunopositivity was analyzed in different cell-containing areas (procerebrum, mesocerebrum,metacerebrum) and in the neuropiles. With all the antibodies raised against peptides, we observedthat only a few neurons, mainly of small and medium size, had immunopositivity during the periodof activity, the patterns of distribution being quite similar to those previously described in Helix orother gastropods. Fibers and varicosities with BAP immunopositivity were found in the procerebraland central neuropiles and sometimes around neurons. Serotonin-immunopositive neurons, includ-ing the giant neuron, were observed in the metacerebrum; numerous fibers and varicositiesimmunopositive for serotonin were present in the neuropile areas. In hibernating snails, the num-ber of fibers with BAP and serotonin immunopositivity decreased in several areas of the neuropiles.Moreover, an increased number of neurons of the metacerebrum (two- to four-fold) and mesocerebrum(8- to 28-fold) had BAP-like immunopositivity, and the intensity of the immunoreaction for serotoninof the metacerebral neurons was also higher than in the active snails. These results are discussed,taking into account two hypotheses. The first hypothesis assumes that the increased immunocy-tochemical staining was really linked to accumulation of BAPs and serotonin. The second hypothesisconsiders that the antibodies for BAPs recognized a preprotein, the synthesis of BAPs being com-pleted during the active period only. Both the hypotheses account for the co-occurrence and co-localization of two or ore peptides and serotonin and stress that the hibernation condition is ofinterest for studies on the actual function of single neurons in the cerebral ganglia. Finally, the dataare consistent with the changes recently found in other markers of the morphological and functionalactivity of neurons, demonstrating that the neuromodulation and the neurotransmission are slowedduring hibernation. J. Exp. Zool. 280:354�367, 1998. © 1998 Wiley-Liss, Inc.
*Correspondence to: Prof. Graziella Bernocchi, Dipartimento diBiologia Animale, Piazza Botta 10, I-27100 Pavia, Italy. E-mail:[email protected]
Received 20 February 1997; Accepted 24 October 1997
A wide spectrum of biological problems can beinvestigated using hibernation as a model. In fact,studies of hibernating animals are of particularinterest in learning about the physiology and me-tabolism of organisms (Hunter, ’64; Machin, ’75;Heller, ’79; Lyman et al., ’82), because they con-tribute to the understanding of some importantaspects of the structure and function of cells(Kolaeva et al., ’80; Bernocchi et al., ’86; Barni etal., ’87; Giacometti et al., ’89; Anderson et al., ’90;Barni and Bernocchi, ’91; Fenoglio et al., ’92;Malatesta et al., ’94, ’95; March and Reisman, ’95).
Our research was focused on studying the
mechanism by which neurons change their me-tabolism when there is suppression or reductionof some physiological activities of the organism(Hunter, ’64; Machin, ’75; Heller, ’79; Lyman etal., ’82). This may be considered a primitive formof cellular adaptability, similar to that used in avariety of situations associated with neuronalplasticity.

HIBERNATION AND NEURONAL ACTIVITY 355
A large number of biactive peptides (BAPs) hasbeen demonstrated in specific cell populations ofboth vertebrates and invertebrates. In particular,in the nervous system, the amino acid sequenceof specific peptides is often similar in differentphyla, suggesting that this class of molecules iswidely distributed and highly conserved (for a re-view, see Fasolo and Clairambault, ’91).
BAPs have a pervasive role in governing essen-tially all physiological activities, and there areprobably only a few biological processes in whichthey are not involved. In particular, in inverte-brates, the role of BAPs in the regulation of a va-riety of activities has been extensively investigatedin mollusks. They are involved in feeding (CCK8/Gastrin, FMRF-amide, see Engelhardt et al., ’82;Vigna et al., ’84; Elekes and Naessel, ’90; Sonettiet al., ’90; Cottrel, ’92; Cottrel et al., ’92), painperception, blood flow regulation, and vasodilation(CGRP, ET, SP, see Osborne et al., ’82; Osborne’84; Breimer et al., ’88; Giaid et al., ’91). In addi-tion, recent studies (Elekes et al., ’93, ’94) on thedistribution of enkephalins and leucokinin in thecentral nervous system of Helix point to an involve-ment of opioid substances in several behavioral phe-nomena (Gutierrez and Asai, ’91; Marchang et al.,’91; Elekes et al., ’93, ’94). Peptides of the VIP (va-soactive intestinal peptide) family are present inthe nervous system of Helix, where they are in-volved in several functions such as vasodilation,water and electrolyte secretion, regulation of me-tabolism, modulation of circadian rhythm, promo-tion of glycogenolysis, etc. (Kaufmann et al., ’95).Recent reports have demonstrated that the num-ber of VIP-immunoreactive cells is lower in ac-tive snails collected in summer in comparison withhibernating animals (Kaufmann et al. ’95).
In addition to BAPs, the biogenic amine seroto-nin, which occurs in relatively high concentrationsin the central nervous system (CNS) of mollusks(Osborne et al., ’82; Osborne, ’84; Hernadi et al.,’89; Elekes, ’91; Vehovszky et al., ’93), can affectand modulate a variety of behavioral and physi-ological processes, such as feeding, withdrawal andescape reaction, blood circulation, and some formsof learning. Some data indicate that, in Helix, thenumber of serotonin-immunoreactive neurons islower during hibernation (Hernadi et al., ’89).
In order to have a histological and histochemi-cal picture of CNS physiology during hibernation,we have carried out a comparative analysis ofchanges in immunopositivity for BAPs, whose dis-tribution is at present unknown in hibernatingHelix aspersa. The BAPs we have analyzed are
involved in many functions accomplished by thecerebral ganglion of H. aspersa, which is a centerof integration of several stimuli (Kandel, ’76;Balaban, ’93) that are at least partially suppressedduring hibernation. The BAP-immunoreactivitychanges were compared with the immunocytochemi-cal pattern of serotonin, one of the first neurotrans-mitters studied by fluorescence histochemistry(Cardot, ’71; Kerkut and Walker, ’75). A preliminaryreport of some of these findings has already ap-peared in abstract form (Bernocchi et al., ’93).
