Embed Size (px)
Citation preview

BAIT FORMULATIONS OF CHEMICAL AND MICROBIAL INSECTICIDES FOR SUPPRESSION
OF CROP-FEEDING GRASSHOPPERS','
J. L. Capinera' and B. E. Hibbard Depal'tment of Entomology Colorado State University
Ft. Collins. Colorado 80523
Abstract: Chemical and microbial insecticides, applied principally in bait formulations. were evaluated for control of crop-feeding grasshoppers, primarily Me[cUlop[US spp .• under field pcn conditions. An aqueous Cannulation of carbaryl was slightly more effective than carbaryltreated wheat bran and rolled wheat baits containing the equivalent amount of insecticide per unit area. Wheat bran was effective hait for grasshoppers in the presence of high and low lo\'els of competing vegetation. Nosema locuslae on wheat bran provided relatively rapid sUI>pression (significant reduction within 1 wk) when applied at. n high rate. Steinememn {e/liae incorporated in calcium nlginnl.e capsules and alfalfa-wheat. pellets provided suppression within 2 days of t.reatment. While not. as effective as aqueous insecticide sprays, baits may be suitable where lower levels of controt are acceptable, or where multiple appJical.ions lIl'e
possible.
Key Words: Melanoplus grasshoppers, baits, Nosema locustae, Steinemema (eltiae, carbaryl.
J. Agric. Entomol 4(·1): 337-344 (October 1987)
Grasshopper management in western states focuses primarily on rangeland species. Tillage, irrigation, and frequent insecticide applications minimize the effects of grasshoppers in most crop agroecosystems. However, relatively undistw'bed areas such as roadsides and irrigation ditches often support very high densities of grasshoppers that may move to crop plants. Occasionally a rangeland species le.g. Melarwplus sanguinipes (F.)I will also infest cropland. Grasshoppers are primarily a rangeland problem, and most recent research has focllsed on management strategies and tactics suitable for this resource; applicability of these management approaches to cropland is questionable. [n addition, recent grasshopper outbreaks have affected suburban gardens and omamental plants, commodities with entirely different monetary values and environmental constraints. Development of management techniques for these latter areas has been almost completely neglected.
Before the advent of aircraft and ultra low volume insecticide application technology, insecticide-treated baits were widely applied for grasshopper control on crop and rangeland (Parker 1939; Shotwell 1942). Although sometimes deemed impractical for use on a large scale, baits have been reported to provide desired population reduction at significantly lower doses of insecticide than required by aqueous formulations. Also, non-target species are less likely to be disrupted (Mukerji et al. 1981). Biological control agents such as Nosema locustae Canning must be consumed to infect grasshoppers, so bait applications are preferred (Henry 1981; Ewell and Mukerji 1980). Because bait formulations seem to offer
I ORTHOPTERA: Acrididae. 2 Accepted for publication 28 October 1987. 3 Current address: Department of Entomology and Kematology. University of Florida, Gainesville, FL
32611.
337

338 J. Agric. EntomoJ. Vol. 4, No. 4 (1987)
advantages in certain cropland and surbw'ban situations, we investigated their comparative advantages, emphasizing use of pathogens.
MATERIALS AND METHODS
Grasshoppers were collected with a sweep net in the vicinity of Fort Collins, Colorado. Collection sites were road-sides, irrigation ditches, and waste areas. The species complex consisted principally of Melanoplus biuitlalus (Say) and M. sanguinipes (F.) (Tlial 1) or of M femunubrum (DeGeerJ. M. differentia/is (ThomasJ. and Phoelaliotes nebrascensis (Thomas) (Trial 2 and 3). Although the nymphs were not identified to species, they were randomly distributed across treatments, so that each species was represented in approximately equal numbers. All tests used grasshopper instars 3 and 4 except the Nosema study, which was conducted with instars 1 and 2.
Grasshoppers were released into pens fol' testing. Pens consist.ed of galvanized metal, 70 cm high, enclosing a 2 by 2 m Brea of soil from which vegetation had been removed. Each pen was seeded with 75 grasshoppers at the initiation of the study, before application of liquid and bait treatments. Food was provided before application of treatments by inserting 2 flats (30 by 50 em) of rye seedlings and one flat of lett.uce seedlings except in the vegetation studies. Pens were censused daily for 3 d after treatment except in the Nosema study, where grasshopper numbers were determined for 3 consecutive weeks. Control populations were established for each study and date, and were provided only with rye and lettuce seedlings for food. Pens had no measurable effect on temperature or humidity, except for a small area immediately adjacent to the metal.
