Embed Size (px)
Citation preview
Author correction
Montross, W. T., Ji, H. and McCrea, P. D. (2000). A �-catenin/engrailed chimera selectively suppresses Wnt signaling. J. CellSci. 113, 1759-1770.
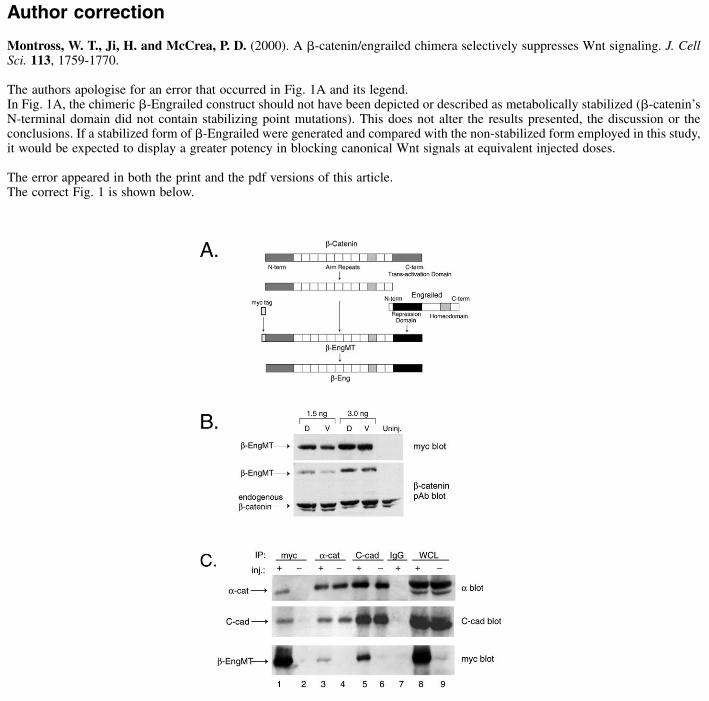
The authors apologise for an error that occurred in Fig. 1A and its legend. In Fig. 1A, the chimeric �-Engrailed construct should not have been depicted or described as metabolically stabilized (�-catenin’sN-terminal domain did not contain stabilizing point mutations). This does not alter the results presented, the discussion or theconclusions. If a stabilized form of �-Engrailed were generated and compared with the non-stabilized form employed in this study,it would be expected to display a greater potency in blocking canonical Wnt signals at equivalent injected doses.
The error appeared in both the print and the pdf versions of this article.The correct Fig. 1 is shown below.
Jour
nal o
f Cel
l Sci
ence

INTRODUCTION
β-catenin, the vertebrate orthologue of Drosophila Armadillo(McCrea et al., 1991; Peifer et al., 1991), is a versatile moleculethat functions in multiple cellular capacities (Barker et al.,2000; Hatzfeld, 1999) including cadherin-dependent cell-celladhesion (Barth et al., 1997; Peifer, 1995; Steinberg andMcNutt, 1999). Within adherens junctions, β-catenin linkscadherin to the cortical actin cytoskeleton in association withadditional proteins such as α-catenin (Herrenknecht et al.,1991; Nagafuchi and Tsukita, 1994) and α-actinin (Knudsenet al., 1995; Ozawa, 1998). In common with integrins and otheradhesion systems, cadherin function is believed to bemodulated by such cytoskeletal interactions (Alattia et al.,1999). In addition to its role at the plasma membrane, β-cateninis an essential participant in the Wnt signal transductionpathway (Dierick and Bejsovec, 1999; Gumbiner, 1995),wherein it physically associates with certain members of theHigh Mobility Group (HMG) of transcription factors, namelyLef/Tcf (Barker et al., 2000; Behrens et al., 1996), to activategene transcription (Cavallo et al., 1997; Clevers and van deWetering, 1997). In its role as a signaling molecule, β-cateninis critical in effecting axis specification and pattern formationin the early vertebrate embryo (Funayama et al., 1995; Millerand Moon, 1996; Molenaar et al., 1996; Sokol, 1999). β-catenin is characterized by a central Armadillo domain that iscomposed of 12 imperfect Arm repeats, each of approximately
42 amino acids, which facilitate interactions with variousprotein partners (Huber et al., 1997; Peifer et al., 1994). Forexample, at the plasma membrane, β-catenin’s Armadillodomain binds a highly conserved region within cadherin’scytoplasmic tail. In the nucleus, β-catenin’s Armadillo domainassociates with the N-terminal region of Lef/Tcf transcriptionfactors (Behrens et al., 1996; Cavallo et al., 1997; van deWetering et al., 1997). β-catenin’s Armadillo domain is flankedon either side by dissimilar N- and C-terminal domains, whichconfer additional functionality (Barth et al., 1996; Munemitsuet al., 1996). For example, the N-terminal domain participatesin regulating β-catenin signaling through its Wnt-dependentphosphorylation by GSK-3 on specific serine residues (Yost etal., 1996), resulting in β-catenin’s destabilization and reducedsignaling capacity (Dominguez et al., 1995; Fagotto et al.,1997; Pierce and Kimelman, 1995).
During early Xenopus development, Wnt signaling functionsto increase the cellular level of β-catenin in dorsal blastomeres(Fagotto et al., 1997; Gradl et al., 1999; Schneider et al., 1996;Wylie et al., 1996), establishing an organizing center throughthe direct activation of specific genes such as the homeoboxgene siamois (Brannon et al., 1997; Fan et al., 1998; Lemaireand Kodjabachian, 1996) and Xnr-3 (McKendry et al., 1997;Smith et al., 1995). Indirectly, β-catenin/Wnt signals alsomodulate expression of the dorsal and organizer specific genesgoosecoid (Cho et al., 1991; Laurent et al., 1997; Steinbeisseret al., 1993) and chordin (Sasai et al., 1994). Collectively, these
1759Journal of Cell Science 113, 1759-1770 (2000)Printed in Great Britain © The Company of Biologists Limited 2000JCS1039
β-catenin plays an integral role in cell-cell adhesion bylinking the cadherin complex of the adherens junction tothe underlying actin cytoskeleton. In addition, β-catenintransduces intracellular signals within the Wntdevelopmental pathway that are crucial to the properestablishment of embryonic axes and pattern formation ofearly mesoderm and ectoderm. For example, in the contextof a defined dorsal ‘organizer’ region of early Xenopusembryos, β-catenin enters the nucleus and associates withtranscription factors of the HMG (High Mobility Group)Lef/Tcf protein family. Consequently, genes such assiamois, a homeobox gene contributing to the specificationof the dorsoanterior axis, are activated. To further examinethe role that β-catenin plays in Wnt signaling, we generateda chimeric protein, β-Engrailed (β-Eng), in which the C-terminal trans-activation domain of β-catenin is replaced
with the transcriptional repression domain of DrosophilaEngrailed. Dorsal overexpression of this mRNA in earlyXenopus embryos leads to suppression of organizer-specificmolecular markers such as siamois, Xnr-3 and goosecoid,corresponding with the dramatic morphologicalventralization of embryos. Ventralized embryos furtherexhibit reduced activity of the Wnt pathway, as indicatedby the loss of the notochord/organizer marker, chordin.Importantly, β-Eng associates and functions normally withthe known components of the cadherin complex, providingthe experimental opportunity to repress β-catenin’ssignaling function apart from its role in cadherin-mediatedcell-cell adhesion.
Key words: Xenopus, β-catenin, Cadherin, Engrailed, Wnt,signalling, Cell-cell adhesion
SUMMARY
A β-catenin/engrailed chimera selectively suppresses Wnt signaling
Werner T. Montross, Hong Ji and Pierre D. McCrea*
Department of Biochemistry and Molecular Biology, University of Texas M. D. Anderson Cancer Center, Houston, TX 77030, USA*Author for correspondence (e-mail: [email protected])
Accepted 2 March; published on WWW 18 April 2000

1760
orchestrated transcriptional events aid in patterning thedorsoventral and anteroposterior axis of the developingembryo. The developmental power of the Wnt pathway hasbeen illustrated by misexpression of each of the knowncomponents of the signaling pathway. For example, theoverexpression of β-catenin in ventral blastomeres inducesdorsal mesoderm and generates an ectopic organizer capableof specifying an ectopic dorsal axis (Funayama et al., 1995;Guger and Gumbiner, 1995; McCrea et al., 1993).Furthermore, expression of β-catenin is capable of rescuing thedorsal axis following antisense depletion of maternal β-cateninor ultra-violet radiation (Funayama et al., 1995; Heasman etal., 1994), or the imposition of upstream blockades of the Wntsignal transduction cascade, demonstrating β-catenin’s key rolein the Wnt signaling pathway and axis specification (Deardorffet al., 1998; Itoh et al., 1998; Sokol, 1996).
In the absence of Wnt signals, the stability of the signalingpool of β-catenin is limited by a large protein complexincluding APC, Axin/Conductin, GSK-3, and components ofthe ubiquitination/proteasome pathway (Bienz, 1999;Easwaran et al., 1999; Ikeda et al., 1998; Kikuchi, 1999;Kishida et al., 1998; Rubinfeld et al., 1996; Salomon et al.,1997). Wnt signals prohibit the activity of GSK-3 within thiscomplex, resulting in β-catenin’s reduced ubiquitin/proteasome-mediated degradation, and thus its increasedstability and signaling capacity (Aberle et al., 1997).
The Wnt family is comprised of secreted glycoproteinligands that bind members of the frizzled family oftransmembrane receptors (Dale, 1998; Moon et al., 1997;Shulman et al., 1998). Frizzled receptor activation leads toGSK-3 inactivation in a manner that is incompletelyunderstood, but which requires the Dishevelled protein, itselfa complex entity capable of activating parallel and distinctsignaling pathways (Boutros and Mlodzik, 1999; Sokol, 1996).β-catenin-dependent Wnt signaling ultimately requires β-catenin’s entry into the nucleus and direct association witharchitectural transcription factors of the HMG box family(Clevers and van de Wetering, 1997; Nusse, 1997).Furthermore, β-catenin’s C-terminal transactivation domain(Vleminckx et al., 1999) facilitates expression of Wnt targetgenes including siamois and Xnr-3.