MATERIALS AND METHODSAdult terrestral snails (Helix aspersa), collected
in the garden of the Section of Ecology (Depart-ment of Genetics and Microbiology, University ofPavia) were used. The active animals were col-lected in September and remained active in thelaboratory for 10 days before killing. The hiber-nating snails were collected in November with theshell closed by the epyphragm and kept in thiscondition at 4°C in a cage until February. A totalof 54 animals of similar size was used.
Fixation and sectioningThe cerebral ganglia were fixed in Bouin solu-
tion (12 for both active and hibernating animals)for 12 h at room temperature or in 4% paraform-aldehyde in 0.1 M phosphate buffer, pH 7.2 (12for both active and hibernating animals), for 12 hat 4°C. After being washed in 70% ethanol for 3days (Bouin) or in phosphate buffer for 12 h(paraformaldehyde), the ganglia were dehydratedand embedded in paraffin. Eight-µm thick hori-zontal or transverse sections were collected on si-lane-coated slides.
Three other ganglia fixed in Bouin for eachexperimental group were cut serially (series of4 slides) for counting immunoreactive neurons(see below).
ImmunocytochemistryImmunocytochemical reactions were carried out
simultaneously for active and hibernating ani-mals. For every antibody and fixative solution, twoslides from at least three animals for both the ac-tive and hibernation periods were used.
On Bouin- and paraformaldehyde-fixed speci-mens, the following rabbit polyclonal antibodieswere used: 1:500 anti-Substance P (SP, Sera Lab,Sussex, U.K.), 1:500 anti-Cholecystokinin/Gastrin(CCK8/Gastrin, Incstar, MN), 1:500 anti-Calcitonin-Gene-related Peptide (CGRP, Peninsula Lab, CA),1:500 anti-Endothelin (ET, Peninsula Lab). Seroto-

356 G. BERNOCCHI ET AL.
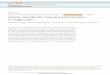
Fig. 1. Immunoreaction for CGRP in paraformaldehyde-fixed ganglia. (a) Active snails: the neurons (n) are immuno-negative; in the central neuropile there are varicosities andstrongly labeled fibers. (b) Hibernating snails: several small-
and medium-sized neurons of the metacerebrum are stronglyimmunopositive, but some areas (stars) of the central neuro-pile do not show immunopositive fibers or varicosities. a: ×90;b: ×100.
nin immunocytochemistry (1:500 antiserotonin,Incstar) was performed on paraformaldehyde-fixedspecimens only. For all antibodies, the incubationtime was 18 h. The peroxidase-antiperoxidase (PAP)method of Sternberger et al. (’70) was used to re-veal the sites of antigen-antibody reaction. Briefly,the sections were incubated in 3% H2O2 in 10%methanol for 7 min; 5% normal goat serum for 45min; primary antibodies; 1:50 goat-anti-rabbit IgGfor 90 min; 1:200 rabbit PAP for 60 min; and 0.05%3,3´diaminobenzidine tetrahydrochloride in 0.05 MTris/HCl buffer, pH 7.6, with 0.01% H2O2 for 20 min.All washings and incubations were in PBS and atroom temperature (20°C). For negative controls,some sections were incubated in normal rabbit se-rum instead of the primary antibodies at the samedilution; for positive staining, control tissues andcells that contain the antigen were used.
Counting neuronsThe number of immunoreactive for BAPs and
immunonegative neurons of the metacerebrumand mesocerebrum was estimated on serial sec-tions of Bouin-fixed ganglia. To avoid multiplecounting of a neuron, its cell body was traced outon consecutive sections. The number of immunore-active neurons was also expressed as percentageof the total neuronal population. The differences
between the active period and hibernation weresignificant by a Mann-Whitney test (P < 0.001).
RESULTSAll the antibodies recognized epitopes in the neu-
ronal perikarya and/or in the fibers of neuropileand interneuronal area (trophospongium) of theland snail CNS. The staining pattern was consis-tent in the different animals of the same period(activity or hibernation). No immunohistochemicallabeling was observed in the cerebral areas andneuropiles in negative control slides.
In the following paragraphs we shall describeanalytically the patterns of the metacerebrum andmesocerebrum (Figs. 1–7) and the central neuro-pile. In the procerebrum, with all the antibodies,no positivity was detectable in the cytoplasm of thesmall neurons during both the active period andthe period of hibernation. During the active period,around the neurons and in the neuropile, there werefibers and varicosities immunopositive for some ofthe antibodies, e.g., serotonin (Fig. 5a), SP, CGRP,ET. This BAP and serotonin immunoreactivity wasless detectable during the hibernation (Fig. 5b).
Influence of fixativesParaformaldehyde permitted a better detection
of the immunopositivity to some BAPs of the fi-bers in the neuropiles and around neurons. For in-

HIBERNATION AND NEURONAL ACTIVITY 357
Fig. 2. Immunoreaction for CGRP in Bouin-fixed ganglia.(a) Active snails: immunopositivity is detectable in some smalland large neurons and the giant neuron (GN, arrow) of the
metacerebrum. (b) Hibernating snails: most neurons of themetacerebrum, including the GN (arrow) are strongly im-munostained. a, b: ×100.
Fig. 3. Immunoreaction for CCK8/gastrin in Bouin-fixedganglia. (a, b) Active snails: the fibers around neurons areimmunopositive (arrows). On the contrary, the neurons arenegative. (c, d) Hibernating snails: in the meso (Ms) and
metacerebrum (Mt), part of the cytoplasm of several neuronsis immunopositive; the fibers around neurons maintainimmunopositivity (arrows). a, c: ×90; b, d: ×360.

358 G. BERNOCCHI ET AL.
Fig. 4. Immunoreaction for SP in Bouin-fixed sections. (a)Active snails: immunopositive fibers (arrows) are presentaround neurons that appear negative. (b) Hibernating snails:
a part of the cytoplasm of some mesocerebral neurons is im-munoreactive (stars). a, b: ×360.
stance, after immunoreaction for CGRP (Fig. 1a),CCK8 (not shown) and SP (not shown) strong posi-tivity was present in the varicosities and fibers ofthe central neuropile of active snails. This positiv-ity was not observed in several areas of the neuro-pile during hibernation (Fig. 1b) and was decreasedafter immunoreaction for SP around neurons.
