Embed Size (px)
Citation preview

of June 9, 2018.This information is current as
of Lysozyme in the Lungs of Transgenic MiceBacterial Killing Is Enhanced by Expression
WeaverHenry T. Akinbi, Ralph Epaud, Hetal Bhatt and Timothy E.
http://www.jimmunol.org/content/165/10/5760doi: 10.4049/jimmunol.165.10.5760
2000; 165:5760-5766; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/10/5760.full#ref-list-1
, 8 of which you can access for free at: cites 33 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 9, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Bacterial Killing Is Enhanced by Expression of Lysozyme inthe Lungs of Transgenic Mice1
Henry T. Akinbi, 2 Ralph Epaud, Hetal Bhatt, and Timothy E. Weaver
To assess the role of lysozyme in pulmonary host defense in vivo, transgenic mice expressing rat lysozyme cDNA in distalrespiratory epithelial cells were generated. Two transgenic mouse lines were established in which the level of lysozyme protein inbronchoalveolar (BAL) lavage fluid was increased 2- or 4-fold relative to that in WT mice. Lung structure and cellular compositionof BAL were not altered by the expression of lysozyme. Lysozyme activity in BAL was significantly increased (6.6- and 17-fold)in 5-wk-old animals from each transgenic line. To determine whether killing of bacteria was enhanced by expression of ratlysozyme, 5-wk-old transgenic mice and WT littermates were infected with 106 CFU of group B streptococci or 107 CFU of amucoid strain of Pseudomonas aeruginosaby intratracheal injection. Killing of group B streptococci was significantly enhanced(2- and 3-fold) in the mouse transgenic lines at 6 h postinfection and was accompanied by a decrease in systemic dissemination ofpathogen. Killing of Pseudomonas aeruginosawas also enhanced in the transgenic lines (5- and 30-fold). Twenty-four hours afteradministration of Pseudomonas aeruginosa, all transgenic mice survived, whereas 20% of the WT mice died. Increased productionof lysozyme in respiratory epithelial cells of transgenic mice enhanced bacterial killing in the lung in vivo, and was associated withdecreased systemic dissemination of pathogen and increased survival following infection.The Journal of Immunology,2000, 165:5760–5766.
Clearance of pathogens in the respiratory tract is mediatedat least in part by the synthesis and secretion of hostdefense molecules into the airway lumen. One of the
most abundant antimicrobial proteins in the lung, lysozyme, is syn-thesized and secreted by glandular serous cells, surface epithelialcells, and macrophages in the human airway (1). Human lysozyme(muramidase,N-acetyl muramide glycanohydrolase, EC 3.2.1.17)is a cationic protein of 148 aa that cleaves glycosidic bonds ofN-acetyl-muramic acid, damaging the bacterial cell wall and ulti-mately killing the organism by lysis (2). Bacteriolytic assays in-dicate that purified lysozyme is active against some Gram-positivebacteria, but has relatively little activity against Gram-negativebacteria, largely because the outer membrane of these organismslimits access of the enzyme to its substrate (3). The concentrationof lysozyme in human airway surface liquid ranges from 20–100mg/ml, a concentration sufficient to kill two important airwaypathogens,Pseudomonas aeruginosaandStaphylococcus aureus,in vitro (4). Despite numerous studies confirming the antibacterialproperties of lysozyme in vitro, there are no studies that have di-rectly assessed the role of lysozyme in the killing of lung patho-gens in vivo. The current study was therefore undertaken to eval-uate the impact of constitutively elevated levels of lysozyme onlung structure and pathogen clearance in transgenic mice.
The spatial expression of pulmonary lysozyme in rodents differsfrom that in humans in that the enzyme is principally synthesizedand secreted by type II alveolar epithelial cells (5, 6). Lysozyme is
one of the most abundant proteins in rat bronchoalveolar lavage(BAL)3 fluid, accounting for as much as 6–7% of the total solubleprotein (5). Although two lysozyme genes have been identified inthe rat, only the type I gene is expressed at detectable levels (7); incontrast, two lysozyme genes are expressed in the mouse, ly-sozyme M and lysozyme P (8, 9). Lysozyme M is the major formexpressed in the lung, and only very low levels of lysozyme PmRNA are detected in mouse lung (8). In the current study ratlysozyme type I, the homologue of mouse lysozyme M, wascloned under control of the human surfactant protein C (SP-C)promoter to direct expression of the transgene to the distal respi-ratory epithelium (10–12). Transgenic mice expressing ratlysozyme exhibited significantly enhanced antimicrobial activity,including enhanced bacterial killing, decreased systemic dissemi-nation of pathogen, and increased survival following infection.