Since β-catenin acts in more than one capacity within thecell, primarily as a mediator of cadherin-dependent cell-celladhesion and signaling via the Wnt pathway, it has beenexperimentally challenging to distinguish between β-catenin’sfunctional contributions in either cellular or developmentalcontexts (Cox et al., 1996; Fagotto et al., 1996; Gumbiner,1995; Zhu and Watt, 1999). Lacking, thus far, has been anexperimental reagent capable of selectively blocking signalingmediated via β-catenin, as opposed to other upstream ordownstream components of the Wnt pathway. To address thisproblem, we have generated a chimeric protein in which the C-terminal transactivation domain of β-catenin has been replacedwith the active repression domain of Drosophila Engrailed(Jaynes and O’Farrell, 1991; Smith and Jaynes, 1996), whileleaving β-catenin’s N-terminal and Armadillo domains intact.Similar approaches employing Engrailed-repressor fusionshave been successfully used to block fushi tarazu (Smith andJaynes, 1996a), Xbra (Conlon et al., 1996) and siamois (Fanand Sokol, 1997; Kessler, 1997) mediated gene transcription.The β-catenin/Engrailed construct contains three S→A and one
T→A, N-terminal mutations (Fig. 1), which prevent itsphosphorylation via GSK-3 and subsequent ubiquitin/proteasome-mediated degradation (Yost et al., 1996). Whileleaving cadherin-mediated adhesion unperturbed, we find thatthis chimera, termed β-Engrailed (or β-Eng), successfullysuppresses β-catenin’s specific function within the Wntsignaling pathway, indicated by the graphic ventralization ofXenopus embryos as assessed by both molecular andmorphological criteria. We expect that β-Engrailed will proveuseful in multiple studies in which the specific repression of β-catenin mediated Wnt signaling is required.
MATERIALS AND METHODS
Xenopus embryo manipulations Xenopus females were induced to produce eggs by injection of 800units of human chorionic gonadotropin at least 10 hours prior to theirisolation. Eggs were obtained by standard means, placed in 0.1×MMR (Marc’s modified Ringers solution, pH 7.4: 100 mM NaCl, 2mM KCl, 2 mM CaCl2, 1 mM MgCl2, 5 mM Hepes), and fertilizedin vitro. The jelly coat was removed by incubation of the embryosprior to initial cleavage by brief room temperature incubation in asolution of 2% cysteine HCl (pH 8.0). Blastula cleavage stages anddorsal versus ventral polarity were determined according to themethod of Nieuwkoop and Faber (1967). Borosilicate glassmicroinjection pipettes (capillary tubes; Sutter Instrument Co.,Novato, CA, USA) were pulled using a P-30 pulling instrument(Sutter), bevelled with a K. T. Brown Type Micropipette beveller(Sutter), and embryos microinjected using the NA-1 oil-drivenmicroinjector (Sutter). Microinjection of RNA was performed at 2-,4- and 8-cell cleavage stages into either one or two blastomeres in thesubequatorial region of the respective marginal zones unless otherwisestated, and embryos placed within a solution of 5% Ficoll in 1× MMRfor at least 60 minutes prior to transfer to 0.1× MMR for culture at14-22°C. mRNAs were injected in volumes ranging from 10-40 nl perblastomere, and in concentrations ranging from 50 pg-3 ng.Embryonic phenotypes were evaluated using a standard binoculardissecting microscope (Nikon, model SMZ-U). Anteroposterior anddorsoventral embryonic axis development was evaluated using theDAI index (Kao and Elinson, 1988).
RNA and in vitro transcriptionThe β-Eng (β-Engrailed) repressor chimera was constructed usingstandard and PCR-based subcloning techniques (Fig. 1A). A constructof Drosophila Engrailed was kindly provided by Heithem El-Hodiri(Jaynes and O’Farrell, 1991). A cDNA construct, β-cateninMT,containing β-catenin’s N-terminal and Armadillo domains andexcluding its C terminus, was obtained from pSP36T+MT (bp’s 1-2085) (Funayama et al., 1995) using PCR-based subcloning (forward:5′-AAG GAA AAA AGC GGC CGC G GCA ACT CAA GCA GAT-3′, reverse: 5′-ATT CCC G GGC CCG GTC TGC AGC CTC ATTCCA-3′). An N-terminal NotI site and a C-terminal ApaI site wereintroduced, and the construct inserted into pCDNA3.1+MT usingcorresponding NotI and ApaI sites, placing the construct immediatelydownstream and in-frame with a myc epitope tag. Next, the N-terminal repressor domain of Engrailed (amino acids 4-300) wasisolated via PCR, employing primers that incorporated ApaI sites intoboth flanking regions and the addition of a stop codon at the 3′ end(forward: 5′-ATT CCC GGG CCC AGG ATC GCT GCA GCC CA-3′, reverse: 5′-CCT AGG GAC ACG CTT ATT CCC GGG-3′). Thisrepressor fragment was blunt-end cloned into pCDNA3.1+MT(Invitrogen, Carlsbad, CA, USA) at the ApaI site, immediatelydownstream of the C-terminal truncated β-catenin. To facilitate the invitro transcription of RNA, the entire myc-tagged β-eng construct was
W. T. Montross, H. Ji and P. D. McCrea

1761β-Engrailed selectively suppresses Wnt signaling
moved into pCS2+ (Rupp et al., 1994) at a StuI site using blunt-endligation, creating β-engMT. The construct’s reading frame was
verified by cDNA sequencing at the institutional core sequencingfacility.
A non-epitope-tagged construct, β-eng, was produced using PCR,permitting the introduction of the restriction site, ClaI (5′ end) andEcoRI (3′ end) restriction sites (forward: 5′-CCATCGAT ATG GCAACT CAA GCA GAT-3′, reverse: 5′-CGGAATTC TTA CTT GCACAG GGA TCC-3′). A control construct consisting of only theEngrailed repressor, engMT, was generated using PCR (forward: 5′-GGGCCC AG GAT CGC TGC AGC CCA-3′; reverse: 5′GGGCCCTTA CTT GCA CAG GGA TTC-3′), introducing ApaI sites tofacilitate insertion into the StuI site of pCS2+MT expression vector.In addition, an untagged construct of the Engrailed repressor domain,engR, was generated via PCR amplification of the repressor region,introducing an 5′ BamHI site (forward: 5′-CG GGA TCC ATG GCCCTG GAG GAT C-3′) and a 3′ EcoRI site (reverse: 5′-G GAA TTCTTA CTT GCA CAG GGA TCC-3′) for the corresponding insertioninto pCS2+. Additionally, XTcf-3/HA, an epitope-tagged version ofXTcf-3 (a kind gift of the Clevers Laboratory, University of Utrecht,The Netherlands) was generated via N-terminal fusion with three HArepeats. The XTcf-3 cDNA was PCR subcloned from the originalplasmid, pCDNAXTcf-3, introducing 5′ and 3′ XbaI sites using theprimers, forward: 5′-GC TCT AGA ATG CCT CAA CTA AAC AGC-3′; reverse: 5′-GC TCT AGA TCA GTC ACT GGA TTT GGT-3′.This fragment was then inserted into pCS2+3HA downstream of threeHA repeats at a convenient XbaI site in the first polylinker. CappedmRNAs encoding the aforementioned constructs were generatedusing the SP6 mMessage mMachine Kit (Ambion, Austin, TX, USA).The cDNA templates were linearized for transcription using thefollowing enzymes: β-engMT (NsiI), β-eng (NsiI), engR (NsiI),engMT (NsiI), pCS2+MT (NotI), β-cateninMT (NotI) and XTcf-3/HA(NotI). The quantity and quality of transcribed RNA products wereevaluated on the basis of migration within 1% agarose formaldehydegels and by optical density (OD 280/260).
Western blots and immunoprecipitationTo evaluate exogenous protein expression following RNAmicroinjection and in vivo translation, embryos were injected at the1- and 2-cell stage with myc- and/or HA-tagged constructs. Embryoswere cultured until stage 9-10, lysed at 4°C in 15 mM Tris, pH 6.8,and extracted with Freon (1,1,2-trichlorotrifluoroethane) to removeyolk proteins. The lysis buffer was supplemented with a proteaseinhibitor cocktail consisting of 1 mM phenylmethylsulfonyl fluoride,4 µg/ml aprotinin, 1 µg/ml pepstatin A, 2 µg/ml leupeptin,10 µg/mlantipain, 50 µg/ml benzamide, 10 µg/ml soybean trypsin inhibitor,100 µg/ml iodoacetamide and 40 µg/ml TLCK. Immunoprecipitationof the myc-tagged constructs, C-cadherin and α-catenin wasperformed with stage 10-12 embryos extracted in 0.5% Triton X-100,10 mM Hepes, pH 7.4, 150 mM NaCl, 2 mM EDTA and 0.02% NaN3,in addition to the aforementioned protease inhibitors. The extract wascleared by centrifugation for 30 minutes, at 4°C, at 14,000 g.Immunoprecipitations of the myc-tagged constructs were performedwith a monoclonal anti-myc antibody 9E10, obtained from theDevelopmental Studies Hybridoma Bank maintained by theUniversity of Iowa. C-cadherin was immunoprecipitated with apolyclonal antibody generated against the extracellular domain ofXenopus C-cadherin (kind gift of W. Brieher and B. Gumbiner,Memorial Sloan-Kettering Cancer Center; Yap et al., 1997). α-Catenin was immunoprecipitated with a monoclonal antibody raisedagainst amino acids 729-906 of mouse α-catenin (TransductionLaboratories, Franklin Lakes, NJ, USA). Immunoprecipitation of thetagged XTcf-3 construct was performed with the monoclonal anti-HAantibody, 12CA5 (Wadzinski et al., 1992). In some experiments,Concanavalin A (Sigma, St Louis, MO, USA) conjugated to sepharosebeads, was used as a control for immunoprecipitation of membraneglycoproteins such as cadherin.
Proteins were resolved on 8% SDS-PAGE and transferred to 0.45µm nitrocellulose membranes. Whole cell lysates were loaded at 2
Fig. 1. Construction of the β-Eng Chimera. (A) The C-terminaltransactivation domain of a stabilized Xenopus β-catenin (*, S→A,T→A mutations in the N-terminal domain) was replaced with the N-terminal repressor domain of Drosophila Engrailed (amino acids 4-300) using PCR-based subcloning techniques (β-Eng). To facilitatebiochemical studies, six myc epitope tags were added to the Nterminus (β-EngMT). (B) Expression of β-Eng protein in Xenopusextracts from gastrula stage (stage 10-12) Xenopus embryos injectedat the 4-cell stage in the dorsal or ventral hemisphere, with either 3.0ng or 1.5 ng of β-engMT mRNA, and detected with either myc 9E10mAb (Top) or β-catenin pAb (Bottom). The bottom panel showsdetection of endogenous β-catenin along with β-EngMT proteins.(C) Immunoprecipitation analysis of gastrula stage (stage 10-12)Xenopus extracts from uninjected control (−) embryos and β-EngMT(+) expressing embryos. β-EngMT expressing embryos were injectedwith a total of 1 ng of mRNA at the 2-cell stage. The extracts wereimmunoprecipitated with antibodies against the β-catenin bindingpartners, β-catenin and C-cadherin, and also with the anti-myc 9E10antibody. β-EngMT precipitates were examined for C-cadherin andα-catenin binding (lane 1). α-catenin and C-cadherin association isdemonstrated in the α-cat blot and the C-cad blot (lanes 3-6). The α-catenin blot indicates a similar association between α-catenin and C-cadherin in both β-EngMT expressing and uninjected embryos (lanes5 and 6). The myc blot shows the association of β-EngMT with α-catenin (lane 3) and demonstrates the binding of β-EngMT by C-cadherin (lane 5). Antibody control appears in lane 7. Whole celllysate (WCL) positive controls for expression of α-catenin, C-cadherin and β-Eng appear in lanes 8 and 9.