In general, as analytically described below,Bouin solution allowed a better visualization ofthe BAP immunocytochemical reaction in the neu-ronal cytoplasms (see Figs. 2–6).
Immunocytochemistry for CGRPIn comparison with the active period (Fig. 2a),
a greater number of neurons were immunopositivein both metacerebrum and mesocerebrum duringhibernation (Fig. 2b). The metacerebral giant neu-ron (GN) was entirely stained.
Immunocytochemistry for CCK8/gastrinA few weakly immunopositive neurons and
immunopositive fibers around neurons were ob-served in the active snails (Fig. 3a,b). Duringhibernation, the fibers maintained some im-munopositivity (Fig. 3c,d). Several large neu-rons of the metacerebrum and mesocerebrum(Fig. 3c,d), including GN (Fig. 6d), were reac-tive and had the immunopositivity often con-fined to a discrete area of the cytoplasm.
Immunocytochemistry for SPOnly a few of the small neurons were strongly
positive in the active animals. The large neurons
of the metacerebrum and mesocerebrum werenegative in the active period (Fig. 4a) but veryreactive in the period of hibernation (Fig. 4b). Insome of the large neurons, the positivity was lo-cated in a discrete area of the cytoplasm. The GNwas weakly positive (not shown).
Immunocytochemistry for ETIn active animals (Fig. 5a,b), the immunopositivity
was in the fibers around the neurons and in thecytoplasm of some neurons (Fig. 5b). Hibernationdetermined a decrease in the immunoreactive fibersand an increase in the number of immunopositiveneuronal cytoplasms (including the GN, Fig. 5c,d;6c) in both the metacerebrum and mesocerebrum.The immunopositivity in neurons was mainly lo-cated in a distinct area of the cytoplasm (Fig. 5c,d).
Coexistence/colocalization of ET and CGRPin the mesocerebrum
Adjacent sections of hibernating snail gangliashowed that in both the left and right mesocerebra,ET-immunopositive neurons (Fig. 6a) were present.The positivity was stronger in the peripheral area ofthe left mesocerebrum. Almost all the neurons of the
Fig. 5. Immunoreaction for ET in Bouin-fixed cerebralganglia. (a, b) Active snails: immunopositive fibers or vari-cosities are found around neurons, of which only a few arelabeled (arrows). (c, d) Hibernating snails: most of the neu-rons, including the GN (arrow), are immunoreactive. Fibersaround neurons are less immunopositive. a, c: ×140; b, d: ×360.

HIBERNATION AND NEURONAL ACTIVITY 359
Figure 5.

360 G. BERNOCCHI ET AL.
Fig. 6. Immunoreaction for ET, CGRP and CCK8/gastrinin Bouin-fixed ganglia of hibernating snails. (a) Immunore-action for ET in the mesocerebrum: in the right (R) meso-cerebrum, all the neurons are immunostained; in the left(L) mesocerebrum, the peripheral neurons are more stronglylabeled than those of the central area. (b) Immunoreactionfor CGRP in the mesocerebrum: in the right mesocerebrum,
a group of central-lateral neurons are immunostained, whilein the left, almost all the neurons are strongly positive.(c) Immunoreaction for ET in the GN: the immunoreaction(arrow) is confined to a segregated cytoplasmic area. (d) Im-munoreaction for CCK8/gastrin in the GN: the same cyto-plasmic area (arrow) as in c is weakly positive. a, b: ×90; c,d: ×580.

HIBERNATION AND NEURONAL ACTIVITY 361
Fig. 7. Immunoreaction for serotonin in paraformalde-hyde-fixed cerebral ganglia. (a, b, c) Active snails: in the neu-ropile of the procerebrum (Pnp), there are immunopositivevaricosities with granular appearance and fine fibers (a); theGN is immunopositive (b), and a large number of labeled fi-bers and varicosities (arrows) are present in the central neu-
ropile (c). (d, e, f) Hibernating snails: in the procerebrum,lesser granular varicosities and fine fibers are evident (d);increased immunoreactivity is shown in the GN (e). In theneuropile surrounding the GN (e) and in the central neuro-pile (f), a reduction in the positive varicosities can be observed.a, b, c, d, e, f: ×360.

362 G. BERNOCCHI ET AL.
left mesocerebrum were also positive for CGRP (Fig.6b). In the right mesocerebrum, on the contrary, onlythe central area was immunoreactive (Fig. 6b).
The GN was immunopositive for ET (Fig. 6c)and CCK8 (Fig. 6d). For both the immunoreac-tions, the distribution of immunoprecipitates wasconfined to the same distinct cytoplasmic area.
Immunocytochemistry for serotoninIn the active snails, the immunopositivity was
observed in the GN (Fig. 7b) and in some neu-rons of the metacerebrum; numerous positive vari-cosities and fibers were also present in the centralneuropile (Fig. 7c).
A marked increase in the intensity of the reac-tion was found in the same neurons, including theGN (Fig. 7e), of the hibernating animals, whereasimmunopositive fibers of the neuropile adjacent theGN were decreased in number (Fig. 7e,f).
Number and percentages of BAP-immunoreactive neurons
For all BAPs, the number (Table 1) and the per-centages (Fig. 8) of immunoreactive neurons dur-ing the active period were very low, mainly in themesocerebrum (from 4 ± 0.9 to a maximum of 12± 2.5 that corresponded to about 2.5% and 7% ofthe neuronal population). In hibernating snails,the percentages were 2- to 4-fold in the metacere-brum and 28-fold in the mesocerebrum. The high-est percentages during hibernation were in themesocerebrum (64–79%).
