Embed Size (px)
Citation preview

PERSPECTIVE
Barbara McClintock’s Unsolved Chromosomal Mysteries: Parallels to Common Rearrangements and Karyotype Evolution
James A. Birchler1 and Fangpu Han2 1Division of Biological Sciences University of Missouri Columbia, MO 65211 2State Key Laboratory of Plant Cell and Chromosome Engineering, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, 100101, China
Corresponding author: James A. Birchler [email protected] Phone: 573-882-4905 FAX: 573-882-0123
Short title: McClintock’s Chromosomal Mysteries
Key words: centromeres, telomeres, centromere misdivision, de novo centromere, X component, deficiency recovery, tiny fragment, BFB cycle, double strand break, centromere inactivation, B chromosome, chromosome fusion, nested chromosome insertion, chromosome evolution
ABSTRACT Two obscure studies on chromosomal behavior by Barbara McClintock are revisited in light of subsequent studies and evolutionary genomics of chromosome number reduction. The phenomenon of deficiency recovery in which adjacent genetic markers lost in the zygote reappear in later developmental sectors is discussed in light of de novo centromere formation on chromosomal fragments. Secondly, McClintock described a small chromosome, which she postulated carried an “X component” that fostered specific types of chromosomal rearrangements mainly involving centromere changes and attachments to the termini of chromosomes. These findings are cast in the context of subsequent studies on centromere misdivision, the tendency of broken fragments to join chromosome ends and the realization from genomic sequences that nested chromosomal insertion and end-to-end chromosomal fusions are common features of karyotype evolution. Together these results suggest a synthesis that centromere breaks, inactivation, and de novo formation together with telomeres - acting under some circumstances as double strand DNA breaks that join with others - is the underlying basis of these chromosomal phenomena.
Plant Cell Advance Publication. Published on March 15, 2018, doi:10.1105/tpc.17.00989
©2018 American Society of Plant Biologists. All Rights Reserved

2
In her Nobel Prize lecture, Barbara McClintock credits a short stay at the University of Missouri
in the summer of 1931 as the beginning of her journey to the discovery of transposable elements (www.nobelprize.org/nobel_prizes/medicine/laureates/1983/mcclintock-lecture). As a
graduate student and subsequently at Cornell University, McClintock had documented how to
identify each of the ten chromosomes of maize. This advance opened the door to the
development of cytogenetics with the fusion of genetic data and chromosomal behavior. Harriet Creighton and McClintock had famously used this melding of fields to demonstrate the
connection between genetic recombination and cytological exchange (Creighton and
McClintock, 1931). McClintock was invited to Missouri by Lewis Stadler, who just a few years earlier had
reported that X-rays caused mutations in maize (Stadler, 1928). Stadler wanted McClintock to
cytologically examine maize materials that had been subjected to X-irradiation. McClintock documented this invitation and brief stay in a series of letters to Charlie Burnham, a colleague
and collaborator from Cornell. These letters are archived in the NIH National Library of Medicine
(https://profiles.nlm.nih.gov/ps/access/LLBBND). In a letter dated February 6, 1931, McClintock
notes that Stadler had arranged a fellowship for the summer at Missouri (at a salary of $500 for two months). She wrote that “I can work on what I please but he wants me to tackle some
deficiency stuff”.
In letters dated June 26, 1931 (Op. cit. LLBBNL) and July 30, 1931 (Op. cit. LLBBNM), after she arrived at Missouri, amongst complaining about the weather (“My arm sticks to the
table (it is so warm) that I push the pen with difficulty—excuse the scrawl.”) and colleagues
(“…the associates here, except for Stadler, have not been exciting at all. It is a one-man show on his part.”), it is revealed that Stadler wanted McClintock to study so-called “deficiency
recovery” upon which he had recently published (Stadler, 1930). She expressed an interest in
this topic but noted the difficulty of the project. McClintock published a 30-page report in the
Missouri Agricultural Research Station Bulletin (McClintock, 1931) about all of the different chromosomal aberrations discovered from X-irradiated materials during this stay but there is no
mention of deficiency recovery.
McClintock went on to Cal Tech and subsequently to Germany for a short stay with a return to Cornell as a research associate (Kass, 2003). In 1936, McClintock returned to Missouri
as a faculty member and again took up the study of the effect of X-rays on chromosomes (Kass,
2003). During this time she discovered the breakage-fusion-bridge (BFB) cycle, which revealed
that normal chromosomes are protected at their ends—the forerunner of the concept of the telomere. This BFB work emerged in part from her study of chromosomal inversions, some of

3
which can recombine within the inverted region to generate dicentric chromosomes prone to
subsequent breakage and acentric fragments that subsequently will be lost. In another treatise (48 pages) in the Missouri Agricultural Research Station Bulletin (McClintock, 1938) describing
the behavior of inversions in meiosis and the subsequent microspores, McClintock alludes to
deficiency recovery and notes that the observed persistence into the zygote of the acentric
fragments produced might be an explanation for deficiency recovery. Her mention of the phenomenon here indicates that there was no progress in understanding it during her 1931 visit.
The phenomenon of deficiency recovery faded away as a research topic among cytogeneticists.
Deficiency Recovery What is deficiency recovery? Stadler reported that X-ray induced or natural deficiencies
that remove multiple genetic markers on a chromosome arm, as revealed by adjacent dominant alleles missing in most of the plant or endosperm, could have a small sector of the plant or
endosperm in which the multiple dominant markers re-appear (Stadler, 1930) (Figure 1). It was
clear from the phenotype that the deficiencies were missing parts of chromosomes so how was
it possible that the dominant markers could re-appear in developmental sectors? This phenomenon was described before DNA was realized to be the genetic material and the nature
of genes was known. Indeed, even after all that is understood today about chromosomes, it still
seems mysterious. If we look back at the phenomenon now, can we imagine a basis for it? A potential
explanation might come from a recent examination of other material from Stadler’s work. Stadler
and Roman (Stadler and Roman, 1948) reported the discovery of a small chromosome called Duplication 3a. This chromosome arose in material in which pollen with dominant markers was
uv-irradiated and crossed to a recessive a1 sh2 tester. Mosaicism for this pigment marker (a1)
and starch content marker (sh2), which are closely linked, signaled the presence of an unstable
chromosome. The instability of this chromosome led to the speculation that it might be a ring chromosome (Stadler and Roman, 1948).
However, a recent examination of this chromosome revealed that it is not in fact a ring,
having telomeres on both ends, and also that it has no detectable canonical centromere DNA (Fu et al., 2013). ChIP-Seq studies using antibodies against the histone variant typical of active
centromeres (CENH3) indicated that the chromosome possessed a de novo centromere (Fu et
al 2013). Other studies have revealed additional cases of de novo centromeres in plants
(Nasuda et al., 2005; Topp et al., 2009; Zhang et al., 2013; Liu et al., 2015; Guo et al., 2016) indicating that their occurrence is not that rare (compared to spontaneous mutation, for