1762
embryo equivalents per lane, while immunoprecipitates were loadedwith at least 2.5 embryo equivalents per lane. To detect proteins inwestern blots, the following antibodies were used at the followingdilutions: 9E10 anti-myc antibody at a 1:10,000 dilution; 12CA5antibody at 1:5,000; anti-α-catenin antibody at 1:2,500; anti-C-cadherin at 1:2,500. β-Eng constructs and endogenous β-catenin werefurther detected with a polyclonal anti-β-catenin antibody raisedagainst the N-terminal domain of Xenopus β-catenin (McCrea et al.,1993). Protein bands were visualized via enhanced chemiluminesence(ECL: Amersham, Buckinghamshire, UK) by reacting with either goatanti-rabbit IgG or goat anti-mouse IgG secondary antibodies (1:3000),each conjugated to horseradish peroxidase (Bio-Rad, Hercules, CA,USA).
Cell adhesion assaysCell adhesion assays were performed according to Torres et al. (1996),with the following minor modifications. Animal caps were excisedfrom stage 9 embryos and dissociated in 1× Ca2+/Mg2+-free modifiedBarth’s solution (CMF-MBS: 88 mM NaCl, 1 mM KCl, 2.4 mMNaHCO3, 10 mM Hepes, pH 7.5) for 1 hour in 35 mm Petri dishescoated with 1% agarose in the aforementioned buffer. Cells werereaggregated at room temperature on an orbital bench-top shaker at60 rpm for 1 hour in 2 mM Ca2+ in the CMF-MBS buffer. Cellreaggregation was observed by digital capture of random cellpopulations from each sample. Aggregates were scored using theparticle analysis utility of NIH Image 1.60 software (1998) (NIH,Bethesda, MD, USA) to scan the digitally captured images for small(5-8 cells) and large (>8 cells) aggregates (Torres et al., 1995).
RT-PCR analysisEmbryos were injected with a total of 0.5 ng of β-engMT mRNA, orβ-galactosidase (or left uninjected), at the 4-cell stage in either twodorsal or two ventral blastomeres, or in all four blastomeres. TotalRNA was obtained by extracting whole embryos cultured to stage 10-12 with Trizol (Gibco-BRL, Gaithersburg, MD, USA) according tothe manufacturer’s protocol, followed by the preparation of mRNAfor the reverse transcriptase reaction (Gibco-BRL). The total cDNAlevels per sample were normalized prior to PCR Supermix (Gibco-BRL) amplification. Controls for DNA contamination were evaluatedusing PCR in the absence of reverse transcriptase (RT−). Molecularmarker primer sequences employed are as follows: Siamois (Fagottoet al., 1997); Histone H4 (Niehrs et al., 1994); Chordin (Sasai et al.,1994); Xbra (Smith et al., 1991); Xnr-3 (Darras et al., 1997); ornithinedecarboxylase (Bassez et al., 1990).
Whole-mount in situ hybridizationWhole-mount in situ hybridizations were undertaken for themolecular marker goosecoid according to Harland (1991), with minormodifications according to El-Hodiri et al. (1997). Embryos wereevaluated at the gastrula stage.
Luciferase assaysLuciferase assays were performed according to the method ofBrannon et al. (1997; reporter plasmids were the kind gift of M.Brannon and David Kimelman, University of Washington). Theexperiments were repeated at least three times, and each assay wasperformed in triplicate. Two reporter plasmids were used: S01324 andS0, which each included the proximal −0.8 kb promoter of the siamoisgene. S01234 contains three Lef/Tcf binding sites (0, 1 and 3) whilethe S0 reporter, a negative control, is mutant at these sites and thusprohibited activation by the β-catenin/Tcf transcriptional machinery(Brannon et al., 1997).
Histology and immunohistochemistryEmbryos were cultured to the required stage for each experiment andfixed in MEMPFA (4% paraformaldehyde, 4 mM Mops, 2 mM EGTA,1 mM MgSO4, pH 7.4) for 1-2 hours at room temperature. Fixed
embryos underwent serial dehydration and were embedded in paraffinaccording to published procedures (Kelly, 1991), and sectioned on arotary microtome (Reichert HistoStat, model 820) at a thickness of 8µm. For visualization of the tissue, samples were stained withHematoxylin and Eosin according to the manufacturer’s instructions(Sigma). To examine the nuclear translocation of β-Eng, embryoswere injected with either 500 pg of β-engMT or 250 pg of pCS2+MT(myc only) mRNA in both blastomeres at the 2-cell stage and cultureduntil stage 9, then embedded in paraffin according to theaforementioned procedure. Paraffin-embedded sections were dewaxedand blocked with 1% BSA (bovine serum albumin) for 60 minutes,followed by overnight incubation with the anti-myc monoclonalantibody 9E10 at a dilution of 1:250 in the blocking solution.Resulting immunocomplexes were visualized using an alkalinephosphatase anti-alkaline phosphatase (APAAP) complex method,according to the manufacturer’s instructions (DAKO APAAP Kit-System 40 K670, DAKO Inc., Carpenteria, CA, USA). Histologicalsections were evaluated using a standard binocular microscope (ZeissAxioskop).
RESULTS
β-Eng is effectively expressed and associatesnormally with the Cadherin complex To verify expression of β-Eng in early Xenopus embryos, weemployed a polyclonal antibody raised against the N terminusof β-catenin (McCrea et al., 1993). A myc-epitope taggedversion of β-Eng (β-EngMT) was evaluated by westernanalysis of injected embryo extracts, revealing that β-EngMTwas effectively expressed in a dose-dependent manner (Fig.1B). The untagged version, β-Eng, was found to be similarlyexpressed in the early embryo.
To assess if β-Eng associated normally with components ofthe cadherin complex (cadherin<>β-catenin<>α-catenin), weconducted immunoprecipitation analysis. 2-cell stage embryoswere injected with a total of 1.0 ng of β-EngMT mRNA intoboth blastomeres and then assayed for the association of β-EngMT protein with endogenous C-cadherin. As anticipated, β-EngMT coprecipitated with endogenous C-cadherin (Fig. 1C)and vice versa. Con A (Sigma), a lectin that binds membraneglycoproteins such as cadherin, likewise efficientlycoimmunoprecipitated β-EngMT (data not shown).Coimmunoprecipitation experiments indicating that β-EngMTcoprecipitated α-catenin and vice versa, confirmed that β-EngMT associated properly with its known binding partnerswithin the cadherin complex (Fig. 1C). Finally, it wasdemonstrated that β-EngMT held no selective advantage ordisadvantage over endogenous β-catenin for incorporationinto the cadherin complex, as indicated in thecoimmunoprecipitation of C-cadherin and α-catenin. Embryosinjected with β-EngMT compared to uninjected embryoscontained equivalent amounts of cadherin complexes measuredas coimmunoprecipitates of C-cadherin in which α-catenin isdetected via western analysis (Fig. 1C, α-catenin blot: lanes 5and 6) and vice versa (Fig. 1C, C-cadherin blot: lanes 3 and 4).
Expression of β-Eng prevents dorsal development inXenopus embryosTo examine whether β-Eng could inhibit Wnt signalingrequired for the formation of the dorsal axis, the chimera wasexpressed in the organizer region of early Xenopus embryos.
W. T. Montross, H. Ji and P. D. McCrea
1763β-Engrailed selectively suppresses Wnt signaling
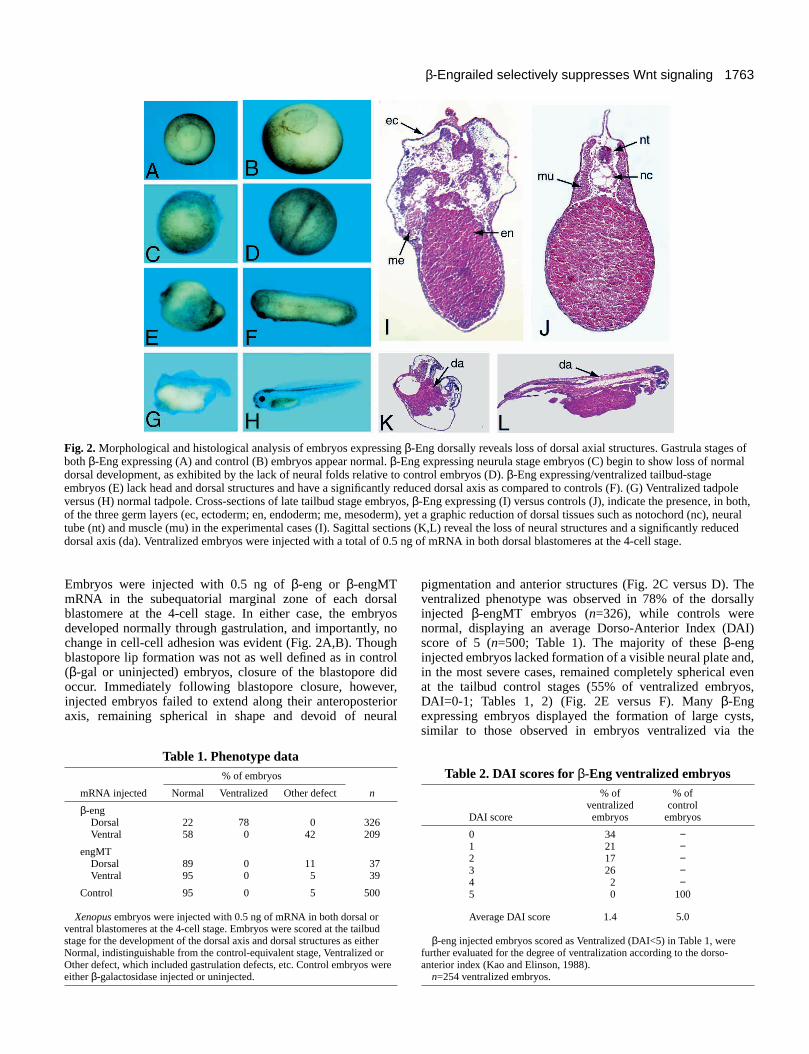
Embryos were injected with 0.5 ng of β-eng or β-engMTmRNA in the subequatorial marginal zone of each dorsalblastomere at the 4-cell stage. In either case, the embryosdeveloped normally through gastrulation, and importantly, nochange in cell-cell adhesion was evident (Fig. 2A,B). Thoughblastopore lip formation was not as well defined as in control(β-gal or uninjected) embryos, closure of the blastopore didoccur. Immediately following blastopore closure, however,injected embryos failed to extend along their anteroposterioraxis, remaining spherical in shape and devoid of neural
pigmentation and anterior structures (Fig. 2C versus D). Theventralized phenotype was observed in 78% of the dorsallyinjected β-engMT embryos (n=326), while controls werenormal, displaying an average Dorso-Anterior Index (DAI)score of 5 (n=500; Table 1). The majority of these β-enginjected embryos lacked formation of a visible neural plate and,in the most severe cases, remained completely spherical evenat the tailbud control stages (55% of ventralized embryos,DAI=0-1; Tables 1, 2) (Fig. 2E versus F). Many β-Engexpressing embryos displayed the formation of large cysts,similar to those observed in embryos ventralized via the
Fig. 2. Morphological and histological analysis of embryos expressing β-Eng dorsally reveals loss of dorsal axial structures. Gastrula stages ofboth β-Eng expressing (A) and control (B) embryos appear normal. β-Eng expressing neurula stage embryos (C) begin to show loss of normaldorsal development, as exhibited by the lack of neural folds relative to control embryos (D). β-Eng expressing/ventralized tailbud-stageembryos (E) lack head and dorsal structures and have a significantly reduced dorsal axis as compared to controls (F). (G) Ventralized tadpoleversus (H) normal tadpole. Cross-sections of late tailbud stage embryos, β-Eng expressing (I) versus controls (J), indicate the presence, in both,of the three germ layers (ec, ectoderm; en, endoderm; me, mesoderm), yet a graphic reduction of dorsal tissues such as notochord (nc), neuraltube (nt) and muscle (mu) in the experimental cases (I). Sagittal sections (K,L) reveal the loss of neural structures and a significantly reduceddorsal axis (da). Ventralized embryos were injected with a total of 0.5 ng of mRNA in both dorsal blastomeres at the 4-cell stage.