Each treatment was conducted in fow' separate pens pel' date, and replicat.ed over time. There were three trials: June-July 1985, July-August 1985, and JulyAugust 1986. The trials differed in that a band of insect adhesive (tanglefoot) was added along the inside top of the metal pen after the first trial, and that the pens were covered with bird netting for the third trial. The tanglefoot was added as a precaution because rain splash contaminated the metal y,rith soil, making it easier for grasshoppers to crawl upwards and escape. The bird netting was not used in 1985 because grasshoppers were numerous at the research site, and therefore the penned grasshoppers were probably not especially attractive to avian predators. However, in 1986 grasshopper density at the site was low, and the grasshoppers might have been attractive to birds. All bait treatments were applied during the early evening hours (7 to 8 p.m.) to minimize interference by high temperatures and sunlight.
Data were analyzed using 2-way analysis of variance; sources of variance were treatment and replication (trial). A separate analysis was conducted for each observation period after treatment. With the exception of the Nosema evaluation, significant interactions occurred, so data for each replicate (trial) are presented. Means were separated with Student·Newman-Keuls (SNI<) multiple range test at E = 0.05 (Zar 1984). Percent corrected control was calculated using Abbott's (1925) fOlmula.
Comparison of carbaryl liquid and bait applications A liquid formulation of carbaryl (Sevin-4-oil) was compared with two commonly
used bait formulations (wheat bran or rolled wheat); each was treated with

339 CAPINERA and HIBBARD: Grasshopper Buits
carbaryl. The liquid formulation was diluted with diesel fuel and applied at a rate of 0.305 kg Altha (AD, in a total volume of 62.5 ltha. Application was made with a compressed air sprayer equipl}ed with a teejet hollow cone nozzle at 2.1 kg/cm2
pressure. The wheat bran was sprayed with undiluted Sevin-4-oil with an air brush to provide 2% AI, and the bait was applied by hand at a rate of 15 kg/ha, resulting in the same rate of insecticide application as the liquid fOimulation. Rolled wheat was treated with carbaryl at 1% .1\1 and applied at 30 kg/ha. Because rolled wheat is twice as heavy as bran, equal volumes were applied, resulting in equivalent amounts of insecticide on an area basis.
Effects of vegetation density on bait efficacy Effects of vegetation density on carbaryl bait efficacy were determined by
establishing high (four flats of rye pel· pen) and low (one nat of rye per pen) levels of vegetation. 'Wheat bran bait \\'as treated with carbaryl as in the aforementioned carbaryl study and applied at the same rates.
Evaluation of protozoan bait Nosema locustae spores were obtained from the 'SDA Rangeland Insect
Laboratory, Bozeman, NIT (1985) and Evans Biocontrol, Durango, CO (1986). Spores were suspended in water and applied by airbrush to wheat bran at a rate of 5 X 109 spores per kg bran. Bran was applied at 15 kg/ha as in the insect.icidetreated plot.s, providing 7.5 X 1010 sporeslha.
Evaluation of entomophagou8 nematode bait Steinemema feLtiae Filipjev (= Neoaplectana carposapsae Weiser) nematodes
(Mexican strain) were cultured in Galleria mellonella (L.) larvae and incorporated in two bait formulations. One bait consisted of 60 mg calcium alginate capsules (I<aya and Nelsen 1985) prepared by Plant Genetics, Inc., Davis, CA. Each capsule contained ca. 400 infective juvenile nematodes and wheat bran (5% by wt); they were applied at a rate of 150 kglha.
The second bait was pellets consisting of a mixture (by wt) of alfalfa meal (23%), wheat flour (11%), wheat bran (11%), corn oil (9%), and water (45.5%). Locust bean gum (0.5%) and nematodes were added to the mixture and extruded through a meat grinder. The pellets produced by brcaking up the extruded material weighted ca. 60-100 mg and contained 6.7 nematodes per mg, equivalent to the calcium alginate capsules. The pellets also were al}plied at 150 kg/ha. Previously (unpublished), we determined that this pellet mixture was acceptable to grasshoppers in laboratory tcsts.
