Embed Size (px)
Citation preview

Developmental Biology 380 (2013) 145–156
Contents lists available at SciVerse ScienceDirect
Developmental Biology
0012-16http://d
n CorrE-m
journal homepage: www.elsevier.com/locate/developmentalbiology
Basigin null mutant male mice are sterile and exhibit impairedinteractions between germ cells and Sertoli cells
Jiajia Bi a, Yanfen Li a, Fengyun Sun b, Anja Saalbach c, Claudia Klein c, David J. Miller a,Rex Hess d, Romana A. Nowak a,n
a Department of Animal Sciences, University of Illinois, Urbana, IL, USAb The Jackson Laboratory, Bar Harbor, Maine, ME, USAc Department of Dermatology, Venereology and Allergology, Leipzig University, Leipzig, Germanyd Department of Comparative Biosciences, University of Illinois, Urbana, IL, USA
a r t i c l e i n f o
Article history:Received 27 September 2012Received in revised form10 May 2013Accepted 14 May 2013Available online 28 May 2013
Keywords:BasiginBSGN-glycansα-Mannosidase IISpermatogenesisBlood-testis barrier
06/$ - see front matter & 2013 Elsevier Inc. Ax.doi.org/10.1016/j.ydbio.2013.05.023
espondence to: 1207 W Gregory Dr. Urbana,ail address: [email protected] (R.A. Nowak
a b s t r a c t
Basigin (BSG) is a multifunctional glycoprotein that plays an important role in male reproduction sincemale knockout (KO) mice are sterile. The Bsg KO testis lacks elongated spermatids and maturespermatozoa, a phenotype similar to that of alpha-mannosidase IIx (MX) KO mice. MX regulatesformation of N-acetylglucosamine (GlcNAc) terminated N-glycans that participate in germ cell-Sertolicell adhesion. Results showed that Bsg KO spermatocytes displayed normal homologous chromosomesynapsis and progression through meiosis. However, only punctate expression of the round spermatidmarker SP-10 in the acrosomal granule of germ cells of Bsg KO mice was detected indicating thatspermatogenesis in Bsg KO mice was arrested at the early round spermatid stages. We observed a largeincrease in the number of germ cells undergoing apoptosis in Bsg KO testes. Using lectin blotting, wedetermined that GlcNAc terminated N-glycans are linked to BSG. GlcNAc terminated N-glycans weresignificantly reduced in Bsg KO testes. These observations indicate that BSG may act as a germ cell-Sertolicell attachment molecule. Loss of BSG significantly reduced adhesion between GC-2 and SF7 cells.Moreover, wild type testes showed strong expression of N-cadherin (CDH2) while expression was greatlyreduced in the testes of Bsg KO mice. In addition, the integrity of the blood-testis barrier (BTB) wascompromised in Bsg KO testes. In conclusion, although some Bsg KO spermatogonia can undergo normalprogression to the spermatocyte stage, BSG-mediated germ cell-Sertoli cell interactions appear to benecessary for integrity of the BTB and spermatocyte progression to mature spermatozoa.
& 2013 Elsevier Inc. All rights reserved.
Introduction
Spermatogenesis is the process of producing sperm (haploid)from initially undifferentiated spermatogonia (diploid). Duringspermatogenesis, there are a group of somatic cells named Sertolicells that have direct contact with all the different types of germcells in the seminiferous tubules. Sertoli cells play a major role inthe regulation of germ cell differentiation and migration. First,these cells form the blood-testis barrier (BTB) in the seminiferousepithelium near the basement membrane which protects thepostmeiotic germ cells from the host immune system in order toavoid attack from the production of anti-sperm antibodies againstgerm cell specific antigens (Dym and Fawcett, 1970; Morrow et al.,2009; Wong et al., 2010). The BTB has been shown to be composedof tight junctions (TJ), basal ectoplasmic specializations (basal ES,
ll rights reserved.
IL 61801. Fax: +217 333 8286.).
a testis-specific type of adherens junction), gap junctions (GJ)and desmosome-like junctions. Several adhesion moleculeshave been identified to play an important role in the functioningof these junctions including claudins and occludins (TJ); cox-sacckie and adenovirus receptor (CXADR), N-cadherin (CDH2)and β-catenin (basal ES); connexin43 (CX43) (GJ); desmoglein-2and desmocollin-2 (desmosome-like junction) (Cheng and Mruk,2009; Komljenovic et al., 2009; Pointis and Segretain, 2005; Wonget al., 2008). Knockouts generated against specific junction pro-teins such as claudin 11 (CLDN11) (Gow et al., 1999) can causesterility in the male mouse, which emphasizes the role thesejunction proteins play in the appropriate functioning of the BTBand also prove that an intact BTB is necessary for successful malefertility.
Sertoli cells are also responsible for the formation and secretionof seminiferous tubule fluid (STF) which consists of a number ofproducts that are involved in spermatogenesis such as transportand binding proteins, proteases and protease inhibitors, hormonesand growth factors (Russell and Griswold, 1993; Skinner and

J. Bi et al. / Developmental Biology 380 (2013) 145–156146
Griswold, 2005). STF is important for maintaining the nutritionaland hormonal microenvironment necessary for the viability anddifferentiation of germ cells during spermatogenesis. It alsoprovides the milieu for developing spermatozoa and the vehicleto transport them from the testis (Rato et al., 2010; Richburg et al.,1994). Furthermore, there is strong evidence that some ligandssecreted by Sertoli cells can modulate gene expression in germcells during spermatogenesis (Tsuruta and O'Brien, 1995). Sertolicells also communicate with germ cells through direct cell-cellcontact and paracrine interactions. The loss of these interactionsbetween Sertoli cells and germ cells leads to the failure ofspermatogenesis. For example, oligosaccharide 310.11 which is aN-acetylglucosamine (GlcNAc) terminated tri-antennary and fuco-sylated N-glycan structure has been shown to play a key role ingerm cell adhesion to Sertoli cells. A 50% reduction in expressionof this N-glycan structure results in almost complete suppressionof fertility (Akama et al., 2002). Although this specific oligosac-charide plays a key role in spermatogenesis, the glycoprotein thatthe oligosaccharide is attached to is still unknown.
Basigin (basic immunoglobulin superfamily; BSG) is a trans-membrane glycoprotein rich in N-glycans that belongs to theimmunoglobulin superfamily (Miyauchi et al., 1990; Tang et al.,2004). This molecule has been identified independently by differ-ent laboratories and is also known as EMMPRIN, CD147 and M6 inhuman, GP42 in mice, HT7 and neurothelin in chicken and OX-47,MC31 and CE9 in the rat (Nehme et al., 1995; Sameshima et al.,2000; Schlosshauer and Herzog, 1990; Seulberger et al., 1992;Wakayama et al., 2000). The molecular weight of BSG lies in arange between 43 kD and 66 kD due to variable glycosylationwhereas the molecular weight of the core protein is around 27 kD(Fossum et al., 1991; Miyauchi et al., 1990; Seulberger et al., 1992).BSG is highly expressed in reproductive tissues and plays a crucialrole in both male and female reproduction (Chen et al., 2010;Igakura et al., 1998; Kuno et al., 1998). In the mouse testis, BSG isexpressed in Sertoli cells, Leydig cells and strongly expressed inspermatocytes and spermatids (Chen et al., 2010; Igakura et al.,1998). Bsg gene KO males, created by targeted disruption, aresterile due to the failure of spermatogenesis (Igakura et al., 1998).However, the underlying mechanisms of Bsg action in mammalianspermatogenesis still remain unclear. The goals of the currentstudy of Bsg gene null mutant (-/-) testes were: (1) to determinethe effect of loss of BSG on the transition of spermatocytes tospermatids; (2) to investigate the importance of BSG in main-tenance of the BTB; and (3) to determine whether BSG is thepredominant protein that binds N-glycans in the testis.
Materials and methods
Animals
Animals used in this research were maintained in accordancewith the guidelines of the Institutional Animal Care and UseCommittee at the University of Illinois. The wild-type C57BL6/Jmale mice were obtained from Jackson Laboratory (Bar Harbor,ME) and were housed under temperature and light controlledconditions (12 h light: 12 h dark) with free access to food andwater. C57BL6/J heterozygous (Bsg +/−) mice were a kind gift fromDr. Takashi Muramatsu, Department of Biochemistry, NagoyaUniversity School of Medicine, Japan. Heterozygote breeding wascarried out in the animal facility of the University of Illinois. At3 weeks of age, male offspring were anesthetized by isofluraneinhalation (Attane; Minrad, Bethlehem, PA) and tail snips werecollected for genotyping. DNA was extracted from the snips usingREDExtract-N-Amp (Sigma, St. Louis, MO), and genotypes deter-mined by PCR using primers for Bsg and neomycin (Igakura et al.,
1998). Offspring with the Bsg −/− phenotype were maintainedunder the same conditions as those for the WT mice. Theincidence of Bsg null mutant offspring is very low due to the factthat most null mutant embryos fail to implant and are lost prior tobirth. Thus we obtained only 1% Bsg null mutant offspring and onlyhalf of these were males. The overall number of Bsg null mutantmale mice utilized for these studies was 15 over a three yearperiod.
