Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED. Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED. BCH 5045 Graduate Survey of Biochemistry Instructor: Charles Guy Producer: Ron Thomas Director: Marsha Durosier Lecture 34 Slide sets available at: http://hort.ifas.ufl.edu/teach/guyweb/bch5045/index.html
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
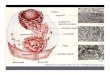
FIGURE 27-39 Synthesis of the core oligosaccharide of glycoproteins. The core oligosaccharide is built up by the successive addition of monosaccharide units. 1,2 The first steps occur on the cytosolic face of the ER. 3 Translocation moves the incomplete oligosaccharide across the membrane (mechanism not shown), and 4 completion of the core oligosaccharide occurs within the lumen of the ER. The precursors that contribute additional mannose and glucose residues to the growing oligosaccharide in the lumen are dolichol phosphate derivatives. In the first step in the construction of the N-linked oligosaccharide moiety of a glycoprotein, 5,6 the core oligosaccharide is transferred from dolichol phosphate to an Asn residue of the protein within the ER lumen. The core oligosaccharide is then further modified in the ER and the Golgi complex in pathways that differ for different proteins. The five sugar residues shown surrounded by a beige screen (after step 7) are retained in the final structure of all N-linked oligosaccharides. 8 The released dolichol pyrophosphate is again translocated so that the pyrophosphate is on the cytosolic face of the ER, then 9 a phosphate is hydrolytically removed to regenerate dolichol phosphate
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Once in the golgi complex sorting must take place to target (send) the protein to its proper destination. Where might the information needed to insure the protein gets to where it should? What are our options?
Presenter
Presentation Notes
FIGURE 27-40 Pathway taken by proteins destined for lysosomes, the plasma membrane, or secretion. Proteins are moved from the ER to the cis side of the Golgi complex in transport vesicles. Sorting occurs primarily in the trans side of the Golgi complex. FIGURE 27-41 Phosphorylation of mannose residues on lysosome-targeted enzymes. N-Acetylglucosamine phosphotransferase recognizes some as yet unidentified structural feature of hydrolases destined for lysosomes.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Proteins must also enter the nucleus. Here the mechanism is a little different than seen for proteins destine for the ER-golgi pathway. Nuclei have large pores, but they are selective. A monopartite NLS consensus sequence has a basic motif of K-K/R-X-K/R amino acids.
Presenter
Presentation Notes
FIGURE 27-42b Targeting of nuclear proteins. (b) Scanning electron micrograph of the surface of the nuclear envelope, showing numerous nuclear pores. FIGURE 27-42a Targeting of nuclear proteins. (a) 1 A protein with an appropriate nuclear localization signal (NLS) is bound by a complex of importin α and β. In addition to the monopartite NLS, there is a bipartite motif of KR[PAATKKAGQA]KKKK which seems to possess two clusters of basic amino acids separated by a spacer of about 10 amino acids. 2 The resulting complex binds to a nuclear pore, and translocates. 3 Inside the nucleus, dissociation of importin β is promoted by the binding of Ran-GTP. 4 Importin α binds to Ran-GTP and CAS (cellular apoptosis susceptibility protein), releasing the nuclear protein. 5 Importin α and β and CAS are transported out of the nucleus and recycled. They are released in the cytosol when Ran hydrolyzes its bound GTP. 6 Ran-GDP is bound by NTF2, and transported back into the nucleus. 7 RanGEF promotes the exchange of GDP for GTP in the nucleus, and Ran-GTP is ready to process another NLS-bearing protein–importin complex.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
While bacteria do not have organelles, they still possess a plasma membrane and export proteins to the outside. They have an export system that is a little bit like what is seen during import of a protein into the ER.