DISCUSSIONCritical aspects
Though we used antibodies that are raisedagainst vertebrate antigens, they recognized epi-
Fig. 8. Percentages of BAP-like immunoreactive neuronsin the metacerebrum (MT) and mesocerebrum (MS) duringactive period (Ac) and hibernation (Hi). CCK8: Cholecystoki-nin; SP: Substance P; CGRP: Calcitonin-gene-related peptide;ET: Endothelin.
TABLE 1. Changes in the number of BAP-likeimmunoreactive neurons of the metacerebrum and
mesocerebrum during active period and hibernation*
Activeperiod Hibernation
CCK8 metacerebrum 26 ± 5 103 ± 5mesocerebrum 10 ± 2 85 ± 8
SP metacerebrum 46 ± 6 100 ± 11mesocerebrum 12 ± 3 114 ± 6
CGRP metacerebrum 40 ± 7 136 ± 10mesocerebrum 4 ± 1 112 ± 7
ET metacerebrum 39 ± 3 150 ± 14mesocerebrum 4 ± 1 92 ± 3
*Values are given as mean ± S.D.; n = 3 per each period; Mann-Whitney test significant level P < 0.001. CCK8, cholecystokinin; SP,substance P; CGRP, calcitonin-gene-related peptide; ET, endothelium.
topes in the Helix aspersa CNS. Indeed, the dataobtained with vertebrate antibodies to reveal in-vertebrate antigens must be interpreted with somecaution. In particular, the results may be affectedby cross-reaction with unrelated molecules, or im-munocytochemical reaction may have detectedparts of a molecule that can be either identical toor shorter or longer than the original antigen be-cause we used polyclonal antibodies that containseveral immunoglobulins against different epitopesof the same immunogenic antigen. As a conse-quence, we describe an “antigen-like immunoreac-tivity.” Nonetheless, as reported below, most of theBAPs considered here and serotonin have beenchemically identified in Helix (for a review, seeElekes et al., ’94) or other gastropods (Vigna et al.,’84; Sonetti et al., ’90; Giaid et al., ’91).
It is known that the type of fixation can influenceimmunoreactivity, in particular of peptidergic cells(Schot et al., ’84; Andriès et al., ’91). We emphasizethat with the two fixatives we used immunolabelingfor BAPs was often detected in different places. Forinstance, after immunoreaction to CGRP, CCK8 andSP, fibers and varicosities were better visualized in

HIBERNATION AND NEURONAL ACTIVITY 363
formaldehyde-fixed specimens, whereas neuronsshowed immunopositivity mainly after fixation withBouin solution. Only for immunoreactions to CCK8and ET were fibers and neurons labeled fromsamples fixed in formaldehyde and those fixed inBouin solution.
Occurrence and function of BAPs andserotonin in gastropods
Several BAPs have been mapped in the ner-vous system of gastropods and in particular inHelix in order to increase the knowledge aboutputative peptidergic pathways. Some BAPs, suchas CCK8/gastrin (Osborne, ’84; Vigna et al., ’84;Gesser and Larsson, ’85; Sonetti et al., ’90),FMRF-amide (Cottrel et al., ’92), myomodulin(Santama et al., ’94), and buccalin (Santama etal., ’94) are implicated in control of feeding be-havior. SP, which has been described in some mol-luskan neurons (Osborne, ’84; Sonetti et al., ’90),is involved in sensory pathways concerned withpain and touch. In mammals, calcitonin andCGRP are involved in fundamental physiologicalfunctions such as pain perception, appetite andblood flow regulation, and vasodilation (Breimeret al., ’88), and in mollusks, CGRP has been foundin slugs (Sasayama et al., ’91). First recognizedas a potent vasoconstrictor, ET has been immu-nocytochemically demonstrated in neurons con-trolling the vascular system and water retentionin Aplysia (Giaid et al., ’91). Opioid peptides areinvolved in a wide spectrum of physiological func-tions, such as thermoregulation, locomotion, painresistance, and immunomodulation (Kavaliersand Hirst, 1984). In particular, neurons contain-ing enkephalins have been recently mapped inganglia of Helix (Marchand et al., ’91; Elekes etal., ’93). Finally, the distribution of peptides ofthe VIP-family, which are involved in many func-tions, has been described in the CNS of Helix(Kaufmann et al., ’95).
With all the antibodies against the peptides wehave considered, individual neurons and fiberswere positive in the cerebral ganglia of H. aspersaduring the active period; the patterns of distribu-tion were quite similar to those previously de-scribed. In gastropods and in land snails of thegenus Helix (Osborne et al., ’82; Osborne, ’84;Cottrel et al., ’92; Elekes and Naessel, ’90; Gutier-rez and Asai, ’91; Marchand et al., ’91; Elekes etal., ’93, ’94; Kaufmann et al., ’95); several peptideshave been localized in the circumesophageal gan-glia, and we have found also fibers immunopositivefor antibodies against SP, CCK8, CGRP, and ET
in the neuropile or around the neurons of the metaand mesocerebrum. Some neuron somata were alsostained in these areas. It must be recalled thatthere are small, medium, and large neurons in themetacerebrum, whereas the mesocerebrum com-prises only large neurons (for a review, see Kerkutand Walker, ’75). In the metacerebrum, the neu-rons that showed SP-, CCK8-, CGRP- and ET-likeimmunoreactivity were mainly small in size, andin the mesocerebrum, the immunopositive neuronsoccurred sporadically.
In invertebrates, including mollusks, serotoninacts as neurotransmitter, modulator, and neuro-hormone (Hernadi et al., ’89; Hernadi and Elekes,’93). In gastropods in particular, serotonin can af-fect and modulate a variety of behavioral andphysiological processes, such as feeding (Weiss etal., ’82; Kupfermann and Weiss, ’82; Kemenes andSrozsa, ’87), withdrawal and escape reaction(Kupfermann and Weiss, ’82), blood circulation(Liebeswar et al., ’75), and some forms of learn-ing (Kandel, ’76; Balaban, ’93). In the Helix centralnervous system, it has been found by immuno-cytochemistry that both the neuropile and the cellbody layer are richly innervated by serotonergicfibers (Hernadi et al., ’89; Elekes, ’91). Accordingly,during the active period, we have found neuronswith serotonin immunoreaction in the Helixmetacerebrum, which sends numerous projectionsto the central neuropile, the body wall, and dif-ferent internal organs. The metacerebral giantneuron, which innervates buccal ganglia, also hadserotonin immunoreactivity, as previously re-ported (Osborne et al., ’82; Osborne, ’84).
