Embed Size (px)
Citation preview

Bacterial MicrocompartmentsThese intracellular structures, formed by polypeptides, confine and helpto retain nonpolar and volatile metabolites
Thomas A. Bobik
Although the carboxysome was fornearly 30 years the only microcom-partment recognized within micro-bial cells, additional microcompart-ments, including some that are
involved in metabolic processes such as degrad-ing carbon sources, are known. Cluster analysisof homologues of microbial microcompart-ment-specific proteins suggests that such enclo-sures could be involved in as many as sevendifferent metabolic processes in various bacte-rial species. Although such microcompartmentsappear to provide barriers for molecules that arepoorly retained by lipid-based membranes, theirother functions, if any, and how they operateremain unknown.
The carboxysome and similar microcompart-ments contain 5–10 different polypeptides thatform a polyhedral shell 100–200 nm across andwhose volume is more than 1,000 times greaterthan that of a ribosome. Their size helps toexplain why early investigators mistook them
for bacteriophage (Fig. 1). The building blocksof bacterial microcompartments are exclusivelyproteins and glycoproteins; there is no publishedevidence that they contain DNA, RNA, or lip-ids. Indeed, electron microscopy shows no lipidmonolayer or bilayer surrounding such micro-compartments, making them the only knownprotein-based metabolic compartments in livingcells. An important unifying property of micro-compartments appears to be that they retainvolatile compounds, according to Joe Penrodand John Roth at the University of California,Davis.
Other Microcompartments,
Carboxysomes Share Features
Carboxysomes were identified during the late1960s, first through electron microscopy studiesof autotrophic bacteria, but soon they were alsofound in cyanobacteria and in chemoau-
totrophs. The shell of a carboxysome con-tains multiple copies of 6 to 10 differentpolypeptides, while the lumen containsmany molecules of ribulose bisphosphatecarboxylase/oxygenase (RuBisCO), theCO2-fixing enzyme of the Calvin cycle. Thecarboxysome apparently concentrates CO2
for RuBisCO, which converts it and ribu-lose bisphosphate into two molecules of3-phosphoglycerate (Fig. 2).
Similar microcompartments are alsofound in nonphotosynthetic bacterial spe-cies where they are involved in other meta-bolic processes. Two distinct microcompart-ments (Fig. 3), found in the heterotrophicenteric bacterium Salmonella enterica, con-tain the components, including coenzymeB12, needed for degrading 1,2-propanediol
Summary
• Some bacterial cells have microcompartments,including the carboxysome, which is involved infixing carbon dioxide, and others that appar-ently help to sequester volatile or toxic metab-olites.
• Although about 25%, or 85 of 337 bacterialgenomes whose sequences are available containgene homologues suggesting that they can makemicrocompartments, no such homologues arefound in Archaea or Eucarya.
• The distinguishing feature of bacterial mic-roompartments is their structure, which con-sists of a complex protein shell that encasesmetabolic enzymes
Thomas A. Bobik isan associate profes-sor in the Depart-ment of Biochemis-try, Biophysics, andMolecular Biology,Iowa State Univer-sity, Ames.
Volume 2, Number 1, 2007 / Microbe Y 25

or ethanolamine. They are important carbonsources for such cells in natural environments.Members of other bacterial genera, includingEscherichia, Klebsiella, Clostridium, Fusobacte-rium, Shigella, Listeria, and Yersinia, also candegrade one or both of these compounds andappear to use microcompartments for this pur-pose.
These catabolic microcompartments werelong overlooked because they do not form whencells are grown on standard laboratory media.Moreover, electron microscopy is required toobserve them. They were discovered after re-searchers realized that the operons specificallyinvolved in degrading 1,2-propanediol and eth-anolamine include genes that encode multiplehomologues of carboxysome shell proteins.