Tissue collection
The wild type and Bsg KO mice (6 days, 12–24 weeks or 15–17months of age) were euthanized with carbon dioxide and testescollected immediately. Tissues for histology were fixed in Bouin'sor 4% paraformaldehyde in PBS overnight and processed forparaffin embedding. Tissues for Western and lectin blottinganalyses were snap-frozen in liquid nitrogen and stored at−80 1C until use.
Radioimmunoassay
Blood samples were collected from wild type and Bsg KO mice(n¼3) and serum samples were stored at −80 1C until the assayswere performed. Testosterone levels were measured using aradioimmunoassay performed by the University of Virginia CoreLigand and Assay Laboratory. The limit of detection for this assaywas 0.1 ng.
Cell culture
GC-2 cells (Hofmann et al., 1995) are an immortalized sperma-tocyte cell line and were purchased from the American TypeCulture Collection (ATCC) (CRL-2196). Cells were cultured inDulbecco's modified Eagle's medium (DMEM) supplemented with10% fetal bovine serum and 1% Penicillin–Streptomycin in a 5% CO2
humidified incubators at 37 1C.SF7 cells (Hofmann et al., 1992) are an immortalized Sertoli cell
line and were a gift from Dr. Hofmann at University of Illinois. Cellswere cultured in DMEM/F12 supplemented with 10% fetal bovineserum, 1% L-glutamine and 1% Penicillin–Streptomycin in a 5% CO2
humidified incubators at 37 1C.
Germ cell isolation and spermatocyte enrichment
Testes were removed from both wild type and Bsg KO mice,detunicated and digested in 0.5 mg/ml collagenase (Sigma) in Krebs-Ringer bicarbonate solution (KRB) [120.1 mM NaCl, 4.8 mM KCl,25.2 mM NaHCO3, 1.2 mM KH2PO4, 1.2 mM MgSO4.7H2O, 1.3 mMCaCl2, 11 mM glucose, 1� essential amino acids, 1� nonessentialamino acids] at 32 1C for 20 min, followed by digestion in 0.5 mg/mltrypsin (Sigma) containing 20 mg/ml DNase I in KRB at 32 1C for13 min. After digestion the cell suspension was filtered through an80-mm mesh filter and washed three times in KRB. Germ cells werethen processed as described below.
Cytological analyses
For immunofluorescence analysis, germ cells from testes ofthree wild type and three Bsg KO mice were collected bycentrifugation, surface-spread in wells of multispot microscopeslides (Shandon, Pittsburgh, PA) and fixed following the procedurepreviously described (Cobb et al., 1999b, 1997). Prior to antibodylabeling, slides were washed three times in washing/blockingbuffer (0.3% BSA, 1% goat serum in phosphate-buffered saline, pH7.4); the second wash included 0.05% Triton-X 100. After draining,the slides were incubated with primary antibodies. Antibodies and

J. Bi et al. / Developmental Biology 380 (2013) 145–156 147
dilution used were: rabbit anti-SYCP1 and SYCP3 (Novus, Littleton,CO) used at 1:100 dilution; guinea pig anti-H1T (Cobb et al., 1999a)at 1:500; rabbit anti-γH2AX (Millipore) at 1:200; Secondaryantibodies against rabbit, rat or mouse IgG and conjugated withAlexa 594 or 488 (Molecular Probes) were used at 1:500 dilution.Images were acquired with a Leica DMRXE epifluorescence micro-scope equipped with a 100X plan-neofluar oil-immersion objec-tive lens and a triple filter (set no. 61000V2 BS&M, ChromaTechnology, Rockingham, VT) for simultaneous visualization ofgreen (Alexa 488), red (Alexa 594), yellow (Alexa 488+Alexa 594)and blue (DAPI) fluorescence. The microscope was linked to aMicromax cooled CCD camera (RS Princeton Instrument) with ahigh-speed shutter driven by a Sutter Lambda 10-2 (Sutter Instru-ment) and Metamorph software (Universal Imaging Corporation)to capture the images.
Histology, immunohistochemistry and immunofluorescence
Paraffin embedded tissues were sectioned at 4 mm andmounted on poly-L-lysine-coated slides. Sections were stainedusing the periodic acid-Schiff (PAS) reaction (Cerri and Sasso-Cerri, 2003) and hematoxylin for histological observation. Afterdeparaffinization and rehydration, sections for immunostainingwere boiled in 10 mM citrate buffer (pH 6.0) for 30 min to promoteantigen retrieval. Endogenous peroxidase activity was blocked inmethanol containing 3% hydrogen peroxide for 15 min. Afterblocking with 2% normal serum (Vector Laboratories, Burlingame,CA) in PBS/Tween 20 (PBS/T) for 60 min, sections were incubatedat 4 1C overnight with: 2 mg/ml goat anti-BSG (R&D, Minneapolis,MN, USA), rabbit anti-TRA 98, 1:1000 (a kind gift of HiromitsuTanaka, Osaka University, Japan), guinea pig anti-SP-10, 1:400(a kind gift of Dr. P. Reddi, University of Virginia), rabbit anti-SOX9, 1:400 (Cosmo Bio Co., LTD, Japan), rabbit 0.2 mg/ml anti-CXADR (Santa Cruz, Santa Cruz, CA), 1 μg/ml rabbit anti-CX43 (CellSignaling Technology, Danvers, MA), 2 μg/ml rabbit anti-CDH2(Santa Cruz, Santa Cruz, CA), and 0.33 μg/ml rabbit anti-CLDN11(abcam, San Francisco, CA). Isotype control IgG was applied as anegative control. After washing in PBS/T, sections were incubatedwith secondary antibody either conjugated with CY3 at 1:1000with PBS/T (Jackson ImmunoResearch, West Grove, PA) or biotin(Vector Laboratories) diluted at 1:200 with PBS/T for 60 min atroom temperature. Sections for immunohistochemistry were thenincubated in ABC solution (Vector Laboratories) for 30 min, reactedwith metal-3,3′-diaminobenzidine (DAB) (Sigma, St. Louis, MO,USA) for minutes, and counterstained with hematoxylin. Sectionsfor immunofluorescence were then mounted using Vectashield(Vector Laboratories). All immunohistochemistry studies wereperformed on testes from three wild type and three Bsg KO mice.
TUNEL staining
Detection of DNA fragmentation of apoptotic cells was per-formed by using an ApopTag Plus Peroxidase In Situ Apoptosis Kit(Chemicon-Millipore, Billerica, MA, USA). Briefly, after deparaffi-nization and rehydration, sections were permeabilized by incuba-tion with proteinase K (20 mg/ml) for 15 min. Endogenousperoxidase activity was blocked in 3% hydrogen peroxide in PBSfor 5 min. After a 10 s incubation in equilibration buffer, sectionswere incubated in the terminal deoxynucleotidyl transferase (TdT)labeling reaction mix for 1hr at 37 1C. After a 10 min wash in stop/wash buffer, sections were incubated with anti-digoxigenin andfinally developed in DAB. For the negative controls, sections wereprocessed without TdT enzyme in the labelling reaction mix.Sections were counterstained with 0.5% (w/v) methyl green for10 min at room temperature. The total number of apoptotic cellswas counted in multiple histological sections of testes from 3 wild
type and 3 Bsg KO testes. We counted the number of apoptoticgerm cells in a minimum of 54 tubules for each testis. Thesenumbers were then divided by the number of seminiferoustubules in each tissue section to obtain the average number ofapoptotic germ cells per semniferous tubule.
Barrier function assays
The permeability of the BTB was assessed using the methoddeveloped by Chen et al. (1997). Briefly, three wild type and threeBsgKO mice (12 weeks old) were anesthetized using ketamine/xylazine (2 mg/0.2 mg). Testes were exposed by a small incisionthrough the scrotal layer. Next, a small opening in the tunicaalbuginea was gently created with fine forceps and 20 ul of 10 mg/mlEZ-Link Sulfo-NHS-LC-Biotin (Fisher Scientific) were injected using aHamilton syringe into the interstitium of one testis. The other testiswas injected with 20 ul of PBS as a control. After 30 min the animalswere euthanized by cervical dislocation, and the testes were imme-diately removed and fixed in 4% paraformaldehyde. Testes were thenprocessed for paraffin embedding. The slides were deparaffinized,rehydrated, boiled in 10 mM citrate buffer (pH 6.0) for 30 min andincubated with Fluorescein Isothiocyanate (FITC) conjugated strepta-vidin (Invitrogen, Grand Island, NY) for 60 min. Slides were thenmounted with vectashield (Vector Laboratories).