RESULTS AND DISCUSSION
All carbaryl treatments significantly reduced grasshopper populations (Table 1), although the liquid application was most efficacious. Wheat bran bait generally appeared to be more effective than rolled wheat bait, although statistically significant differences were infrequent. Average mortality (% corrected control) was 83.8, 87.1, 90.1% for liquid application, 54.4, 69.3, 78.1% for wheat bran, and 42.4, 60.1, 64.3% for rolled wheat, respectively, on days 1 to 3 after treatment. Henry and Oma (1981) compared effectiveness of wheat bran, rolled barley, and rolled oats for inoculation of Melanoplus spp_ with Nosema locustae; wheat bran generally was superior in these studies, also.
340 J. Agric. Entomol. Vol. 4, No. 4 (1987)
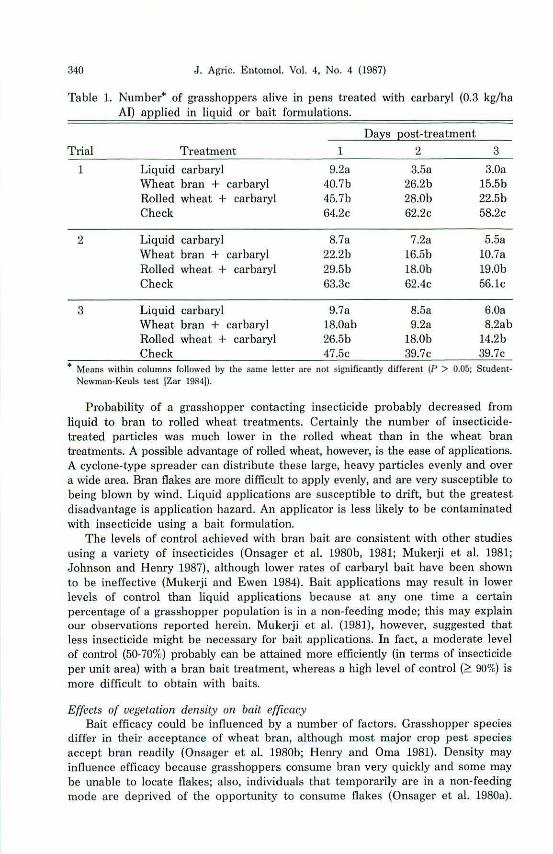
Table 1. Number· of grasshoppers alive in pens treated with carbaryl (0.3 kg/ha AI) applied in liquid 01' bait formulations.
Trial Treatment
1 Liquid carbaryl \Vheat bran + carbaryl Rolled wheat + carbaryl Check
2 Liquid carbaryl Wheat bran + carbaryl Rolled wheat + carbaryl Check
1
9.2a 40.7b 45.7b 64.2c
8.7a 22.2b 29.5b 63.3c
Days post-treatment
2 3
3.5a 3.0a 26.2b 15.5b 28.0b 22.5b 62.2c 58.2c
7.28 5.5a 16.5b 1O.7a 18.0b 19.0b 62.4c 56.ic
3 Liquid carbaryl 9.78 8.5a 6.0a Wheat bran + carbaryl 18.0ab 9.2a 8.2ab Rolled wheat + carbaryl 26.5b 18.0b 14.2b Check 47.5c 39.7c 39.7c
• Means within columns fcllowed hy the same letter are not significantly different (P > 0.05; StudentNewman-Kellis lest IZar 1984]).
Probability of a grasshopper contacting insecticide probably decreased from liquid to bran to rolled wheat treatments. Certainly the number of insecticidetreated particles was much lower in the rolled wheat than in the wheat bran treatments. A possible advantage of rolled wheat, however, is the ease of applications. A cyclone-type spreader can distribute these large, heavy particles evenly and over a wide area. Bran flakes are more difficuJt to apply evenly, and are very susceptible to being blown by wind. Liquid applications are susceptible to drift, but the greatest disadvantage is application hazard. An applicator is less likely to be contaminated with insecticide using a bait formulation.
The levels of control achieved with bran bait are consistent with other studies using a variety of insecticides (DosageI' et al. 1980b, 1981; Mukerji et a1. 1981; Johnson and Hemy 1987), although lower rates of carbaryl bait have been shown to be ineffective (Mukel'ji and Ewen 1984). Bait applications may result in lower levels of control than liquid applications because at anyone time a certain percentage of a grasshopper population is in a non-feeding mode; this may explain our observations reported herein. Mukerji et 01. (1981), however, suggested that less insecticide might be necessary for bait applications. In fact, a moderate level of control (50-70%) probably can be attained more efficiently (in terms of insecticide per unit area) with a bran bait treatment, whereas a high level of control (~ 90%) is more difficult to obtain with baits.