My collaborators and I purified and analyzedthe microcompartment involved in degrading1,2-propanediol in 2003 (Fig. 3). It is composedof at least 14 different polypeptides, includingfour enzymes and seven different putative shellpolypeptides (Fig. 4). Although the ethanol-amine microcompartment has not been purified,it is unlikely to differ dramatically in numbers ofprotein components from those found in theother two microcompartments.
Some investigators suspect that the 1,2-pro-panediol and ethanolamine microcompartmentshelp to retain the two volatile aldehydes, propi-onaldehyde and acetaldehyde, respectively, thatare degradative intermediates in these two path-ways (Fig. 4). Retaining them confers two ben-efits: protecting cytoplasmic components againsttheir toxic effects, and preventing their lossthrough lipid-based membranes.
The Microcompartment
Phenomenon Seems To Be
Restricted to Bacteria
The discovery of microcompartmentsin cyanobacteria, Salmonella, and otherbacteria raised questions about theirdistribution and diversity. How wide-spread is the microcompartment phe-nomenon and how diverse is microcom-partment function?
To address these questions, we re-cently searched GenBank for micro-compartment shell genes and analyzedgene clusters within which they reside.About 25% (85 of 337) of bacterial ge-nomes whose sequences are available inthis database contain shell gene homo-
logues. However, no homologues were foundamong genomic sequences for archaea oreukaryotes.
In most of those 25% of bacterial genomesthat carry those homologue genes, the shellgenes cluster with other genes encoding micro-compartment-associated enzymes. Based on thisconserved arrangement, we define a microcom-partment genomic signature (MGS) as a genecluster that includes multiple shell genes inter-spersed with structural genes encoding enzymes.Further, we assign microcompartment types basedon the enzymes that are encoded within an MGS.For example, signatures that encode shell proteinsas well as B12-dependent diol dehydratase aretyped as a microcompartment for degrading 1,2-propanediol.
Based on the diversity of enzymes withinMGS, we estimate that microcompartments areinvolved in seven or more different metabolicprocesses. Those involved in fixing CO2 or indegrading 1,2-propanediol and ethanolaminecan be considered well-established. The remain-ing four are putative, of unknown function, andmight underestimate the total because our anal-ysis would miss microcompartments containingshell proteins that diverge substantially fromthose that are known. Perhaps, as with viralcapsids, there is a great diversity of microcom-partment shells, serving an array of metabolicpartitioning needs of other microbes.Probing Microcompartment
Structural Components
The structure of the microcompartment—acomplex protein shell encasing metabolic en-
F I G U R E 1
Electron micrograph of polyhedral microcompartments: a. The carboxysomes of H.neapolitanus; b. The microcompartments formed during growth of Salmonella entericaon 1,2-propanediol. Arrowheads point to the microcompartments.
26 Y Microbe / Volume 2, Number 1, 2007

zymes—is unusual and distinguishes bacterialmicrocompartments from all other metabolicmultiprotein complexes.
The microcompartment shell somehow influ-ences the activity of enzymes within the lumen.For instance, mutants that cannot properly formthe carboxysome shell require high CO2 forautotrophic growth, indicating that the shellplays a role in increasing the overall catalyticefficiency of RuBisCO. The 1,2-propanedioland ethanolamine microcompartments containcatabolic enzymes involved in producing and
consuming aldehydes. In these cases, shell mu-tants release massive amounts of aldehydes intothe growth medium, suggesting that these mi-crocompartments are important for retainingaldehydes and possibly for regulating aldehdyeproduction.
Microcompartments also may influence met-abolic reactions by acting as containers for en-zyme substrates. For instance, the carboxysomeshell seems to be a barrier to CO2, concentratingit in the immediate vicinity of RuBisCO and thusenhancing its rate of fixation. Similarly, the 1,2-
Bobik: Dig Deeply into Scientific Problems To Find KnowledgeThomas Aquinas Bobik wanted towork, not attend college, afterhigh school. However, his motherpersuaded him to try college forone year, which “turned intofour, and then into five more ingraduate school,” he says. “Myhigh school chemistry teacher,Mr. Motts, got me interested inscience, and later I went to Indi-ana University at Bloomingtonwhere there was an excellent un-dergraduate program in micro,”he recalls. “I liked the blend ofbiology and chemistry, and I ma-jored in microbiology.”
