Embed Size (px)
Citation preview

BacteriorhodopsinBacteriorhodopsin
The Purple Membrane Protein
Mike GoodreidCHEM*4550 seminar

Outline of PresentationOutline of Presentation
Introduce Bacteriorhodopsin (BR)History of its structural analysisStructural features of the proteinMechanism of actionEnergy involved in action

Source of BRSource of BR
Archaebacteria Halobacteria Salinarium are the source of bacteriorhodopsin
They are halophilic bacteria (found in very salty water e.g. Great Salt Lake)

What is the purple What is the purple membrane?membrane?
The purple membrane patches are areas on the membrane where BR is concentrated
BR absorbs light @ 570 nm (visible green light)
Red and Blue light is reflected, giving membrane its purple colour

So what does BR do?So what does BR do?
BR functions as a proton pumpLong story short: protons are pumped one at
a time from the inside of the cell to the outside
Photons react with a bound retinal group causing conformational change in BR

Photons for ProtonsPhotons for Protons
Bacteriorhodopsin takes energy from photons This energy is converted and creates a proton
gradient by pumping protons outside the cell Protons are allowed back into the cell by an ATP
synthase In a nutshell: Photons are used to power the cell

Milestones in BR Structural Milestones in BR Structural DeterminationDetermination
In order to assess the structure and mechanism of BR, or any membrane protein, we really need to understand its tertiary structure by X-ray crystallography
BUT, membrane proteins don’t crystallize easily

Nobel Prize in Chemistry Nobel Prize in Chemistry (1988)(1988)
Hartmut Michel First to crystallize
BR in 1980 Contribution to
determination of structure of a photosynthetic reaction center earned him a Nobel Prize
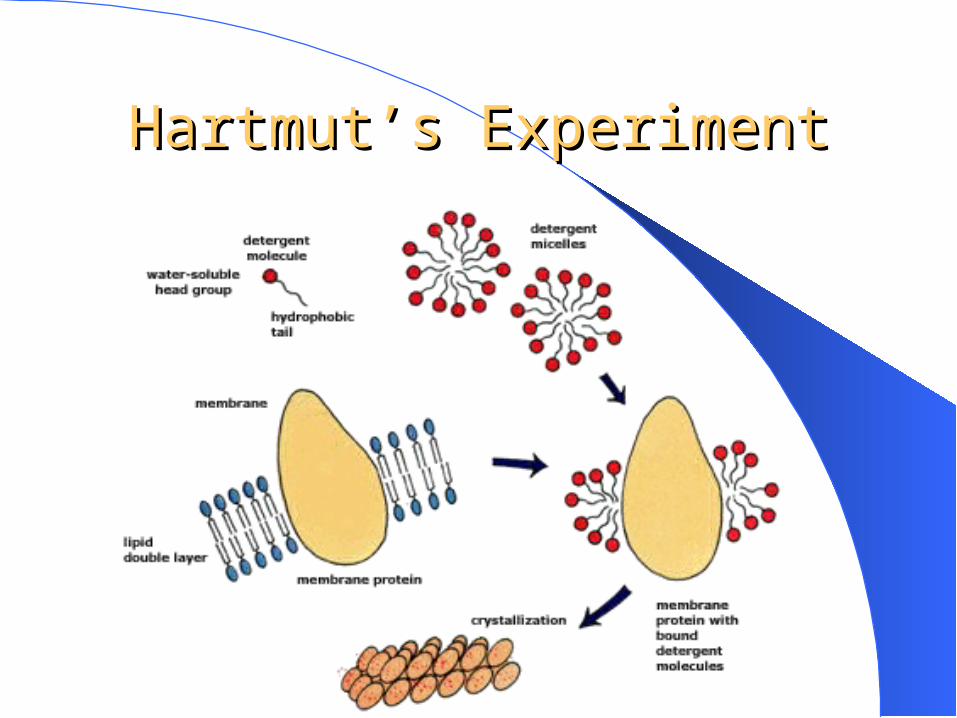
Hartmut’s ExperimentHartmut’s Experiment

FindingsFindings
Could get protein crystallizationCrystals were too small and disordered to
determine tertiary structureResults uncommon because
– BR is a very stable protein– BR forms a 2D lattice in vivo and in vitro
(later)