4
example). Indeed, analysis of many lines of maize found great diversity for the exact positioning
of CENH3 indicating a natural tendency for centromere inactivation (or deletion) with recovery via de novo centromere formation (Schneider et al., 2016).
Looking back at the phenomenon of deficiency recovery in light of these recent
revelations might suggest that it results from de novo centromere formation on the fragment
produced in the generation of the deficiency. If a chromosome is fractured by experimentally induced or natural causes, the remaining portion with a normal centromere will be inherited as
usual to daughter cells. One could speculate that the acentric fragment could languish through
some mitoses into one of two daughter cells, as documented by McClintock for acentric fragments produced from inversion heterozygotes, but upon formation of a de novo centromere
at some point along its length, it would then subsequently be partitioned equally into both
daughter cells in a portion of the remaining developmental lineages. The frequency of de novo centromere formation was realized only recently; previously it was thought that acentric
fragments were certainly destined to be lost. Can de novo centromere formation explain
deficiency recovery? This possibility seems reasonable given the recent documentation of de
novo centromeres on chromosomal fragments (Topp et al., 2009; Fu et al., 2013; Zhang et al., 2013; Liu et al., 2015). However, one might also postulate that the fragment chromatin becomes
attached to a chromosome end (see below) after the early cell divisions of development; but this
would need to occur before de novo telomeres were added to broken ends in the sporophyte (See Tsujimoto, 1993). With modern technologies, it might be possible to test for the presence
of de novo centromeres in the recovered sectors via ChIP-Seq with antibodies against CENH3
or to test whether the released fragments attach to nonhomologous chromosomes ends with imaging techniques, assuming the ability to recognize recovered developmental sectors
amenable to such studies. Indeed, as McClintock noted in her July 30, 1931 letter to Burnham, it
is difficult to find a recovered sector that can be analyzed cytologically. Until such time that the
phenomenon is tackled with modern technologies, it remains a mystery. McClintock’s project that was suggested by Stadler went unsolved, although she was quite productive working on
other material (Kass and Birchler, 2014), which, as McClintock noted, set her on the road to the
discovery of transposable elements.
The X component In a lecture at the University of Missouri during the 1978 Stadler Genetics Symposium
(in honor of L. J. Stadler), McClintock described results of chromosomal rearrangements produced from the BFB cycle (McClintock, 1978). It was in material undergoing the BFB cycle

5
that she discovered activated transposable elements, to which she turned her research activities
for several decades. However, in the 1978 article, she described some earlier studied BFB materials with a concentration on the types of chromosomal aberrations generated. Of particular
focus was a chromosome that produced, repeatedly, specific types of aberrations. Her genetic
analysis mapped the responsible region for this activity “close to or at the centromere of the
initially isolated fragment chromosome 9”. McClintock dubbed this site the “X component”, which she postulated could restructure the genome in precise ways. The material carrying derivatives
of this chromosome still exists but subsequently has never been studied systematically for
chromosomal types produced but it does show aberrant meiotic behavior (Maguire, 1987). Below we speculate on the basis of the specific types of aberrations generated in light of recent
studies of chromosome dynamics and evolutionary genomics.
What are the types of chromosomal aberrations observed in material with the X component chromosome? The initial chromosome produced derivatives that were missing one
arm resulting in a telocentric chromosome. Another type was the production of isochromosomes
in which there is a mirror image duplication of one arm with loss of the other. A third major type
was fusion of broken parts of this chromosome to one end or the other of the progenitor generating pseudo-isochromosomes or terminal reverse duplications. The initial chromosome
was also prone to produce ring chromosomes in which the centromere was fused to one
chromosome end or in which the two ends of the chromosome were fused. In addition to rearrangement within its own structure, the fragment chromosome could
become involved in rearrangements with other members of the complement. One type was the
fusion of the centromere of the fragment chromosome with another centromere creating a centromere-centromere translocation. The other type of rearrangement was fusion of the
fragments to the end of a chromosome arm. The site of breakage of the fragment chromosome
was at the position of the X component, i.e. at or close to the centromere, and missing the
fragment short arm. Of course, other chromosomal changes were found but as McClintock noted a pattern among them was not obvious and likely represents a background of changes. If
the fragment chromosome attached to the end of another chromosome in such a manner that
the fragment retained centromere activity, a dicentric would be produced that could potentially catalyze a chain reaction of rearrangements in the genome that might account for these less
common aberrations.
Let us first consider the changes for the fragment chromosome itself. Because the
centromere region was implicated in initiating the rearrangements, it is possible that the fragment chromosome centromere was prone to misdivision as a univalent in meiosis.

6
Centromere misdivision results from the attachment of a centromere to opposing spindles with a
resulting fission in which the two products can be capable of anaphase movement (e.g. Sears, 1952). It has been documented in several plant species but studied most prominently in maize
and wheat (e.g. Carlson, 1970; Carlson and Chou, 1981; Kaszas and Birchler, 1996; 1998;
Sears, 1952; Lukaszewski, 1993).
Studies of centromere misdivision in maize (Carlson, 1970; 1973a; 1973b; Carlson and Chou, 1981; Kaszas and Birchler, 1996; 1998; Kaszas et al., 2002; Phelps-Durr and Birchler,
2004; Jin et al., 2005) reveal products such as telocentric chromosomes, isochromosomes, and
rings as depicted in Figure 2. Fracture of the centromere can result in loss of one of the two chromosome arms. Alternatively, the broken centromere can fuse upon itself after replication to
produce a mirror image isochromosome. Rings from misdivision result from the joining of the
broken centromere to the end of the same chromosome arm and are a regular product of misdivision (Carlson, 1973b; Kaszas and Birchler, 1996; 1998). An example of a centromere-
telomere fusion to form a ring chromosome is shown in Figure 3. Thus, the behavior of the X
component is similar, if not identical, to that which would occur via centromere misdivision.
What is not explained are the end to end fusions or the centromere to chromosome end fusions. The fragments that were attached to the ends of other chromosomes were typical of
products of centromere misdivision with the broken centromere attaching to a chromosomal
terminus. We postulate that there is a failure of telomere capping that occurs, which could join the DNA double strand break from centromere misdivision to the exposed chromosomal end.
End to end fusion between different chromosomes would also result from such failure. The
nature of telomere fusion is unknown but some fraction of telomeres in Arabidopsis are blunt ended (Kazda et al., 2012; Valuchova et al., 2017) and could serve as a substrate for non-
homologous ending joining repair if the protective proteins were disassociated from the
chromosomal terminus. Telomeres with the canonical structure involving a single strand
overhang might require the action of an exonuclease to be available for fusion. Indeed, mutational reduction of telomerase activity, which is involved with telomere
maintenance, will result in chromosomal end to end fusion (Riha et al., 2001; Riha and Shippen,
2003; Heacock et al., 2004; Kazda et al., 2012; Valuchova et al., 2017). In McClintock’s initial characterization of the BFB cycle she noted that broken chromosomes at meiosis would
continue to fuse and break during the gametophyte generation (and endosperm) but were
“healed” in the sporophyte. The healing occurs when a de novo telomere is added in the
sporophytic generation but this process does not operate in the gametophyte (Wang et al., 1992; Werner et al., 1992; Tsujimoto, 1993; Chao et al., 1996). If centromere misdivision occurs