Since 2004, Bobik has been anassociate professor in the Depart-ment of Biochemistry, Biophysicsand Molecular Biology, at IowaState University, where he focuseson vitamin B12 metabolism inbacteria. “My research focuses onunderstanding bacterial metabo-lism. I chose this area because or-ganic chem was one of my favor-ite subjects,” he says. “I like theorder it brings to chemical reac-tions.” Bobik also is investigatingmicrobial production of fuels, “ahot topic these days,” he says.“Petroleum reserves are limited.Available energy is important tothe quality of human life, so an
alternative to fossil fuels will beneeded in the coming years.”
Bobik grew up in South Bend,Ind., where his father teaches phi-losophy at the University of NotreDame. “Of course, I am an ‘Irish’[Notre Dame] football fan,’’ hesays. As a child, he enjoyed takingthings apart “to find out how theyworked, and then trying to use theparts to build new things—al-though this latter goal was neverfully realized,” Bobik says. Hismother called him “DoubtingThomas’’ because “I questionedwhat I was told, and what peoplesaid.” He has one brother andthree sisters. Although one sisteris a medical technician, “my othersibs did not choose a science-based career,” he says.
Bobik received a B.S. degree inmicrobiology from Indiana Uni-versity, then moved to the Univer-sity of Illinois where he completedan M.S. in 1986, then in 1990 aPh.D., both in microbiology. Af-ter doing postdoctoral research atthe University of Utah in Salt LakeCity, he became an assistant pro-fessor in the department of micro-biology and cell science at theUniversity of Florida in Gaines-ville, where he remained untilmoving to Iowa in 2004, conduct-
ing genetic studies of vitamin B12
metabolism in Salmonella andin higher organisms, and alsoworking on bacterial biosensors.
Bobik credits several mentorswith enriching his appreciation—and respect—for research. “Mygraduate advisor, Ralph Wolfe,taught me the importance of sci-entific rigor,” he says. “He wouldsay that the foundation must bestrong to support the weight offuture advances. He would alsosay that a stool needs at least threelegs to stand up—and that threeindependent lines of evidence areneeded to support any scientificdiscovery.”
His postdoctoral advisor, JohnRoth, taught him important les-sons about the nature of bacterialgenetics. “Bacterial genetics is, inpart, a puzzle where we try to puttogether enough pieces so that wecan make scientific sense out ofphenotypic observations,” Bobiksays. “He would say that ‘there’sa pony in there somewhere,’meaning that if we dig deep into aproblem, we can find new scien-tific knowledge.”
Marlene Cimons
Marlene Cimons is a freelance writerin Bethesda, Md.
Volume 2, Number 1, 2007 / Microbe Y 27
propanediol and ethanolamine microcompart-ments apparently sequester aldehydes, prevent-ing toxicity and carbon loss (Fig. 4). Althoughthese microcompartments respectively restrictthe flow of aldehydes and CO2, they somehowremain selectively permeable to other sub-strates, products, and cofactors.
Some microcompartment shells appear to in-corporate transport systems and selective pores.For example, the enzyme carbonic anhydrase isa component of the carboxysome shell in Halo-thiobacillus, according to Gordon Cannon andhis collaborators at the University of Southern
Mississippi in Hattiesburg. Their find-ing suggests that CO2 molecules entermicrocompartments by means of anhy-drase-catalyzed transport.
In some cases, positive or negativecharges may be used to control the flowof molecules between the cytosol andlumen of microcompartments. ToddYeates and his collaborators at the Uni-versity of California, Los Angeles, re-cently determined the crystal structuresof the carboxysome shell proteinsCcmK2 and CcmK4. Both are wedge-shaped monomers that pack tightly toform a hexamer with a central pore.These hexamers have nearly flat sidesand form tight sheets that could formthe outer face of these microcompart-ment shells. The central pores and thegaps between hexamers are 4–7 Åwide. Because the hexamers have alarge net positive electrostatic potential,the pores might accommodate onlycharged molecules and thus would notallow passage of uncharged compoundssuch as CO2.