Lectin histochemistry
Tissue sections were deparaffinized in xylene three times for atotal of 60 min. The sections were rehydrated in decreasingconcentrations of ethanol. Endogenous peroxidase activity wasblocked in methanol containing 3% hydrogen peroxide for 20 min.After blocking with 1% BSA in PBS/T for 60 min, sections wereincubated at 4 1C overnight with 10 mg/ml biotinylated Grifforniasimplicifolia agglutinin (GSA) II (Vector Laboratories). GSA II thatwas preincubated with N-acetylglucosamine (Sigma, St. Louis, MO)was applied as a negative control. After washing in PBS/T, sectionswere incubated in ABC solution (Vector Laboratories) for 30 min,reacted with DAB for 3 min, and counterstained with hematoxylin.
Immunoprecipitation
Testes from 3 wild type mice were homogenized and proteinsextracted using TNE buffer (1% NP 40, 10 mM Tris pH 8, 1 mMEDTA). 500 μg of protein were incubated with 2 μg of goat poly-clonal antibody against mouse BSG (R&D, Minneapolis, MN) orgoat IgG for 60 min at 4 1C. Protein G plus-Agarose (Santa Cruz)was added into each protein sample overnight at 4 1C. Immuno-precipitates were collected by centrifugation at 2500 rpm at 4 1Cand washed with RIPA buffer (50 mM Tris pH 7.4, 150 mM NaCl,1 mM EDTA, 1% Triton X-100, 1% Sodium deoxycholate, 0.1% SDSand proteinase inhibitor). The protein was removed from thebeads by boiling for 5 min in 1X LSB (2% SDS, 10% glycerol,6.25 mM Tris pH 6.8). Proteins were separated by sodium dodecylsulfate polyacrylamide gel electrophoresis and transferred tonitrocellulose membranes.
Immunoblotting analysis
Membranes were blocked in 5% nonfat dry milk and thenprobed with 0.4 μg/ml goat polyclonal antibody against mouseBSG (R&D) overnight at 4 1C. The membranes were then washedand incubated with horseradish peroxidase-conjugated donkeyanti-goat IgG antibody (abcam, San Francisco, CA) at 1:5000dilution for 60 min at room temperature. The bound secondaryantibody was detected using SuperSignal West Pico Chemilumi-nescent Substrate kit (Fisher Scientific).
J. Bi et al. / Developmental Biology 380 (2013) 145–156148
Lectin blotting analysis
Membranes probed for immunoblotting analysis were strippedwith Restore Western Blot Stripping Buffer (Fisher Scientific). Afterblocking in 1X Carbo-free blocking buffer (Vector Laboratories) foran hour, the membranes were incubated with 1 mg/ml GSA IIovernight at 4 1C. Membranes were then incubated in 0.2 mg/mlImmunopure Avidin Hoseradish Peroxidase Conjugated (avidin-HRP) (Fisher Scientific) for 1 h. The bound avidin-HRP wasdetected using SuperSignal West Pico Chemiluminescent Substratekit (Fisher Scientific).
siRNA transfections and in vitro assay for germ cell-Sertoli celladhesion
Methods for mouse spermatogenic cells (GC-2) and Sertoli cell(SF7) adhesion assays (Akama et al., 2002) were performed aspreviously described with the following modifications. SiRNAscorresponding to mouse Bsg (sense: CGUAGAUUCCCAUCAUACAtt;antisense: UGUAUGAUGGGAAUCUACGgg) were purchased fromambion (Life technologies). The siRNA was transfected into theGC-2 cells following the protocol of siLentFect™ Lipid (Bio-Rad).Briefly, 3 μl of siLentFect Lipid transfection reagent was mixedwith 10 nM of siRNA to form complexes and dispersed into 6-wellcell culture plates at 37 1C for 48 h. Silencer select negative control#1 (Life technologies) was used as a negative control under thesame conditions.
The GC-2 cells were then labeled with [3H] Thymidine (2 μCi/ml)at 37 1C for another 24 h. 4.2�105 [3H]-labeled GC-2 cells wereadded to the SF7 cells and incubated at 37 1C for 4 h. After washing
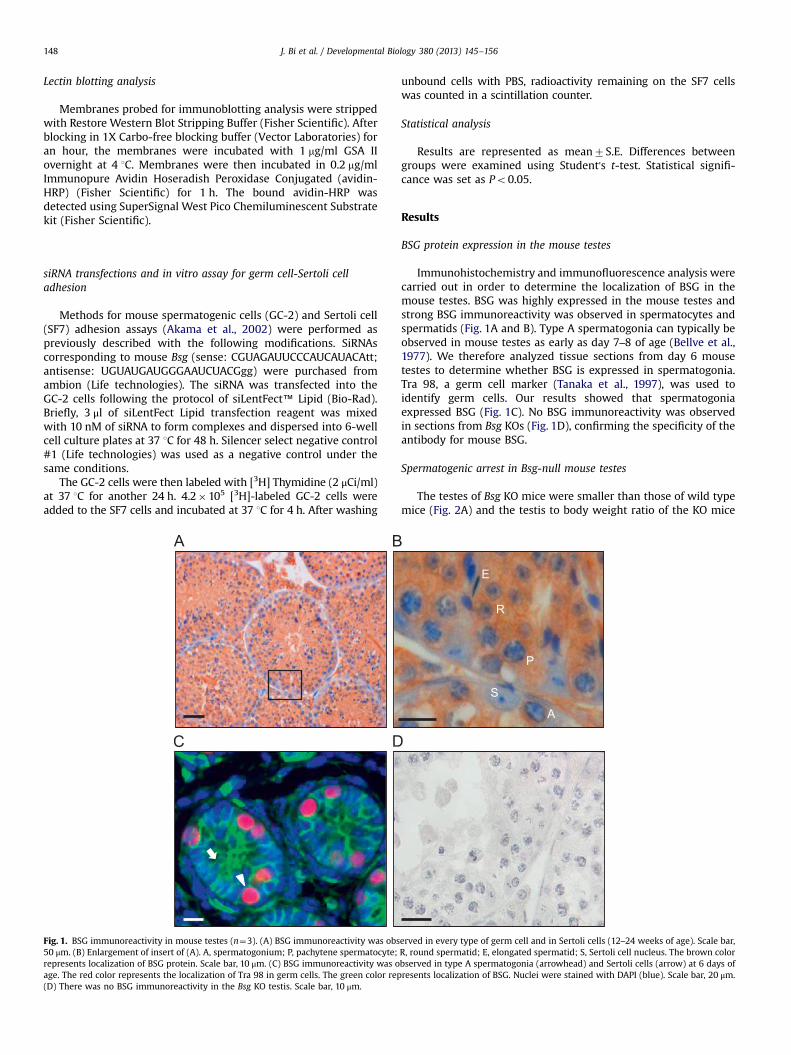
Fig. 1. BSG immunoreactivity in mouse testes (n¼3). (A) BSG immunoreactivity was obs50 μm. (B) Enlargement of insert of (A). A, spermatogonium; P, pachytene spermatocyte;represents localization of BSG protein. Scale bar, 10 μm. (C) BSG immunoreactivity was oage. The red color represents the localization of Tra 98 in germ cells. The green color rep(D) There was no BSG immunoreactivity in the Bsg KO testis. Scale bar, 10 μm.
unbound cells with PBS, radioactivity remaining on the SF7 cellswas counted in a scintillation counter.
Statistical analysis
Results are represented as mean7S.E. Differences betweengroups were examined using Student's t-test. Statistical signifi-cance was set as Po0.05.
Results
BSG protein expression in the mouse testes
Immunohistochemistry and immunofluorescence analysis werecarried out in order to determine the localization of BSG in themouse testes. BSG was highly expressed in the mouse testes andstrong BSG immunoreactivity was observed in spermatocytes andspermatids (Fig. 1A and B). Type A spermatogonia can typically beobserved in mouse testes as early as day 7–8 of age (Bellve et al.,1977). We therefore analyzed tissue sections from day 6 mousetestes to determine whether BSG is expressed in spermatogonia.Tra 98, a germ cell marker (Tanaka et al., 1997), was used toidentify germ cells. Our results showed that spermatogoniaexpressed BSG (Fig. 1C). No BSG immunoreactivity was observedin sections from Bsg KOs (Fig. 1D), confirming the specificity of theantibody for mouse BSG.