Materials and MethodsGeneration of transgenic mice expressing rat lysozyme
The rat lysozyme cDNA was generated from rat type II cell RNA byRT-PCR using upstream primer 59-GAA TTC ATG AAG GCT CTC CTAGTT CTG and downstream primer 59-GAA TTC TCA GAC TCC GCAGTT CCG AAT AT. The amplified 400-bp fragment was confirmed to berat lysozyme by sequence analysis and was cloned into theEcoRI site ofBGI-hSP-C vector, which contained the 3.7-kb human SP-C promoter,rabbitb-globin intron 2, and bovine growth hormone polyadenylation sig-nal (13, 14). The transgene DNA construct was excised from the PUC18vector backbone byNotI/NdeI digestion, gel purified using Qiaex resin(Qiagen, Germany), dialyzed for 48 h against 5 mM Tris (pH 7.4)/1 mMEDTA, and microinjected into fertilized FVB/N oocytes by the Children’sHospital Transgenic Core facility. Founder mice were identified by a trans-gene-specific PCR with primers that amplified a 310-bp fragment spanningthe junction of the rabbitb-globin intron and the rat lysozyme cDNA(upstream primer, 59-AAT TCT GGC TGG CGT GGA AA; downstreamprimer, 59-TTG GTA GGG ATC CCC AAG GCA TT). PCR conditionswere 25 cycles at 55°C annealing temperature with 0.5mM transgene prim-ers, 0.25mM dNTPs, 1 U of AmpliTaq DNA polymerase (Perkin-Elmer/
Division of Pulmonary Biology, Children’s Hospital Medical Center, Cincinnati, OH45229
Received for publication February 7, 2000. Accepted for publication August 17, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants R01-HL-56285S(to T.E.W. and H.T.A.) and the Cystic Fibrosis Foundation RDP Center (to H.T.A.).2 Address correspondence and reprint requests to Dr. Henry T. Akinbi, Division ofPulmonary Biology, Children’s Hospital Medical Center, 3333 Burnet Avenue, Cin-cinnati, OH 45229-3039. E-mail address: [email protected]
3 Abbreviations used in this paper: BAL, bronchoalveolar lavage; SP-C, surfactantprotein C; GBS, group B streptococci; LB, Luria-Bertoni; WT, wild type; MIP-2,macrophage-inflammatory protein-2; m, murine.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Cetus, Foster City, CA), and 13AmpliTaq reaction buffer. PCR resultswere verified by Southern analyses using a32P-labeled probe that hybrid-ized to the rabbitb-globin intron. For all studies, transgenic mice werecompared with wild-type (WT) littermate controls. All mice used in thisstudy were housed in pathogen-free rooms in the animal facility of Chil-dren’s Hospital Medical Center (Cincinnati, OH).
Characterization of transgenic mice
RNA (RT-PCR) analysis.To identify lines that expressed rat lysozymemRNA, lung tissues were collected from 5- to 6-wk-old transgenic miceand control WT littermates. Total cellular RNA was isolated using TRIzolreagent (Life Technologies, Grand Island, NY). One microgram of DNaseI-treated RNA was reverse transcribed using the Superscript kit (Life Tech-nologies) with oligo(dT) as primer. For RT-PCR analyses, rat lysozymecDNA-specific primers (upstream, 59-ACA CAA GCC AGA AAC TACAACC; downstream, 59-GAA TTC TCA GCA TCC GCA GTT CCGAAT) were used with the PCR conditions described above.b-Actin wascoamplified in the PCR reaction as an internal control.Northern analysis. Ten micrograms of total RNA isolated from lung tis-sues of 5-wk-old transgenic mice and control WT littermates was fraction-ated by gel electrophoresis, blotted onto a nylon membrane, probed withbiotin-labeled rat or mouse lysozyme cDNA (provided by Dr. Rainer Ren-kawitz, Max Planck Institut fur Biochemie, Martinsried, Germany), anddetected by chemiluminescence (Pierce, Rockford, IL).Analysis of protein expression.To identify transgenic mouse lines thatexpressed rat lysozyme protein, Western blotting was performed using arabbit anti-human lysozyme Ab (Accurate Chemicals and Scientific Corp,Westbury, NY) that cross-reacts with both mouse and rat lysozyme (Mr,;15K). Lungs were isolated from 5-wk-old offspring of transgenicfounders and homogenized in PBS with 1% (v/v) protease inhibitor mix-ture (Sigma, St. Louis, MO). Protein concentration was determined bybicinchoninic acid protein assay (15). One-half microgram of total lungprotein was analyzed by SDS-PAGE and Western blotting, as previouslydescribed (16). To assess the level of secreted lysozyme, BAL fluid wasobtained from six 5-wk-old transgenic mice and five control WT litter-mates as described below. One-half microgram of total protein of BALfluid was resolved by SDS-PAGE and Western blotted with rabbit anti-human lysozyme Ab. Levels of lysozyme proteins in lung homogenatesand BAL fluids were quantitated by scanning densitometry using Image-Quant software (Molecular Dynamics, Sunnyvale, CA).BAL fluid cell count. Five 5-wk-old transgenic mice and five control WTlittermates were exsanguinated by transecting the abdominal aorta and theinferior vena cava after a lethal i.p. injection of sodium pentobarbital. Thelungs were lavaged three times with 1-ml aliquots of PBS. BAL fluid wascentrifuged at 20003 g for 10 min, and the pellet was resuspended in 0.5ml of PBS. A 50-ml aliquot was stained with an equal volume of 0.4%trypan blue (Life Technologies) for total cell count on a hemocytometer.Differential cell counts were made on cytospin preparations stained withDiff-Quik (Scientific Products, McGraw Park, IL).Lysozyme enzyme activity assay.To assess lysozyme enzyme activity,BAL fluid was obtained from 5-wk-old transgenic mice and control WTlittermates. BAL fluid containing 1mg of total protein was incubated withkilled Micrococcus lysodeikticussuspended in 0.4 M phosphate buffer, pH6.7, at an OD (450 nM) of 1 at 37°C. The change in OD was assessed after1 h of incubation (17). Purified chicken egg lysozyme was used to generatea standard curve (1 U of enzyme activity equals a 0.001 change in OD).Lung morphology and immunohistochemistry.To assess lung morphol-ogy, lungs from four 5-wk-old transgenic mice and four control WT lit-termates from transgenic line 3.5 were inflation-fixed for immunostainingand light microscopy as previously described (18). Immunostaining forlysozyme was performed using antiserum directed against human ly-sozyme (Accurate Chemicals and Scientific Corp.). Parallel lung sectionswere incubated with normal rabbit serum to verify the specificity ofimmunostaining.