Much about Microcompartment
Functions Remains Uncertain
Precisely how microcompartmentswork is uncertain. Of the several differ-ent types of microcompartments, thecarboxysome is best understood. Theirmain function seems to be enhancingCO2 fixation through a CO2-concen-trating mechanism (CCM) (Fig. 2). TheCCM includes an active transport sys-tem for concentrating dissolved CO2 in
the form of HCO3- in the cell cytoplasm. How-
ever, HCO3- and CO2 do not equilibrate in the
cytoplasm because CA is not present there. In-stead, a carboxysome-associated CA convertsHCO3
- to CO2, which is released into the micro-compartment lumen, building to higher levelsuntil it equilibrates with cytoplasmic HCO3
-.Thus, high CO2 levels are maintained in theimmediate vicinity of RuBisCO within the mi-crocompartment lumen, increasing the CO2-fix-ation rate and suppressing photorespiration (anonproductive process in which O2 competi-tively inhibits RuBisCO-catalyzed carboxyla-tion).
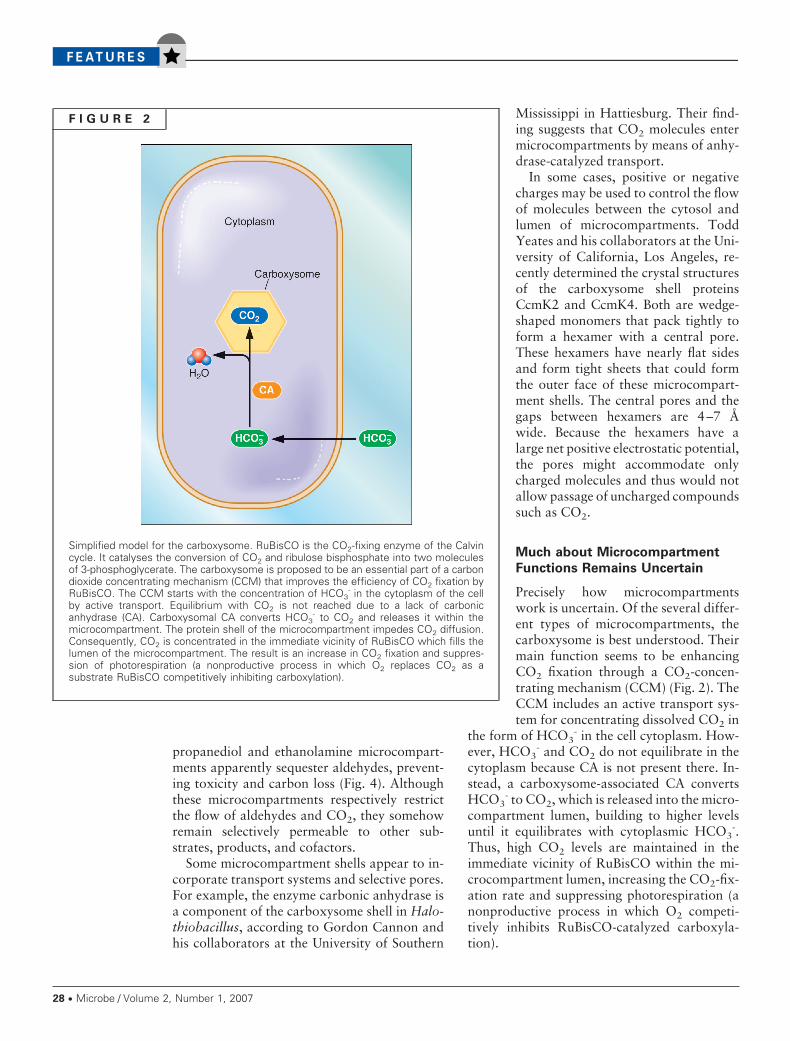
F I G U R E 2
Simplified model for the carboxysome. RuBisCO is the CO2-fixing enzyme of the Calvincycle. It catalyses the conversion of CO2 and ribulose bisphosphate into two moleculesof 3-phosphoglycerate. The carboxysome is proposed to be an essential part of a carbondioxide concentrating mechanism (CCM) that improves the efficiency of CO2 fixation byRuBisCO. The CCM starts with the concentration of HCO3
- in the cytoplasm of the cellby active transport. Equilibrium with CO2 is not reached due to a lack of carbonicanhydrase (CA). Carboxysomal CA converts HCO3
- to CO2 and releases it within themicrocompartment. The protein shell of the microcompartment impedes CO2 diffusion.Consequently, CO2 is concentrated in the immediate vicinity of RuBisCO which fills thelumen of the microcompartment. The result is an increase in CO2 fixation and suppres-sion of photorespiration (a nonproductive process in which O2 replaces CO2 as asubstrate RuBisCO competitively inhibiting carboxylation).