Spermatogenic arrest in Bsg-null mouse testes
The testes of Bsg KO mice were smaller than those of wild typemice (Fig. 2A) and the testis to body weight ratio of the KO mice
E
R
S
P
A
erved in every type of germ cell and in Sertoli cells (12–24 weeks of age). Scale bar,R, round spermatid; E, elongated spermatid; S, Sertoli cell nucleus. The brown colorbserved in type A spermatogonia (arrowhead) and Sertoli cells (arrow) at 6 days ofresents localization of BSG. Nuclei were stained with DAPI (blue). Scale bar, 20 μm.

WT KO
0
1
2
3
4
WT KO
Test
is/B
ody
wt.
(mg/
g)
*
WT KO
Fig. 2. Abnormalities in mouse testes due to loss of BSG expression (12–24 weeks of age; n¼3). (A) Testes from wild type and Bsg KO mice. Scale bar, 1 mm. (B) The testis tobody weight ratio of the KO mice was significantly less than that of the wild type mice. Data are means7S.E. and the star indicates statistical differences (Po0.05). (C) Thetestes from Bsg KO mice exhibit multinucleated giant cells (pachytene spermatocytes, arrowhead; round spermatids, arrow). Scale bar, 20 μm. (D) Abnormal chromatinpatterns in the pachytene spermatocytes of Bsg KO testes. Scale bar, 20 μm. (E) Increased numbers of spermatocytes with pyknotic nuclei. Scale bar, 20 μm. (F) TUNEL assay ofwild type and Bsg KO testes detected marked increases in apoptosis in Bsg KO testes. Apoptotic cells in the testes are stained brown in color. The rectangles located in theright-bottom of the images show enlarged views of the highlighted areas in each panel. Scale bar, 100 μm; scale bar (insert), 20 μm.
J. Bi et al. / Developmental Biology 380 (2013) 145–156 149
was significantly less than that of wild type mice (Fig. 2B).Testosterone levels were determined for wild type and Bsg KOmale mice. The values ranged widely from 0.2 to 1.2 ng/ml for bothgroups and were not different from one another. We nextexamined the histology of both Bsg KO and wild type testes. Bothpachytene spermatocytes (Fig. 2C, arrowhead) and round sperma-tids (Fig. 2C, arrow) formed multinucleated giant cells in contrastto wild type testes where none were found. Moreover, there werea lot of abnormal chromatin patterns, with micronuclear forma-tion, in the pachytene spermatocytes as they enter meiotic division(Fig. 2D, arrowhead) and evidence of cell death based on thepresence of pyknotic nuclei (Fig. 2E, arrowhead). We confirmedprevious studies showing a reduction in the number of spermato-genic cells in Bsg KO mice (Igakura et al., 1998), as null mutanttestes exhibited a complete loss of elongated spermatids andmature spermatozoa. We used a TUNEL assay to evaluate apoptosisof germ cells in Bsg KO mice. As shown in Fig. 2F, there was amarked increase in the number of apoptotic cells/semniferoustubule in the testes of Bsg KO mice (3.1670.29) as compared to
wild type testes (0.1770.01). The position of these apoptotic cellsin the seminiferous epithelium as well as their morphologyindicated that they were spermatocytes.
Because previous studies had not determined the progressionof meiosis or whether meiotic division occurred in Bsg-null testes,we used classical markers for meiotic progression in immuno-fluorescence analyses. Antibodies that recognized SYCP1 (thecentral element of the synaptonemal complex), SYCP3 (the lateralelement of the synaptonemal complex), phosphorylated histoneH2AFX (commonly known as γH2AX), a histone marker of DNAdouble-strand breaks, and H1T (a male germ-cell specific histonemarker for spermatocytes that have reached the mid-pachytenepoint of meiotic prophase) were used to assess whether thespermatocytes of Bsg KO males entered and progressed throughmeiosis (Costa et al., 2005; Inselman et al., 2003). Our resultsshowed that male germ cells of Bsg KO mice displayed normalpatterns of chromosome dynamics (Fig. 3), exhibiting homologouschromosome synapsis (Fig. 3G and H) as well as normal labeling ofthe X–Y bivalent (Fig. 3J), which, as in wild type germ cells, is the

J. Bi et al. / Developmental Biology 380 (2013) 145–156150
only unpaired chromosome (Fig. 3C and I). As seen in Fig. 3K, BsgKO spermatocytes reached the mid-pachytene stage, a time atwhich competence to enter meiotic division arises. This wasconfirmed by the presence of MI spermatocytes (Fig. 3L). Afterchromosomes underwent desynapsis at the end of meiotic pro-phase and entered the MI division phase, SYCP1 labeling disap-peared, and SYCP3 labeling was observed at the centromeric ends
Pachynema
WT
KO
SYCP1
SYCP1 SYCP1 SYCP3
SYCP1 SYCP3
SYCP3
SYCP3
Fig. 3. Surface-spread chromatin preparations documenting normal chromosome pairin(12–24 weeks of age; n¼3). ((A)–(F)) Immunofluorescence with antibodies to SYCP1, SYCantibodies to SYCP1, SYCP3, γH2AX, or H1t in Bsg KO spermatocytes. Autosomal homolocentral element and a lateral element of the SC, respectively; and the merged panel idenwere sequestered in a chromatin domain which was labeled intensely with antibody rehave reached or progressed beyond the mid-pachytene stage. SYCP1 labeling disappears,the centromeric ends of the chromosomes ((F), (L)). There was no difference in localiza
A B
C
Sp10
D
GAP
DH
WT1 WT2 WT3 KO1 KO2 KO3
WT
Fig. 4. Localization and quantitation of SP-10 protein in testes of wild type and Bsg KO mSP-10 in wild type and (B) Bsg KO testis sections. (C) Total proteins were isolated fromdensitometric analysis of the immunoblotting. Bars represent the mean7S.E. of threerectangles located in the right-bottom corners of histological images show enlarged viewprotein. Scale bar, 100 μm; scale bar (enlargement), 20 μm.
of the chromosomes (Fig. 3F and L). In addition, some meiosis IIspermatocytes were also observed in the KO preparations (datanot shown).
To determine whether post-meiotic germ cells are present inBsg KO mice, we immunostained adult testis sections for theacrosome marker SP-10 (known as ACRV1) (Herr et al., 1990;Reddi et al., 1999). This protein only begins to be expressed by
Mid-Pachynema Metaphase I
γH2AX SYCP3
γH2AX SYCP3
H1t SYCP3
H1t SYCP3 SYCP1 SYCP3
SYCP1 SYCP3
g, recombination and chromatin remodeling in wild type and Bsg KO spermatocytesP3, γH2AX, or H1T in wild type spermatocytes. ((G)–(L)) Immunofluorescence withgues were fully synapsed in terms of the labeling patterns of SYCP1 and SYCP3, atified the non-homologous X and Y ((C), (I), arrowhead). The X and Y chromosomescognizing γH2AX. The antibody to H1T protein is a marker for spermatocytes thatwhen chromosomes enter the MI division phase, and the SYCP3 labeling was seen attion of these markers between wild type and Bsg KO spermatocytes.
0
0.2
0.4
0.6
0.8
1
1.2
WT KO
Inte
nsity
of E
xpre
ssio
n
*
KO
ice (12–24 weeks of age; n¼3). (A) Immunohistochemistry for the acrosome markerwhole testes and subjected to immunoblotting. (D) Bar graphs summarizing theindependent mice with the data normalized to the loading control, GAPDH. Thes of the highlighted areas in each panel. The brown color represents localization of

J. Bi et al. / Developmental Biology 380 (2013) 145–156 151
post-meiotic germ cells that are early stage (step 3) roundspermatids (Dr. Prabhu Reddi, personal communication). Asshown in Fig. 4A, in the wild type testes, early stage (step 3)round spermatids showed a punctate expression of SP-10 in theacrosomal granule which progresses into a stronger, crescent-shaped pattern of SP-10 expression in elongated spermatids witha fully developed acrosome. In the testes of Bsg KO mice thepunctate pattern of SP-10 expression in the acrosomal granule ofearly round spermatids was present but overall expression of SP-10was very low because there are no elongated spermatids with fullydeveloped acrosomes in the testes of these KO mice. (Fig. 4B). We
WT
Fig. 5. Abnormalities in testes of aged Bsg KO mice (n¼2). (A) PAS staining in aged wildsections. Numerous vacuoles (arrowhead) were formed within the seminiferous epithespermatogenesis in the seminiferous epithelium of aged Bsg KOs. Scale bar, 20 μm. (E) Sbar, 20 μm. (F) Abundent degenerating, sloughed germ cells were found in the lumen of20 μm. The rectangles located in the right-bottom corner of images are the enlarged vie
used immunoblotting to quantify the level of SP-10 in both wildtype and Bsg KO testes. Our results showed that SP-10 was stronglyexpressed in wild type testes. However, the level of SP-10 in Bsg KOtestes was almost too low to detect which corresponded with ourimmunohistochemistry results (Fig. 4C and D).
