Embed Size (px)
Citation preview

COMMUNICATIONS ARISING
Astrophysics
Supernova-remnantorigin of cosmic rays?
It is thought that Galactic cosmic-raynuclei are gradually accelerated to highenergies (up to about 300 TeV per nucleon,
where 1 TeV is 1012 eV) in the expandingshock waves connected with the remnantsof powerful supernova explosions. Howev-er, this conjecture has eluded direct obser-vational confirmation1,2 since it was firstproposed in 1953 (ref. 3). Enomoto et al.4
claim to have finally found definitive evi-dence that corroborates this model, propos-ing that very-high-energy, TeV-range,g-rays from the supernova remnant (SNR)RX J1713.713946 are due to the inter-actions of energetic nuclei in this region.Here we argue that their claim is not supported by the existing multiwavelengthspectrum of this source. The search for theorigin(s) of Galactic cosmic-ray nuclei maybe closing in on the long-suspected super-nova-remnant sources, but it is not yet over.
We have previously suggested5 that theSNR RX J1713.713946 (dark X-ray con-tours in Fig. 1) might be an accelerator ofcosmic-ray nuclei on the basis of GeV (109
eV) g-ray emission6 (Fig. 1, white contours)seen towards adjacent massive and densemolecular clouds (Fig. 1, false colour), withwhich it seems to be interacting7; the g-ray signature of nucleonic interactions isknown to be greatly amplified in such densemedia. Enomoto et al. have since claimed4
that the g-rays of even higher energy (TeVrange) that they detected directly towards
the more rarefied remnant (red contours inFig. 1) are an unambiguous signature ofnucleonic interactions in this region, thusfinally proving the long-held conjecture.
However, it is well known that SNRsaccelerate electrons (see ref. 8, for example)that can also generate TeV g-rays at thesource sites. It is therefore generally difficultto assign conclusively an electronic ratherthan a nucleonic origin to the observed celestial g-rays9. Fortunately, in this case thediscrimination is not so subtle — if the TeVemission seen in the northwest quadrant ofthe remnant were really due to protons, asproposed4, that part of the SNR would havebeen detected as a bright GeV source in theEnergetic Gamma Ray Experiment Telescope(EGRET) all-sky survey6, but it was not.
The proposed model4 is unsatisfactorybecause it predicts that there will be roughlythree times the GeV g-ray intensity of anearby EGRET source6 (3EG 171413857)at a location where no actual GeV flux hasbeen detected (that is, above the back-ground level), although alternative modelsmight reduce the discrepancy (see also ref.10). In addition, the high gas density (about102 protons cm13) needed to explain theTeV g-rays as nucleonic in origin4 is simplynot compatible –– by several orders of magnitude –– with the X-ray data7 from thesame region of the SNR.
As the peaks of both the TeV (ref. 4) andX-ray (ref. 7) emissions lie in the same low-density7 region of the remnant (Fig. 1), themost plausible explanation is that they botharise from the same population of relativisticelectrons. However, there is an exciting possibility that a minor component of theTeV emission4 could be due to nucleonic
brief communications
NATURE | VOL 418 | 1 AUGUST 2002 | www.nature.com/nature 499
interactions at the location of the adjacentcloud B (Fig. 1), where sufficient molecularmaterial is present5. A suggestive extensionand a local maximum in the TeV significancecontours4 are coincident with this location.
SNR RX J1713.713946 might thereforebe accelerating nuclei to cosmic-ray ener-gies5, but the TeV g-ray signature so fardetected from the northwest quadrant ofthe remnant4 is not proof of this. It is likelythat most — but not all — of the observedTeV emission is actually electronic in origin.
To address the question of the source(s)of the TeV emission properly, better radiodata sampled at several frequencies, as wellas the inclusion of existing low-energyROSAT X-ray data, will be needed to deter-mine the primary synchrotron spectrum.Data of greater spatial resolution from thenext-generation GeV, TeV and neutrinotelescopes will also be important. Onlywhen this multifrequency data set is avail-able can the nature of the contributions(nucleonic or electronic) to the observedTeV emissions start to be disentangled.Yousaf M. Butt*, Diego F. Torres†,Gustavo E. Romero‡, Thomas M. Dame*,Jorge A. Combi‡*Harvard-Smithsonian Center for Astrophysics,Cambridge, Massachusetts 02138, USAe-mail: [email protected]†Department of Physics, Jadwin Hall, PrincetonUniversity, Princeton, New Jersey 08544, USA‡Instituto Argentino de Radioastronomía, CC 5,1894, Villa Elisa, Buenos Aires, Argentina
1. Drury, L. O’C. et al. Space Sci. Rev. 99, 329–352 (2001).
2. Plaga, R. Preprint astro-ph/0111555 at
<http://xxx.lanl.gov> (2001).