28 Y Microbe / Volume 2, Number 1, 2007
According to the CCM model, car-boxysomes are permeable to both ribu-lose bisphosphate and 3-phosphoglyc-erate, the substrate and product of theRuBisCO-catalyzed, CO2–fixing reac-tion. Importantly, the carboxysomesrestrict the movement of CO2. Theircharged pores could provide a conduitfor ribulose bisphosphate and 3-phos-phoglycerate, while restricting themovement of uncharged compoundssuch as CO2. According to this model,CO2 could be brought into the lumenby carbonic anhydrase. Although the1,2-propanediol microcompartmentsare believed to retain the pathway inter-mediate propionaldehyde selectively,its retention during 1,2-propanedioldegradation involves selection among acomplex mix of reactants. The sub-strate for diol dehydratase (1,2-pro-panediol) is uncharged. Moreover, thelumen enzymes within the 1,2-pro-panediol microcompartment requireseveral cofactors, including ATP, NAD,coenzyme B12, and coenzyme A, all ofwhich are too large to fit through smallpores. An analogous situation exists forthe ethanolamine microcompartments.
A possible alternative to the selectivepermeability hypothesis is that the eth-anolamine microcompartments con-centrate the metabolic enzymes andtheir substrates, thereby improvinggrowth, minimizing acetaldehyde tox-icity, and maintaining coenzyme A bal-ances, suggests Jorge Escalante-Semer-ena and his collaborators at the University ofWisconsin, Madison.
Microcompartments Raise
Structural Issues
How are microcompartments constructed? Onepossibility is that they self-assemble with aidfrom chaperone proteins and other assemblyfactors. For instance, the ethanolamine micro-compartment eut operon includes genes thatencode two auxiliary proteins, EutA and EutJ.They appear to be distantly related to the DnaK-family of chaperonins. Although EutA is in-volved in reactivating ethanolamine ammonialyase, the function of EutJ is unknown, meaning
it remains a candidate microcompartment-assembly chaperone.
Similarly, two proteins required for carboxy-some function, OrfA and OrfB, cannot be foundin the final assembly. Thus, they might be minor(and thus undetectable) components that help toassemble these microcompartments.
Not all microcompartment gene clusters en-code proteins that appear to be related to chap-erones. However, other chaperones in cells orother factors might assist in forming microcom-partment structures, even though no such as-sembly factors have been identified.