Table 1. Phenotype data% of embryos
mRNA injected Normal Ventralized Other defect n
β-engDorsal 22 78 0 326Ventral 58 0 42 209
engMTDorsal 89 0 11 37Ventral 95 0 5 39
Control 95 0 5 500
Xenopus embryos were injected with 0.5 ng of mRNA in both dorsal orventral blastomeres at the 4-cell stage. Embryos were scored at the tailbudstage for the development of the dorsal axis and dorsal structures as eitherNormal, indistinguishable from the control-equivalent stage, Ventralized orOther defect, which included gastrulation defects, etc. Control embryos wereeither β-galactosidase injected or uninjected.
Table 2. DAI scores for β-Eng ventralized embryos% of % of
ventralized control DAI score embryos embryos
0 34 −1 21 −2 17 −3 26 −4 2 −5 0 100
Average DAI score 1.4 5.0
β-eng injected embryos scored as Ventralized (DAI<5) in Table 1, werefurther evaluated for the degree of ventralization according to the dorso-anterior index (Kao and Elinson, 1988).
n=254 ventralized embryos.

1764
antisense depletion of β-catenin mRNA (Heasman et al., 1994).Less severely affected embryos were nonetheless ventralized,with obvious perturbations of anterior and dorsal structures,including a crude and diminutive dorsal fin (43% of ventralizedembryos, DAI=2-3; Table 2). Overall, β-Eng expressingembryos have been classified with a DAI score of 4 or less(average DAI=1.4; Table 2; Kao and Elinson, 1988). We expectthat the expression of β-eng mRNA in dorsal blastomeresspecifically inhibits Wnt signaling, and thus dorsal axisdevelopment in Xenopus embryos. While injection of threecontrol constructs for the Engrailed repression domain yieldedsome nonspecific gastrulation abnormalities (distinct fromthose observed in β-Eng expressing embryos), less than 1%produced ventralized embryos (Table 1).
β-Eng does not significantly alter Ca2+-dependentcellular adhesionTo rule out the possibility that the observed phenotype mightresult in part from β-Eng’s perturbation of cadherin-mediatedcell-cell adhesion, Ca2+-dependent cellular adhesion assayswere performed. Embryos were injected after the first cleavagein the animal pole with either β-eng or β-catenin mRNA inboth blastomeres. The embryos were cultured to stage 9 andthen animal caps expressing β-Eng, β-catenin or uninjectedcontrols were each dissociated in a Ca2+-free medium andreaggregated in the presence or absence of Ca2+ (Torres et al.,1995). Relative cell-cell adhesion was evaluated by comparingthe ability of the cells to form small (5-8 cells) versus large(greater than 8 cells) aggregates in the presence of Ca2+
(Kemler et al., 1989). As indicated in Table 3, the ability ofcells expressing β-Eng to form either small or large aggregatesis not significantly altered in comparison to β-cateninexpressing or uninjected cells. The total number of aggregatesformed in the Ca2+(+) case is similar in each assay (Table 3,total aggregates: β-Eng, 98; β-catenin, 97; control, 103). As foruninjected and β-catenin expressing cells (Table 3, control andβ-catenin), the addition of Ca2+ significantly increased cellaggregation in β-Eng expressing cells (Aberle et al., 1996;Fagotto and Gumbiner, 1994), demonstrating the restoration ofcadherin-mediated cellular adhesion. Even cells injected witha twofold increase over the ventralizing dose of β-eng mRNA(1.0 ng) formed aggregates to a similar extent in the presenceof Ca2+ (Table 3, β-Eng).
In further support of the hypothesis that β-Eng does not altercadherin-mediated cell-cell adhesion, it should also be noted
that there are no observable defects in β-Eng expressingembryos as they develop and undergo gastrulation, adevelopmental process known from numerous studies to besensitive to changes in cell-cell adhesion (Kintner, 1992;Levine et al., 1994). Furthermore, manual dissection andsectioning of embryos expressing β-Eng does not reveal anygross internal perturbations of blastomere adhesion. Theseobservations are in contrast to embryos expressing p120catenin or various other cadherin constructs, such as adominant-negative cadherin, which failed gastrulation and hadreadily observable deficiencies in cell-cell adhesion (Lee andGumbiner, 1995; Paulson et al., 1999).
Taken together, the adhesion data suggests that β-Eng doesnot significantly modify cell-cell adhesion by perturbing thecadherin complex, further supporting the hypothesis that β-Eng’s effects are primarily mediated via its suppression of β-catenin-dependent Wnt signaling.
Histological analysis reveals tissue differentiationand loss of dorsal morphology in β-Eng expressingembryosHistological analysis of dorsally injected β-eng embryosrevealed that the three germ layers were present (endo-, meso-and ectoderm; Fig. 2I), while formation of dorsal structuressuch as neural tube, notochord, and somites was significantlyperturbed, and in severe cases, inhibited (Fig. 2I versus J). Incontrol-equivalent tailbud stage embryos (injected dorsallywith β-Eng; DAI<4), anterior structures such as eyes, cementgland and head, were significantly reduced, and histologicalsections revealed severely perturbed neural structures (Fig.2I,K versus J,L) and a greatly reduced dorsal axis. Theseobservations are in congruence with the hypothesis that β-Engis interfering with organizer-dependent dorsalization, but notspecification of the germ layers. Further, histology supports ourmorphological and molecular work indicating that β-Engsuppresses dorsal axial structures and expression of thecorresponding molecular markers.
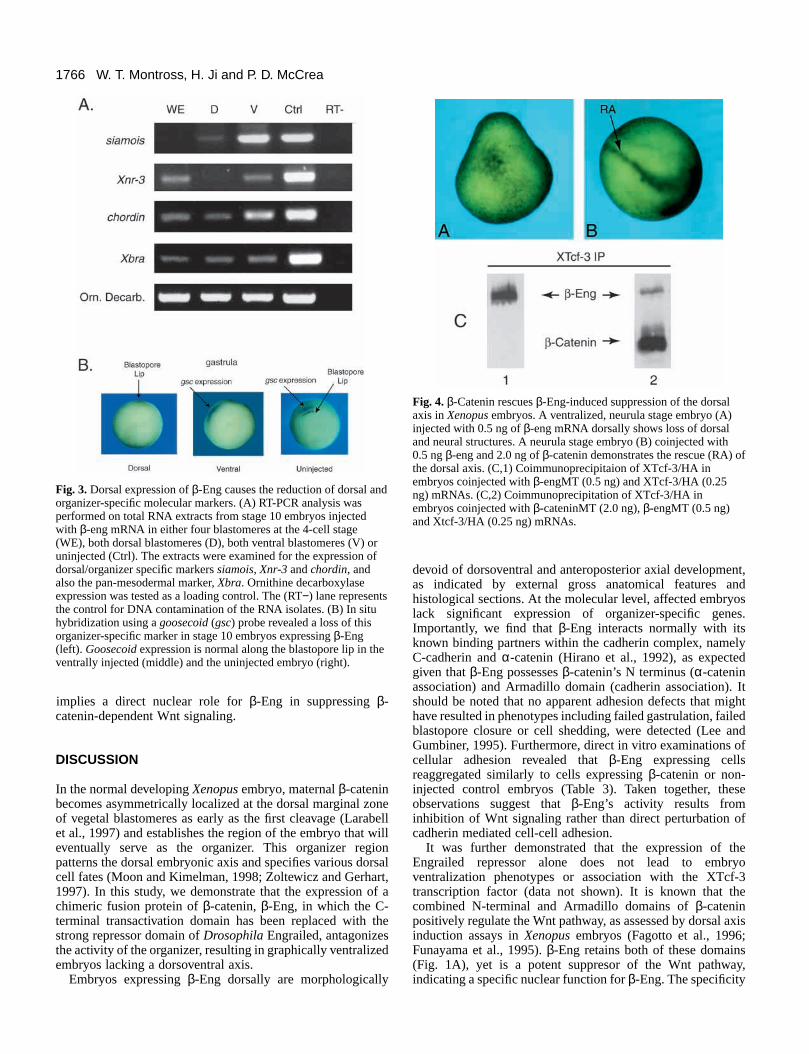
β-Eng expression results in the reduction of dorsaland organizer specific molecular markersTo test for the inhibition of Wnt signaling at the molecularlevel, we examined the effect of expressing β-Eng onorganizer-specific gene expression (Dawid, 1994). Expressionof siamois, a homeobox gene and direct target of Wnt/β-catenin signaling, was evaluated using RT-PCR of stage 10embryos that had been injected with 0.25 ng of β-eng mRNAat the 4-cell stage in each dorsal blastomere (0.50 ng total).Embryos expressing β-Eng protein showed a marked decreaseof siamois when compared to β-galactosidase injected anduninjected controls (Fig. 3A). The organizer specific marker,Xnr-3, a member of the TGFβ superfamily and direct target ofWnt/β-catenin signaling, and chordin, another organizer/dorsal-specific marker, were also significantly reduced in β-engdorsally injected embryos (Fig. 3A). Additionally, expressionof β-Eng led to a marked reduction in the pan-mesodermalmarker, Xbra, indicating a concomitant loss of mesoderminduction with the inhibition of axis formation (Fig. 3A).