Bacteria killing assays
Group B streptococci (GBS).Stock cultures of GBS used in this studywere clinical isolates provided by Dr. J. R. Wright (Duke University,Durham, NC). To minimize variation in virulence, all bacteria used in thisstudy were selected from aliquots of the same passage that had been frozenat270°C in 20% glycerol/PBS. For each experiment, an aliquot of bacteriawas thawed, plated on tryptic soy/5% defibrinated sheep agar plates, andsubsequently inoculated in 4 ml of Todd-Hewitt broth (Difco, Detroit, MI).Bacteria were grown for 14–16 h at 37°C with continuous shaking. Thebacteria were harvested from the broth by centrifugation at 2003 g for 10min, washed, and resuspended in sterile PBS at a concentration of 106
CFU/100ml. The concentration of the inoculum was verified by quantita-tive culture on sheep blood agar plates.Pseudomonas aeruginosa.The strain ofP. aeruginosawas a mucoidsegregant isolated from the sputum of a cystic fibrosis patient (provided byDr. D. J. Hassett, University of Cincinnati College of Medicine, Cincinnati,OH). P. aeruginosaobtained from a single passage was stored in aliquotsat 270°C in 20% glycerol/Luria-Bertoni (LB) broth. For each experiment,an aliquot of bacteria was plated on LB agar followed by inoculation into4 ml of LB broth. Preparation of the inoculum was conducted as describedfor GBS; the concentration (CFU) of the inoculum was verified by quan-titative culture on LB plates.Bacterial infection. The dose of bacteria selected for these studies wasbased on the results of previous studies in the FVB/N mouse strain (19).Mice were anesthetized with isofluorane, the trachea was exposed throughan anterior midline incision, and 106 CFU (GBS) or 107 CFU (P. aerugi-nosa), suspended in 100ml of sterile PBS, was delivered just beneath thecricoid cartilage. The incision was sealed by applying one drop of surgicalglue (Nexabrand, Veterinary Products Laboratories, Phoenix, AZ). As acontrol, 100ml of nonpyrogenic PBS was similarly instilled into transgenicand WT mice. To assess bacterial load at 6 h (GBS) or 24 h (P. aeruginosa)postinfection, mice were anesthetized with i.p. pentobarbital and exsangui-nated by transecting the abdominal aorta, and lung and splenic tissues wereharvested, weighed, and subsequently homogenized in sterile PBS. Serialdilutions of homogenates were plated on blood agar (GBS) or LB (Pseudo-monas) plates and incubated at 37°C overnight. Viable pathogen counts inthe lung and spleen were estimated from the number of colonies after 24 hof quantitative culture and expressed as CFU per gram of tissue.
To determine survival following infection, 20 5- to 6-wk-old transgenicmice from the 3.5 line and 20 WT littermates were infected with 23 107
CFU P. aeruginosaintratracheally, as described above. Water and foodwere provided ad libitum during the period of observation. The number ofsurviving mice was counted every 12 h for up to 60 h, at which time allsurviving mice were sacrificed.
Cytokine levels following infection withP. aeruginosa
Eight 5- to 6-wk-old transgenic mice and eight age-matched WT litter-mates were infected intratracheally with 107 CFU of P. aeruginosasus-pended in 100ml of PBS. Mice were sacrificed at either 6 or 24 h, lungswere lavaged as described above, and lung tissues were homogenized in 1ml of PBS with 1% (v/v) protease inhibitor mixture; the total volume afterhomogenization was adjusted to 2 ml with PBS. Mouse TNF-a, IL-6, mac-rophage inflammatory protein-2 (MIP-2), and RANTES were measured bysandwich enzyme immunoassay (R&D Systems, Minneapolis, MN). Thelimits of sensitivity for each cytokine are,5.1,,3.1, 1.5, and,2 pg/ml,respectively. All samples were assayed in duplicate, and values were nor-malized to total protein in the samples.
Statistical analyses
Differences between groups were assessed by one-way ANOVA, and dif-ferences between means were assessed by contrast comparisons and theStudent-Newman-Keuls test (StatView, Abacus Concepts, Berkeley, CA).Data are expressed as the mean6 SD. Nonparametric survival distribu-tions were estimated to examine differences in survival of transgenic miceand WT littermates. The differences between the two groups were analyzedusing Kaplan-Meier curve statistics.
ResultsGeneration of transgenic mice
To assess the role of lysozyme in pulmonary host defense, trans-genic mouse lines were generated in which rat lysozyme was tar-geted to the distal respiratory epithelium under the direction of the3.7-kb human SP-C promoter. Seven of 21 offspring from fertil-ized oocyte injections were positive for the transgene, as assessedby PCR and confirmed by Southern blot analyses of tail DNA (notshown). Transgenic offspring were indistinct from WT littermateswith respect to body weight, lung weight, longevity, and repro-ductive capability. Rat lysozyme mRNA was detected in the off-spring of two of seven transgenic lines (lines 3.5 and 2.6) by RT-PCR, and only these two lines were further characterized.
Analysis of transgene expression
Expression of the lysozyme transgene was assessed by Northernblot analysis of total RNA isolated from the lungs of 5-wk-old
5761The Journal of Immunology
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

mice. A cDNA probe specific for the rat lysozyme transgene de-tected an approximately 1-kb transcript (Fig. 1A). When the sameblot was probed with a mouse lysozyme cDNA, both the largerendogenous mouse lysozyme mRNA and rat lysozyme transgenemRNA were detected.