Abnormalities in aged mouse testes (15–17 months) caused by loss ofBSG expression
Numerous vacuoles were observed within the seminiferousepithelium of Bsg KO testes of aged mice (Fig. 5C, arrowhead) in
KO
type, (B) young Bsg KO (12–24 weeks) and (C) aged Bsg KO (15–17 months) testislium of aged Bsg KOs. Scale bar, 20 μm. (D) Loss of spermatogonia and failure ofertoli cells (arrowhead) were observed clumped in the lumen of Bsg KO mice. Scaleefferent ductules in Bsg KO mice (arrowhead). Scale bar, 100 μm; scale bar (insert),ws of the highlighted areas in each panel.

J. Bi et al. / Developmental Biology 380 (2013) 145–156152
contrast to wild type (Fig. 5A) and younger Bsg KO (Fig. 5B) testes.Some areas of the seminiferous epithelium showed a significantloss of spermatogonia and failure of spermatogenesis (Fig. 5D). Insome tubules we also observed Sertoli cells clumped in the lumen,appearing to have lost their attachment to the basement mem-brane (Fig. 5E, arrowhead). SOX9, a Sertoli cell marker(Hemendinger et al., 2002), was used to identify these Sertoli cells(red). The lumen of Bsg KO efferent ductules contained degenerat-ing sloughed germ cells and areas of aggregated debris (Fig. 5F,arrowhead). The epididymal lumen was mostly devoid of germcells but some luminal debris was present with occasional degen-erating germ cells.
Integrity of the BTB and expression of the BTB related adhesionmolecules CXADR, CX43, CLDN11 and CDH2 in the testes of wild typeand Bsg KO mice
We next investigated the integrity of the BTB in Bsg KO testes byinjecting a biotin tracer into the testes. In the wild type testes thebiotin tracer was restricted to the interstitium and the basalcompartment of the seminiferous tubules (Fig. 6A and B). In contrast,in Bsg KO mice, the biotin tracer in some tubules penetrated beyondthe BTB and surrounded early pachytene spermatocytes, but did notenter the lumen (Fig. 6H and I, arrowhead).
We therefore carried out immunohistochemical analysis of bothwild type and Bsg KO testes to determine the expression pattern ofsome well-known BTB-related adhesion proteins. We observed thatCXADR immunoreactivity was strongly expressed along the basalcompartment of the seminiferous tubules of wild type mice (Fig. 6C,arrowhead). A very similar localization pattern of CXADR protein wasalso observed in Bsg KO testes (Fig. 6J, arrowhead). CX43 immunor-eactivity in wild type testes was observed in spermatocytes and roundspermatids. Weak immunoreactivity was detected in elongated sper-matids but was undetectable in spermatogonia. A very strong immu-noreactivity was detected at the site of the BTB (Fig. 6D, arrowheads).The localization pattern of CX43 protein in Bsg KO testes was verysimilar (Fig. 6K). CLDN11 immunoreactivity was detected at the siteof the BTB of the seminiferous epithelium in both wild type and
biotin tracer CXADR CX43
WT
KO
Fig. 6. Integrity of the BTB and localization of BTB related adhesion molecules CXADR, Cage; n¼3). A biotin tracer was injected into the testicular interstitial space of both wild tytracer and DAPI. The biotin tracer penetrated around the early pachytene spermatocytes(F)): Immunohistochemistry with antibodies to CXADR, CX43, CLDN11 or CDH2 in wild tyCLDN11 or CDH2 in Bsg KO testes. There was no difference in expression of CXADR, CX43was greatly reduced at the site of the BTB of Bsg KO mice. Panels ((G), (N)): Sections wereThe brown color represents the localization of each protein, while green color representsadhesion molecule at the site of the BTB. Scale bar, 20 μm.
Bsg KO testes and formed an almost continuous ring of expression(Fig. 6E and L).
We also examined expression of CDH2 and saw a significantalteration in expression in Bsg KO testes. CDH2 immunoreactivity inwild type testes was observed at the site of cell-cell contacts betweenSertoli cells and spermatogenic cells (Fig. 6F). However, whereas wildtype testes showed strong expression of CDH2 along the basalcompartment of the seminiferous tubule (Fig. 6F, arrowheads), CDH2expression was greatly reduced in the basal compartment of theseminiferous tubules (the site of the BTB) of Bsg KO mice (Fig. 6M). Nosignal was detected in the negative control sections (Fig. 6G and N).
GlcNAc terminated N-glycans localization in testes of wild type andBsg KO mice
GlcNAc terminated tri-antennary and fucosylated N-glycanstructures have been shown to play a key role in germ cell-Sertoli cell adhesion (Akama et al., 2002). Therefore, we carriedout lectin histochemistry analysis on both wild type and Bsg KOtestes using Griffornia simplicifolia agglutinin (GSA) II whichrecognizes GlcNAc terminated N-glycans (Akama et al., 2002).We found that the expression pattern of GlcNAC terminatedcarbohydrates showed remarkable differences between wild typeand Bsg KO testes. In wild type testes, spermatogenic cells werestrongly stained with GSA II (Fig. 7A). In contrast, immunostainingin the Bsg KO testes was greatly decreased (Fig. 7B). No signal wasfound in the negative control sections (Fig. 7C).
Determining whether GlcNAc terminated N-glycans carbohydratesare linked to BSG
BSG is a highly glycosylated protein rich in N-glycans (Miyauchiet al., 1990; Tang et al., 2004). Lectin blotting analysis was carried outto determine whether GlcNAc terminated N-glycans are one of thecarbohydrates linked to BSG. We first immunoprecipitated proteinlysates from mouse testes using a BSG antibody and then performedimmunoblotting using this same BSG antibody to confirm that wehad successfully pulled down BSG protein (Fig. 8). No BSG signal was
CLDN11 CDH2 ctrl
X43, CLDN11 and CDH2 in the testes of wild type and Bsg KO mice (12–24 weeks ofpe ((A), (B)) and Bsg KO mice ((H), (I)). Panels ((B), (I)): The merged images of biotinin compartment of Bsg KO seminiferous tubules ((H), (I), arrowhead). Panels ((C)–pe testes. Panels ((J)–(M)): Immunohistochemistry with antibodies to CXADR, CX43,and CLDN11 between wild type and Bsg KO testes. However, the expression of CDH2incubated with non-specific IgG as a negative control and there was no any staining.the localization of biotin tracer. Arrowheads indicated the immunoreactivity of each

L+
N
KOWT
Fig. 7. Localizations of GlcNAc terminated N-glycans in the testes of wild type and Bsg KO mice (12–24 weeks of age; n¼3). Localization of GlcNAc-terminated N-glycans inwild type testes (A) and in Bsg KO testes (B). GSA II was strongly expressed in all phases of wild type spermatogenic cells except spermatogonia (arrowhead). This lectinstaining, however; was greatly decreased in the Bsg KO testes. (C) Sections incubated with GSA II that was pretreated with N-acetylglucosamine as a negative control did notshow any positive staining. The rectangles located in the bottom-right corner of images are the enlarged views of the highlighted areas in each panel. The brown colorrepresents localization of GlcNAc-terminated N-glycans. Scale bar, 100 μm; scale bar (insert), 20 μm.
37kD
50kD
IgG Input BSG GSA II
IP - BASIGIN
Fig. 8. GlcNAc-terminated N-glycans are one of the carbohydrates linked to BSG(12–24 weeks of age; n¼3). BSG protein was immunoprecipitated from mousetestes lysates with BSG antibody. Immunoblotting with BSG antibody indicated thatthe protein immunoprecipitated from mouse testes lysates was BSG. No BSG signalwas observed from the IgG control. After stripping the blot, lectin blotting with GSAII was performed to show the presence of GlcNAc terminated N-glycans.
J. Bi et al. / Developmental Biology 380 (2013) 145–156 153
observed from the IgG control, confirming the specificity of theantibody for mouse BSG. These membranes were then stripped andreprobed with GSAII. Our lectin blotting clearly showed the presenceof GlcNAc terminated carbohydrates corresponding with the BSGprotein. These results confirmed that GlcNAc terminated N-glycansare one of the carbohydrates linked to BSG.
Bsg knockdown reduces adhesion of spermatocytes to Sertoli cells
In order to determine whether BSG plays a role in adhesion ofspermatogenic cells to Sertoli cells, we performed an in vitro
adhesion assays with GC-2 and SF7 cells transfected with BsgsiRNA. As shown in Fig. 9A, the expression of BSG protein wasknocked down completely in GC-2 cells transfected with BsgsiRNA, but not in the negative control group 72 h after transfec-tion. Knockdown of Bsg expression in GC-2 cells resulted in asignificant reduction in adhesion (75%) between GC-2 and SF7cells compared with controls as quantitated by the levels oftritiated thymidine bound to SF7 cells (Fig. 9B).