7
in meiosis and telomere capping is compromised in the subsequent gametophyte, ring
chromosome products from misdivision would result from the fusion of the broken centromere and the exposed telomere. Fusions of fragments to chromosome ends could also occur in the
gametophyte and be delivered to the subsequent zygote without any chromosomal breakage.
What about centromere-centromere fusions? In wheat, when genotypes are produced
that foster misdivision in two chromosomes (double monosomics) simultaneously, common products from these experiments are chromosomes in which the broken centromeres from non-
homologous chromosomes join to form a translocation with a junction at the respective
centromeres (Lukaszewski, 1993; 2010; Vega et al., 1998; Zhang et al., 2001; Wang et al., 2017). An example of a hybrid centromere joining wheat and rye chromosome arms produced
by centromere misdivision is shown in Figure 4. With the production of DSB in two different
chromosomes simultaneously as a result of misdivision, the two breaks can be repaired in such a configuration in which chromosome arms from non-homologous chromosomes switch. If in
McClintock’s materials, the univalent fragment chromosome was regularly undergoing
misdivision, any other centromere misdivision would generate the conditions for the recovery of
the described chromosomes. Chromosome end amendment by broken fragments has also been found in maize under
other circumstances and thus is not unique to the X component material. Han and colleagues
(Han et al., 2006) described the recovery of an inactive centromere of the supernumerary B chromosome of maize translocated to the short arm of chromosome 9. The B chromosome is an
extra non-vital chromosome found in some lines of maize. Despite its dispensability, it is
maintained by an accumulation mechanism. This process involves B nondisjunction at the second pollen mitosis so that one of the two sperm has two copies and the other has none
(Roman, 1947). The sperm with the B chromosomes then preferentially fertilizes the egg as
opposed to the polar nuclei in the subsequent fertilization event (Roman, 1948). The centromere
of the B chromosome is obviously the site of nondisjunction but this process requires specific other regions of the chromosome to be present in the nucleus, most notably the distal tip of the
long arm (Ward, 1973). The chromosome 9 with the inactive B centromere does not undergo
nondisjunction because the rest of the B chromosome is missing. However, when B chromosomes are added to the genotype and thus supply the trans-
acting factors for nondisjunction, the inactive B centromere containing chromosome 9 is induced
to attempt nondisjunction (Han et al., 2007). Some nondisjoined chromosomes 9 were in fact
recovered in the next generation but, more frequently, the attempted nondisjunction caused chromosomal breakage of the short arm of chromosome 9 (Han et al., 2007). Released

8
chromosomal fragments were found appended to the short arms of chromosomes 2, 7, and 8.
The process of how these chromosomes were produced is shown in Figure 5 \. The chromosome 7 translocation was recovered and is homozygous viable. Thus, it must not be
missing any vital genes arguing for an attachment at the end or very close to the end of 7S
(Figure 6). Whether there is any resection of the chromosome ends of the recipient
chromosome in these cases is not known. Also, McClintock (McClintock, 1941) in initial studies of the BFB cycle noted a rare case of an otherwise intact chromosome 9 with extra chromatin
attached at the terminus of the short arm beyond the terminal knob heterochromatin. These
products of chromosome rupture with amendment elsewhere in the genome are similar to those from the X component fragment chromosome described by McClintock (McClintock, 1978) but
result from an independent process.
Furthermore, genome sequences of related plant species coupled with comparative evolutionary studies have revealed that some of the most common rearrangements involve
chromosome end to end fusion and the insertion of one chromosome into another at or near the
centromere (Lysak et al., 2006; Luo et al., 2009; Murat et al., 2010; Schubert and Lysak, 2011;
Wang and Bennetzen, 2012; Wang et al., 2014; Hoang and Schubert, 2017; Luo et al., 2017) (Figure 7). If telomere capping is compromised in the gametophyte generations as suggested by
the nature of the BFB cycle described above (McClintock, 1941), end to end fusions together
with inactivation of one centromere could be transmitted to the subsequent generation. If a centromere fracture occurred in meiosis for one chromosome, another chromosome with
compromised telomeres could insert into this single chromosomal break. If the centromere split
results from misdivision, the insertion would have to occur before separation of the breakage products but they might also occur via other circumstances of centromere fracture. Indeed,
centromeres appear to be relatively susceptible to DNA damage due to spindle defects that
foster centromere fission (Guerrero et al., 2010; Schneider et al., 2016). These evolutionary
events would require zero double strand breaks (end to end fusions) or only one (centromere break) and therefore have a higher probability of occurrence than other types of chromosomal
aberrations that require two simultaneous DSB (deletions, duplications, inversions and
translocations). While some cases of chromosome capture appear to be near to as opposed to within centromeres, the exact position of the active centromere shifts across a narrow range in
different lines due to centromere deletion/inactivation and de novo formation over relatively
short evolutionary timeframes (Schneider et al., 2016; Zhao et al., 2017), potentially making
such determinations only an approximation. Subsequent inversions in centromere regions (Lamb et al., 2007) also contribute to an ambiguity to the precise prediction of the site of