Because rat and mouse lysozyme have very similar m.w., it wasnot possible to distinguish the two proteins by SDS-PAGE. Totallevels of lysozyme (rat and mouse) were estimated by Westernblotting of equal amounts of protein from lung homogenates orBAL fluid from 5-wk-old transgenic mice and WT littermates.Total lysozyme protein in mice from transgenic line 2.6 was in-creased 2-fold in both lung homogenate (Fig. 1B) and BAL fluid(not shown); total lysozyme protein in mice from transgenic line3.5 was increased 4-fold compared with that in control WT litter-mates (Fig. 1B). Lysozyme (Mr, 142) was not detected when lunghomogenates and BAL were blotted with nonimmune rabbitserum.
To determine whether increased lysozyme protein levels wereassociated with increased enzyme activity, lysozyme enzyme ac-tivity in BAL fluid from 5-wk-old transgenic mice and wild-typelittermate controls was assessed by a turbidimetric assay usingpurified chicken egg lysozyme as a standard. Lysozyme enzymeactivity in mice from transgenic line 3.5 was increased 17.7-foldcompared with that in wild-type mice (550 U/ng BAL protein vs31 U/ng BAL protein in WT mice;p 5 0.0001; Fig. 2). As pre-
dicted from Western blot analyses, lysozyme enzyme activity inBAL from line 2.6 was increased relative to that in WT controls( p 5 0.02), but was less than that in BAL from transgenic line 3.5.
The pattern of lysozyme expression was characterized by im-munostaining of paraffin-embedded lung sections from 5-wk-oldtransgenic mice and control WT littermates. In WT mice, immu-nostaining of endogenous lysozyme was restricted to alveolar ep-ithelial cells, whereas in transgenic mice, lysozyme staining wasequally prominent in nonciliated bronchiolar cells and alveolar ep-ithelial cells (Fig. 3). No staining was detected in the submucosa orin nonepithelial cells in either WT or transgenic mice or whenparallel lung sections from transgenic mice were incubated withnonimmune rabbit serum. Lysozyme expression in transgenic micetherefore conforms to the previously reported expression patternfor the 3.7-kb human SP-C promoter (10–12).
The histologic features of paraffin-embedded, hematoxylin/eo-sin-stained sections of lungs from uninfected 5-wk-old transgenicmice and control WT littermates were also compared. The archi-tecture of lungs from transgenic mice were indistinct from that oflungs from littermates with respect to septal thickness, type II cellhypertrophy, and hyperplasia (not shown). There was no evidenceof pulmonary edema or vascular congestion, and inflammatorycells were not detected in lung sections from uninfected transgenicmice. In addition, there were no differences in the total cell countor the distribution of cell types recovered from BAL fluid.
Effect of transgene expression on killing of bacteria ininfected mice
Killing of GBS. To determine whether increased lysozyme levelsin the airway enhanced killing of bacteria in the lungs, quantitativecultures of lung homogenates from transgenic mice and WT lit-termate controls were compared following intratracheal injection
FIGURE 2. Lysozyme enzyme activity is significantly increased in theBAL fluid of transgenic mice. Lysozyme enzyme activity was assessed inBAL fluid from 5-wk-old transgenic mice from lines 3.5 and 2.6 and WTlittermates. Ten nanograms of BAL fluid protein was incubated withMi-crococcus lysodeikticussuspended at an OD of 1 at 450 nM. Following 1 hof incubation at 37°C, the change in OD of the suspension was determined.Purified chicken egg white lysozyme was used to generate a standardcurve. Data are the mean6 SEM. n, number of animals per experiment.
FIGURE 1. Expression of rat lysozyme in transgenic mice.A, Repre-sentative Northern blot analysis of 10mg of total cellular RNA from lungtissue isolated from 5-wk-old transgenic mice from line 3.5 (lanes 4and5,left panel;lanes 3and4, right panel) and WT littermates (lanes 1–3,leftpanel;lanes 1,2, and5, right panel). Samples in theleft panelwere probedwith the labeled mouse lysozyme cDNA, which detected both mouse ly-sozyme and the rat lysozyme transgene (;1 kb); samples in theright panelwere probed with the rat lysozyme cDNA, which detected only rat ly-sozyme (1, samples from transgene-positive mice).B, Western blot anal-ysis of 0.5mg of total lung protein from 5-wk-old transgenic mice and WTlittermates. Proteins were fractionated by SDS-PAGE, blotted onto nitro-cellulose membrane, and incubated with anti-human lysozyme Ab, whichdetected both mouse and rat lysozyme,Mr, ;15K (1, samples from trans-gene-positive mice).
5762 LYSOZYME AND PULMONARY HOST DEFENSE
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

of 106 CFU of GBS. All mice survived until sacrifice at 6 h postin-fection. Killing of GBS was enhanced 3.2-fold in mice from trans-genic line 3.5 (2.16 0.13 106 CFU/g protein vs 6.86 0.53 106
in WT mice;p 5 0.01; Fig. 4). The incidence of systemic dissem-ination of infection, as assessed by growth of GBS on plates in-oculated with splenic homogenates, was less in transgenic mice(27 vs 60%;p 5 0.04). Killing of GBS was enhanced 1.7-fold inmice from transgenic line 2.6 (4.26 0.8 3 106 CFU/g protein vs7.1 6 0.6 3 106 in WT mice; p 5 0.04).