Discussion
BSG is a transmembrane glycoprotein that is expressed inseveral reproductive tissues and plays an essential role in bothmale and female fertility (Igakura et al., 1998; Kuno et al., 1998).Previous studies have shown by microscopic analyses that manyBsg KO spermatocytes are arrested and degenerate at the meta-phase of the first meiosis, with only a small number of germ cellsdifferentiating into step 1–2 spermatids (Igakura et al., 1998;Toyama et al., 1999). Our results showed that spermatogenesis inBsg KO mice is arrested at the early round spermatid stages (step3 or 4), before any spermatid differentiation occurred. Moreover,we also found that there was a large increase in the number ofgerm cells undergoing apoptosis in Bsg KO testes. These results areconsistent with previous reports (Chen et al., 2012; Igakura et al.,1998; Toyama et al., 1999) and confirm that BSG is essential for thedevelopment/differentiation of germ cells to round spermatids.

BSG
GAPDH
NC SiRNA
02000400060008000
100001200014000160001800020000
NC SiRNA
CPM
*
Fig. 9. Adhesion of GC-2 spermatocyte cells transfected with Bsg siRNA to SF7Sertoli cells (n¼3). (A) Loss of BSG protein expressions in Bsg siRNA transfectedcells. Protein lysates for BSG were isolated from both Bsg siRNA treated and controlGC-2 cells and subjected to immunoblotting. GAPDH served as a loading control.(B) Adhesion between siRNA treated GC-2 cells and SF7 cells was significantlydecreased in comparison to the control group. Bars represent the CPM7S.E. ofthree independent experiments and the asterisk indicates the statistical difference(Po0.05).
J. Bi et al. / Developmental Biology 380 (2013) 145–156154
In addition, our studies show for the first time that BSG is alsonecessary for maintenance of the BTB in the semniferous tubules.
Although some metaphase spermatocytes are present in Bsg KOtestes, it was important to examine the key events of meioticdivision more carefully in Bsg KO testes. Our results showed thatBsg KO spermatocytes undergo normal homologous chromosomesynapsis and progression to the metaphase of first meiosis asdemonstrated by immunofluorescent labeling of chromosomeswith SYCP1, SYCP3, H1T and γH2AX. However, we observed anumber of abnormal chromatin patterns in the pachytene sper-matocytes as they enter meiotic division, and significant cell deathwas observed in the tubules. We also used an antibody to theround spermatid marker SP-10 to identify spermatids in adulttestes sections. Mouse spermiogenesis is divided into 16 stepsaccording to the shape of the developing acrosome. These stagesare: Golgi phase, steps 1–4; cap phase, steps 5–7; acrosome phase,steps 8–12; and maturation phase, steps 13–16 (Russell, 1990;Segretain and Roussel, 1988). We observed punctate staining forSP-10 in the acrosomal granules of early (step 3) round spermatidsin Bsg KO mice indicating that at least a portion of the germ cellsdid progress to this stage before being lost.
Spermatogenesis begins with mitosis of spermatogonia andformation of early spermatocytes. Whenwe examined testes of oldBsg KO mice (around 15 months old), we observed that numerousvacuoles had formed within the seminiferous epithelium andpachytene spermatocytes showed abnormal nuclei and cell death(data not shown). In some tubules, Sertoli cells were observedclumped in the lumen, appearing to have lost their attachmentto the basement membrane. Moreover, some areas of the
seminiferous epithelium showed significant loss of spermatogoniaand failure of spermatogenesis. The phenotypes described abovewere not found in testes of aged wild type or young Bsg KO males.Thus, in older KO males we observed a more severe phenotypesuggesting that over time the impact of loss of BSG on spermato-genesis becomes more severe and cumulative.
The Sertoli cell is known to secrete fluid to form a seminiferoustubular lumen (Russell et al., 1989). The lumen size in Bsg KO testeswas 65% smaller than that of wild type testes (data not shown).Furthermore, the lumen of Bsg KO efferent ductules containeddegenerating sloughed germ cells and areas of aggregated debris,suggestive of stagnation due to reduced flow of fluid from thetestis. Taken together, these results support that BSG may alsoregulate the secretion of seminiferous tubule fluid from Sertolicells during spermatogenesis.
Integrity of the BTB is essential for survival of the postmeioticgerm cells and injection of biotin tracer is commonly used todetermine the integrity of the BTB (Chen et al., 1997). We observedan altered pattern of biotin localization within the testes of Bsg KOmice indicating that the integrity of the BTB is compromised in BsgKO testes. We further investigated this finding by analyzingimmunolocalization of several BTB component proteins.
The junctional proteins CXADR, CX43, claudins, and CDH2 areimportant for maintenance of BTB integrity (Cheng and Mruk,2009; Komljenovic et al., 2009; Pointis and Segretain, 2005; Wonget al., 2008). We did not observe any difference in localization ofCXADR, CX43 or CLDN11 between wild type and Bsg KO testes.However, the expression of CDH2 was greatly reduced at the basalcompartment of the seminiferous tubule (the site of the BTB) inBsg KO mice. CDH2 belongs to a family of calcium-dependent,homophilic cell adhesion molecules that are expressed in anumber of tissues (Gumbiner, 2005). CDH2, along with β-catenin,is one of the component proteins of the BTB (Cheng and Mruk,2009; Elkin et al., 2010; Lee et al., 2003). Studies have shown thatCDH2 is also involved in spermatogenesis. It mediates Sertoli cell-spermatogenic cell adhesion (Newton et al., 1993), and may alsofacilitate germ cell migration during spermatogenesis (Chunget al., 1998). Since the expression of CDH2 was greatly reducedin the testes of Bsg KO mice this suggests that BSG may regulatethe expression of CDH2 either directly or indirectly. Takentogether, our data show that BTB integrity is compromised butnot destroyed completely in Bsg KO testes. This may be due to thepresence of several other junction proteins such as CXADR, CX43and CLDN11 that are also known to maintain BTB integrity andwhose expression is not altered in Bsg KO testes. Our results show,for the first time, that loss of BSG expression and reducedexpression of the basal ES protein CDH2 disrupts Sertoli cell BTBintegrity.
N-glycans are branched carbohydrates that are attached to theamide nitrogen of asparagine residues of peptides. There are threebroad classes of N-glycans: high mannose, hybrid, and complexand all are found attached to mammalian glycoproteins (Alberts,2002; Fukuda and Akama, 2003). These all share a common corestructure that contains two GlcNac and three mannose, but differin their outer branches. Biosynthesis of N-glycans includes “enbloc” transfer of oligosaccharides made of glucose, mannose andGlcNac utilizing several different enzymes to complete the process.For example, N-acetylglucosaminyltransferase I (GlcNAc-TI) isessential for the conversion of high mannose type N-glycans tohybrid type N-glycans. Mice lacking GlcNAc-TI die at embryonicday 10 due to inhibition of development (Ioffe and Stanley, 1994).The enzyme A-mannosidase II (MX) acts by removing two man-nosyl residues from high mannose type N-glycans (Akama andFukuda, 2006). Mice lacking MX showed a great reduction in thelevel of GlcNAc-terminated N-glycans, and the spermatogeniccells of MX null mice fail to adhere to Sertoli cells leading to the

J. Bi et al. / Developmental Biology 380 (2013) 145–156 155
failure of spermatogenesis (Akama et al., 2002). Expression ofoligosaccharide 310.11 in MX KO testes was reduced by ∼50%compared to that in wild type testes and pretreatment of culturedSertoli cells with this oligosaccharide dramatically blocked thebinding between Sertoli cells and germ cells (Akama et al., 2002)confirming the importance of this GlcNAc-terminated N-glycan forgerm cell adhesion.
BSG is a transmembrane glycoprotein rich in N-glycans (Tanget al., 2004). Our results showed that GlcNAc terminated carbohy-drates were greatly decreased in testes of Bsg KO mice. In addition,lectin blotting clearly showed the presence of GlcNAc terminatedcarbohydrates corresponding with BSG protein. Thus BSG actsas an attachment molecule for GlcNAc-terminated N-glycans.To determine whether BSG plays a role in adhesion of spermato-genic cells to Sertoli cells, we performed in vitro adhesion assayswith SF7 cells and GC-2 cells transfected with Bsg siRNA. We foundthat knockdown of Bsg in GC-2 cells caused a marked reduction(over 75%) in adhesion of GC-2 to SF7 cells. This failure of adhesionwas due to a defect in spermatogenic cells, not in Sertolicells, which is consistent with the earlier reports demonstratingthe importance of GlcNAc-terminated N-glycans expression ingerm cells for attachment to Sertoli cells (Akama et al., 2002).Our findings support that BSG expressed on the surface ofspermatogenic cells is important for germ cell adhesion to Sertolicells.