BAP immunocytochemistry in hibernatingpulmonate snails
The most striking result of the present researchwas the observation that, in the hibernating Helixaspersa, a high number of neurons had immuno-positive cytoplasm for all BAPs. The immunoreac-tivity of fibers decreased for some BAPs in severalareas of the neuropile and for SP and ET aroundneurons. In the mesocerebrum, a high number ofneurons (64% to 78%) were immunopositive forCGRP, ET, CCK8, SP, and in the metacerebrum(30% to 57%) several small- and medium-sized neu-rons were also immunopositive. In the meta-cerebrum, the GN was strongly immunopositive forCGRP and SP, and a part of the cytoplasm was alsointensely immunoreactive for CCK8 and ET.
The high number of immunopositive neurons andthe increase in the intensity of staining in the im-munoreactive cells in hibernating terrestrial snails

364 G. BERNOCCHI ET AL.
suggest an accumulation of reactive epitopes andmolecules in the cytoplasm. This probably gives riseto the co-occurrence of two or more peptides in thesame cells, which were negative during activity, andto the co-localization of differently immunopositiveneurons. Two hypotheses may be put forward asexplanations for these points. The first one assumesthat the immunopositivity is really linked to BAPpresence. Actually, the segregation of immunopre-cipitates—for instance, for CCK8 and ET—in dis-crete areas of the cytoplasm of neurons that areentirely positive for CGRP and SP (e.g., the GN) isconsistent with a storage of BAPs (two or more) inhibernating snail neurons. Therefore, hibernationmay be a tool for studies of the functional role ofdifferent cerebral neurons. In this context, thesmall nervous systems not only give lessons for thestudy of functional organization of cotransmissionsystems (Marder et al., ’95), but the seasonal cycleof some invertebrates also offers an unique oppor-tunity to unravel the true function of brain areas.Though this was not the aim of our study, we can-not exclude the possibility of the co-occurrence oftwo or more peptides, which suggests that a neu-ron can be involved in different functions. Co-oc-currence of neurotransmitters and neuropeptideshas been widely documented (Osborne, ’84; Fasoloand Clairambault, ’91; Marder et al., ’95). More-over, the co-localization of neurotransmitters andneuropeptides in the cerebral ganglia of Helix isevident in the paper by Hernadi and Elekes (’93).
However, co-reactivity does not necessarilymean co-occurrence of two or more peptides. Infact, during the biosynthesis of neuropeptides, apreprotein is incorporated into dense-core vesiclesand subsequently processed to give several BAPsand then transported to the cell terminals. Thus,the preprotein is a precursor for several bioactivepeptides with distinct biological properties. In thisway, all the peptidergic neurons can putativelymanufacture different mixtures of peptides, eachwith a markedly different spectrum of biologicalactivity. The recent findings of Kaufmann et al.(’95) show that hibernating snails contained thehighest number of VIP and mainly of prepro-VIPimmunoreactive neurons. In this light, the posi-tivity of the neurons for two or more antibodiesagainst bioactive peptides also may be ascribedto the presence of a preprotein molecule, synthe-sized in the cell cytoplasm and blocked in this lo-calization. In this case, all the neurons may havethe capability and possess the preteolytic enzymesto manufacture several peptides. The hypothesiswill be tested by further research.
According to this interpretation, during hiberna-tion the neurons are ready to elaborate and releasemore than one bioactive peptide. The eventual di-versification of neurons occurs only during the pe-riod of activity. This places under discussion themultipotential role of neurons and the compartmen-talization of ganglia in invertebrates, as tentativelyanalyzed by Hernadi and Elekes (’93).
During hibernation the large neurons of themesocerebrum, which has a role in mating andavoidance behavior (for a review, see Chase andLi, ’94; Li and Chase, ’95), showed immunoreac-tivity for ET, CGRP, SP, and CCK8. This indicatesthat all these BAPs are involved in these activi-ties or that the mesocerebral neurons have morenumerous roles than expected. The same consid-eration may be made for GN, which controls thefeeding behavior (Chase and Tidd, ’91; Chase andTolloczko, ’92) and is immunopositive not only toCCK8, but also to CGRP and ET.
Finally, it is particularly interesting that thereare also differences in the immunoreactivity of thepaired mesocerebral lobes during hibernation; thissupports a different involvement of the right andleft mesocerebrum in the hormonal and/or neuronalactivities (for a review, see Chase and Li, ’94).
Serotonin immunocytochemistry inhibernating pulmonate snails
Another interesting result concerns the changes,not in only number but also in intensity in im-munoreactivity for serotoninergic neurons of themetacerebrum. In particular, the metacerebral GNwas described as a typical serotonin-containingneuron (Osborne et al., ’82; Osborne, ’84; Hernadiet al., ’89). This neuron increased its immunore-activity during hibernation, suggesting an accu-mulation of serotonin in the neuronal cytoplasm,as observed for peptides. It has been shown, how-ever, that 3H-serotonin injected into serotoninergicsomata accumulates in both synaptic vesicles andlarge lysosomes (Cottrel and Osborne, ’70; Schkol-nik and Schwartz, ’80). Consequently, it has beensuggested that serotoninergic synaptic vesiclesthat are not released are ultimately transportedback to the soma, where they are degraded by ly-sosomes. Electron immunocytochemistry willclarify whether the increase in the immuno-positivity of the soma of the GN is due to the stor-age of the neurotransmitter before the release orbefore being degraded by lysosomes. It must bestressed that the immunoreactivity of serotoninwas decreased in the nearby neuropile.

HIBERNATION AND NEURONAL ACTIVITY 365
Functional changes in the CNSduring hibernation
The physiology and metabolism of the organ-ism change during hibernation in both vertebratesand invertebrates (Hunter, ’64; Machin, ’75; Heller,’79; Lyman et al., ’82), to different extents, accord-ing to the modality of hibernation and, probably,to the systematic position.