19901990
Henderson et al. use cryo-crystallography to study BR
Crystallization occurredFirst instances of structural determinationHowever, some areas of the protein could
not be resolved

1990 First structure of BR1990 First structure of BR
First structures of BR from side and top/bottom

1996: E.M Landau 1996: E.M Landau & J.P. Rosenbusch & J.P. Rosenbusch
Paradigm shift in crystallization of membrane proteins
Use Cubic Lipid Phase MatrixFirst complete structural determination of
BR

Intro to CLPIntro to CLP
CLP matrix (bicontinuous cubic phase)
Involves -high lipid content monoolein
(1-monooleoyl-rac-glycerol, C18:1c9, = MO) -aqueous pores that
penetrate membrane -proteins embedded
At high concentrations of lipids, more complex phasebehaviour occurs (say goodbye to micelles and bilayers)

Seeding and FeedingSeeding and Feeding
Purple membrane patches (or BR monomers) diffuse into the CLP
Addition of Sorensen salt increases curvature of the CLP’s membranes

Seeding and FeedingSeeding and Feeding
Protein separates into planar domains (crystal formation)
Mature crystals co-exist with BR depleted cubic phase
Hydration (dilution of Sorensen salt solution) reverses the crystallization process (crystals dissolve back into CLP matrix)

ResultsResultsHexagonal crystals from MO bicontinuous lipid phase lead to complete structural determination of BR(3.7 Angstrom resolution)

BR gene expressionBR gene expression
786 nt structural gene13 AA precursor sequence
+248 AA in mature BR
+1 AA (D) at C-terminal sequenceNo intervening sequencesNo prokaryotic promoter (yet?)

Brp has role in retinal synthesis from beta-caroteneBlh has a similar role(?)

Structural Features of BRStructural Features of BR
Extracellular matrix
Cytosol
H+ | |V

Structural InfoStructural Info
7 TM helices Forms a homotrimer Homotrimers
aggregate to form the purple membrane
Stability of trimer by:– G113, I117, L48– Most stability comes
from surrounding lipids

Are There Any Highly-Are There Any Highly-conserved Residues?conserved Residues?
You’d better believe it!L. Brown, 2001:-Upon BLASTing the H. Salinarium BR, found very high homology among all BR from a number of different Halobacterium -Around the K216 schiff base, there is no deviation in AA composition for a good 4.5 Angstroms-This type of analysis shows the entire retinal binding pocket is highly conserved. Therefore, MANY of the AAs in BR are structurally and/or catalytically important. SDM is a useful tool for validating this statement.

PhotocyclePhotocycle
A lesson in pushing protons

Photocycle of BR begins with absorption of a photon withwavelength of 550 nm. All-trans retinal13-cis retinal
1313 |CHO
All-trans retinal (blue)Carbon 13 (red)

Photocycle of BR begins with absorption of a photon withwavelength of 550 nm. All-trans retinal13-cis retinal
1313 |CHO
13-cis retinal (blue+cyan)Carbon 13 (red)
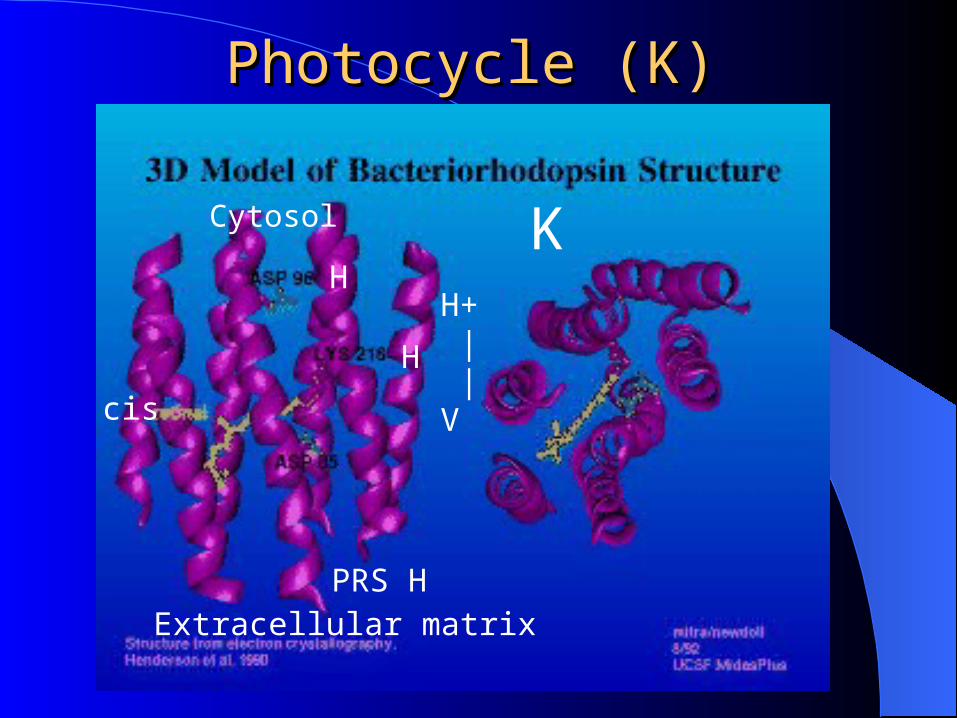
Photocycle (K)Photocycle (K)
Extracellular matrix
Cytosol
H+ | |V
K
H
H
PRS H
cis