9
chromosome capture within the vicinity of the centromere (Schneider et al., 2016). These
chromosomal events over evolutionary time appear related to those documented experimentally by McClintock (McClintock, 1978) and others (Carlson, 1970; 1973a; 1973b; Kaszas and
Birchler, 1996; 1998; Phelps-Durr and Birchler, 2004; Han et al., 2006; 2007; Gao et al., 2011).
One potential explanation for these chromosomal changes is that some type of
recombination ties together nonhomologous chromosomes at their ends. This scenario would require identical inverted tandem repeat sequences at the ends of all chromosomes in order to
align sequences in the proper register for crossing over to occur. We are not aware that such
structures, which would have to permit recombination, are universally present near the ends of plant chromosomes. Moreover, the parallels to experimentally induced rearrangements
involving chromosomal ends that do not involve recombination would suggest otherwise.
Further, while it is possible that end to end fusions and nested chromosomal insertions might be common because other types of aberrations are selected against to some degree, the
prevalence of related structures in unselected materials from experimental work and the lower
order kinetics for their formation argues against such a scenario. Indeed, genome sequencing of
Aegilops tauschii revealed the remnants of telomere sequences at the junction of an evolutionary end to end chromosomal fusion (Luo et al., 2017).
McClintock noted that heterochromatic knobs could be preferred sites for chromosomal
rearrangements in addition to centromeres and end to end fusions. In this regard, Rhoades and colleagues (Rhoades et al., 1967; Rhoades and Dempsey, 1972) described a genotype (called
Hi Loss) that fostered chromosomal breaks at knob sites at the second pollen mitosis. Knobs
are visible landmarks on maize chromosomes that are composed of thousands of copies of a unit repeat (Peacock et al., 1981). They are typically highly condensed chromosomal sites that
are late replicating in the cell cycle (Pryor et al., 1980). In the Hi Loss background, the presence
of B chromosomes would induce knobbed chromosome arms to break at the second pollen
mitosis in the gametophyte generation. Although never examined experimentally to our knowledge, such knob breakage in the presence of a misdivided centromere or other broken
fragments is likely to produce fusions at knob sites. The relationship of the B chromosome
nondisjunction and knob breakage at the same specific mitosis was noted (Pryor et al., 1980) with the suggestion that both result from delayed replication of the respective structures beyond
the timeframe of the initiation of anaphase. Such delayed replication for the B centromere would
result in nondisjunction but chromosomal breakage for the interstitial knobs. Because the B
chromosome can occasionally nondisjoin in the endosperm and other tissues (Carlson, 1973b), it is possible that knob breakage might also occur occasionally under other circumstances.

10
In conclusion, we speculate that there is a convergence of observations from
experimental studies of chromosomal breakage about the preponderance of broken centromeres and fusions to the ends of chromosomes that explain several common karyotypic
changes revealed by comparative genomics. Chromosomal breakage initiated in meiosis, such
as centromere misdivision, would occur immediately prior to the gametophyte generation that
produces the gametes. If telomere capping of the fractured chromosome fails to protect the break from DNA repair pathways, as suggested by the behavior of the BFB cycle, the most
common chromosomal changes noted by McClintock would be formed and directly introduced
into the next generation. As opposed to chromosomal changes in somatic cells, this progression of events will lead to heritable genomic re-arrangements.
Here, we have re-examined problems studied by Barbara McClintock that have all but
been forgotten in the background of her many other brilliant contributions. Many aspects of these topics deserve further experimental investigation such as determining the frequency and
developmental timing of the apparent occasional failure of telomere capping. It is often a useful
exercise to place recent discoveries such as centromere epigenetics and comparative genomics
in the context of the historical literature, in which hidden mysteries are clarified and a greater synthesis can potentially emerge.
Acknowledgements Research on this topic is supported by National Science Foundation Plant Genome grant NSF
IOS-1444514 (JB) and by the National Natural Science Foundation of China (31630049 and
31320103192)(FH). We thank Lee Kass and Dorothy Shippen for discussion.
REFERENCES
Carlson, W. (1970). Nondisjunction and isochromosome formation in the B chromosome of maize. Chromosoma 30: 356-365.
Carlson, W. (1973a). A procedure for localizing genetic factors controlling mitotic nondisjunction in the B chromosome of maize. Chromosoma 42: 127-136.
Carlson, W. (1973b). Instability of the maize B chromosome. Theoretical and Applied Genetics 43: 147-150.
Carlson, W.R., and Chou, T.S. (1981). B chromosome nondisjunction in corn: Control by factors near the centromere. Genetics 97: 379-89.
Chao, S., Gardiner, J.M., Melia-Hancock, S., and Coe, E.H., Jr. (1996). Physical and genetic mapping of chromosome 9S in maize using mutations with terminal deficiencies. Genetics 143: 1785-1794.
Creighton, H.B., and McClintock, B. (1931). A correlation of cytological and genetical crossing-over in Zea mays. Proc. Natl. Acad. Sci. USA 17: 492-497.

11
Fu, S., Lv, Z., Gao, Z., Wu, H., Pang, J., Zhang, B., Dong, Q., Guo, X., Wang, X., and Birchler, J.A. (2013). De novo centromere formation on a chromosome fragment in maize. Proc. Natl. Acad. Sci. USA 110: 6033-6036.
Gao, Z., Fu, S., Dong, Q., Han, F., and Birchler, J.A. (2011). Inactivation of a centromere during the formation of a translocation in maize. Chromosome Res. 19: 755-761.
Guerrerro, A.A., Gamero, M.C., Trachana, V., Futterer, A., Pacios-Bras, C., Diaz-Concha, N.P., Cigudosa, J.C., Martinez-A., C., and van Wely, K.H.M. (2010). Centromere-localized breaks indicate the generation of DNA damage by the mitotic spindle. Proc. Natl. Acad. Sci. USA 107: 4159-4164.
Guo, X., Su, H., Shi, Q., Fu, S., Wang, J., Hu, Z., and Han, F. (2016). De novo centromere formation and centromere sequence expansion in wheat and its wide hybrids. PLoS Genetics 12: e1005997.
Han, F., Lamb, J.C., and Birchler, J.A. (2006). High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 103: 524-533.
Han, F., Lamb, J.C., Yu, W., Gao, Z. and Birchler, J.A. (2007). Centromere function and nondisjunction are independent components of the maize B chromosome accumulation mechanism. Plant Cell 19: 524-33.
Han, F., Gao, Z., and Birchler, J.A. (2009). Reactivation of an inactive centromere reveals epigenetic and structural components for centromere specification in maize. Plant Cell 21: 1929-1939.
Heacock, M., Spangler, E., Riha, K., Puizina, J., and Shippen, D. E. (2004). Molecular analysis of telomere fusions in Arabidopsis: multiple pathways for chromosome end-joining. The EMBO Journal 23: 2304-2313.
Hoang, P.T.N. and Schubert, I. (2017). Reconstruction of chromosome rearrangements between the two most ancestral duckweed species Spirodela polyrhiza and S. intermedia. Chromosoma 126: 929-739.
Jin, W., Lamb, J.C., Vega, J.M., Dawe, R.K., Birchler, J.A., and Jiang, J. (2005). Molecular and functional dissection of the maize B chromosome centromere. Plant Cell 17: 1412-23.
Kass, L.B. (2003). Records and recollections: A new look at Barbara McClintock, Nobel-Prize-Winning Geneticist. Genetics 164: 1251-1260.
Kass, L.B. and Birchler, J.A. (2014). Setting up a prepared mind: Summary and analysis of "Cytological observations of deficiencies involving known genes, translocations and an inversion in Zea mays. IN: Perspectives on Nobel Laureate Barbara McClintock's Publications (1926-1984): A companion volume. The Internet-First University Press. URL: http://hdl.handle.net/1813/34897.
Kaszás, E., and Birchler, J.A. (1996). Misdivision analysis of centromere structure in maize. EMBO J 15: 5246-55.
Kaszás, E., and Birchler, J.A. (1998). Meiotic transmission rates correlate with physical features of rearranged centromeres in maize. Genetics 150: 1683-92.
Kaszás, E., Kato, A., and Birchler, J.A. (2002). Cytological and molecular analysis of centromere misdivision in maize. Genome 45: 759-68.
Kazda, A., Zellinger, B., Rossler, M., Derboen, E., Kusenda, B., and Riha, K. (2012). Chromosome end protection by blunt-ended telomeres. Genes and Development 26: 1703-1713.
Lamb, J.C., Meyer, J.M., and Birchler, J.A. (2007). A hemicentric inversion in the maize line knobless Tama flint created two sites of centromeric elements and moved the kinetochore-forming region. Chromosoma 116: 237-247.
Liu, Y., Su, H., Pang, J., Gao, Z., Wang, X.J., Birchler, J.A., and Han, F. (2015). Sequential de novo centromere formation and inactivation on a chromosomal fragment in maize. Proc. Natl. Acad. Sci. USA 112: E1263-1271.