3. Shklovskii, I. S. Dokl. Akad. Nauk SSSR 91, 475–478 (1953).
4. Enomoto, R. et al. (CANGAROO collaboration) Nature 416,
823–826 (2002).
5. Butt, Y. M., Torres, D. F., Combi, J. A., Dame, T. &
Romero, G. E. Astrophys. J. 562, L167–L171 (2001).
6. Hartman, R. C. et al. Astrophys. J. (suppl.) 123, 79–202 (1999).
7. Slane, P. et al. Astrophys. J. 525, 357–367 (1999).
8. Koyama, K. et al. Nature 378, 255–258 (1995).
9. Pohl, M. Astron. Astrophys. 307, L57–L59 (1996).
10.Reimer, O. & Pohl, M. Preprint astro-ph/0205256 at
<http://xxx.lanl.gov> (2002); Astron. Astrophys. (in the press).
COMMUNICATIONS ARISING
Behavioural evolution
Does similarity breedcooperation?
Reciprocity1, whether direct2 or indirect3,is thought to be the key to establishingcooperation among non-relatives. But
Riolo et al.4 have presented a model in whichcooperation is instead based on similarity:agents donate only when their partner’s ‘tag’lies within a ‘tolerance’ range around theirown. Here we point out that their modelrequires individuals with identical tags tocooperate with each other, and show thatcooperation tends to collapse when individ-uals bearing identical tags are given the
Cloud A
Cloud B
349.5° 349.0° 348.5° 348.0° 347.5° 347.0° 346.5°
–0.5°
0°
–0.5°
–1.0°
60
50
40
30
20
10
Galactic longitude
Gal
actic
latit
ude
103
WC
O (K
m s
–1)
Figure 1 An overlay map in Galactic coordinates showing supernova remnant (SNR) RX J1713.713946 (G347.310.5) in dark contours
(ROSAT PSPC X-ray; from ref. 7). Red, TeV significance contours4; white, location-probability contours of the GeV (109 eV) EGRET source
3EG J171413857 (ref. 6); colour scale indicates the intensity of CO (J41→0) emission, and consequently the column density of the
ambient molecular clouds, in the velocity interval v41105 to 180 km s11 associated with the SNR, corresponding to a kinematic
distance of 6.350.4 kpc (ref. 5). Note the local maximum of the TeV significance contours at cloud B, which may indicate a hadronic
origin of the TeV flux at that location. For further details, see ref. 5.
© 2002 Nature Publishing Group
option of not donating. We therefore questiontheir mechanism for maintaining coopera-tion without reciprocity.
What makes cooperation so challengingfor theorists is explaining how it can persistin the face of more exploitative strategies. Inthe system of Riolo et al., clusters of coopera-ting agents with similar tags arise inter-mittently, only to be undermined by agentsthat reduce their tolerance level, T, such that they accept more donations than they offer. However, there is a limit to such cheatingimposed by the minimum T40. This meansthat when individuals with identical tagsinteract, they must always donate. A strikingcharacteristic of Riolo and colleagues’ simu-lations was the formation, through differen-tial reproduction, of clusters of agents withidentical tags. Most individuals in their sim-ulations (up to 97% of the population4) weretherefore ultimately constrained to cooperate.