Using whole-mount in situ hybridization methods (Harland,1991), we further tested if goosecoid, an indirect target ofWnt/β-catenin signaling, was repressed. Goosecoid is anorganizer-specific marker normally expressed along the dorsal
W. T. Montross, H. Ji and P. D. McCrea
Table 3. Adhesion assays: cell adhesion analysismRNA injected
β-eng β-catenin Control
Ca2+: + − + − + −Number of aggregates
5-8 cell 25 12 36 31 35 18>8 cell 73 2 61 19 68 17
Total aggregates 98 14 97 50 103 35
Two-cell embryos were injected with a total of either 1 ng β-eng mRNA, 2ng of β-catenin mRNA, or were uninjected (control). All assays wereperformed in both the presence (+) and absence (−) of calcium. The valuesindicate the number of aggregates of given sizes counted in three separatesamples of each assay. Total aggregates, the number of aggregates counted inthree separate samples of each assay.

1765β-Engrailed selectively suppresses Wnt signaling
blastopore lip of gastrulating embryos (Fig. 3B). Embryos atthe 4-cell stage injected with 0.5 ng of β-eng into both dorsalor ventral blastomeres were probed with a digoxigin-labeledgoosecoid probe (Harland, 1991). Dorsally injected embryosshowed a dramatic decrease in goosecoid expression along thedorsal blastopore lip, reflecting the noted loss of organizer-specific gene expression (Fig. 3B). Ventrally injected embryos,along with uninjected controls, demonstrated no loss ofgoosecoid expression at the gastrula stage (Fig. 3B). Later indevelopment, goosecoid is normally expressed in anterior/headstructures and along the dorsal aspect of the embryo. Atcontrol-equivalent tailbud stages, β-eng injected embryos stillshowed a loss of goosecoid expression, while no loss wasapparent in the ventrally injected and control embryos (data notshown). In summary, the reduction of siamois, chordin,goosecoid and Xnr-3 expression in β-eng injected embryosis consistent with the morphological loss of dorsoanterioraxial structures (embryonic ventralization), indicating directeffects upon Wnt/β-catenin-dependent signaling and axisinduction.
Exogenous β-catenin restores axis development inβ-eng injected embryosβ-catenin-mediated rescue of normal axial development inembryos in which Wnt signals have been suppressed providesa powerful specificity test for Wnt pathway inhibitors(Heasman et al., 1994; Kessler, 1997). If β-Eng directlyinterferes with the normal activity of β-catenin-dependent Wntsignaling within the dorsal organizer, then exogenous β-cateninwould be anticipated to restore normal axial development.Indeed, coexpression of 2 ng of β-catenin with 0.5 ng of β-engresults in normal axial development in over 90% (n=49) ofinjected embryos (Table 4; Fig. 4A versus B), indicatingthat β-Eng specifically suppresses β-catenin-mediated Wntsignaling.
To further investigate the mechanism by which this rescuetakes place, stage 10 embryos expressing either β-Eng (0.5 ngof mRNA) and XTcf-3 (0.25 ng mRNA), or β-Eng (0.5 ng),XTcf-3 (0.25 ng) and β-catenin (2.0 ng), were assayed viaprotein coimmunoprecipitation (Fig. 4C, see also Fig. 6B). Themolecules were epitope tagged (β-Eng and β-catenin with myc;XTcf-3 with HA) to facilitate the selective biochemical assayof the exogenous proteins and thereby provide an assessmentof their relative extent of association. Immunoprecipitation ofXTcf-3 indicated that it binds β-Eng (Fig. 4C,1). To asignificant extent, coinjection of β-catenin with β-eng revealedβ-catenin’s competition for XTcf-3, illustrated by the relativereduction of β-Eng protein coprecipitating with XTcf-3 (Fig.4C,2). This evidence suggests that the β-catenin-mediatedrescue of β-eng injected embryos occurs via the displacement/competition of β-Eng from XTcf-3, and that β-Eng’s repressiveaction upon Wnt/β-catenin gene targets is specific.
The specificity of β-Eng in suppression of β-catenin/Wnt signaling is supported by luciferasereporter assays of the known Wnt target, siamoisTo further address the specificity and efficacy of β-Eng ininhibiting β-catenin-dependent Wnt signaling, we testedsuppression of transcription from siamois reporter constructs.These reporters, generously provided by the laboratory ofDavid Kimelman (University of Washington), consisted of the
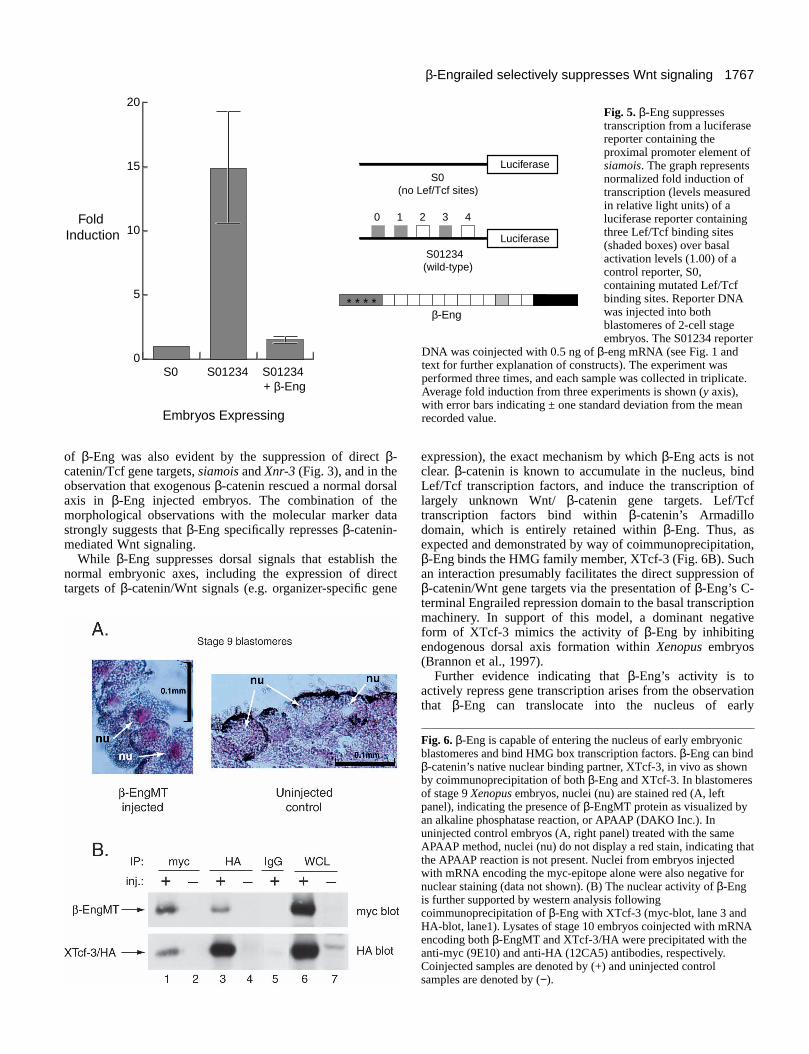
–0.8 kb proximal promoter of the siamois gene (Brannon et al.,1997). The first construct, S01234, included three Tcf bindingsites required for gene activation via β-catenin/Tcf and twogeneral enhancing elements. The second construct, S0, wasmutant in all five of these binding sites, and served as a negativecontrol (Fig. 5). The reporters were injected into the vegetalhemisphere of dorsal blastomeres, a region of high endogenousβ-catenin signaling. Dorsal injection of the S0 reporter plasmidDNA established the basal level of luciferase transcription,while dorsal injection of S01234 resulted in nearly a 15-foldinduction of luciferase activity over basal levels. Coexpressionof the S01234 reporter with β-Eng abolished luciferase activity,resulting in an overall induction of only a fraction overbasal levels (Fig. 5). Thus, β-Eng effectively suppressed theexpression of an established target of β-catenin/Wnt signaling,likely via a direct repressive effect upon its promoter.
Nuclear translocation of β-EngThus far, we have provided evidence that β-Eng specificallysuppresses Wnt/β-catenin transcription. To further assess β-Eng’s function, we tested if β-Eng entered the nucleus to bindXTcf-3. Using immunohistochemical techniques, we foundthat Xenopus embryos injected with β-engMT mRNA stainedpositively for the inclusion of β-Eng within their nuclei (Fig.6A, left panel). When compared to control embryos, eitheruninjected (Fig. 6A, right panel), or with molar equivalents ofmRNA encoding only the myc tag (data not shown), nuclearstaining was not detected (Fig. 6A, right panel), indicating thatthe β-Eng protein is capable of translocating to the nucleus tosuppress Wnt/β-catenin-mediated transcription. This resultwas anticipated because β-Eng retains β-catenin’s entireArmadillo domain, which can bind Lef/Tcf transcriptionfactors harboring nuclear localization sequences (NLS) (Huberet al., 1996). Furthermore, it has been shown that β-catenin’sArmadillo repeat domain is the minimal structural requirementfor its NLS-independent (Lef/Tcf independent) and importin-independent nuclear localization (Fagotto et al., 1998;Funayama et al., 1995).
β-Eng’s capacity to bind XTcf-3 in vivo is again supportedvia western analysis, demonstrating a reciprocal associationbetween the two proteins (Fig. 6B). Embryos were coinjectedwith 500 pg of β-engMT mRNA and 250 pg of XTcf-3/HA inboth blastomeres at the 2-cell stage. At stage 9, the embryoswere lysed and coimmunoprecipitated using the monoclonalanti-HA 12CA5 antibody and the anti-myc 9E10 antibody.Western analysis of the coprecipitation revealed a strong andreciprocating association between β-Eng and XTcf-3. Thisevidence, taken along with the nuclear immunohistochemistry,
Table 4. β-Catenin rescues β-Eng induced dorsal axissuppression
% of embryos
mRNA injected Normal Abnormal Ventralized Rescued n
β-eng 18 0 82 − 57β-eng/β-catenin 91 9 0 91 45Control 87 13 0 − 121
Xenopus embryos injected with 0.5 ng of mRNA dorsally, yieldedventralized phenotypes (Fig. 4A). Coinjection of 0.5 ng of β-eng mRNA with2.0 ng of β-catenin mRNA restored the dorsal axis (RA) in over 90% of theinjected embryos (Fig. 4B).
1766
implies a direct nuclear role for β-Eng in suppressing β-catenin-dependent Wnt signaling.