Effects of uegetation density on bait efficacy Bait efficacy could be innuenced by a number of factors. Grasshopper species
differ in their acceptance of wheat bran, although most major crop pest species accept bran readily (Onsager et al. 1980b; Henry and Dma 1981). Density may influence efficacy because grasshoppers consume bran very quickly and some may be unable to locate flakes; also, individuals that temporarily are in a non-feeding mode are deprived of the opportunity to consume flakes (Onsager et al. 1980a).
341 CAPINERA and I-UBBARD: Grasshopper Baits
Increasing the rate of application of insecticide-treated bran invariably increases the level of mortality, although it may not be cost-efficient to do so (Dnsager et al. 19808; Mukel'ji el 81. 1981).
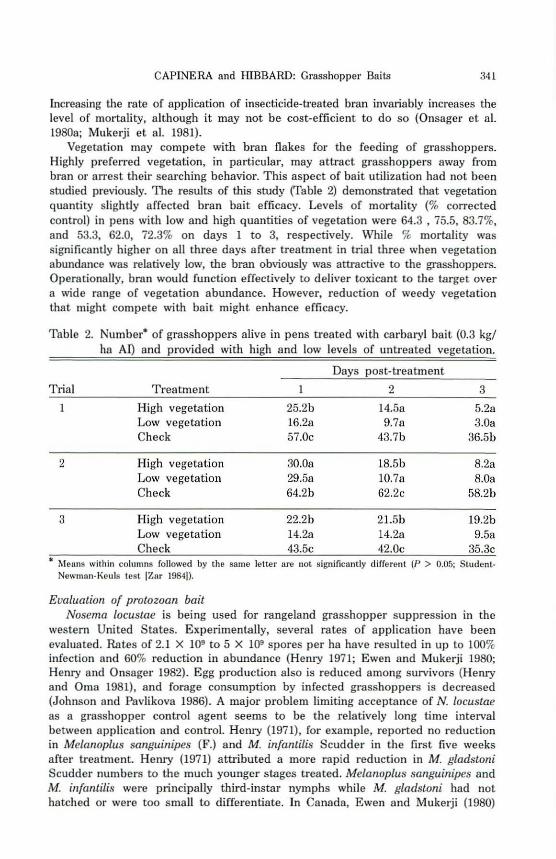
Vegetation may compete with bran flakes for the feeding of grasshoppers. Highly preferred vegetation, in particular, may attract grasshoppers away from bran or arrest their searching behavior. This aspect of bait utilization had not been studied previously. The results of this study (Table 2) demonsb'ated that vegetation quantity slightly affected bran bait efficacy. Levels of mortality (% corrected control) in pens with low and high quantities of vegetation were 64.3 , 75.5, 83.7%, and 53.3, 62.0, 72.3% on days 1 to 3, respectively. 'While % mortality was significantly higher on all three days after treatment in trial three when vegetat.ion abundance was relatively low, the bran obviously was attractive to the grasshoppers. Operationally, bran would function effectively to deliver toxicant to the target over a wide range of vegetation abundance. However, reduction of weedy vegetation that might compete with bait might enhance efficacy.
Table 2. Number- of grasshoppers alive in pens treated with carbaryl bait (0.3 kgl ha AI) and provided with high and low levels of untreated vegetation.
D8Ys post-treatment
Trial Treatment 1 2 3
High vegetation Low vegetation Check
25.2b 16.2a 57.0c
l4.58 9.78
43.7b
5.28 3.0a
36.5b
2 High vegetation 30.08 18.5b 8.28 Low vegetation 29.58 1O.7a 8.0a Check 64.2b 62.2c 58.2b
3 High vegetation 22.2b 21.5b 19.2b Low vegetation 14.2a 14.2a 9.5a Check 43.5c 42.0c 35.3c
• r.lenns within columns followed by the same leiter are not sib'llificantly different (p > 0.05; Sludent.Newman-Kellis test IZar 19841).