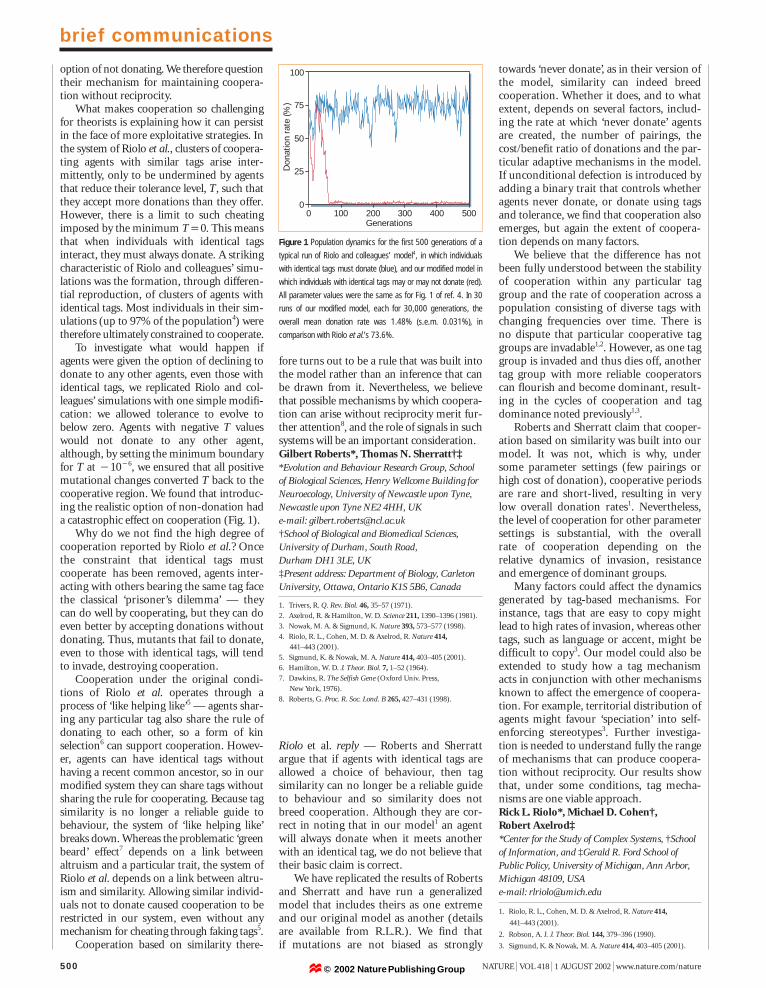
To investigate what would happen ifagents were given the option of declining todonate to any other agents, even those withidentical tags, we replicated Riolo and col-leagues’ simulations with one simple modifi-cation: we allowed tolerance to evolve tobelow zero. Agents with negative T valueswould not donate to any other agent,although, by setting the minimum boundaryfor T at 11016, we ensured that all positivemutational changes converted T back to thecooperative region. We found that introduc-ing the realistic option of non-donation hada catastrophic effect on cooperation (Fig. 1).
Why do we not find the high degree ofcooperation reported by Riolo et al.? Oncethe constraint that identical tags mustcooperate has been removed, agents inter-acting with others bearing the same tag facethe classical ‘prisoner’s dilemma’ — theycan do well by cooperating, but they can doeven better by accepting donations withoutdonating. Thus, mutants that fail to donate,even to those with identical tags, will tendto invade, destroying cooperation.
Cooperation under the original condi-tions of Riolo et al. operates through aprocess of ‘like helping like’5 — agents shar-ing any particular tag also share the rule ofdonating to each other, so a form of kin selection6 can support cooperation. Howev-er, agents can have identical tags withouthaving a recent common ancestor, so in ourmodified system they can share tags withoutsharing the rule for cooperating. Because tagsimilarity is no longer a reliable guide tobehaviour, the system of ‘like helping like’breaks down. Whereas the problematic ‘greenbeard’ effect7 depends on a link betweenaltruism and a particular trait, the system ofRiolo et al. depends on a link between altru-ism and similarity. Allowing similar individ-uals not to donate caused cooperation to berestricted in our system, even without anymechanism for cheating through faking tags5.
Cooperation based on similarity there-
fore turns out to be a rule that was built intothe model rather than an inference that canbe drawn from it. Nevertheless, we believethat possible mechanisms by which coopera-tion can arise without reciprocity merit fur-ther attention8, and the role of signals in suchsystems will be an important consideration.Gilbert Roberts*, Thomas N. Sherratt†‡*Evolution and Behaviour Research Group, Schoolof Biological Sciences, Henry Wellcome Building forNeuroecology, University of Newcastle upon Tyne,Newcastle upon Tyne NE2 4HH, UKe-mail: [email protected]†School of Biological and Biomedical Sciences,University of Durham, South Road, Durham DH1 3LE, UK‡Present address: Department of Biology, CarletonUniversity, Ottawa, Ontario K1S 5B6, Canada
1. Trivers, R. Q. Rev. Biol. 46, 35–57 (1971).
2. Axelrod, R. & Hamilton, W. D. Science 211, 1390–1396 (1981).
3. Nowak, M. A. & Sigmund, K. Nature 393, 573–577 (1998).
4. Riolo, R. L., Cohen, M. D. & Axelrod, R. Nature 414,
441–443 (2001).