Another puzzle is how proteins are targetedand find their way to the lumen of these protein-based compartments. Perhaps the shell forms
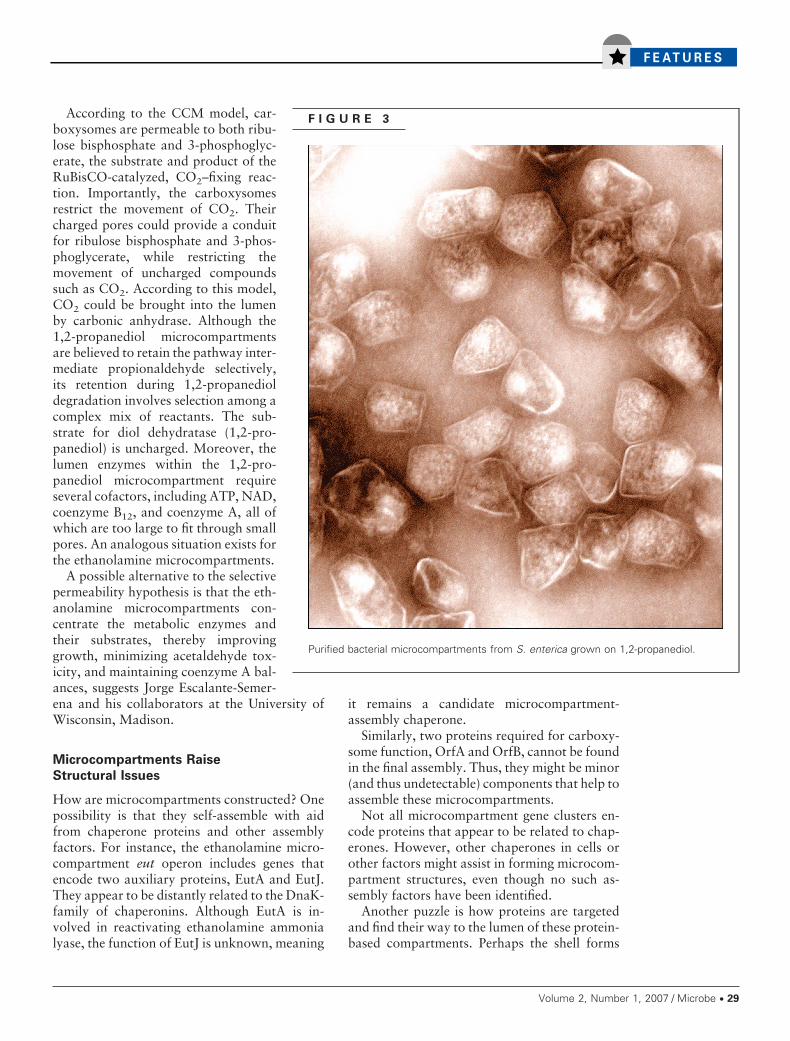
F I G U R E 3
Purified bacterial microcompartments from S. enterica grown on 1,2-propanediol.
Volume 2, Number 1, 2007 / Microbe Y 29
around the lumen enzymes, and maybe they actas a scaffold for that shell. Although this expla-nation is plausible for carboxysomes in whichthe small subunit of RuBisCO is required forproper assembly, it does not seem to fit the caseof the 1,2-propanediol microcompartments. Forthem, deleting the genes encoding each lumenenzyme one by one does not prevent the shellfrom forming. Thus, 1,2-propanediol micro-compartments are unlikely to depend on ahighly structured scaffold composed of lumenenzymes.
Several proteins found in the lumen of micro-compartments have N- or C-terminal exten-sions. For example, two of the three subunits ofB12-dependent diol dehydratase within the lu-men of the 1,2-propanediol microcompartmentshave short N-terminal extensions compared totheir homologues that are confined to the cyto-plasm. Deleting these extensions dramaticallyincreases the solubility of this diol dehydratase,but does not appreciably affect its catalytic ac-tivity, according to Tetsuo Toraya and his col-laborators at Okayama University in Japan.They speculate that these sequences are involved
in assembling microcompartments, presumablythrough protein-protein interactions.
Although microcompartment shells resembleviral capsids, electron microscopy reconstruc-tions indicate that carboxysomes are not icosa-hedra but are D6 polyhedra, a symmetry that isnot found among known viral capsids. More-over, Blast, PSI-Blast, and direct structuralsearches uncover no significant similarity be-tween viral capsid and microcompartment shellproteins. Thus, microcompartment shells andviral capsids appear to be unrelated to one an-other.