Presenter
Presentation Notes
FIGURE 27-43 Signal sequences that target proteins to different locations in bacteria. Basic amino acids (blue) near the amino terminus and hydrophobic core amino acids (yellow) are highlighted. The cleavage sites marking the ends of the signal sequences are indicated by red arrows. Note that the inner bacterial cell membrane (see Figure 1-6) is where phage fd coat proteins and DNA are assembled into phage particles. OmpA is outer membrane protein A; LamB is a cell surface receptor protein for λ phage.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Protein export in bacteria
Outside
Presenter
Presentation Notes
FIGURE 27-44 Model for protein export in bacteria. 1 A newly translated polypeptide binds to the cytosolic chaperone protein SecB, which 2 delivers it to SecA, a protein associated with the translocation complex (SecYEG) in the bacterial cell membrane. 3 SecB is released, and SecA inserts itself into the membrane, forcing about 20 amino acid residues of the protein to be exported through the translocation complex. 4 Hydrolysis of an ATP by SecA provides the energy for a conformational change that causes SecA to withdraw from the membrane, releasing the polypeptide. 5 SecA binds another ATP, and the next stretch of 20 amino acid residues is pushed across the membrane through the translocation complex. Steps 4 and 5 are repeated until 6 the entire protein has passed through and is released to the periplasm. The electrochemical potential across the membrane (denoted by + and –) also provides some of the driving force required for protein translocation.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Protein import pathways into mitochondria.
Wiedemann N et al. J. Biol. Chem. 2004;279:14473-14476
Protein import pathways into mitochondria. Most mitochondrial proteins are synthesized in the cytosol. With the help of cytosolic chaperones, mitochondrial precursor proteins are transferred to the general entry gate of mitochondria, the TOM complex, from where they are subsequently sorted into one of the mitochondrial sub-compartments. The precursors of β-barrel outer membrane proteins require the SAM complex. Preproteins destined for the matrix depend on the presequence translocase (TIM23 complex) and its associated import motor (PAM complex) for their transport across the inner mitochondrial membrane. Carrier proteins are inserted into the inner membrane with the help of the carrier translocase (TIM22 complex). OM, outer membrane; IMS, intermembrane space; IM, inner membrane.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Protein translocation across and into the outer mitochondrial membrane.
Wiedemann N et al. J. Biol. Chem. 2004;279:14473-14476
Import of presequence proteins into the mitochondrial matrix.
Presenter
Presentation Notes
Protein translocation across and into the outer mitochondrial membrane. Mitochondrial precursor proteins possess either an N-terminal presequence (positively charged amphipathic α-helix) or contain internal targeting signals. The presequence is recognized by the import receptors Tom20 and Tom22, after which the preprotein is transferred with the help of Tom5 to the general import pore formed by Tom40. The TOM complex contains two Tom40 pores (not shown in the figure). After presequence proteins emerge from the Tom40 channel they bind to the intermembrane space domain of Tom22 prior to their transfer to the presequence translocase of the inner membrane. Carrier proteins with internal targeting signals are initially recognized by the receptor Tom70. With the help of the other receptors, Tom20, Tom22, and Tom5, internal loops of the carrier proteins are translocated through the Tom40 channel and bind to the Tim9-Tim10 complex of the intermembrane space (IMS). The import of outer membrane proteins initially depends on the receptors Tom20 and Tom22. After transport through the TOM machinery, the SAM complex is required for the insertion of β-barrel proteins into the outer membrane (OM). Import of presequence proteins into the mitochondrial matrix. After passing through the channel of the TOM complex, presequence-carrying preproteins are recognized by an intermembrane space (IMS) domain of Tom22. From here, they are brought into contact with Tim50, which then assists in guiding them to the presequence translocase (TIM23 complex). Their subsequent insertion into the channel, formed by Tim23, requires the membrane potential (Δψ) across the inner membrane (IM). A further driving force is provided by the PAM in the matrix. The central component of this motor, mtHsp70, is transiently anchored at the translocase by Tim44 and requires assistance from Pam18, Pam16, and Mge1 for promotion of the reaction cycle. The presequences of the precursor proteins are cleaved off by the MPP in the matrix. OM, outer membrane.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Protein-import pathways into chloroplasts. Preproteins with an amino-terminal transit sequence form a cytosolic guidance complex. The complex consists of an HSP70 (heat shock protein-70) chaperone and a 14-3-3 dimer. Preproteins without a cleavable transit sequence are targeted and insert into the outer envelope. IE, inner envelope; IES, inter-envelope space; OE, outer envelope;
OEP, outer-envelope protein; SEC, secretory pathway; SRP; signal-recognition particle; TAT, twin-arginine translocase; TIC, translocon of the inner chloroplast envelope.