Studies carried out mainly on organs, tissues, andcells from vertebrate species (Kolaeva et al., ’80;Barni et al., ’87; Bernocchi et al., ’86; Giacometti etal., ’89; Anderson et al., ;90; Barni and Bernocchi,’91; Fenoglio et al., ’92; Malatesta et al., ’94, ’95;March and Reisman, ’95) have explained some con-trol mechanisms of adaptation of the animals to dif-ferent environmental conditions and have providedinteresting contributions to the knowledge of thestructure and function of quiescent cells. In the ver-tebrate CNS, information has been acquired con-cerning some structural and functional changes ofneurons during natural hibernation, mainly regard-ing the condensation state of chromatin (Bernocchiet al., ’86) and nucleolar and nuclear organization(Giacometti et al., ’89; Malatesta et al., ’94, ’95).These studies showed a lower activity of the nucleusboth in amphibians and in mammals.
There are few data about seasonal variation inserotonin, dopamine and enkephalin contents ingastropod ganglia (Cardot, ’71; Hiripi and Salanki,’73; Hernadi et al., ’89, ’93; Gutierrez and Asai,’91). The biochemical analysis demonstrated thatlevels of serotonin and dopamine were higher inthe spring than in winter (Hernadi et al., ’89;Juhos et al., ’93).
Furthermore, there are few studies about the his-tochemistry of circumesophageal ganglia during theseasonal cycle (Gesser and Larsson, ’85; Hernadiet al., ’89; Kaufmann et al., ’95). In particular,Hernadi et al. (’93) observed a notable amount ofdopamine in the CNS of Helix, accompanied by alow number of dopamine-immunopositive neuronsin summer, but the highest number of dopamine-immunopositive neurons was present in winter,when the dopamine content was lower. The differ-ent distribution and intensity of immunopositivitywe have found in the neuropile and in neuronalsomata could explain this discrepancy.
The immunocytochemistry of some markers ofsystems that regulate neuronal morphology andactivity showed clear changes in the cerebral gan-glion of H. aspersa during hibernation in com-parison with the active period (Vignola et al., ’95).In hibernating snails, there was an increased im-munoreactivity for calmodulin, in agreement with
an increase of the intracellular Ca2+ concentra-tion. Calmodulin is the major intracellular Ca2+
recipient in most eukaryotic cells and is thus animportant mediator for many basic cellularprocesses, including the synthesis and release ofneurotransmitters. This suggests that, during hi-bernation, neurons decrease or suppress the Ca2+-dependent mechanisms. The lowered Ca2+-ATPaseactivity (Vignola et al., ’95), which also plays arole in regulating the intracellular ionized calciumconcentration, supports this interpretation.
Finally, the immunocytochemical expression ofsome cytoskeletal components also changed dur-ing hibernation (Vignola et al., ’95). There was anincrease in the high-molecular-weight phosphory-lated neurofilament-like immunoreactivity in thecytoplasm of some clusters of neurons and, in con-trast, a decrease in the immunopositivity for al-pha-tubulin and microtubule-associated proteins2 in the neuropile, which suggests that the trans-port of vesicles in the axons may be slowed dur-ing hibernation (Vignola et al., ’95). This isconsistent with the modification in the pattern ofneurotransmission and neuromodulation shown inthe present investigation.
ACKNOWLEDGMENTSThe research was supported by Italian M.U.R.S.T.
(40% and 60%) and C.N.R. grants to G. Bernocchi.We are grateful to Mrs. P. Veneroni and Mr. S.Veneroni for their excellent technical assistance.Special thanks are also due to Mrs. T. Bosini fortyping the manuscript.
REFERENCESAnderson, D.G., G.A. Lopez, D. Bewernick, S. Brazal, J. Pon-
der, and G.M. Russom (1990) Changes in renal morphologyand renin secretion in the golden-mantled ground squirrel(Spermophilus lateralis). Cell Tissue Res., 262:99–104.
Andriès, J.C., G. Belemtougri, and G. Tramu (1991) Multiplepeptide immunoreactivities in the nervous system of Aeschnacyanea (Insecta, Odonata). Histochemistry, 96:139–148.
Balaban, P. (1993) Behavioral neurobiology of learning in ter-restrial snails. Progr. Neurobiol., 41:1–19.
Barni, S., and G. Bernocchi (1991) Internalization of eryth-rocytes into liver parenchymal cells in naturally hibernat-ing frogs (Rana esculenta L.). J. Exp. Zool., 258:143–150.
Barni, S., G. Bernocchi, and G. Gerzeli (1987). Nuclearchanges and morphology of the epidermis in the hibernat-ing frog. Tissue Cell, 19:817–825.
Bernocchi, G., S. Barni, and E. Scherini (1986) The annualcycle of Erinaceus europaeus L. as a model for a furtherstudy of cytochemical heterogeneity in Purkinje neuron nu-clei. Neuroscience, 17:427–437.
Bernocchi, G., C. Vignola, B. Lomazzi, and E. Scherini (1993)Il ganglio cerebrale di Helix aspersa durante l’ibernazione.Un modello per lo studio dei sistemi intracellulari cheregolano la neuroarchitettura. 55th Congr. UZI, Torino, 27/9-2/10/1993, p 40.

366 G. BERNOCCHI ET AL.
Breimer, L.H., I. MacIntyre, and M. Zaidi (1988) Peptidesfrom the calcitonin genes: Molecular genetics, structure andfunction. Biochem. J., 255:377–390.
Cardot, J. (1971) Variations saisonnières de la 5-hydrox-ytryptamine dans les tissus nerveux et cardiaque chez leMollusque Helix promatia. C.R. Soc. Biol., 165:338–341.
Chase, R., and G. Li (1994) Mesocerebral neurons and theirrole in the control of mating behaviour. Neth. J. Zool.,44:212–222.