However, bacterial microcompartment shellsare not the only protein sheets known in biol-ogy. Viral capsids and cell envelopes of pro-karyotes and eukaryotes in many cases arepartly composed of protein sheets that help toprotect the cell or viral nucleic acid and providereceptors for substances binding to those cells.Bacterial microcompartments may indicate thatsuch protein sheets are more functionally so-phisticated than is appreciated. If so, the princi-ples of microcompartment design and functioncould have broad relevance in biology.
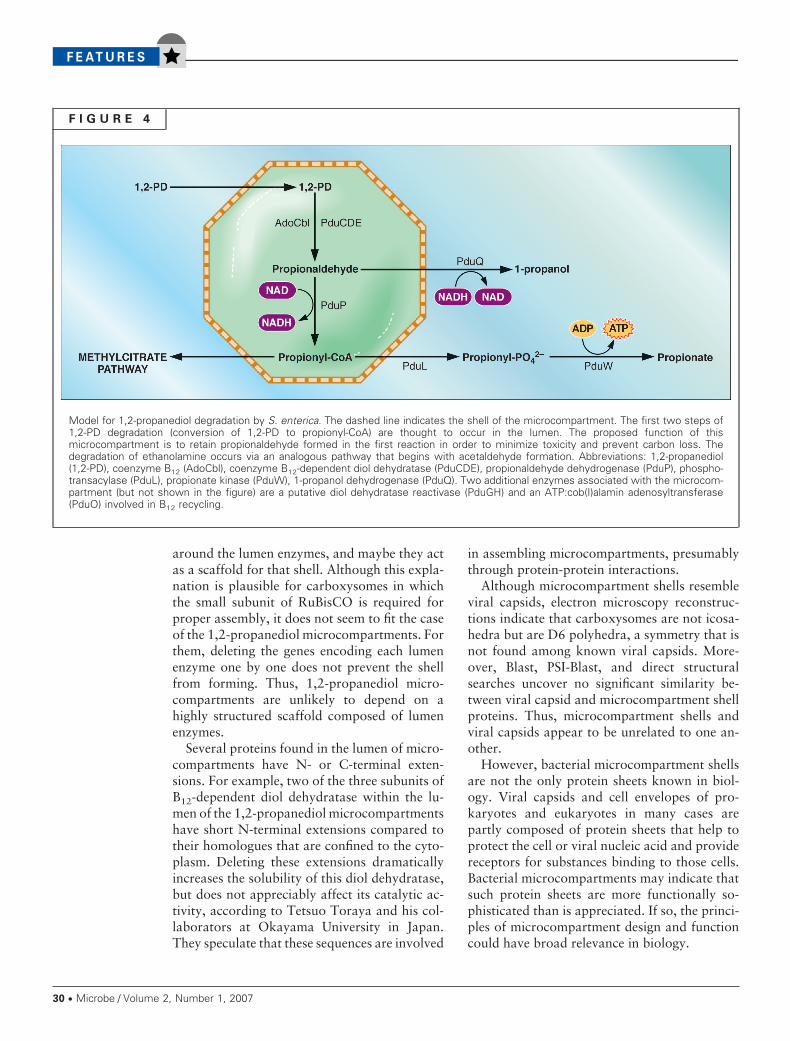
F I G U R E 4
Model for 1,2-propanediol degradation by S. enterica. The dashed line indicates the shell of the microcompartment. The first two steps of1,2-PD degradation (conversion of 1,2-PD to propionyl-CoA) are thought to occur in the lumen. The proposed function of thismicrocompartment is to retain propionaldehyde formed in the first reaction in order to minimize toxicity and prevent carbon loss. Thedegradation of ethanolamine occurs via an analogous pathway that begins with acetaldehyde formation. Abbreviations: 1,2-propanediol(1,2-PD), coenzyme B12 (AdoCbl), coenzyme B12-dependent diol dehydratase (PduCDE), propionaldehyde dehydrogenase (PduP), phospho-transacylase (PduL), propionate kinase (PduW), 1-propanol dehydrogenase (PduQ). Two additional enzymes associated with the microcom-partment (but not shown in the figure) are a putative diol dehydratase reactivase (PduGH) and an ATP:cob(I)alamin adenosyltransferase(PduO) involved in B12 recycling.