DISCUSSION
In the normal developing Xenopus embryo, maternal β-cateninbecomes asymmetrically localized at the dorsal marginal zoneof vegetal blastomeres as early as the first cleavage (Larabellet al., 1997) and establishes the region of the embryo that willeventually serve as the organizer. This organizer regionpatterns the dorsal embryonic axis and specifies various dorsalcell fates (Moon and Kimelman, 1998; Zoltewicz and Gerhart,1997). In this study, we demonstrate that the expression of achimeric fusion protein of β-catenin, β-Eng, in which the C-terminal transactivation domain has been replaced with thestrong repressor domain of Drosophila Engrailed, antagonizesthe activity of the organizer, resulting in graphically ventralizedembryos lacking a dorsoventral axis.
Embryos expressing β-Eng dorsally are morphologically
devoid of dorsoventral and anteroposterior axial development,as indicated by external gross anatomical features andhistological sections. At the molecular level, affected embryoslack significant expression of organizer-specific genes.Importantly, we find that β-Eng interacts normally with itsknown binding partners within the cadherin complex, namelyC-cadherin and α-catenin (Hirano et al., 1992), as expectedgiven that β-Eng possesses β-catenin’s N terminus (α-cateninassociation) and Armadillo domain (cadherin association). Itshould be noted that no apparent adhesion defects that mighthave resulted in phenotypes including failed gastrulation, failedblastopore closure or cell shedding, were detected (Lee andGumbiner, 1995). Furthermore, direct in vitro examinations ofcellular adhesion revealed that β-Eng expressing cellsreaggregated similarly to cells expressing β-catenin or non-injected control embryos (Table 3). Taken together, theseobservations suggest that β-Eng’s activity results frominhibition of Wnt signaling rather than direct perturbation ofcadherin mediated cell-cell adhesion.
It was further demonstrated that the expression of theEngrailed repressor alone does not lead to embryoventralization phenotypes or association with the XTcf-3transcription factor (data not shown). It is known that thecombined N-terminal and Armadillo domains of β-cateninpositively regulate the Wnt pathway, as assessed by dorsal axisinduction assays in Xenopus embryos (Fagotto et al., 1996;Funayama et al., 1995). β-Eng retains both of these domains(Fig. 1A), yet is a potent suppresor of the Wnt pathway,indicating a specific nuclear function for β-Eng. The specificity
W. T. Montross, H. Ji and P. D. McCrea
Fig. 3. Dorsal expression of β-Eng causes the reduction of dorsal andorganizer-specific molecular markers. (A) RT-PCR analysis wasperformed on total RNA extracts from stage 10 embryos injectedwith β-eng mRNA in either four blastomeres at the 4-cell stage(WE), both dorsal blastomeres (D), both ventral blastomeres (V) oruninjected (Ctrl). The extracts were examined for the expression ofdorsal/organizer specific markers siamois, Xnr-3 and chordin, andalso the pan-mesodermal marker, Xbra. Ornithine decarboxylaseexpression was tested as a loading control. The (RT−) lane representsthe control for DNA contamination of the RNA isolates. (B) In situhybridization using a goosecoid (gsc) probe revealed a loss of thisorganizer-specific marker in stage 10 embryos expressing β-Eng(left). Goosecoid expression is normal along the blastopore lip in theventrally injected (middle) and the uninjected embryo (right).
Fig. 4. β-Catenin rescues β-Eng-induced suppression of the dorsalaxis in Xenopus embryos. A ventralized, neurula stage embryo (A)injected with 0.5 ng of β-eng mRNA dorsally shows loss of dorsaland neural structures. A neurula stage embryo (B) coinjected with0.5 ng β-eng and 2.0 ng of β-catenin demonstrates the rescue (RA) ofthe dorsal axis. (C,1) Coimmunoprecipitaion of XTcf-3/HA inembryos coinjected with β-engMT (0.5 ng) and XTcf-3/HA (0.25ng) mRNAs. (C,2) Coimmunoprecipitation of XTcf-3/HA inembryos coinjected with β-cateninMT (2.0 ng), β-engMT (0.5 ng)and Xtcf-3/HA (0.25 ng) mRNAs.
1767β-Engrailed selectively suppresses Wnt signaling
of β-Eng was also evident by the suppression of direct β-catenin/Tcf gene targets, siamois and Xnr-3 (Fig. 3), and in theobservation that exogenous β-catenin rescued a normal dorsalaxis in β-Eng injected embryos. The combination of themorphological observations with the molecular marker datastrongly suggests that β-Eng specifically represses β-catenin-mediated Wnt signaling.
While β-Eng suppresses dorsal signals that establish thenormal embryonic axes, including the expression of directtargets of β-catenin/Wnt signals (e.g. organizer-specific gene
expression), the exact mechanism by which β-Eng acts is notclear. β-catenin is known to accumulate in the nucleus, bindLef/Tcf transcription factors, and induce the transcription oflargely unknown Wnt/ β-catenin gene targets. Lef/Tcftranscription factors bind within β-catenin’s Armadillodomain, which is entirely retained within β-Eng. Thus, asexpected and demonstrated by way of coimmunoprecipitation,β-Eng binds the HMG family member, XTcf-3 (Fig. 6B). Suchan interaction presumably facilitates the direct suppression ofβ-catenin/Wnt gene targets via the presentation of β-Eng’s C-terminal Engrailed repression domain to the basal transcriptionmachinery. In support of this model, a dominant negativeform of XTcf-3 mimics the activity of β-Eng by inhibitingendogenous dorsal axis formation within Xenopus embryos(Brannon et al., 1997).
Further evidence indicating that β-Eng’s activity is toactively repress gene transcription arises from the observationthat β-Eng can translocate into the nucleus of early
β-Eng* * * *
Luciferase S0(no Lef/Tcf sites)
Luciferase S01234(wild-type)
0 1 2 3 4
0
5
10
15
20
S01234 + β-Eng
S01234S0
Fold Induction
Embryos Expressing
Fig. 5. β-Eng suppressestranscription from a luciferasereporter containing theproximal promoter element ofsiamois. The graph representsnormalized fold induction oftranscription (levels measuredin relative light units) of aluciferase reporter containingthree Lef/Tcf binding sites(shaded boxes) over basalactivation levels (1.00) of acontrol reporter, S0,containing mutated Lef/Tcfbinding sites. Reporter DNAwas injected into bothblastomeres of 2-cell stageembryos. The S01234 reporter
DNA was coinjected with 0.5 ng of β-eng mRNA (see Fig. 1 andtext for further explanation of constructs). The experiment wasperformed three times, and each sample was collected in triplicate.Average fold induction from three experiments is shown (y axis),with error bars indicating ± one standard deviation from the meanrecorded value.
Fig. 6. β-Eng is capable of entering the nucleus of early embryonicblastomeres and bind HMG box transcription factors. β-Eng can bindβ-catenin’s native nuclear binding partner, XTcf-3, in vivo as shownby coimmunoprecipitation of both β-Eng and XTcf-3. In blastomeresof stage 9 Xenopus embryos, nuclei (nu) are stained red (A, leftpanel), indicating the presence of β-EngMT protein as visualized byan alkaline phosphatase reaction, or APAAP (DAKO Inc.). Inuninjected control embryos (A, right panel) treated with the sameAPAAP method, nuclei (nu) do not display a red stain, indicating thatthe APAAP reaction is not present. Nuclei from embryos injectedwith mRNA encoding the myc-epitope alone were also negative fornuclear staining (data not shown). (B) The nuclear activity of β-Engis further supported by western analysis followingcoimmunoprecipitation of β-Eng with XTcf-3 (myc-blot, lane 3 andHA-blot, lane1). Lysates of stage 10 embryos coinjected with mRNAencoding both β-EngMT and XTcf-3/HA were precipitated with theanti-myc (9E10) and anti-HA (12CA5) antibodies, respectively.Coinjected samples are denoted by (+) and uninjected controlsamples are denoted by (−).

1768
blastomeres, as revealed by immunohistochemical techniques(Fig. 6A). This result was also expected because β-Eng retainsa complete Armadillo repeat region capable of binding both theNLS-bearing Lef/Tcf factors (Huber et al., 1996), and nuclearpore components via NLS and importin-independentmechanisms (Fagotto et al., 1998). Collectively, thecoimmunoprecipitation data and the nuclear translocationevidence strengthens the argument that β-Eng possesses aspecific nuclear activity.
We have demonstrated that β-Eng suppresses β-catenin-dependent Wnt signaling, as shown by the morphologicalventralization of Xenopus embryos in addition to β-Eng’ssuppression of dorsal and organizer-specific molecular markerssuch as Xnr-3, siamois, goosecoid and chordin. While both thesignaling and adhesive function of β-catenin appear to beindependent (Heasman et al., 1994; Peifer, 1995), selectivesuppression of β-catenin’s signaling function has previouslybeen difficult to achieve. We expect that β-Eng will proveuseful to various researchers working upon β-catenin-mediatedWnt signaling. For example, unpublished work in ourlaboratory includes use of β-Eng to inhibit β-catenin-dependent signaling in ventral Xenopus development andsuggests that β-catenin may play a significant role in thepatterning and formation of ventral structures. Unlike the useof antisense technology to deplete signaling pools of β-cateninwithin the entire embryo (Heasman et al., 1994), β-Eng maybe expressed within discrete tissues or regions in the contextof an otherwise normal embryo. Further, work in progress hasdemonstrated that β-Eng effectively suppresses β-catenin-mediated Wnt signaling in mammalian cells (canine MDCKkidney cells), and we thus expect that its tissue-specificexpression will prove useful in a number of model animalsystems, and perhaps even in the suppression of pathologicalWnt signaling contributing to the progression of variouscarcinomas (Bullions and Levine, 1998; Miller et al., 1999;Peifer, 1997; Polakis, 1999).
We are grateful to D. Kimelman and M. Brannon (University ofWashington) for the gift of the Siamois/Luciferase Reporterconstructs, to B. Gumbiner and to W. Brieher (Memorial Sloan-Kettering Cancer Center) for the C-cadherin polyclonal antibody, andto L. Etkin and H. El-Hodiri (The University of Texas M. D. AndersonCancer Center) for the Drosophila Engrailed DNA. We also gratefullyacknowledge the current and past sources of support that havecontributed to this work: NIH RO1 Grant GM 52112; Texas ARPGrant 15-135; March of Dimes Basil O’Connor Grant 5-0926;Kleberg Foundation Award; CCSG Developmental Funds CCSG-CA16672 and University of Texas M. D. Anderson Cancer Center NCIcore grant CA-16672. W. Montross was partially supported by aMinority Research Supplement to NIH RO1 Grant GM 52112.
REFERENCES
Aberle, H., Bauer, A., Stappert, J., Kispert, A. and Kemler, R. (1997). β-catenin is a target for the ubiquitin-proteasome pathway. EMBO J. 16, 3797-3804.
Aberle, H., Schwartz, H. and Kemler, R. (1996). Cadherin-catenin complex– protein interactions and their implications for cadherin function. J. Cell.Biochem. 61, 514-523.