Evaluation of protozoan bait Nosema locustae is being used for rangeland grasshopper suppression in the
western United States. Experimentally, several rates of application have been evaluated. Rates of 2.1 X 109 to 5 X 109 spores per ha have resulted in up to 100% infection and 60% reduction in abundance (Henry 1971; Ewen and Mukerji 1980; Henry and Dnsager 1982). Egg production also is reduced among survivors (Henry and Oma 1981), and forage consumption by infected grasshoppers is decreased (Johnson and Pavlikova 1986). A major problem limiting acceptance of N. locustae as a grasshopper control agent seems to be the relatively long time interval between application and control. Henry (1971), for example, reported no reduction in Melanoplus sanguinipes (F.) and M. infantilis Scudder in the first five weeks after treatment. Henry (1971) attributed a more rapid reduction in M. gladstoni Scudder numbers to the much younger stages treated. Melanoplus sanguinipes and M. infantilis were principally third-instar nymphs while M. gladstoni had not hatched or were too small to differentiate. In Canada, Ewell and Mukel'ji (1980)
342 J. Agric. Enlomol. Vol. 4, No. 4 (I98?)
treated third-instal' nymphs and reported 20% reductions in abundance 4 wk after b·eatment. Since farmers and gardeners typically wait until damage by grasshoppers is imminent before attempting control, a four to five week interval after treatment with relatively low level of subsequent mortality is unacceptable. Therefore, we sought to ascertain whether more rapid population reduction could be obtained by increasing the pathogen dose.
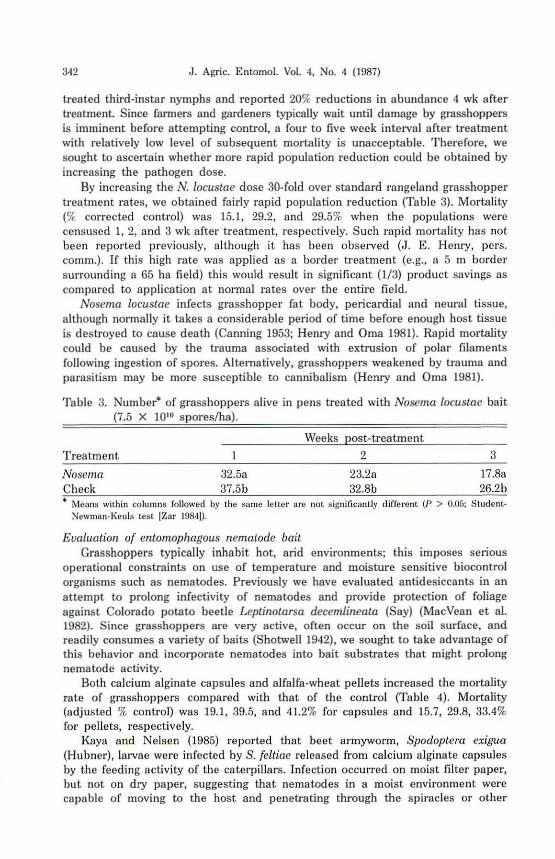
By increasing the N. locustae dose 30·fold over standard rangeland grasshopper treatment rates, we obtained fairly rapid population reduction (Table 3). Mortality (% corrected control) was 15.1, 29.2, and 29.5% when the populations were censllsed 1,2, and 3 wk after treatment, respectively. Such rapid mortality has not been reported previously, although it has been observed (J. E. Henry, pel's. comm.). If this high rate was applied as a border treatment (e.g., H 5 m border surrounding a 65 ha field) this would result in significant (1/3) product savings as compared to application at normal rates over the entire field.
Nosema locustae infects grasshopper fat body, pericardial and neural tissue, although nonnally it takes a considerable period of time before enough host tissue is destroyed [0 cause death (Canning 1953; Henry and Oma 1981). Rapid mortality could be caused by the trauma associated with extrusion of polar filaments following ingestion of spores. Altematively, grasshoppers weakened by trauma and parasitism may be more susceptible to cannibalism (Henry and Oma 1981).
Table 3. Number· of grasshoppers alive in pens treated with Nosema locustae bait (7.5 X 10" sporesth.).
Weeks post-treatment
Treatment 1 2 3
Nosema 32.5a 23.2. 17.8a Check 37.5b 32.8b 26.2b • Mellns within colullIns followed by the same leller are not sib'nilicantly difFllrent (fJ > 0.05; Student
NewlIllllI·j{euls test JZar 198·11).