Killing of P. aeruginosa. All transgenic mice survived until sac-rifice at 24 h postinfection following intratracheal infection with107 CFU of P. aeruginosa; in contrast, 20% of infected WT micedied. Bacterial killing was enhanced 30.6-fold in mice from trans-genic line 3.5 (1.066 0.053 106 CFU/g protein compared with32.4 6 0.41 3 106 in WT littermate controls;p 5 0.03; Fig. 5).Bacterial killing was enhanced approximately 6.2-fold in micefrom transgenic line 2.6 (6.526 0.713 106 CFU/g protein;p 50.05). Systemic dissemination was not detected in surviving miceat 24 h postinfection. Following intratracheal infection with 23
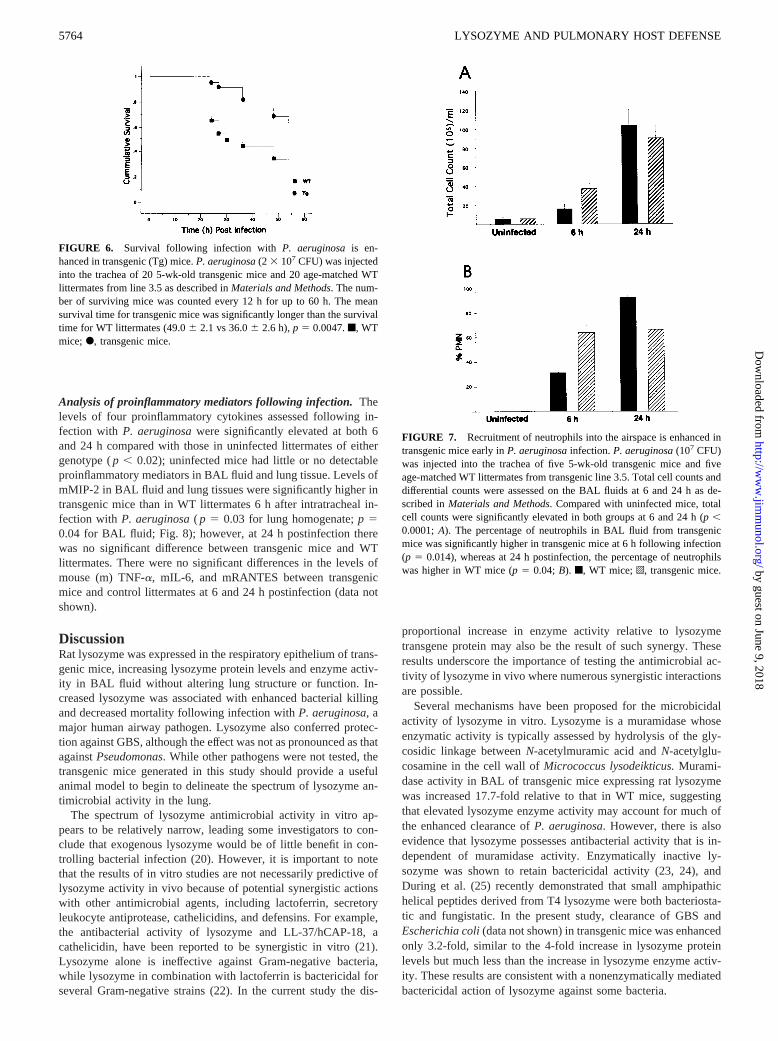
107 CFU of P. aeruginosa, the mean survival time for transgenicmice (49.06 2.1 h) was significantly longer than that of WTlittermates (36.06 2.6 h;p 5 0.0047; Fig. 6).
Neutrophil recruitment followingP. aeruginosainfection. Fol-lowing intratracheal infection with 107 CFU of Pseudomonasaeruginosa, total cell count and neutrophil influx were assessed inthe lungs of transgenic mice and WT littermate controls. Total cellcounts were significantly increased in WT and transgenic micecompared with uninfected littermates at 6 and 24 h postinfection( p , 0.0001; Fig. 7A). The percentage of neutrophils in BAL fluidwas significantly higher in transgenic mice than in WT littermates(61.56 5.6 vs 29.96 7.2; p 5 0.014) at 6 h postinfection; how-ever, at 24 h postinfection, the percentage of neutrophils in theBAL fluid was higher in WT mice (88.96 1.9 vs 63.76 8.0;p 50.04; Fig. 7B).
FIGURE 5. Killing of P. aeruginosaat 24 h postinfection is enhancedin transgenic mice.P. aeruginosa(107 CFU) was injected into the tracheaof 5-wk-old transgenic mice and control WT littermates from lines 3.5 and2.6 as described inMaterials and Methods. Bacterial counts were deter-mined by quantitative cultures of lung homogenates at 24 h postinfection.Data are expressed as CFU per gram of lung tissue6 SEM. n, Number ofanimals per experiment.p 5 0.03 for WT vs transgenic line 3.5;p 5 0.05for WT vs transgenic line 2.6.
FIGURE 3. Lysozyme protein is expressed in alveolar cells and bronchiolar epithelial cells in transgenic mice. Cellular localization of lysozyme proteinin transgenic mice was assessed by immunohistochemistry on lung sections from 5-wk-old transgenic mice from line 3.5 (C) and WT littermates (B) usinganti-human lysozyme Ab. No staining was detected when parallel sections from transgenic mice were incubated with nonimmune rabbit serum (A).Lysozyme was detected in alveolar epithelial cells (arrow) in WT and transgenic mice. Lysozyme expression was also detected in bronchiolar epithelialcells of transgenic mice, but not WT mice (arrowheads). Magnification,3240.
FIGURE 4. Killing of GBS at 6 h postinfection is enhanced in trans-genic mice. GBS (106 CFU) was injected into the trachea of 5-wk-oldtransgenic mice from lines 3.5 and 2.6 and control WT littermates as de-scribed inMaterials and Methods. Bacterial counts were determined byquantitative culture of equal volumes of lung homogenates 6 h postinfec-tion. Data are expressed as CFU per gram of lung tissue6 SEM.n, numberof animals per experiment.p 5 0.01 for WT vs transgenic;p 5 3.5 and0.04 for WT vs transgenic line 2.6.