12
Lukaszewski, A. J. (1993). Reconstruction in wheat of complete chromosomes 1B and 1R from the 1RS.1BL translocation of ‘Kavkaz’ origin. Genome 36: 821-824.
Lukaszewski, A.J. (2010). Behavior of centromeres in univalent and centric misdivision in wheat. Cytogenetics and Genome Research 129: 97-109.
Luo, M.C., Deal, K.R., Akhunov, E.D., Akhunova, A. R., Anderson, O.D., Anderson, J. A., Blake, N., Clegg, M.T., Coleman-Derr, D., Conley, E.J., Crossman, C.C., Dubcovsky, J., Gill, B. S., Gu, Y.Q., Hadam, J., Heo, H.Y., Huo, N., Lazo, G., Ma, Y., Matthews, D.E., McGuire, P.E., Morrell,P. L., Qualset, C.O., Renfro, J., Tabanao, D., Talbert, L.E., Tian, C., Toleno, D.M., Warburton,M. L., You, F.M., Zhang, W. and Dvorak, J. (2009). Genome comparisons reveal a dominantmechanisms of chromosome number reduction in grasses and accelerated genome evolutionin Triticeae. Proc. Natl. Acad. Sci. USA 106: 15780-15785.
Luo, M.C., Gu, Y.Q., Puiu, D., Wang, H., Twardziok, S.O. et al. (2017). Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature, doi 10.1038/nature24486.
Lysak, M.A., Berr, A., Pecinka, A., Schmidt, R., McBreen, K., and Schubert, I. (2006). Mechanisms of chromosome number reduction in Arabidopsis and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 103: 5224-5229.
Maguire, M.P. (1987). Meiotic behavior of a tiny fragment chromosome that carries a transposed centromere. Genome 29: 744-747.
McClintock, B. (1931). Cytological observations of deficiencies involving known genes, translocations and an inversion in Zea mays. Missouri Agricultural Experiment Station Research Bulletin 163: 1-30.
McClintock, B. (1938). The fusion of broken ends of sister half-chromatids following chromatid breakage at meiotic anaphases. Missouri Agricultural Experiment Station Research Bulletin 290: 1-48.
McClintock, B. (1941). The stability of broken ends of chromosomes in Zea mays. Genetics 26: 234-282.
McClintock, B. (1978). Mechanisms that rapidly reorganize the genome. Stadler Symposium 10: 25-47.
Murat, F., Xu, J. H., Tannier, E., Abrouk, M., Guilhot, N., Pont, C., Messing, J. and Salse, J. (2010). Ancestral grass karyotype reconstruction unravels new mechanisms of genome shuffling as a source of plant evolution. Genome Research 20: 1545-1557.
Nasuda, S., Hudakova, S., Schubert, I., Houben, A., and Endo, T.R. (2005). Stable barley chromosomes without centromeric repeats. Proc. Natl. Acad. Sci. USA 102: 9842-9847.
Peacock, W.J., Dennis, E.S., Rhoades, M.M. and Pryor, A.J. (1981). Highly repeated DNA sequence limited to knob heterochromatin in maize. Proc. Natl. Acad. Sci. USA 78: 4490-4494.
Phelps-Durr, T.L., and Birchler, J.A. (2004). An asymptotic determination of minimum centromere size for the maize B chromosome. Cytogenet. Genome Res. 106: 309-13.
Pryor, A., Faulkner, K., Rhoades, M.M. and Peacock, W.J. (1980). Asynchronous replication of heterochromatin in maize. Proc. Natl. Acad. Sci. USA 77: 6705-6709.
Rhoades, M.M. and Dempsey, E. (1972). On the mechanism of chromatin loss induced by the B chromosome of maize. Genetics 71: 73-96.
Rhoades, M.M., Dempsey, E., and Ghidoni, E. (1967). Chromosome elimination in maize induced by supernumerary B chromosomes. Proc. Natl. Acad. Sci. USA 57: 1626-1632.
Riha, K., McKnight, T.D., Griffing, L.R. and Shippen, D.E. (2001). Living with genome instability: Plant responses to telomere dysfunction. Science 291: 1797-1800.
Riha, K. and Shippen, D. E. (2003). Ku is required for telomeric C-rich strand maintenance but not for end-to-end chromosome fusions in Arabidopsis. Proc. Natl. Acad. Sci. USA 100: 611-615.
Roman, H. (1947). Mitotic nondisjunction in the case of interchanges involving the B-type chromosome in maize. Genetics 32: 391-409.
Roman, H. (1948). Directed fertilization in maize. Proc. Natl. Acad. Sci. USA 34: 36-42.