Photocycle (L)Photocycle (L)
Extracellular matrix
Cytosol
H+ | |V
KL
H
H -Partial retinal relaxation-Subtle changes in protein conformation
PRS H
cis

Photocycle (M)Photocycle (M)
Extracellular matrix
Cytosol
H+ | |V
LM
H
H -K216 (schiff base deprotonated)-D85 picks up proton (perhaps via H2O intermediate)-Proton lost from PRS
PRS
cis

Photocycle (N)Photocycle (N)
Extracellular matrix
Cytosol
H+ | |V
MN
H
H
-D96 deprotonated-K216 picks up proton
PRS
cis

Photocycle (O)Photocycle (O)
Extracellular matrix
Cytosol
H+ | |V
NO
H
H
-Retinal reisomerizes back to All-Trans -D96 reprotonated from cytosol
PRS
H

Photocycle (final step)Photocycle (final step)
Extracellular matrix
Cytosol
H+ | |V
OK
H
H
-D85 deprotonated -PRS reprotonated-back to square 1 until another proton isomerizes the All-trans retinal
PRS
H

Basic BiophysicsBasic Biophysics
And now for something completely different

Thermodynamics of TransportThermodynamics of Transport
Energy of a photon:
E=hc/lambda
let lambda = 550 nm
Ephoton=3.61x10^(-19) J
Energy req’d to move H+
/\G=RTln([H+out]/[H+in])
-zF/\psi
let: H+out=10,000 H+in, T=295K
/\G=3.75E-20(J/H+) - zF/\psi
let: Vm=-60mV (an estimate)
/\G=(3.75(E-20) – 9.61E(-21)) J/H+
/\G = + 4.7E-20JSince Ephoton>/\G, we can see that the photon is sufficiently energized to move the proton

What promise does BR hold?
Bioengineering:
-Scaffold for a light poweredCation pump
-Facilitate environmentalcleanup of heavymetals
-Cheap, easy way of accumulating protons:
-Industry-Fuel cell cars

ReferencesReferencesLanyi, J.K. (2001) Biochemistry (Moscow) 66, 1477-1482
Brown, L.S. (2001) Biochemistry (Moscow) 66, 1546-1552
Dunn, R., McCoy, J., Simsek, M., Majumdar, A., Chang, S.H., Rajbhandary, U.L., and Khorana, H.G..(1981) Proc Natl Acad Sci USA.78, 6744-6748
Jagannathan, K., Chang, R., and Yethiraj, A. (2002) Biophys J 83, 1902-1916
Peck, R.F., Echavarri-Erasun, C., Johnson, E.A., Ng, W.V., Kennedy S.P., Hood, L., DasSarma, S., and Krebs, M.P. (2001) J Biol Chem. 23, 5739-5744
Landau, E.M.and Rosenbusch, J.P. (1996) Proc. Natl. Acad. Sci. USA93, 14532-14535
Nollert, P. Qiu, H., Caffrey, M., Rosenbusch, J.P., and Landau, E.M. (2001)FEBS Lett. 504, 179-186