Testes of Bsg KO mice also exhibited multinucleated giant cells.These multinucleated giant cells may form due to the failure ofgerm cells to lose their intercellular bridges. Similar multinu-cleated giant cells have been observed in the testes of MX KO mice(Akama et al., 2002), cAMP responsive element modulator (Crem)KO mice (Nantel et al., 1996) and Bcl-w KO mice (Print et al., 1998),which also show elevated apoptosis of male germ cells similar toBsg KO males. The phenotype of the Crem KO mouse is very similarto the Bsg KO mouse (Nantel et al., 1996). For example, latespermatids are completely absent and there is a significantincrease in apoptotic germ cells. BCL-W is a pro-survival proteinbelonging to the BCL2 family (Print et al., 1998). Bcl-w KO mice alsoshow a marked reduction in the number of elongating spermatids.Understanding the mechanism of action of both CREM and BCL-Win the testis would definitely provide insights into the functions ofBSG during spermatogenesis.
In conclusion, our studies found that spermatocytes lacking BSGexpression showed normal homologous chromosome synapsis andprogression to the metaphase of first meiosis. However, spermato-genesis in Bsg KO males was arrested at the early (step 3 or 4) roundspermatid stage. The loss of BSG in Bsg KO testes led to greatlyreduced expression of CDH2 in the basal compartment of theseminiferous tubule and to disruption of the integrity of the BTB.Furthermore, BSG also appears to serve as an important attachmentmolecule between Sertoli cells and germ cells since inhibition of BSGexpression in GC-2 cells significantly reduced adhesion between GC-2 and SF7 cells. This attachment may be mediated by the N-glycanscontained in BSG. Thus the loss of BSG significantly impairs interac-tions between gametes and Sertoli cells. This is the reason for thelarge increase in the number of germ cells undergoing apoptosis inBsg KO testes resulting in azoospermia. Our results provide adirection for future studies of the interaction between Sertoli cellsand germ cells. Elucidation of the specific functions of BSG duringspermatogenesis may lead to the development of new therapeutictargets for reversing azoospermia.
Acknowledgements
We would like to thank Dr. Prabhu Reddi for his thoughtful andvery helpful discussion during the course of writing this manuscript.
Grant Support: This research was supported by the EuniceKennedy Shriver NICHD/NIH through cooperative agreement [U54HD40093] to R.A.N. as part of the Specialized Cooperative CentersProgram in Reproduction and Infertility Research and The Uni-versity of Virginia Center for Research in Reproduction LigandAssay and Analysis Core is supported by the Eunice KennedxyShriver NICHD/NIH (SCCPIR) Grant U54-HD28934.
References
Akama, T.O., Fukuda, M.N., 2006. N-glycan structure analysis using lectins and analpha-mannosidase activity assay. Methods Enzymol. 416, 304–314.
Akama, T.O., Nakagawa, H., Sugihara, K., Narisawa, S., Ohyama, C., Nishimura, S.,O'Brien, D.A., Moremen, K.W., Millan, J.L., Fukuda, M.N., 2002. Germ cell survivalthrough carbohydrate-mediated interaction with Sertoli cells. Science 295,124–127.
Alberts, B., 2002. Molecular Biology of the Cell, fourth ed. Garland Science, NewYork.
Bellve, A.R., Cavicchia, J.C., Millette, C.F., O'Brien, D.A., Bhatnagar, Y.M., Dym, M.,1977. Spermatogenic cells of the prepuberal mouse. Isolation and morpholo-gical characterization. J. Cell Biol. 74, 68–85.
Cerri, P.S., Sasso-Cerri, E., 2003. Staining methods applied to glycol methacrylateembedded tissue sections. Micron 34, 365–372.
Chen, H., Fok, K.L., Jiang, X., Jiang, J., Chen, Z., Gui, Y., Chan, H.C., Cai, Z., 2012. CD147regulates apoptosis in mouse spermatocytes but not spermatogonia. Hum.Reprod. 27, 1568–1576.
Chen, L., Bi, J., Nakai, M., Bunick, D., Couse, J.F., Korach, K.S., Nowak, R.A., 2010.Expression of basigin in reproductive tissues of estrogen receptor-{alpha} or-{beta} null mice. Reproduction 139, 1057–1066.
Chen, Y., Merzdorf, C., Paul, D.L., Goodenough, D.A., 1997. COOH terminus ofoccludin is required for tight junction barrier function in early Xenopusembryos. J. Cell Biol. 138, 891–899.
Cheng, C.Y., Mruk, D.D., 2009. An intracellular trafficking pathway in the semi-niferous epithelium regulating spermatogenesis: a biochemical and molecularperspective. Crit. Rev. Biochem. Mol. Biol. 44, 245–263.
Chung, S.S., Mo, M.Y., Silvestrini, B., Lee, W.M., Cheng, C.Y., 1998. Rat testicularN-cadherin: its complementary deoxyribonucleic acid cloning and regulation.Endocrinology 139, 1853–1862.
Cobb, J., Cargile, B., Handel, M.A., 1999a. Acquisition of competence to condensemetaphase I chromosomes during spermatogenesis. Dev. Biol. 205, 49–64.
Cobb, J., Miyaike, M., Kikuchi, A., Handel, M.A., 1999b. Meiotic events at thecentromeric heterochromatin: histone H3 phosphorylation, topoisomerase IIalpha localization and chromosome condensation. Chromosoma 108, 412–425.
Cobb, J., Reddy, R.K., Park, C., Handel, M.A., 1997. Analysis of expression andfunction of topoisomerase I and II during meiosis in male mice. Mol. Reprod.Dev 46, 489–498.
Costa, Y., Speed, R., Ollinger, R., Alsheimer, M., Semple, C.A., Gautier, P., Maratou, K.,Novak, I., Hoog, C., Benavente, R., Cooke, H.J., 2005. Two novel proteinsrecruited by synaptonemal complex protein 1 (SYCP1) are at the centre ofmeiosis. J. Cell Sci. 118, 2755–2762.
Dym, M., Fawcett, D.W., 1970. The blood-testis barrier in the rat and thephysiological compartmentation of the seminiferous epithelium. Biol. Reprod.3, 308–326.
Elkin, N.D., Piner, J.A., Sharpe, R.M., 2010. Toxicant-induced leakage of germ cell-specific proteins from seminiferous tubules in the rat: relationship to blood-testis barrier integrity and prospects for biomonitoring. Toxicol. Sci. 117,439–448.
Fossum, S., Mallett, S., Barclay, A.N., 1991. The MRC OX-47 antigen is a member ofthe immunoglobulin superfamily with an unusual transmembrane sequence.Eur. J. Immunol. 21, 671–679.
Fukuda, M.N., Akama, T.O., 2003. The role of N-glycans in spermatogenesis.Cytogenet. Genome Res. 103, 302–306.
Gow, A., Southwood, C.M., Li, J.S., Pariali, M., Riordan, G.P., Brodie, S.E., Danias, J.,Bronstein, J.M., Kachar, B., Lazzarini, R.A., 1999. CNS myelin and Sertoli cell tightjunction strands are absent in Osp/claudin-11 null mice. Cell 99, 649–659.
Gumbiner, B.M., 2005. Regulation of cadherin-mediated adhesion in morphogen-esis. Nat. Rev. Mol. Cell Biol. 6, 622–634.
Hemendinger, R.A., Gores, P., Blacksten, L., Harley, V., Halberstadt, C., 2002.Identification of a specific Sertoli cell marker, Sox9, for use in transplantation.Cell Transplant. 11, 499–505.
Herr, J.C., Flickinger, C.J., Homyk, M., Klotz, K., John, E., 1990. Biochemical andmorphological characterization of the intra-acrosomal antigen SP-10 fromhuman sperm. Biol. Reprod. 42, 181–193.
Hofmann, M.C., Abramian, D., Millan, J.L., 1995. A haploid and a diploid cell coexistin an in vitro immortalized spermatogenic cell line. Dev. Genet. 16, 119–127.
Hofmann, M.C., Narisawa, S., Hess, R.A., Millan, J.L., 1992. Immortalization of germcells and somatic testicular cells using the SV40 large T antigen. Exp. Cell. Res.201, 417–435.
Igakura, T., Kadomatsu, K., Kaname, T., Muramatsu, H., Fan, Q.W., Miyauchi, T.,Toyama, Y., Kuno, N., Yuasa, S., Takahashi, M., Senda, T., Taguchi, O., Yamamura,K., Arimura, K., Muramatsu, T., 1998. A null mutation in basigin, an

J. Bi et al. / Developmental Biology 380 (2013) 145–156156
immunoglobulin superfamily member, indicates its important roles in peri-implantation development and spermatogenesis. Dev. Biol. 194, 152–165.