13
Schneider, K.L., Xie, Z.D., Wolfgruber, T.K. and Presting, G.G. (2016). Inbreeding drives maize centromere evolution. Proc. Natl. Acad. Sci. USA 113: E987-996.
Schubert, I., and Lysak, M.A. (2011). Interpretation of karyotype evolution should consider chromosome structural constraints. Trends in Genetics 27: 207-216.
Sears, E.R. (1952). Misdivision of univalents in common wheat. Chromosoma 4: 535-550. Stadler, L.J. (1928). Genetic effects of X-rays in maize. Proc. Natl. Acad. Sci. USA 14: 69-75. Stadler, L.J. (1930). Recovery following genetic deficiency in maize. Proc. Natl. Acad. Sci. USA
16: 714-720. Stadler, L.J., and Roman, H. (1948). The effect of X-rays upon mutation of the gene A in maize.
Genetics 33: 273–303. Topp, C.N., Okagaki, R.J., Melo, J.R., Kynast, R.G., Phillips, R.L., and Dawe, R.K. (2009).
Identification of a maize neocentromere in an oat-maize addition line. Cytogenetic and Genome Research 124: 228-238.
Tsujimoto, H. (1993). Molecular cytological evidence for gradual telomere synthesis at the broken chromosome ends in wheat. J. Plant Research 106: 239-244.
Valuchova, S., Fulnecek, J., Prokop, Z., Stolt-Bergner, P., Janouskova, E., Hofr, C., and Riha, K. (2017). Protection of Arabidopsis blunt-ended telomeres is mediated by a physical association with the Ku heterodimer. The Plant Cell 29: 1533-1545.
Vega, J.M., and Feldman, M. (1998). Effect of the pairing gene Ph1 on centromere misdivision in common wheat. Genetics 148: 1285-1294.
Wang, H., and Bennetzen, J.L. (2012). Centromere retention and loss during the descent of maize from a tetraploid ancestor. Proc. Natl. Acad. Sci. USA 109: 21004-9.
Wang, X., Jin, D., Wang, Z., Guo, H., Zhang, L., Wang, L., Li, J., and Paterson, A. H. (2014). Telomere-centric genome repatterning determines recurring chromosome number reductions during the evolution of eukaryotes. New Phytologist 205: 378-389.
Wang, J., Liu, Y., Su, H., Guo, X., and Han, F. (2017). Centromere structure and function analysis in wheat-rye translocation lines. Plant Journal 91: 199-207.
Wang, S., Lapitan, N.L.V., Roder, M., and Tsuchiya, T. (1992). Characterization of telomeres in Hordeum vulgare chromosomes by in situ hybridization. II. Healed broken chromosomes in telotrisomic 4L and acrotrisomic 4L4S lines. Genome 35: 975-980.
Ward, E.J. (1973). Nondisjunction: localization of the controlling site in the maize B chromosome. Genetics 73: 387-391.
Werner, J.E., Kota, R.S., Gill, B.S., and Endo, T.R. (1992). Distribution of telomeric repeats and their role in the healing of broken chromosome ends in wheat. Genome 35: 844-848.
Zhang, B., Lv, Z., Pang, J., Liu, Y., Guo, X., Fu, S., Li, J., Dong, Q., Wu, H.J., Gao, Z., Wang, X.J., and Han, F. (2013). Formation of a functional maize centromere after loss of centromeric sequences and gain of ectopic sequences. Plant Cell 25: 1979-89.
Zhang, P., Friebe, B., Lukaszewski, A. J., and Gill, B. S. (2001). The centromere structure in Robertsonian wheat-rye translocation chromosomes indicates that centric breakage-fusion can occur at different positions within the primary constriction. Chromosoma 110: 335-344.
Zhao, H.N., Zeng, Z.X., Koo, D.-H., Gill, B.S., Birchler, J.A., and Jiang, J.M. (2017). Recurrent establishment of de novo centromeres in the pericentromeric region of maize chromosome 3. Chromosome Research 25: 299-311.