5763The Journal of Immunology
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Analysis of proinflammatory mediators following infection.Thelevels of four proinflammatory cytokines assessed following in-fection with P. aeruginosawere significantly elevated at both 6and 24 h compared with those in uninfected littermates of eithergenotype (p , 0.02); uninfected mice had little or no detectableproinflammatory mediators in BAL fluid and lung tissue. Levels ofmMIP-2 in BAL fluid and lung tissues were significantly higher intransgenic mice than in WT littermates 6 h after intratracheal in-fection with P. aeruginosa( p 5 0.03 for lung homogenate;p 50.04 for BAL fluid; Fig. 8); however, at 24 h postinfection therewas no significant difference between transgenic mice and WTlittermates. There were no significant differences in the levels ofmouse (m) TNF-a, mIL-6, and mRANTES between transgenicmice and control littermates at 6 and 24 h postinfection (data notshown).
DiscussionRat lysozyme was expressed in the respiratory epithelium of trans-genic mice, increasing lysozyme protein levels and enzyme activ-ity in BAL fluid without altering lung structure or function. In-creased lysozyme was associated with enhanced bacterial killingand decreased mortality following infection withP. aeruginosa, amajor human airway pathogen. Lysozyme also conferred protec-tion against GBS, although the effect was not as pronounced as thatagainstPseudomonas. While other pathogens were not tested, thetransgenic mice generated in this study should provide a usefulanimal model to begin to delineate the spectrum of lysozyme an-timicrobial activity in the lung.
The spectrum of lysozyme antimicrobial activity in vitro ap-pears to be relatively narrow, leading some investigators to con-clude that exogenous lysozyme would be of little benefit in con-trolling bacterial infection (20). However, it is important to notethat the results of in vitro studies are not necessarily predictive oflysozyme activity in vivo because of potential synergistic actionswith other antimicrobial agents, including lactoferrin, secretoryleukocyte antiprotease, cathelicidins, and defensins. For example,the antibacterial activity of lysozyme and LL-37/hCAP-18, acathelicidin, have been reported to be synergistic in vitro (21).Lysozyme alone is ineffective against Gram-negative bacteria,while lysozyme in combination with lactoferrin is bactericidal forseveral Gram-negative strains (22). In the current study the dis-
proportional increase in enzyme activity relative to lysozymetransgene protein may also be the result of such synergy. Theseresults underscore the importance of testing the antimicrobial ac-tivity of lysozyme in vivo where numerous synergistic interactionsare possible.
Several mechanisms have been proposed for the microbicidalactivity of lysozyme in vitro. Lysozyme is a muramidase whoseenzymatic activity is typically assessed by hydrolysis of the gly-cosidic linkage betweenN-acetylmuramic acid andN-acetylglu-cosamine in the cell wall ofMicrococcus lysodeikticus.Murami-dase activity in BAL of transgenic mice expressing rat lysozymewas increased 17.7-fold relative to that in WT mice, suggestingthat elevated lysozyme enzyme activity may account for much ofthe enhanced clearance ofP. aeruginosa. However, there is alsoevidence that lysozyme possesses antibacterial activity that is in-dependent of muramidase activity. Enzymatically inactive ly-sozyme was shown to retain bactericidal activity (23, 24), andDuring et al. (25) recently demonstrated that small amphipathichelical peptides derived from T4 lysozyme were both bacteriosta-tic and fungistatic. In the present study, clearance of GBS andEscherichia coli(data not shown) in transgenic mice was enhancedonly 3.2-fold, similar to the 4-fold increase in lysozyme proteinlevels but much less than the increase in lysozyme enzyme activ-ity. These results are consistent with a nonenzymatically mediatedbactericidal action of lysozyme against some bacteria.
FIGURE 6. Survival following infection with P. aeruginosais en-hanced in transgenic (Tg) mice.P. aeruginosa(2 3 107 CFU) was injectedinto the trachea of 20 5-wk-old transgenic mice and 20 age-matched WTlittermates from line 3.5 as described inMaterials and Methods. The num-ber of surviving mice was counted every 12 h for up to 60 h. The meansurvival time for transgenic mice was significantly longer than the survivaltime for WT littermates (49.06 2.1 vs 36.06 2.6 h),p 5 0.0047.f, WTmice;F, transgenic mice.
FIGURE 7. Recruitment of neutrophils into the airspace is enhanced intransgenic mice early inP. aeruginosainfection.P. aeruginosa(107 CFU)was injected into the trachea of five 5-wk-old transgenic mice and fiveage-matched WT littermates from transgenic line 3.5. Total cell counts anddifferential counts were assessed on the BAL fluids at 6 and 24 h as de-scribed inMaterials and Methods. Compared with uninfected mice, totalcell counts were significantly elevated in both groups at 6 and 24 h (p,0.0001;A). The percentage of neutrophils in BAL fluid from transgenicmice was significantly higher in transgenic mice at 6 h following infection(p 5 0.014), whereas at 24 h postinfection, the percentage of neutrophilswas higher in WT mice (p5 0.04;B). f, WT mice;o, transgenic mice.
5764 LYSOZYME AND PULMONARY HOST DEFENSE
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Enhanced bacterial killing in transgenic mice may be partly dueto increased recruitment of neutrophils early in the course of in-fection. Neutrophil influx into the airspaces was slightly, but sig-nificantly, higher in transgenic mice at 6 h postinfection; further,the level of mMIP-2, a neutrophil chemotactic factor, was signif-icantly elevated in transgenic mice at this time point. In contrast,neutrophil numbers and mMIP-2 levels in unchallenged transgenicmice were not different from those in WT littermates. These datasuggest that constitutive overexpression of lysozyme in the air-ways may enhance neutrophil recruitment following infection. Al-though it is reasonable to speculate that increased neutrophil influxmay contribute to enhanced bacterial killing in these transgenicmice, this hypothesis has not been directly tested.