Inselman, A., Eaker, S., Handel, M.A., 2003. Temporal expression of cell cycle-relatedproteins during spermatogenesis: establishing a timeline for onset of themeiotic divisions. Cytogenet. Genome Res. 103, 277–284.
Ioffe, E., Stanley, P., 1994. Mice lacking N-acetylglucosaminyltransferase I activitydie at mid-gestation, revealing an essential role for complex or hybrid N-linkedcarbohydrates. Proc. Natl. Acad. Sci. U.S.A. 91, 728–732.
Komljenovic, D., Sandhoff, R., Teigler, A., Heid, H., Just, W.W., Gorgas, K., 2009.Disruption of blood-testis barrier dynamics in ether-lipid-deficient mice. CellTissue Res. 337, 281–299.
Kuno, N., Kadomatsu, K., Fan, Q.W., Hagihara, M., Senda, T., Mizutani, S., Muramatsu,T., 1998. Female sterility in mice lacking the basigin gene, which encodes atransmembrane glycoprotein belonging to the immunoglobulin superfamily.FEBS Lett. 425, 191–194.
Lee, N.P., Mruk, D., Lee, W.M., Cheng, C.Y., 2003. Is the cadherin/catenin complex afunctional unit of cell-cell actin-based adherens junctions in the rat testis? Biol.Reprod. 68, 489–508.
Miyauchi, T., Kanekura, T., Yamaoka, A., Ozawa, M., Miyazawa, S., Muramatsu, T.,1990. Basigin, a new, broadly distributed member of the immunoglobulinsuperfamily, has strong homology with both the immunoglobulin V domainand the beta-chain of major histocompatibility complex class II antigen.J. Biochem. 107, 316–323.
Morrow, C.M., Tyagi, G., Simon, L., Carnes, K., Murphy, K.M., Cooke, P.S., Hofmann,M.C., Hess, R.A., 2009. Claudin 5 expression in mouse seminiferous epitheliumis dependent upon the transcription factor ets variant 5 and contributes toblood-testis barrier function. Biol. Reprod. 81, 871–879.
Nantel, F., Monaco, L., Foulkes, N.S., Masquilier, D., LeMeur, M., Henriksen, K.,Dierich, A., Parvinen, M., Sassone-Corsi, P., 1996. Spermiogenesis deficiency andgerm-cell apoptosis in CREM-mutant mice. Nature 380, 159–162.
Nehme, C.L., Fayos, B.E., Bartles, J.R., 1995. Distribution of the integral plasmamembrane glycoprotein CE9 (MRC OX-47) among rat tissues and its inductionby diverse stimuli of metabolic activation. Biochem. J. 310 (Pt 2), 693–698.
Newton, S.C., Blaschuk, O.W., Millette, C.F., 1993. N-cadherin mediates Sertoli cell-spermatogenic cell adhesion. Dev. Dyn. 197, 1–13.
Pointis, G., Segretain, D., 2005. Role of connexin-based gap junction channels intestis. Trends Endocrinol. Metab. 16, 300–306.
Print, C.G., Loveland, K.L., Gibson, L., Meehan, T., Stylianou, A., Wreford, N., deKretser, D., Metcalf, D., Kontgen, F., Adams, J.M., Cory, S., 1998. Apoptosisregulator bcl-w is essential for spermatogenesis but appears otherwise redun-dant. Proc. Natl. Acad. Sci. U.S.A. 95, 12424–12431.
Rato, L., Socorro, S., Cavaco, J.E., Oliveira, P.F., 2010. Tubular fluid secretion in theseminiferous epithelium: ion transporters and aquaporins in Sertoli cells.J. Membr. Biol. 236, 215–224.
Reddi, P.P., Flickinger, C.J., Herr, J.C., 1999. Round spermatid-specific transcription ofthe mouse SP-10 gene is mediated by a 294-base pair proximal promoter. Biol.Reprod. 61, 1256–1266.
Richburg, J.H., Redenbach, D.M., Boekelheide, K., 1994. Seminiferous tubule fluidsecretion is a Sertoli cell microtubule-dependent process inhibited by 2,5-hexanedione exposure. Toxicol. Appl. Pharmacol. 128, 302–309.
Russell, L.D., 1990. Histological and Histopathological Evaluation of the Testis, firsted. Cache River Press, Clearwater, FL.
Russell, L.D., Bartke, A., Goh, J.C., 1989. Postnatal development of the Sertoli cellbarrier, tubular lumen, and cytoskeleton of Sertoli and myoid cells in the rat,and their relationship to tubular fluid secretion and flow. Am. J. Anat. 184,179–189.
Russell, L.D., Griswold, M.D., 1993. The Sertoli Cell, first ed. Cache River Press,Clearwater, FL.
Sameshima, T., Nabeshima, K., Toole, B.P., Yokogami, K., Okada, Y., Goya, T., Koono,M., Wakisaka, S., 2000. Glioma cell extracellular matrix metalloproteinaseinducer (EMMPRIN) (CD147) stimulates production of membrane-type matrixmetalloproteinases and activated gelatinase A in co-cultures with brain-derivedfibroblasts. Cancer Lett. 157, 177–184.
Schlosshauer, B., Herzog, K.H., 1990. Neurothelin: an inducible cell surface glyco-protein of blood–brain barrier-specific endothelial cells and distinct neurons.J. Cell Biol. 110, 1261–1274.
Segretain, D., Roussel, C., 1988. Endocytic origin for periaxonemal vesicles along theflagellum during mouse spermiogenesis. Gamete Res. 21, 451–463.
Seulberger, H., Unger, C.M., Risau, W., 1992. HT7, Neurothelin, Basigin, gp42 and OX-47–many names for one developmentally regulated immuno-globulin-likesurface glycoprotein on blood–brain barrier endothelium, epithelial tissuebarriers and neurons. Neurosci. Lett. 140, 93–97.
Skinner, M.K., Griswold, M.D., 2005. Sertoli Cell Biology. Elsevier Academic Press,Amsterdam; Boston.
Tanaka, H., Pereira, L.A., Nozaki, M., Tsuchida, J., Sawada, K., Mori, H., Nishimune, Y.,1997. A germ cell-specific nuclear antigen recognized by a monoclonal anti-body raised against mouse testicular germ cells. Int. J. Androl. 20, 361–366.
Tang, W., Chang, S.B., Hemler, M.E., 2004. Links between CD147 function, glycosyla-tion, and caveolin-1. Mol. Biol. Cell 15, 4043–4050.
Toyama, Y., Maekawa, M., Kadomatsu, K., Miyauchi, T., Muramatsu, T., Yuasa, S.,1999. Histological characterization of defective spermatogenesis in mice lack-ing the basigin gene. Anat. Histol. Embryol. 28, 205–213.
Tsuruta, J.K., O'Brien, D.A., 1995. Sertoli cell-spermatogenic cell interaction: theinsulin-like growth factor-II/cation-independent mannose 6-phosphate recep-tor mediates changes in spermatogenic cell gene expression in mice. Biol.Reprod. 53, 1454–1464.
Wakayama, T., Nagata, K., Ohashi, K., Mizuno, K., Tanii, I., Yoshinaga, K., Oh-Oka, T.,Toshimori, K., 2000. The expression and cellular localization of the spermflagellar protein MC31/CE9 in the rat testis: possible posttranscriptionalregulation during rat spermiogenesis. Arch. Histol. Cytol. 63, 33–41.
Wong, E.W., Mruk, D.D., Cheng, C.Y., 2008. Biology and regulation of ectoplasmicspecialization, an atypical adherens junction type, in the testis. BBA 1778,692–708.
Wong, E.W., Mruk, D.D., Lee, W.M., Cheng, C.Y., 2010. Regulation of blood-testisbarrier dynamics by TGF-beta3 is a Cdc42-dependent protein trafficking event.Proc. Nat. Acad. Sci. U.S.A. 107, 11399–11404.
















![KD-A645 / KD-R640 / KD-R540 / KD-R440 - Car Audio ...santafeautosound.com/uploads/product-manuals/JVC KD-R540.pdfKD-A645 / KD-R640 / KD-R540 / KD-R440 GET0829-001A [J/JW] ENGLISH ESPAÑOL](https://img.pdfslide.us/doc/110x75/5aaf5da87f8b9a25088d67c4/kd-a645-kd-r640-kd-r540-kd-r440-car-audio-kd-r540pdfkd-a645-kd-r640.jpg)