30 Y Microbe / Volume 2, Number 1, 2007

SUGGESTED READING
Badger, M. R., and G. D. Price. 2003. CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversityand evolution. J. Exp. Bot. 54:609–622.Bobik, T. A. 2006. Polyhedral organelles compartmenting bacterial metabolic processes. Appl. Microbiol. Biotechnol.70:517–525.Brinsmade, S. R., T. Paldon, and J. C. Escalante-Semerena. 2005. Minimal functions and physiological conditions required forgrowth of Salmonella enterica on ethanolamine in the absence of the metabolosome. J. Bacteriol. 187:8039–8046.Kerfeld, C. A., M. R. Sawaya, S. Tanaka, C. V. Nguyen, M. Phillips, M. Beeby, and T. O. Yeates. 2005. Protein structuresforming the shell of primitive bacterial organelles. Science 309:936–938.Penrod, J. T., and J. R. Roth. 2006. Conserving a volatile metabolite: a role for carboxysome-like organelles in Salmonellaenterica. J. Bacteriol. 188:2865–2874.Shively, J. M., G. van Keulen, and W. G. Meijer. 1998. Something from almost nothing: carbon dioxide fixation inchemoautotrophs. Annu. Rev. Microbiol. 52:191–230.So, A. K., G. S. Espie, E. B. Williams, J. M. Shively, S. Heinhorst, and G. C. Cannon. 2004. A novel evolutionary lineage ofcarbonic anhydrase (epsilon class) is a component of the carboxysome shell. J. Bacteriol. 186:623–630.
ASM Pre-Meeting Workshop Scholarships
The ASM International Education Committee is accepting applications for the 2007 ASM Pre-Meeting Workshop Scholarships. The Scholarship is available to ASM members and non-members currently residing in developing countries who are pursuing, or who are within five years of having received, their Masters, PhD, or equivalent academic degree.
This Scholarship will provide two successful applicants with registration to the pre-meeting workshop of their choice at the ASM General Meeting. A list of this year’s workshops will shortly be available on the ASM General Meeting website at http://gm.asm.org/workshops/workshop-overview.shtm.
Open to ASM members and non-members alike, applicants must have submitted an abstract to the meeting and received their acceptance e-mail. They must also have pre-registered to attend the ASM General Meeting.
Application Deadline – March 1, 2007
Program information and application form are on the ASM web page at
http://www.asm.org/International/index.asp?bid=18369
Volume 2, Number 1, 2007 / Microbe Y 31










![BACTERIOPHAGE-RESISTANT AND BACTERIOPHAGE-SENSITIVE ...halsmith/phagemutantsubmitted_2.pdf · BACTERIOPHAGE-RESISTANT AND BACTERIOPHAGE-SENSITIVE BACTERIA IN A CHEMOSTAT ... [22],](https://img.pdfslide.us/doc/110x75/5b3839687f8b9a5a518d2ce1/bacteriophage-resistant-and-bacteriophage-sensitive-halsmithphagemutantsubmitted2pdf.jpg)




![University of Bath · Recent Advances in Therapeutic Delivery Systems of Bacteriophage and Bacteriophage-Encoded Endolysins ... drug resistant bacterial isolates [27,28], and the](https://img.pdfslide.us/doc/110x75/5f0f3e237e708231d443313d/university-of-bath-recent-advances-in-therapeutic-delivery-systems-of-bacteriophage.jpg)