Lysozyme was previously identified in rat alveolar type II epi-thelial cells by immunohistochemistry (5, 6). In the present studylysozyme was amplified by PCR using cDNA generated fromhighly purified isolated rat type II epithelial cells; the deducedamino acid sequence of the amplified product was identical withthe previously reported sequence for rat lysozyme (26) with thesingle exception of a substitution of glycine for tryptophan at po-sition 46. Immunogold labeling and subcellular fraction of type IIepithelial cells localized lysozyme to lamellar bodies, secretorygranules in which pulmonary surfactant is stored (27–29). Follow-ing secretion, lysozyme was detected in association with tubularmyelin, a lattice-like network of surfactant membranes (27, 28).Interestingly SP-A, another lamellar body protein involved in air-way host defense (30, 31), was also associated with tubular myelin(28, 32), raising the possibility that this structure may serve as ascaffold for host defense proteins involved in alveolar surveillance.
Although the present study has not defined the precise mecha-nism by which lysozyme promotes bacterial killing in vivo, theseresults have important implications for enhancing airway host de-fense. Lysozyme levels in transgenic mice were constitutively el-evated without affecting lung structure, suggesting that lysozymecould be safely administered for extended therapy. Most impor-tantly, elevated levels of lysozyme significantly enhanced killingof P. aeruginosa, the most common pathogen associated withchronic colonization of the airway in cystic fibrosis patients. Ly-sozyme administration alone or in combination with other agents,such as elastase inhibitors (33), may provide an important adjunctto treatment of chronic lung infections with antibiotics.
AcknowledgmentsWe thank Dr. Jeffrey Whitsett for insightful suggestions and for review ofthe manuscript.
References1. Konstan, M. W., P. W. Chen, J. M. Sherman, M. J. Thomassen, R. E. Wood, and
T. F. Boat. 1981. Human lung lysozyme: sources and properties.Am. Rev. Respir.Dis. 123:120.
2. Imoto, T., L. N. Johnson, A. C. T. North, D. C. Phillips, and J. A. Rupley. 1972.Vertebrate lysozymes. InThe Enzymes, Vol. 7. P. D. Boyer, ed. Academic Press,New York, p. 665.
3. Coonrod, J. D. 1986. The role of extracellular bactericidal factors in pulmonaryhost defense.Semin. Respir. Infect. 1:118.
4. Travis, S. M., B. A. Conway, J. Zabner, J. J. Smith, N. N. Anderson, P. K. Singh,E. Peter Greenberg, and M. J. Welsh. 1999. Activity of abundant antimicrobialsof the human airway.Am. J. Respir. Cell Mol. Biol. 20:872.
5. Singh, G., S. L. Katyal, W. E. Brown, D. L. Collins, and R. J. Mason. 1988.Pulmonary lysozyme: a secretory protein of type II pneumocytes in the rat.Am.Rev. Respir. Dis. 138:1261.
6. Spicer, S. S., R. Frayser, G. Virella, and B. J. Hall. 1977. Immunocytochemicallocalization of lysozymes in respiratory and other tissues.Lab. Invest. 36:282.
7. Yeh, T. C., A. C. Wilson, and D. M. Irwin. 1993. Evolution of rodent lysozymes:isolation and sequence of the rat lysozyme genes.Mol. Phylogenet. Evol. 2:65.
8. Cross, M., I. Mangelsdorf, A. Wedel, and R. Renkawitz. 1988. Mouse lysozymeM gene: isolation, characterization, and expression studies.Proc. Natl. Acad. Sci.USA 85:6232.
9. Cortopassi, G. A., and A. C. Wilson. 1990. Recent origin of the P lysozyme genein mice.Nucleic Acids Res. 18:1911.
10. Glasser, S. W., T. R. Korfhagen, S. E. Wert, M. D. Bruno, K. M. McWilliams,D. K. Vorbroker, and J. A. Whitsett. 1991. Genetic element from human surfac-tant protein SP-C gene confers bronchiolar-alveolar cell specificity in transgenicmice.Am. J. Physiol. 261:L349.
11. Wert, S. E., S. W. Glasser, T. R. Korfhagen, and J. A. Whitsett. 1993. Transcrip-tional elements from the human SP-C gene direct expression in the primordialrespiratory epithelium of transgenic mice.Dev. Biol. 156:426.
12. Glasser, S. W., M. S. Burhans, S. K. Eszterhas, M. D. Bruno, andT. R. Korfhagen. 2000. Human SP-C gene sequences that confer lung epithelium-specific expression in transgenic mice.Am. J. Physiol. 278:L933.
13. Beck, D. C., C. L. Na, J. A. Whitsett, and T. E. Weaver. 2000. Ablation of acritical surfactant protein B intramolecular disulfide bond in transgenic mice.J. Biol. Chem. 275:3371.
14. Korfhagen, T. R., S. W. Glasser, S. E. Wert, M. D. Bruno, C. C. Daugherty,J. D. McNeish, J. L. Stock, S. S. Potter, and J. A. Whitsett. 1990.Cis-actingsequences from a human surfactant protein gene confer pulmonary-specific geneexpression in transgenic mice.Proc. Natl. Acad. Sci. USA 87:6122.
15. Smith, P. K., R. I. Krohn, G. T. Hermanson, A. K. Mallia, F. H. Gartner,M. D. Provenzano, E. K. Fujimoto, N. M. Goeke, B. J. Olson, and D. C. Klenk.1985. Measurement of protein using bicinchoninic acid.Anal. Biochem. 150:76.