5. Sigmund, K. & Nowak, M. A. Nature 414, 403–405 (2001).
6. Hamilton, W. D. J. Theor. Biol. 7, 1–52 (1964).
7. Dawkins, R. The Selfish Gene (Oxford Univ. Press,
New York, 1976).
8. Roberts, G. Proc. R. Soc. Lond. B 265, 427–431 (1998).
Riolo et al. reply — Roberts and Sherrattargue that if agents with identical tags areallowed a choice of behaviour, then tag similarity can no longer be a reliable guideto behaviour and so similarity does notbreed cooperation. Although they are cor-rect in noting that in our model1 an agentwill always donate when it meets anotherwith an identical tag, we do not believe thattheir basic claim is correct.
We have replicated the results of Robertsand Sherratt and have run a generalizedmodel that includes theirs as one extremeand our original model as another (detailsare available from R.L.R.). We find that if mutations are not biased as strongly
towards ‘never donate’, as in their version ofthe model, similarity can indeed breedcooperation. Whether it does, and to whatextent, depends on several factors, includ-ing the rate at which ‘never donate’ agentsare created, the number of pairings, thecost/benefit ratio of donations and the par-ticular adaptive mechanisms in the model.If unconditional defection is introduced byadding a binary trait that controls whetheragents never donate, or donate using tagsand tolerance, we find that cooperation alsoemerges, but again the extent of coopera-tion depends on many factors.
We believe that the difference has notbeen fully understood between the stabilityof cooperation within any particular taggroup and the rate of cooperation across apopulation consisting of diverse tags withchanging frequencies over time. There is no dispute that particular cooperative taggroups are invadable1,2. However, as one taggroup is invaded and thus dies off, anothertag group with more reliable cooperatorscan flourish and become dominant, result-ing in the cycles of cooperation and tagdominance noted previously1,3.
Roberts and Sherratt claim that cooper-ation based on similarity was built into ourmodel. It was not, which is why, undersome parameter settings (few pairings orhigh cost of donation), cooperative periodsare rare and short-lived, resulting in verylow overall donation rates1. Nevertheless,the level of cooperation for other parametersettings is substantial, with the overall rate of cooperation depending on the relative dynamics of invasion, resistanceand emergence of dominant groups.
Many factors could affect the dynamicsgenerated by tag-based mechanisms. Forinstance, tags that are easy to copy mightlead to high rates of invasion, whereas othertags, such as language or accent, might bedifficult to copy3. Our model could also beextended to study how a tag mechanismacts in conjunction with other mechanismsknown to affect the emergence of coopera-tion. For example, territorial distribution ofagents might favour ‘speciation’ into self-enforcing stereotypes3. Further investiga-tion is needed to understand fully the rangeof mechanisms that can produce coopera-tion without reciprocity. Our results showthat, under some conditions, tag mecha-nisms are one viable approach.Rick L. Riolo*, Michael D. Cohen†,Robert Axelrod‡*Center for the Study of Complex Systems, †Schoolof Information, and ‡Gerald R. Ford School ofPublic Policy, University of Michigan, Ann Arbor,Michigan 48109, USAe-mail: [email protected]
1. Riolo, R. L., Cohen, M. D. & Axelrod, R. Nature 414,
441–443 (2001).
2. Robson, A. J. J. Theor. Biol. 144, 379–396 (1990).
3. Sigmund, K. & Nowak, M. A. Nature 414, 403–405 (2001).
brief communications
500 NATURE | VOL 418 | 1 AUGUST 2002 | www.nature.com/nature
Figure 1 Population dynamics for the first 500 generations of a
typical run of Riolo and colleagues’ model4, in which individuals
with identical tags must donate (blue), and our modified model in
which individuals with identical tags may or may not donate (red).
All parameter values were the same as for Fig. 1 of ref. 4. In 30
runs of our modified model, each for 30,000 generations, the
overall mean donation rate was 1.48% (s.e.m. 0.031%), in
comparison with Riolo et al.’s 73.6%.
0
25
50
75
100
0 100 200 300 400 500Generations
Don
atio
n ra
te (%
)
© 2002 Nature Publishing Group





