Chase, R., and C. Tidd (1991) Stability of dendritic mass dur-ing aestivation. J. Comp. Neurol., 303:150–158.
Chase, R., and B. Tolloczko (1992) Synaptic innervation ofthe giant cerebral neuron in sated and hungry snails. J.Comp. Neurol., 318:93–102.
Cottrell, G.A. (1992) Roles and actions of neuropeptide mes-sengers; insights from studies with FMRFamide-related pep-tides and molluscan neurons. C.R. Acad. Sci. (Paris) t. 314,Série III, Suppl., 9:81–82.
Cottrell, G.A., and N.N. Osborne (1970) Subcellular localiza-tion of serotonin in an identified serotonin-containing neu-ron. Nature, 225:470–472.
Cottrell, G.A., D.A. Price, K.E. Doble, S. Hettle, J. Sommer-ville, and M. MacDonald (1992) Identified Helix neurons:Mutually exclusive expression of the tetrapeptide andheptapeptide members of the FMRFamide family. Biol. Bull.,183:113–122.
Elekes, K. (1991) Serotonin-immunoreactive varicosities inthe cell body region and neural sheath of the snail, Helixpomatia, ganglia: An electron microscopic immunocy-tochemical study. Neuroscience, 42:583–591.
Elekes, K., and D.R. Naessel (1990) Distribution of FMRF-amide-like immunoreactive neurons in the central nervoussystem of the snail Helix pomatia. Cell Tissue Res.,262:177–190.
Elekes, K., G.B. Stefano, and D.O. Carpenter (1993) Enkepha-lin-like immunoreactive neurons in the central nervous sys-tem of gastropods (Helix pomatia, Lymnaea stagnalis,Aplysia californica): A comparative immunocytochemicalstudy. Cell Tissue Res., 272:329–341.
Elekes, K., L. Hernadi, J.E. Muren, and D.R. Naessel (1994)Peptidergic neurons in the snail Helix pomatia: Distribu-tion of neurons in the central and peripheral nervous sys-tems that react with an antibody raised to the insectneuropeptide, Leucokinin I. J. Comp. Neurol., 341:257–272.
Engelhardt, R.P., N. Dhainaut-Courtois, and G. Tramu (1982)Immunohistochemical demonstration of a CCK-like peptidein the nervous system of a marine annelid worm, Nereisdiversicolor O.F. Muller. Cell Tissue Res., 227:401–411.
Fasolo, A., and P. Clairambault (1991) Zoologie et BiologieAnimale. Quelques aspects des neuropeptides. Ann. Sci. Nat.(Paris, 13e série, 12:107–155.
Fenoglio, C., G. Bernocchi, and S. Barni (1992) Frog hepato-cyte modifications induced by seasonal variations: A mor-phological and cytochemical study. Tissue Cell, 24:17–29.
Gesser, B.P., and L.I. Larsson (1985) Changes of enkephalin-like to gastrin/cholecystokinin-like immunoreactivity in snailneurons. J. Neurosci., 5:1412–1417.
Giacometti, S., E. Scherini, and G. Bernocchi (1989) Seasonalchanges in the nucleoli of Purkinje cells of the hedgehogcerebellum. Brain Res., 488:365–368.
Giaid, A., T. Masaki, T. Ouimet, M. Yanagisawa, L. Gaspar, M.Cantin, S. Kimura, and V.F. Castellucci (1991) Expression ofendothelin-like peptide in the nervous system of the marinemollusk Aplysia. J. Cardiovasc. Pharmacol., 17:S449–S451.
Gutierrez, R., and M. Asai (1991) Ir-met and ir-leu-enkephalin
content in the perioesophageal ganglia of Helix aspersa. Sea-sonal variations. Comp. Biochem. Physiol., 100C:609–613.
Heller, H.C. (1979) Hibernation: Neural aspects. Annu. Rev.Physiol., 41:305–321.
Hernadi, L., and K. Elekes (1993) Peptidergic and aminergiccenters in the Helix cerebral ganglia: Somatotopy and im-munocytochemistry. Acta Biol. Hung., 44:89–92.
Hernadi, L., K. Elekes, and K. Srozsa (1989) Distribution ofserotonin-containing neurons in the central nervous systemof the snail Helix pomatia. Cell Tissue Res., 257:313–323.
Hernadi, L., S. Juhos, and K. Elekes (1993) Distribution oftyrosine-hydroxylase immunoreactive and dopamine immu-noreactive neurons in the central nervous system of thesnail Helix pomatia. Cell Tissue Res., 274:503–513.
Hiripi, L., and J. Salanki (1973) Seasonal and activity-de-pendent changes of the serotonin level in the CNS and heartof the snail (Helix pomatia L.). Comp. Gen. Pharmacol.,4:285–292.
Hunter, W.R. (1964) Ecology in non marine forms. In: Physi-ology of Mollusca. K.M. Wilbur, and C.M. Yonge, eds. Aca-demic Press, New York, Vol. 1, pp. 83–126.
Juhos, S.Z., L. Hiripi, and K. Elekes (1993) Biochemical char-acterization of a serotonergic system in the neural sheathof Helix ganglia. Acta Biol. Hung., 44:83–87.
Kandel, E.R. (1976) Cellular Basis of Behavior. Freeman andCo., San Francisco.
Kaufmann, W., H.H. Kerschbaum, C. Hauser-Kronberg, G.W.Hacker, and A. Hermann (1995) Distribution and seasonalvariation of vasoactive intestina (VIP)-like peptides in thenervous system of Helix pomatia. Brain Res., 695:125–136.
Kavaliers, M., and M. Hirst (1984) The presence of an opioidsystem mediating behavioral thermoregulation in the ter-restrial snail Cepea nemoralis. Neuropharmacology, 23:1285–1289.
Kemenes, G., and K. Srozsa (1987) The role of serotonergicmechanisms in food-induced arousal of the snail Helixpomatia L. In: Neurobiology, Molluscan Models. H.H. Boer,W.P.M. Geraerts, and J. Joosse, eds. Amsterdam: North Hol-land, pp. 277–286.