Figure 1. Deficiency Recovery. The kernel at the right illustrates an apparent spontaneous case of deficiency recovery. The kernel at the left has a recessive genetic marker for anthocyanin production in the endosperm (c1) and is representative of the maternal parent. The center kernel has the dominant allele for the pigment gene (C1) and is representative of the male parent. In the kernel at the right, the dominant marker is missing on most of the endosperm but a small sector with light pigmentation typical of a c1/c1/C1 endosperm is present. The dominant marker must have been delivered to the zygote, but is missing in much of the endosperm and recovers only in a sector. Photo by Zhi Gao.
Figure 2. Diagrammatic representation of products of centromere misdivision. Top, left) Misdivision of the centromere occurs when the kinetochore attaches to the spindle from both poles and fractures the centromere of a univalent (Sears, 1952). The chromosome depicted is a univalent in meiosis I with sister chromatids of a chromosome with arms of different lengths. The different arms are shown in orange and blue. Top, right) The attachment of the chromosome from both poles breaks the chromosome at the centromere and the replicated chromatids of the different arms progress to opposite poles. Molecular studies indicate that centromere misdivision cleaves the underlying DNA sequences (Kaszas and Birchler, 1996; 1998; Jin et al., 2005). Bottom) Products of centromere misdivision include telocentric chromosomes derived from either arm (blue or orange) of the progenitor chromosome, isochromosomes derived from either chromosome arms but which are fused at the site of centromere breakage and ring chromosomes that join the broken centromere to the end of the same chromosome (Carlson, 1970; 1973a; 1973b; Carlson and Chou, 1981; Kaszas and Birchler, 1996; 1998; Kaszas et al., 2002; Phelps-Durr and Birchler, 2004; Jin et al., 2005).
2
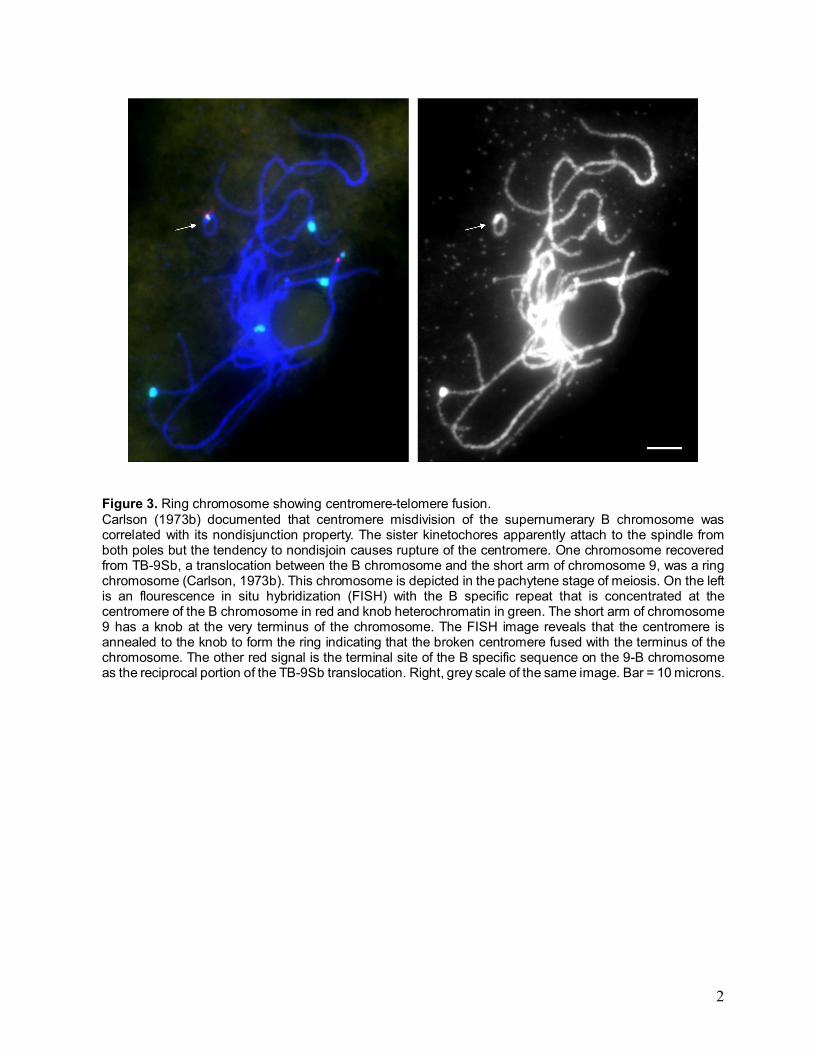
Figure 3. Ring chromosome showing centromere-telomere fusion. Carlson (1973b) documented that centromere misdivision of the supernumerary B chromosome was correlated with its nondisjunction property. The sister kinetochores apparently attach to the spindle from both poles but the tendency to nondisjoin causes rupture of the centromere. One chromosome recovered from TB-9Sb, a translocation between the B chromosome and the short arm of chromosome 9, was a ring chromosome (Carlson, 1973b). This chromosome is depicted in the pachytene stage of meiosis. On the left is an flourescence in situ hybridization (FISH) with the B specific repeat that is concentrated at the centromere of the B chromosome in red and knob heterochromatin in green. The short arm of chromosome 9 has a knob at the very terminus of the chromosome. The FISH image reveals that the centromere is annealed to the knob to form the ring indicating that the broken centromere fused with the terminus of the chromosome. The other red signal is the terminal site of the B specific sequence on the 9-B chromosome as the reciprocal portion of the TB-9Sb translocation. Right, grey scale of the same image. Bar = 10 microns.
3
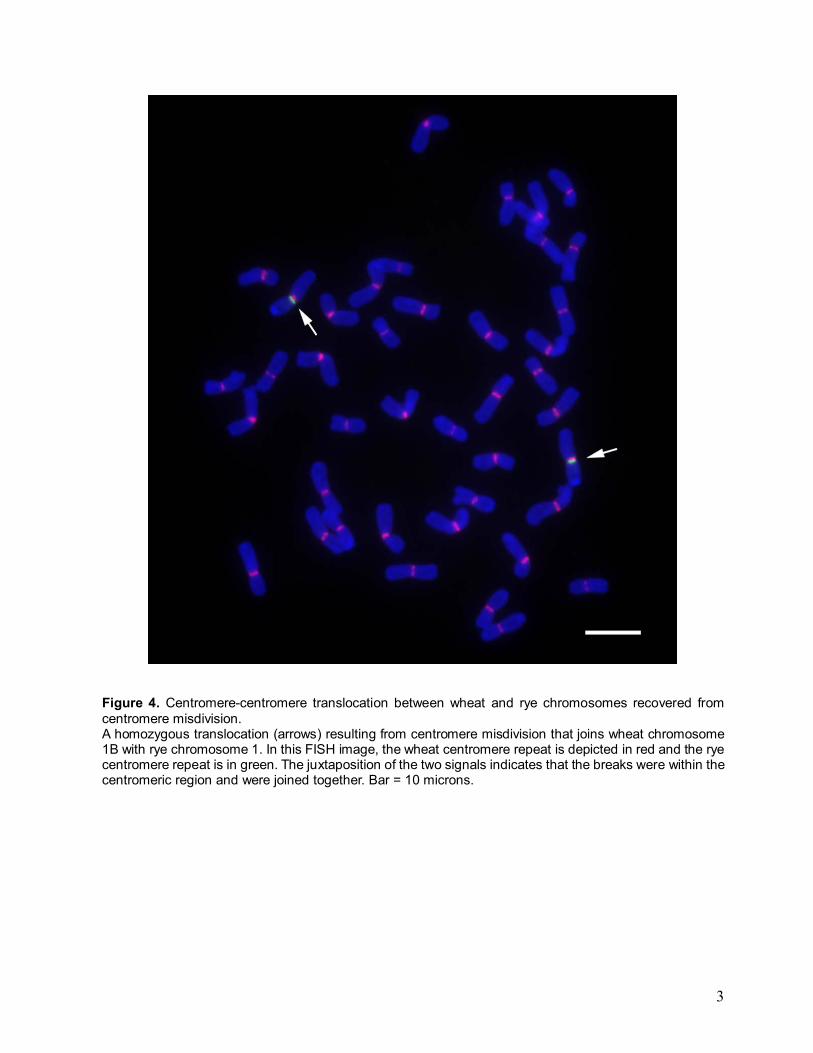
Figure 4. Centromere-centromere translocation between wheat and rye chromosomes recovered from centromere misdivision. A homozygous translocation (arrows) resulting from centromere misdivision that joins wheat chromosome 1B with rye chromosome 1. In this FISH image, the wheat centromere repeat is depicted in red and the rye centromere repeat is in green. The juxtaposition of the two signals indicates that the breaks were within the centromeric region and were joined together. Bar = 10 microns.
4
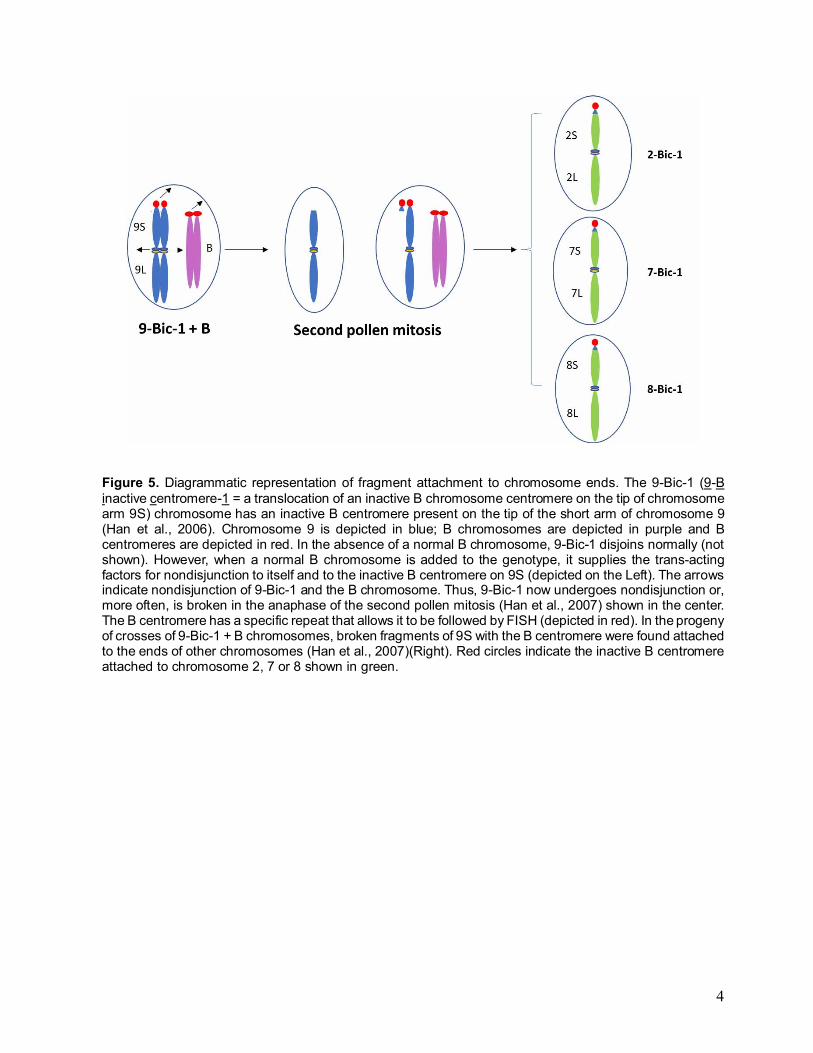
Figure 5. Diagrammatic representation of fragment attachment to chromosome ends. The 9-Bic-1 (9-B inactive centromere-1 = a translocation of an inactive B chromosome centromere on the tip of chromosome arm 9S) chromosome has an inactive B centromere present on the tip of the short arm of chromosome 9 (Han et al., 2006). Chromosome 9 is depicted in blue; B chromosomes are depicted in purple and B centromeres are depicted in red. In the absence of a normal B chromosome, 9-Bic-1 disjoins normally (not shown). However, when a normal B chromosome is added to the genotype, it supplies the trans-acting factors for nondisjunction to itself and to the inactive B centromere on 9S (depicted on the Left). The arrows indicate nondisjunction of 9-Bic-1 and the B chromosome. Thus, 9-Bic-1 now undergoes nondisjunction or, more often, is broken in the anaphase of the second pollen mitosis (Han et al., 2007) shown in the center. The B centromere has a specific repeat that allows it to be followed by FISH (depicted in red). In the progeny of crosses of 9-Bic-1 + B chromosomes, broken fragments of 9S with the B centromere were found attached to the ends of other chromosomes (Han et al., 2007)(Right). Red circles indicate the inactive B centromere attached to chromosome 2, 7 or 8 shown in green.
5
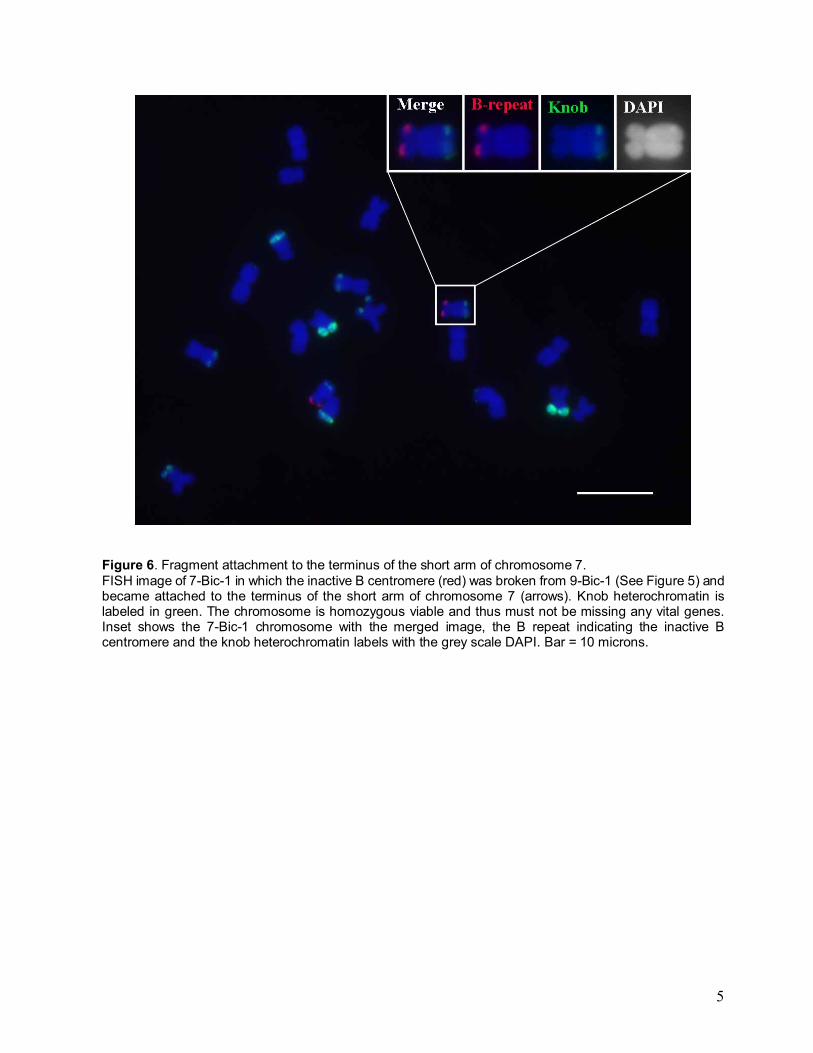
Figure 6. Fragment attachment to the terminus of the short arm of chromosome 7. FISH image of 7-Bic-1 in which the inactive B centromere (red) was broken from 9-Bic-1 (See Figure 5) and became attached to the terminus of the short arm of chromosome 7 (arrows). Knob heterochromatin is labeled in green. The chromosome is homozygous viable and thus must not be missing any vital genes. Inset shows the 7-Bic-1 chromosome with the merged image, the B repeat indicating the inactive B centromere and the knob heterochromatin labels with the grey scale DAPI. Bar = 10 microns.