Alattia, J. R., Kurokawa, H. and Ikura, M. (1999). Structural view ofadherin-mediated cell-cell adhesion. Cell. Mol. Life Sci. 55, 359-367.
Barker, N., Morin, P. J. and Clevers, H. (2000). The Yin-Yang of TCF/β-catenin signaling. Adv. Cancer Res. 77, 1-24.
Barth, A. I., Nathke, I. S. and Nelson, W. J. (1997). Cadherins, catenins andAPC protein: interplay between cytoskeletal complexes and signalingpathways. Curr. Opin. Cell Biol. 9, 683-690.
Barth, A. I. M., Pollack, A. L., Altschuler, Y., Mostov, K. E. and Nelson,W. J. (1996). NH2-terminal deletion of β-catenin results in stablecolocalization of mutant β-catenin with adenamatous polyposis coli proteinand altered MDCK cell adhesion. J. Cell Biol. 136, 693-706.
Bassez, T., Paris, J., Omilli, F., Dorel, C. and Osborne, H. B. (1990). Post-transcriptional regulation of ornithine decarboxylase in Xenopus laevisoocytes. Development 110, 955-962.
Behrens, J., von Kries, J. P., Kuhl, M., Bruhn, L., Wedlich, D., Grosschedl,R. and Birchmeier, W. (1996). Functional interaction of β-catenin with thetranscription factor LEF-1. Nature 382, 638-642.
Bienz, M. (1999). APC: the plot thickens. Curr. Opin. Genet. Dev. 9, 595-603.Boutros, M. and Mlodzik, M. (1999). Dishevelled: at the crossroads of
divergent intracellular signaling pathways. Mech. Dev. 83, 27-37.Brannon, M., Gomperts, M., Sumoy, L., Moon, R. T. and Kimelman, D.
(1997). A β-catenin/XTcf-3 complex binds to the siamois promoterto regulate dorsal axis specification in Xenopus. Genes Dev. 11, 2359-2370.
Bullions, L. C. and Levine, A. J. (1998). The role of β-catenin in celladhesion, signal transduction, and cancer. Curr. Opin. Oncol. 10, 81-87.
Cavallo, R., Rubenstein, D. and Peifer, M. (1997). Armadillo and dTCF: amarriage made in the nucleus. [Review] Curr. Opin. Genet. Dev. 7, 459-466.
Cho, K. W. Y., Blumberg, B., Steinbeisser, H. and De Robertis, E. M.(1991). Molecular nature of Spemann’s organizer: The role of the Xenopushomeobox gene goosecoid. Cell 67, 1111-1120.
Christian, J. L. and Moon, R. T. (1993). Interactions between Xwnt-8 andSpemann organizer signaling pathways generate dorsoventral pattern in theembryonic mesoderm of Xenopus. Genes Dev. 7, 13-28.
Clevers, H. and van de Wetering, M. (1997). TCF/LEF factor earn theirwings. Trends Genet. 13, 485-489.
Conlon, F. L., Sedgwick, S. G., Weston, K. M. and Smith, J. C. (1996).Inhibition of Xbra transcription activation causes defects in mesodermalpatterning and reveals autoregulation of Xbra in dorsal mesoderm.Development 122, 2427-2435.
Cox, R. T., Kirkpatrick, C. and Peifer, M. (1996). Armadillo is required foradherens junction assembly, cell polarity, and morphogenesis duringDrosophila embryogenesis. J. Cell Biol. 134, 133-148.
Dale, T. C. (1998). Signal transduction by the Wnt family of ligands. Biochem.J. 329, 209-223.
Darras, S., Marikawa, Y., Elinson, R. P. and Lemaire, P. (1997). Animaland vegetal pole cells of early Xenopus embryos respond differently tomaternal dorsal determinants: implications for the patterning of theorganizer. Development 124, 4275-4286.
Dawid, I. B. (1994). Intercellular signaling and gene regulation during earlyembryogenesis of Xenopus laevis. J. Biol. Chem. 269, 6259-6262.
Deardorff, M. A., Tan, C., Conrad, L. J. and Klein, P. S. (1998). Frizzled-8 is expressed in the Spemann organizer and plays a role in earlymorphogenesis. Development 125, 2687-700.
Dierick, H. and Bejsovec, A. (1999). Cellular mechanisms of wingless/Wntsignal transduction. Curr. Top. Dev. Biol. 43, 153-190.
Dominguez, I., Itoh, K. and Sokol, S. Y. (1995). Role of Glycogen SynthaseKinase-3β as a negative regulator of dorsoventral axis formation in Xenopusembryos. Proc. Natl. Acad. Sci. USA 92, 8498-8502.
Easwaran, V., Song, V., Polakis, P. and Byers, S. (1999). The ubiquitin-proteasome pathway and serine kinase activity modulate adenomatouspolyposis coli protein-mediated regulation of β-catenin- lymphocyteenhancer-binding factor signaling. J. Biol. Chem. 274, 16641-1665.
El-Hodiri, H. M., Shou, W. and Etkin, L. D. (1997). Xnf7 functions indorsal-ventral patterning of the Xenopus embryo. Dev. Biol. 190, 1-17.
Fagotto, F., Funayama, N., Gluck, U. and Gumbiner, B. M. (1996). Bindingto cadherins antagonizes the signaling activity of β-catenin during axisformation in Xenopus. J. Cell Biol. 132, 1105-1114.
Fagotto, F., Gluck, U. and Gumbiner, B. M. (1998). Nuclear localizationsignal-independent and importin/karyopherin- independent nuclear importof β-catenin. Curr. Biol. 8, 181-90.
Fagotto, F., Guger, K. and Gumbiner, B. M. (1997). Induction of the primarydorsalizing center in Xenopus by the Wnt/GSK/β-catenin signaling pathway,but not by Vg1, Activin or Noggin. Development 124, 453-460.
Fagotto, F. and Gumbiner, B. M. (1994). β-catenin localization duringXenopus embryogenesis: accumulation at tissue and somite boundaries.Development. 120, 3667-3679.
Fan, M. J., Gruning, W., Walz, G. and Sokol, S. Y. (1998). Wnt signaling
W. T. Montross, H. Ji and P. D. McCrea

1769β-Engrailed selectively suppresses Wnt signaling
and transcriptional control of Siamois in Xenopus embryos. Proc Natl. AcadSci USA 95, 5626-5631.
Fan, M. J. and Sokol, S. Y. (1997). A role for Siamois in Spemann organizerformation. Development 124, 2581-2589.
Funayama, N., Fagotto, F., McCrea, P. D. and Gumbiner, B. M. (1995).Embryonic axis induction by the Armadillo repeat domain of β-catenin:evidence for intracellular signaling. J. Cell Biol. 128, 959-968.
Gradl, D., Kuhl, M. and Wedlich, D. (1999). Keeping a close eye on Wnt-1/wg signaling in Xenopus. Mech. Dev. 86, 3-15.
Guger, K. A. and Gumbiner, B. M. (1995). β-catenin has Wnt-like activityand mimics the Nieuwkoop signaling center in Xenopus dorsal-ventralpatterning. Devl. Biol. 172, 115-125.
Gumbiner, B. M. (1995). Signal transduction of β-catenin. Curr. Opin. CellBiol. 7, 634-640.
Harland, R. M. (1991). In Situ Hybridization: An Improved Whole MountMethod for Xenopus Embryos. San Diego: Academic Press.
Hatzfeld, M. (1999). The Armadillo family of structural proteins. Int. Rev.Cytol. 186, 179-224.
Heasman, J., Crawford, A., Goldstone, K., Garner-Hamrick, P.,Gumbiner, B., McCrea, P., Kintner, C., Yoshida Noro, C. and Wylie, C.(1994). Overexpression of cadherins and underexpression of β-catenininhibit dorsal mesoderm induction in early Xenopus embryos. Cell 79, 791-803.
Herrenknecht, K., Ozawa, M., Eckerskorn, C., Lottspeich, F., Lentner, M.and Kemler, R. (1991). The uvomorulin-anchorage protein α-catenin is avinculin homologue. Proc. Natl. Acad. Sci. USA 88, 9156-9160.
Hirano, S., Kimoto, N., Shimoyama, Y., Hirohashi, S. and Takeichi, M.(1992). Identification of a neural α-catenin as a key regulator of cadherinfunction and multicellular organization. Cell 70, 293-301.
Huber, A. H., Nelson, W. J. and Weis, W. I. (1997). Three-dimensionalstructure of the Armadillo repeat region of β-catenin. Cell 90, 871-882.
Huber, O., Korn, R., McLaughlin, J., Ohsugi, M., Herrmann, B. G. andKemler, R. (1996). Nuclear localization of β-catenin by interaction withtranscription factor LEF-1. Mech. Dev. 59, 3-10.
Ikeda, S., Kishida, S., Yamamoto, H., Murai, H., Koyama, S. and Kikuchi,A. (1998). Axin, a negative regulator of the Wnt signaling pathway, formsa complex with GSK-3β and β-catenin and promotes GSK-3β – dependentphosphorylation of β-catenin. EMBO J. 17, 1371-1384.
Itoh, K., Krupnik, V. E. and Sokol, S. Y. (1998). Axis determination inXenopus involves biochemical interactions of Axin, Glycogen SynthaseKinase-3 and β-catenin. Curr. Biol. 8, 591-594.
Jaynes, J. B. and O’Farrell, P. H. (1991). Active repression of transcriptionby the Engrailed homeodomain protein. EMBO J. 10, 1427-1433.
Kao, K. R. and Elinson, R. P. (1988). The entire mesodermal mantle behavesas Spemann’s organizer in dorsoanterior enhanced Xenopus laevis embryos.Dev. Biol. 127, 64-77.
Kelly, G. M., Eib, D. W. and Moon, R. T. (1991). Histological preparationof Xenopus laevis oocytes and embryos. In Xenopus laevis: Practical Usesin Cell and Molecular Biology (ed. B. K. Kay and H. B. Peng), pp. 389-417. Academic Press, San Diego.
Kemler, R., Ozawa, M. and Ringwald, M. (1989). Calcium-dependent celladhesion molecules. Curr. Opin. Cell Biol. 1, 892-897.
Kessler, D. S. (1997). Siamois is required for formation of Spemann’sorganizer. Proc. Natl. Acad. Sci. USA 94, 13017-13022.
Kikuchi, A. (1999). Roles of Axin in the Wnt signalling pathway. Cell Signal.11, 777-788.
Kintner, C. (1992). Regulation of embryonic cell adhesion by the cadherincytoplasmic domain. Cell 69, 225-236.
Kishida, S., Yamamoto, H., Ikeda, S., Kishida, M., Sakamoto, I., Koyama,S. and Kikuchi, A. (1998). Axin, a negative regulator of the Wnt signalingpathway, directly interacts with adenomatous polyposis coli and regulatesthe stabilization of β-catenin. J. Biol. Chem. 273, 10823-10826.