Evaluation of entomophagous nematode bait Grasshoppers typically inhabit hot, arid environments; this imposes serious
operational constraints on use of temperature and moisture sensitive biocontrol organisms such as nematodes. Previously we have evaluated antidesiccants in an attempt to prolong infectivity of nematodes and provide protection of foliage against Colorado potato beetle Leplinotarsa decemlineat.a (Say) (MacVean et al. 1982)_ Since grasshoppers are very active, often occur on the soil surface, and readily consumes a variety of baits (Shotwell 1942), we sought to take advantage of this behavior and incorporate nematodes into bait substrates that might prolong nematode activity.
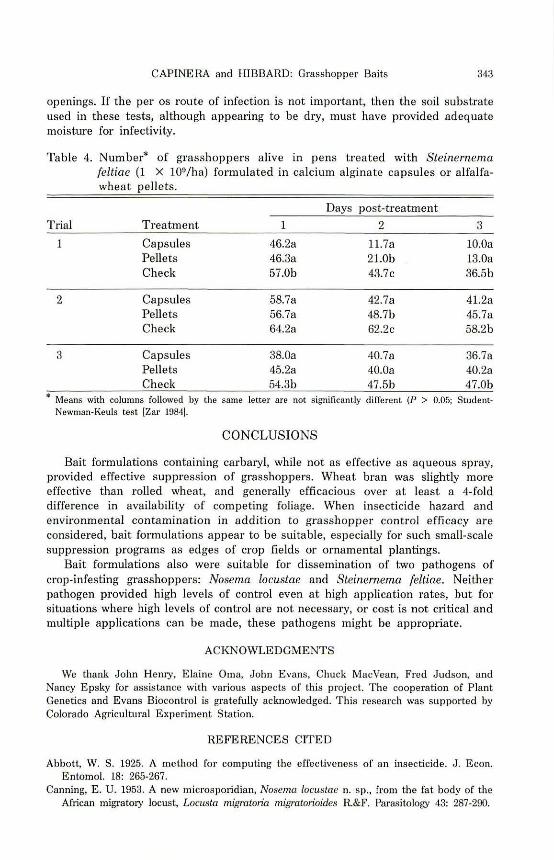
Both calcium alginate capsules and alfalfa-wheat pellets increased the mortality rate of grasshoppers compared with that of the control (Table 4). Mortality (adjusted % control) was 19.1, 39.5, and 41.2% for capsules and 15.7, 29.8, 33.4% for pellets, respectively.
Kayo and Nelsen (1985) reported thot beet armyworm, Spodoplera exigua (Hubner), larvae were infected by S. [eltiae released from calcium alginate capsules by the feeding activity of the caterpillars. Infection occurred on moist filter paper, but not on dry paper, suggesting that nematodes in a moist environment were capable of moving to the host and penetrating through the spiracles 01' other
343 CAPINE RA and HIBBARD: Grasshopper Baits
openings. If the per os route of infection is not important, then the soil substrate used in these tests, although appearing to be dry, must have provided adequate moisture for infectivity.
Table 4. Number'" of grasshoppers alive in pens treated with Steinernema feltiae (1 X 109/ha) formulated in calcium alginate capsules or alfalfa· wheat pellets.
Days post-treatment
Trial Treatment 2 3
1 Capsules 46.28 1L7a 10.0. Pellets 46.3. 21.0b 13.0. Check 57.0b 43.7c 36.5b
2 Capsules 58.7a 42.78 41.2a Pellets 56.7a 48.7b 45.7a Cbeck 64.2a 62.2c 58.2b
3 Capsules 38.0a 40.7a 36.7a Pellets 45.2a 40.08 40.2. Check 54.3b 47.5b 47.0b
.. Means with columns followed by the same letter are not signiliculltly different (P > 0.05; StudentNewman-Keuls test tZar 198·11.
CONCLUSIONS
Bait formulations containing carbaryl, while not as effective as aqueous spray, provided effective suppression of grasshoppers. Wheat bran was slightly more effective than rolled wheat, and generally efficacious over at least a 4-fold difference in availability of competing foliage. When insecticide hazard and environmental contamination in addition to grasshopper control efficacy are considered, bait formulations appear to be suitable, especially for such small·scale suppression programs as edges of crop fields or ornamental plantings.
Bait formulations also were suitable for dissemination of two pathogens of crop-infesting grasshoppers: Nosema locustae and Steinemema feltiae. Neither pathogen provided high levels of control even at high application rates, but for situations where high levels of control are not necessary, 01' cost is not critical and multiple applications can be made, these pathogens might be appropriate.