6
Figure 7. Diagrammatic representation of nested chromosomal insertion and end to end fusions—common chromosome number reduction events in evolution. Two of the most common chromosomal rearrangements during karyotype evolution are end to end fusions and nested chromosomal insertions (Lysak et al., 2006; Luo et al., 2009; Murat et al., 2010; Schubert and Lysak, 2011; Wang and Bennetzen, 2012; Wang et al., 2014; Hoang and Schubert, 2017; Luo et al., 2017). At the left, two nonhomologous chromosomes are depicted in orange and blue with the centromere depicted as gray ovals. End to end fusions result from the joining of the termini of the two chromosomes, which is likely associated with deletion or inactivation of one of the two centromeres such that the result is not functionally dicentric, which would lead to its fracture. Experimentally, centromere inactivation can occur within the span of a cell cycle, or at least a few, and then the inactive state is perpetuated over generations (Han et al., 2009; Gao et al., 2011). Nested chromosomal insertion occurs when one chromosome inserts at or near the centromere of another chromosome. Again, only one centromere would retain function for the integrity of the new chromosome to be maintained.

DOI 10.1105/tpc.17.00989; originally published online March 15, 2018;Plant CellJames A. Birchler and Fangpu Han
karyotype evolutionBarbara McClintock's Unsolved Chromosomal Mysteries: Parallels to common rearrangements and
This information is current as of August 9, 2020
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