16. Akinbi, H. T., J. S. Breslin, M. Ikegami, H. S. Iwamoto, J. C. Clark,J. A. Whitsett, A. H. Jobe, and T. E. Weaver. 1997. Rescue of SP-B knockoutmice with a truncated SP-B proprotein: function of the C-terminal propeptide.J. Biol. Chem. 272:9640.
17. Shugar, D. 1952. Measurement of lysozyme activity and ultra-violet inactivationof lysozyme.Biochim. Biophys. Acta 8:302.
18. Lin, S., C. L. Na, H. T. Akinbi, K. S. Apsley, J. A. Whitsett, and T. E. Weaver.1999. Surfactant protein B (SP-B)2/2 mice are rescued by restoration of SP-Bexpression in alveolar type II cells but not Clara cells.J. Biol. Chem. 274:19168.
19. Akinbi, H. T., H. Bhatt, W. M. Hull, and T. E. Weaver. 1999. Altered surfactantprotein B levels in transgenic mice do not affect clearance of bacteria from thelungs.Pediatr. Res. 46:530.
20. Neu, H. C., J. d. Dreyfus, and R. E. Canfield. 1968. Effect of human lysozyme onGram-positive and Gram-negative bacteria.Antimicrob. Agents Chemother. 8:442.
21. Bals, R., X. Wang, M. Zasloff, and J. M. Wilson. 1998. The peptide antibiotic
FIGURE 8. Levels of mMIP-2 are signifi-cantly increased in transgenic mice 6 h postin-fection. P. aeruginosa(107 CFU) was injectedinto the trachea of eight 5-wk-old transgenicmice and eight age-matched control WT litter-mates from transgenic line 3.5. The levels ofproinflammatory mediators were assessed inBAL fluid and lung tissues at 6 and 24 h postin-fection and in an equal number of uninfected lit-termates as described inMaterials and Methods.Levels of mTNF-a, mIL-6, and mRANTESwere not significantly different between trans-genic mice and control WT littermates. Levels ofmMIP-2 were significantly higher in transgenicmice at 6 h in both BAL (p5 0.04;B) and lunghomogenates (p5 0.03; A). f, WT mice; M,transgenic mice.
5765The Journal of Immunology
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

LL-37/hCAP-18 is expressed in epithelia of the human lung where it has broadantimicrobial activity at the airway surface.Proc. Natl. Acad. Sci. USA 95:9541.
22. Ellison, R. T. D., and T. J. Giehl. 1991. Killing of Gram-negative bacteria bylactoferrin and lysozyme.J. Clin. Invest. 88:1080.
23. Laible, N. J., and G. R. Germaine. 1985. Bactericidal activity of human ly-sozyme, muramidase-inactive lysozyme, and cationic polypeptides againstStrep-tococcus sanguisand Streptococcus faecalis: inhibition by chitin oligosaccha-rides.Infect. Immun. 48:720.
24. Pellegrini, A., U. Thomas, R. von Fellenberg, and P. Wild. 1992. Bactericidalactivities of lysozyme and aprotinin against Gram-negative and Gram-positivebacteria related to their basic character.J. Appl. Bacteriol. 72:180.
25. During, K., P. Porsch, A. Mahn, O. Brinkmann, and W. Gieffers. 1999. Thenon-enzymatic microbicidal activity of lysozymes.FEBS Lett. 449:93.
26. White, T. J., G. A. Mross, E. F. Osserman, and A. C. Wilson. 1977. Primarystructure of rat lysozyme.Biochemistry 16:1430.
27. Power, J. H. T., H. A. Barr, and T. E. Nicholas. 1993. Characterization andimmunohistochemical localization of the 15 kD protein isolated from rat lunglamellar bodies.Am. J. Respir. Cell Mol. Biol. 8:98.
28. Haller, E. M., S. A. Shelley, M. R. Montgomery, and J. U. Balis. 1992. Immu-nocytochemical localization of lysozyme and surfactant protein-A in rat type-IIcells and extracellular surfactant forms.J. Histochem. Cytochem. 40:1491.
29. Beers, M. F., C. Y. Kim, C. Dodia, and A. B. Fisher. 1994. Synthesis of type IIcell lamellar body lysozyme 15 kD protein (lbl-15) by perfused rat lung.Am. J. Respir. Cell Mol. Biol. 11:240.
30. LeVine, A. M., M. D. Bruno, K. M. Huelsman, G. F. Ross, J. A. Whitsett, andT. R. Korfhagen. 1997. Surfactant protein A-deficient mice are susceptible togroup streptococcal infection.J. Immunol. 158:4336.
31. LeVine, A. M., K. E. Kurak, M. D. Bruno, J. M. Stark, J. A. Whitsett, andT. R. Korfhagen. 1998. Surfactant protein-A-deficient mice are susceptible toPseudomonas aeruginosainfection.Am. J. Respir. Cell Mol. Biol. 19:700.
32. Voorhout, W. F., T. Veenendaal, H. P. Haagsman, A. J. Verkleij,L. M. G. van Golde, and H. J. Geuze. 1991. Surfactant protein A is localized atthe corners of the pulmonary tubular myelin lattice.J. Histochem. Cytochem.39:1331.
33. Cantin, A. M., and D. E. Woods. 1999. Aerosolized prolastin suppresses bacterialproliferation in a model of chronic pseudomonas aeruginosa lung infection.Am. J. Respir. Crit. Care Med. 160:1130.
5766 LYSOZYME AND PULMONARY HOST DEFENSE
by guest on June 9, 2018http://w
ww
.jimm
unol.org/D
ownloaded from