Kerkut, G.A., and R.J. Walker (1975) Nervous system, eye andstatocyst. In: Functional Anatomy and Physiology. W. Fretter,and J. Peake, eds. Academic Press, London, pp. 165–244.
Kolaeva, S.G., L.I. Kramarova, E.N. Ilyasova, and F.E. Ilyasov(1980) The kinetics and metabolism of the cell. Int. Rev.Cytol., 66:147–170.
Kupfermann, I., and K.R. Weiss (1982) Activity of an identi-fied serotonergic neuron in free moving Aplysia correlatedwith behavioral arousal. Brain Res., 241:334–337.
Li, G., and R. Chase (1995) Correlation of axon projectionsand peptide immunoreactivity in nesocerebral neurons ofthe snail Helix aspersa. J. Comp. Nerol., 353:9–17.
Liebeswar, G., J.E. Goldman, J. Koester, and E. Mayeri (1975)Neural control of circulation in Aplysia. III: Neurotrans-mitters. J. Neurophysiol., 38:767–779.
Lyman, C.P., J.S. Willis, A. Malan, and L.C.H. Wang (1982)Hibernation and Torpor in Mammals and Birds. AcademicPress, New York.
Machin, J. (1975) Water relationships. In: Pulmonates. Vol.1: Functional Anatomy and Physiology. V. Fretter and J.Peake, eds. Academic Press, London, pp. 105–163.
Malatesta M., C. Zancanaro, T.E. Martin, E.K.L. Chan, F.Amairic, R. Luehrmann, P. Vogel, and S. Fakan (1994)Cytochemical and immunocytochemical characterizationof nuclear bodies during hibernation. Eur. J. Cell Biol.,65:82–93.

HIBERNATION AND NEURONAL ACTIVITY 367
Malatesta M., C. Zancanaro, M. Tamburini, T.E. Martin, X.D.Fu, P. Vogel, and S. Fakan (1995) Novel nuclear ribonucle-oprotein structure components in the dormouse adrenal cor-tex during hibernation. Chromosoma, 104:121–128.
March, P.E., and H.M. Reisman (1995) Seasonal changes inhepatocyte ultrastructure correlated with the cyclic synthe-sis of secretory proteins in the winter flounder (Pleuronectesamericanus). Cell Tissue Res., 281:153–161.
Marchand, C.R., B. Griffond, K. Mounzih, and C. Colard(1991) Distribution of methionine-enkephalin-like andFMRFamide-like immunoreactivities in the central nervoussystem (including dorsal bodies) of the snail Helix aspersaMueller. Zool. Sci., 8:905–913.
Marder E., A.E. Christie, and V.L. Kilman (1995) Functionalorganization of cotransmission systems: Lessons from smallnervous system. Invertebrate Neuroscience, 1:105–112.
Osborne, N.N. (1984) Putative neurotransmitters and theircoexistence in gastropod mollusks. In: Coexistence of neu-roactive substances in neurons. V. Chan-Palay and S.L.Palay, eds., John Wiley & Sons, New York, pp. 395–409.
Osborne, N.N., A.C. Cuello, and G.J. Dockray (1982). Sub-stance P and cholecystokinin-like peptides in Helix neuronsand cholecystokinin and serotonin in a giant neuron. Sci-ence, 216:409–411.
Santama, N., M. Brierley, J.F. Burke, and P.R. Benjamin(1994) Neural network controlling feeding in Lymnaeastagnalis: immunocytochemical localization of myomodulin,small cardioactive peptide, buccalin, and FMRFamide-re-lated peptides. J. Comp. Neurol., 342:352–365.
Sasayama, Y., A. Katoh, C. Oguro, A. Kambegawa, and A.Yoshizawa (1991) Cells showing immunoreactivity for cal-citonin or calcitonin gene-related peptide (CGRP) in the cen-tral nervous system of some invertebrates. Gen. Comp.Endocrinol., 83:406–414.
Schkolnik, L., and J.H. Schwartz (1980) Genesis and matu-
ration of serotonergic vesicles in identified giant cerebralneurons of Aplysia. J. Neurophysiol., 43:945–967.
Schot, L.P.C., H.H. Boer, and C. Montagne-Wajer (1984)Characterization of multiple immunoreactive neurons inthe central nervous system of the pond snail Lymnaeastagnalis with different fixatives and antisera absorbedwith the homologous and the heterologous antigens. His-tochemistry, 81:373–378.
Sonetti, D., C. Lusvardi, and A. Fasolo (1990) Immunohis-tochemical localization of some vertebrate-like neuropep-tides (SP, NPY, CGRP, CCK) in the central nervous systemof the freshwater snail Planorbarius corneus. Cell TissueRes., 260:435–448.
Sternberger, L.A., P.H. Hardy, J.J. Cuculis, and J.D. Meyer(1970) The unlabelled antibody-enzyme method of immuno-cytochemistry: Preparation and properties of soluble antigenantibody complex (HRP-anti-HRP) and its use in identifica-tion of spirochetes. J. Histochem. Cytochem., 10:315–328.
Vehovszky, A., L. Hernadi, K. Elekes, and P. Balaban (1993)Serotonergic input on identified command neurons in He-lix. Acta Biol. Hung., 44:97–101.
Vigna, S.R., J.L.M. Morgan, and T.M. Thomas (1984) Local-ization and characterization of gastrin/cholecystokinin-likeimmunoreactivity in the central nervous system of Aplysiacalifornica. J. Neurosci., 4:1370–1377.
Vignola, C., C. Fenoglio, E. Scherini, and G. Bernocchi (1995)The cerebral neurons of Helix aspersa during hibernation.Changes in the cytochemical detection of calmodulin,cytoskeletal components and phosphatases. Tissue Cell,27:185–196.
Weiss, K.R., U.T. Koch, J. Koester, S.C. Rosen, and I.Kupfermann (1982) The role of arousal in modulating feed-ing behavior of Aplysia: Neuron and behavioral studies. In:The Neural Basis of Feeding and Reward. B.G. Hoebel andD. Novin, eds. Haer Institute, Brunswick, N.J., pp. 25–57.