Soll and Schleiff, Nature Reviews | Molecular Cell Biology (2004) 5: 198-208
Presenter
Presentation Notes
In the cytosol, preproteins with an amino-terminal presequence or transit sequence (yellow) can form a cytosolic guidance complex on phosphorylation in the cytosol. This guidance complex consists of an HSP70 (heat shock protein-70) chaperone and a 14-3-3 dimer. Altenatively, non-phosphorylated preproteins can associate with TOC159 (translocon of the outer chloroplast envelope-159) or with HSP70. All these complexes bind to TOC receptors in a GTP-dependent manner. A soluble TOC159 receptor might shuttle between the cytosol and the TOC complex bringing preproteins to the organelle surface. In a joint effort between TOC and TIC, preproteins are imported across both membranes in an NTP-dependent manner. The transit sequences are cleaved off by the stromal processing peptidase (SPP). Mature proteins either fold and assemble in the stroma or are directed to the thylakoids by various pathways. Preproteins without a cleavable transit sequence are mostly bound by HSP70 and targeted to the outer envelope. They can insert spontaneously into the membrane in vitro, although the process might be facilitated by proteinaceous factors in vivo, which still need to be identified (highlighted by a question mark). ALB3, albino3; IE, inner envelope; IES, inter-envelope space; OE, outer envelope; OEP, outer-envelope protein; SEC, secretory pathway; SRP; signal-recognition particle; TAT, twin-arginine translocase; TIC, translocon of the inner chloroplast envelope.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
Endocytosis pathways in eukaryotic cells
Presenter
Presentation Notes
FIGURE 27-45 Summary of endocytosis pathways in eukaryotic cells. Pathways dependent on clathrin or caveolin make use of the GTPase dynamin to pinch vesicles from the plasma membrane. Some pathways do not use clathrin or caveolin; some of these make use of dynamin and some do not.
Images from the Text are protected by Copyright (c) 2008 by W. H. Freeman and Company, and by the licensors of W. H. Freeman and Company. Living Graphs software (c) 2008 Sumanas, Inc. ALL RIGHTS RESERVED.
Commentary by the instructor is protected by Copyright (c) 2011. ALL RIGHTS RESERVED.
It is every bit as important for proteins to be degraded to their constituent amino acids as it is for proteins to be synthesized. Without protein turnover, regulation of enzymatic activity becomes rather limited. Ubiquitin is a small (76 aa) highly conserved protein used to tag other proteins for termination by the 26S proteosome.
PDB 2D5G
Presenter
Presentation Notes
FIGURE 27-47 Three-step pathway by which ubiquitin is attached to a protein. Two different enzyme-ubiquitin intermediates are involved. The free carboxyl group of ubiquitin's carboxyl-terminal Gly residue is ultimately linked through an amide (isopeptide) bond to an ε-amino group of a Lys residue of the target protein. Additional cycles produce polyubiquitin, a covalent polymer of ubiquitin subunits that targets the attached protein for destruction in eukaryotes. FIGURE 27-48 Three-dimensional structure of the eukaryotic proteasome. The 26S proteasome is highly conserved in all eukaryotes. The two subassemblies are the 20S core particle and the 19S regulatory particle. (a) (PDB ID 1IRU) The core particle consists of four rings arranged to form a barrel-like structure. Each of the inner rings has seven different β subunits (light blue), three of which have protease activities (dark blue). The outer rings each have seven different α subunits (gray). (b) A regulatory particle forms a cap on each end of the core particle. The core particle is colored as in (a). The base and lid segments of each regulatory particle are presented in different shades of pink. The regulatory particle unfolds ubiquitinated proteins (blue) and translocates them into the core particle, as shown.