Knudsen, K. A., Soler, A. P., Johnson, K. R. and Wheelock, M. J. (1995).Interaction of α-actinin with the cadherin/catenin cell-cell adhesion complexvia α-catenin. J. Cell Biol. 130, 67-77.
Larabell, C. A., Torres, M., Rowning, B. A., Yost, C., Miller, J. R., Wu,M., Kimelman, D. and Moon, R. T. (1997). Establishment of the dorso-ventral axis in Xenopus embryos is presaged by early asymmetries in β-catenin that are modulated by the Wnt signaling pathway. J. Cell Biol. 136,1123-1136.
Laurent, M. N., Blitz, I. L., Hashimoto, C., Rothbacher, U. and Cho, K.W. (1997). The Xenopus homeobox gene twin mediates Wnt induction ofgoosecoid in establishment of Spemann’s organizer. Development 124,4905-4916.
Lee, C. H. and Gumbiner, B. M. (1995). Disruption of gastrulationmovements in Xenopus by a dominant-negative mutant for C-cadherin. Dev.Biol. 171, 363-373.
Lemaire, P. and Kodjabachian, L. (1996). The vertebrate organizer: structureand molecules. Trends Genet. 12, 525-531.
Levine, E., Lee, C. H., Kintner, C. and Gumbiner, B. M. (1994). Selectivedisruption of E-cadherin function in early Xenopus embryos by a dominantnegative mutant. Development 120, 901-909.
McCrea, P. D., Brieher, W. M. and Gumbiner, B. M. (1993). Induction ofa secondary body axis in Xenopus by antibodies to β-catenin. J. Cell Biol.123, 477-484.
McCrea, P. D., Turck, C. W. and Gumbiner, B. (1991). A homolog of theDrosophila protein Armadillo (Plakoglobin) associated with E-cadherin.Science 254, 1359-1361.
McKendry, R., Hsu, S. C., Harland, R. M. and Grosschedl, R. (1997). LEF-1/TCF proteins mediate Wnt-inducible transcription from the Xenopusnodal-related-3 promoter. Dev. Biol. 192, 420-431.
Miller, J. R., Hocking, A. M., Brown, J. D. and Moon, R. T. (1999).Mechanism and function of signal transduction by the Wnt/ β-catenin andWnt/Ca2+ pathways. Oncogene 18, 7860-7872.
Miller, J. R. and Moon, R. T. (1996). Signal transduction through β-cateninand specification of cell fate during embryogenesis. Genes Dev. 10, 2527-2539.
Molenaar, M., van de Wetering, M., Oosterwegel, M., Peterson-Maduro,J., Godsave, S., Korinek, V., Roose, J., Destree, O. and Clevers, H.(1996). XTcf-3 transcription factor mediates β-catenin-induced axisformation in Xenopus embryos. Cell 86, 391-399.
Moon, R. T., Brown, J. D. and Torres, M. (1997). WNTs modulate cell fateand behavior during vertebrate development. Trends Genet. 13, 157-162.
Moon, R. T. and Kimelman, D. (1998). From cortical rotation to organizergene expression: toward a molecular explanation of axis specification inXenopus. BioEssays 20, 536-545.
Munemitsu, S., Albert, I., Rubinfeld, B. and Polakis, P. (1996). Deletion ofan amino-terminal sequence of β-catenin in vivo promotes thehyperphosphorylation of the adenomatous polyposis coli tumor suppressorprotein. Mol. Cell Biol. 16, 4088-4094.
Nagafuchi, A. and Tsukita, S. (1994). The loss of the expression of α-catenin,the 102 kD cadherin associated protein, in central nervous tissues duringdevelopment. Dev. Growth Diff. 36, 59-71.
Niehrs, C., Steinbeisser, H. and De Robertis, E. M. (1994). Mesodermalpatterning by a gradient of the vertebrate homeobox gene goosecoid. Science263, 817-820.
Nieuwkoop, P. D. and Faber, J. (1967). Normal Table of Xenopus laevis(Daudin). Amsterdam: Elsevier North-Holland Biomedical Press.
Nusse, R. (1997). A versatile transcriptional effector of wingless signaling.Cell 89, 321-323.
Ozawa, M. (1998). Identification of the region of α-catenin that plays anessential role in cadherin-mediated cell adhesion. J. Biol. Chem. 273, 29524-29529.
Paulson, A. F., Fang, X., Ji, H., Reynolds, A. B. and McCrea, P. D. (1999).Misexpression of the catenin p120(ctn)1A perturbs Xenopus gastrulationbut does not elicit Wnt-directed axis specification. Dev. Biol. 207, 350-363.
Peifer, M. (1995). Cell adhesion and signal transduction: The Armadilloconnection. Trends Cell Biol. 5, 224-229.
Peifer, M. (1997). β-catenin as oncogene: the smoking gun. Science 275, 1752-1753.
Peifer, M., Berg, S. and Reynolds, A. B. (1994). A repeating amino acid motifshared by proteins with diverse cellular roles. Cell 76, 789-791.
Peifer, M., Rauskolb, C., Williams, M., Riggleman, B. and Wieschaus, E.(1991). The segment polarity gene armadillo interacts with the winglesssignaling pathway in both embryonic and adult pattern formation.Development 111, 1029-1043.
Pierce, S. B. and Kimelman, D. (1995). Regulation of Spemann organizerformation by the intracellular kinase Xgsk-3. Development 121, 755-765.
Polakis, P. (1999). The oncogenic activation of β-catenin. Curr. Opin. Genet.Dev. 9, 15-21.
Rubinfeld, B., Albert, I., Porfiri, E., Fiol, C., Munemitsu, S. and Polakis,P. (1996). Binding of GSK-3β to the APC-β-catenin complex and regulationof complex assembly. Science 272, 1023-1026.
Rupp, R. A. W., Snider, L. and Weintraub, H. (1994). Xenopus embryosregulate the nuclear localization of XMyoD. Genes Dev. 8.
Salomon, D., Sacco, P. A., Roy, S. G., Simcha, I., Johnson, K. R., Wheelock,M. J. and Ben-Ze’ev, A. (1997). Regulation of β-catenin levels and

1770
localization by overexpression of plakoglobin and inhibition of theubiquitin-proteasome system. J. Cell Biol. 139, 1325-1335.
Sasai, Y., Lu, B., Steinbeisser, H., Geissert, D., Gont, L. K. and DeRobertis, E. M. (1994). Xenopus chordin: a novel dorsalizing factoractivated by organizer-specific homeobox genes. Cell 79, 779-790.
Schneider, S., Steinbeisser, H., Warga, R. M. and Hausen, P. (1996). β-Catenin translocation into nuclei demarcates the dorsalizing centers in frogand fish embryos. Mech. Dev. 57, 191-198.
Shulman, J. M., Perrimon, N. and Axelrod, J. D. (1998). Frizzledsignaling and the developmental control of cell polarity. Trends Genet. 14,452-458.
Smith, J. C., Price, B. M., Green, J. B. A., Weigel, D. and Herrman, B. G.(1991). Expression of a Xenopus homolog of Brachyury (T) is an immediateearly response to mesoderm induction. Cell 67, 79-87.
Smith, S. T. and Jaynes, J. B. (1996). A conserved region of Engrailed, sharedamong all en-, gsc-, Nk1-, Nk2- and msh-class homeoproteins, mediatesactive transcriptional repression in vivo. Development 122, 3141-3150.
Smith, W. C., McKendry, R., Ribisi, S., Jr. and Harland, R. M. (1995). Anodal-related gene defines a physical and functional domain within theSpemann organizer. Cell 82, 37-46.
Sokol, S. Y. (1996). Analysis of Dishevelled signalling pathways duringXenopus development. Curr. Biol. 6, 1456-1467.
Sokol, S. Y. (1999). Wnt signaling and dorso-ventral axis specification invertebrates. Curr. Opin. Genet. Dev. 9, 405-410.
Steinbeisser, H., De Roberts, E. M., Ku, M., Kessler, D. S. and Melton, D.(1993). Xenopus axis formation: induction of goosecoid by injected Xwnt-8 and activin mRNAs. Development 118, 499-507.
Steinberg, M. S. and McNutt, P. M. (1999). Cadherins and their connections:adhesion junctions have broader functions. Curr. Opin. Cell Biol. 11, 554-560.
Torres, M. A., Yang-Snyder, J. A., Purcell, S. M., DeMarais, A. A.
and McGrew, L. L. (1995). Activities of the Wnt-1 class of secretedsignaling factors are antagonized by the Wnt-5A class and by a dominantnegative cadherin in early Xenopus development. J. Cell Biol. 133, 1123-1137.
van de Wetering, M., Cavallo, R., Dooijes, D., van Beest, M., van Es, J.,Loureiro, J., Ypma, A., Hursh, D., Jones, T., Bejsovec, A. (1997).Armadillo coactivates transcription driven by the product of the Drosophilasegment polarity gene dTCF. Cell 88, 789-799.
Vleminckx, K., Kemler, R. and Hecht, A. (1999). The C-terminaltransactivation domain of β-catenin is necessary and sufficient for signalingby the LEF-1/ β-catenin complex in Xenopus laevis. Mech. Dev. 81, 65-74.
Wadzinski, B. E., Eisfelder, B. J., Peruski, L. F., Jr., Mumby, M. C. andJohnson, G. L. (1992). NH2-terminal modification of the phosphatase 2Acatalytic subunit allows functional expression in mammalian cells. J. Biol.Chem. 267, 16883-16888.
Wylie, C., Kofron, M., Payne, C., Anderson, R., Hosobuchi, M., Joseph,E. and Heasman, J. (1996). Maternal β-catenin establishes a ‘dorsal signal’in early Xenopus embryos. Development 122, 2987-2996.
Yap, A. S., Brieher, W. M., Pruschy, M. and Gumbiner, B. M. (1997).Lateral clustering of the adhesive ectodomain: a fundamental determinantof cadherin function. Curr. Biol. 7, 308-315.
Yost, C., Torres, M., Miller, J. R., Huang, E., Kimelman, D. and Moon, R.(1996). The axis-inducing activity, stability, and subcellular distribution ofβ-catenin is regulated in Xenopus embryos by Glycogen Synthase Kinase-3. Genes Dev. 10, 1443-1454.
Zhu, A. J. and Watt, F. M. (1999). β-Catenin signalling modulatesproliferative potential of human epidermal keratinocytes independently ofintercellular adhesion. Development 126, 2285-2298.
Zoltewicz, J. S. and Gerhart, J. C. (1997). The Spemann organizer ofXenopus is patterned along its anteroposterior axis at the earliest gastrulastage. Dev. Biol. 192, 482-491.
W. T. Montross, H. Ji and P. D. McCrea





