ACKNOWLEDGMENTS
We thank John Henry, Elaine Oma, John Evans, Chuck r\'IacVean, Fred Judson, and Nanc)' Epsky for assistance with various aspects of this project. The cooperation of Plant Genetics and Evans Biocont.1'01 is gratefully acknowledged. This research was supported by Colorado Agricultural Experiment Station.
REFERENCES C1TED
Abbott, \V. S. 1925. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 18: 265-267.
Canning, E. U. 1953. A new microsporidian, Nosema locuslae ll. sp., from the fat body of the Abican migratory locust, Locu.sta migral0l1a migratorioides R.&F. Parasitology 43: 287-290.

344 J. Agric. Entomol. Vol. 4, No.4 (1987)
Ewen, A. R, and M. K. MukcJji. 1980. Evaluation of Nosema locustae (Microsporida) as a control agent of grasshopper populations in Saskatchewlill. J. Invertebr. Pathol. 35: 295303.
Henry, J. E. 1971. Experimental application Nosema Joct/sloe for control of grasshoppers. J. Invertebr. Pathal. 18: 339-394.
HenI)', H. E. 1981. Natuml and applied control of insects by protozoa. Annu. Rev. Entomol. 26, 49-73.
Henry, J. E., and E. A. Oma. 1981. Pest control by Nosema. /ocuslae, a pathogen of grasshoppers and crickets, pp. 573-586. In H. D. Burges, Ed. Microbial control of Pests and Plant Diseases 1970-1980. Academic Press, London.
Henry, J. E., and J. A. Onsager. 1982. Large-scale test of control of grasshoppers on rangeland with Nosema locus/ae. J. Econ. Entomol. 75: 31·35.
Johnson, D. L., and E. Pavlikova. 1986. Reduction of consumption by grasshoppers (Orthoptera: Aclididae) infected with Nosema locus/ae Canning (Microspolida: Nosemutidae). J. Invertebr. Pathol. 48: 232-238.
Johnson, D. 1., and J. E. Henry. 1987. Low rates of insecticides and Nosema locus/at: (Microsporidia: Nosemlltidae) on baits applied to roadsides for I,."rasshopper (Orthoptera: Acrididae) control. J. Econ. Entomo!. 80: 685-689.
Kaya, H. 1<., and C. E. Nelsen. 1985. Encapsulation of steinernematid and heterorhabditid nematodes with calcium alginate: a new approach for insect control and other applications. Environ. Entomol. 14: 572-574.
MacVean, C. l\'1., J. W. Brewer, and J. 1. Capinera. 1982. Field tests of ant.idesiccants to extend the infection period of an entomogenous nematode, Neoaplectana carpocapsac, against the Colorado potato beetle. J. Econ. Entomol. 75: 97-101.
Mukerji, M. K., and A. B. Ewen. 1984. Field evaluation of cypermethrin and carbaryl as sprays and baits for grasshopper (Orthoptera: Acrididae) control in Saskatchewan. Cmt. Entomo1.: 116: 5-9.
Mukerji, M. K., A. B. Ewen, C. H. Craig, and R. J. Ford. 1981. Evaluation of insecticidetreated bran baits for grasshopper control in Saskatchewan (Orthoptera: Acrididae). Can. EntomoL 113: 705-710.
Onsager, J. A., J. E. Henry, and R. N. Foster. 1980a. A model for predicting efficacy of carbaryl bait for control of rangeland grasshoppers. J. Econ. Entomol. 73: 726-729.
Onsager, J. A., J. E. Henry, R. N. Foster, and R. T. Staten. 1980b. Accepbnce of wheat bl'llll bait by species of nlllgeland grasshoppers. J. Econ. Entomol. 73: 548-551.
Onsager, J_ A., N. E. Rees, J. E. Henry, and R. N. Foster. 1981. Integration of bllit fonnulations of Nosema. locustlle and carbaryl for cont.rol of mngeland grasshoppers. J. Econ. Entomol. 74: 183-187.
Parker, J. R. 1939. Grasshoppers and their control. USDA ji'al'mcrs' Bull. 1828. Shotwell, R. L. 1942. Evalullt.iol1 of baits and bait ingredients used in grasshopper control.
USDA Tech. Bull. 793. Zur, J. H. 1984. Biostatisticul Analysis, second edition. Prentice-Hall, Englewood Cliffs,
NJ.





























